

-
从地理梯度上看,纬度多样性梯度格局(Latitudinal Diversity Gradients,LDG)是地球上最广泛、最重要的生物多样性分布格局之一[1]。生物多样性在生态学研究中占据着重要地位,在以往研究中,植物群落多样性沿地理梯度变化的格局,一直以来都是生物多样性研究中的一个重要课题[2-3]。在群落生态学研究中,生物多样性的空间分布格局常常受到各种生态梯度因素制约的影响[4]。其中,物种多度分布格局通常也会随不同的气候环境而发生改变。群落多样性指数可以将群落结构数量化,进而分析出群落中的物种多度格局,在深入讨论群落性质上具有进一步的阐释意义。众多研究学者对物种丰富度的纬度分布格局做了大量研究,有结果显示,物种丰富度会在中纬度地区达到峰值,随纬度梯度变化整体趋势不明显[5-6],然而也有研究发现,在从两极到赤道,即随纬度下降,物种丰富度呈显著增加趋势[7-9],物种多度分布格局也随之发生相应变化。在热带森林群落中,中性理论模拟群落物种相对丰度、物种-面积关系和物种共存模式的效果表现得尤为突出[10-12];至温带森林群落,生态位分化过程在群落结构中起到了更为重要的主导作用[13]。一般来说,群落多度研究也能更全面地反映出不同群落之间的差异[14-15],若将群落的物种多度格局与多样性指数结合分析,达到相辅相成之效,同时也对群落结构的进一步研究具有更深远的意义。本研究以中国境内温带跨越至热带的4个森林群落为研究对象,选取3种种多度分布模型,对样地内木本植物群落物种多度曲线进行拟合,试图阐述木本植物群落物种多度格局随纬度梯度的变化规律,探讨各物种多度分布模型的拟合优度,探讨群落结构形成的生态学过程,以期对纬度梯度上的森林物种多样性维持机制有进一步了解。
-
本研究选择了位于国内温带至热带具有代表性的4个样地,分别为温带地区的长白山(Changbai Mountains, CBS)、中亚热带地区的古田山(Gutian Mountain, GTS)、南亚热带的鼎湖山(Dinghu Mountain, DHS)和热带北缘地区的西双版纳(Xishuangbanna, XSB)20~25 hm2不等的固定监测大样地,各固定监测样地的具体情况如下:
(1)吉林省长白山阔叶红松林25 hm2样地,位于42°N,植物群落是以红松(Pinus koraiensis)为主的针阔混交林,偶混有亚热带和亚寒带的植物成分,群落整体具有明显的成层现象和优势树种。该区域内主要受季风影响,会形成温带大陆性气候,年平均气温 3.6℃,年平均降水量700 mm,降水多集中在夏季,夏季短暂且较温暖,冬季漫长且寒冷。该植物群落内土壤分布多为山地森林土,颜色多呈暗棕色[16-17]。
(2)浙江省古田山亚热带常绿阔叶林24 hm2样地,位于29°N,植物群落是以黄山松(Pinus taiwanensis)针阔混交林为主,属于亚热带季风气候区。年平均气温为 15.3℃,最高气温常出现在 7 月,为 38.1℃,最低气温出现在 1 月,为−6.8℃,年均降雨量可达 1963.7 mm。在夏季因受季风影响,降水充沛,冬季季风因被山体遮挡,气候会稍显干燥,故容易形成冬季温暖、夏季凉爽湿润的气候特点。该植物群落中土壤因地形环境较复杂,沿海拔从下至上主要包括红壤、黄红壤、红黄壤和沼泽土4类,其中,红壤、黄红壤、红黄壤垂直分布于海拔 500~1000 m,而沼泽土则多分布于地形低洼地带[18-19]。
(3)广东省鼎湖山南亚热带常绿阔叶林20 hm2样地,位于24°N,植物群落垂直结构比较复杂,植被具有较为显著的地上成层现象,是北回归线附近保存较为完整的地带性植被。该区域属于亚热带向热带的过渡区域,为南亚热带季风湿润型气候,夏季和冬季气候交替明显,年平均气温为20.9℃,最热月常为7月,平均气温为28.0℃,最冷月为1月,平均气温为12.6℃,年降雨量为1 985 mm,且多集中于4—9月,多伴有台风。在海拔300 m以下的大部分丘陵地带多分布着赤红壤,并且土层厚度可达到40~80 cm;黄壤多见于海拔 300~900 m地区,土层厚度40~90 cm,当海拔高度超过900 m,多为山地灌丛草甸土,其厚度较薄,约20~30 cm [20]。
(4)云南省西双版纳热带季节雨林20 hm2样地,位于21°N,森林类型属热带季节雨林植被亚型,植物群落类型中多以望天树(Parashorea chinensis)为优势,属于北热带季风气候,常年受印度季风影响,气候温暖湿润,具有较强的季节性,且干湿季明显,年平均气温为 21.7℃,最高温常出现在6月,最低温出现在1月,年平均降雨量达1 531.9 mm,5—10 月为雨季,11月至翌年4月为干季。群落内的土壤分布类型多为砖红壤[21-24]。
-
本研究所选样地均严格按照 CTFS(Centre for Tropical Forest Science)[25] 的技术标准建设,并对样地内 DBH ≥ 1 cm 的木本植物进行统计分析,计算各样地群落内的α 多样性和β多样性,分析各样地中整体群落的物种多度分布格局;采用频度定义的方法[26-27],将各群落中频度大于25%的物种定义为常见种,而频度小于5%的物种定义为稀有种,并分析群落中常见种和稀有种各自的种多度分布格局。
-
(1)多样性指数常作为群落结构种的一种量化指标,可用于反映群落中的组成结构,也可为研究群落中各物种或个体分布之间相关关系提供系统性理解。基于各群落种实测所得的树木个体数据,α 多样性指数的计算公式如下:
Shannon-Wiener 多样性指数[28]:
$$ H=-\sum\nolimits_{i=1}^S P_i \log P_i ; $$ Simpson 优势度指数[29]:
$$ D_s=1-\sum\nolimits_{i=1}^s P_i^2 ; $$ 式中,Pi为群落中各植物种的重要值,S 为群落中的物种数。
(2)Bray-curtis 相异度常用于衡量不同群落间物种组成的差异度,可基于群落中不同物种组成的数量特征如多度、重要值等进行计算,公式如下:
$$ d^{B C D}=\frac{ \displaystyle \sum\nolimits_{i=1}^k\left|y_{i, k}-y_{j, k}\right|}{ \displaystyle \sum\nolimits_{i=1}^k\left(y_{i, k}+y_{j, k}\right)} $$ 式中,k 是物种数,y 表示物种多度,i 和 j 表示相比较的2个样地[30]。
-
(1)纯统计模型
对数级数模型:
$$ S(n)=\alpha \frac{X^n}{n} $$ 式中,S(n)为群落中多度为n的物种数,α代表群落内物种多样性;X为常数(0<X≤1)[31]。
对数正态模型:
$$ \begin{split} & A_i=e^{\log (\mu)+\log (\delta)} \varPhi \\& (\mathrm{i}=1,2,3, \cdots, S); \end{split}$$ 式中,Ai为第 i 个物种的多度,µ 和 δ 分别表示正态分布的均值和方差,Φ 为正态偏差[32]。
(2)生态位理论模型
生态位优先占领模型:
$$ \begin{split} & A_I=A_1(1-k)^{i-1} \\& (i=1,2,3, \cdots, S) ; \end{split}$$ 式中,模型假定种 1 先占用群落总生态位的 k 份,种 2 占用剩下的k(1−k)份,种 3 再继续占据余下的k(1−k)2 份,依次类推,直至剩余资源无法继续维持另一个物种的生存[33]。
断棍模型:
