


-
海豚链球菌(Streptococcus iniae)是一种重要的水生动物致病菌,1976年Pier等[1]从亚马逊江豚(Inia geoffrensis)中首次分离获得,之后陆续发现它可感染除南极洲以外的所有鱼类[2]。我国也有海豚链球菌感染卵形鲳鲹(Trachinotus ovatus)[3-4]、尼罗罗非鱼(Oreochromis niloticus)[5-6]、鲟鱼(Acipenser sinensis)[7-8]、银鼓鱼(Selenotoca multifasciata)[9]、小黄鱼(Larimichthys polyactis)[10]、黄鳍鲷(Acanthopagrus latus)[11]等发病的相关报道。海豚链球菌具有感染宿主范围广、传染性强、导致死亡率高等特点,给养殖业造成巨大的经济损失,并受到了国内外水产行业工作者的高度关注。
海豚链球菌病的暴发除了与病原菌本身的毒力有关,一些环境因子在疾病发生发展过程中亦发挥重要作用,如温度、pH值等[12],尤其是养殖的环境温度与该病的发生呈现比较明显的相关性,发病时水温通常在27 ℃以上[13]。祝璟琳等[14]开展的流行病学研究表明,在32 ℃以上的温度下罗非鱼更容易爆发链球菌病,罗非鱼免疫力在高温条件下受到明显的抑制[15],链球菌对宿主的粘附、定植、入侵和抵抗免疫清除能力明显增强[16-17]。同时,较高温度时海豚链球菌产生的胞外产物中可能含有较多致病因子,从而影响细菌毒力[18],尼罗罗非鱼在感染海豚链球菌后其肝脏、肾脏在高温时病理损伤更严重[19]。以上研究均表明在较高的温度下病原链球菌的毒力显著增强,从而引起链球菌病的爆发。
转录组测序(RNA-Seq)技术是一项能够检测有机体整体转录水平的新兴技术手段,目前已有学者利用转录组技术研究细菌的毒力调控。如:Li等[20]利用RNA-seq技术分析了提高温度和营养成分对副溶血性弧菌毒力的影响,结果发现温度和营养成分的提高可显著提高副溶血性弧菌的毒力;王松艳[21]对不同温度下培养的无乳链球菌进行转录组测序,筛选出7个毒力相关基因;师若萍[22]运用高通量RNA测序和非靶向代谢组学的研究方法,探究大肠杆菌在不同温度下的生长及其在转录和代谢水平的变化,了解其对于不同温度的生理调节机制,以上研究结果表明温度可广泛影响细菌毒力和基因的表达。但目前尚未见温度对海豚链球菌致病性的影响以及其基因转录表达的研究报道。卵形鲳鲹是我国海水养殖鱼类中产量位居第二的优良品种,高温期易暴发海豚链球菌病[3-4]。本研究以卵形鲳鲹源海豚链球菌Tozj-1为研究对象,选用RNA-Seq技术对不同温度(25 ℃和35 ℃)培养的海豚链球菌进行高通量测序分析,筛选差异表达基因,对差异表达基因进行GO功能和KEGG通路富集分析,探究温度对海豚链球菌基因转录表达的影响及富集的功能和信号通路,以期为揭示温度与海豚链球菌致病性积累基础数据,为后续进一步研究海豚链球菌的致病机理提供理论依据。
-
本研究测序和试验用菌株Tozj-1来源于广东省水产动物病害防控与健康养殖重点实验室。Tozj-1为实验室于2020年8月从广东省湛江市某深海网箱养殖的患病卵形鲳鲹(Trachinotus ovatus)分离获得。分离的菌株经16S rRNA测序、生理生化鉴定为海豚链球菌(S. iniae),通过人工感染卵形鲳鲹确认其为病原菌,并保存于广东省水产动物病害防控与健康养殖重点实验室。
-
将Tozj-1菌株划线接种于BHI固体培养皿上,置于28 ℃恒温培养箱中培养24 h。用无菌接种环挑取单个菌落接种于新鲜的无菌BHI液体培养基,于28 ℃,120 r·min−1下振荡过夜扩大培养。将上述菌液以1∶50(V/V)的比例接种到无菌的BHI培养基,分别置于25 ℃和35 ℃恒温摇床,150 r·min−1振荡培养,每个温度下设置3个重复。每隔1 h取样,测定菌液的吸光值(OD600),并绘制不同温度下的海豚链球菌生长曲线。
-
为了探究不同温度对卵形鲳鲹源海豚链球菌毒力的影响,本研究使用卵形鲳鲹进行体内攻毒试验。实验前,将28 ℃扩大培养的海豚链球菌以1∶50(V/V)的比例接种到新鲜BHI培养基中,分别置于25、35 ℃恒温摇床中培养至对数生长期(OD600 =0.9~1.1),收集菌体,用无菌PBS缓冲液将菌液稀释至1.0×107 CFU·mL−1备用。
将360尾健康的卵形鲳鲹[体质量(50±6)g·尾]随机分为4组(2个实验组和2个对照组),每组设3个重复,每个重复30尾鱼。25 ℃实验组于水温25 ℃饲养,每尾腹腔注射25 ℃体外培养的Tozj-1菌悬液,对照组注射无菌PBS;35 ℃实验组于水温35 ℃饲养,每尾腹腔注射35 ℃体外培养的Tozj-1菌悬液,对照组注射等量无菌PBS。观察攻毒后14 d内试验鱼的发病情况,记录并统计死亡鱼数量,通过解剖濒临死亡鱼并进行病原分离鉴定,以确定感染病原,鉴定方法同1.1。
-
将28 ℃下培养的菌株Tozj-1以1∶50(V/V)的比例稀释于新鲜BHI肉汤中,分别于35 ℃和25 ℃条件下培养至对数生长期(OD600 =0.9~1.1)。每个温度设置3个生物学重复。其中,以25 ℃培养的海豚链球菌作为对照组,标记为CK-1、CK-2、CK-3;以35 ℃培养的海豚链球菌作为实验组,标记为Test-1、Test-2、Test-3。每个样品取1 mL菌液,12 000 r·min−1离心2 min,弃去上清,加入100 μL溶菌酶,混匀,置于37 ℃裂解15 min,按照Trizol试剂盒操作说明进行菌体的总RNA抽提,使用Agilent 2100生物分析仪评估RNA质量并通过RNase free琼脂糖凝胶电泳进行检测,经质量检测合格后纯化总RNA,cDNA文库构建和转录组测序工作委托广州基迪奥生物科技有限公司完成。
-
原始数据的存储采用FASTQ格式,通过fastp[23](
https://github.com/OpenGene/fastp )对下机数据进行质量检测和数据过滤,去除原始数据中质量低、接头污染、全部都是A碱基以及未知碱基N含量过高的Reads后最终获得Clean reads。利用工具Bowtie 2[24]建立参考基因组索引,将质控分析后的高质量测序数据比对到海豚链球菌参考基因组SF1(GenBank登录号CP005941.1)上。采用FPKM法计算每个注释基因的表达水平,使用FDR与log2FC来筛选差异基因,筛选条件为FDR<0.05且|log2FC|>1。 -
根据每组筛选的差异表达基因,使用R语言Pheatmap软件包对差异基因进行双向聚类分析。首先,得到的差异表达基因在GO(The Gene Ontology)数据库中的各个项目进行映射,然后通过GO功能显著性富集分析[25],在宏观上了解该物种的基因功能分布特征。然后,基于KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库,对每个KEGG pathway不同层次上存在的差异表达基因数量进行统计,确定这些差异表达基因的主要代谢路径和信号通路[26],进一步了解基因的生物功能。最后,利用超几何学方法,以Qvalue≤0.1的Pathway作为在差异表达基因中显著富集的Pathway。
