



-
兰科(Orchidaceae)植物类群庞大,多具有观赏、药用、食用等经济价值。由于其对生境的独特需求,多以小种群形式分布,较其他植物更容易受到生境退化或丧失的威胁[1]。目前,世界所有兰科植物已被列入《濒危野生动植物物种国际贸易公约》加以保护,成为植物保护中的1个“旗舰”类群[2]。在国家林业和草原局、农业农村部于2021年9月最新公布的《国家重点保护野生植物名录》中,野生兰科植物共涉及到23属349种,野生兰科植物的保护现已受到空前重视。
附生型兰科植物(Epiphytic Orchids)(以下简称附生兰)为没有地下生长阶段,通过气生根附着于岩石或树干上的兰科植物。因为生长的依附性,附生兰科植物对附生环境和基质要求更为严格,对环境变化更为敏感,更容易形成斑块状分布[3]。同时,附生兰生长缓慢,通常1个生长轴1年仅发1个新芽,从萌发到成熟需要超过1年的时间,当生境和种群遭到破坏后,恢复极为缓慢[4]。附生兰约占全部附生植物种数的10%~25% [5-6],而附生植物是热带、亚热带湿性山地森林和少部分温带雨林的重要组成部分,对维持森林生态系统生物多样性、水分平衡、养分循环、为动物提供食物和庇护场所等方面的贡献不可忽略[7]。因此,研究附生兰的分布特征可对附生兰科植物的生境保护和植株繁育提供理论依据,进而对生态系统的生物多样性和功能的维持意义重大。近年来,对于附生兰分布的研究多集中于地区附生兰物种多样性与组成[8-13],或侧重个别环境因子如海拔[14-16]、温度[9, 16-17]、降水[9, 16-17]、植被类型[17-20]等对附生兰科植物分布的影响,或少数附生兰如华石斛(Dendrobium sinense)等的适宜生境模拟,而鲜有综合研究附生兰分布的多因素作用的报道[17, 21],这对于附生兰生境保护与多样性维持是不利的。与地生植物不同,附生兰远离土壤基质,更多地受到附生基质与气候因子的影响,因而附生兰的影响因子分析中应将诸多的气候因子与附生基质(树皮)特征纳入其中。相关的少许研究仅局限于年平均气温与年降雨量的变化上,时间尺度上过于宽泛,不利于解释附生兰生境的气候差异[13, 17],附生基质对附生兰分布影响的研究报道同样少见[20]。附生兰在世界范围内大多面临着各种人为因素的生存威胁。要制定地方乃至全国的附生兰科植物保育及资源合理开发利用策略,通过系统、综合的附生植物与环境因子的关联分析研究来建立理论指导是亟待解决的问题。本研究拟结合附生兰分布状况与分布点上对附生兰生存起重要作用的各类生态因子如海拔、地形、植被类型、气候等,综合分析鹦哥岭附生兰科植物的生境适应规律,旨在为海南热带雨林国家公园内附生兰的保育、适宜生境选择、气候变化背景下的种群数量预测及未来的资源开发利用提供理论依据。
-
鹦哥岭自然保护区,现为海南热带雨林国家公园管理局鹦哥岭分局所辖的自然保护地,位于海南岛中南部,地理坐标为109° 11′ 27″~109° 34′ 06″ E,18° 49′ 30″~19° 08′ 41″ N,总面积50 464 hm2。地形以山地为主,保护区内最高峰为鹦哥岭,海拔1 812 m,是海南第2高峰。保护区属热带海洋性季风气候,年均气温20~24 ℃,在相同高度上,年均气温由东北向西南递增。年平均降雨量1 400~2 200 mm,但季节分配不均,东南坡降雨明显高于西北坡。依据杨小波等[22]的《海南植被志》,研究区域林区的植被可划分为热带针叶林、高山云雾林、山地雨林、低地雨林、低地雨林次生林、季雨林、旱地农业生产植被等植被型或植被亚型。
卢刚等[23]通过多年的考察,出版了《海南鹦哥岭兰科植物图鉴》,共记录到野生兰科植物88属204种。其中,附生兰科植物104种,占总数的51.00%。该调查成果为本次研究提供了宝贵资料。
-
调查前查阅已有资料记录,结合野外踩点,根据鹦哥岭自然保护区的地形、地貌、附生兰科植物多样性概况,在各类地形、海拔段、植被区、小生境、附生兰丰富的区域设置样线。
-
以每条进入林区的路径为样线,以样线两侧5 m内出现的有兰科植物附生的乔木或藤本作为调查样木,记录附生兰物种及其宿主信息。参考《海南植物图志》[24]、《鹦哥岭兰科植物图鉴》[23]、《中国野生兰科植物原色图鉴》[25]确定宿主树、附生兰科植物的物种,中文名称、拉丁学名均参照《中国植物志》[26]2019网页版中的植物名称。样木所在地理坐标及海拔用GPS记录,坡向和坡度分别用罗盘、坡度仪进行实测,植被郁闭度采用抬头望法估测。
于2020年8月至2021年8月期间分不同季节进行调查,样木分布情况见图1。共设置样木477个,调查到附生兰40属78种。
图 1 野外调查样木分布图
Figure 1. Diagram of the sample trees
-
气温和雨量数据来自世界气候数据网站(
http: //www.worldclim.org/ )。 -
应用公式
$S_{1}=(1- $ $\cos (({\text{π}} / 180)(S-30))) / 2 $ (式中,S为坡向原始数据,S1为转化后的坡向数据)将坡向数据转换为0~1之间的数据,转换后的数据越大表示生境条件越干热,越小则说明生境条件越湿冷;坡向在0°~22.5°和337.5°~360°为北坡,22.5°~67.5°为东北坡,67.5°~112.5°为东坡,112.5°~157.5°为东南坡,157.5°~202.5°为南坡,202.5°~247.5°为西南坡,47.5°~292.5°为西坡,292.5°~337.5°为西北坡;附生兰物种丰富度由附生兰物种数表示,附生兰多度由附生兰个体数量表示。 -
排序及制图由Canoco for Windows 5.0 完成。排序的物种数据由78种附生兰物种在447个样木内出现或者不出现(0,1)组成,其矩阵组成(样方×物种)为447行×78列;环境数据矩阵由实测的6种环境因子和来自世界气候网站的19种气候因子组成,为447行×25列。
-
