

 下载:
下载:

-
海南岛地处华南热带与亚洲热带北缘,其中部与南部山区孕育了丰富的动植物资源,属于“海南热带雨林国家公园”辖区,也是受到国际社会密切关注的生物多样性保护热点区域[1]。海南岛受季风影响而分为干、湿2季,但在霸王岭、尖峰岭、黎母山和五指山等海拔 1 200 m以上的山顶和山脊地段,终年云雾缭绕,是我国主要的热带山地云雾林分布区[2-3]。云雾林内拥有很高的植物多样性和特有性,尤其是丰富的树附生苔藓植物,构成了云雾林植物群落的一大特色。
树附生苔藓是指附生于维管植物的茎干、树枝等表面的苔藓植物,主要包括苔类植物和藓类植物两大类群。多数树附生苔藓沿附主的茎和枝干匍匐或蔓延生长,形成平铺型、交织型或悬垂型等多种生活型,在森林的水分涵养、物质循环等方面执行着重要功能[4]。树附生苔藓不仅对附主植物有强烈的依赖性,而且具有变水植物的特点[5],对环境湿度的变化十分敏感。当今全球气候持续变暖,空气污染加重,使得树附生苔藓面临着生存的多重压力,其多样性易迅速减少而难以恢复。然而海南热带雨林国家公园树附生藓类植物的资源分布情况,以及气候条件变化对树附生藓类物种多样性是否有影响尚未见报道,为此,笔者于2019—2020年多次对霸王岭自然保护区热带山地云雾林的树附生藓类植物进行野外调查、标本采集和鉴定,同时查阅了相关文献,以期为海南热带雨林国家公园苔藓植物的研究与保护提供新资料。
-
Yakushimabryum H.Akiyama, Ying Chang, T.Yamag. & B.C.Tan, J. Bryol. 33(1): 42. 2011.
模式种:Y. longissimum H.Akiyama, Ying Chang, T.Yamag. & B.C.Tan
植物体淡绿色至黄绿色,无光泽或具光泽。主茎匍匐,紧贴基质,有密集和不规则的分枝;支茎短,匍匐或倾立。茎不具中轴。枝上部叶腋常着生丝状芽孢。鳞毛缺。具假鳞毛或无。假根浅褐色至红褐色,成束状,光滑或稀疏具疣,多着生于主茎上。茎叶与枝叶形态相近,或茎叶略小。叶卵圆状至长圆状披针形,先端渐尖或锐尖,基部略狭;中肋缺或具不明显2短中肋,边缘具齿或仅上部边缘具微齿;叶细胞线形或长梭形,壁薄,稍波状,光滑;角细胞多列,方形,黄色至红褐色,壁厚,形成明显分化的细胞群[6]。