-
为了验证海豚链球菌转录组数据的可靠性,将差异表达基因与毒力因子数据库(Virulence Factors of Pathogenic Bacteria,VFDB)进行比对,筛选出22个与毒力相关的差异表达基因,使用primer 5.0软件设计特异性定量引物(基因名称和引物序列见表1),引物委托生工生物工程(上海)股份有限公司合成。根据cDNA反转录试剂盒的使用说明书将质检合格的总RNA反转录为cDNA,应用实时荧光定量PCR(Real-time quantitative PCR, qPCR)技术对其进行转录水平的验证,qPCR反应程序为95 ℃预变性1 min,95 ℃变性15 s,60 ℃退火20 s,72 ℃延伸30 s,共计40个循环结束,60 ℃读取熔解曲线。以16S rRNA基因为内参基因,采用2–ΔΔCt方法分析计算各毒力基因的相对表达量。
表 1 检测海豚链球菌毒力基因的qRT-PCR特异性引物
功能分类 引物名称 上游引物序列(5′→3′) 下游引物序列(5′→3′) 内参基因 16S rRNA CCAAGGCGACGATACATAG TTGCCGAAGATTCCCTACT ABC转运蛋白 srtF GGAGCAGGAAAGTCAACG TGGATAGATCGCAGGATT oppF AAACCACTGTTGGTCGTGC TCTTTGCGGAAATCTCGTA oppA GGAAGTAATCTATTGCGTGTC TACTGCCATCAGACCATTT 双组份系统 ciaR AATGCGTATTCAAGCCCTCT AATGGTAGTGTCTCCGACT ciaH TGCTTTGAAATACACGGACTC AACACGATAGAAGCGGTCA phoR AGGCTAAACAACTCCACTT TCATCGGTGTACCTAACAG phoP ACAGCAACAGATGGCAGAT AGACCTCAATACCGTCCAA 溶血素 sagI ATGTCATTGTTATGCCAACT GTAAGGGCCTGCACTCAA CAMP因子 camp TGCCTCAACATCAAACAG ATGGCTTGAACAGCATCA 酶 htrA TCAGGTTCAAATGGAGGAG AAGGCAGGACGAATCACT danK CAGTTCGTCGTGCCCTTTC TTGGCTCTTTACCAGTTTCAG clpE GTCCCTCACAAACTCCAGC CTCTTCAAATTGACCACGA nanA GCCGTAACCGCTCACCACT TTTCAATCCCGTCAAGTCC hylB AACCCTGCTTCAACACTCT CCAATCTCATAATCCCACC C5α肽酶 scpB GGGTTGAGTGTCCGTAATT CTGACGCATCATCATCTGC 分泌系统 essC GCCGTCAACGCCTCTTCTC GGCATTGGCTCTGTCACTT secA2 AGCTGGAATTGAATACGC AGATAATGAGCCCGAAGG secY2 GCATTAAACCAGGACAAGC GATATTACCGCCACCAGAA secY ATAGCAGCTCAACAAACCC ATGCCTTCAATACCAGTCG 转录调节因子 cysB CGAAATCCTAAGGGCATCA CGTTCCTCCAACAAAGAG galR GAGGCTGGTATCACTGTTC GCAGTAGTTCCCATTTCCT 生物膜 glgA TAGCGTTTGCTCTTGATGT TAAGCCTGACTTGCCGTAT -
海豚链球菌在本试验的2个培养温度下,35 ℃生长速度明显快于25 ℃,能够快速进入生长对数期,35 ℃在3 h左右即达到对数生长期,25 ℃在9 h左右才达到对数生长期。到达生长平台期后,35 ℃的吸光值略高于25 ℃的吸光值。当吸光值OD600在1.0左右时,2个培养温度的海豚链球菌均处于对数生长期(图1)。
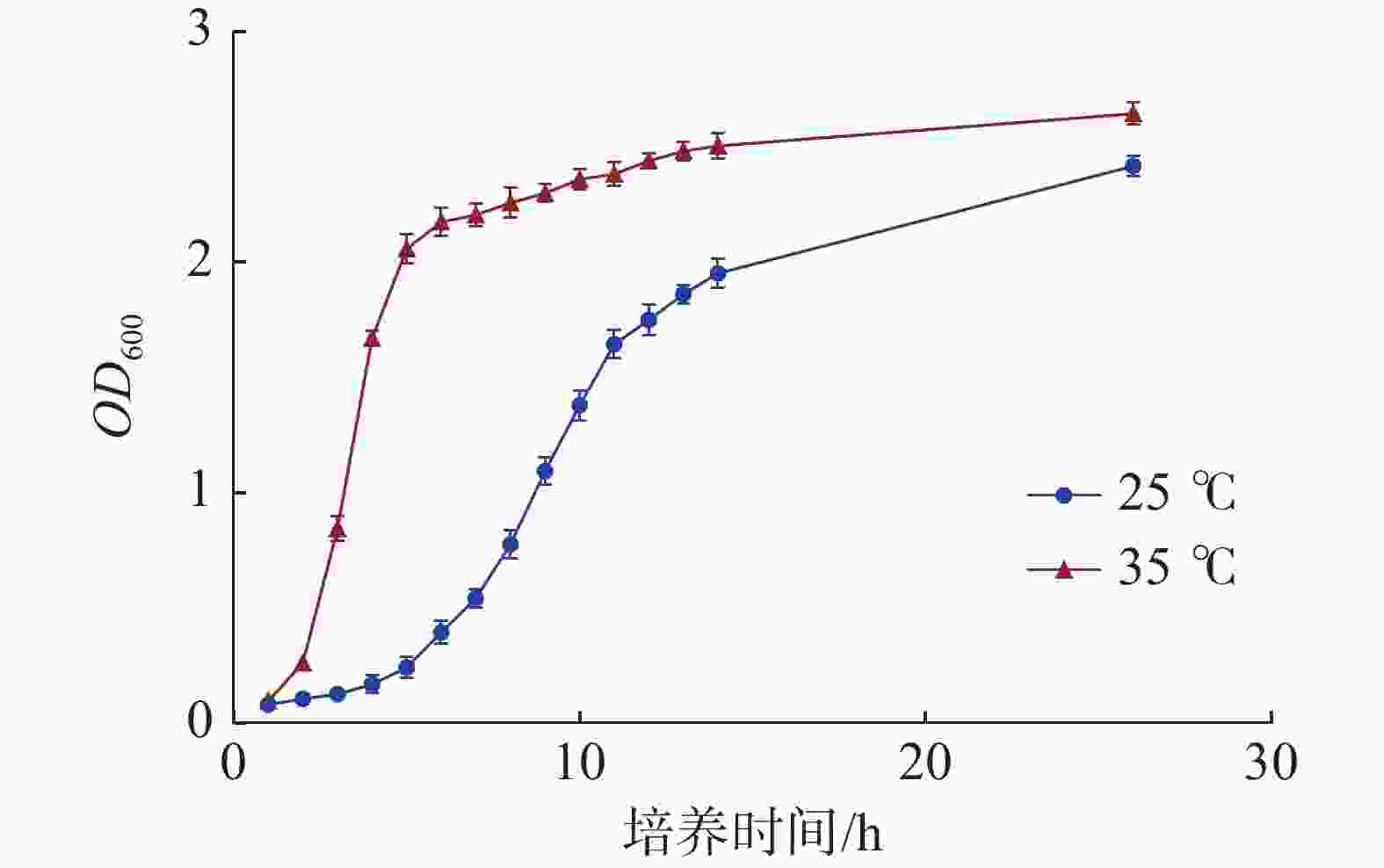
图 1 海豚链球菌在不同培养温度下的生长曲线
-
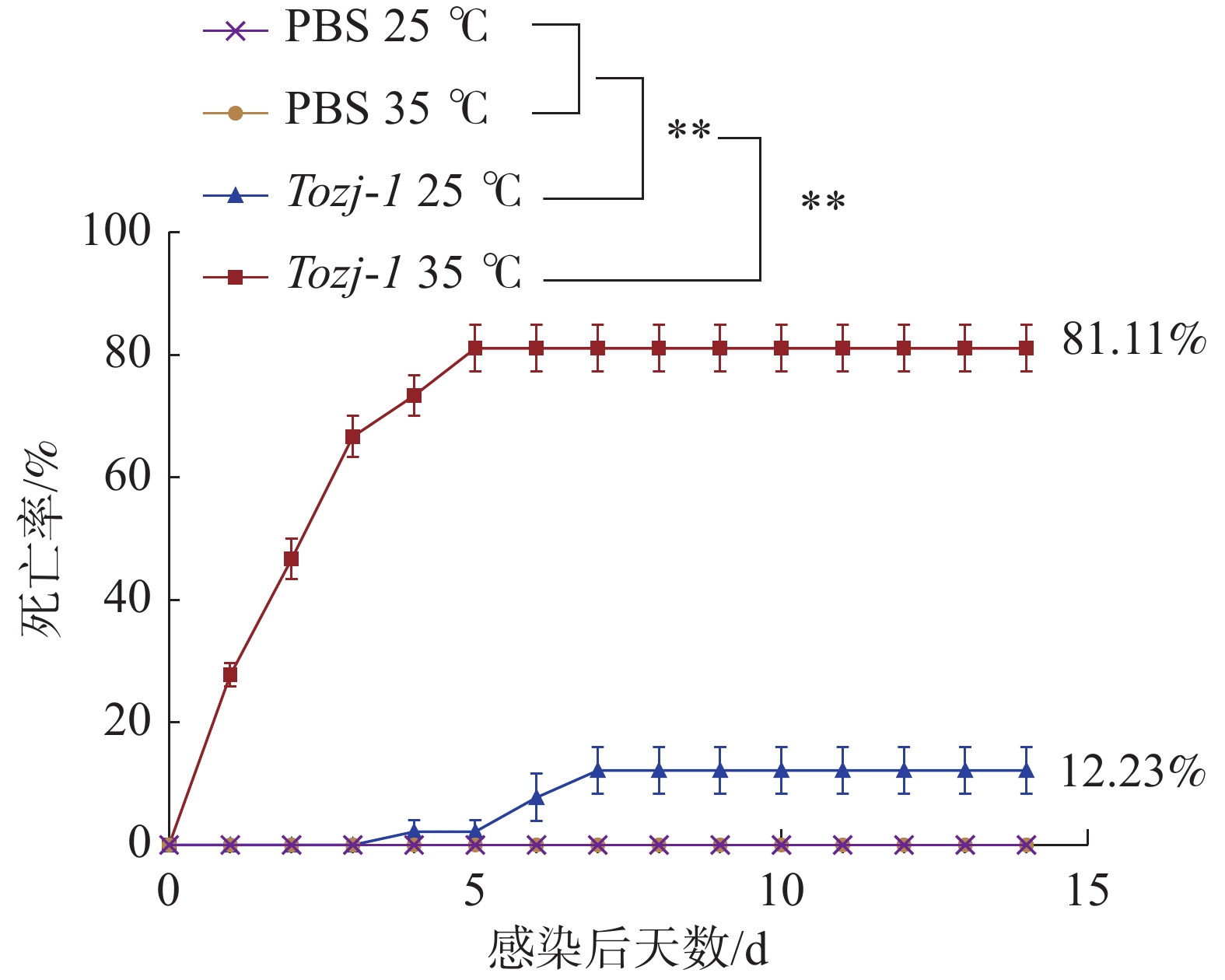
不同温度人工感染结果显示(图2),35 ℃时,在感染后1~3 d卵形鲳鲹即出现大量死亡,之后死亡量逐渐趋于稳定,第5 天停止死亡,患病卵形鲳鲹死亡前出现游泳异常、眼球突出、鳍条基部充血等症状,病原菌经分离培养,16S rRNA鉴定为海豚链球菌。25 ℃时,卵形鲳鲹在感染后3 天内未出现死亡,第4 天开始出现少量死亡,死亡率较低,第7天停止死亡。海豚链球菌在水温35 ℃和25 ℃造成的卵形鲳鲹的累积死亡率分别为81.11%和12.23%,PBS对照组均未出现死亡。
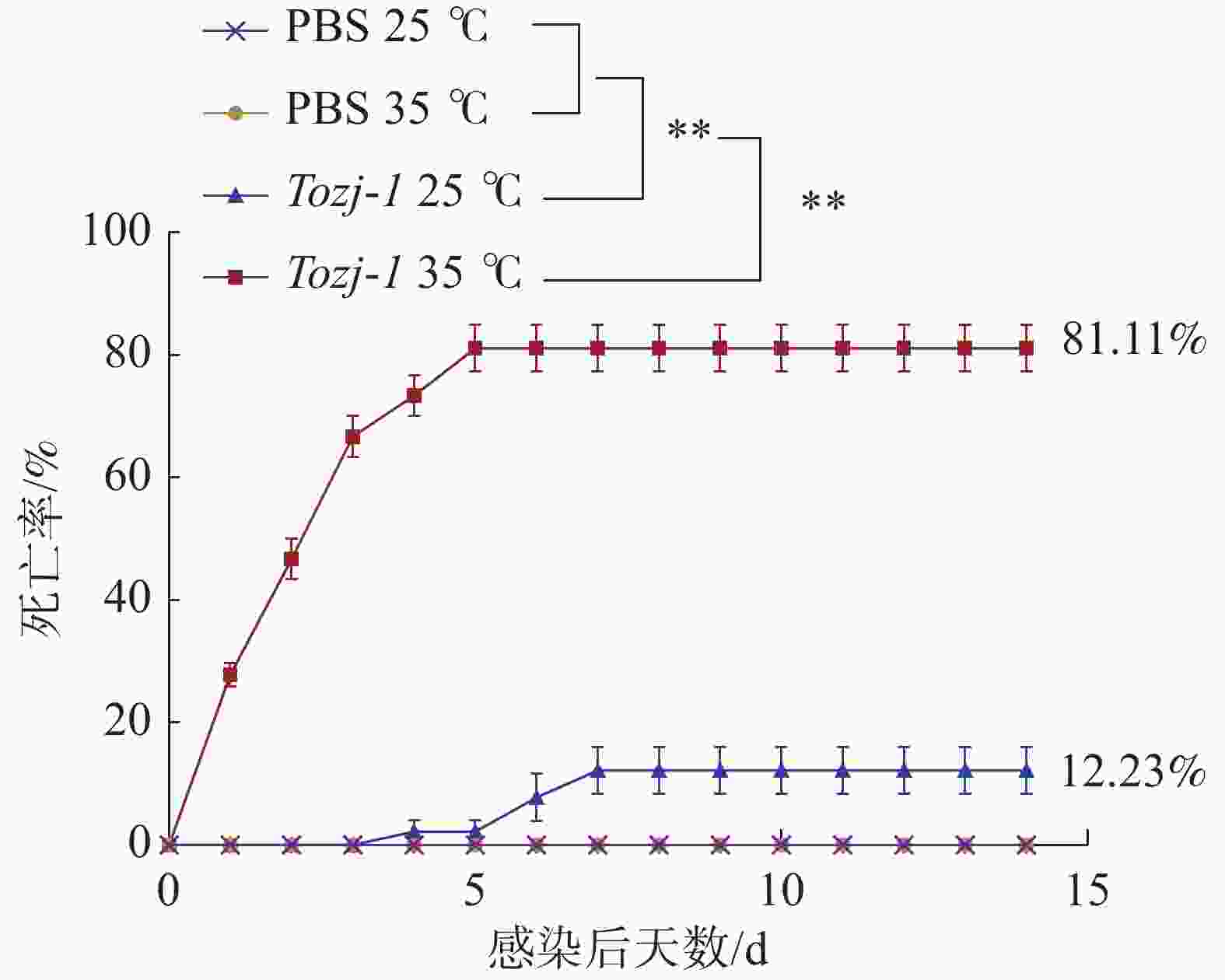
图 2 环境温度对卵形鲳鲹海豚链球菌病死亡率的影响
-
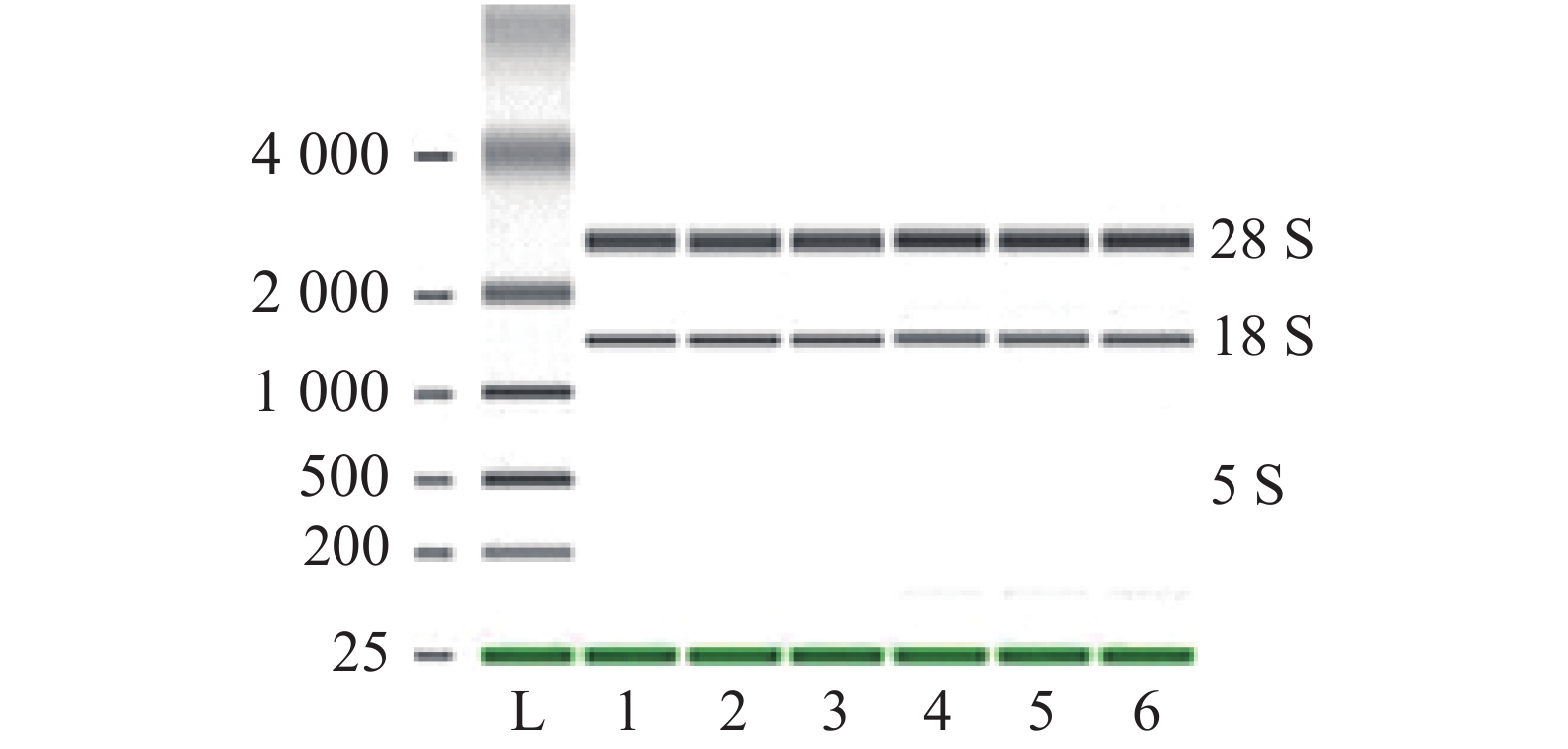
提取培养至对数生长期(OD600 =0.9~1.1)的海豚链球菌的总RNA,1%琼脂糖凝胶电泳结果如图3所示,28 S和18 S的条带清晰无弥散,RNA完整性好,无降解。核酸蛋白检测仪测定结果显示样品的A260 /A280比值在2.0左右,浓度为400~700 ng·μL−1,RNA质量符合转录组测序的要求。

图 3 1%琼脂糖凝胶电泳检测海豚链球菌Tozj-1株总RNA
-
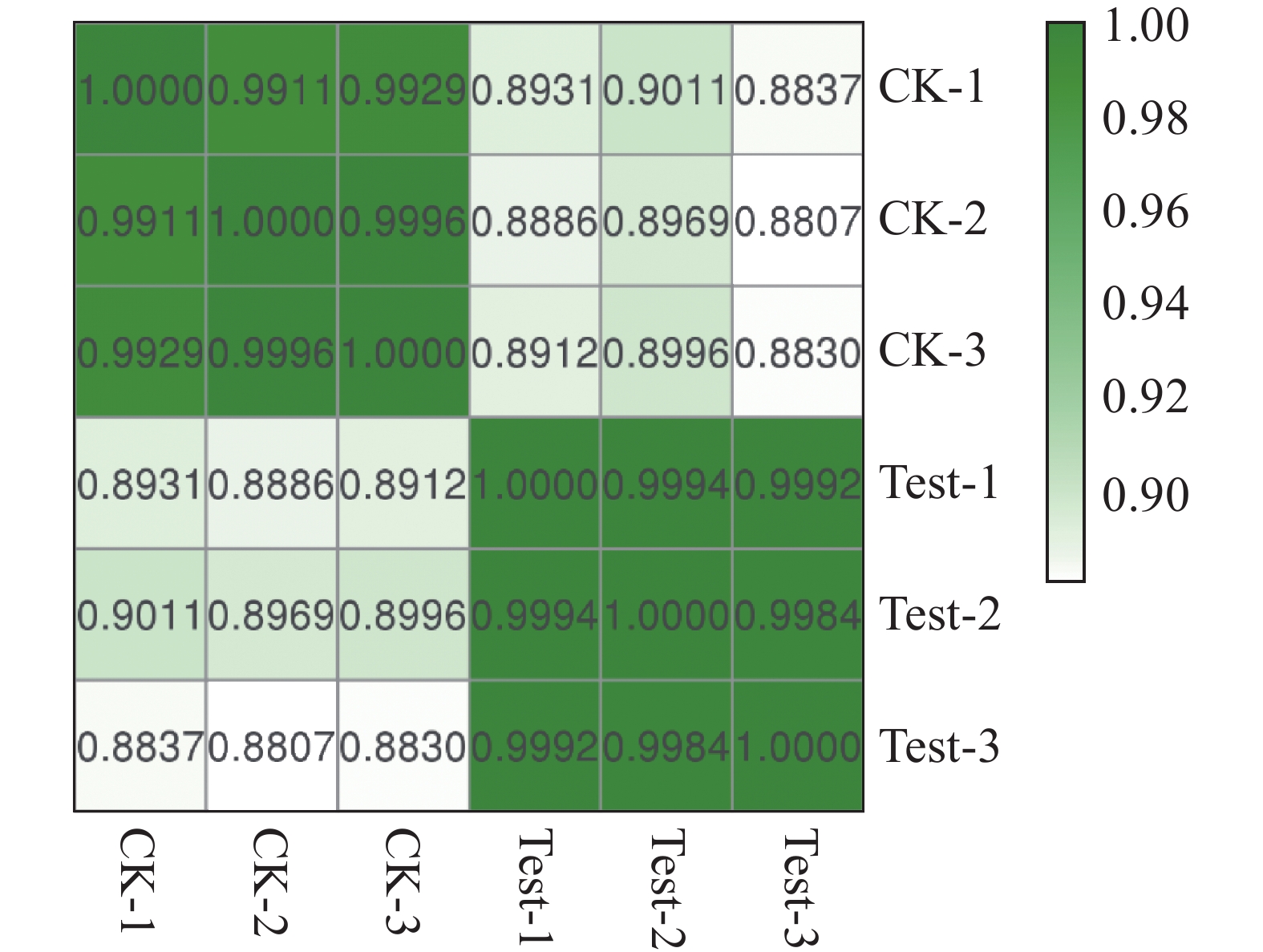
本试验中每个温度设有3个生物学重复,其中CK-1、CK-2、CK-3分别表示25 ℃的3个平行样品,Test-1、Test-2、Test-3分别表示35 ℃的3个平行样品。如图4所示,同一温度的3个平行样品相关性系数在0.998 0以上,表明来自3个重复性试验的转录组数据具有高度可重复性,可用于后续分析。
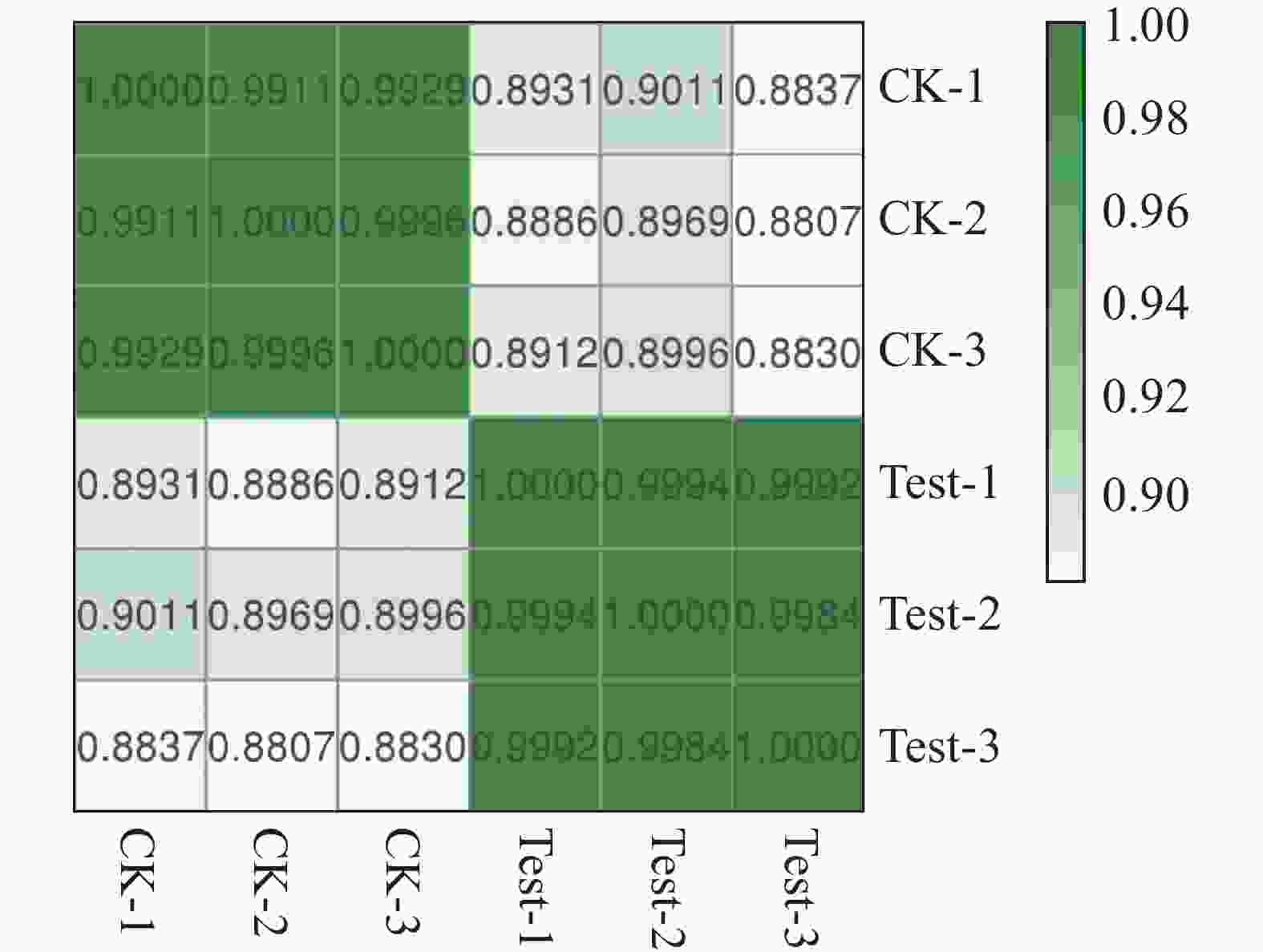
图 4 样品相关性分析热图
-
Illumina测序得到的数据经过质量检测和数据过滤,最终获得Clean reads,各样本的Clean reads与Raw reads的Q20和Q30分别达到了97%和93%以上,GC含量在各样本中较为一致,为40.19%~41.22%(表2)。以上数据均表明转录组测序的数据良好,可以用于后续进一步分析。
表 2 过滤前后碱基信息统计
样品 过滤前 过滤后 Raw Data/bp Q20/% Q30/% GC含量/% Raw Data/bp Q20/% Q30/% GC含量/% CK-1 3283961700 98.06 93.98 40.78 3202330021 98.11 94.08 40.49 CK-2 2481964500 98.20 94.34 41.20 2397726314 98.27 94.47 40.83 CK-3 2190706200 98.03 93.95 41.22 2118495261 98.11 94.09 40.85 Test-1 2753256900 98.03 93.97 40.76 2655059882 98.11 94.12 40.35 Test-2 2777076900 97.89 93.63 40.60 2677608106 97.98 93.80 40.19 Test-3 2516073000 97.77 93.33 41.10 2415735833 97.88 93.53 40.66 -