$$\begin{split} & A_i=\frac{J}{S} \sum_{x=i}^S \frac{1}{x} \\& (i=1,2,3, \cdots, S); \end{split} $$ 式中,该模型假设某一群落内的总生态位是长度为 1 的一条短棍,若将其分割为S 段,则代表生态位被S个物种所占有,模型假定S个物种的分类地位和竞争能力均相似,且在群落中同时出现。J表示群落内物种的个体总数,Ai 表示第Ai物种的多度[34]。
(3)中性理论模型
复合群落零和多项式模型:
$$ \begin{split} & S(n)=\frac{\theta}{n} \int_0^J f_{n, 1}(y)\left(1-\frac{y}{J}\right) ; \\& f_{n, \delta}(y)=\frac{1}{\varGamma(n) \delta^n} \exp \left(-\frac{y}{\delta}\right) y^{n-1} \end{split}$$ 式中,S(n)为群落中多度为 n 的物种数,θ 代表群落内物种多样性;J为取样点内个体数[35]。该模型假定某一群落的物种多度分布由自中性复合群落的随机漂变过程主导。
Volkov模型:
$$ S(n)=\theta \frac{J !}{n !(J-n)} \frac{\varGamma(\gamma)}{\varGamma(J+\gamma)} \int_0^\gamma \frac{\varGamma(n+y)}{\varGamma(1+y)} \frac{\varGamma(J-n+\gamma-y)}{\varGamma(\gamma-y)} d y \text {; } $$ $$ \varGamma(z)=\int_0^{\infty} t^{z-1} e^{-t} d t $$ $$ \gamma=\frac{m(J-1)}{1-m} $$ 式中,相较于复合群落零和多项式模型增加了迁移系数 m,S(n)为群落中多度为 n 的物种数,γ 表示迁移到局域群落内的个体数[36]。
-
本研究将采用 K-S 检验[37-38]对模型的拟合效果进行验证,同时还利用赤池信息准则(Akaike information criterion, AIC)来比较模型拟合的优劣。K-S 检验是一种常用的非参数检验方法,它通过计算2个样本经验分布函数(Empirical distribution function)之间的距离(统计量 D ),以此来判断二者是否存在显著差异。为了避免 K-S 检验中常发生的数据连结问题,在处理数据时通过优化 bootstrap,之后再进行 K-S 检验(其中,bootstrap 的次数为 1 000 次,在所得结果中,若当 P<0.05 时,则表明该模型被拒绝)[39]。同时还会利用赤池信息准则(Akaike information criterion, AIC)对各模型的拟合优度进行判断,在 AIC 检验中,若 AIC 的数值越小,则表示该模型拟合效果越好。
以上数据处理分析及优度检验均基于 R-4.2.1 软件(R Core Team, 2022),其中模型拟合使用 sads 软件包完成;基于 bootstrap 的 K-S 检验利用 Matching 软件包完成。
-
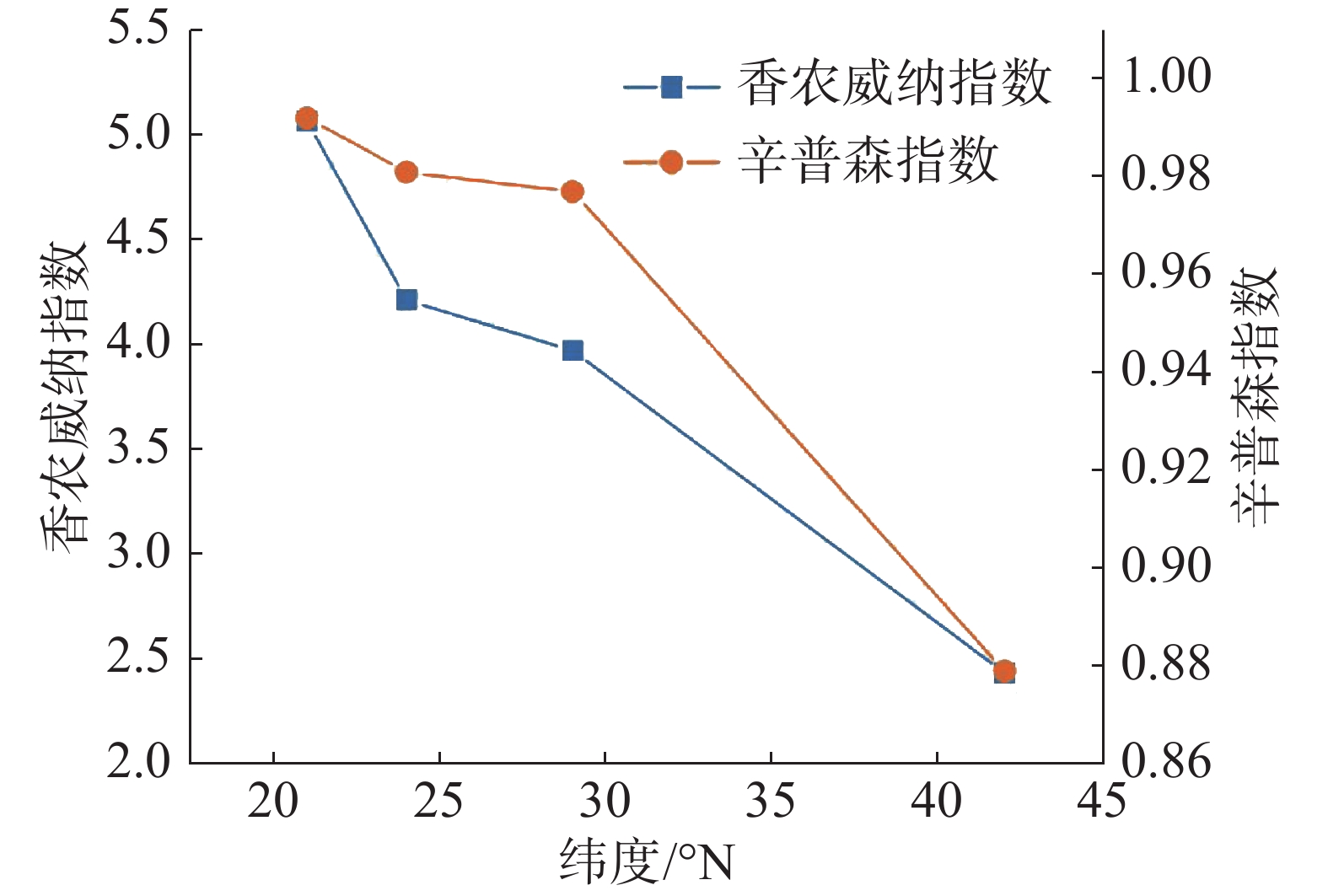
从图1 中可以看出,随纬度的上升,森林群落中α多样性呈现逐渐减小的趋势,在热带森林群落中,α多样性最高,在温带森林群落中则最小。其中Shannon-Wiener指数与Simpson指数的变化趋势基本保持一致。在β多样性方面,从图2 可以看出,沿纬度梯度,Bray-curtis 相异度逐渐增大,且在位于低纬度森林群落中,热带森林群落与纬度相邻的亚热带森林相异度小于其余纬度梯度群落之间的相异度。
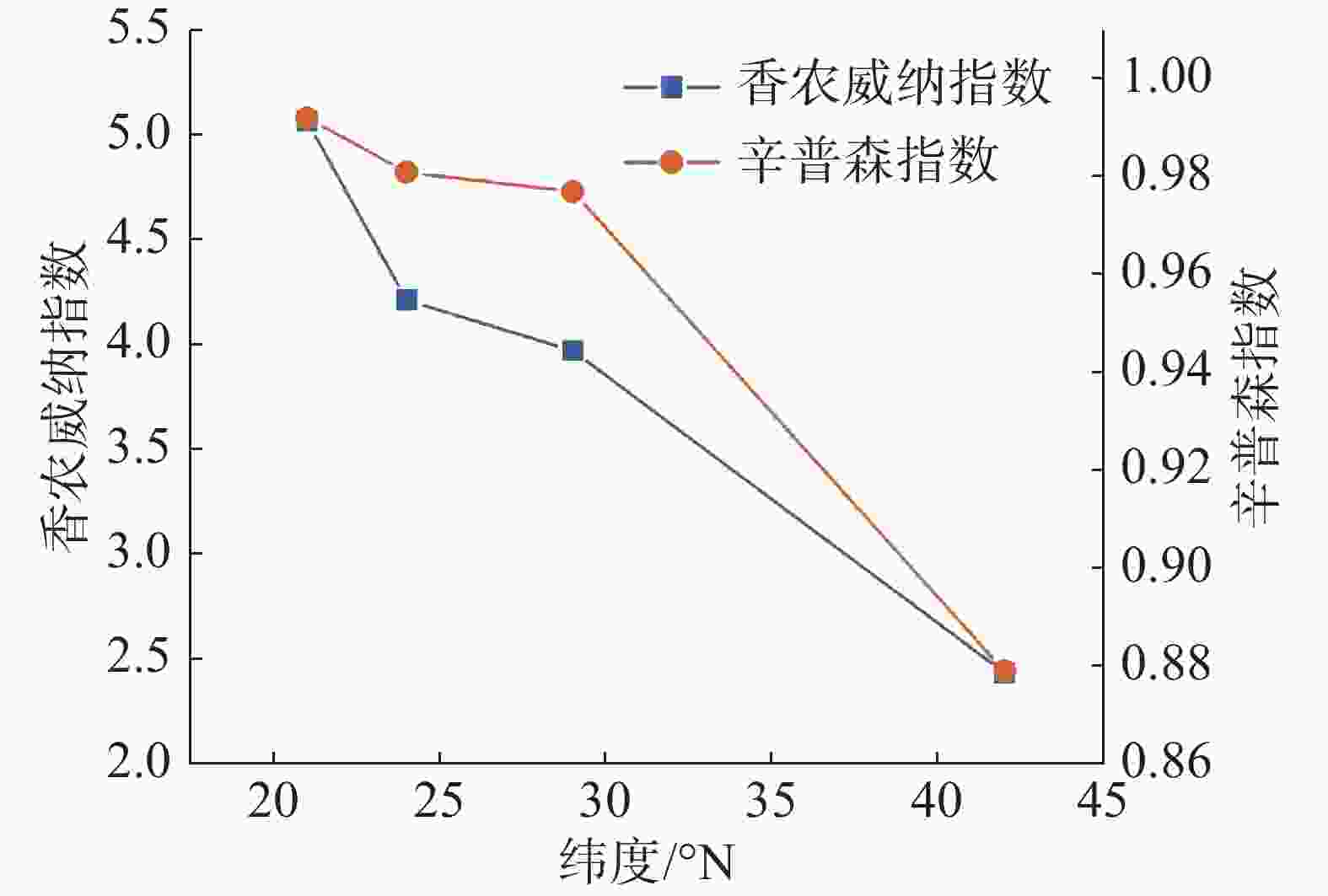
图 1 纬度梯度群落α多样性变化特征
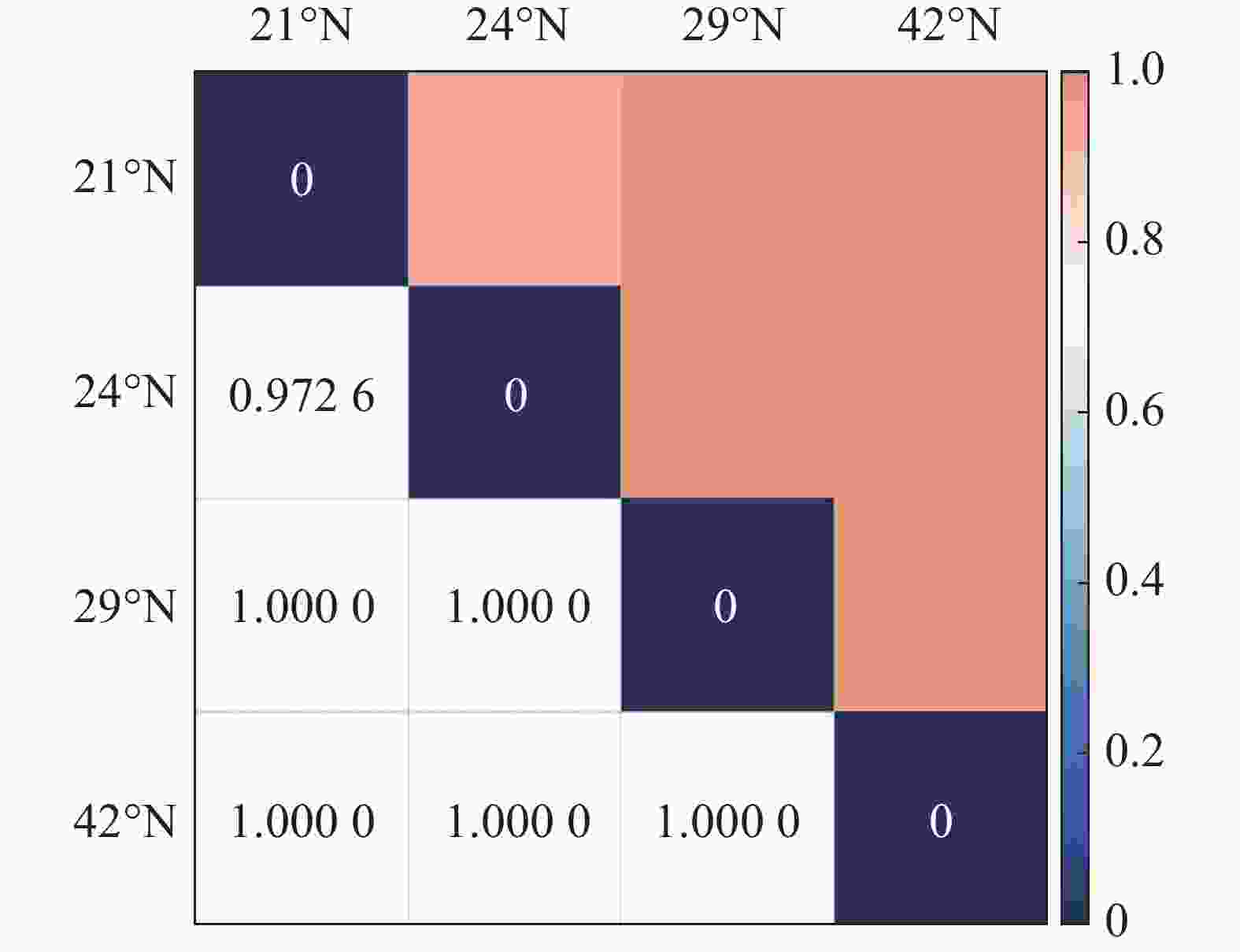
图 2 纬度梯度各群落 Bray-curtis 相异性
-
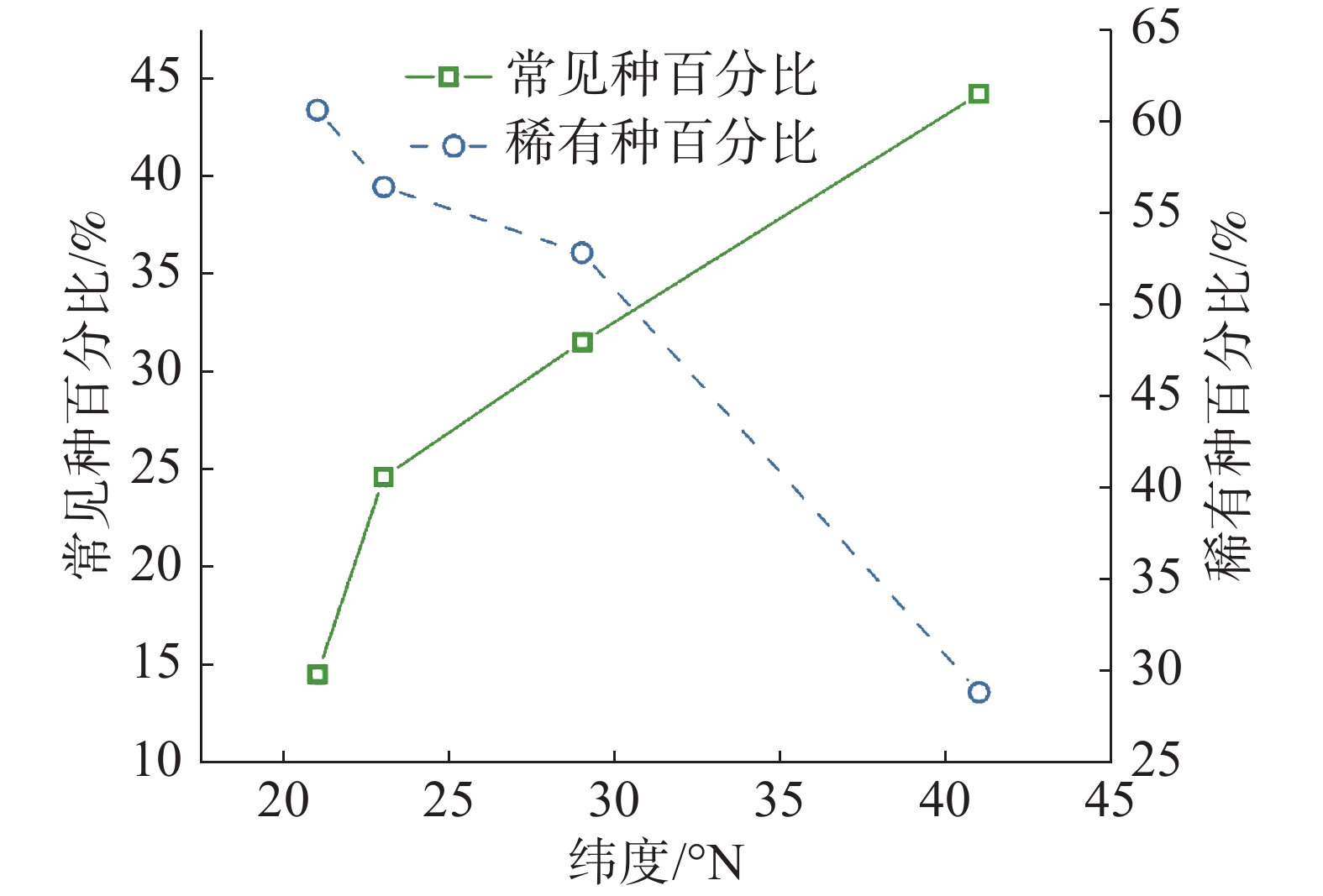
从统计结果可以看出(图3,表1),常见种物种数变化趋势与群落内物种总数变化趋势基本相符,即纬度越高,常见种物种数逐渐变少。但不同纬度群落中常见种种数相比较可以看出,随纬度从赤道向北增加,常见种占总树种数分别为 14.5%、24.6%、31.5%、44.2%,呈增长趋势。从稀有种变化情况可知,沿纬度梯度,稀有种数目变化趋势与群落内物种总数变化趋势一致,即纬度越高,稀有种数目变少;从不同纬度群落中稀有种种数相比较可看出,随纬度从赤道向北增加,稀有种占总树种数分别为 60.6%、56.4%、52.8%、28.8%,也呈减少趋势。
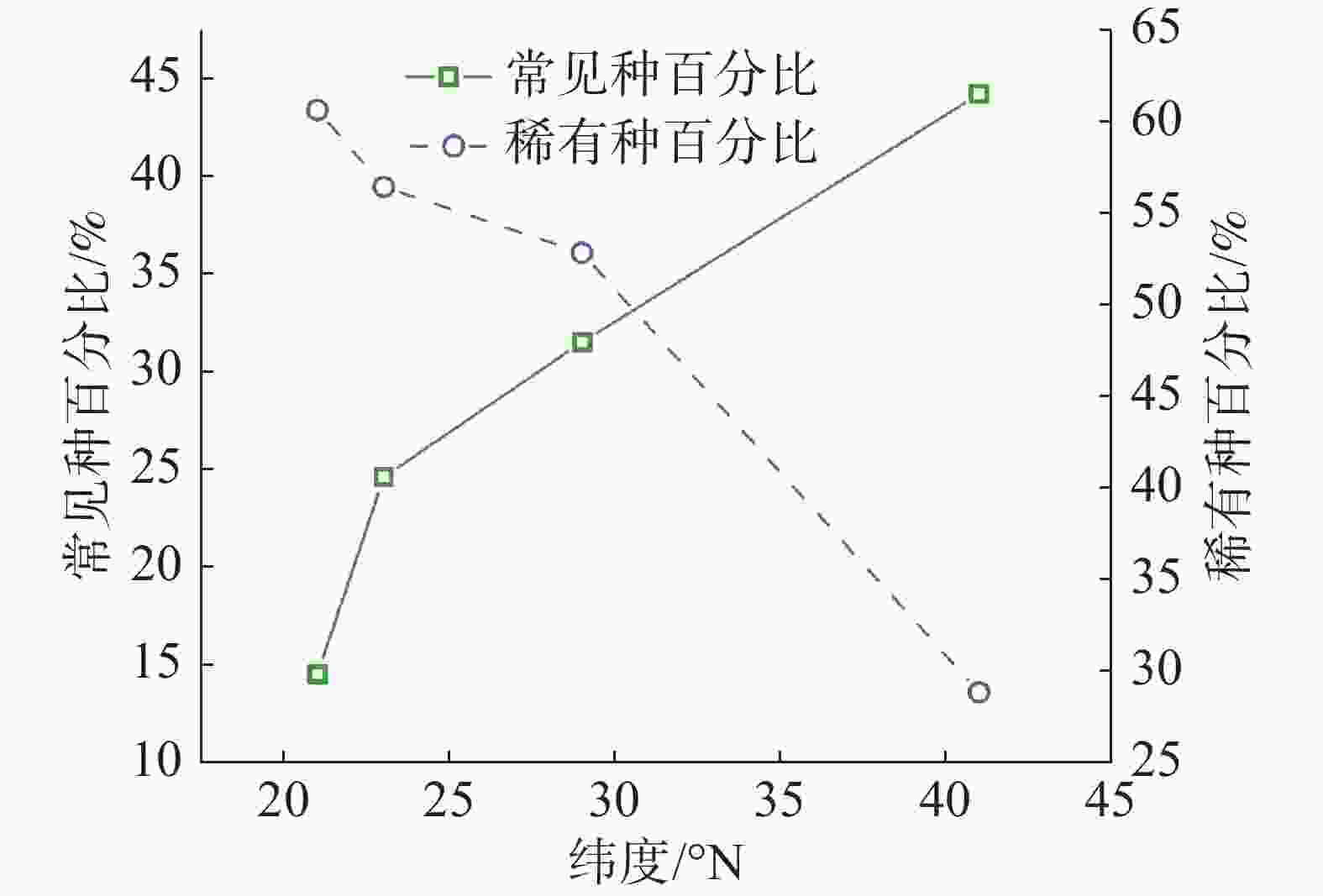
图 3 各群落中常见种和稀有种所占百分比沿纬度梯度变化情况
表 1 纬度梯度各群落树种数情况
样地 纬度/°N 物种总数/种 常见种/种 稀有种/种 CBS 41 52 23(44.23%) 15(18.85%) GTS 29 159 50(31.45%) 84(52.83%) DHS 23 195 48(30.19%) 110(56.41%) XSB 21 469 68(14.50%) 284(60.55%) 注:括号内为常见种和稀有种在总树种数中所占百分比。 -
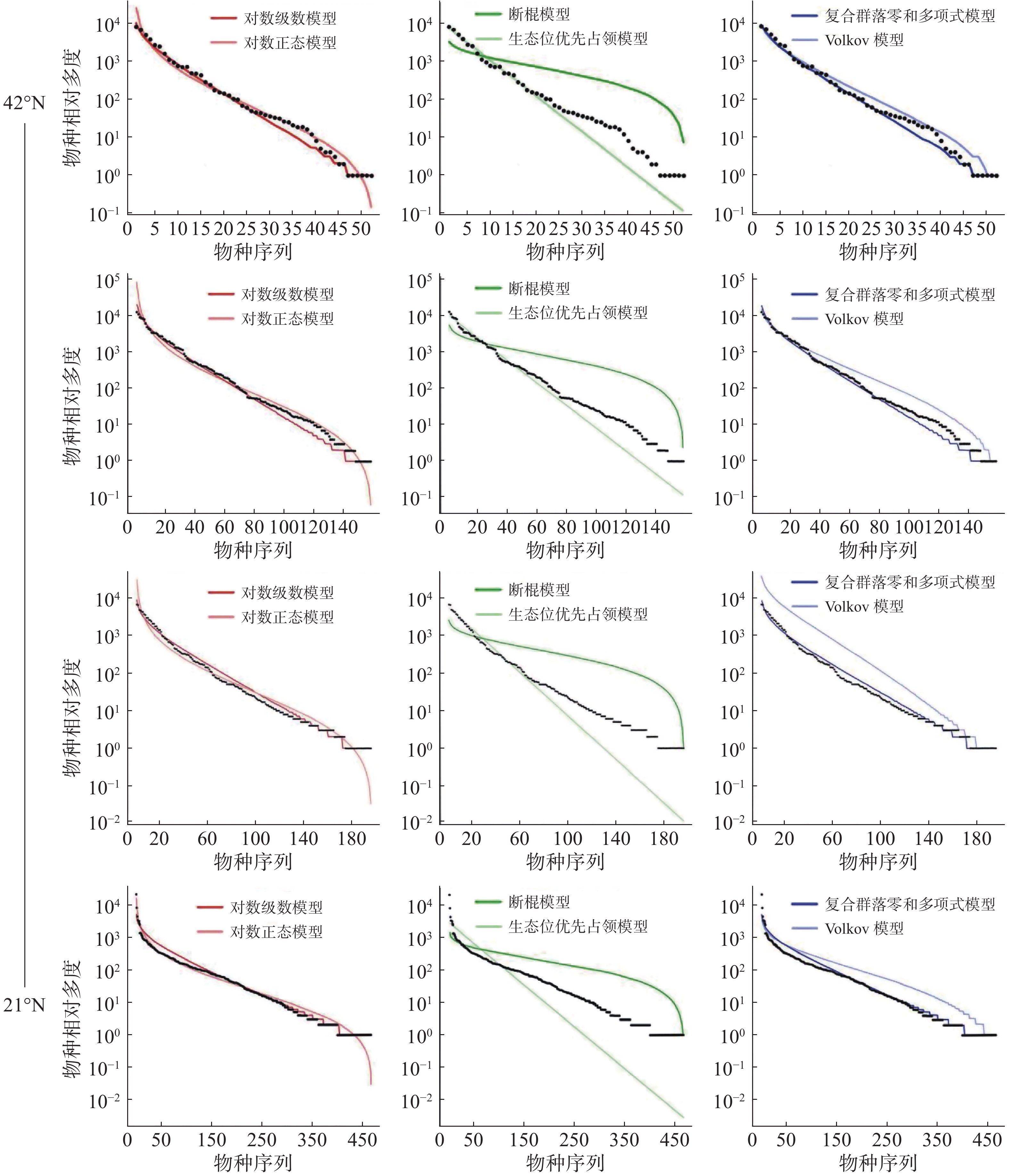
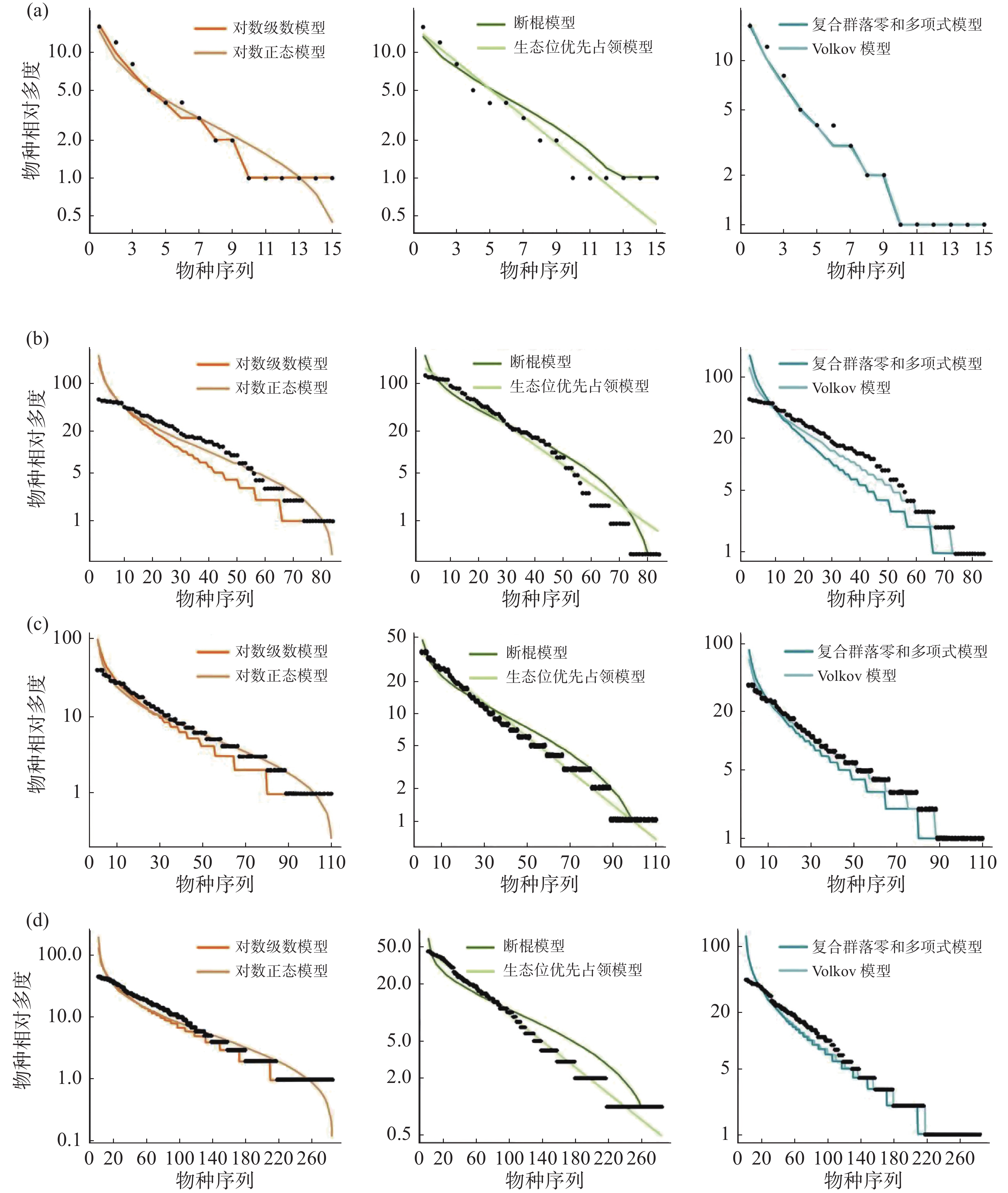
通过统计分析4个样地实测数据(表1)可发现,纬度从南至北,群落内物种数有逐渐减少的趋势,热带西双版纳森林群落中的物种数最多,有 469 种,物种数最少为温带长白山森林群落,只有 59 种,利用6个物种多度分布模型拟合结果表明(图4,表2~3):在较低纬度的西双版纳(21°N)和鼎湖山(24°N)群落中,生态位优先占领模型无法通过 K-S 检验(P < 0.05),西双版纳和鼎湖山群落的物种多度分布可由对数级数模型、对数正态模型、断棍模型、复合群落零和多项式模型和 Volkov 模型拟合,其中均显示出复合群落零和多项式模型和对数级数模型的拟合效果最优,西双版纳群落种多度模型拟合效果排序为:复合群落零和多项式模型 > 对数级数模型 > 对数正态模型 > Volkov 模型 > 断棍模型,鼎湖山群落种多度模型拟合效果排序为:复合群落零和多项式模型 > 对数级数模型 > Volkov 模型 > 对数正态模型 > 断棍模型。

图 4 纬度梯度整体群落物种多度分布模型拟合结果
表 2 纬度梯度上群落整体物种多度模型拟合优度检验结果
样地 检验方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 XSB AIC 4859.61 4921.56 5960.97 867769.40 4863.14 4976.28 P 0.99 1 0.93 <0.05 1 1 D 0.06 0.06 0.10 0.30 0.06 0.06 DHS AIC 2207.81 2247.37 2744.62 597060.90 2207.72 2246.64 P 0.98 1 0.60 <0.05 0.98 0.89 D 0.08 0.04 0.16 0.28 0.09 0.11 GTS AIC 2103.45 2124.18 2480.24 1030196.00 2103.15 2118.66 P 0.99 1 0.42 0.09 0.98 0.06 D 0.08 0.04 0.17 0.22 0.08 0.08 CBS AIC 665.50 676.00 791.37 187239.20 665.17 669.29 P 0.98 0.99 0.54 0.09 0.99 0.98 D 0.09 0.07 0.16 0.22 0.09 0.09 注:AIC 是赤池信息准则;D 是 K-S 检验,P为显著度,XSB表示西双版纳、DHS表示鼎湖山、GTS表示古田山、CBS表示长白山。 古田山(29°N)和长白山(41°N)群落中物种多度拟合结果显示所有模型都被接受(P > 0.05),种多度分布模型拟合优度排序为:复合群落零和多项式模型 > 对数级数模型 > Volkov 模型 > 对数正态模型 > 断棍模型 > 生态位优先占领模型,其中生态位优先占领模型的拟合效果依然最差。
