

-
红脉穗螟[Tirathaba rufivena (Walker)]属鳞翅目(Lepidoptera)螟蛾科(Pyralidae),是槟榔的主要害虫之一,主要危害槟榔的花穗和果实,造成严重的落花落果。雌虫通常在佛焰苞基部产卵,幼虫孵化后钻入未展开的花苞内取食,使花苞枯萎,或从部分开放的佛焰苞入侵,啃食幼嫩的花穗,导致槟榔产量下降。当前对红脉穗螟的防治以喷洒农药为主。但槟榔树高可达10 m,且树干笔直,无枝条。依据棕榈科植物的特点,红脉穗螟为害的花苞部位随植株生长而随之长高。因此喷洒农药的操作难度大,成本高。且红脉穗螟幼虫大多隐藏在花苞中,喷洒的农药难以直接和虫体接触从而进行灭杀。使用传统农药还会促使昆虫产生抗药性,导致农药的防治效果减弱,并产生农药残留问题,对生态环境、人体健康等均产生有害作用,这不符合绿色植保理念。随着槟榔种植面积的扩大及绿色发展的需要,红脉穗螟的绿色防控已成为必要。当前对红脉穗螟绿色农药的开发中,植物源药剂如印楝素、鱼藤酮、茶皂素、烟碱已被证明对红脉穗螟具有生物活性[1-4]。此外,对红脉穗螟天敌的应用也是防治的一环。寄生蜂是在害虫防治中的一类常用天敌。目前已发现扁股小蜂[Elasmus punctulatus (VermaHayat)]、褐带卷蛾茧蜂[Bracon adoxophyesi (Mimanikawa)]、麦蛾柔茧蜂(Habrobracon hebetor)等对红脉穗螟都有寄生关系[5-7]。在寄生蜂之外,绿僵菌(Metarhizium)、垫跗螋[Chelisoches morio (Fabricius)]等均是可防治红脉穗螟的潜在生物天敌[8-9]。近年来,基于害虫嗅觉识别开发出的具有引诱或驱避作用的药剂也逐渐出现,在稻纵卷叶螟(Cnaphalocrocis medinalis) [10]、番茄潜叶蛾[Tuta absoluta(Meyrick)][11]、梨小食心虫(Grapholita molesta)[12]等害虫防治中均得到应用。这类药剂具有成本低、特异性强、防治效果好、对环境无污染的特点,是理想的防控措施。但目前基于红脉穗螟嗅觉识别机制开发其引诱或驱避剂的相关报道较少,该领域尚待研究。
红脉穗螟通过嗅觉识别槟榔花挥发物及信息素等信息化合物,精确定位寄主植物,并完成交配、产卵等关键生命活动[13-15]。嗅觉在昆虫寄主植物识别、交配、产卵和躲避天敌等行为中发挥着关键的作用[16]。昆虫嗅觉系统涉及多种嗅觉相关蛋白的参与,包括气味结合蛋白(odorant-binding protein,OBPs)、化学感觉蛋白(chemosensory protein,CSPs)、气味受体(odorant receptor,ORs)、离子型受体(ionotropic receptor,IRs)和感觉神经元膜蛋白(sensory neuron membrane protein,SNMP)[17-18]。气味结合蛋白OBPs在嗅觉信号传导过程中起着关键作用,其内部由多个疏水性氨基酸形成特定的结合位点,可结合一种或多种气味分子[19],并将其递送至感觉神经元树突上的嗅觉受体,实现化学信号向电信号的转换,进而将信号传递给更高一级神经中枢。OBPs是一类小分子水溶性球状蛋白,通常由150个左右的氨基酸组成,其N末端含有约20个氨基酸的信号肽序列。根据序列中半胱氨酸的数量可以将OBP分为以下类型:1)Classic OBP,含6个保守半胱氨酸残基,符合“C1-X25–30-C2-X3-C3-X36–42-C4-X8–14-C5-X8-C6”模式;2)Atypical OBP,含6个保守半胱氨酸残基,但其位置不符合上述模式;3)Plus-C OBP,保守半胱氨酸残基数量多于6个;4)Minus-C OBP:保守半胱氨酸残基缺失1~2个[20, 21]。在鳞翅目昆虫中,OBPs还可根据氨基酸序列的同源性划分为性信息素结合蛋白(pheromone binding protein,PBP)、普通气味结合蛋白Ⅰ(generalodorant-binding protein 1,GOBP1)和普通气味结合蛋白Ⅱ(general odorant-binding protein 2,GOBP2)[22]。目前,鳞翅目昆虫OBP的研究已较为深入,如草地贪夜蛾(Spodoptera frugiperda)[23]、李小食心虫(Grapholita funebrana)[24]、美国白蛾(Hyphantria cunea)[25]、栗叶瘤丛螟(Orthaga achatina)[26]、二点委夜蛾(Athetis lepigone)[27]、柑橘凤蝶(Papilio xuthus)[28]、桃蛀螟(Conogethes pinicolalis)[29]等。