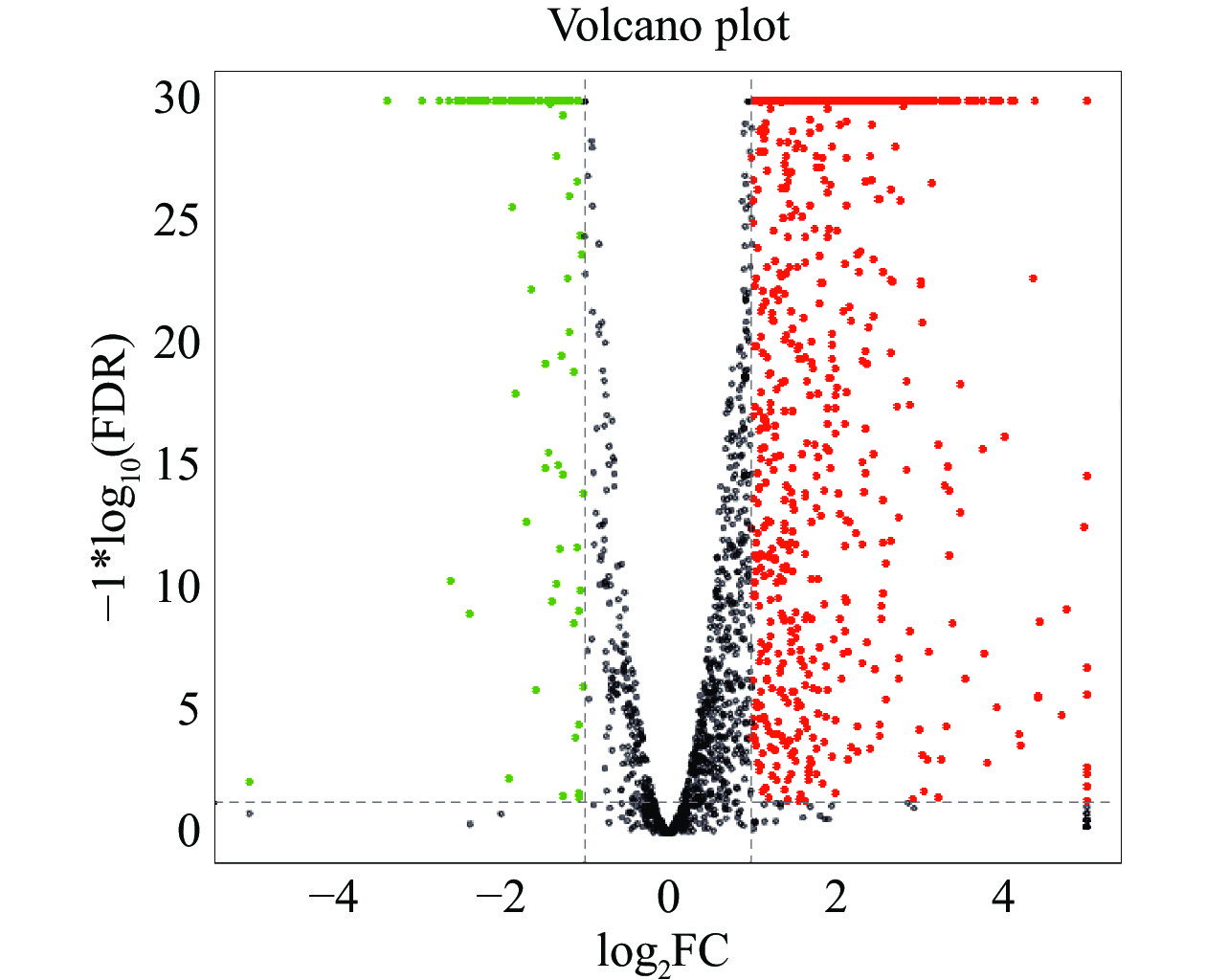
使用Bowtie 2软件将6个转录组数据映射到海豚链球菌SF1基因组,结果显示,所有样品数据皆可映射到基因组上,并且这些样本的映射率均在98%以上(表3)。说明该转录组数据可靠,可以用于后续的试验分析。差异表达基因结果如图5所示,共筛选到927个显著差异表达基因,其中820个上调表达,107个下调表达。
表 3 原始数据比对到基因组
样品 总reads数 匹配上的reads数 匹配比例/% CK-1 18162776 17938781 98.77 CK-2 13888834 13717694 98.77 CK-3 12276544 12129610 98.80 Test-1 16739444 16498781 98.56 Test-2 17315536 17063899 98.55 Test-3 14858622 14630754 98.47 
图 5 差异基因统计火山图
-
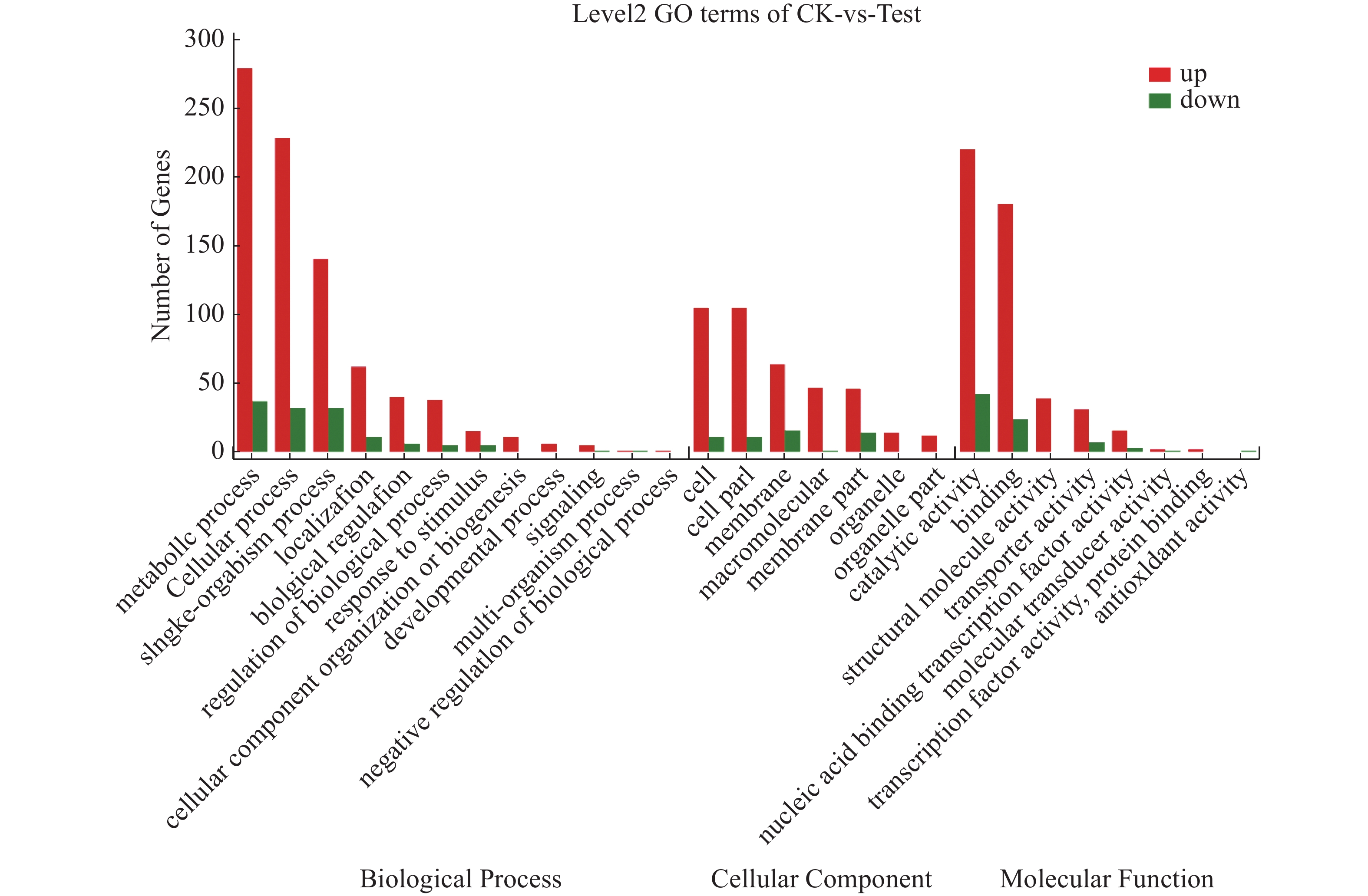
对获得的差异表达基因进行GO注释分析,共得到1 611个GO功能注释,其中生物学过程(biological process)959个、细胞组分(cellular component)446个和分子功能(molecular function)570个。如图6所示,差异基因注释在生物学过程有关的GO terms数量最多,共计12个,主要有代谢过程(metabolic process)、细胞过程(cellular process)和单一有机过程(single-organism process)等;其次是与分子功能有关的terms,共计8个,主要有催化活性(catalytic activity)、结合(binding)等;与细胞组分有关的terms最少,为7个,主要有细胞(cell)、细胞成分(cell part)和膜(membrane)等。海豚链球菌代谢过程、细胞过程、单一有机过程、结合过程和催化活性这几个亚类包含的差异基因比较多,它们均受温度调控。
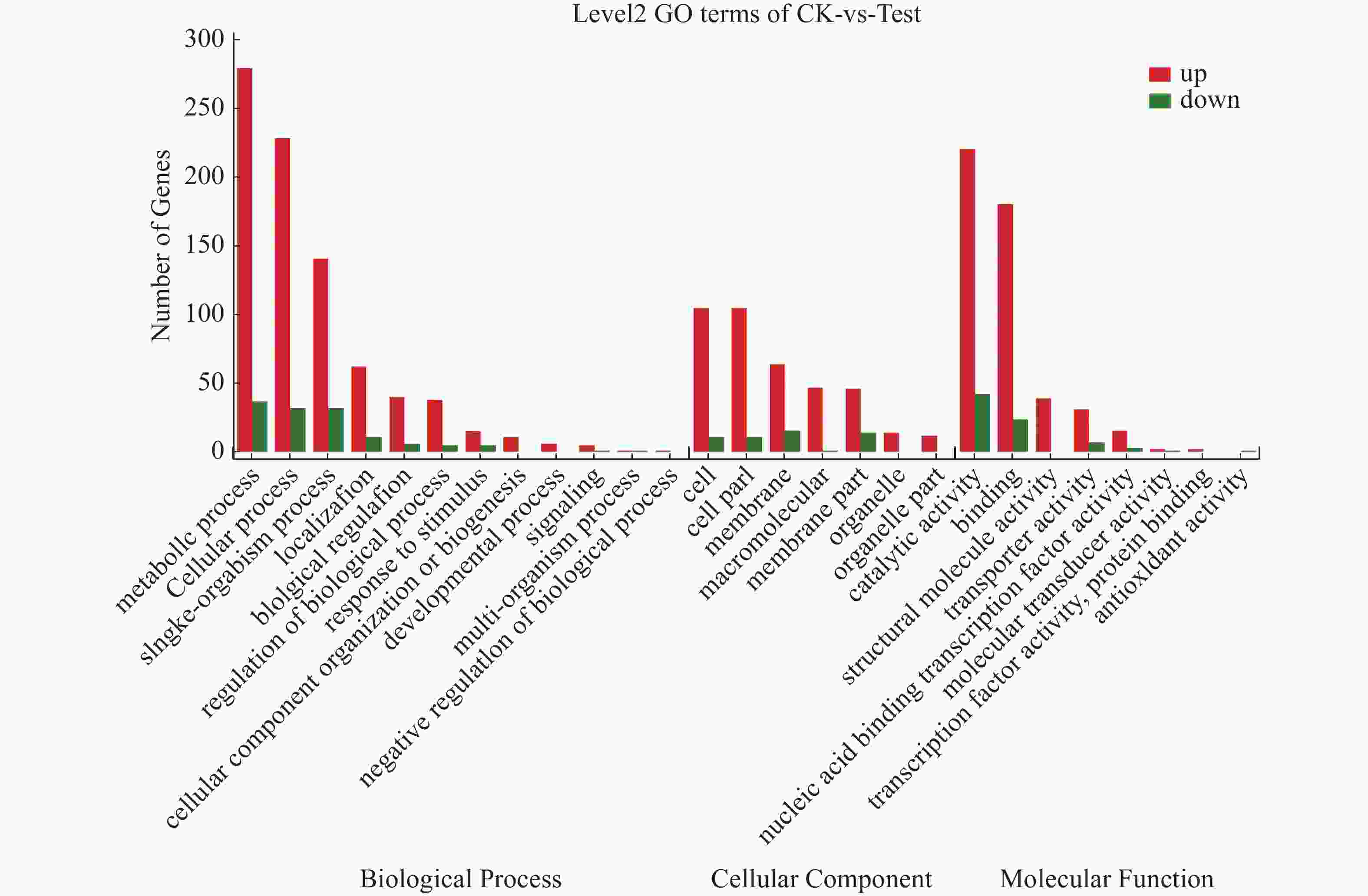
图 6 CK与Test差异基因GO分类
-
将2个样本的差异基因进行KEGG富集分析,结果显示,820个上调表达基因中有289个基因可以富集到KEGG的98条pathways,其中显著富集的pathways有5个。这些显著上调的基因主要与信号转导、跨膜转运和毒力因子表达有关,包括ABC转运、核糖体通路、群体感应、脂肪酸代谢和合成通路(图7-a)。除此之外,还包括少量与叶酸和淀粉蔗糖代谢的相关基因。值得注意的是,这些显著上调的pathways中有许多重复出现的基因(表4),特别是srtF基因(K710_0263)在ABC转运、群体感应系统和双组份调控通路中均上调表达。
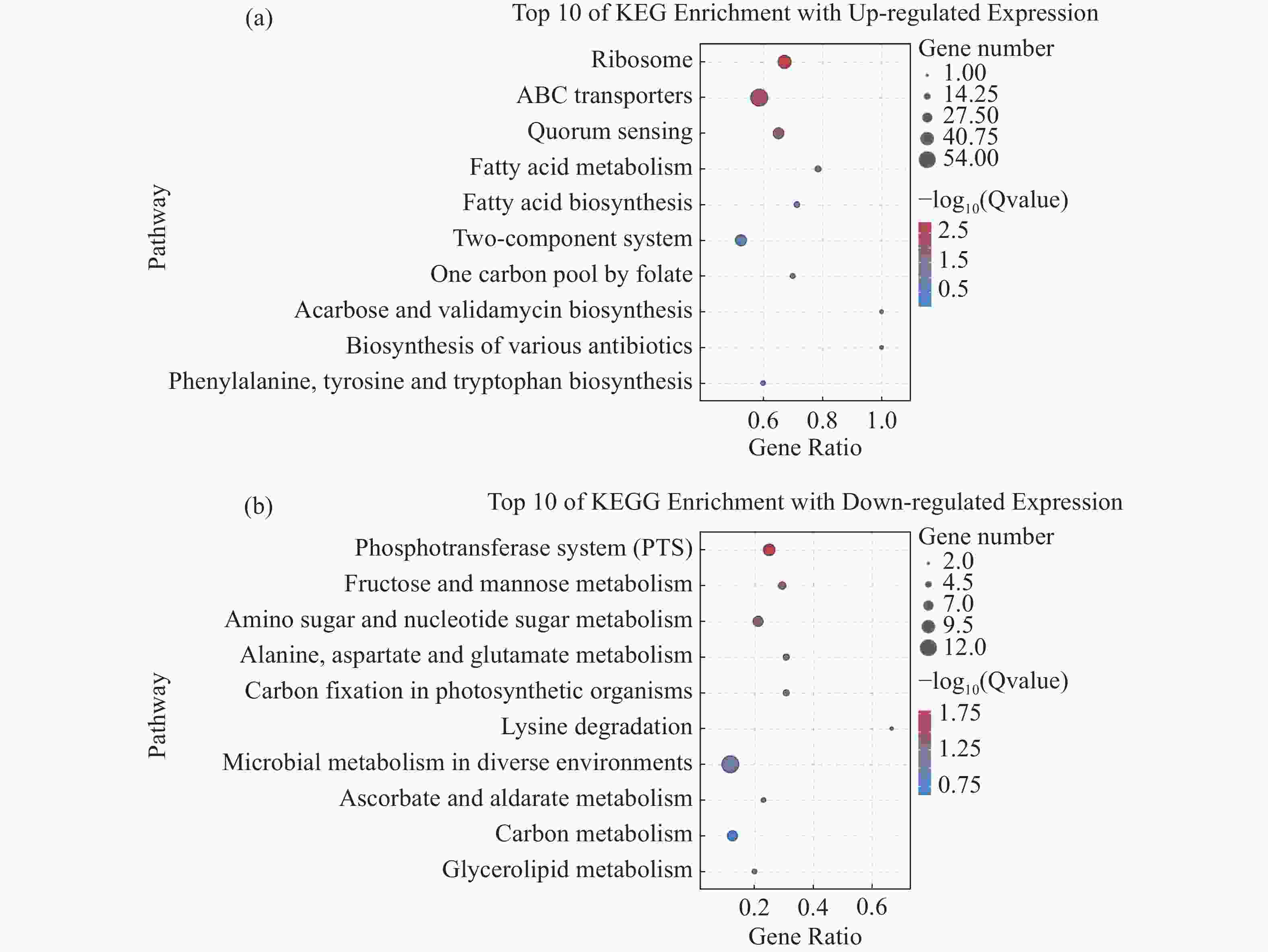
图 7 差异表达基因的KEGG富集分析(35 ℃相对于25 ℃)
表 4 同时参与多条通路的上调表达基因
基因编号 基因 log2FC P 值 信号通路 K710_0263 srtF 3.545569062 4.87E-07 ABC、QS、TCS K710_0976 pstS1 1.033494031 3.50E-21 ABC、TCS K710_1395 pstS2 1.95493017 0.000493456 K710_0264 -- 1.435244618 0.033047506 K710_0434 aliA 1.773511395 4.04E-88 ABC、QS K710_0435 amiC 2.144477475 2.01E-147 K710_0436 amiD 1.999414258 2.79E-108 K710_0437 amiE 2.644802657 2.2E-135 K710_0438 oppF 1.546252336 5.51E-34 K710_1372 oppA 2.841219614 9.87E-36 K710_1541 livF 3.018078137 7.12E-24 K710_1542 livG 2.774570548 3.04E-27 K710_1543 braE 3.172328349 5.42E-46 K710_1544 livH 2.123233661 6.80E-14 K710_1545 braC 1.739006903 4.55E-08 K710_1899 oppF 1.465280432 2.26E-60 K710_1900 oppD 1.390808961 2.33E-58 K710_1901 oppC 1.860690382 5.27E-90 K710_1902 oppB 1.838812786 3.31E-75 K710_0983 ciaR 1.254553 2.19E-23 QS、TCS K710_0984 ciaH 2.680568694 2.45E-103 K710_1962 -- 1.18165149 4.63E-08 K710_1963 -- 1.490948251 1.20E-14 K710_1964 -- 1.597587531 1.1E-20 K710_0266 spaR 1.788729367 7.42E-07 K710_0267 spaK 1.534275087 2.29E-06 K710_0700 lcfB 1.690794566 4.46E-27 FAM、FAB K710_1743 accA 1.562040684 3.24E-57 K710_1744 accD 2.053546645 5.86E-106 K710_1745 accC1 2.160376735 1.45E-128 K710_1746 fabZ 1.80677952 8.39E-86 K710_1747 accB 2.817952264 2.85E-41 K710_1748 fabF 1.760435914 1.04E-111 K710_1749 CLKR27 1.345256763 7.69E-51 K710_1750 fabD 2.368803207 9.24E-116 K710_1751 PA1024 1.624187665 5.32E-40 注:ABC指ABC转运通路;QS指群体感应通路;TCS指双组份系统;FAM指脂肪酸代谢通路;FAB指脂肪酸合成通路; log2FC为相对表达倍数。 35 ℃相对于25 ℃共有45个基因下调表达,总计可以映射到KEGG的38条pathways,按照阈值过滤后筛选到5个显著富集的pathways,这些显著下调的基因几乎都与各种糖类和碳类的获取和代谢相关,包括磷酸转移酶系统、果糖和蔗糖代谢、氨基糖和核糖代谢等通路(图7-b),另外还包括33条不显著富集的pathways。
