





-
叶面积指数(Leaf area index,LAI,记作ILA )是草地生态系统重要的状态变量[1- 2],它又称叶面积系数,指单位草地面积上植物叶面积的总和,通常用于描述植被覆盖的密度和茂密程度,数值越大代表植被覆盖越密集[3]。它是反映草地生长状态的重要参数,其变化能够直接或者间接揭示牧草在不同放牧方式下的变化规律。LAI也用来描述植被的生长状况和生态功能,草地的许多生物过程都受到叶面积指数的影响,草地叶面积指数能够直接反映草地生长状况和生产力水平,并且能够指示草地生长状况[4]。因此,叶面积指数常用于研究草地群落组成与生态系统之间的反馈机制过程[5],以及在小空间尺度上衡量冠层生物量和异质性。在多草种混合草地草原上,LAI经常被用来监测碳平衡和评价草的质量和生产力[6-8]。近几十年来,无人机结合植被指数的经验模型法使其在草地遥感监测方面成为一项极具前途的技术[9]。经验模型法认为叶面积指数与植被指数之间存在函数关系,根据此种函数关系可以进行叶面积指数反演。二十世纪七十年代,Bunnik利用遥感技术成功提取出叶面积指数,奠定了遥感影像反演LAI的基础[10]。无人机在图像质量、传感效率和运营成本之间都可以实现适当的平衡。各级草地管理单位及研究人员建立了众多的草地状况监测站点,结合遥感或无人机技术,能够提供大范围且精确的草地状况基础数据[11]。基于主动多光谱冠层传感器,Li等[12]提出一种利用无人机主动传感估算水稻LAI的新方法,相较于辐射传输模型的不确定性及辐射信号携带的有限信息,无人机所制备的植被指数是根据光谱信号的目标信息计算出来的,更加能够准确地反映出短时间内草地受到放牧干扰的生长情况。
放牧方式直接或者间接干扰牧草生长,代表性的放牧方式有轮牧和连续放牧。适当的轮牧充分利用牧草的生物学特性,降低了牲畜的选择性采食行为与践踏对草地的干扰强度,使草地的生产力在整个生产过程能够达到最大化[13-14]。然而,过度轮牧会造成地表被过度践踏而使草地裸露并导致裸露地表破碎化,改变土壤的物理性状,从而抑制草地植物的生长并降低草地物种多样性,改变草地生态系统原有的结构和功能[15-16]。因此,过度集约利用草原是许多植物物种消失的主要原因。而中等强度轮牧可以有效地缓和由于牲畜啃食造成的草地叶片减少而引起的光合作用减少,缓解草地有机物的急骤减少[17],促进草地牧草的生长,草地叶面积指数反而因放牧升高,这表明在适当强度的轮牧干扰下,有利于维持草地叶面积处于较高状态,草地生物的多样性和丰富度得以维持甚至有所增加。轻度的连续放牧由于食物来源的限制,在某一特定区域有序活动,可以加快草地无效掉落物的分解,增加土壤氮和有机物的含量,减少土壤硝酸盐的可提取磷进而降低土壤污染[18],从而促进草地牧草生长。彭祺等[19]却认为连续放牧是一种无计划、粗放的经营方式,过度地连续放牧会导致草地生物多样性和稳定性降低,牲畜的过度踩踏效应会使土壤压缩从而导致草地生产力降低,质量下降,甚至引起草地不可逆的退化[20],但是连续放牧并不意味着家畜对个体的连续无休止采食,通过轻度或者中度连续放牧也能使草层保持较高的有机物积累水平,刺激并促进草地的生长,维持草地的稳定[21],适当地将食草动物引入草原能对植物群落的富集和多样性有积极影响,有助于保持和增加其植物多样性,使草地叶面积指数保持稳定。热带地区不同放牧方式与放牧强度下,叶面积指数的动态变化机制是怎样的,值得深入研究。
本研究按照不同放牧方式(轮牧和连续放牧)、不同放牧强度(重度、中度和轻度)的放牧策略进行放牧试验,利用多光谱无人机对放牧期间的草地LAI按日尺度进行监测,获得每日放牧前和放牧后的多光谱影像,分析不同放牧策略对草地LAI的影响,揭示不同放牧策略对草地叶面积指数影响的变化规律,在微观层面优选出抑制草地退化适应性管理策略。
-
放牧实验区位于海南省文昌市锦山镇锦华村,草地主要由地毯草和狗牙根组成,该地区位于海南岛东北部,气候类型属热带海岛季风气候,每年11月到次年4月为干季,全年平均温度在22.5~25.6 ℃之间,年均降雨量在900~2 500 mm之间[22],该地区温暖潮湿的气候和较高的降雨量,为畜牧业提供了充足的水源和气候条件。经实地走访调查和村委会提供的数据,发现村内家庭牧场普遍采取散养的放牧模式,散养牛的总数量为212头。并且该村土地撂荒情况严重,位于锦华村西部的大片水田无人管理,出现水田和草地无序放牧的情况。
-
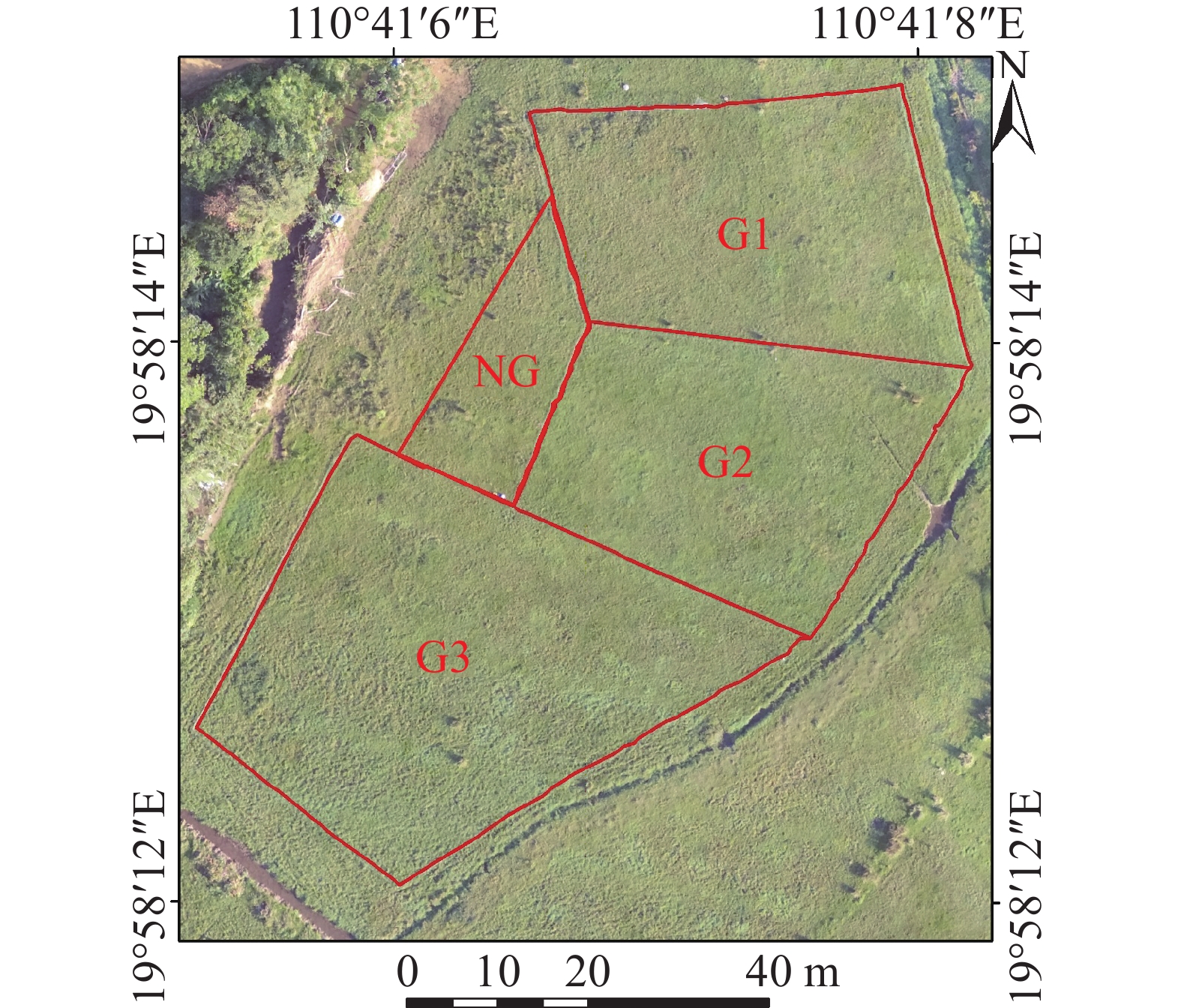
按照不同放牧方式(轮牧和连续放牧)、不同放牧强度(重度、中度和轻度)的管理方式进行试验(表1)。放牧强度通过调节样地牛头数来控制,利用多光谱无人机对放牧期间的草地LAI按日尺度进行监测。实验草地面积为0.39 hm2,分成3个放牧样地:G1、G2和G3,面积分别为0.11、0.11和0.17 hm2,在3个草地的周围再选取1个样地作为不放牧草地NG,以便进行对照,将G1作为轮牧的第一块样地,G2作为轮牧的第二块样地,按照时间顺序依次进行5 d的放牧,G3作为连续放牧的样地,进行10 d的放牧,放牧时间为每日8∶30—17∶30,其余时间将牛赶回牛棚,同一个时间段保证放牧强度相同,放牧强度的设定参考Costa等[23]对热带牧场放牧强度的分析和当地25头·hm−2的生产实际,总共进行3次放牧试验(图1)。
表 1 放牧实验设计
放牧强度 放牧日期 轮牧G1、
G2/头连续放
牧G3/头载畜量/
(头·hm−2)重度放牧 2022-11-10—11-19 9 7 41 中度放牧 2022-12-9—12-18 5 4 23 轻度放牧 2022-12-20—12-29 3 2 13 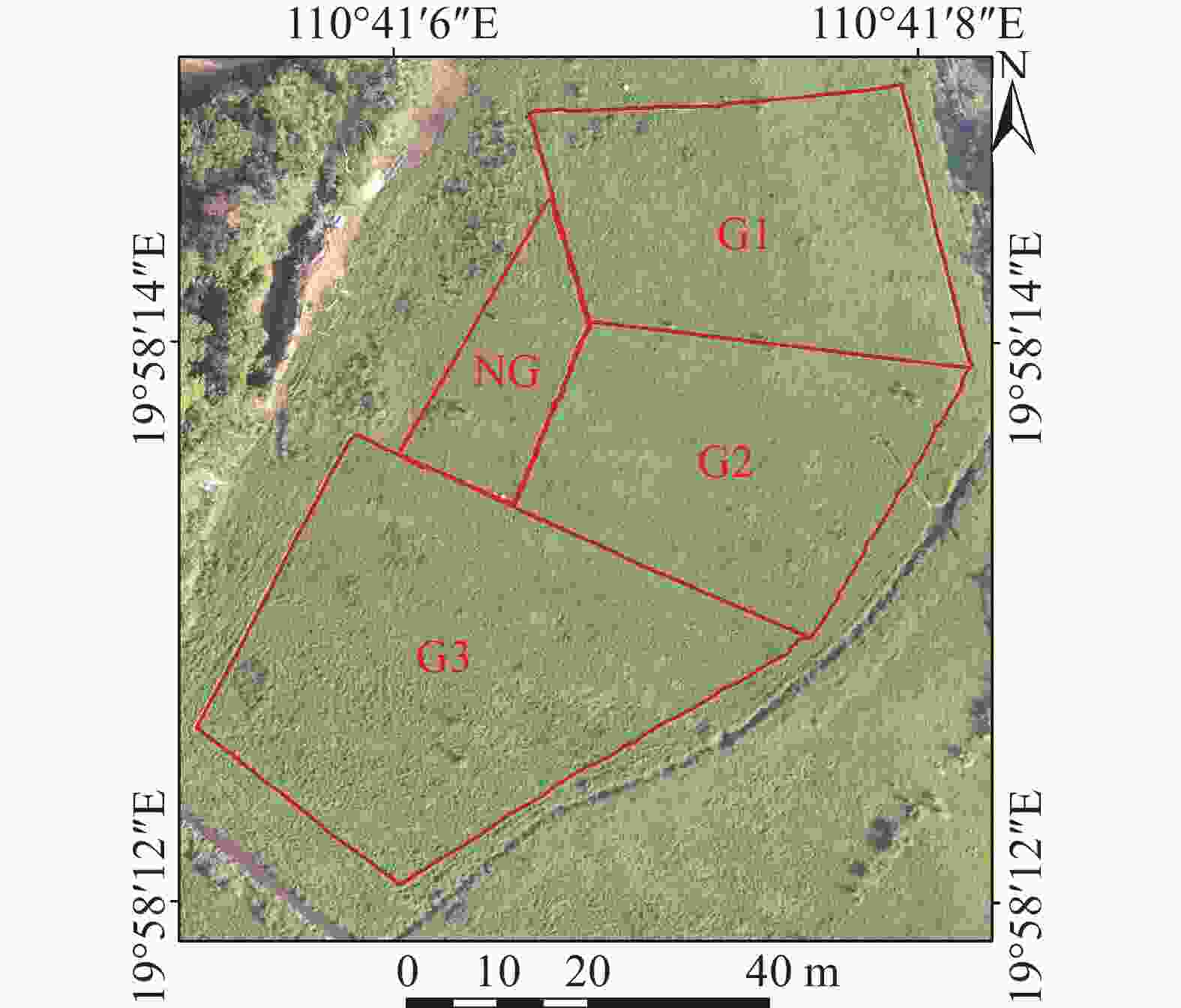
图 1 放牧实验设计图
-
采用LAI2200-C植被冠层分析仪观测LAI。LAI2200-C是一种常用于测量植被叶面积指数的仪器,其工作原理是基于直接测量植被冠层中的光线透过程度,该仪器内部还装有全球定位系统(GPS)模组。在稳定的天空条件下,对研究区的草地按照对角线的方式选择两条线路做好标记,以1.5 m的等距离测量,进行测量时使用270°视野遮盖帽,将传感器放在植被冠层上方,所得到的天空亮度记为A值,然后在冠层下方读取4个B值。测量完成后,LAI2200-C会自动将测量结果保存在其内部存储卡中,将仪器与计算机连接来读取、导出测量结果,利用LAI2200-C方法计算ILA,计算公式为:
$$ I_{LA}=2\sum _{i=1}^{5}\overline {{K}}_{i}{W}_{i} ,$$ (1) 式中,ILA是叶片(所有遮光体)面积指数,
$\overline {{K}}_{i} $ 是i环的平均接触频率,Wi是i环的权重因子。平均接触频率是透射率和路径长度的函数,可以通过n个Above/Below的观测值计算获得:
$$ {\overline K _i} = \dfrac{{\dfrac{1}{n} \displaystyle \sum\limits_{i = 1}^n { - {\text{ln}}\left( {\dfrac{{{B_i}}}{A}} \right)} }}{{{S_i}}}。 $$ (2) $$ {W}_{i}=\Delta {\theta }_{i}\mathrm{sin}\overline {{\theta }_{i}}, $$ (3) 权重因子Wi由公式(3)计算得出,其中:
$ \overline {{\theta }_{i}} $ 是平均天顶角,$ \Delta {\theta }_{i} $ 是与第$ i $ 环相关的环宽度(弧度)。$ \Delta {\theta }_{i} $ 的值由表2给出。权重因子被标准化为总和等于1,所以当一个或多个环被遮盖,其余环的权重因子会增大。表 2 环宽度和权重因子
环 宽度/(°) 权重因子 1 12.2 −0.041 2 12.2 0.131 3 11.8 0.201 4 13.2 0.290 5 13.2 0.337 -