雌雄异株。雄苞着生于枝顶端或侧短枝上,雄苞叶宽卵圆形,具尖,无中肋。内雌苞叶无中肋,窄披针形,向先端渐狭。蒴柄细,平滑,红棕色。孢蒴直立或稍倾斜,窄卵圆形至圆柱形,蒴盖细长,环带不分化,干时蒴口下部多少内缢。蒴齿2层。外齿层齿片披针形,长渐尖,光滑或具密疣。内齿层齿条线形,具密疣;无齿毛;基膜低。孢子圆球形,绿色,壁稍厚,具细疣[7]。
生境:树干或树枝生。
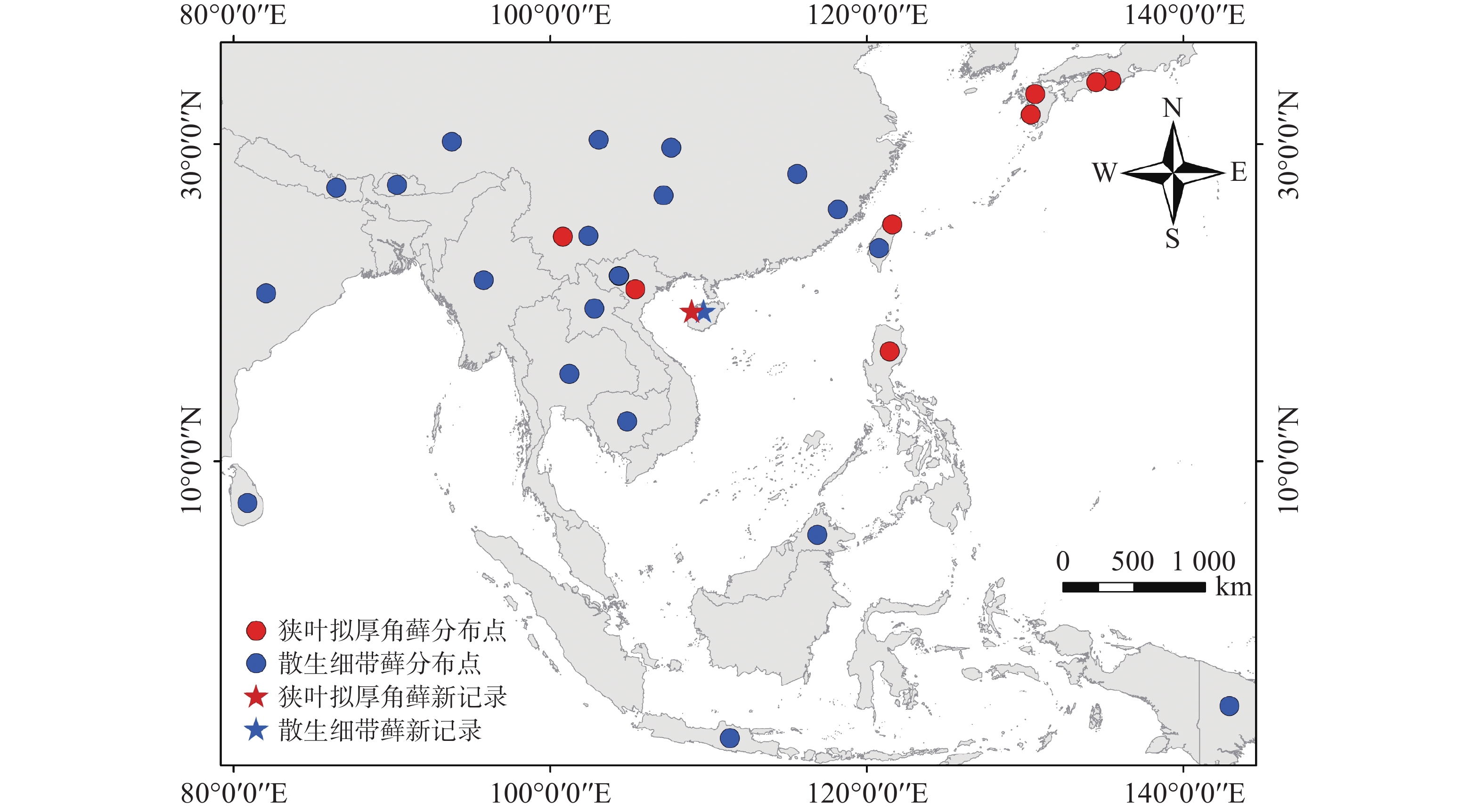
分布:中国(云南、香港、台湾),日本,马来西亚,菲律宾,越南[8],海南首次记录。
拟厚角藓属最初以Y. longissimum为模式标本,建立在毛锦藓科(Pylaisiadelphaceae)之下[6],该属与同科的竹藓属[Aptychella (Broth.) Herzog]、厚角藓属(Gammiella Broth.)等近缘属在形态上相近,因而属间关系一直存在争议[9-10]。为探讨属间以及属下分类群的系统地位,AKIYAMA 等[8]依据cpDNA片段构建分子系统树,并结合形态学证据,发现竹藓属的Aptychella subintegra Tixier与厚角藓属的狭叶厚角藓[Gammiella tonkinensis (Broth. & Paris) B.C.Tan]都隶属于拟厚角藓属,从而将上述2个种重新组合为:亚全缘拟厚角藓(新拟)[Y. subintegrum (Tixier) H.Akiyama]和狭叶拟厚角藓(新拟)[Y. tonkinense (Broth. & Paris) H.Akiyama],将Y. longissimum处理为Y. subintegrum的异名,同时发表1个新种:Y. brevigemmium H.Akiyama。
2020年,HAN 和 JIA [11]采用分子标记和形态学方法,整理修订了广义锦藓科(Sematophyllaceae s.l.,即狭义锦藓科+毛锦藓科)家族的分类阶元关系,结果显示毛锦藓科为并系,包含在广义锦藓科这一单系类群中。在此基础上,广义锦藓科被划分为6个亚科,拟厚角藓属划归为同叶藓亚科(新拟)(Isopterygoideae W.Han & Y.Jia)。目前,拟厚角藓属在全世界有3种,中国记载有2种:亚全缘拟厚角藓、狭叶拟厚角藓。
-
Yakushimabryum tonkinense (Broth. & Paris) H.Akiyama, Acta Phytotax. Geobot. 68(3): 165. 2017.