-
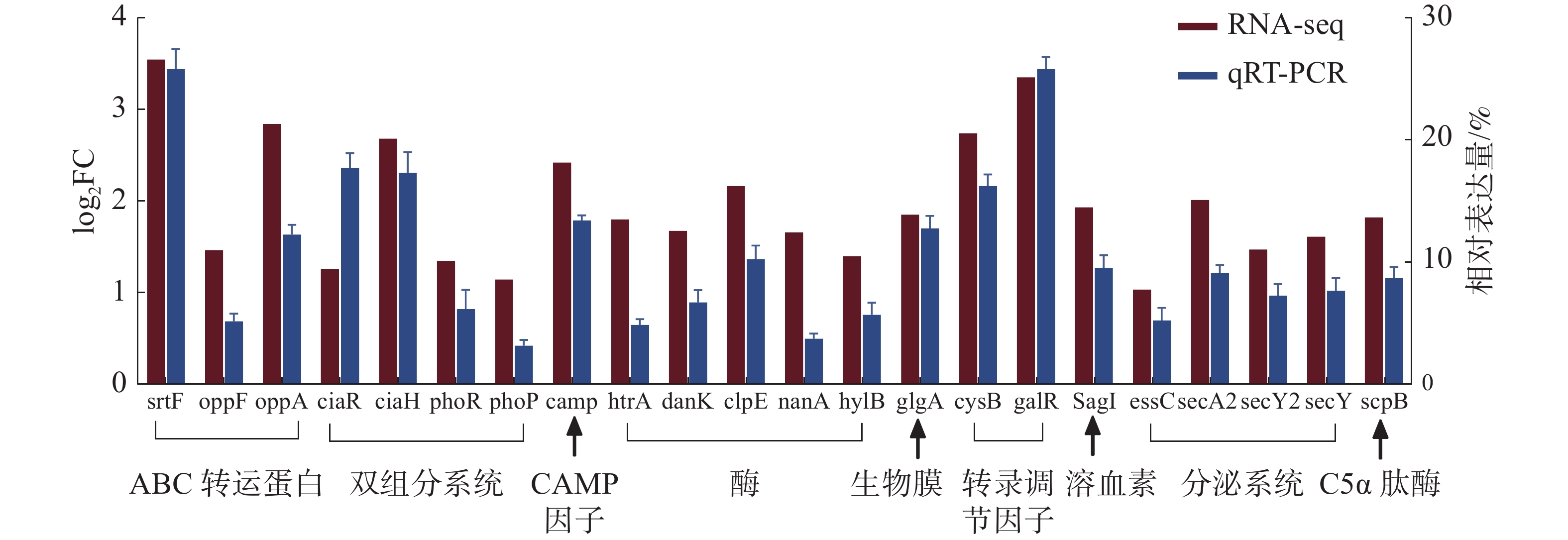
为了评估转录组数据的可靠性,随机筛选22个差异表达的重要毒力基因,采用实时荧光定量PCR的方法检测毒力基因的转录水平,每个基因设置3次生物学重复,以保证数据的准确性,结果如图8所示。2种检测方法经SPSS软件的相关性分析,相关系数达到了0.824,相关极显著(P<0.001),说明qRT-PCR验证结果与RNA-Seq数据具有较好的一致性,并且基因表达变化的趋势基本相同,进一步验证了转录组数据的可靠性。
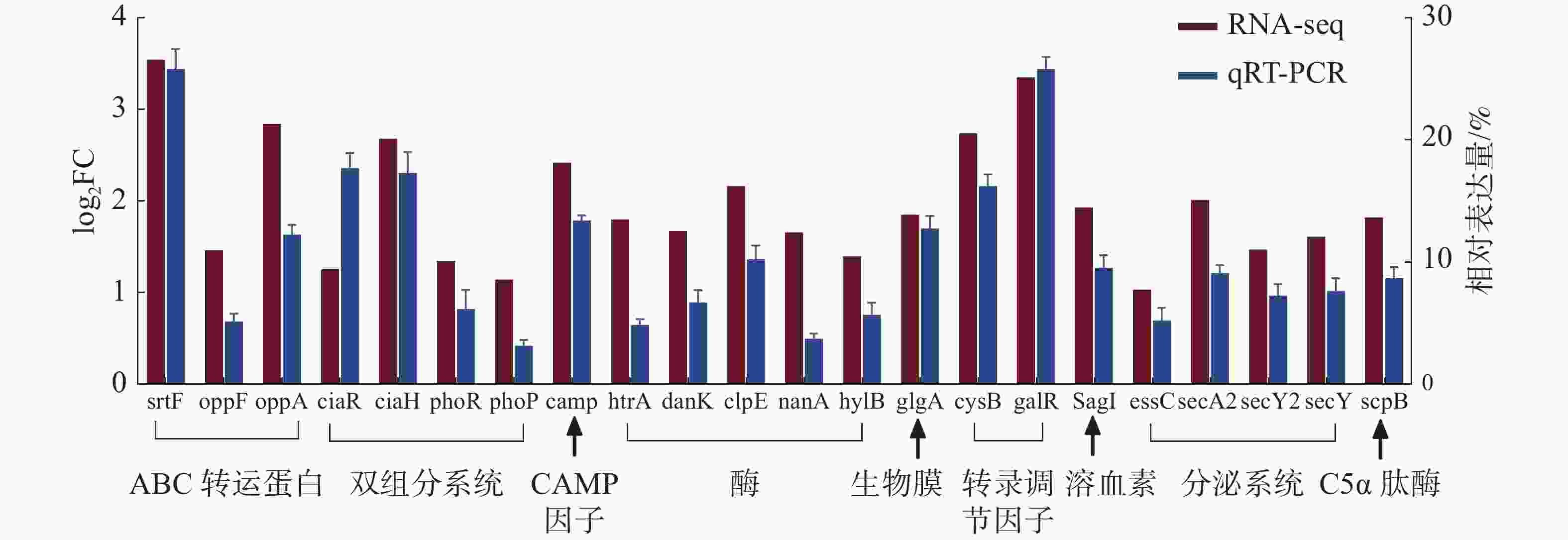
图 8 海豚链球菌在25 ℃和35 ℃条件下毒力相关基因的相对表达量
-
根据文献[27-28]以及VFDB数据库的筛选,发现约58个当前已知或预测的海豚链球菌毒力因子。由表5可知,上调表达的毒力因子有46个,特别是溶血素(SagI)、黏附素(fap1)、转录调节因子(cysB、galR)和C5α肽酶(scpB)等重要的毒力因子,在35 ℃培养条件下毒力因子基因的表达量与25 ℃培养条件下相比明显升高(log2FC>1.6);下调表达的毒力因子基因相对较少,仅有12个(表6),其中包含海豚链球菌毒力标志基因simA。
表 5 上调表达的细菌毒力因子
基因编号 基因 功能注释 log2FC P 值 致病性 K710_0202 adcA zinc ABC transporter solute-binding protein 0.880109591 4.58E-07 黏附 K710_1317 adcA ABC transporter, substrate-binding protein 0.700787773 0.000713 K710_0463 fap1 LPXTG cell wall anchor domain-containing protein 2.813413469 6.74E-158 K710_0464 -- cell wall-anchored protein, partial 3.049435798 7.41E-62 K710_0465 -- liver stage antigen 3 3.180292852 1.65E-112 K710_0466 fap1 accessory Sec-dependent serine-rich glycoprotein adhesin 1.86831671 2.47E-45 K710_0469 -- accessory Sec-dependent serine-rich glycoprotein adhesin 2.140592405 1.16E-84 K710_1067 strA sortase 0.88369926168 1.36E-19 K710_1757 dnak molecular chaperone DnaK 1.67489208905 K710_1013 yplQ hemolysin Ⅲ family protein 1.695832888 1.03E-62 链球菌溶血素S K710_1282 -- SagI 1.930380188 1E-24 K710_1283 -- SagI 1.63917186 5.55E-37 K710_0589 -- ISSag7 transposase 3.804604311 0.000922 K710_1286 -- SagF 1.48053786 1.87E-18 K710_1289 -- streptolysin associated protein SagC 0.646170474 2.38E-05 K710_1290 -- SagB/ThcOx family dehydrogenase 0.13695252 0.3274135 K710_0950 -- CAMP factor family pore-forming toxin 2.420733487 5.35E-78 CAMP
因子K710_1575 scpB segregation/condensation protein B 1.822257336 7.87E-24 C5α肽酶 K710_0833 cpsB cpsB 0.541827357 2.13E-06 荚膜多糖 K710_0834 cpsC cpsC 0.54955598 4.33E-08 K710_0835 cpsD tyrosine-protein kinase 0.65100143348 8.62E-10 K710_1854 essC type Ⅶ secretion protein EssC 1.036654657 4.94E-37 分泌系统 K710_1857 -- type Ⅶ secretion protein EssA 1.792524646 7.30E-06 K710_1858 esaA type Ⅶ secretion protein EsaA 1.555802255 3.85E-51 K710_0099 secY preprotein translocase subunit SecY 1.61255272 1.13E-91 K710_0471 secY2 accessory Sec system protein translocase subunit SecY2 1.470711097 3.92E-15 K710_0475 secA2 SecA2 2.013221387 2.85E-80 K710_0153 -- type Ⅱ secretion system protein 4.212993723 0.0001727 K710_0831 cysB LysR family transcriptional regulator 2.737240401 9.72E-171 转录调节因子 K710_0832 tagU SAK_1262 - regulatory protein CpsX 0.411128745 2.42E-05 K710_0293 galR LacI family DNA-binding transcriptional regulator 3.35070055 4.01E-216 K710_0307 BCE_2019 MarR family transcriptional regulator 1.172169347 1.77E-15 K710_0113 -- TetR family transcriptional regulator 1.396816216 0.0127906 K710_0142 adcR MarR family transcriptional regulator 0.601342176 6.69E-05 K710_0159 MJ0272 helix-turn-helix transcriptional regulator 2.609316032 3.61E-12 K710_0190 yciT DeoR/GlpR transcriptional regulator 2.072378929 3E-96 K710_0216 