为避免因气候因子间存在多元共线性而导致的排序结果的不可靠,首先采用前向选择(forward-selection)通过自变量效应由大到小手动选取气候因子。被选取的气候因子有年平均温度、每月最高气温与最低气温差值的平均值(以下简称平均月温差)、最干月降雨量。预先进行除趋势对应分析(detrended correspondence analysis, DCA),结果显示,最长梯度大于4,说明物种组成数据存在较大差异,故选用典范对应分析(canonical correspondence analysis, CCA)研究环境因子对物种组成的影响。
-
以选取的3种气候因子和6种生境因子进行重分析。为明确各个因子对兰科植物物种组成影响的相对重要性,参照文献[28]的方法计算每一个环境因子对兰科植物物种组成的总效应与净效应,仅以1个环境因子作为解释变量进行CCA排序所得的物种组成变异的解释量为此环境因子的总效应,而以1个环境因子作为解释变量,其他环境因子作为协变量进行CCA排序所得的物种组成变异的解释量为此环境因子的净效应,所有排序的显著性均由Monte Carlo随机置换(999次)进行检验。其余作图由Origin 2018完成。
-
用于CCA排序的9个环境因子共解释了鹦哥岭附生兰组成总变异的7.8%,基于所有典范特征值之和的Monte Carlo随机置换检验为极显著(P < 0.01)(表1),说明物种与所选环境因子之间有着强烈的相关性。CCA前两轴有比第3和第4轴更高的特征值,说明前两轴在对物种组成变异的解释中起主导作用,第1轴解释了物种组成变异的1.82%,第2轴解释了物种组成变异的1.5%(表1)。CCA第1轴和第2轴代表了不同的环境梯度。CCA 1轴中,最干月降水量、海拔、山地雨林及热带针叶林与之呈显著正相关,年平均气温、平均月温差、树皮粗糙、低地雨林次生林、旱地农业生产植被、低地雨林及坡向与之呈显著负相关;主要为海拔梯度,温度梯度及降雨量梯度(高的相关系数)。CCA 2轴中年均气温、树皮有覆盖物、低地雨林次生林、热带针叶林与之呈显著正相关关系,最干月降水量、海拔、郁闭度山地雨林等与之呈显著负相关关系;表明该轴为1个综合轴,反映了气温、植被类型等的变化。
表 1 环境因子与典范对应分析(CCA)前4排序轴的相关系数及排序概要
Table 1. Correlation coefficients between environmental variables and the first four ordination axes generated by canonical correspondence analysis (CCA), and ordination summary
环境因子及排序概要 第1轴 第2轴 第3轴 第4轴 年平均气温 −0.7244 0.3695 0.2917 0.1838 每月最高气温与最低气温的差值平均值 −0.2874 0.1346 −0.7005 0.2767 最干月降雨量 0.7794 −0.3117 −0.2100 0.1618 海拔 0.9193 −0.2272 −0.1120 0.0002 郁闭度 0.0730 −0.2669 0.3062 0.5140 树皮光滑 −0.0066 −0.0514 0.2120 0.3365 树皮粗糙 −0.3236 −0.1897 −0.1731 −0.2646 树皮有覆盖物 0.4709 0.3302 0.0056 −0.0055 低地雨林次生林 −0.4168 0.2685 −0.1919 −0.1721 旱地农业生产植被 −0.2349 0.1465 0.2825 −0.0761 热带针叶林 0.6876 0.7125 0.0110 −0.0517 低地雨林 −0.2243 0.0781 −0.2343 0.2861 山地雨林 0.6125 −0.7359 0.2186 −0.0218 坡向 −0.2970 0.1531 0.3493 −0.0937 坡度 0.0600 −0.2970 −0.1318 0.0011 CCA排序概要 特征值 0.8445 0.6955 0.4556 0.3077 物种组成变异累积解释率 1.82 3.32 4.3 4.96 典范轴的相关性 0.9537 0.9082 0.7298 0.6377 物种−环境关系累积解释率 23.26 42.42 54.97 63.45 注:所有典范轴的显著性检验:pseudo-F = 2.4; P = 0.001。 基于Monte Carlo随机置换的检验结果表明(表2),除坡度因子外,其余生境因子均显著地解释了附生兰组成的变异。净效应由大到小依次为植被类型、树皮类型、年平均气温、每月最高气温与最低气温的差值平均值、最干月降雨量、海拔、郁闭度、坡向、坡度,仅坡度的净效应未达到显著。
表 2 环境因子对物种组成的总效应与净效应
Table 2. Gross and net effects of environmental variables on species composition
环境因子 总效应 pseudo-F P 净效应 pseudo-F P 植被类型 2.3 2.5 0.001 2.3 2.5 0.001 树皮类型 1.2 2.7 0.001 0.7 1.6 0.008 年平均气温 1.3 6 0.001 0.6 2.7 0.001 每月最高气温与最低气温的差值平均值 0.8 3.6 0.001 0.6 2.4 0.001 最干月降雨量 1.4 6.3 0.001 0.6 2.5 0.001 海拔 1.7 7.5 0.001 0.5 2.2 0.001 郁闭度 0.6 2.7 0.001 0.4 1.7 0.003 坡向 0.6 2.5 0.001 0.4 1.7 0.001 坡度 0.4 1.7 0.003 0.3 1.2 0.177 -
在进行排序分析的78种兰科植物中,分布较广的种有金塔隔距兰(Cleisostoma filiforme)、钗子股(Luisia morsei)、海南石斛(Dendrobium hainanense)、纯色万代兰(Vanda subconcolor)、大序隔距兰(Cleisostoma paniculatum)等,发生样木数分别为55、50、41、40、32,在总样木数(447)中的占比分别为12.30%、11.86%、9.17%、8.95%、7.16%。而分布样木数仅为1的稀有种类也达到了14种,包括胼胝兰(Biermannia calcarata)、蛇舌兰(Diploprora championii)、短耳鸢尾兰(Oberonia falconeri)、厚唇兰(Epigeneium clemensiae)、镰叶盆距兰(Gastrochilus acinacifolius)等。