本研究利用无人机对不同放牧策略下的草地叶面积指数按日尺度进行监测。所用的DJI精灵4多光谱版无人机,可以拍摄5个波段的图像:蓝光、绿光、红光、红边和近红外,提供高精度的定位信息。本试验利用无人机划定飞行区域,设置93.9 m的飞行高度,飞行速度14.6 m·s−1,等时间间隔2.2 s拍照,每日在早上牛入栏前和下午牛出栏后进行飞行,以保证获得草地5 cm分辨率的可见光成像(JPEG格式)和多光谱成像(TIFF)。
-
采用目标取样法在轮牧草地和连续放牧草地中分别选择具有健康状况良好且具有代表性的一头牛,人工目视全天跟踪观测。图2为牛在各种行为下的样本图像,利用连续记录法按每5 s一个频次,采集在放牧时间内(8∶30—17∶30)放牧牛采食、站立、慢走、躺卧和反刍的运动行为时间。

图 2 牛的行为示例图
-
无人机影像处理和制备NDVI均基于大疆智图完成。将影像导入到大疆智图软件中,进行包括相机标定、畸变校正、图像匹配、飞行姿态校正等预处理操作,对原始影像进行正射,将图像像素的灰度值转换为地物反射率或光谱反射率,并根据像元的位置计算出地物的空间分布,以消除地形起伏等因素对影像的影响,获得正射影像。选取最常用的植被指数归一化植被指数(NDVI)进行LAI反演,反演过程参照Hassan等[24]的研究,归一化植被指数NDVI的计算公式为:
$$ {I_{{\text{NDV}}}} = \frac{{{A_{{\text{ref}}}}({\text{NIR}}) - {A_{{\text{ref}}}}({\text{Red}})}}{{{A_{{\text{ref}}}}({\text{NIR}}) + {A_{{\text{ref}}}}({\text{Red}})}} 。$$ (4) 其中:Aref(NIR)和Aref(Red)分别表示NIR波段和Red波段的反照率。
将Qinc和Qref分别代表波段上的入射光量和反射光量,太阳光(入射光)照射到地面后发生反射,多光谱相机采集地面反射光线的信号并产生影像,而多光谱传感器则采集入射光线的信号,从而得出下列公式:
$${A_{{\text{ref}}}}\left( {{\text{NIR}}} \right) = \frac{{{Q_{{\text{ref}}}}\left( {{\text{NIR}}} \right)}}{{{Q_{{\text{inc}}}}\left( {{\text{NIR}}} \right)}} = \frac{{{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} \times {\rho _{{\text{NIR}}}}, $$ (5) $$ {A_{{\text{ref}}}}\left( {{\text{Red}}} \right) = \frac{{{Q_{{\text{ref}}}}\left( {{\text{Red}}} \right)}}{{{Q_{{\text{inc}}}}\left( {{\text{Red}}} \right)}} = \frac{{{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{{L_{\text{s}}}\left( {{\text{Red}}} \right)}} \times {\rho _{{\text{Red}}}}。$$ (6) Qinc(NIR)和Qref(NIR)分别表示在NIR波段上的入射光量和反射光量,Qinc(Red)和Qref(Red)则分别表示在Red波段上的反射光量和入射光量,Lcam(NIR)和Lcam(Red)分别表示从NIR和Red波段的多光谱图像中获得的图像信号值,而Ls(NIR)和Ls(Red)则分别表示从NIR和Red波段的多光谱光强传感器获得的感光信号值,ρNIR和ρRed表示调节NIR波段和Red波段的调节图像信号与多光谱光强传感器信号之间相互转化的参数。
在使用NIR波段相机作为标准时,其他波段的相机和多光谱光强传感器的感度都需要进行校准,需要将其他波段的相机和多光谱光强传感器的感度调整到与NIR波段的相机和多光谱光强传感器相匹配。ρx作为调节图像信号与多光谱光强传感器信号之间相互转化的参数,校准参数分别为pLcamX和pLSX。通过校准可以定义:
$${\rho _{\text{x}}} = {\rho _{{\text{NIR}}}}\frac{{p{L_{{\text{cam}}}}X}}{{p{L_{\text{s}}}X}}。 $$ (7) 可以得到:
$$ \begin{split} \,{A_{{\text{ref}}}}\left( {{\text{NIR}}} \right) & = \frac{{{Q_{{\text{ref}}}}\left( {{\text{NIR}}} \right)}}{{{Q_{{\text{inc}}}}\left( {{\text{NIR}}} \right)}} = \frac{{{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} \times {\rho _{{\text{NIR}}}} \times \frac{{p{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{p{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} \\ &= \frac{{{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{{L_{\text{s}}}\left( {{\text{NIR}}} \right) \times p{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} \times {\rho _{{\text{NIR}}}} , \\[-4pt] \end{split}$$ (8) $$ \begin{split} \, {A_{{\text{ref}}}}\left( {{\text{Red}}} \right) & = \frac{{{Q_{{\text{ref}}}}\left( {{\text{Red}}} \right)}}{{{Q_{{\text{inc}}}}\left( {{\text{Red}}} \right)}} = \frac{{{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{{L_{\text{s}}}\left( {{\text{Red}}} \right)}} \times {\rho _{{\text{NIR}}}} \times \frac{{p{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{p{L_{\text{s}}}\left( {{\text{Red}}} \right)}} \\ &= \frac{{{L_{{\text{cam}}}}\left( {{\text{Red}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{{L_{\text{s}}}\left( {{\text{Red}}} \right) \times p{L_{\text{s}}}\left( {{\text{Red}}} \right)}} \times {\rho _{{\text{NIR}}}} 。 \\[-4pt] \end{split} $$ (9) 因此,使用无人机影像计算NDVI的公式如下:
$$ \begin{split} & {I_{{\text{NDV}}}} = \frac{{{A_{{\text{ref}}}}\left( {{\text{NIR}}} \right) - {A_{{\text{ref}}}}\left( {{\text{Red}}} \right)}}{{{A_{{\text{ref}}}}\left( {{\text{NIR}}} \right) + {A_{{\text{ref}}}}\left( {{\text{Red}}} \right)}} = \\ &\left( {\frac{{{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{{L_{\text{s}}}\left( {{\text{NIR}}} \right) \times p{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} - \frac{{{L_{{\text{cam}}}}\left( {{\text{Red}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{{L_{\text{s}}}\left( {{\text{Red}}} \right) \times p{L_{\text{s}}}\left( {{\text{Red}}} \right)}}} \right)\Bigg/ \\ &\left( {\frac{{{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{NIR}}} \right)}}{{{L_{\text{s}}}\left( {{\text{NIR}}} \right) \times p{L_{\text{s}}}\left( {{\text{NIR}}} \right)}} + \frac{{{L_{{\text{cam}}}}\left( {{\text{Red}}} \right) \times p{L_{{\text{cam}}}}\left( {{\text{Red}}} \right)}}{{{L_{\text{s}}}\left( {{\text{Red}}} \right) \times p{L_{\text{s}}}\left( {{\text{Red}}} \right)}}} \right) 。 \end{split}$$ (10) -
归一化植被指数(NDVI)作为最常用的LAI反演指数,利用了冠层反射或辐射中的红光和近红外波段信息。本研究通过多光谱无人机数据生产NDVI,在3个不同放牧区域内分别选取30个监测点,使用植被冠层分析仪实地测量草地叶面积指数(LAI),分析监测点所对应NDVI与所测量LAI的相关关系,以此制备整个研究区的LAI数据。利用归一化植被指数(NDVI)估算LAI的通常步骤是先NDVI与LAI的经验关系,并使用植被冠层分析仪的实地观测数据进行拟合,再利用拟合好的模型进行估算,表达NDVI-LAI之间的经验关系L=f(x)主要有以下几种形式[25-26]:
$$ L=A X^3+B X^2+C X+D ,$$ (11) $$ L=A+B X^C ,$$ (12) $$ L=-1 / 2 A \ln (1-x) , $$ (13) $$ L=A e^{B x} , $$ (14) 式中,X为归一化植被指数NDVI,参数A、B、C和D为经验值,不同植被类型参数有所不同。
采用Origin统计软件对LAI和NDVI进行关系拟合。将同一时间通过LAI-2200C所测出来的LAI与基于无人机遥感影像所制备的NDVI进行一一对应,根据常用的经验模型,将NDVI和LAI进行组合,逐步拟合回归,依次输入模型方程,在进行拟合回归时,要考虑到NDVI的变化对于LAI变化的显著性程度大小,对于与LAI没有显著性关系的模型方程,直接排除该模型。通过实测的数据依次对上述方程进行拟合,获得最佳拟合方程和给定系数后,便可将公式应用到遥感影像上来进行LAI空间分布的制图。
-
基于经验模型反演的LAI具有较高的精度。选择73个样点进行经验模型关系拟合,部分样点NDVI和LAI值如表3所示。从图3(a)可以看出,NDVI与LAI具有较好的相关性,当NDVI接近0.8时,NDVI值趋于缓和,该LAI反演模型的相关系数R2为0.88,P值小于0.01,表明用该模型拟合NDVI和LAI的效果较好。图3(b)对实测LAI和经验模型模拟出来的LAI进行了验证,相关系数R2=0.74,两组数据之间的整体偏差较小,当LAI靠近1.5时,反演结果会与实测值出现偏差,但在其他取值范围偏差较小,并且从整体来看,两者具有大致相同的变化趋势,表明该经验模型反演的LAI误差较小,反演效果较好。
表 3 部分样点归一化植被指数(NDVI)和叶面积指数(LAI)值
NDVI LAI NDVI LAI 0.645 94 −0.041 0.747 80 1.56 0.658 05 0.131 0.751 99 1.62 0.672 03 0.201 0.764 08 1.56 0.681 73 0.290 0.766 22 1.25 0.692 09 0.337 0.774 08 1.78 0.697 78 1.6 0.783 46 1.44 0.705 51 1.18 0.787 54 1.99 0.718 57 1.95 0.795 80 2.16 0.725 85 1.38 0.797 35 2.16 0.727 22 1.41 0.807 38 2.25 0.737 78 1.46 0.810 44 2.35 …… …… …… …… 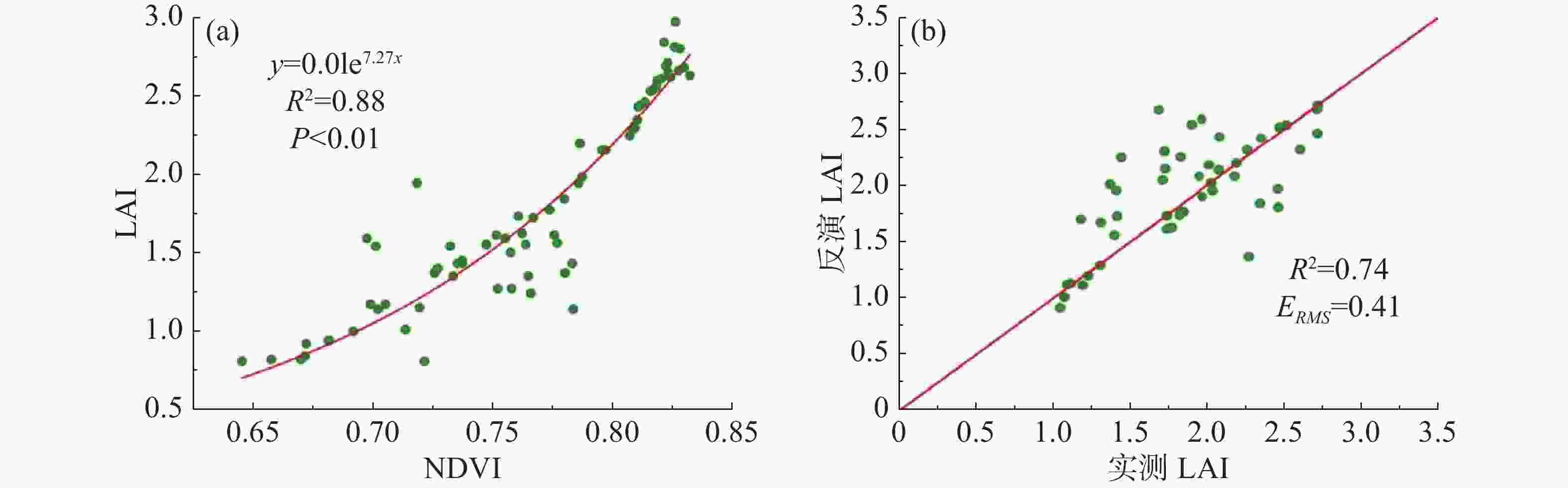
图 3 基于经验模型的草地LAI反演
-
重度放牧下,轮牧和连续放牧的草地LAI上升的区域占草地总面积的比例均比下降的区域占比小;中度放牧下,轮牧的草地LAI上升的区域占草地总面积的比例比下降的区域占比大,但连续放牧的草地LAI上升的区域占草地总面积的比例比下降的区域占比小;轻度放牧下,轮牧和连续放牧的草地LAI上升的区域占草地总面积的比例均比下降的区域占比大。
不同放牧方式和强度下各草地的LAI变化如图4所示,从G1(a,b,c)和G2(a,b,c)可以看出,在轮牧方式下,重度放牧时G1的LAI显著上升的区域极少呈现零散分布,下降的区域占比高达94.40%,G2的LAI上升区域几乎为0,下降的区域占比高达99.19%;中度放牧时,G1草地右下角处LAI呈现显著上升状态,整体来看草地有53.37%的草地区域呈上升趋势,G2则仅有47.91%的区域呈上升的趋势,上升区域主要集中在右上角的牛栏入口处,从平均水平上看,G1和G2两块草地平均上升的区域占比为50.65%;轻度放牧时,G1各处LAI有所上升,有高达64.21%的草地区域LAI值呈现明显上升的趋势,G2有57.83%的区域LAI呈现上升趋势,上升区域分布较为均匀。从G3(a,b,c)可以看出,在连续放牧方式下,重度放牧时G3的LAI下降区域占比87.35%,比G1和G2的都要低,中度放牧时25.83%的区域LAI呈现上升趋势,虽然下降的区域占比仍然较大,但是与重度放牧相比下降程度有所减轻,而在轻度放牧时,有60.37%的草地区域LAI呈现了上升趋势。图中的NG(a,b,c)体现出,在没有放牧干扰的情况下,同期未放牧草地LAI上升区域的占比始终比下降区域高,分别为67.96%、60.59%、53.83%,但可能是由于气候因素,上升区域占比逐渐减小。

图 4 不同放牧方式与强度下草地LAI变化动态
由此可得,重度放牧下将牛集中在G1和G2分别进行集中的5天放牧时,由于G1和G2面积较小,牛的可食用草量少,两个草地的LAI变化降幅较大,虽然G3的面积比G1和G2大,但是由于放牧头数过多和时间过长,导致3块草地LAI下降的区域占比都较大。中度放牧下轮牧有助于提高草地的LAI,而连续放牧由于畜牧长时间的啃食,导致即使是放牧强度降低,草地LAI上升区域仍然较小。而轻度放牧下草地承载的畜牧量足够低时,无论是基于轮牧还是连续放牧制度,草地状况都呈现出相同的向好趋势,LAI的上升区域占比也变得更大。
-
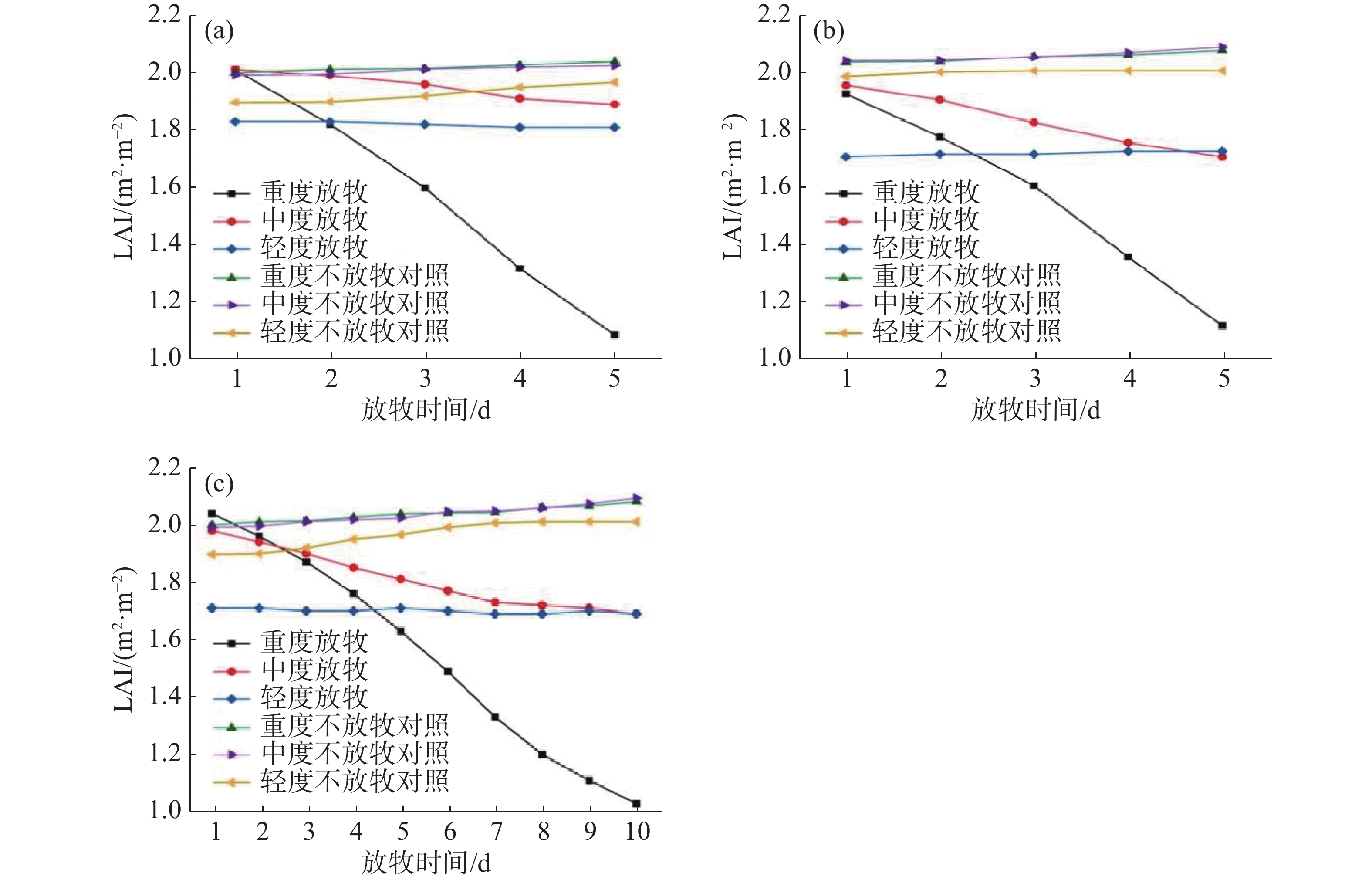