目前关于红脉穗螟气味结合蛋白的研究中,王正利[30]用转录组测序技术获得红脉穗螟111个嗅觉相关蛋白基因序列,并在其中鉴定到11个OBP基因。周祥等[31]针对在雌虫中表达量较高的TrufOBP4进行了克隆与结合模式分析,揭示了TrufOBP4在寄主识别中的潜在功能。本研究针对两个在触角中表达量相对较高的TrufOBP1和TrufOBP5进行序列分析及原核表达,通过荧光竞争结合实验解析其与寄主植物挥发物及信息素的结合特性,并通过风洞试验验证筛选到的潜在配体对红脉穗螟的行为调控作用,初步阐明红脉穗螟识别寄主植物的分子机制,为开发基于行为调控的红脉穗螟绿色防控技术提供理论支撑。
-
红脉穗螟幼虫采集自海南省儋州市海南大学儋州校区槟榔林(19.51° N,109.49° E),将有受害状的槟榔花苞带回实验室后挑取将幼虫在饲养盒中进行人工饲养,环境条件为温度为(28±1) ℃,湿度为(80±10)%,光周期(L:D)14 h:10 h。幼虫饲喂人工饲料,成虫提供10%的蜂蜜水。
-
收集羽化后雌成虫,使用Trizol法提取红脉穗螟总RNA,使用Nanodrop 2000超微量分光光度计(ThermoFisher, Massachusetts,美国)检测测定RNA的纯度及浓度,RNA样品存储于−80 ℃以备后续实验。以提取的RNA为模板,利用PrimeScript RT reagent Kit试剂盒(Takara,北京,中国)进行cDNA合成,储存于−20 ℃。
-
利用NCBI ORF Finder(
https://www.ncbi.nlm.nih.gov/orffinder )预测TrufOBP基因的开放阅读框(Openreading frame,ORF);使用SignalP v4.1(https://services.healthtech.dtu.dk/services/SignalP-4.1/ )预测TrufOBP的信号肽;通过Expasy ProtParam(https://web.expasy.org/protparam )预测TrufOBP分子质量和等电点;利用 SOPMA(http://npsapbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_so pma.html )对TrufOBP氨基酸序列二级结构进行预测[31]。利用NCBI BLAST模块进行同源性搜索,并与数据库中鳞翅目昆虫的OBP序列进行多重比对,并使用软件MEGA v11构建系统发育树[32]。使用软件Snapgene设计特异性引物并构建原核表达载体,引物由擎科生物公司进行引物合成(表1)。表 1 本研究主要引物
Table 1. Primers in this study
引物 Primer 序列(5'-3') Sequence (5'-3') 内切酶 Endonuclease TrufOBP1-F CGCCATATGTTAACAGATGAGCAGAAAGAG Nde Ⅰ TrufOBP1-R AAATATGCGGCCGCCAGGAAGATGGCGTGCTT Not Ⅰ TrufOBP5-F CCGGAATTCGAAATTGAACAACTACAACC EcoR Ⅰ TrufOBP5-R AAATATGCGGCCGCGCATATCCATTCCGCAGC Not Ⅰ 注:酶切位点用下划线表示。
Note: The cleavage sites are indicated by underscores. -
以合成cDNA为模板,使用特异性引物对目的基因进行PCR扩增,扩增产物经琼脂糖凝胶电泳回收纯化后,与质粒载体进行双酶切(37 ℃ 15 min,65 ℃ 20 min),获得带黏性末端的目的基因片段。随后,使用T4 DNA ligase在24 ℃连接3 h,并将连接产物转化至大肠杆菌DH5α感受态细胞(天根,北京,中国)。挑取阳性单菌落进行测序验证。
-
将测序正确的质粒转化进BL21感受态细胞(天根,北京,中国)中。将成功转化质粒的BL21感受态接种于含卡那霉素(100 µg·mL−1)的LB液体培养基内,摇床(37 ℃,220 r·min−1)中培养12 h获得高浓度的菌液。在超净工作台中将菌液以1:100接入至放有LB液体培养基的培养瓶中,在37 ℃,220 r·min−1条件下培养至一定浓度,使用分光光度计在600 nm 波长下测量光密度,OD600在0.6~0.8,诱导蛋白表达。在TrufOBP1菌液中加入0.5 mmol·L−1 IPTG,16 ℃下诱导9 h,在TrufOBP5菌液中加入1 mmol·L−1 IPTG,20 ℃下诱导12 h。培养结束后离心收集沉淀。TrufOBP1使用NTA-0缓冲液(pH7.9)重悬菌体;TrufOBP5使用含8 mol·L−1尿素的PBS缓冲液(pH7.4)重悬菌液[33],超声破碎,取诱导前、后菌液、超声破碎上清和沉淀进行SDS-PAGE蛋白电泳检测。使用镍–亲和层析柱纯化TrufOBP,对各浓度洗脱液接取到各组分进行SDS-PAGE电泳检测,明确蛋白纯化情况。使用透析膜对蛋白进行纯化并进行SDS-PAGE电泳检测,最后使用考马斯亮蓝法测定蛋白浓度。