沿纬度梯度,6个模型的拟合优度在各样地中的检验结果均显示中性理论模型对物种多度分布格局的拟合效果优于生态位理论模型,且其中复合群落零和多项式模型拟合结果均为最佳,除生态位优先占领模型之外,其他5种模型均可以拟合不同纬度下的物种多度分布,且拟合效果会随纬度上升变得更好。
表 3 整体物种种多度分布模型AIC值标准化
样地 标准化AIC 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 XSB AICs 0.00560 0.00567 0.00687 1 0.00560 0.00573 DHS AICs 0.00369 0.00376 0.00459 1 0.00369 0.00376 GTS AICs 0.00204 0.00206 0.00241 1 0.00204 0.00205 CBS AICs 0.00355 0.00361 0.00422 1 0.00355 0.00357 -
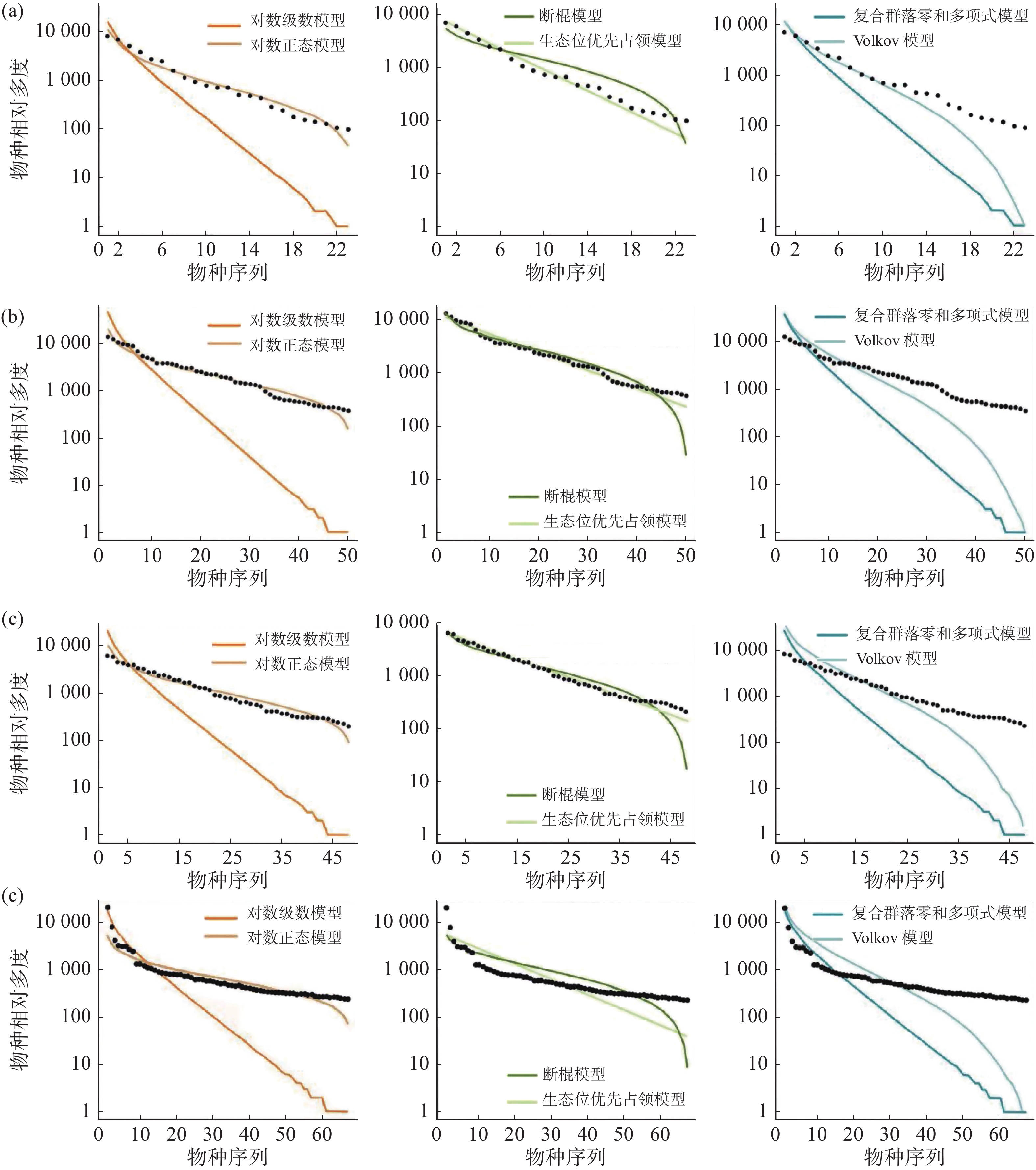
在常见种的拟合效果上(图5,表4),所有模型均被接受(P > 0.05),其中生态位理论模型中的生态位优先占领模型的拟合效果较差,所有群落中6个模型拟合效果排序均为:对数正态模型 > 断棍模型 > Volkov 模型 > 复合群落零和多项式模型 > 对数级数模型 > 生态位优先占领模型。
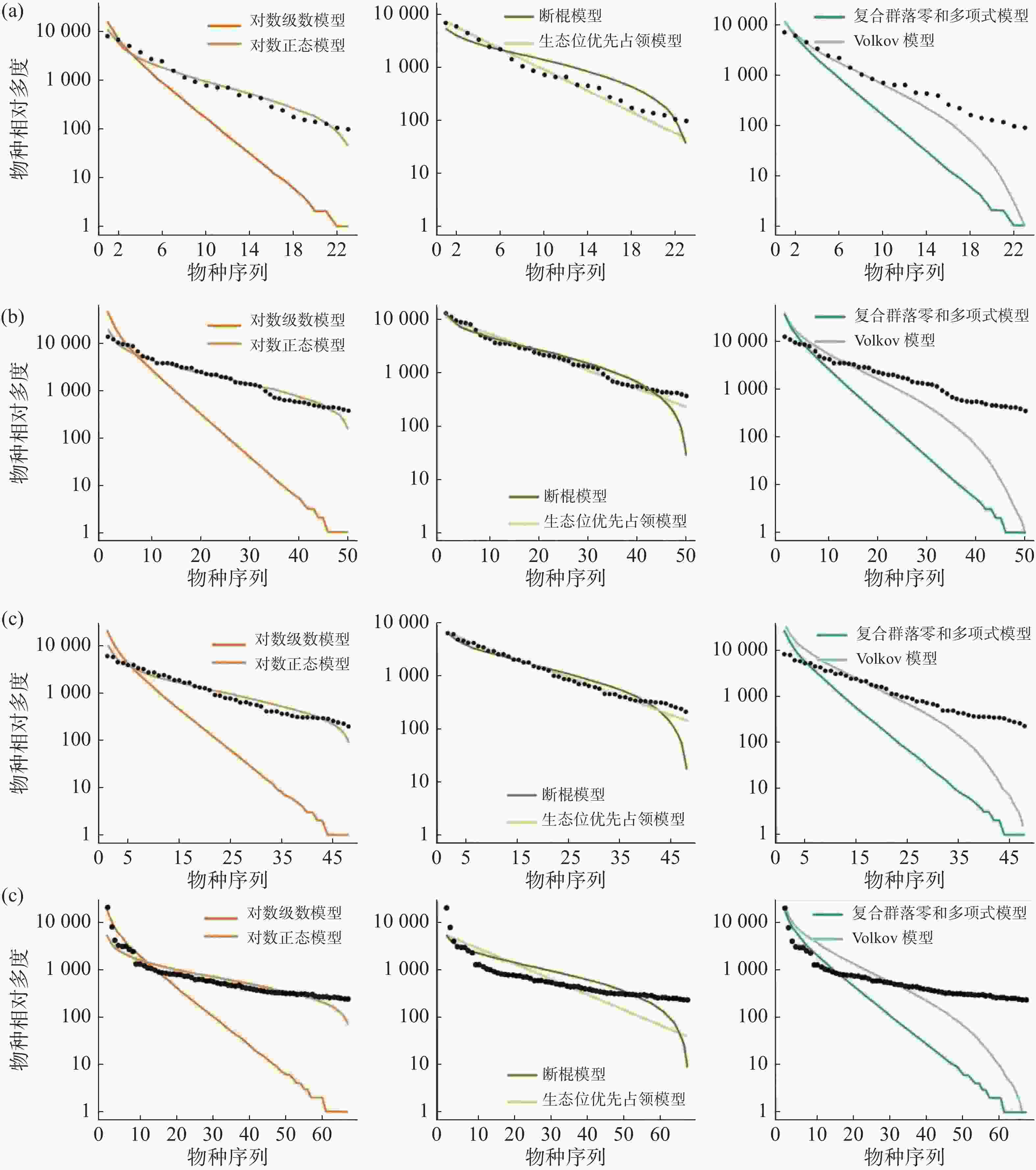
图 5 纬度梯度上群落中常见种物种多度分布拟合结果
表 4 纬度梯度上群落中常见种物种多度模型拟合优度检验
样地 检验方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AIC 410.09 382.11 385.70 177469.40 409.29 393.40 P 0.40 0.40 0.40 1 0.40 0.41 D 0.18 0.18 0.18 0.07 0.18 0.18 GTS AIC 982.17 886.67 889.97 930143.00 981.34 933.30 P 0.17 0.19 0.21 1 0.21 0.21 D 0.21 0.21 0.21 0.04 0.21 0.21 DHS AIC 888.03 800.41 803.18 520499.80 887.21 844.25 P 0.27 0.28 0.26 1 0.29 0.27 D 0.20 0.20 0.20 0.04 0.20 0.20 XSB AIC 1176.23 1039.42 1087.66 867769.40 1175.92 1116.54 P 0.24 0.22 0.24 0.893 0.25 0.24 D 0.20 0.20 0.20 0.09 0.20 0.20 通过标准化之后的 AIC 可更直观地发现(表5),在常见种的种多度分布拟合中,生态位理论模型中的断棍模型较中性理论模型拟合效果更优,且对数正态模型拟合效果优于对数级数模型,沿纬度从赤道向北增加,断棍模型对群落中常见种种多度分布格局的拟合效果均表现最好,除古田山群落(29°N)之外,断棍模型的拟合优度也呈逐渐增加趋势。
表 5 常见种模型拟合AIC值标准化
样地 标准化AIC 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AICs 0.00231 0.00215 0.00217 1 0.00231 0.00220 GTS AICs 0.00106 0.00095 0.00096 1 0.00106 0.00100 DHS AICs 0.00171 0.00154 0.00154 1 0.00171 0.00162 XSB AICs 0.00136 0.00120 0.00125 1 0.00136 0.00129 -
在稀有种的拟合效果上(图6,表6),所有模型均被接受(P > 0.05),生态位理论模型中的生态位优先占领模型拟合效果较差,西双版纳群落模型拟合效果排序为:复合群落零和多项式模型 > 对数级数模型 > Volkov 模型 > 对数正态模型 > 断棍模型 > 生态位优先占领模型;鼎湖山群落模型拟合效果排序为:Volkov 模型 > 复合群落零和多项式模型 > 对数级数模型 > 对数正态模型 > 断棍模型 > 生态位优先占领模型,古田山群落模型拟合效果排序为:对数级数模型 > Volkov 模型 > 断棍模型 > 复合群落零和多项式模型 > 对数正态模型 > 生态位优先占领模型,长白山群落模型拟合效果排序为:复合群落零和多项式模型 > 对数级数模型 > Volkov 模型 > 断棍模型 > 对数正态模型 > 生态位优先占领模型,总体上,中性理论模型对稀有种的拟合效果更佳,且对数级数模型拟合效果优于对数正态模型。通过标准化 AIC 也可看出(表7),沿纬度梯度,中性理论模型对群落中稀有种种多度分布的拟合效果均更好。
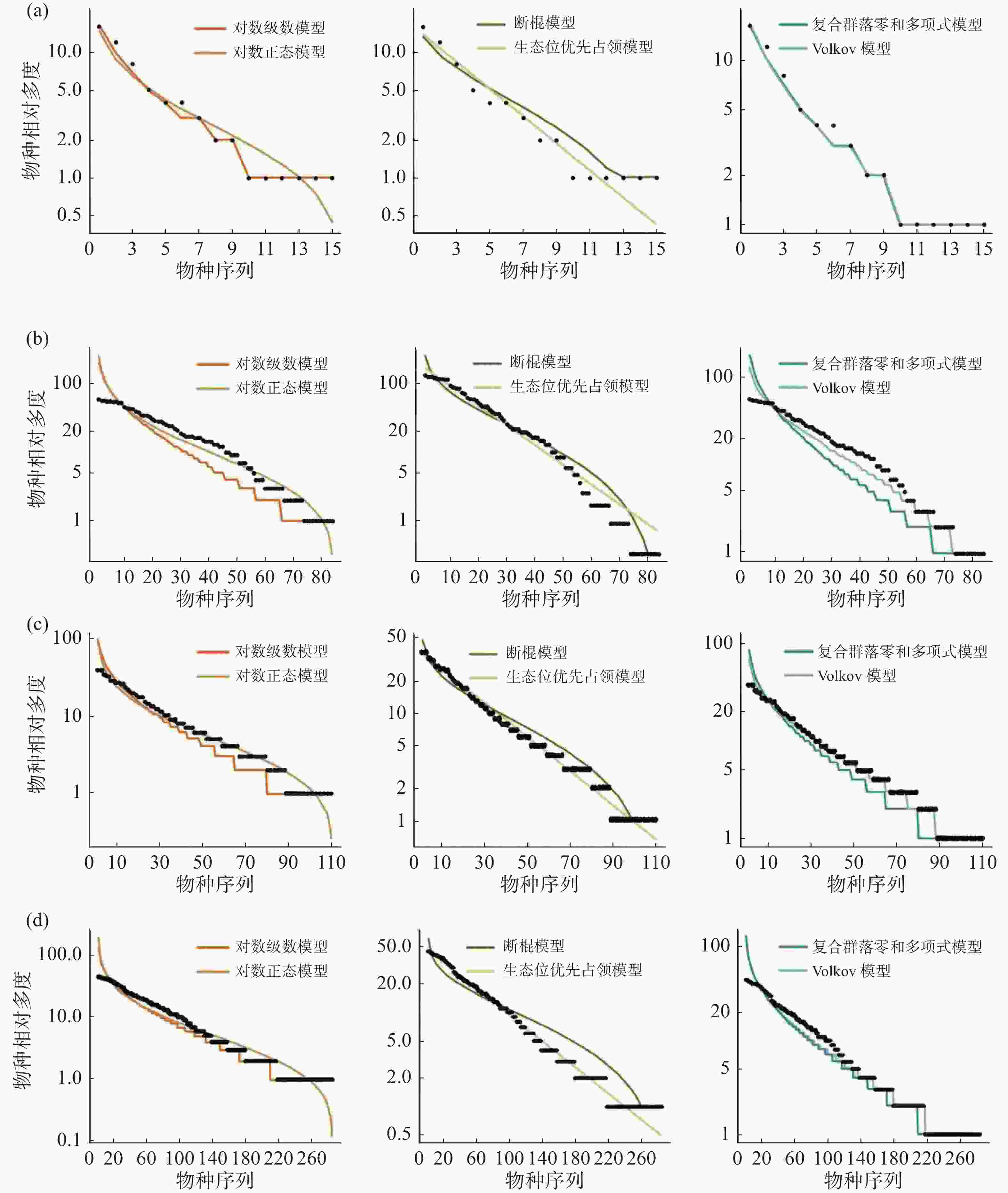
图 6 纬度梯度上群落中稀有种物种多度分布拟合结果
表 6 纬度梯度上群落中稀有种物种多度模型拟合优度检验
样地 检测方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AIC 68.46 73.21 72.77 284.13 68.24 70.44 P 0.99 0.99 0.99 0.54 0.99 0.99 D 0.11 0.11 0.11 0.19 0.11 0.11 GTS AIC 602.97 663.41 654.62 12115.59 662.39 649.12 P 0.99 1 1 0.99 1 0.97 D 0.08 0.04 0.08 0.08 0.08 0.13 DHS AIC 691.13 694.83 701.08 8259.98 690.72 683.22 P 0.97 0.99 1 0.83 0.97 1 D 0.13 0.08 0.08 0.14 0.13 0.08 XSB AIC 1794.92 1845.50 1887.33 29364.89 1794.54 1795.46 P 1 1 1 0.79 1 1 D 0.83 0.83 0.83 0.15 0.83 0.83 表 7 稀有种模型拟合AIC值标准化
样地 标准化
AIC对数级
数模型对数正态
模型断棍
模型生态位优先
占领模型复合群落零和
多项式模型Volkov
模型CBS AICs 0.24094 0.25766 0.25612 1 0.24017 0.24791 GTS AICs 0.04977 0.05476 0.05403 1 0.05467 0.05358 DHS AICs 0.00171 0.08412 0.08488 1 0.08362 0.08271 XSB AICs 0.06113 0.06285 0.06427 1 0.06111 0.06114 -