重度和中度放牧下,无论是轮牧还是连续放牧,草地LAI均值都呈现下降的趋势,下降幅度较大,而在轻度放牧下,轮牧的草地LAI均值呈现上升趋势,连续放牧的草地LAI均值依旧下降,下降幅度较低。各草地的LAI变化如图5所示,重度放牧下G1第一天的LAI均值为2.01,第五天LAI的均值为1.09,LAI均值下降了约45.78%;中度放牧下G1第一天的LAI均值为2.01,而第五天的LAI均值为1.89,相对于放牧前的LAI均值下降了约5.97%,然而有53.37%的草地区域呈显著上升的趋势;轻度放牧下G1第一天的LAI均值为1.83,而第五天的平均LAI均值仅为1.81,相对于放牧前下降了约1.09%,但是却有高达64.21%的草地区域LAI值呈现明显上升的趋势;同期未放牧草地NG的LAI分别从2.00、1.99和1.89上升到了2.04、2.02和1.97,上升幅度分别为1.97%、1.62%和3.66%。重度放牧下G2第一天的LAI均值为1.93,第五天LAI的均值为1.12,LAI均值下降了约41.97%;中度放牧下G2第一天的LAI均值为1.98,而第五天的LAI均值为1.71,相对于放牧前的LAI均值下降了约12.76%;轻度放牧下G2放牧第一天的LAI均值为1.71,而放牧第五天的LAI均值为1.73,相对于放牧前上升了约1.17%;同期未放牧草地NG的LAI分别从2.04、2.05和2.00上升到了2.08、2.09和2.01,上升幅度分别为2.01%、2.31%和1.01%。重度放牧下G3第一天的LAI均值为2.04,第十天的LAI均值为1.03,LAI下降了约49.51%;中度放牧下G3第一天的LAI均值为1.98,而第十天的LAI均值为1.69,变化了0.24,相对于放牧前的LAI均值下降了约14.65%;轻度放牧下G3第一天的平均LAI为1.71,而第十天的平均LAI为1.70,相对于放牧前下降了约0.58%,但依旧有60.37%的草地区域LAI呈现上升趋势;同期未放牧草地NG的LAI分别从2.00、1.99和1.89上升到了2.08、2.09和2.01,上升幅度分别为4.15%、5.12%和6.04%。
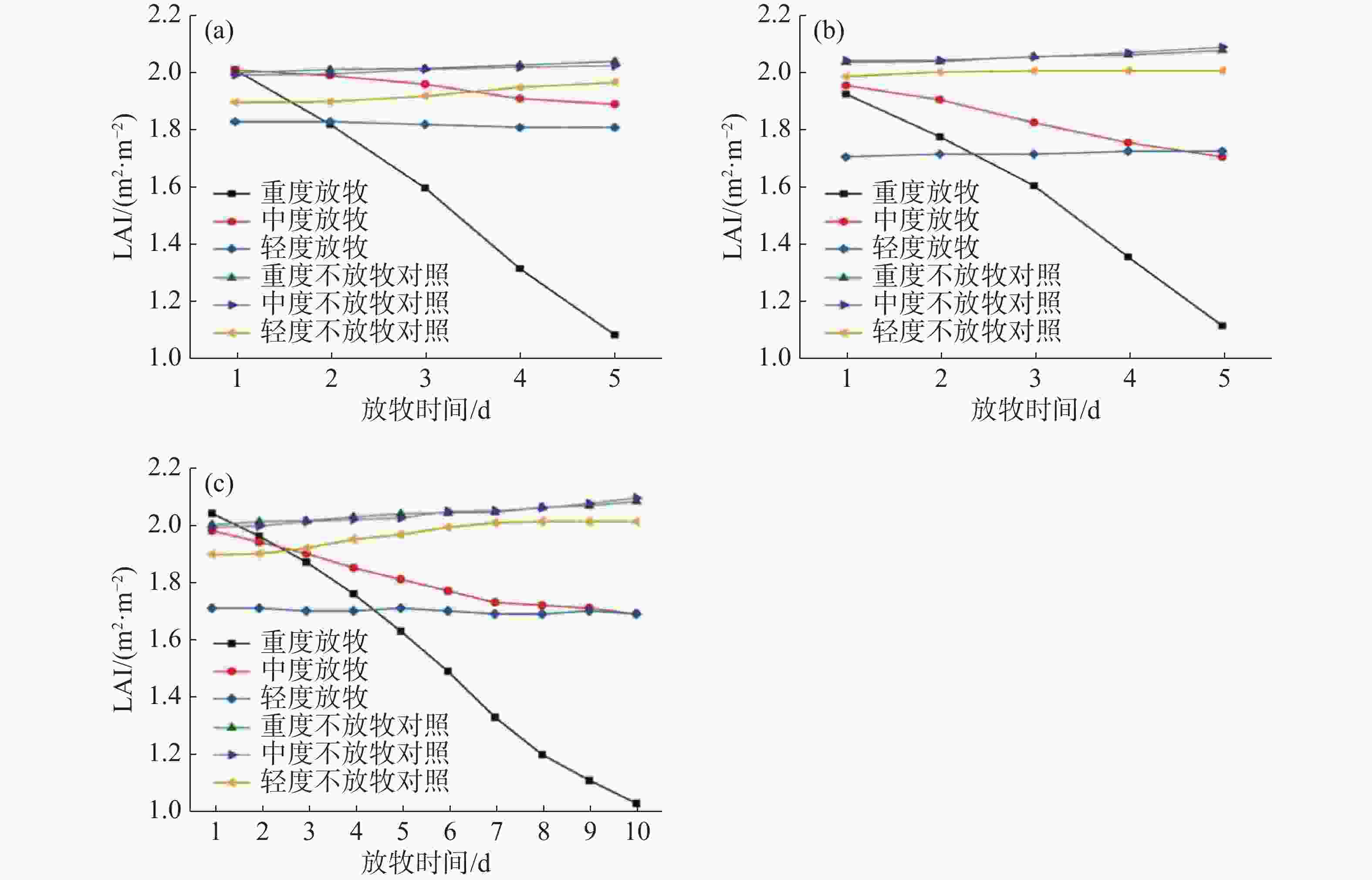
图 5 不同放牧强度下草地LAI均值变化
-
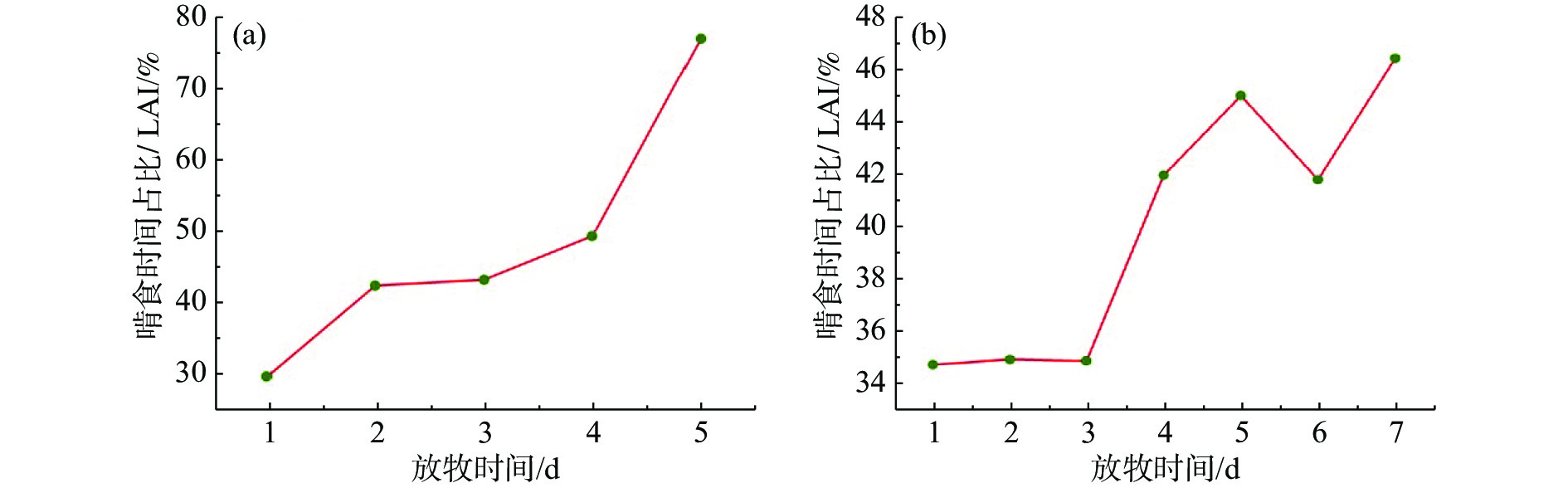
重度放牧下,无论是轮牧还是连续放牧都会增加牛的采食时间。根据连续记录法按每5 s一个频次,采集牛采食、站立、慢走、躺卧和反刍的运动行为时间,图6为采集的各个行为的比例,在G1中,采食的时间呈现出先增加后减少再增加的趋势,G1放牧的最后一天采食时间占比高达84.17%,较第一天增加了24.29%;躺卧的时间则呈现相反的趋势,最后一天躺卧占比仅有7.07%,较第一天减少了11.61%;站立的时间在放牧第一天和最后一天大致相同,分别为7.81%和8.57%,仅相差0.76%,但在其他时间极低;反刍的时间则在放牧最后一天达到最低,其变化趋势与采食时间完全相反,这可能是因为采食时间占用了反刍的时间;慢走时间占比始终最低,最后一天站立时间占比仅有0.19%,较第一天减少了1.62%。在G3中,采食时间呈现出先减少后增加再减少的趋势,在放牧第四天采食时间占比达到最高值73.91%,但每日采食时间差额不大,变化较为平缓;躺卧时间虽呈现相反的趋势,但变化同样较小;站立时间在放牧后期占比较大,最高可达11.83%,较第一天增长了6.93%;反刍时间呈现出先减少后增加的趋势,在采食时间最低时反刍时间达到最高9.53%;慢走时间占比始终最低,最后一天的站立时间较第一天减少了2.51%。
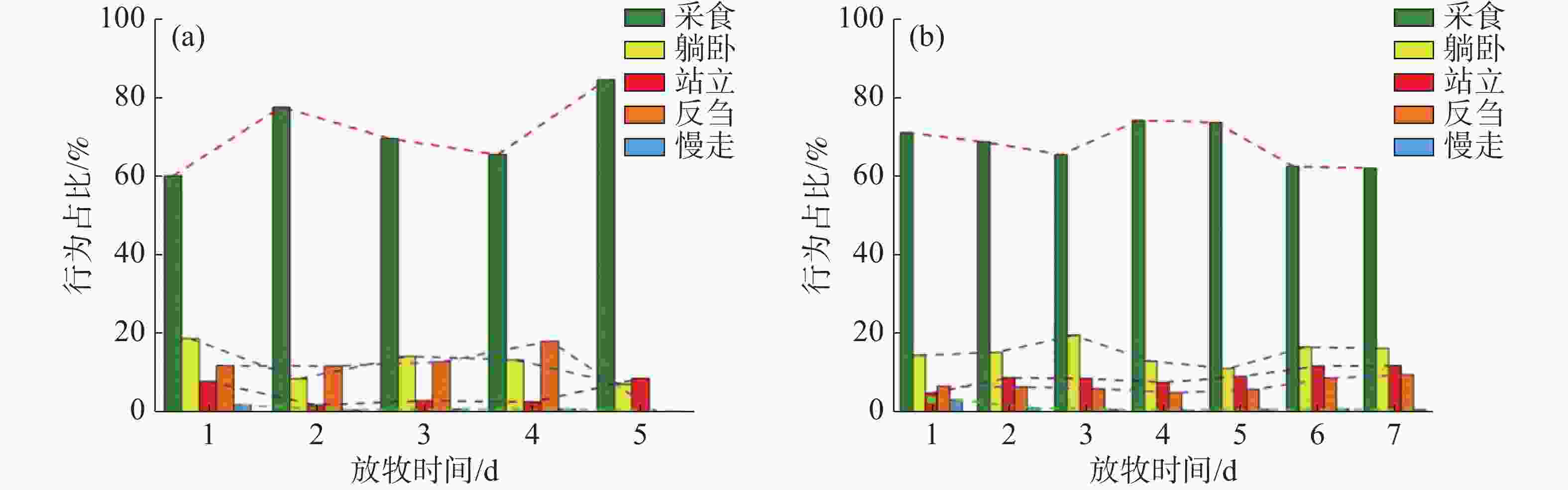
图 6 重度放牧条件下牛的行为时间占比
以啃食时间占比与LAI的比值来监测牛采食叶面积指数的量,发现在G1和G3中,随着放牧时间的变化,日均LAI都越来越小,虽然G1中啃食时间占比呈现了先增加后减少再增加的趋势,G3中啃食时间占比则先减少后增加再减少,但是从图7可以看出,两种放牧方式下草地的啃食时间占比与LAI的比值都越来越大,表明牛采食同样的LAI投入时间越来越长。综合来看,G1平均每天采食的时间占比为71.20%,G3为68.02%,无论是在轮牧草地还是在连续放牧草地,采食的时间占比始终最高,表明高强度放牧会显著增加畜牧采食时间。

图 7 重度放牧条件下牛的啃食时间占比/LAI
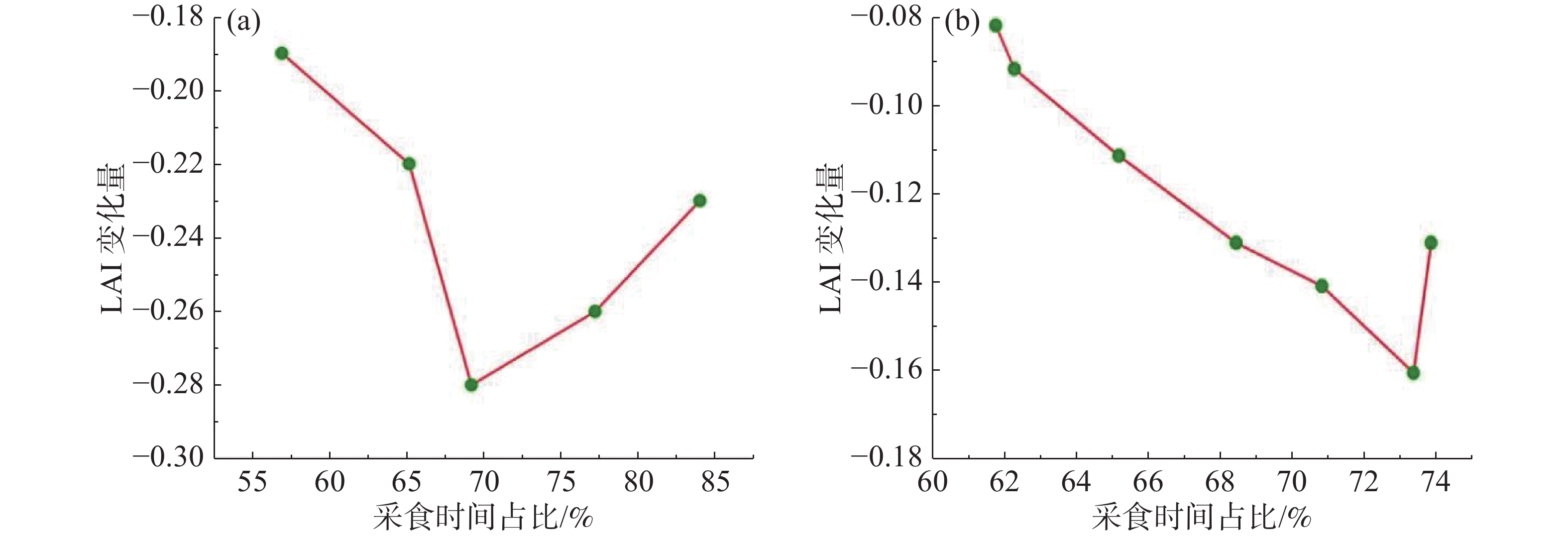
当牛的采食时间增加时,LAI减少量也随之增加,增加到一定程度就会到达草地的供给极限。图8为牛每日采食占比与LAI变化量的关系,LAI变化量由每日放牧结束后的LAI减去放牧开始前的LAI计算获得。由图8可知,在G1中,当采食时间占比逐渐增加时,LAI的变化量越大,减少量级越大,但当采食时间占比增加至70.88%左右时,LAI变化量逐渐回弹,不再降低;在G3中,最初的LAI变化量也是随着采食时间占比的增大而增大,但当采食时间占比增加至73.42%左右时,LAI变化量也逐渐呈现出上升的趋势。可见,无论是在轮牧还是连续放牧的制度下,当采食时间占比达到70.88%~73.42%左右时,已经到达了LAI变化的极限。此时,草地叶面积指数下降到初始状态(第一天啃食前)的79.60%~79.90%,即使啃食时间占比增加,LAI减少量也不再增加。

图 8 牛的采食时间占比与LAI变化量关系
-
通过放牧实验,本研究发现重度放牧对所有草地的LAI上升不利,所有草地的LAI上升的区域要远小于下降的区域;而在中度放牧强度下,轮牧制度的草地LAI上升的区域占草地总面积的比例比下降的比例大,而连续放牧地块的草地LAI上升的区域占草地总面积的比例比下降的比例小;轻度放牧强度下,轮牧和连续放牧的草地LAI上升的区域占草地总面积的比例均比下降的比例大。过度放牧一直以来都被认为是植被叶面积指数下降的主要负驱动力[27-28]。在过去的几十年里,中国西北广大草原的环境退化程度不断加剧,过度放牧是主要原因[29]。考虑到年际气候变化也会影响到牧场草地的生长,Hao等[30]将放牧对LAI的影响与年际气候变化的影响分开,同样证明了放牧对植被叶面积指数的负面影响(减少LAI)极大地抵消了为草地带来的正面影响(增加LAI)。而随着放牧强度的降低,草地冠层盖度等生物指标和丰富度增加,中度放牧被证明可以显著改善植被覆盖和促进物种恢复[31-32]。Deng等[33]在草原牧场,发现中度放牧管理可用于实现植物功能群落的理想转变,合理的放牧强度对牧场生态系统的可持续性有积极影响,通过中等强度放牧对草地进行长期有效管理既可以抵消当地的退化,也有益于草地的功能恢复。高寒草地野外观测也发现,在水热条件适宜的情况下,轻、中度放牧压力比不放牧更有利于提高叶面积指数,即轻、中度放牧区的叶面积指数高于非放牧区的叶面积指数[34]。因此,轻、中度压力的放牧对草地的贡献是积极的,有利于提高草地LAI,促进草地生长。这可能是由于草地的生长速率与草地的现存草量密切相关,重度放牧下草地现存草量少,草地再生长所需的营养物质和能量得不到充足的供应,草地再生速率降低,而在轻度放牧下,草地现存草量多,草地的营养物质维持了植被的呼吸作用,消耗掉大量能量,只有在适度放牧下,才能将植被高度和覆盖度控制在一定范围内,可以使得光线更加均匀地分布到草地内部,提高光合作用的速率和草地生长效率,牧草得到及时更新。
-
轮牧制度的草地在重度放牧下呈现出下降趋势,但在中度和轻度放牧下草地的LAI上升区域面积占比与下降的区域相比较大。连续放牧制度下的草地LAI在重度和中度放牧下呈现相同的下降趋势,只有在轻度放牧时LAI上升区域占比高于下降区域占比。当放牧强度适中时,在严格的放牧管理下如轮牧制度,可以使LAI保持相对稳定,草地的叶面积指数出现微增。这是因为植物能够维持当前体内有机物的持续流量,通过体内的有机储备在使得草地保持在最低LAI标准以上和不受到生长条件的限制[35],Trindaded等[36]已发现了叶面积与消化率和营养价值呈正相关,轮牧制度通过更换放牧草地来增加放牧间隔时间,促进叶片的营养生长和草茎的伸长,保证牛的啃食量。一般来说,连续放牧的草地会导致草地管理不当,Da等[37]发现轮牧和连续放牧之间的牧草积累和草地LAI存在明显的差异,连续放牧所带来的高强度放牧会使草地LAI减少,这表明在连续放牧条件下,畜牧的过度踩踏无法为植物提供可以维持生长的有机储备。Sbrissia等[38]的结果也表明,在连续放牧的条件下,由于地上所产生的同化物会使得牧草积累,此时草地叶片的质量就对植物的生长起着至关重要的作用,甚至会阻碍植物生长,叶面积指数就会变得越低。
-