CCA双序图(图2)中展示的受生境因子影响最显著的20种兰科植物。位于排序图右上角的物种,如白唇槽舌兰(Holcoglossum subulifolium)、乐东石豆兰(Bulbophyllum ledungense)、伏生石豆兰(Bulbophyllum reptans)、黑毛石斛(Dendrobium williamsonii)等附生兰,对应的生境条件为中高海拔、较多的降雨量、较低的年均气温及较小的平均月温差,其多生于湿冷、树皮有厚覆盖物的的热带针叶林中的宿主上;位于排序图中下部的物种,如石斛(Dendrobium nobile)、狭叶金石斛(Flickingeria angustifolia)、石仙桃(Pholidota chinensis)、粗茎苹兰(Pinalia amica)、指叶拟毛兰(Mycaranthes pannea)、匙唇兰(Schoenorchis gemmata)、牛角兰(Ceratostylis hainanensis)等附生兰,对应的生境为中低海拔地段、坡度大、郁闭度较高、降雨较为充沛的山地雨林;位于双序图左部的附生兰,如多花脆兰(Acampe rigida)、海南石斛(Dendrobium hainanense)、钗子股(Luisia morsei)、金塔隔距兰(Cleisostoma filiforme)、大尖囊蝴蝶兰(Phalaenopsis deliciosa)等,对应的生境条件为低海拔、较高的年平均温度、较大的平均月温差,其分布区多为旱地农业生产植被或低地雨林次生林,往往生长于阳坡。其他未展示的大部分种类出现在排序图中心点附近,它们的分布受到生境因子的综合影响相对较小。
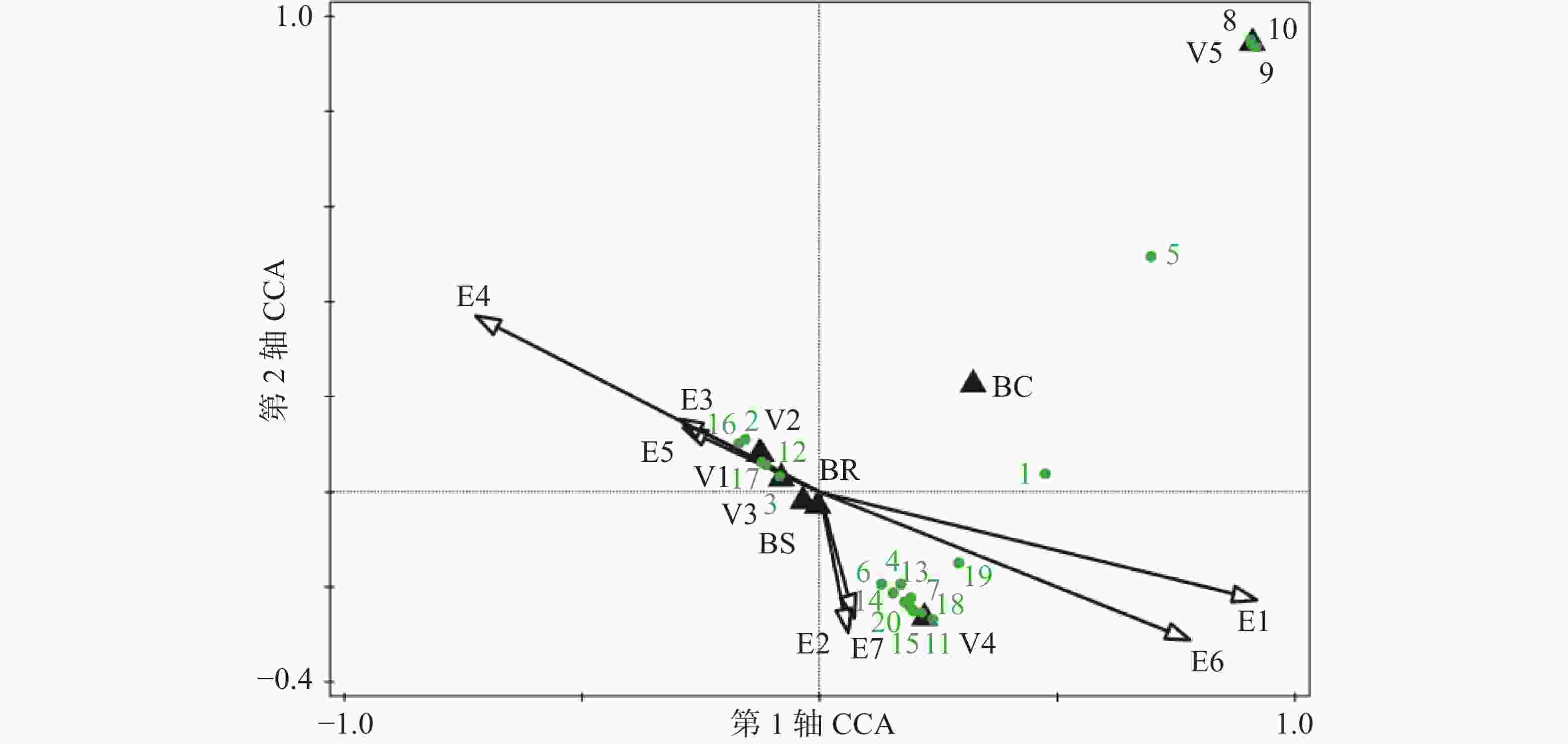
图 2 物种与环境因子的典范对应分析(CCA)双序图
Figure 2. Canonical correspondence analysis (CCA) biplot of species and environmental variables
-
在垂直水平上,把鹦哥岭林区划分为8个海拔段,各海拔段附生兰物种丰富度及物种多度情况见图3-A。随着海拔的升高,附生兰物种数呈现出先上升后下降的趋势。700~900 m海拔段出现峰值,生长有47种附生兰,占调查到的总物种数的60.26%;1300~1500 m海拔段附生兰物种数最少,仅芳香石豆兰、流苏贝母兰、厚唇兰、黑毛石斛4种,占总物种数的5.13%。附生兰个体数随着海拔的升高而递增。300 m以下海拔段平均每木有附生兰33.51株,而在海拔1500 m以上平均每木附生兰达686株。
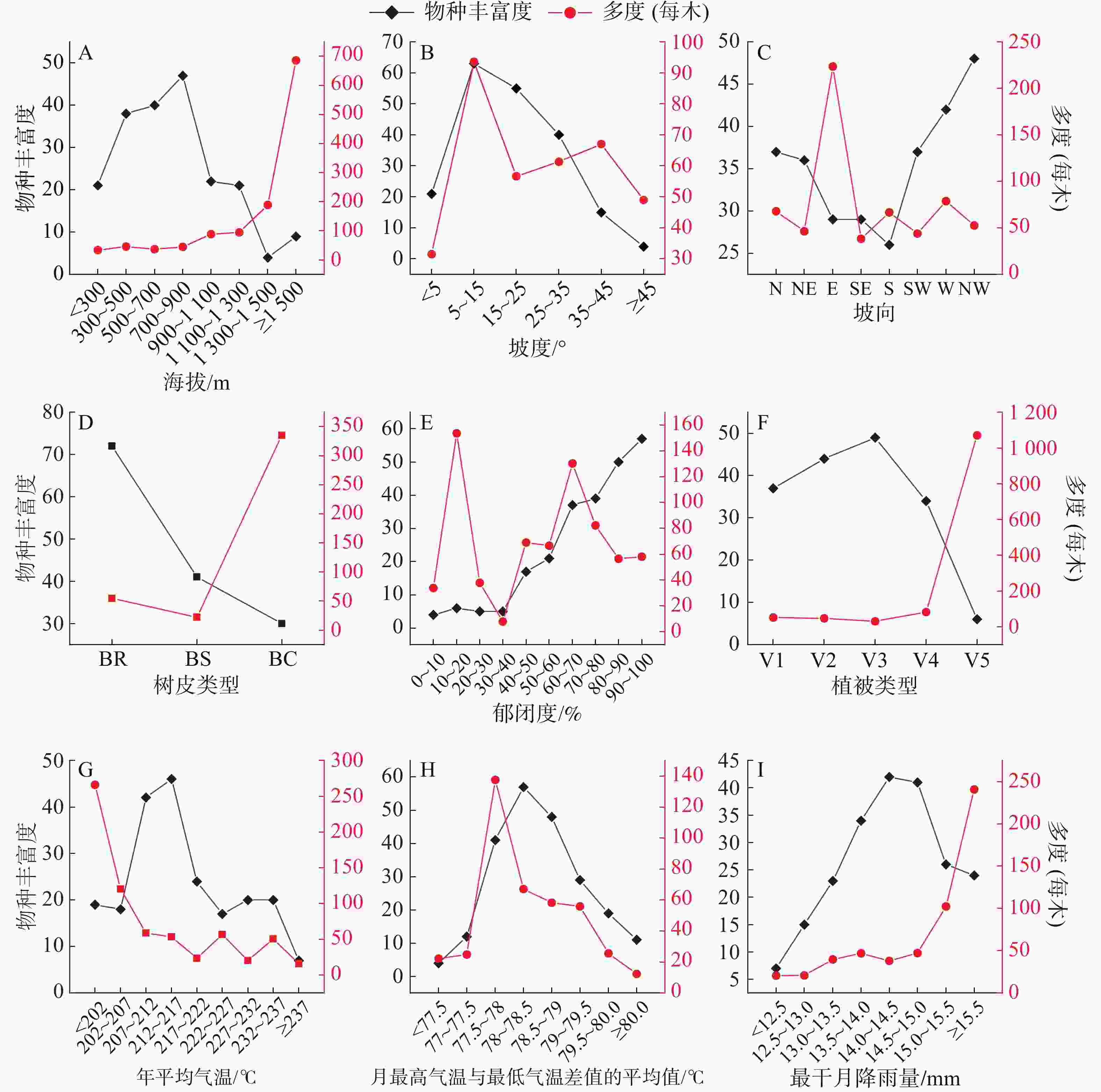
图 3 附生兰物种丰富度、多度与9个生态因子的关系
Figure 3. The relationship between the species richness and abundance of epiphytic orchids and 9 ecological variables
研究区域的坡度可划分为6个等级(图3-B),5°以下为平坡,5°~15°为缓坡,15°~25°为斜坡,25°~35°为陡坡,35°~45°为急坡,45°以上为险坡(含最小值不含最大值,下同)。缓坡的附生兰物种数最多,达63种,占总物种数的80.77%;陡坡的附生兰物种数最少,仅4种,分别为藓叶卷瓣兰、石斛、白点兰、金塔隔距兰,占总物种数的5.13%。附生兰个体数目的峰值同样出现在缓坡地段,平均每木着生有附生兰93.59株,平坡地段平均每木附生兰个体数最少,为31.52株。
图3-C显示,在研究区域的8个坡向地段中,附生兰物种分布数目较多的为西北坡、西坡、北坡,而南坡、东南坡、东坡物种分布数目相对较少,南坡分布的附生兰物种数最少,为26种,占总物种数的33.33%,多为相对耐旱物种,如钗子股、硬叶兰,多花脆兰,金塔隔距兰、海南石斛等。东坡分布的附生兰个体数量最大,平均每木达223.40株,其余坡向上附生兰个体数量差别不明显。
根据树皮表面特征,将附生兰样木树皮划分为粗糙,光滑,有覆盖物(如苔藓植物)3类(图3-D)。树皮粗糙的样木上分布有附生兰达72种,占总物种数目的92.31%。而树皮上有覆盖物的样木着生有个体数量较多的附生兰,平均每木可达335株,大量着生于有覆盖物树皮上的代表物种有乐东石豆兰、伏生石豆兰、黑毛石斛、指叶拟毛兰、芳香石豆兰等。
附生兰在不同郁闭度区间的分布情况见图3-E。附生兰丰富度随着郁闭度的增大而递增,郁闭度在10%以下的林区仅分布有绒兰、海南石斛、红花隔距兰、短耳鸢尾兰等4种附生兰,占总物种数的5.13%,而在郁闭度90%以上的林区则分布有57种附生兰,占总物种数的73.77%。附生兰个体数量在10%~20%、60%~70%两个郁闭度区间出现峰值,暗示着不同的附生兰对郁闭度有不同的偏好。
在本研究区域内,低地雨林中附生兰物种丰富度最高,为49种,占总物种数的62.83%,代表物种有纯色万代兰、海南钻喙兰、白点兰、红花隔距兰、海南石斛等,热带针叶林中分布的附生兰物种最少,仅伏生石豆兰、白唇槽舌兰、流苏贝母兰、黑毛石斛、乐东石豆兰、粗茎苹兰等6种,占总物种数的7.69%。在热带针叶林种,附生兰多度明显较高,平均每木可达1 070株,表明热带针叶林中的附生兰具有较高的优势度。
图3-G和3-H分别展示了附生兰物种丰富度、物种多度随生境年平均气温、平均月温差变化的情况。研究区域附生兰物种丰富度随年平均气温的升高呈现先升高后降低的趋势,在21.2~21.7 ℃段出现峰值,物种丰富度达47种,占总物种数的60.26%,年平均气温达到23.7 ℃以上时,附生兰物种丰富度最低,仅大尖囊蝴蝶兰、寄树兰、齿瓣鸢尾兰、短序脆兰、多花脆兰、大叶寄树兰、硬叶兰等7种,占总物种数的8.97%;附生兰多度随着年平均气温的升高呈现出递减的趋势。随着平均月温差的增大,附生兰物种丰富度和多度均呈现出先升高后降低的趋势,温差在7.75~7.90之间的地段附生兰物种丰富度及多度较高。