endR LacI family DNA-binding transcriptional regulator 1.007490952 7.92E-15 K710_0221 yvfU response regulator transcription factor 0.526432544 0.0436715 K710_0223 ydfL MerR family transcriptional regulator 0.879891334 8.93E-06 K710_0252 rgg Rgg/GadR/MutR family transcriptional regulator 0.554807421 0.1430867 K710_0266 spaR response regulator transcription factor 1.788729367 7.42E-07 K710_0307 BCE_2019 MarR family transcriptional regulator 1.172169347 1.77E-15 K710_0983 ciaR response regulator transcription factor 1.254553 2.19E-23 K710_0984 ciaH HAMP domain-containing histidine kinase 2.680568694 2.45E-103 K710_1396 phoR two-component sensor histidine kinase 1.349739342 8.62E-40 K710_1397 phoP response regulator transcription factor 1.144507733 5.47E-17 表 6 下调表达的细菌毒力因子
基因编号 基因 功能注释 log2FC P 值 致病性 K710_0182 -- simA - SiM protein / LPXTG cell wall anchor domain-containing protein −0.886343218 0.0550742 SiM蛋白 K710_0298 -- preprotein translocase subunit YajC −0.527790103 5.45E-07 分泌系统 K710_0255 -- ISSag8, transposase −0.381375332 0.065939 链球菌
溶血素SK710_1553 -- ISSag8, transposase −1.000831387 4.81E-15 K710_1011 -- ISSag8, transposase −1.202452481 4.88E-24 K710_1641 -- ISSag7 transposase −1.993544978 0.138188 K710_0249 hssR response regulator transcription factor −0.092622415 0.551351 转录调节
因子K710_0288 lrp PucR family transcriptional regulator −0.452055279 0.003169 K710_0486 lacR AraC family transcriptional regulator −1.038713833 5.09E-11 K710_0744 purR LacI family transcriptional regulator −0.816241597 1.03E-13 K710_1606 arlR response regulator transcription factor −0.566354947 1.75E-08 K710_1905 glnR MerR family transcriptional regulator −0.402293774 5.01E-06 -
温度是影响细菌毒力的重要环境因子。由病原菌引发的鱼类疾病的暴发往往与水温有着极为紧密的关联性。郭富强等[29]的研究结果表明,随着水温的不断升高罗非鱼感染无乳链球菌的累积死亡率也随之升高,在33 ℃死亡率最高;刘志刚等[17]发现无乳链球菌感染罗非鱼在37 ℃死亡率最高;Zamri-Saad等[30]认为链球菌水温在32~37 ℃时易出现高发病率和高死亡率,而水温26 ℃以下时较少发病。在本研究中,人工回归感染试验结果表明,卵形鲳鲹感染海豚链球菌后在35 ℃下的死亡率高达81.11%,而在25 ℃下的死亡率仅为12.23%,2个温度的PBS对照组均未出现死亡,说明海豚链球菌在35 ℃时毒力更强,导致宿主死亡率更高,发病症状更为明显。同时,本课题组在卵形鲳鲹链球菌病的流行病学调查研究中发现,卵形鲳鲹在25 ℃以下几乎不感染发病,在30~35 ℃时死亡率较高。因此,本研究设置25 ℃培养的海豚链球菌作为对照组,35 ℃培养的海豚链球菌为实验组,探究其在不同温度下基因转录水平上的差异表达,揭示温度对海豚链球菌毒力基因的影响。
在本研究中,当设置FDR<0.05且|log2FC|>1时,与25 ℃培养的海豚链球菌相比,35 ℃培养的海豚链球菌显著上调的基因有820个,而显著下调的基因只有107个。从整体来看,在较高的培养温度下(35 ℃)菌株大多数基因处于上调状态,菌株表现为更加“活跃”的状态,而在较低的温度下(25 ℃)菌株大多数基因则显著下调,菌株显得相对不那么“活跃”,具体表现为与糖类、碳类的代谢相关的基因显著下调,而与细菌毒力相关的基因则显著上调。Mereghetti[31]等在研究无乳链球菌全基因转录组测序时也有类似的发现,编码毒力的相关基因在40 ℃时上调表达,而处于生长稳定期的大量的新陈代谢相关基因在30 ℃时上调表达。
ABC转运蛋白是已知的最大蛋白家族之一,并在细菌中广泛存在,其将ATP水解与各种底物(例如蛋白质、脂质、肽、固醇、糖、离子及药物)的主动运输结合在一起[32-33],这将导致细胞质膜两侧的物质频繁交换。在本研究中,海豚链球菌在35 ℃时ABC转运通路相关的基因显著上调,说明35 ℃海豚链球菌可能涉及各种物质参与的能量交换,以及物质的合成与代谢。群体感应是一种协同调节系统,它以一种被称为AIP(autoinducing peptide)的自诱导肽作为信号分子,通过感知菌群密度和环境因子的变化,并把周围的环境信息传递到双组份系统当中,再由双组份系统对相关基因进行表达调控[34]。一些AIP的前体肽在核糖体中合成,但是AIP无法自由地穿透细胞壁,因此必须通过ABC运输系统或其他的膜通道蛋白进入胞外发挥作用。因此部分上调表达的基因同时参与了ABC转运通路、双组份信号系统通路、群体感应通路、脂肪酸代谢和合成通路当中的2~3个通路,特别是显著上调的srtF(K710_0263)基因同时参与了3个通路的表达调控,具有重要的研究意义。研究发现,srtF属于C类sortase家族,主要作用是在细菌表面形成菌毛并将其固定在细胞壁上,与细胞黏附、定植、生物膜形成等环境过程相互作用[35]。