在重度放牧下,无论是轮牧还是连续放牧都会增加牛的采食时间,LAI减少量也随之增加,增加到一定程度就会到达草地的供给极限。当牛的采食时间占比达到70.88%~73.42%左右时,也就是草地能够单日供给的极限后,草地可食用牧草大幅减少,牲畜个体采食竞争加剧,整体采食时间占比提高,草地叶面积快速下降。以往的研究也证明了由于牲畜对草地的直接啃食,草地地上生物量有所下降[39-40],这可能是由于畜牧在采食过程中不断踩踏,导致土壤压实从而限制了草地根系的伸长和发育[41-42]。此外也有研究表明,牲畜的踩踏和践踏会通过压实增加表土的容重[43],表土结构的这些变化导致含水量下降[44],渗透因此减少,加上植被覆盖减少将导致蒸腾减少[45]。畜牧的采食时间过长会导致植被产量和生物量减少,氨化、硝化作用和土壤肥力下降[46]。过度放牧下畜牧的高强度啃食已被认为是中国西北部草地覆盖率和质量下降、生物多样性丧失及水土流失导致土地退化的主要原因之一[47]。因此,了解草地退化背后驱动力的影响对于应对全球变化适应具有重要的生态系统管理意义[48]。为了提高草原生态系统的服务功能,应减少放牧压力,特别是在不利气候和过度放牧双重负面影响加剧的地区,轮牧应该纳入管理计划,规定在整个放牧区域的放牧时间,调整牛的啃食时间,有助于评估该地区最近大规模生态恢复活动的有效性,未来的研究也可以侧重于通过控制牛的啃食时间来了解不同放牧制度下的草地LAI动态的生物物理过程。
-
基于多光谱无人机反演的不同放牧策略下草地叶面积指数结果显示,在中度放牧时轮牧草地LAI上升的区域占草地总面积的比例比下降的区域大,但连续放牧草地的LAI上升区域占草地总面积的比例比下降的区域小,这表明中度放牧条件下的轮牧有利于草地LAI的提升。而在重度放牧下,牛的采食时间占总行为比始终最高,无论是轮牧还是连续放牧都会增加牛的采食时间,LAI减少量也随之增加,增加到一定程度就会到达草地的供给极限,即使采食时间增加,LAI减少量也不再增加。当然,放牧行为对草地的生长可能存在着滞后效应,这种效应在未来将会成为进行长期草畜监测研究的重要问题。
基于多光谱无人机的不同放牧策略对草地叶面积指数变化动态解析
DOI: 10.15886/j.cnki.rdswxb.20230047
 CSTR: 32425.14.j.cnki.rdswxb.20230047
CSTR: 32425.14.j.cnki.rdswxb.20230047
Dynamic analysis of grassland leaf area index changes under different grazing strategies based on multispectral UAV
-
摘要: 针对海南岛耕地撂荒形成的草地无序放牧较为普遍的现象,以不同放牧方式(轮牧和连续放牧)、不同放牧强度(重度、中度和轻度)进行草地放牧试验,利用多光谱无人机和植被冠层分析仪,获取放牧期间日尺度草地叶面积指数(Leaf area index, LAI)信息,定量分析不同放牧策略对草地叶面积指数及牧牛行为变化的影响。研究结果表明:(1)中度放牧条件下的轮牧有利于草地LAI的提升。重度放牧时,轮牧和连续放牧草地的LAI上升的区域占草地总面积的比例分别为3.21%和12.65%;中度放牧时,轮牧和连续放牧草地的LAI上升的区域占草地总面积的比例分别为52.01%和25.83%;轻度放牧时,轮牧和连续放牧草地的LAI上升的区域占草地总面积的比例分别为61.02%和60.37%。(2)重度放牧条件下,牛的采食时间占比始终最高,随着采食时间占比的增加,草地LAI的减少量也随之增加,但当采食时间占比增加至70.88%~73.42%时,草地LAI的减少量逐渐降低。此时,草地叶面积指数下降到初始状态(第一天啃食前)的79.60%~79.90%,即牛的啃食量已经到达了草地LAI当日能够供给的极限,当超过这个极限时,草地可食用牧草大幅减少,牲畜个体采食竞争加剧,同时牛的啃食行为时间占比大大提高。研究结果有助于优选出牧场尺度下最佳的草畜管理措施,为热带草地畜牧系统的可持续发展从新的角度提供理论方法和决策支撑,助力国家生态文明试验区(海南)的建设。Abstract: Disorderly grazing in grassland on abandoned cultivated land hindered the sustainable development of the grassland in Hainan Island. Grassland grazing experiments were conducted according to different grazing methods (rotational grazing and continuous grazing) and different grazing intensities (severe, moderate, and slight). Multispectral UAVs and vegetation canopy analyzers were used to obtain daily-scale grassland leaf area index (LAI) during the grazing period, and the impact of different grazing strategies on changes in grassland leaf area index and cattle behavior were also quantitatively analyzed. Results showed that rotational grazing under moderate grazing intensity was beneficial to improving grassland LAI. Under severe grazing intensity the proportion of areas where LAI increased in rotational grazing and continuous grazing grassland accounted for 3.21% and 12.65% of the total grassland area, respectively. Under moderate grazing intensity the proportion of areas where LAI increased in rotational grazing and continuous grazing grassland accounted for 52.01% and 25.83% of the total grassland area, respectively. Under slight grazing intensity the proportion of areas where LAI increased in rotational grazing and continuous grazing grassland accounted for 61.02% and 60.37% of the total grassland area, respectively. Under severe grazing intensity the proportion of cattle's feeding time to the total behavior was always the highest. As the proportion of feeding time increased, the reduction of grassland LAI was also increased, but when the proportion of feeding time increasing to 70.88%-73.42%, the reduction of grassland LAI was gradually decreased. At this time, the leaf area index of the grassland dropped to 79.60%-79.90% of the initial leaf