附生兰物种丰富度及多度随研究区域最干月降雨量的变化情况见图3-I。附生兰物种丰富度随最干月降雨量增多而先升高后降低,在最干月降雨量为14~15 mm地段附生兰物种丰富度最高,达40种以上。在12.5 mm以下附生兰分布很少,仅大尖囊蝴蝶兰、寄树兰、齿瓣鸢尾兰、短序脆兰、多花脆兰、大叶寄树兰、硬叶兰等7种,占总物种数的8.79%,而这些附生兰均生长在沟谷旁林内。随着最干月降雨量的增多,附生兰多度呈现出递增的趋势。
-
本研究中的环境因子可划分为地形因子如海拔、坡度和坡向等,气候因子如年平均气温、平均月温差和最干月降雨量等,生物因子如植被郁闭度、植被类型和树皮类型等。
-
CCA分析确定了研究区域种类组成的主要梯度,并用解释变量对其进行解释。虽然环境梯度具有明显的生态解释,但CCA排序轴解释的变异很低(7.8%)。采用间接梯度分析DCA排序进一步探索物种组成数据,结果显示4个轴对物种组成变异的解释量为8.34,高于CCA排序轴(4.96),表明该研究对部分生态因子如生境湿度,群落的发育历史、树皮的化学特性等的影响没有考虑到,但是DCA分析也表现出较低的变异量,表明潜在的缺失变量并不比包含的4个变量更重要。同时,也说明研究区域的附生兰分化较深入,物种组成十分复杂。物种组成解释率较低的可能原因是:物种组成数据矩阵是由(0,1)组成,且物种数据矩阵中有较多的0值;排序轴的多项式的失真[27]。但是,低的解释率并不影响所选环境因子对附生兰物种组成解释的高显著性。
-
海拔、坡度、坡向等地形因子一般通过影响湿度、光照、温度、降雨等能量环境间接对陆生植物的分布产生影响,且地形因子其较其他环境因子易于测量,因此常用来解释植物物种组成的变异[28-29]。本研究结果表明,海拔为影响鹦哥岭附生兰分布的重要环境因子,沿海拔梯度附生兰组成出现明显变异,基于总效应与净效应的结果也清晰表明了其对物种组成影响的显著性,相似的结论在其他地区的研究中也有报道[17, 20-21]。有研究认为,坡向与坡度的变化,可引起生境光照,湿度的变化进而影响物种组成[30-31],在本研究中,坡向的总效应和净效应均达显著水平,而坡度尽管总效应达到了显著水平,其净效应却不显著。可能的原因是,附生兰远离土壤基质,常着生在高大乔木的冠层区,坡度对其的影响显然要比地生植物小很多,余东莉等[12]和周康等[21]对其他地区兰科植物的研究也有相似的结果。
光照、温度、水分等生态因子直接影响着植物的生理过程,对植物的分布起着决定性的作用。对于附生兰科植物而言,林区郁闭度决定了生境的光照条件,温度则限制着附生兰的分布,降水则更多的是对林区空气湿度及附生基质(宿主树皮)含水量的调节。有人通过研究得出年平均温度、年降雨量等气候因子对兰科植物的物种组成具有显著影响[17]。本研究中,郁闭度、年平均温度、平均月温差及最干月降雨量的总效应及净效应均达到了高的显著性,可见不同的附生兰对气候因子的需求相异,表明了研究区域的附生兰较高的分化程度。
植被类型本身是特定区域的各类植物与诸多环境因子、植物之间长期相互作用的结果,附生兰的存在即表明其在当前的植被类型中占据一定的生态位,因而植被类型的变化能够很好的反映附生兰在物种组成上的变异。据笔者观察,植被类型往往比其他环境因子具更高的解释率,如热带针叶林中的黑毛石斛+乐东石豆兰+伏生石豆兰组合、低地雨林中的大序隔距兰+纯色万代兰+窄唇蜘蛛兰组合等,亦有相关研究支持了此观点[21]。区别于众多地生植物所依赖的土壤,宿主树皮是附生兰着生的载体,其对附生兰的生长发育的作用至今尚未完全明晰[11, 20],但不难推断的是,树皮的表面特征决定了附生兰附着及种子萌发的难易程度[5]。本研究结果表明,植被类型为影响附生兰物种组成最重要的因子,在所选的9个环境因子中,植被类型对兰科植物物种组成变异的解释率最高并且也达到了高的显著水平,其次则为树皮类型。
-
对于热带兰科植物的多数研究表明,其垂直分布格局为“中间膨胀型”[15-16, 19]。本研究结果显示,鹦哥岭附生兰在中低海拔段(700~900 m)有着最高物种丰富度,这与许多学者的研究一致,其原因可能是低海拔地区人为干扰强度较大,附生兰和其生境易遭破坏,而高海拔地区极端的环境条件不利于多数附生兰的生长。少数适应于高海拔湿冷条件的附生兰如黑毛石斛、伏生石豆兰、流苏贝母兰等,由于竞争小,环境条件稳定且极少受到人类活动干扰,因而个体数量庞大,在高海拔地区占有绝对的优势。坡度对不同类型的植物分布具有不同的影响[32],对于远离土壤的附生兰而言,其产生的作用则更加微妙复杂。本研究结果表明,缓坡地段,附生兰物种丰富度较高,而物种多度的变化在坡度梯度上呈现出先升高后降低的趋势。可能的原因是,缓坡地段土壤肥沃且排水良好,有利于高大粗壮宿主树的长成,可为附生兰提供更多的生长环境。在山地区域,坡向的变化影响了环境的光照条件和水分的分配,从而间接影响了植物的分布。兰科植物多为喜湿耐阴的草本植物。本研究结果显示,附生兰在阴坡与半阴坡的兰科物种数和个体数量要多于阳坡与半阳坡,证明附生兰对光照的低的需求度和对空气湿度的高的需求。
气候是影响植物分布的主导因素,表现在热量是植物生命活动能量来源、水分是影响植物生理活动的因素和植物基本成分[33]。一般而言,附生型兰科植物多产于热带或亚热带,可在较高温度的环境下生长。在本研究中,附生兰物种丰富度随年平均气温的升高呈现先升高后降低的趋势,在21.2~21.7 ℃段出现峰值,物种丰富度最高,年平均气温达到23.7 ℃以上时,附生兰物种丰富度很低,类似的结论在同为中国热带地区的西双版纳的兰科植物分布研究中亦见报道[17]。温差可影响植物生理活动中有机物质的合成量、开花率[34]等,从而影响植物的分布与繁殖力。本研究表明,温差在7.8~7.85 ℃之间的地段附生兰物种丰富度较高,该研究结果对于当地附生兰集中保护区域的选取具重要的参考意义。对于附生兰而言,降雨影响着附生基质(树皮)的含水量和林分的湿度。本研究得出,附生兰物种丰富度随最干月降雨量增多而先升高后降低,在最干月降雨量为14~15 mm地段附生兰物种丰富度最高,随着最干月降雨量的增多,附生兰多度呈现出递增的趋势,可见,多数附生兰分布的适宜生境为降雨量较大的林区。