在下调表达基因中比较典型的是双组份系统中编码柠檬酸合酶(Citrate lyase)相关的基因(K710_0369、K710_0374、K710_0375、K710_0379和K710_1345),由此表明,在35 ℃时,Tozj-1菌株的三羧酸循环被抑制,对葡萄糖的代谢能力明显降低,而在25 ℃相对较低的环境温度中,其对葡萄糖的代谢能力表现良好。这与胡文婷[16]研究的无乳链球菌和刘韬[36]研究的鲁氏耶尔森菌结论一致,低温更有利于病原菌葡萄糖的代谢。并且在35 ℃时,与磷酸转移酶系统、果糖和蔗糖代谢、氨基糖和核苷酸糖代谢相关的基因显著下调,这证明糖类、碳类的代谢更适合在较低温度下进行。
病原菌的致病过程与毒力因子的共同介导作用密切相关。海豚链球菌几种重要的毒力因子包括M蛋白、磷酸葡萄糖苷酶(pgmA)、溶血素S、C5α肽酶、脱乙酰基酶、CAMP因子。M蛋白[37]是一种可以通过荚膜延伸到菌体表面形成菌毛的蛋白,它在细菌粘附、感染、抵御吞噬等方面起关键作用;磷酸葡萄糖苷酶(pgmA)[38]促进细胞壁和荚膜的生物合成,并能抵抗正电性抗菌肽;溶血素S[39-40]能破坏宿主红细胞、单核细胞和粒细胞,有可能对脑血管造成损伤;C5α肽酶能水解中性粒细胞化学诱导物补体因子C5α,从而破坏宿主细胞的抗侵袭能力;脱乙酰基酶[41]能够提高病原菌对溶菌酶的耐受性,逃避宿主免疫,黏附并入侵宿主的上皮细胞;CAMP因子[42]通过与FC区域的免疫球蛋白结合,从而有助于拦截抗体向补体呈递抗原。这些毒力因子相互作用协助S.iniae侵入宿主,逃避宿主的免疫防御。在本研究中,不同温度下培养的海豚链球菌,其毒力因子呈差异表达,在35 ℃时,除了溶血素、黏附素、转录调节因子等毒力因子转录上调之外,抗逆因子和一些酶类基因的转录表达量也大幅上升,只有少量的毒力因子下调表达。刘志刚等[17]、王松艳[21]和郭富强[29]对无乳链球菌的研究亦发现类似的结果。以上相关研究结果均表明,高温有助于海豚链球菌毒力相关基因表达,导致其毒力增强,有助于其黏附、定植、入侵、胞内存活和扩散等致病过程,引起宿主发病。
本研究结果初步证实温度能够广泛影响海豚链球菌基因的转录水平,进而调控海豚链球菌的致病性,为进一步研究海豚链球菌的致病机理提供理论依据,但其调控机制仍需要进一步研究加以阐明。
Transcriptome analysis of Streptococcus iniae from fish at different culture temperatures
-
摘要: 为鉴定海豚链球菌(Streptococcus iniae)在不同温度的转录水平差异,通过生长曲线测定和人工感染试验分析该菌在25 ℃和35 ℃下的生长情况和致病性,采用链特异性转录组测序(Strand-specific RNA-seq)技术对25 ℃和35 ℃培养的海豚链球菌Tozj-1菌株进行测序分析,筛选差异表达基因,通过GO(Gene Ontology)数据库和KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库对差异表达基因进行GO功能和KEGG通路富集分析,然后利用VFDB(Virulence factor database)数据库筛选差异表达的重要毒力基因并使用实时荧光定量PCR进行验证。结果显示,海豚链球菌在35 ℃条件下有更快的生长速度和更强的毒力;转录组共筛选获得927个显著差异表达基因(P<0.05),其中包括820个上调基因和107个下调基因;GO功能富集分析发现,差异基因主要富集在代谢、细胞、催化及结合过程;KEGG富集分析发现,差异基因主要富集在核糖体通路、ABC转运通路、群体感应等信号通路。以上结果表明温度调控海豚链球菌基因的转录表达及相关通路的富集,为后续海豚链球菌致病机理的研究提供数据支持。Abstract: To identify the differences in transcription level of Streptococcus iniae at different temperatures, the growth and pathogenicity of S. iniae cultured at 25 ℃ and 35 ℃ were analyzed by using growth curve and artificial infection test, and strand-specific RNA-seq technology was used for sequencing analysis of S. iniae Tozj-1 strain cultured at 25 ℃ and 35 ℃. The differentially expressed genes (DEGs) were screened and their GO function and KEGG pathway enrichment were analyzed based on the GO (Gene Ontology) database and KEGG (Kyoto Encyclopedia of Genes and Genomes) database. The key differentially expressed virulence genes were filtrated using virulence factor database (VFDB) and verified by real-time quantitative PCR. The results showed that S. iniae grew faster with a higher virulence at 35 ℃. A total of 927 significantly differentially expressed genes were screened (P<0.05), including 820 up-regulated genes and 107 down-regulated genes. GO functional enrichment analysis showed that the DEGs were mainly enriched in metabolic process, cellular process, binding process and catalytic activity. KEGG enrichment analysis revealed that the DEGs were mainly enriched in ribosome, quorum sensing, ABC transporters and other signaling pathways. These results indicated that temperature regulated the transcription and expression of S .iniae genes and the enrichment of related pathways, which provided data support for further research in the pathogenesis of S .iniae.