-
使用多功能酶标仪进行荧光竞争结合实验,设置荧光激发波长337 nm,发射波长380 nm,终止波长530 nm,间隔5 nm[34]。选择1-NPN作为荧光探针,以甲醇作为溶剂溶解1-NPN(400 μmol·L−1)。向200 μL蛋白溶液(2 μmol·L−1)中逐次加入1 μL 1-NPN探针,使探针终浓度为2~24 μmol·L−1。待测化合物用甲醇溶解,配置成400 μmol·L−1的工作液。将200 μL蛋白(2 μmol·L−1)与1 μL 1-NPN溶液(400 μmol·L−1)混匀静置2 min后测定荧光值,向混合液中逐次加入1 μL待测化合物,使化合物终浓度为2~24 μmol·L−1。每次混匀静置2 min后测量荧光强度,记录荧光值最大时的荧光波长与荧光值,重复3次[35]。使用斯卡查得方程计算1-NPN探针与TrufOBPs的结合常数(Kd)。根据配体的IC50(荧光值下降到一半时配体的浓度)和Kd值,计算配体与TrufOBPs结合的解离常数(Ki),计算公式:Ki=[IC50]/(1+[1-NPN]/Kd),其中[1-NPN]为游离荧光探针1-NPN的浓度。Ki<20表示蛋白与配体有强结合能力,20≤Ki<50表示蛋白与配体有弱结合能力,Ki≥50表示蛋白与配体不结合[36]。数据处理使用Graphpad v8.0.8软件进行计算与作图。
-
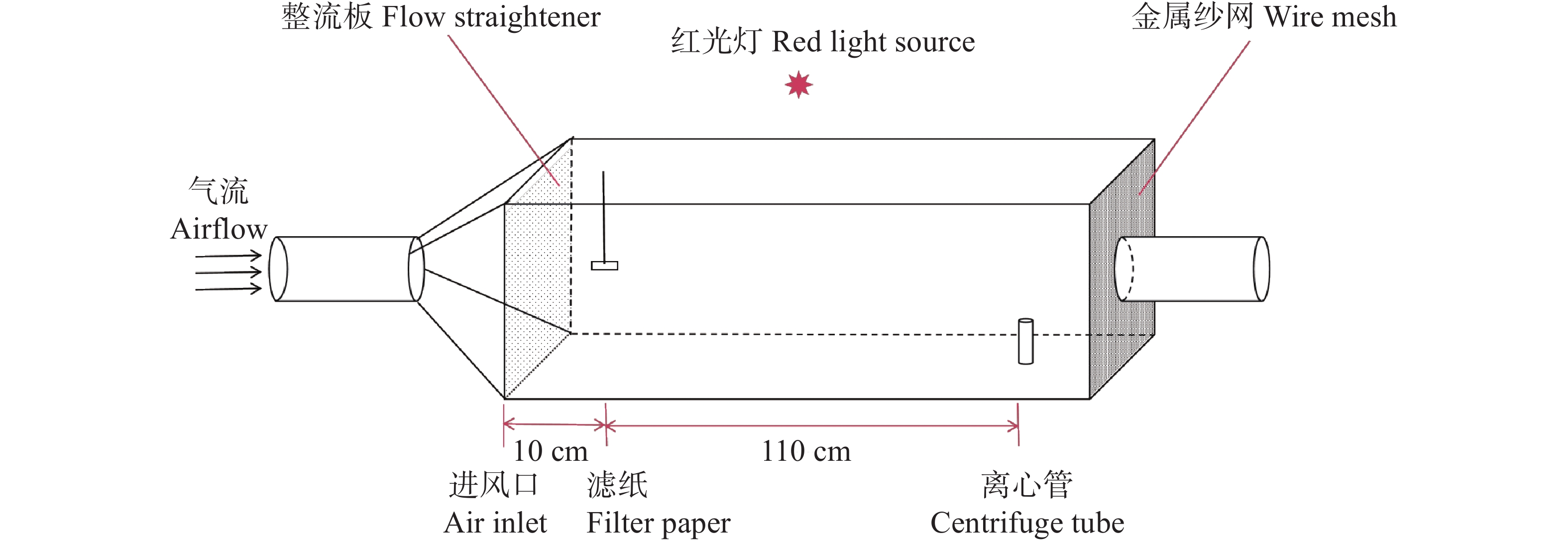
在150 cm × 60 cm × 60 cm的风洞中测定红脉穗螟雌成虫对不同化合物的行为反应(图1)。实验前用50 mL离心管装入2日龄未交配红脉穗螟雌成虫进行1 h的暗处理。以正己烷为溶剂,将各化合物配置成1 g·L−1测试质量浓度,将50 μL待测样品溶液滴于橡胶诱芯,并悬挂于距进风口中央。将暗处理后的红脉穗螟置于下风口,风速0.5 m·s−1,记录5 min内红脉穗螟的行为并计算各行为的反应,按以下标准进行记录:静止:试虫静止不动;振翅:试虫扇动翅膀但未离开离心管;起飞:试虫飞出离心管但与气味源的水平距离>90 cm;逆风飞行:90 cm≤试虫与气味源的水平距离<10 cm;接触源:试虫与气味源的水平距离≤10 cm [37]。最后计算红脉穗螟的各行为的反应率,使用SPSS v26软件进行卡方检验差异化分析[38],使用Graphpad v8.0.8软件进行作图。计算公式如下:
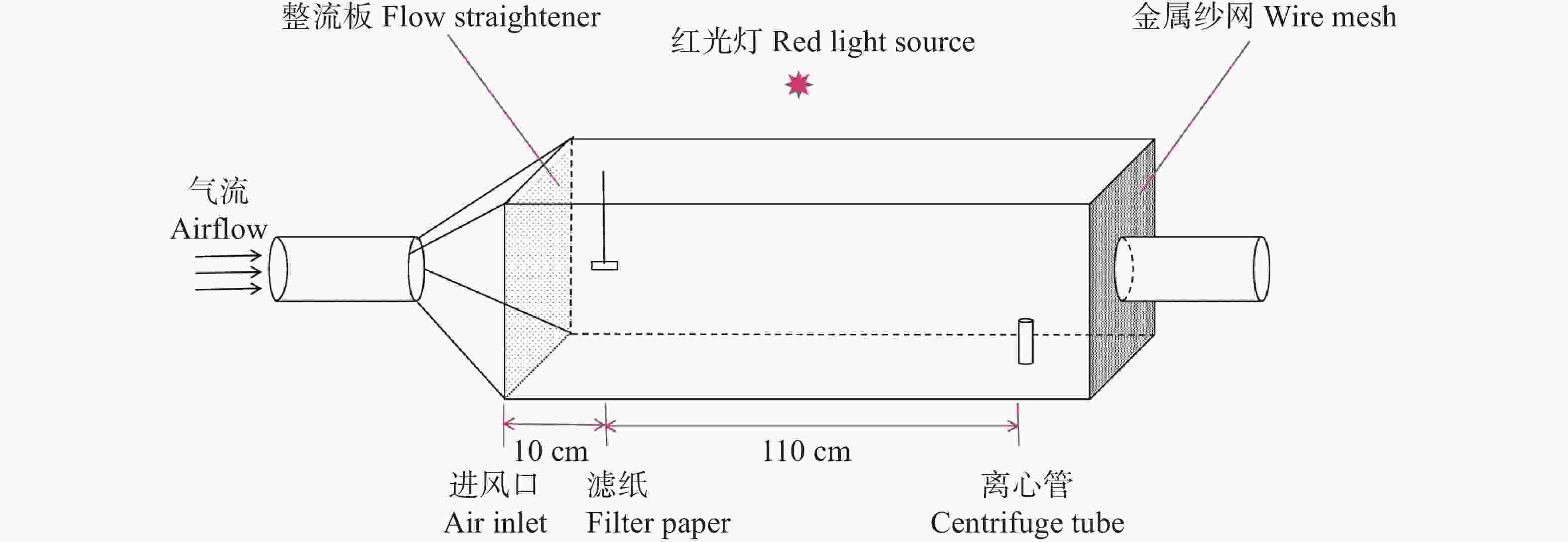
图 1 风洞装置示意图
Figure 1. Diagram of wind tunnel installation
选择率=(选择行为虫数/总虫数)×100%
-
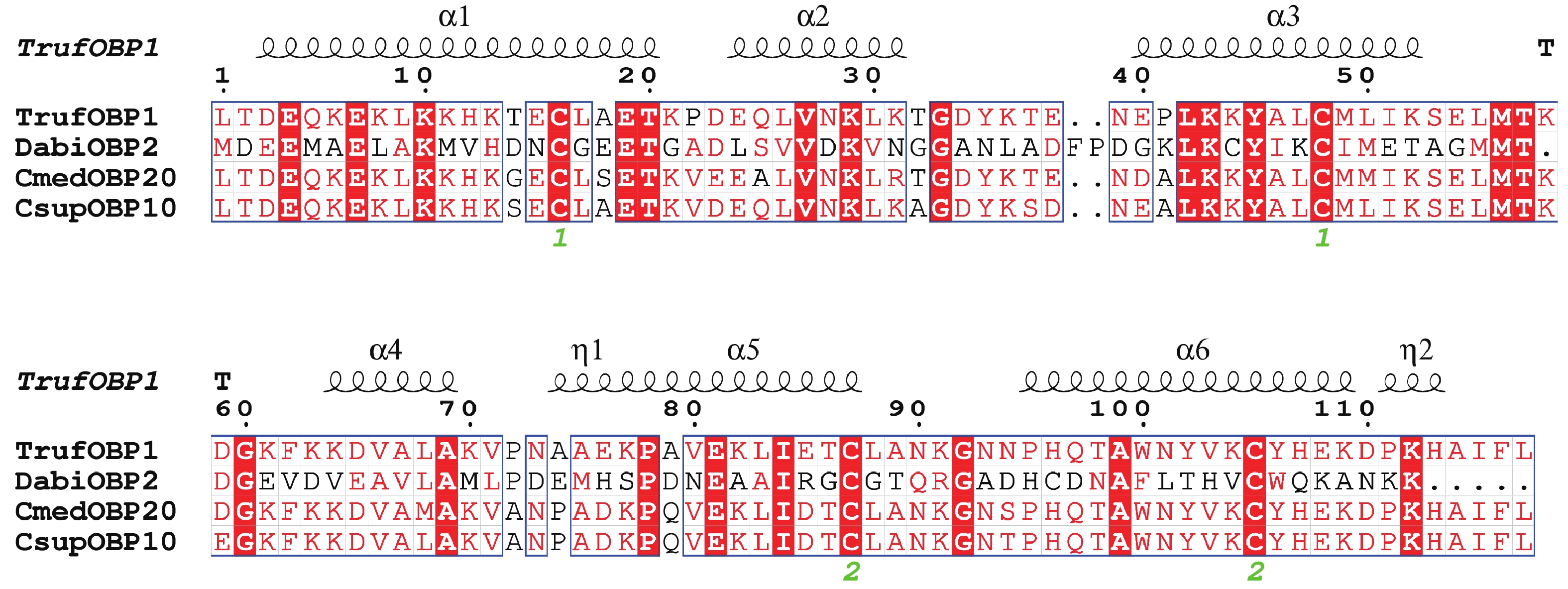
TrufOBP1基因的cDNA序列全长922 bp,其中完整开放阅读框ORF为402 bp,共编码133个氨基酸。预测其分子质量为14.8 kDa,理论等电点(pI)为9.00,为碱性蛋白。序列分析显示,该蛋白含有4个保守的Cys位点,属于Minus-C型OBP。信号肽预测表明其由16个氨基酸残基组成。总平均疏水性为−0.528,表明TrufOBP1具有亲水性。二级结构分析显示,该蛋白结构主要包括α−螺旋(56.03%)、β−转角(9.48%)、无规卷曲(30.17%)及延长链(4.31%)(图2)。
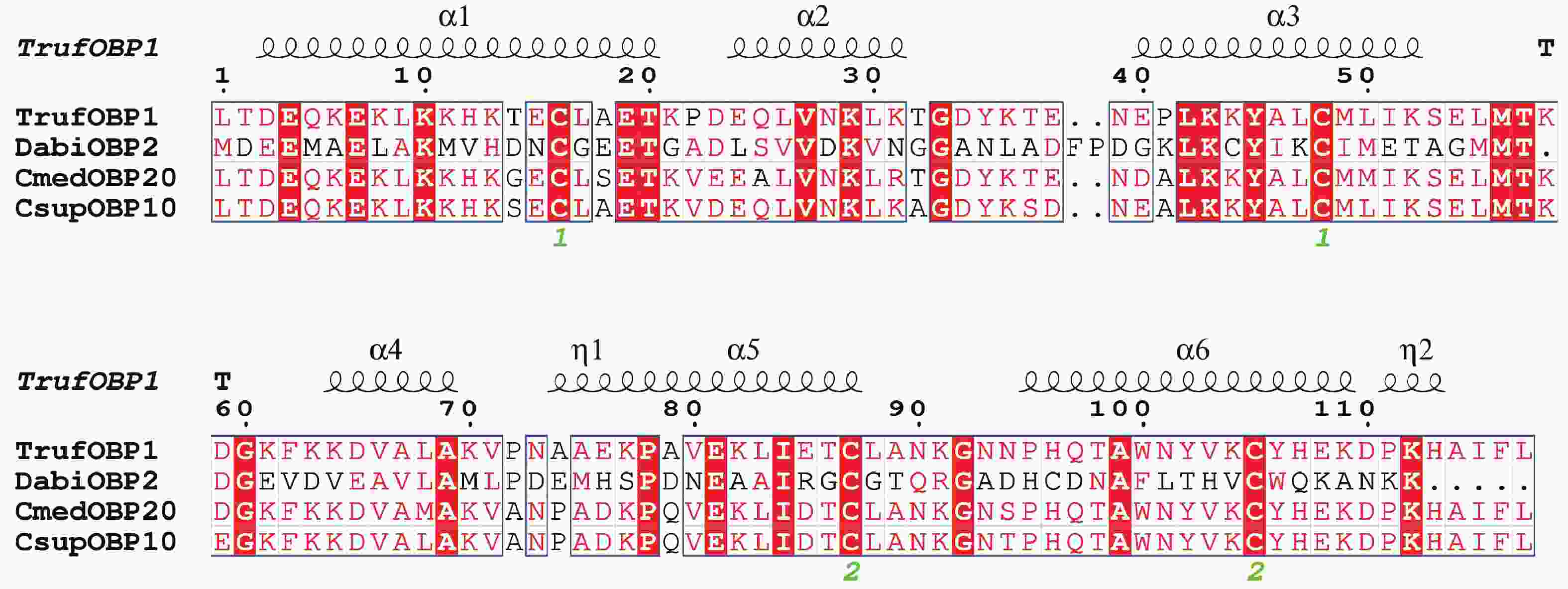
图 2 TrufOBP1与其他螟蛾科昆虫OBP的多序列比对
Figure 2. Alignment of TrufOBP1 with OBP genes from other insect species of Pyraloidae
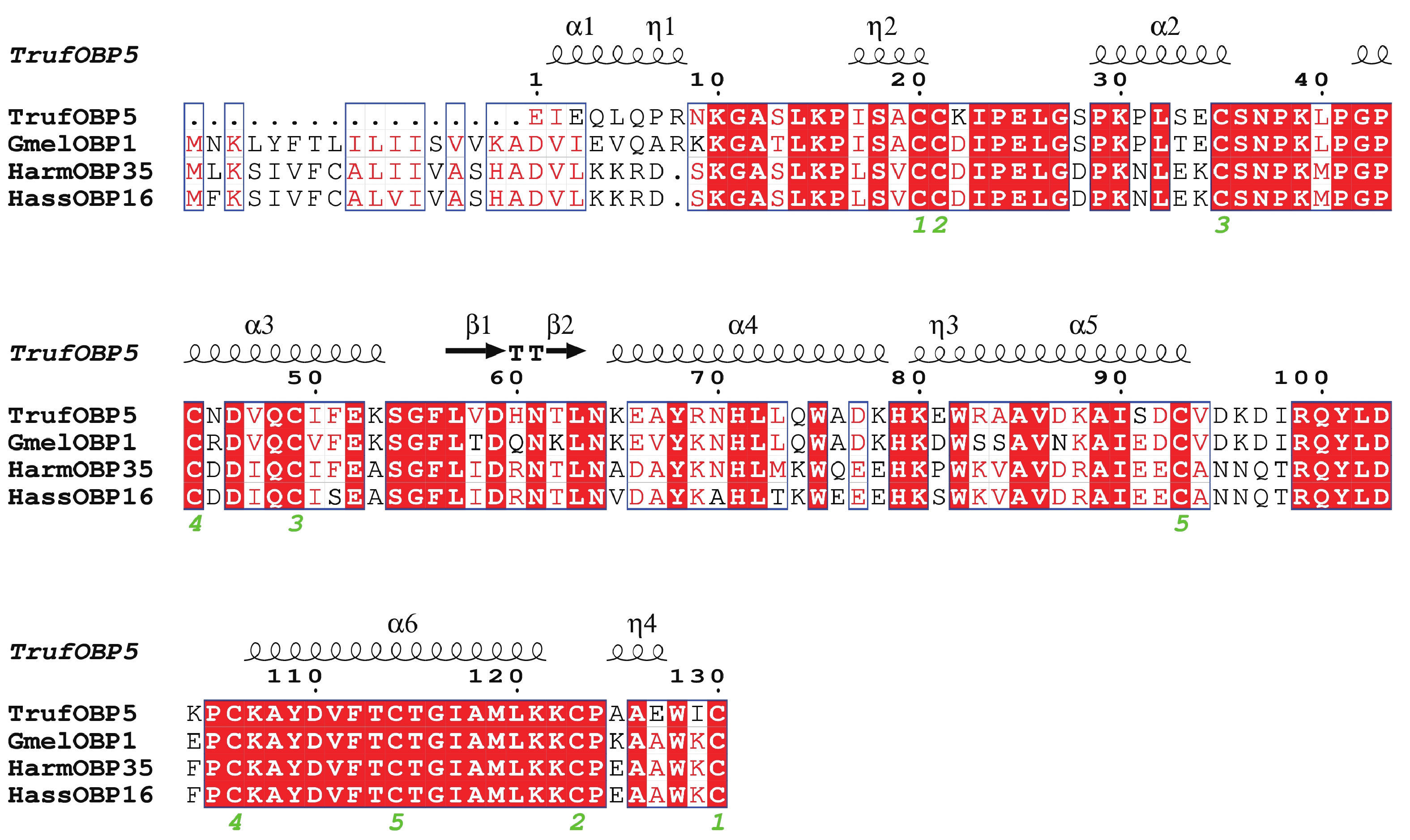
TrufOBP5基因cDNA序列全长为686 bp,开放阅读框全长为444 bp,编码147个氨基酸,分子量为21.0 kDa,理论等电点为8.3,为碱性蛋白。序列中有10个保守的Cys位点,为Plus-C型OBP。10个半胱氨酸形成五对二硫键(图3)。信号肽预测其含有17个氨基酸残基。总平均疏水性为−0.481,推测蛋白为亲水性蛋白。二级结构结果显示TrufOBP5蛋白主要结构为无规卷曲(50.0%),其次是α−螺旋(36.92%)、β−转角(6.15%)、延长链(6.92%)(图3)。
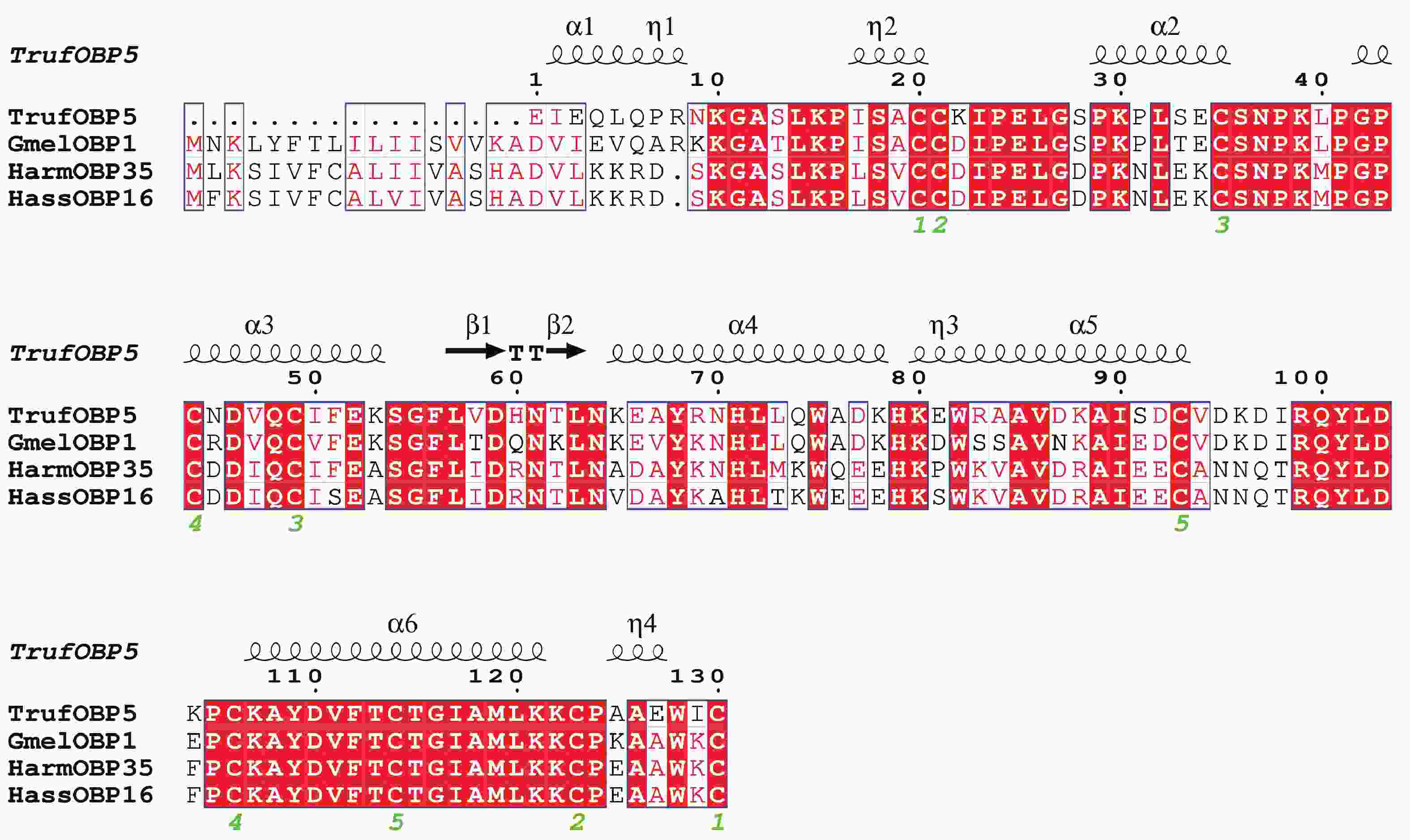
图 3 TrufOBP5与其他鳞翅目昆虫OBP的多序列比对
Figure 3. Alignment of TrufOBP5 with OBP genes from other insect species of Lepidoptera.