Basionym: Clastobryum tonkinense Broth. & Paris, Rev. Bryol. 35: 47. 1908 ≡ Aptychella tonkinensis (Broth. & Paris) Broth., Nat. Pflanzenfam. (ed. 2), 11: 406. 1925 ≡ Gammiella tonkinensis (Broth. & Paris) B.C.Tan, Bryologist 93: 433. 1990; Moss Fl. China 8: 33. 2005.
Aptychella glomeratopropagulifera (Toyama) Seki, J. Sci. Hiroshima Univ., Ser. B, Div. 2, Bot. 12: 72. 1968 ≡ Clastobryum glomeratopropagulifera Toyama, Acta Phytotax. Geobot. 4: 214. f. 2. 1935.
植物体淡绿至黄绿色,老标本变棕黄色,有时稍具光泽。主茎匍匐,紧贴基质,密集分枝。分枝长度常小于1 cm,平伏或稍斜倾立,叶片着生略呈圆条形,先端着芽部分易断裂。茎不具中轴,直径达0.1 mm。假根淡褐色至红褐色,成束状,光滑或具细疣。假鳞毛缺或短丝状,长3~5个细胞。叶新鲜或干时伸展,卵圆状至长圆状披针形,长0.8~1.9 mm,宽0.2~0.5 mm;先端骤缩呈锐尖或渐尖,内凹,基部较狭,不下延;具不明显2短中肋或无中肋;边缘多少内曲,具明显细齿或微齿。叶片上部细胞窄长梭形至线形,长约50~100 µm,宽约5 µm,壁薄,光滑;中部细胞线形,壁薄或略加厚,长80~125 µm,宽约5 µm,光滑;角细胞分化明显,具2~4 列方形或长方形细胞,壁厚,黄色至黄褐色,最外部1或2列细胞多少膨大,上方有少数短小的细胞。孢蒴未见。形态特征详见图1。

图 1 狭叶拟厚角藓
Figure 1. Yakushimabryum tonkinense (Broth. & Paris) H.Akiyama
标本鉴定:海南昌江县霸王岭国家级自然保护区松林顶,19°4′57.83″ N,109°12′43.59″ E,海拔1 336 m,徐雪艳等20190425-36B;19°5′12.18″ N,109°12′40.52″ E,海拔1 378 m,徐雪艳等20190819-23D;19°5′12.55″ N,109°12′40.16″ E,海拔1 383 m,徐雪艳等20190819-34B。