从热带跨越至温带,物种之间的相互作用,以及环境因子的影响都会使群落向不同的结构组成发展进化,使得植物群落的组成结构出现差异化。众多研究表明沿赤道向更高纬度发展,植物物种丰富度和多样性通常被认为呈逐渐下降趋势[40],在本研究结果中也出现了群落中的物种丰富度向赤道方向逐渐增加的相似变化趋势,在离赤道较近的低纬度西双版纳森林群落因为气候季节间和年际间变化较小,群落物种多样性较高,亦或因其群落进化时间较长,环境条件更加稳定,故物种丰富度也会更高;鼎湖山森林群落因纬度位置处于亚热带与热带过渡区[20],热带物种丰富,所以物种多样性会略高于同为亚热带群落的古田山植物群落,同样也因纬度与西双版纳森林群落较接近,具有一定数量的共有种,故群落之间的相异度较低;而纬度位置较高的长白山森林群落气候环境较特殊,自然选择主要受物理因素控制,且具有广适性的植物更容易生存,故群落内物种数下降且多样性较小。
关于物种多样性沿纬度梯度分布格局有很多假说试图解释[41],如“竞争假说”[42-43]、“环境异质性假说”[44]、“生态位假说”[45]等,其中众多假说中所涉及到的生态位分化理论和群落中性理论是目前从物种相似度的视角来阐述物种共存和物种多样性维持的两种主要理论,大部分的研究结果表明,在低纬度热带地区,群落中的物种多样性较高,无法通过生态位分化来形成相应的群落结构,故群落中物种多度的变化更倾向于受随机过程的可能性较大[46-48];沿纬度升高,群落中物种多样性较低纬度群落低,可供分配的资源充足,生态位分化得更明显。
-
物种多度模型拟合效果方面,在本研究纬度梯度上,模型拟合结果显示4个纬度梯度群落中均为中性理论模型拟合效果最佳,其次为对数级数模型。中性理论包括了2种物种相对多度的分布情况,一是在集合群落中,通过点突变形成新物种的对数级数分布;其二是局域群落中,因受扩散限制而随机产生新物种的零和多项式分布,且此分布模式的解释度在热带雨林群落中较高[49]。从此结果来看,零和多项式模型与其他众多模型相比,具有拟合曲线的尾部较长的特点,结合热带群落中物种多度情况,也能更好地说明该模型在低纬度的热带群落中拟合优度更高。
物种多度模型拟合的结果通常具有一定的尺度效应[50],不同尺度的群落中主导的生态学过程可能不同,故模型拟合的结果也会存在差异[51]。以往研究发现,随着研究群落样地尺度的增大,最优的拟合模型可能会发生变化,当选样尺度越小(40 m×40 m 或 60 m×60 m以下),生态位理论模型拟合效果越好[52],而中性理论模型在大尺度(100 m× 100 m)群落的物种多度格局的拟合方面往往会优于生态位模型,能更好地预测群落植物物种多度分布[53-54]。综上,也能更好地解释在此森林群落中中性理论模型拟合效果更优。在本研究中较高纬度的长白山群落物种多样性显著下降,但依然显示出复合群落零和多项式模型和对数级数模型拟合效果较好。
-
过往研究中,常见种与稀有种的多度分布格局同群落整体多度分布的模式一直存在争议:Vazquez 等[55-56]发现,常见种的多度分布通常与群落整体多度分布模式保持一致;但也有学者在研究中发现植物群落中的稀有种对整体物种多样性及多度的贡献度高于非稀有种[57]。本研究得出的结果与此相似,纬度沿赤道向北增加,常见种和稀有种种树同群落中总树种数的变化趋势一致,均为逐渐减少,但其各自占总树种数的百分比有所差异:稀有种所占百分比随纬度增大而减少,而常见种所占百分比却随纬度增大而增大;从模型拟合效果来看,低纬度至高纬度地区,在常见种群落中断棍模型均具有较好的拟合效果,而对稀有种物种多度的拟合则表现出中性理论模型效果更佳,且稀有种种多度分布最优拟合模型与群落整体种多度分布最优模型一致,均为中性理论模型。由此可说明,常见种种多度分布格局与群落中的资源分配有关,而稀有种的种多度分布格局形成过程更倾向于生态位等价下,具有随机性的物种生态漂变[58],且稀有种的种多度分布可能是群落整体物种多度分布格局形成的主导因素。
对于群落中的稀有种来说,不同纬度上中性理论模型拟合的效果有所差别。复合群落零和多项式模型对低纬度的西双版纳群落和较高纬度的长白山群落中稀有种的种多度分布格局拟合效果更好,而增加了密度限制因子的 Volkov 模型却对位于中等纬度的古田山森林群落和鼎湖山森林群落中稀有种种多度分布格局拟合更优。因为不同群落的分布格局往往会有不同的形成机制,如遗传漂变和物种迁移等基本生态过程[59]均会导致向中性理论模型分布,且在群落的不同的生活史阶段,生态位与中性过程的相对重要性是在不断变化的[60],同时也有研究发现,稀有种在群落中会具有较高的丧失率和迁移率[51]。基于此,可猜测在本研究中不同纬度群落中稀有种的种多度格局形成过程存在差异。
沿纬度上升,常见种多度分布模型拟合结果呈现出对数正态模型和断棍模型效果最优,虽然与群落整体物种多度分布结果不一致,但群落中常见种占群落总树种数百分比却呈增加趋势。一般来说,在群落中常见种会比稀有种具有更高的稳定性[61],在低纬度的群落中,物种多样性高,在资源分配方面,常见种个体在群落中往往占据的资源更多,故导致常见种种树数会低于稀有种;而在高纬度地区中的常见种因能适应严苛的环境,逐渐成为了纬度梯度上的特化种,在高纬度的环境中占据较高的生态位,故群落中物种种树会高于稀有种,且占比高于稀有种。故结合中性理论模型的拟合特点分析,在高纬度的群落中,起主导作用的生态学过程不能单从稀有种的种多度模型拟合效果看,关于常见种和稀有种在群落中物种多度分布方面的研究还亟待更深入的探究。
本研究对纬度21°~40°N范围内森林群落种多度分布格局进行研究,结果发现:纬度沿赤道至北,群落内物种数有逐渐减少的趋势,低纬度群落中植物物种数最多,高纬度群落物种数最少。且沿纬度梯度的未发现物种多度格局分布的明显差异,但不代表各群落中的生态过程相同,结果表明在各森林群落中种多度分布模型拟合优度检验结果均显示出中性理论模型对现阶段物种多度分布格局的拟合效果优于生态位理论模型,除生态位优先占领模型之外,其余模型均可以拟合不同纬度下的物种多度分布,且拟合效果随纬度上升变得更好;森林群落中常见种和稀有种物种数变化趋势与群落内物种总数变化趋势基本符合,即纬度越高,物种数逐渐变少,但常见种在总树种数中的占比呈增长趋势,断棍模型对群落中常见种种多度分布格局的拟合效果均表现最好;而稀有种数占总树种数比值呈逐渐减小趋势,中性理论模型对稀有种种多度分布的拟合效果均更好。
Latitudinal gradient changes in species rank and abundance of forest communities in tropical to temperate China
-
摘要: 为了深入了解森林群落物种多度分布格局在纬度梯度上的分布特征,以位于 21~42°N 纬度范围内的4个具有代表性的森林群落为研究对象,采用纯统计、中性理论和生态位理论三类物种多度分布模型进行拟合优度检验,结合群落多样性的变化特征进一步分析,结果表明:在此纬度梯度上,各森林群落中 α 多样性均随纬度升高逐渐减小;在种多度模型拟合方面并未显示出纬度梯度上的差异,结果均为中性理论模型中的复合群落零和多项式模型的拟合效果最优;随纬度上升,群落中常见种和稀有种物种数均逐渐减少,稀有种种多度分布拟合结果与整体群落种多度分布最优模型一致,而常见种种多度分布拟合结果则显示出生态位理论模型中的断棍模型效果更优。热带雨林群落中的稀有种比例较高,当受到干扰时更容易造成物种的丧失,从而具有更高的保护价值。