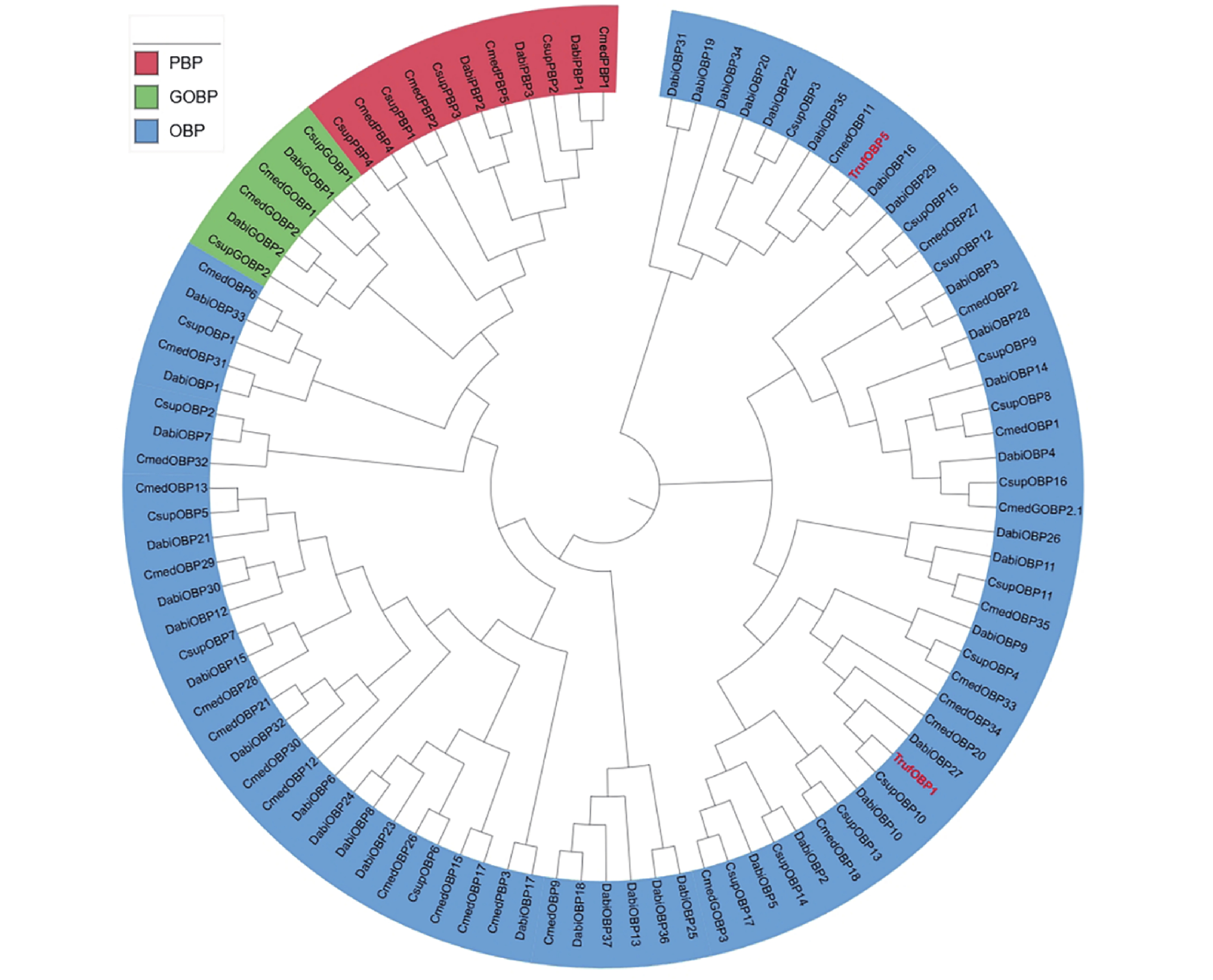
针对TrufOBP1、TrufOBP5和其他4种鳞翅目昆虫的OBPs构建了系统发育树,结果表明鳞翅目昆虫GOBP与PBP分别形成单独的聚类,其他OBP呈现分散的进化模式。TrufOBP1与TrufOBP5分散于OBP分支中,不属于GOBP或PBP类型。TrufOBP1与CsupOBP10聚为一支,关系最近,而TrufOBP5与DabiOBP16聚为一支(图4)。
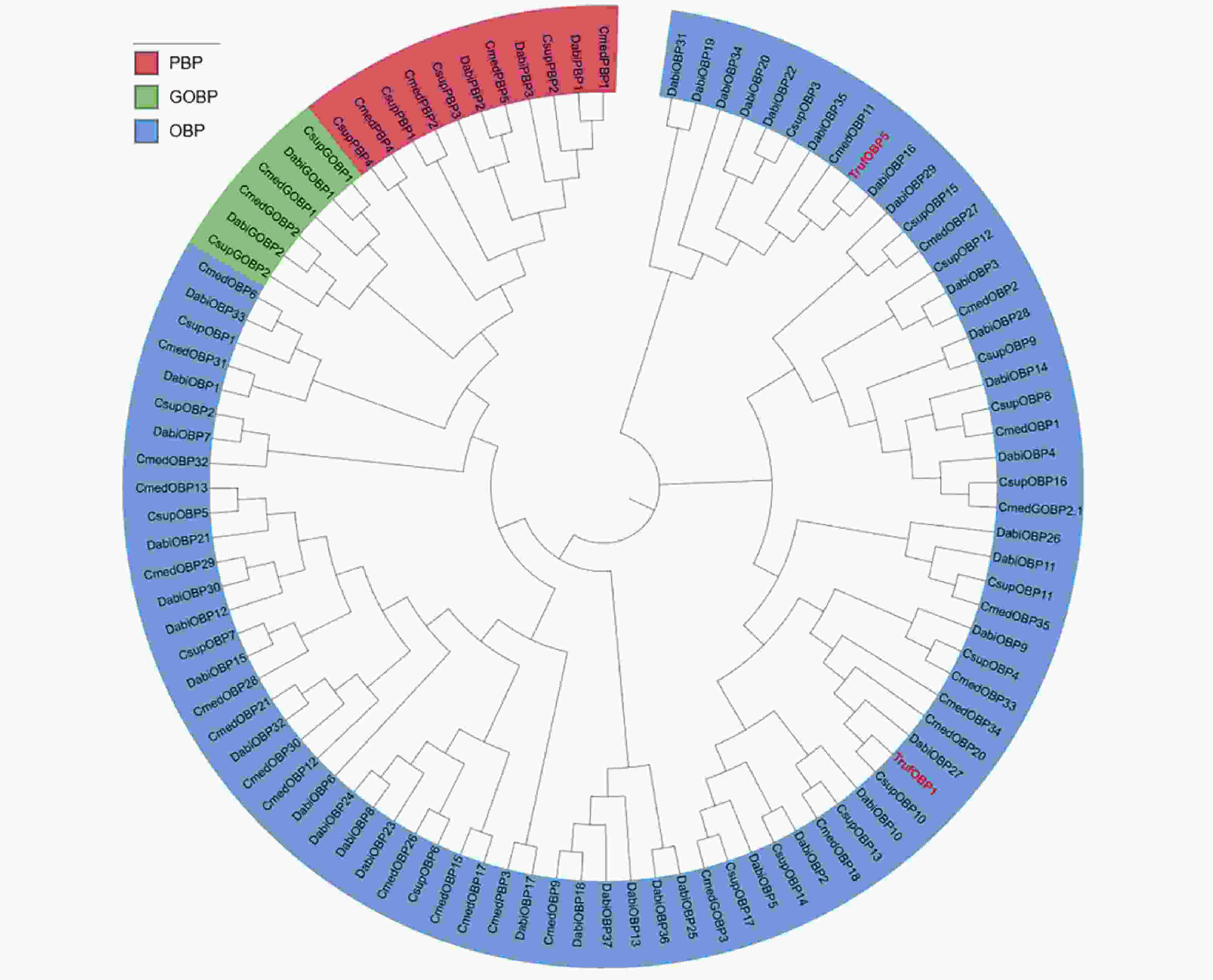
图 4 TrufOBP1和TrufOBP5的系统发育树
Figure 4. Phylogenetic tree of TrufOBP1 and TrufOBP5
-
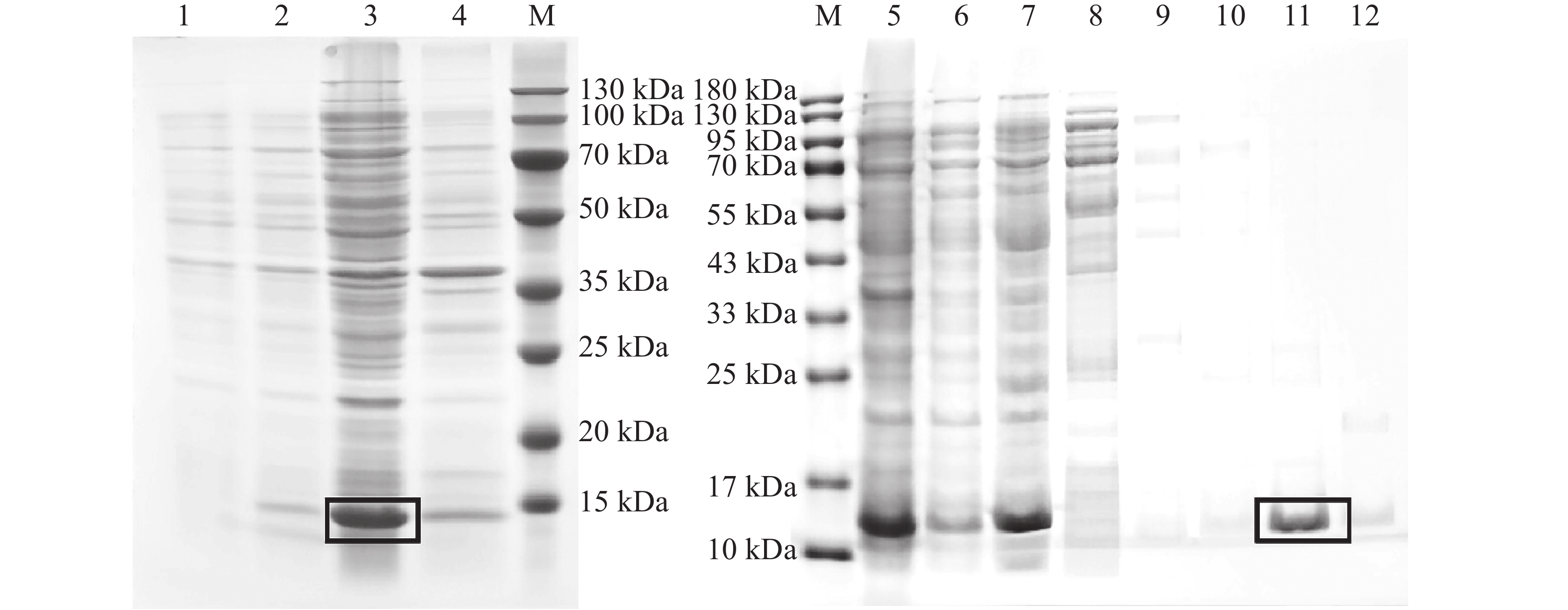
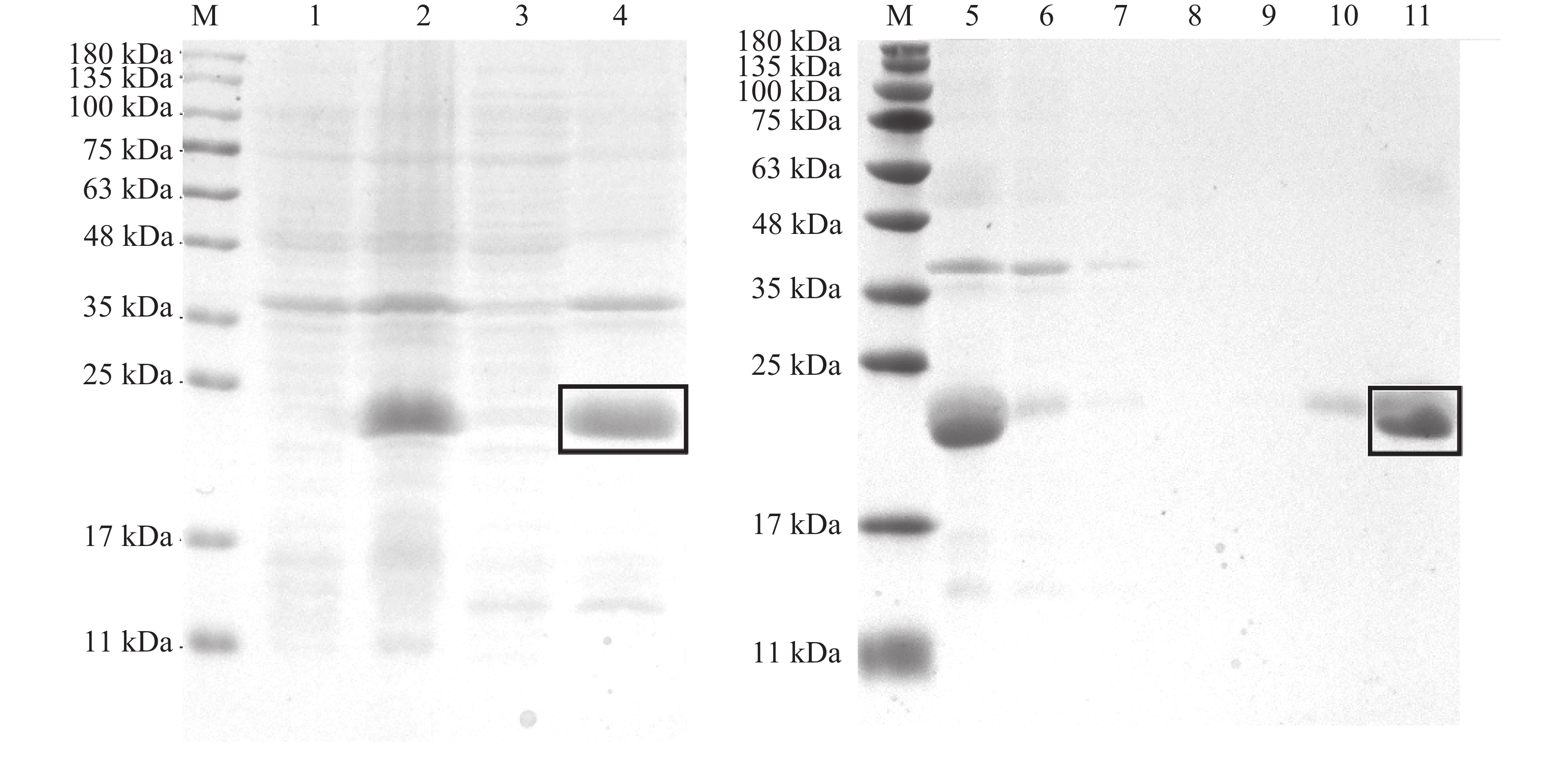
构建pET-30a(+)-TrufOBP1、pET-30a(+)-TrufOBP5重组质粒,将其转入BL21(DE3)感受态细胞后诱导表达。SDS-PAGE电泳结果显示TrufOBP1和TrufOBP5在对应分子质量位置均获得了清晰条带,表明蛋白成功表达,TrufOBP1主要在上清中,为可溶状态。TrufOBP5主要分布在沉淀中,为包涵体。蛋白纯化结果表明,TrufOBP1可被100 mmol·L−1咪唑洗脱液成功洗脱获得单一目的蛋白(图5)。TrufOBP5在200 mmol·L−1咪唑洗脱液成功洗脱(图6)。
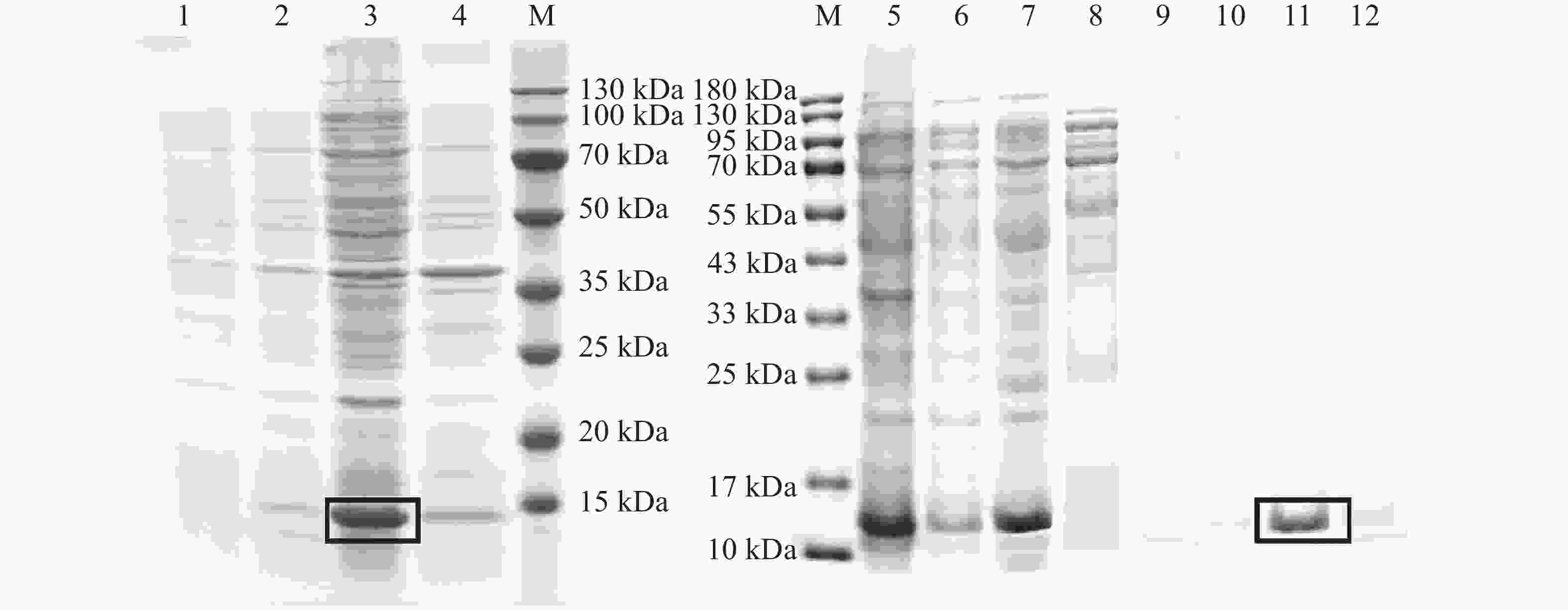
图 5 TrufOBP1诱导表达以及纯化效果分析
Figure 5. Expression and purification of TrufOBP1
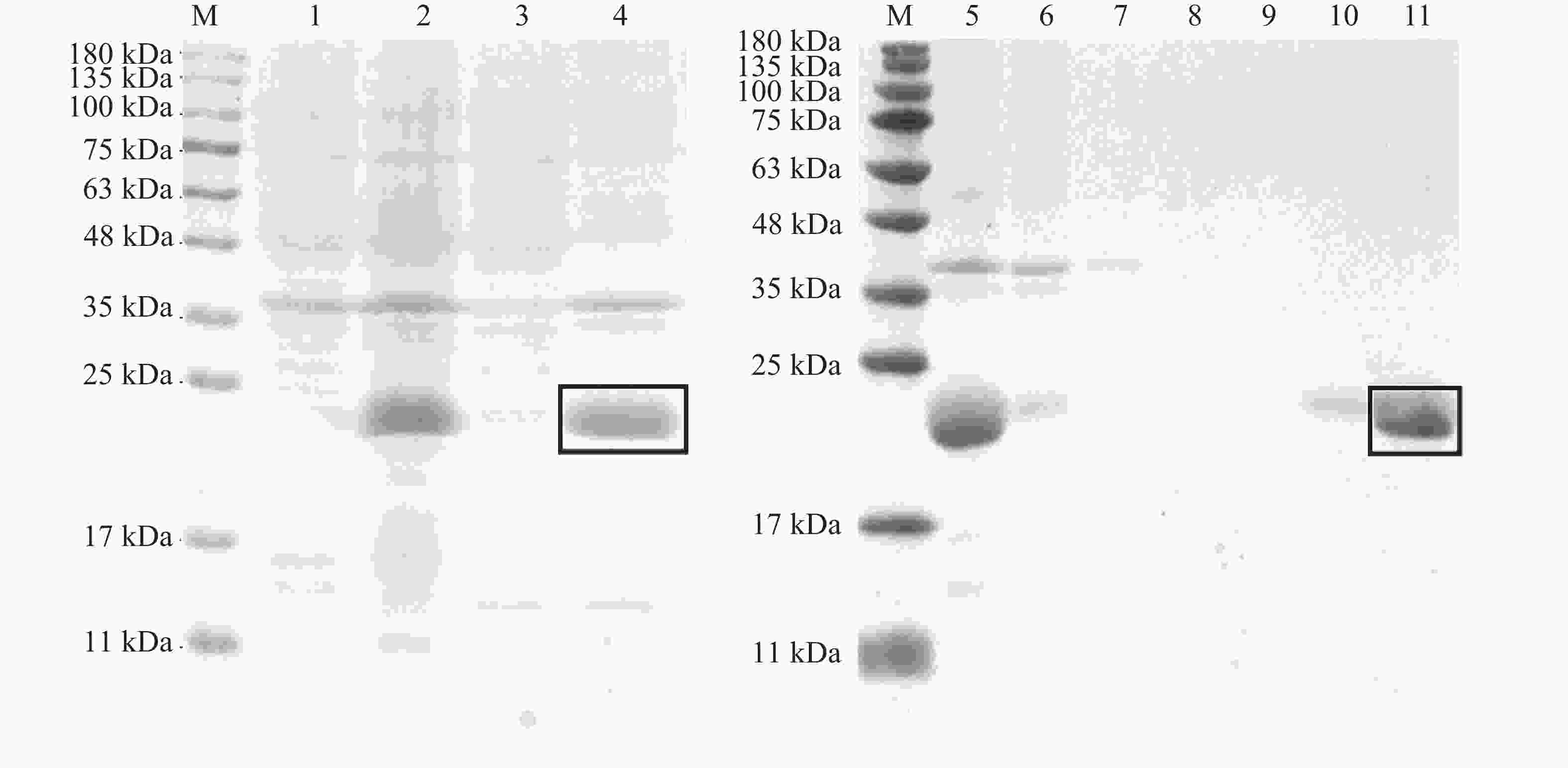
图 6 TrufOBP5诱导表达以及纯化效果分析
Figure 6. Expression and purification of TrufOBP5
-
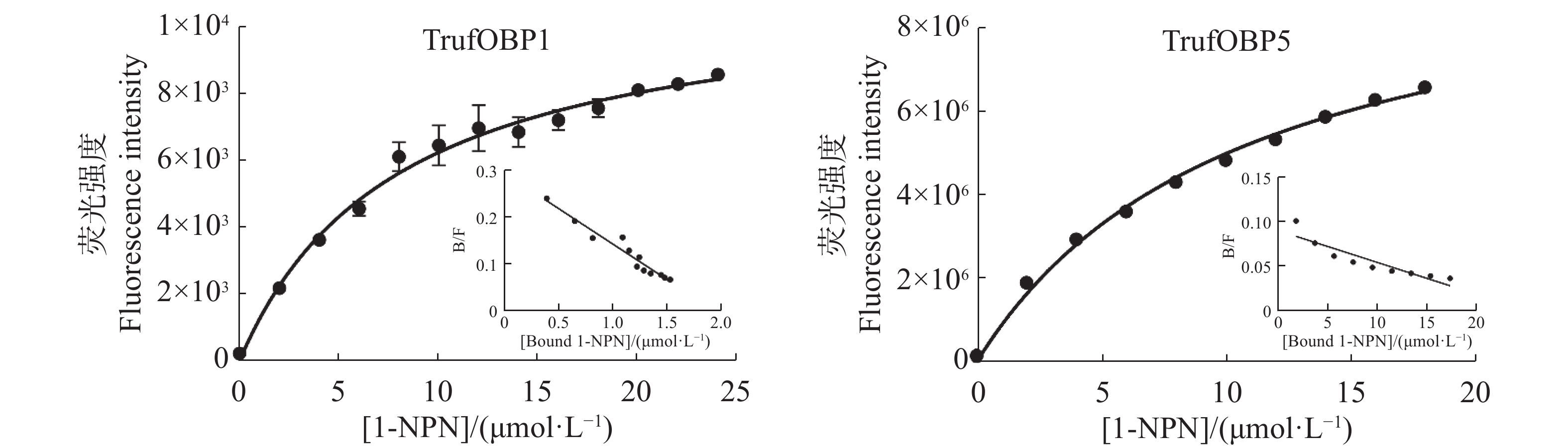
首先测定TrufOBP1、TrufOBP5与荧光探针1-NPN的结合常数,在337 nm的激发光下,TrufOBP1、TrufOBP5与1-NPN的发射光谱在峰值位于415 nm。使用Scatchard方程对数据进行拟合,计算得出TrufOBP1的Kd值为8.00 μmol·L−1,TrufOBP5的Kd值为10.68 μmol·L−1。1-NPN与两个蛋白结合存在线性关系,可用于后续竞争结合实验(图7)。
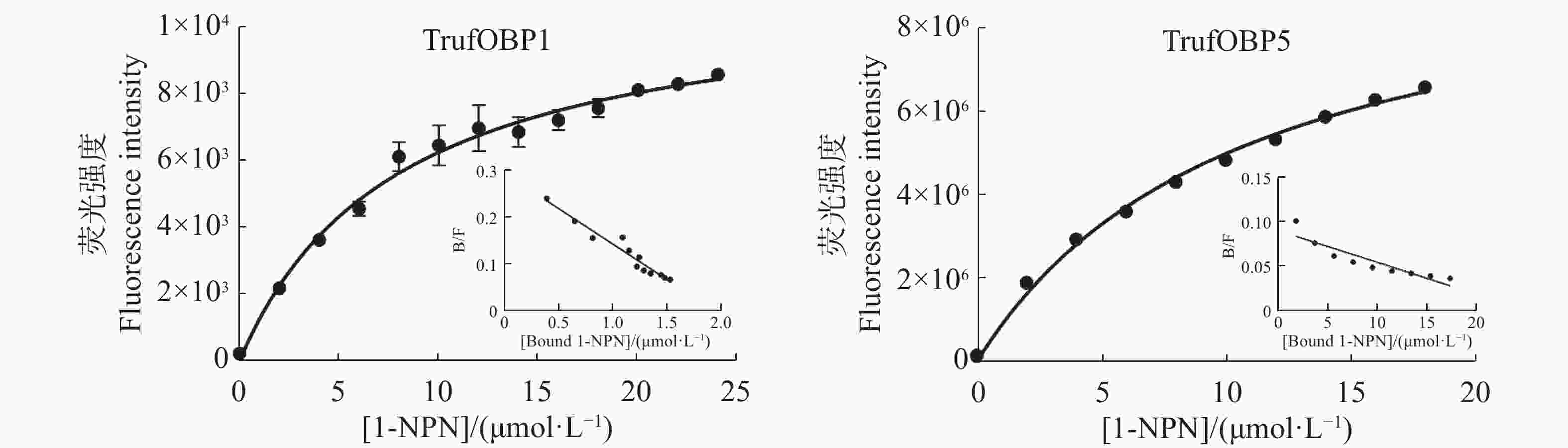
图 7 TrufOBP1、TrufOBP5与1-NPN探针的结合曲线和斯卡查德分析
Figure 7. The binding parameters of 1-NPN to TrufOBP1/TrufOBP5 and the Scatchard plots
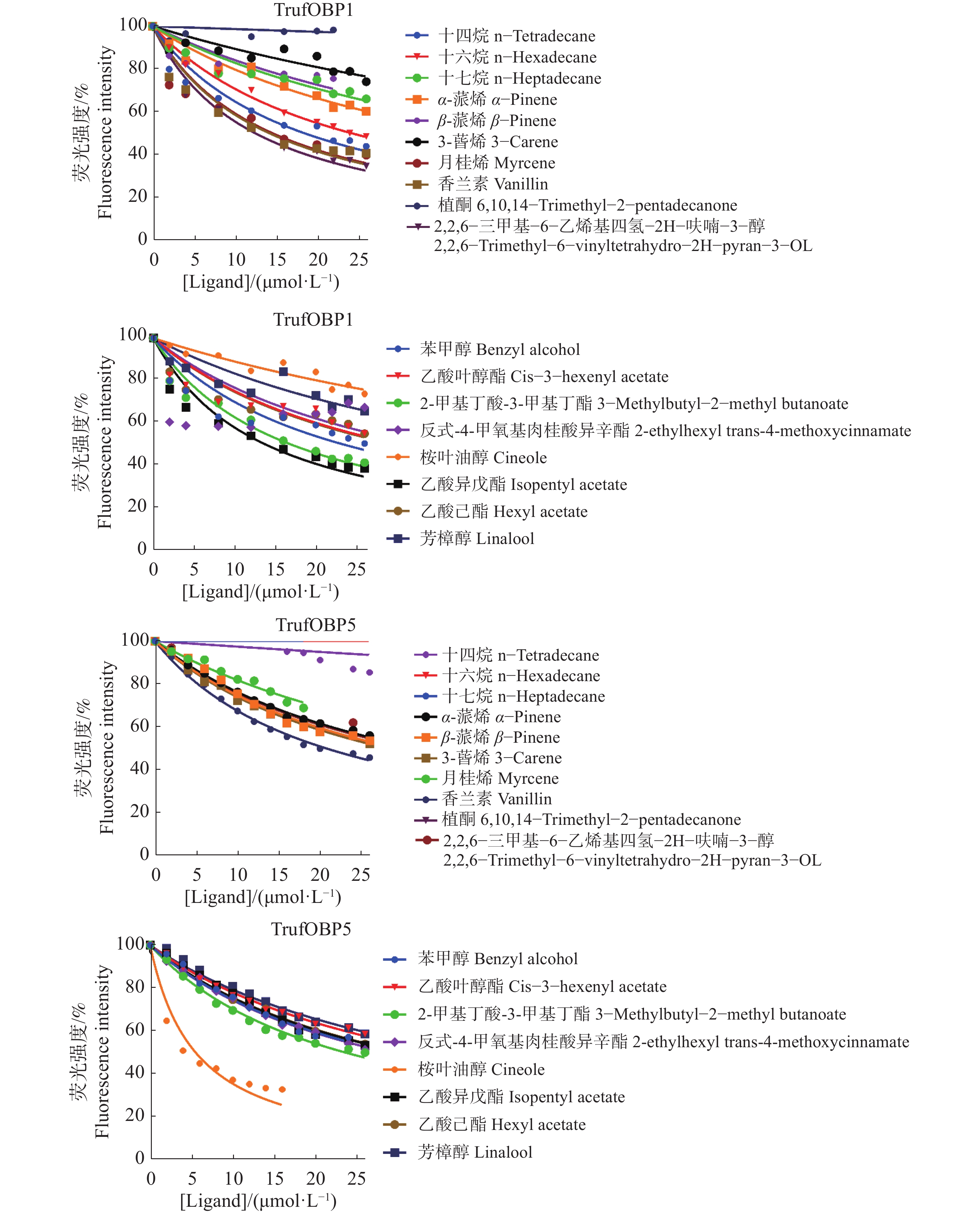
对TrufOBP1、TrufOBP5与18种配体进行荧光竞争结合分析,结果表明,TrufOBP1对11种配体表现出较强结合能力(表2)。其中与2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇结合能力最强。同时与5种配体呈现弱结合能力,与植酮、3−蒈烯不结合(图8);TrufOBP5与反式−4−甲氧基肉桂酸异辛酯有较强的结合能力[Ki = (5.16±1.05) μmol·L−1],其次为香兰素[Ki =(19.12±0.66) μmol·L−1]。此外,TrufOBP5与12种配体呈现弱结合能力(图8)。
表 2 TrufOBP与18种配体的荧光竞争亲和力
Table 2. Fluorescence competitive affinity of TrufOBP with 18 ligands
序号
Serial Number化合物
CompoundCAS Ki/(μmol·L−1) TrufOBP1 TrufOBP5 1 十四烷 n−tetradecane 629-59-4 11.34±1.10 >50 2 十六烷 n−hexadecane 544-76-3 14.58 ±1.22 >50 3 十七烷 n−heptadecane 629-78-7 28.74 ±1.51 >50 4 α−蒎烯 α−pinene 80-56-8 23.46±1.43 29.36±0.33 5 β−蒎烯 β−pinene 127-91-3 29.90 ±1.53 27.07±1.77 6 β−月桂烯 β−myrcene 123-35-3 8.83±1.35 41.34±3.79 7 3−蒈烯 3−carene 13466 -78-9>50 25.95±0.94 8 反式−4−甲氧基肉桂酸异辛酯