area index (before grazing on the first day), which meant that the number of leaves fed by the cattle was upto the limit that the LAI of the grassland could supply on the day. When this limit was exceeded, individual livestock competition would be intensified. At the same time, the proportion of time spent by cattle on gnawing behavior greatly increased. All these results will help to select the best grass and livestock management methods at the pasture scale, to provide theoretical methods and decision-making support for the sustainable development of tropical grassland livestock systems from a new perspective, and to assist the construction of the National Pilot Zone for Ecological Conservation (Hainan).
-
Key words:
- grazing strategy /
- grassland /
- leaf area index /
- multispectral /
- Hainan Island
-
表 1 放牧实验设计
放牧强度 放牧日期 轮牧G1、
G2/头连续放
牧G3/头载畜量/
(头·hm−2)重度放牧 2022-11-10—11-19 9 7 41 中度放牧 2022-12-9—12-18 5 4 23 轻度放牧 2022-12-20—12-29 3 2 13  下载: 导出CSV
下载: 导出CSV
表 3 部分样点归一化植被指数(NDVI)和叶面积指数(LAI)值
NDVI LAI NDVI LAI 0.645 94 −0.041 0.747 80 1.56 0.658 05 0.131 0.751 99 1.62 0.672 03 0.201 0.764 08 1.56 0.681 73 0.290 0.766 22 1.25 0.692 09 0.337 0.774 08 1.78 0.697 78 1.6 0.783 46 1.44 0.705 51 1.18 0.787 54 1.99 0.718 57 1.95 0.795 80 2.16 0.725 85 1.38 0.797 35 2.16 0.727 22 1.41 0.807 38 2.25 0.737 78 1.46 0.810 44 2.35 …… …… …… ……
下载: 导出CSV
-
[1] DEBLONDE G, PENNER M, ROYER A. Measuring leaf area index with the Li-cor LAI-2000 in pine stands[J]. Ecology, 1994, 75(5): 1507 − 1511. doi: 10.2307/1937474 [2] LANE D R, COFFIN D P, LAUENROTH W K. Changes in grassland canopy structure across a precipitation gradient[J]. Journal of Vegetation Science, 2000, 11(3): 359 − 368. doi: 10.2307/3236628 [3] WATSON D J. Comparative physiological studies on the growth of field crops: I. variation in net assimilation rate and leaf area between species and varieties, and within and between years[J]. Annals of Botany, 1947, 11(1): 41 − 76. doi: 10.1093/oxfordjournals.aob.a083148 [4] CHEN J M, CIHLAR J. Retrieving leaf area index of boreal conifer forests using Landsat TM images[J]. Remote Sensing of Environment, 1996, 55(2): 153 − 162. doi: 10.1016/0034-4257(95)00195-6 [5] CHAPIN F S III, SALA O E, BURKE I C, et al. Ecosystem consequences of changing biodiversity[J]. BioScience, 1998, 48(1): 45 − 52. doi: 10.2307/1313227 [6] GUO X, ZHANG C, WILMSHURST J, et al. Monitoring grassland health with remote sensing approaches[J]. Prairie Perspectives, 2005, 8: 11 − 22. [7] BLACK S C. Estimation of grass photosynthesis rates in mixed-grass prairie using field and remote sensing approaches [D], Saskatoon: University of Saskatchewan, 2006. [8] ZHANG C, GUO X, WILMSHURST J, et al. Application of RADARSAT imagery to grassland biophysical heterogeneity assessment[J]. Canadian Journal of Remote Sensing, 2006, 32(4): 281 − 287. doi: 10.5589/m06-025 [9] COLOMINA I, MOLINA P. Unmanned aerial systems for photogrammetry and remote sensing: a review[J]. ISPRS Journal of Photogrammetry and Remote Sensing, 2014, 92: 79 − 97. doi: 10.1016/j.isprsjprs.2014.02.013 [10] BUNNIK N J J The multispectral reflectance of shortwave radiation by agricultural crops in relation with their morphological and optical properties. 1978. [11] 唐芳林, 周红斌, 朱丽艳, 等. 构建林草融合的草原调查监测体系[J]. 林业建设, 2020(5): 11 − 16. [12] LI S, DING X, KUANG Q, et al. Potential of UAV-based active sensing for monitoring rice leaf nitrogen status[J]. Frontiers in Plant Science, 2018, 9: 1834.??. doi: 10.3389/fpls.2018.01834 [13] UNDERSANDER D, ALBERT B, COSGROVE D, et al. Pastures for profit: A guide to rotational grazing [M]. Madison: Cooperative Extensiton Publications, University of Wisconsin-Extension, 2002. [14] 韩国栋, 卫智军, 许志信. 