Abstract: The study of species abundance in forest community can not only reveal the maintenance mechanism of community biodiversity, but also provide theoretical basis for biodiversity conservation. In order to have an in-depth understanding of the distribution characteristics of species abundance patterns in forest communities along the latitude gradient, four representative forest communities within the latitude range of 21°-42°N were selected. Species abundance distribution models based on pure statistics, neutral diversity theory and niche theory were adopted for fitting analysis, and further analysis was carried out in combination with the change characteristics of community diversity. The results showed that α diversity in each forest community decreased with the increase of latitude at this latitude gradient. In terms of species abundance model fitting, there was no difference in latitude gradient, and the fitting results showed that the metacommunity zero-sum multinomial distribution model in the neutral biodiversity theory had the best fitting effect. With the increase of latitude, the number of common and rare species in the community decreased gradually. The fitting results of rare species abundance distribution were consistent with the optimal community species abundance distribution model, while the fitting results of common species abundance distribution showed that the broken stick model in the niche theory had a better fitting effect. The proportion of rare species in tropical rainforest community is higher, which is more likely to cause species loss when disturbed, and thus has higher conservation value.
-
表 1 纬度梯度各群落树种数情况
样地 纬度/°N 物种总数/种 常见种/种 稀有种/种 CBS 41 52 23(44.23%) 15(18.85%) GTS 29 159 50(31.45%) 84(52.83%) DHS 23 195 48(30.19%) 110(56.41%) XSB 21 469 68(14.50%) 284(60.55%) 注:括号内为常见种和稀有种在总树种数中所占百分比。  下载: 导出CSV
下载: 导出CSV
表 2 纬度梯度上群落整体物种多度模型拟合优度检验结果
样地 检验方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 XSB AIC 4859.61 4921.56 5960.97 867769.40 4863.14 4976.28 P 0.99 1 0.93 <0.05 1 1 D 0.06 0.06 0.10 0.30 0.06 0.06 DHS AIC 2207.81 2247.37 2744.62 597060.90 2207.72 2246.64 P 0.98 1 0.60 <0.05 0.98 0.89 D 0.08 0.04 0.16 0.28 0.09 0.11 GTS AIC 2103.45 2124.18 2480.24 1030196.00 2103.15 2118.66 P 0.99 1 0.42 0.09 0.98 0.06 D 0.08 0.04 0.17 0.22 0.08 0.08 CBS AIC 665.50 676.00 791.37 187239.20 665.17 669.29 P 0.98 0.99 0.54 0.09 0.99 0.98 D 0.09 0.07 0.16 0.22 0.09 0.09 注:AIC 是赤池信息准则;D 是 K-S 检验,P为显著度,XSB表示西双版纳、DHS表示鼎湖山、GTS表示古田山、CBS表示长白山。
下载: 导出CSV
表 3 整体物种种多度分布模型AIC值标准化
样地 标准化AIC 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 XSB AICs 0.00560 0.00567 0.00687 1 0.00560 0.00573 DHS AICs 0.00369 0.00376 0.00459 1 0.00369 0.00376 GTS AICs 0.00204 0.00206 0.00241 1 0.00204 0.00205 CBS AICs 0.00355 0.00361 0.00422 1 0.00355 0.00357
下载: 导出CSV
表 4 纬度梯度上群落中常见种物种多度模型拟合优度检验
样地 检验方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AIC 410.09 382.11 385.70 177469.40 409.29 393.40 P 0.40 0.40 0.40 1 0.40 0.41 D 0.18 0.18 0.18 0.07 0.18 0.18 GTS AIC 982.17 886.67 889.97 930143.00 981.34 933.30 P 0.17 0.19 0.21 1 0.21 0.21 D 0.21 0.21 0.21 0.04 0.21 0.21 DHS AIC 888.03 800.41 803.18 520499.80 887.21 844.25 P 0.27 0.28 0.26 1 0.29 0.27 D 0.20 0.20 0.20 0.04 0.20 0.20 XSB AIC 1176.23 1039.42 1087.66 867769.40 1175.92 1116.54 P 0.24 0.22 0.24 0.893 0.25 0.24 D 0.20 0.20 0.20 0.09 0.20 0.20
下载: 导出CSV
表 5 常见种模型拟合AIC值标准化
样地 标准化AIC 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AICs 0.00231 0.00215 0.00217 1 0.00231 0.00220 GTS AICs 0.00106 0.00095 0.00096 1 0.00106 0.00100 DHS AICs 0.00171 0.00154 0.00154 1 0.00171 0.00162 XSB AICs 0.00136 0.00120 0.00125 1 0.00136 0.00129