2-ethylhexyl trans-4-methoxycinnamate83834 -59-718.66 ±1.32 5.16±1.05 9 苯甲醇 benzyl alcohol 100-51-6 13.79 ±1.14 27.05±1.60 10 芳樟醇 linalool 78-70-6 29.52 ±1.52 26.61±0.76 11 桉叶油醇 cineole 470-82-6 47.88 ±1.73 28.24±1.49 12 乙酸异戊酯 isopentyl acetate 123-92-2 8.34 ±0.97 28.52±1.23 13 2−甲基丁酸−3−甲基丁酯
3−methylbutyl−2−methyl butanoate27625 -35-010.14 ±1.06 21.75±0.94 14 乙酸己酯 hexyl acetate 142-92-7 17.40 ±1.30 35.26±2.11 15 乙酸叶醇酯 cis−3−hexenyl acetate 3681 -71-817.75±1.3 32.59±1.09 16 香兰素 vanillin 121-33-5 8.65 ±0.99 19.12±0.66 17 2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇
2,2,6−trimethyl−6−vinyltetrahydro−2H−pyran−3−OL14049 -11-77.66 ±0.94 28.83±1.79 18 植酮 6,10,14−trimethyl−2−pentadecanone 502-69-2 >50 >50 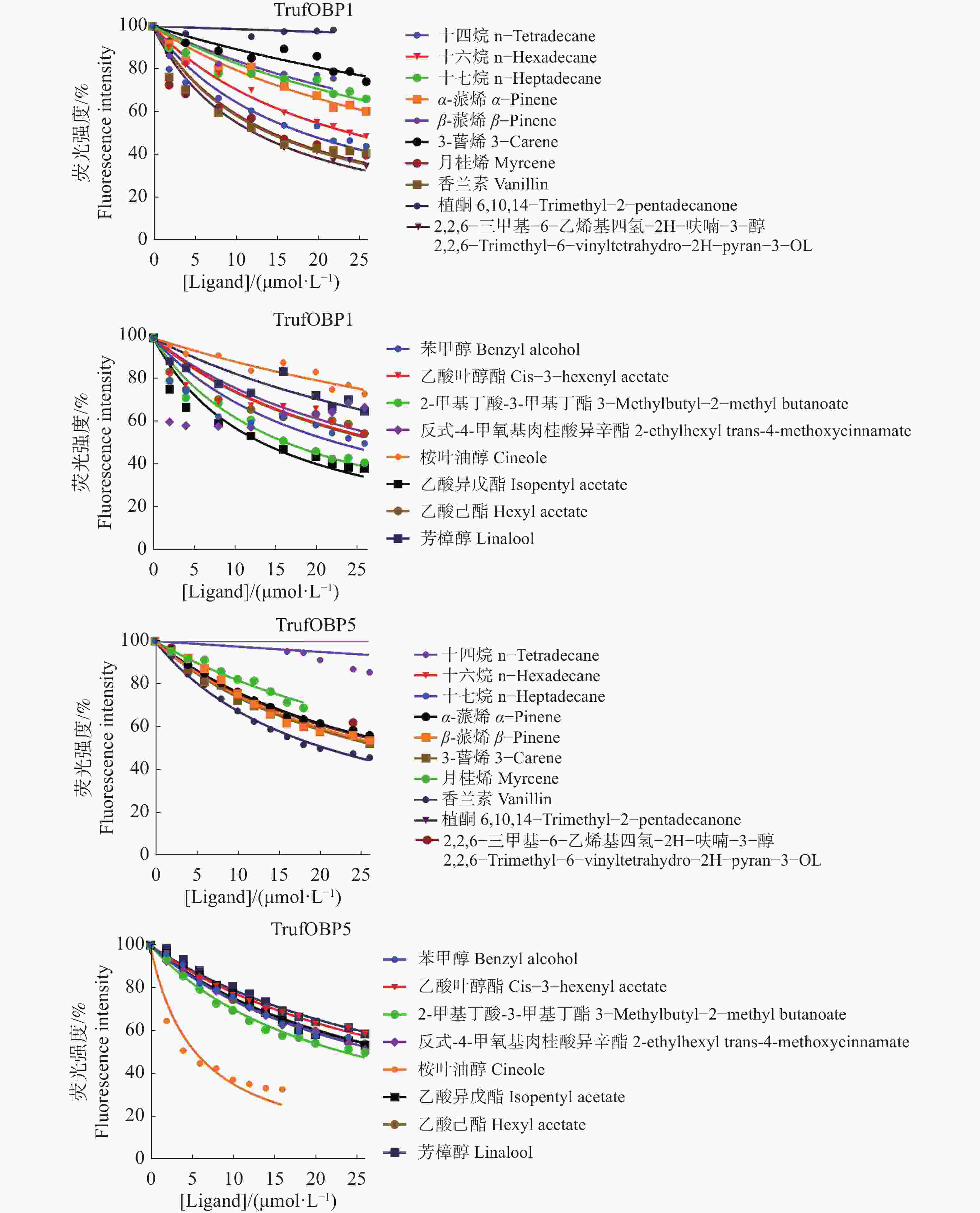
图 8 TrufOBP的荧光竞争结合曲线
Figure 8. Fluorescence competitive binding curve of TrufOBP
-
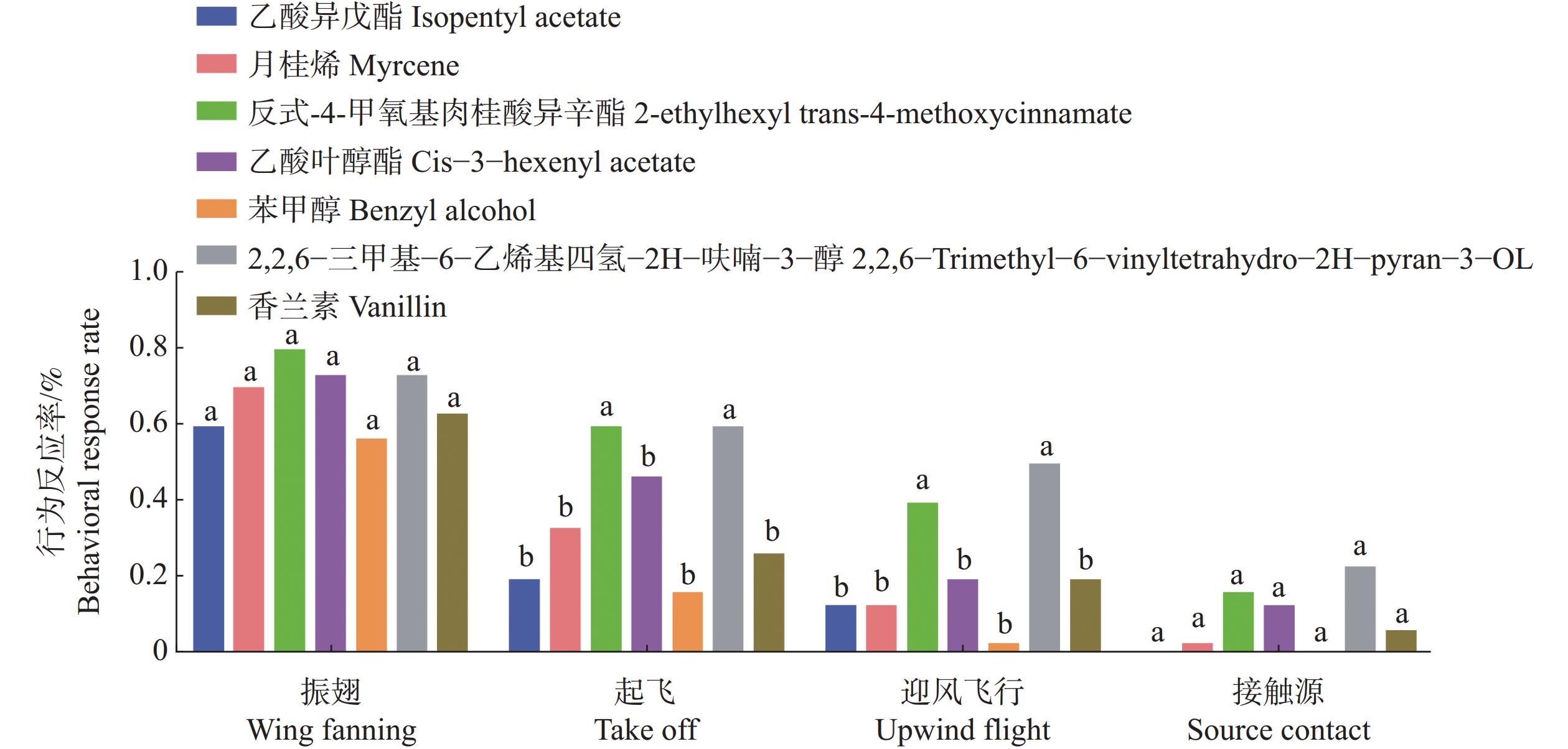
基于荧光竞争结合实验结果,筛选了7种与TrufOBP1、TrufOBP5具有较强结合能力的化合物进行红脉穗螟雌成虫的风洞行为实验。实验结果表明反式−4−甲氧基肉桂酸异辛酯和2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇对红脉穗螟表现出较强引诱作用,在起飞和迎风飞行的行为选择中和其他化合物有显著差异。且接触源的行为选择也高于其他化合物。其中2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇的引诱作用最强(图9)。
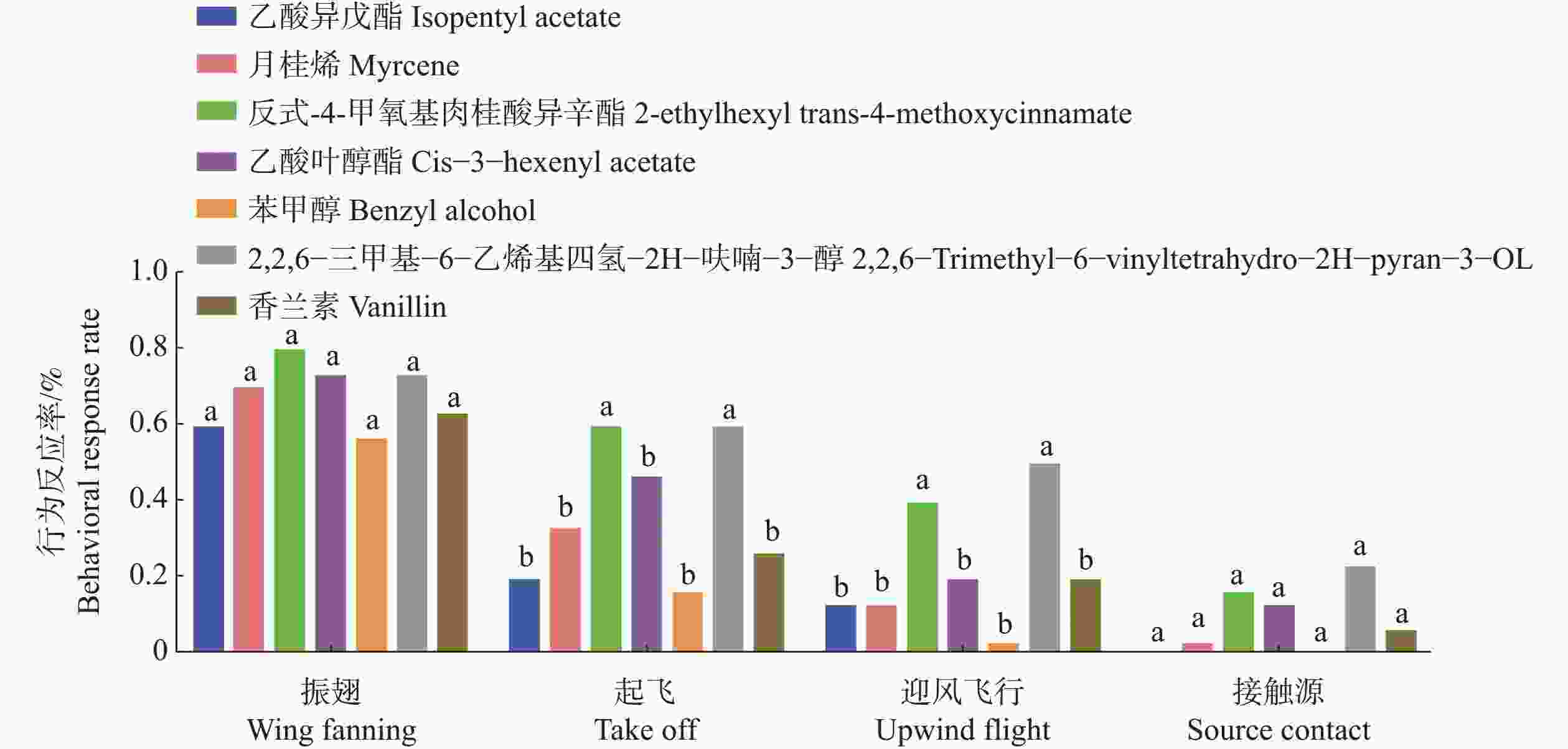
图 9 雌成虫对7种化合物的风洞行为反应
Figure 9. Wind tunnel behavioral responses of female adults to 7 compounds
-
气味结合蛋白在昆虫嗅觉识别过程中发挥着重要作用,介导昆虫对寄主植物和性信息素的感知。本研究对红脉穗螟两个气味结合蛋白TrufOBP1和TrufOBP5进行了序列特征及结合特性研究,并结合风洞行为实验,探讨其在寄主识别和性信息素感知中的潜在作用。序列分析结果表明,TrufOBP1具有4个保守半胱氨酸位点,与经典OBP(Classic OBP)相比,其第2位与第5位半胱氨酸被其他氨基酸取代,符合Minus-C 型OBP的特征。在桔小实蝇(Bactrocera dorsalis)[39]、枣实蝇(Carpomya vesuviana)[40]、云斑天牛(Batocera horsfieldi)[41]、西方蜜蜂(Apis mellifera)[42]及棉铃虫[43]等昆虫中均有关于Minus-C OBP的研究。研究表明,Minus-C型OBP在较为原始的昆虫中较为丰富,而Plus-C型OBP在进化程度较高的昆虫中分布较广,Plus-C型和Classic型的OBP可能是Minus-C型OBP的进化衍生物[42, 44]。在二化螟[Chilo suppressalis (Walker)]中鉴定到1个Minus-C型的CsupOBP1,研究发现其对寄主挥发物β-紫罗兰酮有较强结合能力[45];梨小食心虫GmolOBP14被鉴定为Minus-C型OBP,对寄主挥发物梨酯和十二醛表现出强烈的结合活性,对其他性信息素和气味物质不结合[46],以上研究表明这些Minus-C型OBP在寄主植物气味识别中有高度特异性结合的作用。相比之下,TrufOBP5具有10个保守的半胱氨酸残基,可形成5对二硫键,属于Plus-C型OBP。这类OBP主要分布于进化程度较高的昆虫,相关研究相对较少。暗黑鳃金龟(Holotrichia parallela)的HparOBP14作为Plus-C OBP,能够结合植物挥发物和有机肥挥发物,并在昆虫产卵地选择中发挥重要作用[47]。