区域植被类型是一定区域内植物群落与地形、气候、人为因素等长时间相互作用的结果,往往能综合反映该区域的环境条件与植物特征。附生兰是热性林的代表成分之一,因而在位于热带边缘的海南岛,附生兰物种丰富度一定程度上反映了该地段热带属性的强弱[35-36]。在研究区域内,低地雨林中附生兰物种丰富度最高,热带针叶林中附生兰物种丰富度最低。该结果与刘广福等[19]在海南霸王岭对附生兰在不同类型森林植被中的分布结论略有不同,原因可能是,尽管附生兰的分布依赖于特定的植被类型,但在确定的植被类型内部,附生兰生境小气候很容易受到各种因素复杂的影响。实际上,不论是在哪种植被类型的林区,均存在有大片面积的地段中观察不到附生兰的情况,而附生兰如灰尘般的种子的传播能力是毋庸置疑的。因此,笔者认为,附生兰对的生长对小气候有着极高的要求,大多数环境难以满足附生兰种子萌发及幼苗生长的要求[37]。植被郁闭度一方面塑造了附生兰的光照环境,另一方面影响了林分的空气湿度。本研究得出,郁闭度较大的林区具有略高的附生兰物种丰富度和个体数量,表明多数附生兰对光照需求很低,这与其他地区对附生兰的研究结论一致[17]。树皮类型决定了附生兰种子附着与萌发的难易程度,从而影响了附生兰的分布。研究区域内附生兰广泛分布于粗糙树皮上,而高海拔区域的覆盖有厚苔藓的树皮上,往往分布有数目庞大的附生兰,可见树皮基质对于附生兰的生长具有重要作用。树皮可能包含诸多对附生兰生长起重要作用的因素,如含水量、化学物质、微生物群落、pH等等,相关的研究尚不深入[20, 38-40]。
总的来说,自然环境中附生兰的分布受多种生态因子的共同作用,其作用机制也十分复杂,在不同的环境中,附生兰对各单一的生态因子的需求也有所变化。植被类型较大程度包含了附生兰分布的生境需求,实际上也代表了各个环境因子的适宜范围。
Relationship between species composition and distribution of epiphytic orchids and ecological factors in Yinggeling Mountain, Hainan
-
摘要: 为了研究附生兰的物种组成、分布及与生态因子的关系,对海南热带雨林国家公园中的鹦哥岭地区的附生兰进行了调查,采用CCA探索了环境因子对物种组成的影响,计算了各环境因子对物种组成的总效应和净效应,并分析了附生兰物种丰富度和多度在不同环境梯度的变化情况。结果表明:(1) 所选9个环境因子共解释了物种组成变异的7.8%,其净效应由大到小依次为植被类型、树皮类型、年平均气温、每月最高气温与最低气温的差值平均值、最干月降雨量、海拔、郁闭度、坡向、坡度,仅坡度的净效应未达到显著。(2) 随海拔升高,附生兰物种丰富度呈现出先上升后下降的趋势,中低海拔段(700~900 m)物种最丰富,附生兰多度呈现出递增的趋势;缓坡地段,附生兰物种丰富度与多度均较高,阳坡和半阳坡的附生兰物种丰富度与多度较阴坡与半阴坡少;粗糙的树皮更易着生附生兰,覆盖物为树皮上的附生兰提供了良好的生长环境;多数附生兰倾向于生长在弱光环境中;低地雨林中附生兰物种最丰富,热带针叶林中的附生兰具较高的优势度;附生兰倾向于分布在较低的气温、每月最高气温与最低气温差值在(7.8~7.85)℃之间的生境中;在降雨量较多的环境中,附生兰物种丰富度和多度较高。Abstract: The relationship between species composition and distribution of epiphytic orchids and ecological factors is of great significance for epiphytic orchids conservation and habitat protection. A survey was made of the epiphytic orchid species in the Yinggeling Mountain section of Hainan Tropical Rainforest National Park. The influence of ecological factors on the composition of epiphytic orchids was analyzed by using Canonical Correspondence Analysis (CCA) and the total effect and the net effect of the factors on the epiphytic orchid species were calculated by using partial CCA. The epiphytic orchid species richness and abundance in environmental changes at different gradients were analyzed. Results showed that the selected 9 ecological factors explained 7.8% of the variation of the species, with their net effect being in the order of vegetation type > bark type > annual average air temperature > average of monthly difference between the highest and lowest temperatures > precipitation in the driest month > altitude > canopy density > slope aspect > slope gradient. The factors except the slope gradient had significant net effect. The species richness