下载: 导出CSV
表 6 纬度梯度上群落中稀有种物种多度模型拟合优度检验
样地 检测方法 对数级数模型 对数正态模型 断棍模型 生态位优先占领模型 复合群落零和多项式模型 Volkov模型 CBS AIC 68.46 73.21 72.77 284.13 68.24 70.44 P 0.99 0.99 0.99 0.54 0.99 0.99 D 0.11 0.11 0.11 0.19 0.11 0.11 GTS AIC 602.97 663.41 654.62 12115.59 662.39 649.12 P 0.99 1 1 0.99 1 0.97 D 0.08 0.04 0.08 0.08 0.08 0.13 DHS AIC 691.13 694.83 701.08 8259.98 690.72 683.22 P 0.97 0.99 1 0.83 0.97 1 D 0.13 0.08 0.08 0.14 0.13 0.08 XSB AIC 1794.92 1845.50 1887.33 29364.89 1794.54 1795.46 P 1 1 1 0.79 1 1 D 0.83 0.83 0.83 0.15 0.83 0.83
下载: 导出CSV
表 7 稀有种模型拟合AIC值标准化
样地 标准化
AIC对数级
数模型对数正态
模型断棍
模型生态位优先
占领模型复合群落零和
多项式模型Volkov
模型CBS AICs 0.24094 0.25766 0.25612 1 0.24017 0.24791 GTS AICs 0.04977 0.05476 0.05403 1 0.05467 0.05358 DHS AICs 0.00171 0.08412 0.08488 1 0.08362 0.08271 XSB AICs 0.06113 0.06285 0.06427 1 0.06111 0.06114
下载: 导出CSV
-
[1] ZHANG Y Z, MA J, LIANG S L, et al. A stacking ensemble algorithm for improving the biases of forest aboveground biomass estimations from multiple remotely sensed datasets [J]. GIScience & Remote Sensing, 2022, 59(1): 234 − 249. [2] EBRU C F, SIRIN U. The changes related with altitudinal gradients and seasonal variation in the species composition of Carabidae (Coleoptera) in Turkmen Mountain (Eskisehir, Turkey) [J]. Ekoloji., 2016, 25(98): 17 − 24. [3] 李林, 魏识广, 练琚愉, 等. 亚热带不同纬度植物群落物种多样性分布规律[J]. 生态学报, 2020, 40(4): 1249 − 1257. [4] HUSTON M. Biological diversity: the coexistence of species on changing landscape [J]. Journal of the Marine Biological Association of the United Kingdom, 1994, 75(1): 261 − 261. [5] COATES M. A comparison of intertidal assemblages on exposed and sheltered tropical and temperate rocky shores [J]. Global Ecology Biogeography., 1998, 7(2): 115 − 125. doi: 10.2307/2997814 [6] UYSAL H, UNVER S, KIZILET H. The effects of neonicotinoids on the longevity of the male and female populations of Drosophila melanogaster [J]. Ekoloji., 2015, 24(96): 57 − 63. [7] GASTON K J. Global patterns in biodiversity [J]. Nature, 2000(405): 220 − 227. [8] RICKLEFS R E, HE F. Region effects influence local tree species diversity [J]. Proceedings of the National Academy of Sciences of the United States of America, 2016(113): 674 − 679. [9] KINLOCK N L, PROWANT L, HERSTOFF E M, et al. Explaining global variation in the latitudinal diversity gradients: meta-analysis confirms known patterns and uncovers new ones [J]. Global Ecology Biogeography, 2018(27): 125 − 141. [10] VOLKOV I, BANAVAR J R, HE F, et al. Density dependence explains tree species abundance and diversity in tropical forests [J]. Nature, 2005(438): 658 − 661. [11] HUBBELL S P. Neutral theory and the evolution of ecological equivalence [J]. Ecology, 2006(87): 1387 − 1398. [12] ROSINDELL J, CORNELL S J. Species-area curves, neutral models, and long-distance dispersal [J]. Ecology, 2009(90): 1743 − 1750. [13] FURNISS T J, LARSON A J, LUTZ J A. Reconciling niches and neutrality in a subalpine temperate forest [J]. Ecosphere, 2017, 8(6): e01847. [14] 姜隽. 生物群落的物种多样性研究综述[J]. 科技情报开发与经济, 2009, 19(27): 131 − 133. [15] 李巧, 涂璟, 熊忠平, 等. 物种多度格局研究概况物种多度格局研究概况[J]. 云南农业大学学报, 2011, 26(1): 117 − 123. [16] MI Z. Climatic dynamics of broadleaved Korean pine forest in Changbai Mountain during the last 22 Years [J]. Chinese Journal of Ecology, 2005(24): 1007 − 1012. [17] 张姗, 蔺菲, 原作强, 等. 