生境:树干或树枝附生。
地理分布:中国(云南、台湾),日本,菲律宾,越南[8],海南首次记录(图2)。
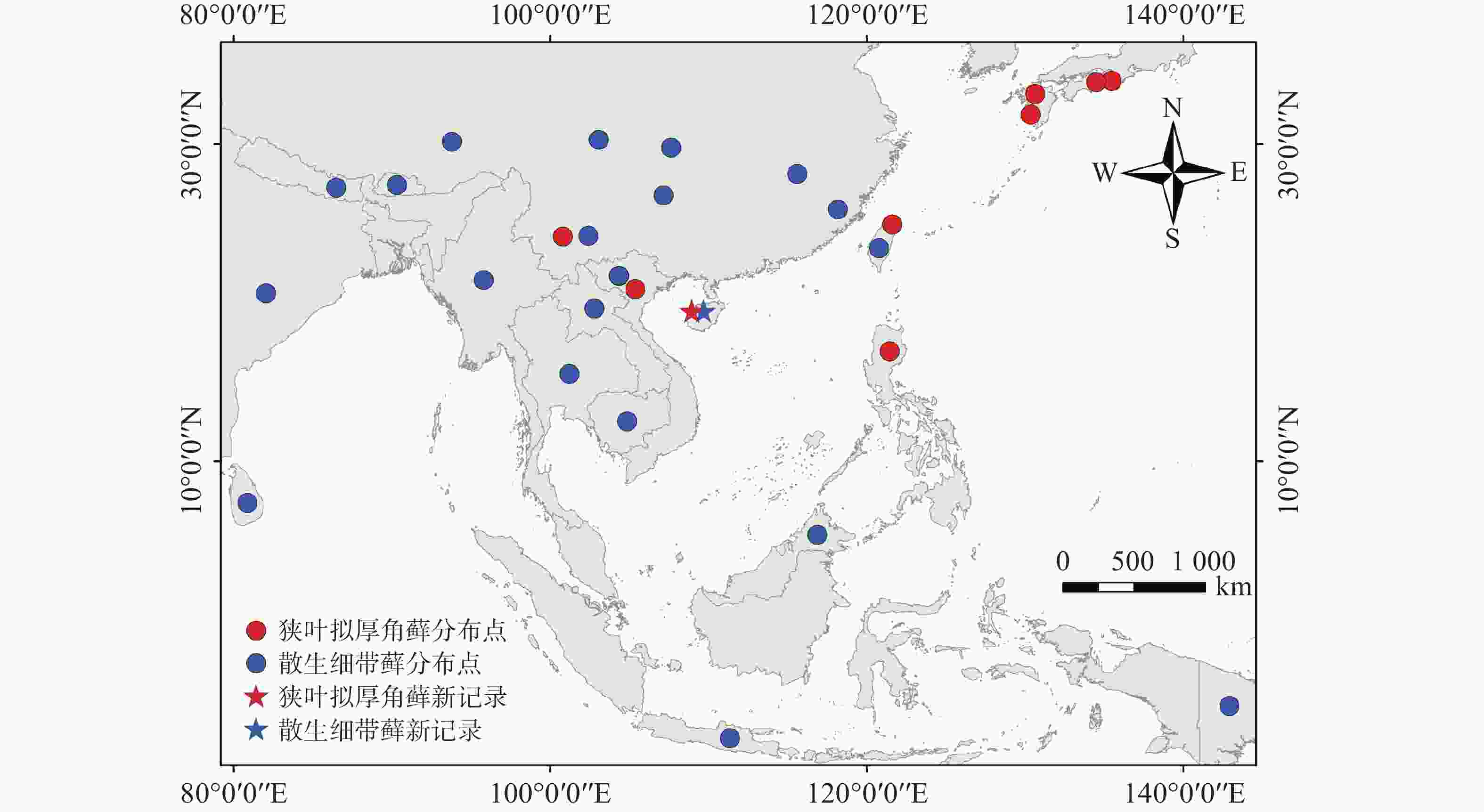
图 2 狭叶拟厚角藓和散生细带藓的世界分布图
Figure 2. Global distribution map of Yakushimabryum tonkinense and Trachycladiella sparsa
据载本种常在叶腋处着生丝状芽孢作为无性繁殖器官[8],但本研究中3份标本均未发现芽孢,而观察到多数植株在分枝的先端即生长芽的部分极易发生断裂(图1-A),形成带芽的断枝。这种情况也存在于同属的亚全缘拟厚角藓居群中[8],因此推测这些居群依靠枝条的断离片段而非芽孢作为无性繁殖器官进行营养繁殖。
狭叶拟厚角藓最早被列入疣胞藓属(Clastobryum Dozy & Molk.),TAN[12]将该种组合为狭叶厚角藓,AKIYAMA[8]又将其移入拟厚角藓属。狭叶拟厚角藓与亚全缘拟厚角藓在形态上相近而容易混淆,后者与前者的主要区别为分枝具细长悬垂末梢,叶基部边缘通常不具齿,丝状芽孢细胞为长矩形,壁薄而光滑。狭叶拟厚角藓与小厚角藓[Gammiella ceylonensis (Broth.) B.C.Tan & W.R.Buck]也较为相似,但后者个体较小(长约1 mm),叶为窄卵圆形至长圆形,叶先端具锐尖、急尖或短渐尖,可以从形态特点上进行区分。
《中国苔藓志》(第八卷)[13]和《广东苔藓志》[14]记载本种叶边缘有微齿或近全缘,生境为土面或岩石;而根据AKIYAMA的研究[8],狭叶拟厚角藓叶边缘均具齿,多数附生于森林内乔木或灌木的茎干和树枝上,稀岩面生;两者的形态学描述及生境记载有一定差异。推测由于前期本种的归属不清,且在形态上与近缘种极易混淆,亚洲及中国的部分记录可能存在对形态相近种的错误鉴定[8],因而笔者采用AKIYAMA最新的地理分布记录[8]。
-
Trachycladiella sparsa (Mitt.) M.Menzel, J. Hattori Bot. Lab. 75: 78. 1994; Moss Fl. China 5: 314. 2011.