短花针茅草原划区轮牧试验研究[J]. 内蒙古农业大学学报(自然科学版), 2001, 22(1): 60 − 67. [15] 马骏骥, 李钢铁, 刘慧, 等. 放牧方式对浑善达克沙地东缘草地植被特征影响[J]. 内蒙古林业科技, 2016, 42(2): 30 − 34. [16] 许志信, 赵萌莉. 过度放牧对草原土壤侵蚀的影响[J]. 中国草地, 2001, 23(6): 60 − 64. [17] TEAGUE W R, DOWHOWER S L. Patch dynamics under rotational and continuous grazing management in large, heterogeneous paddocks[J]. Journal of Arid Environments, 2003, 53(2): 211 − 229. doi: 10.1006/jare.2002.1036 [18] SANJARI G, GHADIRI H, CIESIOLKA C A A, et al. Comparing the effects of continuous and time-controlled grazing systems on soil characteristics in Southeast Queensland[J]. Soil Research, 2008, 46(4): 348 − 358. doi: 10.1071/SR07220 [19] 彭祺, 王宁. 不同放牧制度对草地植被的影响[J]. 农业科学研究, 2005, 26(1): 27 − 30. [20] 张铜会, 赵哈林, 大黑俊哉, 等. 连续放牧对沙质草地植被盖度、土壤性质及其空间分布的影响[J]. 干旱区资源与环境, 2003, 17(4): 117 − 121. [21] SEMIADI G, MUIR P D, BARRY T N, et al. Grazing patterns of sambar Deer (Cervus unicolor) and red Deer (Cervus elaphus) in captivity[J]. New Zealand Journal of Agricultural Research, 1993, 36(2): 253 − 260. doi: 10.1080/00288233.1993.10417761 [22] 罗红霞, 戴声佩, 刘恩平, 等. 2001—2014年海南岛植被覆盖时空变化特征[J]. 水土保持研究, 2018, 25(5): 343 − 350. [23] COSTA C M, DIFANTE G S, COSTA A B G, et al. Grazing intensity as a management strategy in tropical grasses for beef cattle production: a meta-analysis[J]. Animal, 2021, 15(4): 100192. doi: 10.1016/j.animal.2021.100192 [24] HASSAN M A, YANG M, RASHEED A, et al. A rapid monitoring of NDVI across the wheat growth cycle for grain yield prediction using a multi-spectral UAV platform[J]. Plant Science, 2019, 282: 95 − 103. doi: 10.1016/j.plantsci.2018.10.022 [25] QI J, CHEHBOUNI A, HUETE A R, et al. A modified soil adjusted vegetation index[J]. Remote Sensing of Environment, 1994, 48(2): 119 − 126. doi: 10.1016/0034-4257(94)90134-1 [26] SHAFIAN S, RAJAN N, SCHNELL R, et al. Unmanned aerial systems-based remote sensing for monitoring sorghum growth and development[J]. PLoS One, 2018, 13(5): e0196605. doi: 10.1371/journal.pone.0196605 [27] HAO L, PAN C, LIU P, et al. Detection of the coupling between vegetation leaf area and climate in a multifunctional watershed, northwestern China[J]. Remote Sensing, 2016, 8(12): 1032 − 1055. doi: 10.3390/rs8121032 [28] LIU P, HAO L, PAN C, et al. Combined effects of climate and land management on watershed vegetation dynamics in an arid environment[J]. Science of the Total Environment, 2017, 589: 73 − 88. doi: 10.1016/j.scitotenv.2017.02.210 [29] BARANOVA A, SCHICKHOFF U, WANG S, et al. Mountain pastures of Qilian Shan: plant communities, grazing impact and degradation status (Gansu Province, NW China)[J]. Hacquetia, 2016, 15(2): 21 − 35. doi: 10.1515/hacq-2016-0014 [30] HAO L, PAN C, FANG D, et al. Quantifying the effects of overgrazing on mountainous watershed vegetation dynamics under a changing climate[J]. Science of the Total Environment, 2018, 639: 1408 − 1420. doi: 10.1016/j.scitotenv.2018.05.224 [31] LIANG Y, HAN G, ZHOU H, et al. Grazing intensity on vegetation dynamics of a typical steppe in northeast inner Mongolia[J]. Rangeland Ecology & Management, 2009, 62(4): 328 − 336. [32] ZHAO L P, SU J S, WU G L, et al. Long-term effects of grazing exclusion on aboveground and belowground plant species diversity in a steppe of the Loess Plateau, China[J]. Plant Ecology and Evolution, 2011, 144(3): 313 − 320. doi: 10.5091/plecevo.2011.617 [33] DENG L, SWEENEY S, SHANGGUAN Z P. Grassland responses to grazing disturbance: plant diversity changes with grazing intensity in a desert steppe[J]. Grass and Forage Science, 2014, 69(3): 524 − 533. doi: 10.1111/gfs.12065 [34] 苏爱玲, 张振华, 汪诗平, 等. 