of the epiphytic orchids increased and then decreased with the increase of altitude, and the species richness was the highest at the altitude of between 700 m and 900 m, but the abundance of the epiphytic orchids showed an increasing trend. The species richness and abundance of the epiphytic orchids were higher on the gentle slopes, and lower on the sunny and semi-sunny slopes than on the shady and semi-shady slopes. The rough bark was more likely to give rise to epiphytic orchids, and the covering on the barks provided comfortable growth environment for epiphytic orchids. Most epiphytic orchids tended to grow in a low-light environment. The epiphytic orchid species were the most abundant in tropical lowland rainforest, and the epiphytic orchids had higher dominance in tropical coniferous forests. The epiphytic orchids tended to be distributed in the habitats at low temperature, and the difference between monthly maximum temperature and monthly minimum temperature was (7.8 − 7.85) ℃. The species richness and abundance of the epiphytic orchids were higher in the environment with higher rainfall in the Yinggeling mountain.
-
Key words:
- epiphytic orchid /
- species composition /
- richness /
- abundance /
- ecological factor
-
图 2 物种与环境因子的典范对应分析(CCA)双序图
E1.海拔;E2.坡度;E3.坡向;E4.年平均气温;E5.每月最高气温与最低气温差值的平均值;E6.最干月降雨量;E7.郁闭度;V1.旱地农业生产植被;V2.低地雨林次生林;V3.低地雨林;V4.山地雨林;V5.热带针叶林;BR.树皮粗糙;BS.树皮光滑;BC.树皮有覆盖物;1.流苏贝母兰;2.多花脆兰;3.海南石斛;4.石斛;5.黑毛石斛;6.狭叶金石斛;7.芳香石豆兰;8.白唇槽舌兰;9,乐东石豆兰;10,伏生石豆兰;11,藓叶卷瓣兰;12,钗子股;13,钟兰;14,牛角兰;15,指叶拟毛兰;16.大尖囊蝴蝶兰;17.金塔隔距兰;18.石仙桃;19.粗茎苹兰;20.匙唇兰。
Fig. 2 Canonical correspondence analysis (CCA) biplot of species and environmental variables
E1. Altitude; E2. Slope gradient; E3. Slope aspect; E4. Annual average air temperature; E5. Average of monthly difference between the highest and lowest temperatures; E6. Precipitation in the driest month; E7. Crown density; V1. Dryland agricultural vegetation; V2. Tropical lowland rainforest secondary forest; V3. Tropical lowland rainforest; V4. Mountain rainforest; V5. Tropical coniferous forest; BR. Bark rough; BS. Bark smooth; BC. Bark with coverings; 1. Coelogyne fimbriata; 2. Acampe rigida; 3. Dendrobium hainanense; 4. Dendrobium nobile; 5. Dendrobium williamsonii; 6. Flickingeria angustifolia; 7. Bulbophyllum ambrosia; 8. Holcoglossum subulifolium; 9. Bulbophyllum ledungense; 10. Bulbophyllum reptans; 11. Bulbophyllum retusiusculum; 12. Luisia morsei; 13. Campanulorchis thao; 14. Ceratostylis hainanensis; 15. Mycaranthes pannea; 16. Phalaenopsis deliciosa; 17. Cleisostoma filiforme; 18. Pholidota chinensis; 19. Pinalia amica; 20. Schoenorchis gemmata.