Basionym: Meteorium sparsum Mitt., J. Proc. Linn. Soc., Bot., Suppl. 1(2): 158. 1859 ≡ Papillaria sparsa (Mitt.) Paris, Index Bryol. 909. 1897 ≡ Floribundaria sparsa (Mitt.) Broth., Nat. Pflanzenfam. I(3): 822. 1906.
Floribundaria chloronema (Müll.Hal.) Broth., Nat. Pflanzenfam. I(3): 822. 1906 ≡ Papillaria chloronema Müll.Hal., Bull. Soc. Roy. Bot. Belgique 34(2): 69. 1896.
Papillaria formosana Nog., Trans. Nat. Hist. Soc. Formosa 24: 119. f. l. 1934.
植物体黄绿色至深绿色,老标本变黄褐色,无光泽。主茎葡匐,长10 cm以上,稀疏分枝;分枝长0.3~2.5 cm,略呈圆条形,钝端。茎叶中上部伸展,阔心脏状卵形,或卵状披针形,常具波纹和弱纵褶,长0.5~2.4 mm,宽0.2~0.8 mm;基部宽阔,两侧圆钝,略下延,向上渐尖或骤缩呈长毛尖;边缘常波曲,具细齿;中肋单一,细弱,消失于叶中部以上,其上部常具细疣。叶细胞不透明,长菱形至线形,长30~70 μm,宽3~6 μm,壁厚,沿两侧壁密被细疣,边缘细胞均具密疣;基部细胞长方形,透明无疣,胞壁厚,具壁孔;角部细胞近方形,平滑。枝叶略小于茎叶,形态与茎叶相似。孢蒴未见。形态特征详见图3。

图 3 散生细带藓
Figure 3. Trachycladiella sparsa (Mitt.) M.Menzel
标本鉴定:海南昌江县霸王岭国家级自然保护区松林顶,19°5′1.44″ N,109°12′43.50″ E,海拔1 331 m,徐雪艳等20201119-106。
生境:树干附生。
地理分布:中国(江西、四川、重庆、贵州、云南、西藏、福建、台湾),尼泊尔,不丹,印度,缅甸,斯里兰卡,泰国,印度尼西亚,老挝,巴布亚新几内亚[15-16],马来西亚(婆罗洲)[17],柬埔寨[18],越南[19],海南首次记录(图2)。
细带藓属全世界有2种:细带藓[Trachycladiella aurea (Mitt.) M.Menzel)]和散生细带藓(T. sparsa),仅分布于亚洲热带和亚热带南部,在中国均有记载。它们共同的特征是叶细胞均狭长呈长菱形或线形,细胞腔窄,侧壁上密被细疣,使得细胞轮廓模糊。主要的识别点为:散生细带藓叶边缘细胞均具密疣(图3-N),而细带藓叶的边缘细胞一般透明无疣,形成明显的边缘。
-
狭叶拟厚角藓为东南亚−东亚分布,多见于海拔30~1 000 m,散生细带藓广泛分布于南海周边的热带和亚热带南部山区。海南前期仅报道尖峰岭有细带藓1种[20],此后未出现拟厚角藓属和细带藓属在海南分布的记录。近年来,笔者在海南热带山地云雾林发现狭叶拟厚角藓和散生细带藓的踪迹,推测可能因为气候变暖等环境变化,使得部分低海拔分布的物种向更高海拔山区以至山顶迁移,山地云雾林能够截留云雾中的水分,凉爽和湿润的生境为向上迁移的苔藓物种提供了适宜的生存场所,使得云雾林成为它们的避难所[21]。
致谢:海南热带雨林国家公园管理局霸王岭分局为本研究开展野外调查提供了帮助;中国科学院植物研究所贾渝研究员为拟厚角藓属新拟中文名并对本文撰写给予了指导。一并致谢。
New Data of Epiphytic Mosses in Hainan, China
-