不同季节放牧对矮嵩草草甸植物叶面积指数的影响[J]. 草原与草坪, 2010, 30(1): 50 − 55. [35] BARON V S, MAPFUMO E, DICK A C, et al. Grazing intensity impacts on pasture carbon and nitrogen flow[J]. Journal of Range Management, 2002, 55(6): 535 − 541. doi: 10.2307/4003996 [36] DA TRINDADE J K, DA SILVA S C, DE SOUZA S J JR, et al. Composição morfológica da forragem consumida por bovinos de corte durante o rebaixamento do capim-marandu submetido a estratégias de pastejo rotativo[J]. Pesquisa Agropecuária Brasileira, 2007, 42(6): 883 − 890. [37] DA SILVA S C, PEREIRA L E T, SBRISSIA A F, et al. Carbon and nitrogen reserves in marandu palisade grass subjected to intensities of continuous stocking management[J]. The Journal of Agricultural Science, 2015, 153(8): 1449 − 1463. doi: 10.1017/S0021859614001130 [38] SBRISSIA A F, DA SILVA S C, SARMENTO D O L, et al. Tillering dynamics in palisadegrass swards continuously stocked by cattle[J]. Plant Ecology, 2010, 206(2): 349 − 359. doi: 10.1007/s11258-009-9647-7 [39] LIN X, ZHANG Z, WANG S, et al. Response of ecosystem respiration to warming and grazing during the growing seasons in the alpine meadow on the Tibetan Plateau[J]. Agricultural and Forest Meteorology, 2011, 151(7): 792 − 802. doi: 10.1016/j.agrformet.2011.01.009 [40] DENG L, ZHANG Z, SHANGGUAN Z. Long-term fencing effects on plant diversity and soil properties in China[J]. Soil and Tillage Research, 2014, 137: 7 − 15. doi: 10.1016/j.still.2013.11.002 [41] SUMANTA B, RITCHIE MARK E. Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition[J]. Ecology Letters, 2010, 13(8): 959 − 68. doi: 10.1111/j.1461-0248.2010.01486.x [42] BAI W, FANG Y, ZHOU M, et al. Heavily intensified grazing reduces root production in an Inner Mongolia temperate steppe[J]. Agriculture, Ecosystems & Environment, 2015, 200: 143 − 150. [43] FATICHI S, ZEEMAN M J, FUHRER J, et al. Ecohydrological effects of management on subalpine grasslands: from local to catchment scale[J]. Water Resources Research, 2014, 50(1): 148 − 164. doi: 10.1002/2013WR014535 [44] KAIRIS O, KARAVITIS C, SALVATI L, et al. Exploring the impact of overgrazing on soil erosion and land degradation in a dry Mediterranean agro-forest landscape (crete, Greece)[J]. Arid Land Research and Management, 2015, 29(3): 360 − 374. doi: 10.1080/15324982.2014.968691 [45] PEREYRA D A, BUCCI S J, ARIAS N S, et al. Grazing increases evapotranspiration without the cost of lowering soil water storages in arid ecosystems[J]. Ecohydrology, 2017, 10(6): e1850. doi: 10.1002/eco.1850 [46] STEINFELD H, WASSENAAR T. The role of livestock production in carbon and nitrogen cycles[J]. Annual Review of Environment and Resources, 2007, 32: 271 − 294. doi: 10.1146/annurev.energy.32.041806.143508 [47] SUN L, YANG L, HAO L, et al. Hydrological effects of vegetation cover degradation and environmental implications in a semiarid temperate steppe, China[J]. Sustainability, 2017, 9(2): 281 − 301. doi: 10.3390/su9020281 [48] GAO J. Analysis and assessment of the risk of snow and freezing disaster in China[J]. International Journal of Disaster Risk Reduction, 2016, 19: 334 − 340. doi: 10.1016/j.ijdrr.2016.09.007 -

 点击查看大图
点击查看大图
图(8) / 表 (3)
计量
- 文章访问数: 1153
- HTML全文浏览量: 625
- PDF下载量: 49
- 被引次数: 0



