图 3 附生兰物种丰富度、多度与9个生态因子的关系
A.海拔;B.坡度;C.坡向;D.树皮类型;E.郁闭度;F.植被类型;G.年平均气温;H.每月最高气温与最低气温差值的平均值;I.最干月降雨量;V1.旱地农业生产植被;V2.低地雨林次生林;V3.低地雨林;V4.山地雨林;V5.热带针叶林;BR.树皮粗糙;BS.树皮光滑;BC.树皮有覆盖物。
Fig. 3 The relationship between the species richness and abundance of epiphytic orchids and 9 ecological variables
A. Altitude; B. Slope gradient; C. Slope aspect; D. Bark type; E. Crown density; F. Vegetation type; G. Annual average air temperature; H. Average of monthly difference between the highest and lowest temperatures; I. Precipitation of the driest month; V1. Dryland agricultural production vegetation; V2. Tropical lowland rainforest secondary forest; V3. Tropical lowland rainforest; V4. Mountain rainforest; V5. Tropical coniferous forest; BR. Bark rough; BS. Bark smooth; BC. Bark with coverings.
表 1 环境因子与典范对应分析(CCA)前4排序轴的相关系数及排序概要
Table 1 Correlation coefficients between environmental variables and the first four ordination axes generated by canonical correspondence analysis (CCA), and ordination summary
环境因子及排序概要 第1轴 第2轴 第3轴 第4轴 年平均气温 −0.7244 0.3695 0.2917 0.1838 每月最高气温与最低气温的差值平均值 −0.2874 0.1346 −0.7005 0.2767 最干月降雨量 0.7794 −0.3117 −0.2100 0.1618 海拔 0.9193 −0.2272 −0.1120 0.0002 郁闭度 0.0730 −0.2669 0.3062 0.5140 树皮光滑 −0.0066 −0.0514 0.2120 0.3365 树皮粗糙 −0.3236 −0.1897 −0.1731 −0.2646 树皮有覆盖物 0.4709 0.3302 0.0056 −0.0055 低地雨林次生林 −0.4168 0.2685 −0.1919 −0.1721 旱地农业生产植被 −0.2349 0.1465 0.2825 −0.0761 热带针叶林 0.6876 0.7125 0.0110 −0.0517 低地雨林 −0.2243 0.0781 −0.2343 0.2861 山地雨林 0.6125 −0.7359 0.2186 −0.0218 坡向 −0.2970 0.1531 0.3493 −0.0937 坡度 0.0600 −0.2970 −0.1318 0.0011 CCA排序概要 特征值 0.8445 0.6955 0.4556 0.3077 物种组成变异累积解释率 1.82 3.32 4.3 4.96 典范轴的相关性 0.9537 0.9082 0.7298 0.6377 物种−环境关系累积解释率 23.26 42.42 54.97 63.45 注:所有典范轴的显著性检验:pseudo-F = 2.4; P = 0.001。  下载: 导出CSV
下载: 导出CSV
表 2 环境因子对物种组成的总效应与净效应
Table 2 Gross and net effects of environmental variables on species composition
环境因子 总效应 pseudo-F P 净效应 pseudo-F P 植被类型 2.3 2.5 0.001 2.3 2.5 0.001 树皮类型 1.2 2.7 0.001 0.7 1.6 0.008 年平均气温 1.3 6 0.001 0.6 2.7 0.001 每月最高气温与最低气温的差值平均值 0.8 3.6 0.001 0.6 2.4 0.001 最干月降雨量 1.4 6.3 0.001 0.6 2.5 0.001 海拔 1.7 7.5 0.001 0.5 2.2 0.001 郁闭度 0.6 2.7 0.001 0.4 1.7 0.003 坡向 0.6 2.5 0.001 0.4 1.7 0.001 坡度 0.4 1.7 0.003 0.3 1.2 0.177
下载: 导出CSV
-
[1] SWARTS N D, DIXON K W. Terrestrial orchid conservation in the age of extinction [J]. Ann Bot, 2009(3): 543 − 556. [2] 罗毅波, 贾建生, 王春玲. 中国兰科植物保育的现状和展望[J]. 生物多样性, 2003(1): 70 − 77. doi: 10.3321/j.issn:1005-0094.2003.01.010 [3] TREMBLAY R L, MELENDEZ A E, KAPAN D. Do epiphytic orchids behave as metapopulations? Evidence from colonization, extinction rates and asynchronous population dynamics [J]. Biological Conservation, 2006, 129(1): 70 − 81. doi: 10.1016/j.biocon.2005.11.017 [4] SCHMIDT G, ZOTZ G. Inherently slow growth in two Caribbean epiphytic species: A demographic approach [J]. Journal of Vegetation Science, 2002, 13(4): 527 − 534. doi: 10.1111/j.1654-1103.2002.tb02079.x [5] BENZING D H. Vascular Epiphytes: General Biology and Associated Biota[M]. UK: Cambridge University Press, 1990: 1 − 43. [6] 赵明旭, GEEKIYANAGE N, HARRISON R D, 等. 西双版纳布龙自然保护区勐宋片区附生维管植物多样性与分布特征(英文)[J]. 植物分类与资源学报, 2015, 37(3): 327 − 338. [7] 施文艳, 龙光强, 杨生超, 等. 附生植物生理生态适应性研究进展[J]. 热带亚热带植物学报, 2018, 26(1): 92 − 106. doi: 10.11926/jtsb.3752 [8] HU X, ZHU J K, SONG X Q, et al. Orchid diversity in China’s Hainan Island: Distribution and conservation [J]. Collectanea Botanica, 2015, 34: e7. doi: 10.3989/collectbot.2015.v34.007 [9] NURFADILAH S. Diversity of epiphytic orchids and host trees (Phorophytes) in secondary forest of Coban Trisula, Malang Regency, East Java, Indonesia [J]. Biotropla, 2015, 22(2): 120 − 128. [10] PATRICK M. Diversity and distribution of Epiphytic Orchids in Kibale National Park, Uganda [J]. Selbyana, 2008, 29(2): 217 − 225. [11] TIMSINA B, ROKAYA M B, MUNZBERGOVA Z, et al. Diversity, distribution and host-species associations of epiphytic orchids in Nepal [J]. Biodiversity and Conservation, 2016, 25(13): 2803 − 2819. doi: 10.1007/s10531-016-1205-8 [12] 余东莉, 杨正斌, 宋志勇, 等. 