摘要: 为了解海南热带雨林国家公园树附生藓类植物的资源分布和物种多样性,笔者于 2019—2020年多次在霸王岭国家级自然保护区热带山地云雾林开展树附生藓类植物资源调查。在野外采集、标本鉴定和查阅文献的基础上,发现拟厚角藓属(Yakushimabryum H.Akiyama, Ying Chang, T.Yamag. & B.C.Tan)为海南藓类植物新记录属,狭叶拟厚角藓[Yakushimabryum tonkinense (Broth. & Paris) H.Akiyama]和散生细带藓[Trachycladiella sparsa (Mitt.) M.Menzel]为海南新记录种。笔者对新记录属和种的形态特征进行了描述并提供了特征图版,对生境以及地理分布等信息进行了介绍。新记录的发现丰富了我国和海南树附生苔藓植物的生物多样性。Abstract: Field investigations were conducted in tropical montane cloud forests of Bawangling National Nature Reserve from 2019 to 2020 to explore the distribution and species diversity of epiphytic mosses in Hainan Tropical Rainforest National Park. Here, we firstly found and reported the genus Yakushimabryum H.Akiyama, Ying Chang, T.Yamag. & B.C.Tan and two species, namely Yakushimabryum tonkinense (Broth. & Paris) H.Akiyama and Trachycladiella sparsa (Mitt.) M.Menzel based on species taxonomic identification and pertinent literature, which were new additions to the bryoflora of Hainan Province, China. Their detailed information, including the description and pictures of morphological characteristics, habitats and geographical distribution were provided. The new records enriched the diversity of epiphytic bryophytes in Hainan and China.
-
Key words:
- new record /
- epiphytic mosses /
- Yakushimabryum /
- tropical montane cloud forest /
- Hainan
-
图 2 狭叶拟厚角藓和散生细带藓的世界分布图
Fig. 2 Global distribution map of Yakushimabryum tonkinense and Trachycladiella sparsa
图 3 散生细带藓
A. 干燥植株一段;B & C. 茎叶;D. 枝叶;E. 叶先端细胞;F. 叶上部细胞;G. 叶中部细胞;H. 叶中部具齿边缘;I & J. 叶基部中央细胞;K. 叶基部边缘细胞; L & M. 叶中部细胞具多疣;N. 叶边缘细胞具疣;O. 茎横切面。凭证标本:徐雪艳等20201119-106。
Fig. 3 Trachycladiella sparsa (Mitt.) M.Menzel
A. Portion of dry sterile plant. B & C. Stem leaves. D. Branch leaf. E. Apical cells of leaf. F. Upper cells of leaf. G. Median cells of leaf. H. Margin serrulate at mid-leaf. I & J. Central basal cells of leaf. K. Marginal cells of leaf base. L & M. Median laminal cells pluripapillose. N. Marginal laminal cells pluripapillose. O. Cross-section of stem. All from X.-Y. Xu et al. 20201119-106.