长白山阔叶红松林草本层物种多度分布格局及其季节动态[J]. 生物多样性, 2015, 23(5): 641 − 648. doi: 10.17520/biods.2015089 [18] 胡正华, 于明坚, 丁炳扬, 等. 古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究[J]. 应用与环境生物学报, 2003(4): 341 − 345. doi: 10.3321/j.issn:1006-687X.2003.04.003 [19] 祝燕, 赵谷风, 张俪文, 等. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构[J]. 植物生态学报, 2008, 32(2): 262 − 273. doi: 10.3773/j.issn.1005-264x.2008.02.004 [20] 叶万辉, 曹洪麟, 黄忠良, 等. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J]. 植物生态学报, 2007, 32(2): 274 − 286. [21] CAO M, ZOU X M, WANG M, et al. Tropical forests of Xishuangbanna, China [J]. Biotropica, 2006(38): 306 − 309. [22] ZHU H. Forest vegetation of Xishuangbanna, south China [J]. Forestry Studies in China, 2006(2): 1 − 58. [23] 兰国玉, 胡跃华, 曹敏, 等. 西双版纳热带森林动态监测样地——树种组成与空间分布格局[J]. 植物生态学报, 2008, 32(2): 287 − 298. doi: 10.3773/j.issn.1005-264x.2008.02.006 [24] 兰国玉, 朱华, 曹敏. 西双版纳热带季节雨林树种的区系组成成分分析[J]. 西北林学院学报, 2013, 28(1): 33 − 38. doi: 10.3969/j.issn.1001-7461.2013.01.06 [25] CONDIT R. The CTFS and the Standardization of Methodology [M]. Berlin: Springer-Verlag, 1998. [26] 王世雄, 赵亮, 李娜, 等. 稀有种和常见种对植物群落物种丰富度格局的相对贡献[J]. 生物多样性, 2016, 24(6): 658 − 664. doi: 10.17520/biods.2015239 [27] 王世雄, 王孝安, 郭华, 等. 稀有种和常见种对黄土高原辽东栎群落物种多样性贡献的多尺度分析[J]. 生态学报, 2018, 38(22): 8060 − 8069. [28] 张金屯. 植被数量生态学方法 [M]. 北京: 中国科技出版社, 1995. [29] 谢晋阳, 陈灵芝. 中国暖温带若干灌丛群落多样性问题的研究[J]. 植物生态学报, 1997, 21(3): 197 − 207. doi: 10.3321/j.issn:1005-264X.1997.03.001 [30] BRAY J R, CURTIS J T. An ordination of upland forest communities of southern Wisconsin [J]. Ecological Monographs, 1957, 27: 325 − 349. doi: 10.2307/1942268 [31] FISHER R A, CORBET A S, WILLIAMS C B. The relation between the number of species and the number of individual in a random sample of an animal population [J]. Journal of Animal Ecology, 1943, 12(1): 42 − 58. doi: 10.2307/1411 [32] PRESTON F W. The canonical distribution of commonness and Rarity: Part I [J]. Ecology, 1962, 43(2): 185 − 215. doi: 10.2307/1931976 [33] MOTOMARA I. A statistical treatment of ecological communities [J]. Zoological Magazine, 1932, 44: 379 − 383. [34] MACARTHUR R H. On the relative abundance of bird species [J]. Proceedings of the National Academy of Sciences, 1957, 43(3): 293 − 295. doi: 10.1073/pnas.43.3.293 [35] ALONSO D, MCKANE A J. Sampling Hubbell's neutral theory of biodiversity [J]. Ecology Letters, 2004, 7(10): 901 − 910. doi: 10.1111/j.1461-0248.2004.00640.x [36] VOLKOV I, BANAVAR J R, HUBBELL S P, et al. Neutral theory and relative species abundance in ecology [J]. Nature, 2003, 424(6952): 1035 − 1037. doi: 10.1038/nature01883 [37] MCGILL B J. A test of the unified neutral theory of biodiversity [J]. Nature, 2003, 422: 881 − 885. doi: 10.1038/nature01583 [38] 高利霞, 毕润成, 闫明. 山西霍山油松林的物种多度分布格局[J]. 植物生态学报, 2011, 35(12): 1256 − 1270. [39] SEKHON J S. Multivariate and Propensity Score Matching Software with Automated Balance Optimization: The Matching package for R [J]. Journal of Statistical Software, 2011, 42(7): 1 − 52. [40] SALAZAR L, HOMEIER J, KESSLER M, et al. Diversity patterns of ferns along elevational gradients in Andean tropical forests [J]. Plant Ecology and Diversity, 2015(8): 13 − 24. [41] CHU C J, LUTZ J A, KRAL K, et al. Direct and indirect effects of climate on richness drive the latitudinal diversity