西双版纳石灰山季雨林附生兰科植物种群动态研究[J]. 林业调查规划, 2018, 43(3): 34 − 39. doi: 10.3969/j.issn.1671-3168.2018.03.007 [13] 余东莉, 杨正斌, 宋志勇. 西双版纳坝区独生古树附生兰科植物多样性研究[J]. 林业调查规划, 2018, 43(4): 207 − 212. doi: 10.3969/j.issn.1671-3168.2018.04.047 [14] YULIA N D, BUDIHARTA S, YULISTYARINI T. Analysis of epiphytic orchid diversity and its host tree at three gradients of altitudes in Mount Lawu, Java [J]. Biodiversitas, 2011, 12(4): 294. [15] 董全英. 海南五指山国家级自然保护区附生兰科植物生态习性研究[D]. 上海: 华东师范大学, 2013. [16] 王喜龙, 刘强, 余东莉, 等. 西双版纳片段化石灰岩森林附生兰科植物多样性研究[J]. 广西植物, 2016, 36(2): 162 − 169. doi: 10.11931/guihaia.gxzw201412036 [17] 余东莉, 杨鸿培, 杨正斌. 环境因子对西双版纳野生兰科植物物种组成和丰富度的影响[J]. 林业调查规划, 2016, 41(6): 35 − 41,48. doi: 10.3969/j.issn.1671-3168.2016.06.007 [18] 张玉武, 喻理飞. 贵州梵净山生物圈保护区原生兰科植物生态特征初步研究[J]. 山地农业生物学报, 2009, 28(4): 288 − 293. doi: 10.3969/j.issn.1008-0457.2009.04.002 [19] 刘广福, 臧润国, 丁易, 等. 海南霸王岭不同森林类型附生兰科植物的多样性和分布[J]. 植物生态学报, 2010, 34(4): 396 − 408. doi: 10.3773/j.issn.1005-264x.2010.04.005 [20] YAGYA P A, ANTON F, HAGEN S F, et al. Diversity, composition and host-species relationships of epiphytic orchids and ferns in two forests in Nepal [J]. Journal of Mountain Science, 2017, 14(6): 1065 − 1075. doi: 10.1007/s11629-016-4194-x [21] 周康, 张哲, 宋希强, 等. 海南主要自然保护地兰科植物多样性与生境的关联分析[J/OL]. 广西植物, (2021−6−03) [2022−01−25]. http://kns.cnki.net/kcms/detail/45.1134.Q.20210602.1642.004.html. [22] 杨小波, 陈宗铸, 李东海, 等. 海南植被志(第一卷)[M]. 北京: 科学出版社, 2019: 124 − 155. [23] 卢刚, 方赞山, 刘磊, 等. 海南鹦哥岭兰科植物图鉴[M]. 海口: 海南出版社, 2018. [24] 杨小波. 海南植物图志[M]. 北京: 科学出版社, 2015. [25] 金效华, 李剑武, 叶德平. 中国野生兰科植物原色图鉴[M]. 郑州: 河南科学技术出版社, 2019. [26] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1993. [27] OKLAND R H. On the variation explained by ordination and constrained ordination axes [J]. Journal of Vegetation Science, 1999, 10(1): 131 − 136. doi: 10.2307/3237168 [28] 刘冠成, 黄雅曦, 王庆贵, 等. 环境因子对植物物种多样性的影响研究进展[J]. 中国农学通报, 2018, 34(13): 89 − 95. [29] 杨学成, 吴卓翎, 周庆, 等. 康禾自然保护区森林苔藓植物丰富度对地形因子的响应[J]. 生态环境学报, 2019(10): 1974 − 1981. [30] 张昌顺, 谢高地, 包维楷, 等. 地形对澜沧江源区高寒草甸植物丰富度及其分布格局的影响[J]. 生态学杂志, 2012, 31(11): 2767 − 2774. [31] 袁铁象, 张合平. 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响[J]. 应用生态学报, 2014, 25(10): 2803 − 2810. [32] 区余端, 苏志尧, 李镇魁, 等. 地形因子对粤北山地森林不同生长型地表植物分布格局的影响[J]. 应用生态学报, 2011, 22(5): 1107 − 1113. [33] 吕佳佳, 吴建国. 气候变化对植物及植被分布的影响研究进展[J]. 环境科学与技术, 2009, 32(6): 85 − 95. doi: 10.3969/j.issn.1003-6504.2009.06.020 [34] 苗鹭. 长春市植物物候对城市地表温度的响应[D]. 长春: 东北师范大学, 2009. [35] MORRIS J, WELLS T, WILLEMS J H. Population ecology of terrestrial orchids [J]. Journal of Ecology, 1991, 81(1): 202. [36] ROMERO G A. LIBROS: Dressler: Phylogeny and classification of the orchid family[M]. Portland: Dioscorides Press, 1993: 59-75. [37] 谢泰祥, 郑清冬, 王艺, 等. 兰科植物与共生真菌互作关系的研究进展[J]. 分子植物育种, (2021−07−19): 1 − 9. [2022−01−21]. http://kns.cnki.net/kcms/detail/46.1068.S.20210716.1720.016.html. [38] 李静静. 海南霸王岭拟石斛附生特性及其根部内生菌多样性的研究[D]. 海口: 海南大学, 2016. [39] WANG XM, LI YJ, SONG XQ, et al. Influence of host tree species on isolation and communities of mycorrhizal and endophytic fungi from roots of a tropical epiphytic orchid, Dendrobium sinense ( Orchidaceae) [J]. Mycorrhiza, 2017, 27(7): 709 − 718. doi: 10.1007/s00572-017-0787-7 [40] AWASTHIA O P, SHARMA E, L. PALNI L M. Stemflow: A source of nutrients in some naturally growing epiphytic orchids of the Sikkim Himalaya [J]. Annals of Botany, 1995, 75(1): 5 − 11. doi: 10.1016/S0305-7364(05)80003-4 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 647
- HTML全文浏览量: 239
- PDF下载量: 14
- 被引次数: 0