-
[1] MYERS N, MITTERMEIER R A, MITTERMEIER C G, et al. Biodiversity hotspots for conservation priorities [J]. Nature, 2000, 403(6772): 853 − 858. doi: 10.1038/35002501 [2] ALDRICH M, BILLINGTON C, EDWARDS M, et al. A global directory of tropical montane cloud forests [M]. Cambridge: UNEP-World Conservation Monitoring Center, 1997: 205–208. [3] 龙文兴, 丁易, 臧润国, 等. 海南岛霸王岭热带云雾林雨季环境特征[J]. 植物生态学报, 2011, 35(2): 137 − 146. [4] FREGO K A. Bryophytes as potential indicators of forest integrity [J]. Forest Ecology and Management, 2007, 242(1): 65 − 75. doi: 10.1016/j.foreco.2007.01.030 [5] BERDUGO M B, QUANT J M, WASON J W, et al. Latitudinal patterns and environmental drivers of moss layer cover in extratropical forests [J]. Global Ecology and Biogeography, 2018, 27(10): 1213 − 1224. doi: 10.1111/geb.12778 [6] AKIYAMA H, CHIANG Y, YAMAGUCHI T, et al. Yakushimabryum longissimum (Pylaisiadelphaceae) gen. & sp. nov., from the Yakushima Island, Japan [J]. Journal of Bryology, 2011, 33(1): 42 − 49. doi: 10.1179/1743282010Y.0000000002 [7] AKIYAMA H. Newly found sporophytes and male plants of Yakushimabryum longissimum (Pylaisiadelphaceae, Musci) and a significant extension of its distribution [J]. Bryological Research, 2014, 11(3): 63 − 69. [8] AKIYAMA H. Systematic study of the Yakushimabryum and related genera in the Pylaisiadelphaceae (Bryophyta) [J]. Acta Phytotaxonomica et Geobotanica, 2017, 68(3): 145 − 174. [9] SUZUKI T, INOUE Y, TSUBOTA H, et al. Notes on Aptychella (Sematophyllaceae, Bryopsida): Yakushimabryum longissimum, syn. nov. [J]. Hattoria, 2013, 4: 107 − 118. [10] AKIYAMA H, SCHÄFER-VERWIMP A, PRINTARAKUL N, et al. Phylogenetic study of the genus Aptychella (Pylaisiadelphaceae, Musci) [J]. The Bryologist, 2015, 118(3): 273 − 283. doi: 10.1639/0007-2745-118.3.273 [11] HAN W, JIA Y. Phylogeny and classification of the Sematophyllaceae s. l. (Hypnales, Bryophyta) [J]. Journal of Systematics and Evolution, 2020, 59(3): 524 − 540. [12] TAN B C. Six new taxa of Malesian mosses [J]. The Bryologist, 1990, 93(4): 429 − 437. doi: 10.2307/3243608 [13] JIA Y, WU P C, TAN B C. Sematophyllaceae [M] // WU P C, CROSBY M R, HE S. Moss Flora of China. New York: Missouri Botanical Garden Press, 2005: 33. [14] 吴德邻, 张力. 广东苔藓志[M]. 广州: 广东科技出版社, 2013: 461–462. [15] 贾渝, 何思. 中国生物物种名录: 第一卷 植物, 苔藓植物[M]. 北京: 科学出版社, 2013: 226. [16] 王立松, 贾渝, 张宪春, 等. 中国生物物种名录: 第一卷 植物, 总名录(上册) [M]. 北京: 科学出版社, 2018: 85. [17] ANDI M A M, SULEIMAN M. New and noteworthy Hypnales (Bryophyta) records from the Nuluhon Trusmadi Forest Reserve in Borneo [J]. Check List, 2020, 16(6): 1663 − 1671. doi: 10.15560/16.6.1663 [18] HIGUCHI M, SUZUKI T, BUCK W R, et al. A contribution to the knowledge of moss flora of Koh-Kong and Mondulkiri Provinces, Cambodia [J]. Bulletin of the National Museum of Nature and Science. Ser. B, 2020, 46(1): 9 − 15. [19] HE S, KHANG N S. New records and an updated checklist of the mosses of Vietnam [J]. Tropical Bryology, 2012, 34(1): 32 − 88. doi: 10.1179/1743282011Y.0000000038 [20] LIN P J, PIIPPO S, KOPONEN T, et al. Bryophyte flora of Jianfengling Mts, Hainan Island, China [J]. Bryobrothera, 1992, 1: 195 − 214. [21] BUBB P, MAY I, MILES L, et al. Cloud Forest Agenda [M]. Cambridge: UNEP-World Conservation Monitoring Centre, 2004: 1–32. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1131
- HTML全文浏览量: 591
- PDF下载量: 64
- 被引次数: 0