gradients in forest trees [J]. Ecology Letters, 2019(22): 245 − 255. [42] PIANKA E R. Latitudinal gradients in species diversity: a review of concepts [J]. The American Naturalist, 1966, 100: 33 − 46. doi: 10.1086/282398 [43] SCHEMSKE D W, MITTELBACH G G. “Latitudinal gradients in species diversity”: reflections on Pianka’s 1966 article and a look forward [J]. The American Naturalist, 2017, 189: 599 − 603. doi: 10.1086/691719 [44] STEIN A, GERSTNER K, KREFT H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales [J]. Ecology Letters, 2014, 17: 866 − 880. doi: 10.1111/ele.12277 [45] QIAN H, JIN Y, RICKLEFS R E. Patterns of phylogenetic relatedness of angiosperm woody plants across biomes and life-history stages [J]. Biogeography, 2017, 44: 1383 − 1392. doi: 10.1111/jbi.12936 [46] HUBBELL S P, FOSTER R B. Tropical rain forest: ecology and management [M]. London: The British Ecological Society, 1983: 25-41. [47] HUBBELL S P. Neutral theory in community ecology and the hypothesis of functional equivalence. Functional [J]. Ecology, 2005, 19(1): 166 − 172. [48] HUBBELL S P. The neutral theory of biodiversity and biogeography and Stephen Jay Gould [J]. Paleobiology, 2005, 31(5): 122 − 132. [49] 周淑荣, 张大勇. 群落生态学的中性理论[J]. 植物生态学报, 2006, 30(5): 868 − 877. doi: 10.3321/j.issn:1005-264X.2006.05.018 [50] LINDO Z, WINCHESTER N. Scale dependent diversity patterns in arboreal and terrestrial oribatid mite (Acari: Oribatida) communities [J]. Ecography, 2008, 31(1): 53 − 60. doi: 10.1111/j.2007.0906-7590.05320.x [51] HARPOLE W S, TILMAN D. Non-neutral patterns of species abundance in grassland communities[J]. Ecology Letters, 2006(9): 15-23. [52] 陈俊, 艾训儒, 姚兰, 等. 木林子天然次生林典型群落物种多度分布格局的尺度效应[J]. 湖北民族学院学报 (自然科学版), 2018, 36(2): 130 − 133. [53] 程佳佳, 米湘成, 马克平, 等. 亚热带常绿阔叶林群落物种多度分布格局对取样尺度的响应[J]. 生物多样性, 2011, 19(2): 168 − 177. [54] FISHEER C K, MEHT P. The transition between the niche and neutral regimes in ecology [J]. Proceedings of the National Academy of Sciences, 2014, 111(36): 13111 − 13116. doi: 10.1073/pnas.1405637111 [55] VAZQUEZ L B, GASTON K J. Rarity, commonness, and patterns of species richness: the mammals of Mexico [J]. Global Ecology and Biogeography, 2004, 13: 535 − 542. doi: 10.1111/j.1466-822X.2004.00126.x [56] PEARMAN P B, WEBER D. Common species determine richness patterns in biodiversity indicator taxa [J]. Biological Conservation, 2007, 138(1): 109 − 119. [57] 刘旻霞, 李全弟, 蒋晓轩,等. 甘南亚高寒草甸稀有种对物种多样性和物种多度分布格局的贡献[J]. 生物多样性, 2020, 28(2): 107 − 116. doi: 10.17520/biods.2019297 [58] VELLEND M. Conceptual synthesis in community ecology [J]. Quarterly Review of Biology, 2010, 85(2): 183 − 206. doi: 10.1086/652373 [59] 刘金亮, 于明坚. 片段化森林群落构建的生态过程及其检验方法[J]. 植物生态学报, 2019, 43(11): 929 − 945. doi: 10.17521/cjpe.2019.0155 [60] HU Y H, SHA L Q, BLANCHET F G, et al. Dominant species and dispersal limitation regulate tree species distributions in a 20-ha plot in Xishuangbanna, southwest China [J]. Oikos, 2012, 121(6): 952 − 960. doi: 10.1111/j.1600-0706.2011.19831.x [61] MOUILLOT D, LEPRETRE A, ANDREI-RUIZ M C, et al. The fractal model: a new model to describe the species accumulation process and relative abundance distribution (RAD) [J]. Oikos, 2000, 90(2): 333 − 342. doi: 10.1034/j.1600-0706.2000.900214.x -

 点击查看大图
点击查看大图
图(6) / 表 (7)
计量
- 文章访问数: 575
- HTML全文浏览量: 203
- PDF下载量: 19
- 被引次数: 0