-
Key words:
- Streptococcus iniae /
- transcriptome sequencing /
- temperature /
- virulence factor /
- qRT-PCR
-
表 1 检测海豚链球菌毒力基因的qRT-PCR特异性引物
功能分类 引物名称 上游引物序列(5′→3′) 下游引物序列(5′→3′) 内参基因 16S rRNA CCAAGGCGACGATACATAG TTGCCGAAGATTCCCTACT ABC转运蛋白 srtF GGAGCAGGAAAGTCAACG TGGATAGATCGCAGGATT oppF AAACCACTGTTGGTCGTGC TCTTTGCGGAAATCTCGTA oppA GGAAGTAATCTATTGCGTGTC TACTGCCATCAGACCATTT 双组份系统 ciaR AATGCGTATTCAAGCCCTCT AATGGTAGTGTCTCCGACT ciaH TGCTTTGAAATACACGGACTC AACACGATAGAAGCGGTCA phoR AGGCTAAACAACTCCACTT TCATCGGTGTACCTAACAG phoP ACAGCAACAGATGGCAGAT AGACCTCAATACCGTCCAA 溶血素 sagI ATGTCATTGTTATGCCAACT GTAAGGGCCTGCACTCAA CAMP因子 camp TGCCTCAACATCAAACAG ATGGCTTGAACAGCATCA 酶 htrA TCAGGTTCAAATGGAGGAG AAGGCAGGACGAATCACT danK CAGTTCGTCGTGCCCTTTC TTGGCTCTTTACCAGTTTCAG clpE GTCCCTCACAAACTCCAGC CTCTTCAAATTGACCACGA nanA GCCGTAACCGCTCACCACT TTTCAATCCCGTCAAGTCC hylB AACCCTGCTTCAACACTCT CCAATCTCATAATCCCACC C5α肽酶 scpB GGGTTGAGTGTCCGTAATT CTGACGCATCATCATCTGC 分泌系统 essC GCCGTCAACGCCTCTTCTC GGCATTGGCTCTGTCACTT secA2 AGCTGGAATTGAATACGC AGATAATGAGCCCGAAGG secY2 GCATTAAACCAGGACAAGC GATATTACCGCCACCAGAA secY ATAGCAGCTCAACAAACCC ATGCCTTCAATACCAGTCG 转录调节因子 cysB CGAAATCCTAAGGGCATCA CGTTCCTCCAACAAAGAG galR GAGGCTGGTATCACTGTTC GCAGTAGTTCCCATTTCCT 生物膜 glgA TAGCGTTTGCTCTTGATGT TAAGCCTGACTTGCCGTAT  下载: 导出CSV
下载: 导出CSV
表 2 过滤前后碱基信息统计
样品 过滤前 过滤后 Raw Data/bp Q20/% Q30/% GC含量/% Raw Data/bp Q20/% Q30/% GC含量/% CK-1 3283961700 98.06 93.98 40.78 3202330021 98.11 94.08 40.49 CK-2 2481964500 98.20 94.34 41.20 2397726314 98.27 94.47 40.83 CK-3 2190706200 98.03 93.95 41.22 2118495261 98.11 94.09 40.85 Test-1 2753256900 98.03 93.97 40.76 2655059882 98.11 94.12 40.35 Test-2 2777076900 97.89 93.63 40.60 2677608106 97.98 93.80 40.19 Test-3 2516073000 97.77 93.33 41.10 2415735833 97.88 93.53 40.66
下载: 导出CSV
表 3 原始数据比对到基因组
样品 总reads数 匹配上的reads数 匹配比例/% CK-1 18162776 17938781 98.77 CK-2 13888834 13717694 98.77 CK-3 12276544 12129610 98.80 Test-1 16739444 16498781 98.56 Test-2 17315536 17063899 98.55 Test-3 14858622 14630754 98.47
下载: 导出CSV
表 4 同时参与多条通路的上调表达基因
基因编号 基因 log2FC P 值 信号通路 K710_0263 srtF 3.545569062 4.87E-07 ABC、QS、TCS K710_0976 pstS1 1.033494031 3.50E-21 ABC、TCS K710_1395 pstS2 1.95493017 0.000493456 K710_0264 -- 1.435244618 0.033047506 K710_0434 aliA 1.773511395 4.04E-88 ABC、QS K710_0435 amiC 2.144477475 2.01E-147 K710_0436 amiD 1.999414258 2.79E-108 K710_0437 amiE 2.644802657 2.2E-135 K710_0438 oppF 1.546252336 5.51E-34 K710_1372 oppA 2.841219614 9.87E-36 K710_1541 livF 3.018078137 7.12E-24 K710_1542 livG 2.774570548 3.04E-27 K710_1543 braE 3.172328349 5.42E-46 K710_1544 livH 2.123233661 6.80E-14 K710_1545 braC 1.739006903 4.55E-08 K710_1899 oppF 1.465280432 2.26E-60 K710_1900 oppD 1.390808961 2.33E-58 K710_1901 oppC 1.860690382 5.27E-90 K710_1902 oppB 1.838812786 3.31E-75 K710_0983 ciaR 1.254553 2.19E-23 QS、TCS K710_0984 ciaH 2.680568694 2.45E-103 K710_1962 -- 1.18165149 4.63E-08 K710_1963 -- 1.490948251 1.20E-14 K710_1964 -- 1.597587531 1.1E-20 K710_0266 spaR 1.788729367 7.42E-07 K710_0267 spaK 1.534275087 2.29E-06 K710_0700 lcfB 1.690794566 4.46E-27 FAM、FAB K710_1743 accA 1.562040684 3.24E-57 K710_1744 accD 2.053546645 5.86E-106 K710_1745 accC1 2.160376735 1.45E-128 K710_1746 fabZ 1.80677952 8.39E-86 K710_1747 accB 2.817952264 2.85E-41 K710_1748 fabF 1.760435914 1.04E-111 K710_1749 CLKR27 1.345256763 7.69E-51 K710_1750 fabD 2.368803207 9.24E-116 K710_1751 PA1024 1.624187665 5.32E-40 注:ABC指ABC转运通路;QS指群体感应通路;TCS指双组份系统;FAM指脂肪酸代谢通路;FAB指脂肪酸合成通路; log2FC为相对表达倍数。
下载: 导出CSV
表 5 上调表达的细菌毒力因子
基因编号 基因 功能注释 log2FC P 值 致病性 K710_0202 adcA zinc ABC transporter solute-binding protein 0.880109591 4.58E-07 黏附 K710_1317 adcA ABC transporter, substrate-binding protein 0.700787773 0.000713 K710_0463 fap1 LPXTG cell wall anchor domain-containing protein 2.813413469 6.74E-158 K710_0464 -- cell wall-anchored protein, partial 3.049435798 7.41E-62 K710_0465 -- liver stage antigen 3 3.180292852 1.65E-112 K710_0466 fap1 accessory Sec-dependent serine-rich glycoprotein adhesin 1.86831671 2.47E-45 K710_0469 -- accessory Sec-dependent serine-rich glycoprotein adhesin 2.140592405 1.16E-84 K710_1067 strA sortase 0.88369926168 1.36E-19 K710_1757 dnak molecular chaperone DnaK 1.67489208905 K710_1013 yplQ hemolysin Ⅲ family protein 1.695832888 1.03E-62 链球菌溶血素S K710_1282 -- SagI 1.930380188 1E-24 K710_1283 -- SagI 1.63917186 5.55E-37 K710_0589 -- ISSag7 transposase 3.804604311 0.000922 K710_1286 -- SagF 1.48053786 1.87E-18 K710_1289 -- streptolysin associated protein SagC 0.646170474 2.38E-05 K710_1290 -- SagB/ThcOx family dehydrogenase 0.13695252 0.3274135 K710_0950 -- CAMP factor family pore-forming toxin 2.420733487 5.35E-78 CAMP
因子K710_1575 scpB segregation/condensation protein B 1.822257336 7.87E-24 C5α肽酶 K710_0833 cpsB cpsB 0.541827357 2.13E-06 荚膜多糖 K710_0834 cpsC cpsC 0.54955598 4.33E-08 K710_0835 cpsD tyrosine-protein kinase 0.65100143348 8.62E-10 K710_1854 essC type Ⅶ secretion protein EssC 1.036654657 4.94E-37 分泌系统 K710_1857 -- type Ⅶ secretion protein EssA 1.792524646 7.30E-06 K710_1858 esaA type Ⅶ secretion protein EsaA 1.555802255 3.85E-51 K710_0099 secY preprotein translocase subunit SecY 1.61255272 1.13E-91 K710_0471 secY2 accessory Sec system protein translocase subunit SecY2 1.470711097 3.92E-15 K710_0475 secA2 SecA2 2.013221387 2.85E-80 K710_0153 -- type Ⅱ secretion system protein 4.212993723 0.0001727 K710_0831 cysB LysR family transcriptional regulator 2.737240401 9.72E-171 转录调节因子 K710_0832 tagU SAK_1262 - regulatory protein CpsX 0.411128745 2.42E-05 K710_0293 galR LacI family DNA-binding transcriptional regulator 3.35070055 4.01E-216 K710_0307 BCE_2019 MarR family transcriptional regulator 1.172169347 1.77E-15 K710_0113 -- TetR family transcriptional regulator 1.396816216 0.0127906 K710_0142 adcR MarR family transcriptional regulator 0.601342176 6.69E-05 K710_0159 MJ0272 helix-turn-helix transcriptional regulator 2.609316032 3.61E-12 K710_0190 yciT DeoR/GlpR transcriptional regulator 2.072378929 3E-96 K710_0216 endR LacI family DNA-binding transcriptional regulator 1.007490952 7.92E-15 K710_0221 yvfU response regulator transcription factor 0.526432544 0.0436715 K710_0223 ydfL MerR family transcriptional regulator 0.879891334 8.93E-06 K710_0252 rgg Rgg/GadR/MutR family transcriptional regulator 0.554807421 0.1430867 K710_0266 spaR response regulator transcription factor 1.788729367 7.42E-07 K710_0307 BCE_2019 MarR family transcriptional regulator 1.172169347 1.77E-15 K710_0983 ciaR response regulator transcription factor 1.254553 2.19E-23 K710_0984 ciaH HAMP domain-containing histidine kinase 2.680568694 2.45E-103 K710_1396 phoR two-component sensor histidine kinase 1.349739342 8.62E-40 K710_1397 phoP response regulator transcription factor 1.144507733 5.47E-17
下载: 导出CSV
表 6 下调表达的细菌毒力因子
基因编号 基因 功能注释 log2FC P 值 致病性 K710_0182 -- simA - SiM protein / LPXTG cell wall anchor domain-containing protein −0.886343218 0.0550742 SiM蛋白 K710_0298 -- preprotein translocase subunit YajC −0.527790103 5.45E-07 分泌系统 K710_0255 -- ISSag8, transposase −0.381375332 0.065939 链球菌
溶血素SK710_1553 -- ISSag8, transposase −1.000831387 4.81E-15 K710_1011 -- ISSag8, transposase −1.202452481 4.88E-24 K710_1641 -- ISSag7 transposase −1.993544978 0.138188 K710_0249 hssR response regulator transcription factor −0.092622415 0.551351 转录调节