荧光竞争结合实验表明,TrufOBP1与11种配体表现出较强结合能力,包括9种寄主挥发物和2种信息素组分,表现出较宽的结合谱,与其他一些鳞翅目昆虫存在差异。TrufOBP5仅与两种化合物呈现出强结合能力,其中与反式−4−甲氧基肉桂酸异辛酯的Ki值为(5.16±1.05)μmol·L−1,结合能力远高于其他化合物。这种特异性结合能力可能与其结构有关,Plus-C型OBP由于额外的二硫键可能形成独特的配体结合腔,从而提高了对特定化合物的亲和力。TrufOBP1也对反式−4−甲氧基肉桂酸异辛酯表现出较强的结合能力,表明该化合物在红脉穗螟寄主识别过程中可能具有重要作用。王正[30]发现TrufOBP4对性信息素组分中的植酮和香兰素具有强结合能力,都梓萌[48]研究表明TrufOBP7也能够与香兰素强结合。结合本研究结果,香兰素可能是性信息素中的主要组分。
风洞行为实验结果明确了5种化合物对红脉穗螟雌成虫有较好引诱活性,包括3种植物挥发物和2种信息素。实验发现寄主挥发物中β−月桂烯、反式−4−甲氧基肉桂酸异辛酯、乙酸叶醇酯均可成功引诱红脉穗螟雌成虫、其中反式−4−甲氧基肉桂酸异辛酯的引诱效果最强,这一结果与荧光竞争结合实验相一致,表明TrufOBP1和TrufOBP5可能在昆虫感知寄主植物挥发物的过程中协同作用。这些寄主挥发物在昆虫寄主选择过程中可能发挥关键作用,可用于未来害虫防控中的行为调控。此外,2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇和香兰素在风洞实验中也表现出较强的引诱活性。Sasaerila等[49]研究发现,在红脉穗螟的近缘种油棕蛀果穗螟(Tirathaba mundella)中,这两种化合物也被鉴定为其性信息素成分,该信息素组分在近缘物种中具有一定的保守性。这一结果进一步验证了2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇和香兰素可能是红脉穗螟性信息素的主要组分。TrufOBP1对这两种化合物均具有较高的结合能力,表明其不仅在红脉穗螟识别寄主挥发物中发挥作用,也可能在性信息素感知中发挥重要作用[50-51]。能够同时结合寄主植物挥发物和性信息素的OBP 在昆虫中多有报道。例如,二点委夜蛾雄蛾的AlepGOBP2能够结合性信息素Z7-12∶Ac、Z9-14∶ Ac及玉米挥发物罗勒烯和(E)−β−法尼烯[27];而小菜蛾(Plutella xylostella)幼虫中的PxylGOBP2对性信息素和寄主植物气味均表现出较好的结合能力[52],这表明部分OBP可能在昆虫的多重感知过程中发挥作用。
本研究通过分析两个TrufOBPs的序列特征、配体结合能力及行为反应,系统探讨了其在红脉穗螟寄主识别和性信息素感知中的作用,对开发高效的红脉穗螟绿色农药具有重要意义。目前同源建模、分子对接技术等在探究OBP与小分子物质结合模式的分析中已广泛应用。后续将通过更丰富的研究技术解析气味结合蛋白与配体的结合机制,为利用昆虫嗅觉调控技术进行害虫防治提供理论依据。
红脉穗螟两个气味结合蛋白的序列特征及结合特性分析
DOI: 10.15886/j.cnki.rdswxb.20250041
 CSTR: 32425.14.j.cnki.rdswxb.20250041
CSTR: 32425.14.j.cnki.rdswxb.20250041
Sequence characterization and ligand binding analysis of two odorant-binding proteins in Tirathaba rufivena (Walker)
-