因子K710_0288 lrp PucR family transcriptional regulator −0.452055279 0.003169 K710_0486 lacR AraC family transcriptional regulator −1.038713833 5.09E-11 K710_0744 purR LacI family transcriptional regulator −0.816241597 1.03E-13 K710_1606 arlR response regulator transcription factor −0.566354947 1.75E-08 K710_1905 glnR MerR family transcriptional regulator −0.402293774 5.01E-06
下载: 导出CSV
-
[1] PIER G B, MADIN S H. Streptococcus iniae sp. nov. a beta-hemolytic streptococcus isolated from an Amazon freshwater dolphin, Inia geoffrensis[J]. International Journal of Systematic Bacteriology, 1976, 26(4): 545 − 553. doi: 10.1099/00207713-26-4-545 [2] YOUNG E J, BANNISTER J, BULLER N B, et al. Streptococcus iniae associated mass marine fish kill off Western Australia[J]. Diseases of Aquatic Organisms, 2020, 142: 197 − 201. doi: 10.3354/dao03545 [3] XIONG X, PENG Y, CHEN R, et al. Efficacy and transcriptome analysis of golden pompano (Trachinotus ovatus) immunized with a formalin-inactived vaccine against Streptococcus iniae[J]. Fish & Shellfish Immunology, 2023, 134: 108489. [4] 武沛文, 宋大伟, 郭丹玲, 等. 湛江卵形鲳鲹致病链球菌的分离、鉴定及药敏实验[J]. 基因组学与应用生物学, 2019, 38(12): 5403 − 5411. [5] HECKMAN T I, SHAHIN K, HENDERSON E E, et al. Development and efficacy of Streptococcus iniae live-attenuated vaccines in Nile Tilapia, Oreochromis niloticus[J]. Fish & Shellfish Immunology, 2022, 121: 152 − 162. [6] AL-HARBI A H. Whole-genome sequence of Streptococcus iniae strain AH1, isolated from hybrid tilapia (Oreochromis niloticus × Oreochromis aureus)[J]. Microbiology Resource Announcements, 2022, 11(6): e0008722. doi: 10.1128/mra.00087-22 [7] FENG Y, BAI M, GENG Y, et al. The potential risk of antibiotic resistance of Streptococcus iniae in sturgeon cultivation in Sichuan, China[J]. Environmental Science and Pollution Research, 2021, 28(48): 171 − 180. [8] MUGETTI D, COLUSSI S, PASTORINO P, et al. Episode of mortality associated with isolation of Streptococcus iniae in Adriatic sturgeon (Acipenser naccarii Bonaparte, 1836) reared in Northern Italy[J]. Journal of Fish Diseases, 2022, 45(6): 939 − 942. doi: 10.1111/jfd.13608 [9] 杨林狄, 贾新蕾, 黄增朝, 等. 银鼓鱼海豚链球菌的分离、鉴定及毒力基因检测[J]. 大连海洋大学学报, 2023, 38(2): 1 − 9. [10] 徐伟, 施慧, 汪玮, 等. 小黄鱼海豚链球菌的分离鉴定[J]. 中国预防兽医学报, 2022, 44(7): 725 − 730. [11] 王汉清, 黄郁葱, 林潮峰, 等. 黄鳍鲷海豚链球菌的分离·鉴定及药敏试验[J]. 安徽农业科学, 2018, 46(21): 100 − 102. doi: 10.3969/j.issn.0517-6611.2018.21.028 [12] ZHOU K, CUI T T, LI P L, et al. Modelling and predicting the effect of temperature, water activity and pH on growth of Streptococcus iniaein Tilapia[J]. Journal of Applied Microbiology, 2008, 105(6): 1956 − 1965. doi: 10.1111/j.1365-2672.2008.03969.x [13] MAULU S, HASIMUNA O J, MPHANDE J, et al. Prevention and control of streptococcosis in Tilapia culture: a systematic review[J]. Journal of Aquatic Animal Health, 2021, 33(3): 162 − 177. doi: 10.1002/aah.10132 [14] 祝璟琳, 杨弘, 邹芝英, 等. 海南养殖罗非鱼(Oreochromis niloticus)致病链球菌的分离、鉴定及其药敏试验[J]. 海洋与湖沼, 2010, 41(4): 590 − 596. [15] 陈家长, 臧学磊, 瞿建宏, 等. 温度胁迫下罗非鱼(GIFT Oreochromis niloticus)机体免疫力的变化及其对海豚链球菌易感性的影响[J]. 农业环境科学学报, 2011, 30(9): 1896 − 1901. [16] 胡文婷. 基于代谢组学的温度对罗非鱼链球菌病的影响和调控方法研究[D]. 海口: 海南大学, 2017. [17] 刘志刚, 可小丽, 卢迈新, 等. 温度对尼罗罗非鱼无乳链球菌毒力的影响[J]. 水产学报, 2013, 37(11): 1733 − 1741. [18] 刘海波, 绳秀珍, 唐小千, 等. 培养条件对海豚链球菌的生长及其胞外产物蛋白组成的影响[J]. 中国动物检疫, 2016, 33(5): 85 − 89. [19] 柒壮林, 祝璟琳, 杨弘, 等. 不同温度下罗非鱼人工感染海豚链球菌后血液生化指标及组织病理[J]. 广东海洋大学学报, 2012, 32(4): 34 − 41. [20] LI L, LU J, ZHAN P, et al. RNA-seq analysis unveils temperature and nutrient adaptation mechanisms relevant for pathogenicity in Vibrio parahaemolyticus[J]. Aquaculture, 2022, 558: 738397. doi: 10.1016/j.aquaculture.2022.738397 [21] 王松艳. 鱼源无乳链球菌转录组分析及其毒力相关基因筛选和sRNA预测[D]. 湛江: 广东海洋大学, 2016. [22] 师若萍. 大肠杆菌NCM3722在不同温度中的转录组和代谢组的研究[D]. 深圳: 深圳大学, 2019. [23] CHEN S, ZHOU Y, CHEN Y, et al. Fastp: an ultra-fast all-in-one FASTQ preprocessor[J]. Bioinformatics, 2018, 34(17): i884 − i890. doi: 10.1093/bioinformatics/bty560 [24] LANGMEAD B, SALZBERG S L. Fast gapped-read alignment with bowtie 2[J]. Nature Methods, 2012, 9(4): 357 − 379. doi: 10.1038/nmeth.1923 [25] GOLKAR-NARENJI A, ANTOSIK P, NOLIN S, et al. Gene ontology groups and signaling pathways regulating the process of avian satellite cell differentiation[J]. Genes, 2022, 13(2): 242. doi: 10.3390/genes13020242 [26] KANEHISA M, FURUMICHI M, TANABE M, et al. KEGG: new perspectives on genomes, pathways, diseases and drugs[J]. Nucleic Acids Research, 2017, 45(D1): D353 − D361. doi: 10.1093/nar/gkw1092 [27] COLUSSI S, PASTORINO P, MUGETTI D, et al. Isolation and genetic characterization of Streptococcus iniae virulence factors in adriatic sturgeon (Acipenser naccarii)[J]. Microorganisms, 2022, 10(5): 883. doi: 10.3390/microorganisms10050883 [28] BAIANO J C F, BARNES A C. Towards control of Streptococcus iniae[J]. Emerging Infectious Diseases, 2009, 15(12): 1891 − 1896. doi: 10.3201/eid1512.090232 [29] 郭富强. 温度对罗非鱼无乳链球菌致病机制影响的研究[D]. 上海: 上海海洋大学, 2018. [30] ZAMRI-SAAD M, AMAL M N A, SITI-ZAHRAH A. Pathological changes in red tilapias (Oreochromis spp.) naturally infected by Streptococcus agalactiae[J]. Journal of Comparative Pathology, 2010, 143(2/3): 227 − 229. [31] MEREGHETTI L, SITKIEWICZ I, GREEN N M, et al. Remodeling of the Streptococcus agalactiae transcriptome in response to growth temperature[J]. PLoS One, 2008, 3(7): e2785. doi: 10.1371/journal.pone.0002785 [32] BENDA M, SCHULZ L M, STüLKE J, et al. Influence of the ABC transporter YtrBCDEF of Bacillus subtilis on competence, biofilm formation and cell wall thickness[J]. Frontiers in Microbiology, 2021, 12: 587035. doi: 10.3389/fmicb.2021.587035 [33] MALLICK S, KUMAR A, DODIA H, et al. Biochemical characterization of anE. colicell division factor FtsE shows ATPase cycles similar to the NBDs of ABC-transporters[J]. Bioscience Reports, 2021, 41(1): BSR20203034. doi: 10.1042/BSR20203034 [34] 饶贤才, 胡福泉. 分子微生物学前沿[J]. 农药学学报, 2013, 15(4): 444. [35] FAULDS-PAIN A, SHAW H A, TERRA V S, et al. The Streptococcos suis sortases SrtB and SrtF are essential for disease in pigs[J]. Microbiology, 2019, 165(2): 163 − 173. doi: 10.1099/mic.0.000752 [36] 刘韬, 魏文燕, 刘家星, 等. 不同温度条件下鲁氏耶尔森氏菌的链特异性转录组分析[J]. 水生生物学报, 2019, 43(5): 969 − 976. [37] KIM D, BECK B R, LEE S M, et al. Pellet feed adsorbed with the recombinant Lactococcus lactis BFE920 expressing SiMA antigen induced strong recall vaccine effects against Streptococcus iniae infection in olive flounder (Paralichthys olivaceus)[J]. Fish & Shellfish Immunology, 2016, 55: 374 − 383. [38] 马丽, 吴金英, 高凇泽, 等. 海豚链球菌simA和pgmA真核表达质粒对尼罗罗非鱼免疫保护的研究[J]. 南方水产科学, 2020, 16(3): 38 − 46. [39] ETHICA S N, DARMAWATI S, DEWI S S, et al. Streptolysin encoding genes sagC and sagD as biomarkers of fish pathogen Streptococcus iniae: an in silico study[J]. Squalen Bulletin of Marine and Fisheries Postharvest and Biotechnology, 2020, 15(1): 21 − 39. [40] LIU C, HU X, CAO Z, et al. Construction and characterization of a DNA vaccine encoding the SagH against Streptococcus iniae[J]. Fish & Shellfish Immunology, 2019, 89: 71 − 75. [41] 白明焕, 耿毅, 赵若璇, 等. 四川鲟源海豚链球菌的毒力基因谱及分子分型[J]. 华南农业大学学报, 2020, 41(5): 36 − 42. [42] SHENG X, GAO J, LIU H, et al. Recombinant phosphoglucomutase and CAMP factor as potential subunit vaccine antigens induced high protection against Streptococcus iniae infection in flounder (Paralichthys olivaceus)[J]. Journal of Applied Microbiology, 2018, 125(4): 997 − 1007. doi: 10.1111/jam.13948 -

 点击查看大图
点击查看大图

图(8) / 表 (6)
计量
- 文章访问数: 2440
- HTML全文浏览量: 868
- PDF下载量: 59
- 被引次数: 0