摘要: 昆虫嗅觉系统在寄主识别、交配和产卵等行为中发挥关键作用,其中气味结合蛋白(odorant-binding proteins,OBPs)在嗅觉信号传递过程中起重要作用。本研究针对红脉穗螟(Tirathaba rufivena)两个气味结合蛋白TrufOBP1和TrufOBP5进行了序列特征及与配体结合特性分析,并通过风洞实验验证其在行为调控中的作用。序列分析表明,TrufOBP1具有4个保守半胱氨酸,属于Minus-C型OBP;TrufOBP5具有10个保守半胱氨酸,属于Plus-C型OBP。荧光竞争结合实验结果表明,TrufOBP1具有较广的结合谱,与11种配体结合,其中包括9种槟榔花挥发物和2种性信息素;TrufOBP5结合特异性强,仅与反式−4−甲氧基肉桂酸异辛酯和香兰素结合,推测其可能在寄主植物识别过程中发挥重要作用。风洞实验发现:寄主挥发物中,β−月桂烯、反式−4−甲氧基肉桂酸异辛酯和乙酸叶醇酯可引诱红脉穗螟雌成虫。性信息素组分2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇引诱效果最强,此外,香兰素也表现出引诱活性。本研究揭示了TrufOBPs在红脉穗螟寄主识别和性信息素感受中的作用,为害虫行为调控提供了理论依据,并为基于嗅觉的绿色防控策略开发提供支持。Abstract: The insect olfactory system plays a pivotal role in host recognition, mating, and oviposition, with odorant-binding proteins (OBPs) serving critical functions in olfactory signal transduction. An attempt was made to analyze the sequence characteristics and ligand-binding properties of two OBPs (TrufOBP1 and TrufOBP5) in Tirathaba rufivena, a major pest infesting inflorescences and fruit of arecanut (Areca catechu), and to validate their behavioral regulatory roles through wind tunnel experiments. Sequence analysis revealed that TrufOBP1 contains four conserved cysteines and is classified as a Minus-C OBP, whereas TrufOBP5 possesses ten conserved cysteines and is categorized as a Plus-C OBP. Fluorescence competitive binding assays demonstrated that TrufOBP1 exhibited broad ligand specificity and was bound to 11 compounds, including nine volatiles from areca nut (Areca catechu) inflorescences and two sex pheromones. In contrast, TrufOBP5 displayed high binding specificity, and interacted exclusively with octyl p-methoxycinnamate and vanillin, suggesting its potential role in host plant recognition. Wind tunnel experiments identified β-myrcene, octyl p-methoxycinnamate, and 3-hexenyl acetate as attractants for female adults from host volatiles. Among sex pheromone components, 2,2,6-trimethyl-6-vinyltetrahydro-2H-pyran-3-OL exhibited the highest attraction activity, while vanillin also demonstrated significant attractant effects. This study elucidates the functional roles of TrufOBP1 and TrufOBP5 in host recognition and sex pheromone perception in T. rufivena, providing a theoretical foundation for pest behavior regulation and supporting the development of olfaction-based green control strategies.
-
图 2 TrufOBP1与其他螟蛾科昆虫OBP的多序列比对
Fig. 2 Alignment of TrufOBP1 with OBP genes from other insect species of Pyraloidae
冷杉梢斑螟 Dioryctria abietella (Dabi);稻纵卷叶螟 Cnaphalocrocis medinalis (Cmed);二化螟 Chilo suppressalis (Csup)。
图 3 TrufOBP5与其他鳞翅目昆虫OBP的多序列比对
Fig. 3 Alignment of TrufOBP5 with OBP genes from other insect species of Lepidoptera.
大蜡螟 Galleria mellonella (Gmel);棉铃虫 Helicoverpa armigera (Harm);烟青虫 Helicoverpa assulta (Hass)。
图 4 TrufOBP1和TrufOBP5的系统发育树
Fig. 4 Phylogenetic tree of TrufOBP1 and TrufOBP5
二化螟 Chilo suppressalis (Csup);稻纵卷叶螟 Cnaphalocrocis medinalis (Cmed);棉铃虫 Helicoverpa armigera (Harm);冷杉梢斑螟 Dioryctria abietella (Dabi)。
图 5 TrufOBP1诱导表达以及纯化效果分析
注:M,蛋白Marker;1~2,未诱导和诱导菌液;3~4,菌液破碎后上清液和沉淀;5~6,诱导菌液破碎后上清液和流穿液;7~12,0、20、40、60、100、200 mmol·L−1咪唑洗脱蛋白。目的蛋白被方框标记。
Fig. 5 Expression and purification of TrufOBP1
Note: M, Protein molecular weight standard; 1−2, Protein sample without induction and protein sample with induction; 3−4, The supernatant of protein sample with induction after ultrasonic crushing and precipitation; 5−6, The supernatant of protein sample with induction after ultrasonic crushing, Protein sample without induction; 7−12, The protein was eluted with imidazole at several concentrations to give the protein samples (0, 20, 40, 60, 100 and 200 mmol·L−1). Label the target protein with a rectangle.
图 6 TrufOBP5诱导表达以及纯化效果分析
注:M,蛋白Marker;1~2,未诱导和诱导菌液;3~4,菌液破碎后上清液和沉淀;5~6,诱导菌液破碎后上清液和流穿液;7~11,0、20、40、100、200 mmol·L−1咪唑洗脱蛋白。目的蛋白被方框标记。
Fig. 6 Expression and purification of TrufOBP5
Note: M, Protein molecular weight standard; 1−2, Protein sample without induction and protein sample with induction; 3−4, The supernatant of protein sample with induction after ultrasonic crushing and precipitation; 5−6, The supernatant of protein sample with induction after ultrasonic crushing, and protein sample without induction; 7−11, The protein was eluted with imidazole at several concentrations to give the protein samples (0, 20, 40, 100 and 200 mmol·L−1). Label the target protein with a rectangle.
图 7 TrufOBP1、TrufOBP5与1-NPN探针的结合曲线和斯卡查德分析
注:非线性拟合为TrufOBP 与1-NPN探针的结合曲线,线性拟合为斯卡查德分析。
Fig. 7 The binding parameters of 1-NPN to TrufOBP1/TrufOBP5 and the Scatchard plots
Note: The curve fitting in the figure is the combination curve of TrufOBP and 1-NPN probe, and the linear fitting is Scatchard analysis.
图 9 雌成虫对7种化合物的风洞行为反应
注:不同小写字母表示在同一行为中,不同化合物处理间的行为反应率差异显著。
Fig. 9 Wind tunnel behavioral responses of female adults to 7 compounds
Note: Different lowercase letters indicate significant differences in behavioral response rates among different compound treatments within the same behavioral type.
表 1 本研究主要引物
Table 1 Primers in this study
引物 Primer 序列(5'-3') Sequence (5'-3') 内切酶 Endonuclease TrufOBP1-F CGCCATATGTTAACAGATGAGCAGAAAGAG Nde Ⅰ TrufOBP1-R AAATATGCGGCCGCCAGGAAGATGGCGTGCTT Not Ⅰ TrufOBP5-F CCGGAATTCGAAATTGAACAACTACAACC EcoR Ⅰ TrufOBP5-R AAATATGCGGCCGCGCATATCCATTCCGCAGC Not Ⅰ 注:酶切位点用下划线表示。
Note: The cleavage sites are indicated by underscores. 下载: 导出CSV
下载: 导出CSV
表 2 TrufOBP与18种配体的荧光竞争亲和力
Table 2 Fluorescence competitive affinity of TrufOBP with 18 ligands
序号
Serial Number化合物
CompoundCAS Ki/(μmol·L−1) TrufOBP1 TrufOBP5 1 十四烷 n−tetradecane 629-59-4 11.34±1.10 >50 2 十六烷 n−hexadecane 544-76-3 14.58 ±1.22 >50 3 十七烷 n−heptadecane 629-78-7 28.74 ±1.51 >50 4 α−蒎烯 α−pinene 80-56-8 23.46±1.43 29.36±0.33 5 β−蒎烯 β−pinene 127-91-3 29.90 ±1.53 27.07±1.77 6 β−月桂烯 β−myrcene 123-35-3 8.83±1.35 41.34±3.79 7 3−蒈烯 3−carene 13466 -78-9>50 25.95±0.94 8 反式−4−甲氧基肉桂酸异辛酯
2-ethylhexyl trans-4-methoxycinnamate83834 -59-718.66 ±1.32 5.16±1.05 9 苯甲醇 benzyl alcohol 100-51-6 13.79 ±1.14 27.05±1.60 10 芳樟醇 linalool 78-70-6 29.52 ±1.52 26.61±0.76 11 桉叶油醇 cineole 470-82-6 47.88 ±1.73 28.24±1.49 12 乙酸异戊酯 isopentyl acetate 123-92-2 8.34 ±0.97 28.52±1.23 13 2−甲基丁酸−3−甲基丁酯
3−methylbutyl−2−methyl butanoate27625 -35-010.14 ±1.06 21.75±0.94 14 乙酸己酯 hexyl acetate 142-92-7 17.40 ±1.30 35.26±2.11 15 乙酸叶醇酯 cis−3−hexenyl acetate 3681 -71-817.75±1.3 32.59±1.09 16 香兰素 vanillin 121-33-5 8.65 ±0.99 19.12±0.66 17 2,2,6−三甲基−6−乙烯基四氢−2H−呋喃−3−醇
2,2,6−trimethyl−6−vinyltetrahydro−2H−pyran−3−OL14049 -11-77.66 ±0.94 28.83±1.79 18 植酮 6,10,14−trimethyl−2−pentadecanone 502-69-2 >50 >50
下载: 导出CSV
-
[1] Zhong B, Lv C, Qin W. Effectiveness of the botanical insecticide azadirachtin against Tirathaba rufivena (Lepidoptera: Pyralidae) [J]. The Florida Entomologist, 2017, 100(2): 215−218. https://doi.org/10.1653/024.100.0215 doi: 10.1653/024.100.0215 [2] 钟宝珠, 吕朝军, 李朝绪, 等. 鱼藤酮对红脉穗螟的控制作用研究[J]. 中国植保导刊, 2023, 43(9): 5−9. https://doi.org/10.3969/j.issn.1672-6820.2023.09.001 doi: 10.3969/j.issn.1672-6820.2023.09.001 [3] 吕朝军, 钟宝珠, 苟志辉, 等. 鱼藤酮和茶皂素对槟榔红脉穗螟的联合毒力[J]. 生物安全学报, 2015, 24(3): 241−243. [4] 吕朝军, 钟宝珠, 钱军, 等. 烟碱对槟榔红脉穗螟生长发育和存活的影响[J]. 生物安全学报, 2013, 22(3): 201−205. [5] 甘炳春, 周亚奎, 杨新全, 等. 红脉穗螟寄生性天敌扁股小蜂的人工大量繁殖技术[J]. 中国森林病虫, 2011, 30(4): 29−30. https://doi.org/10.3969/j.issn.1671-0886.2011.04.010 doi: 10.3969/j.issn.1671-0886.2011.04.010 [6] 钟宝珠, 吕朝军, 李朝绪, 等. 褐带卷蛾茧蜂对不同龄期红脉穗螟幼虫的选择性和适应性[J]. 山西农业大学学报(自然科学版), 2023, 43(1): 97−103. [7] 陈运雷, 闫三强, 吕宝乾, 等. 不同寄主对麦蛾柔茧蜂寄生、发育和繁殖的影响[J]. 热带农业科学, 2022, 42(4): 77−80. https://doi.org/10.12008/j.issn.1009-2196.2022.04.013 doi: 10.12008/j.issn.1009-2196.2022.04.013 [8] 钟宝珠, 孙晓东, 吕朝军, 等. 分离自红棕象甲虫尸上的绿僵菌菌株鉴定及其对红脉穗螟的致病力[J]. 中国植保导刊, 2021, 41(7): 21−25+47. https://doi.org/10.3969/j.issn.1672-6820.2021.07.004 doi: 10.3969/j.issn.1672-6820.2021.07.004 [9] 李娇, 吕朝军. 红脉穗螟天敌垫跗螋的饲养方法[J]. 热带农业科学, 2017, 37(11): 65−68. [10] 李爱国, 许莉, 刘晓娜. 双性引诱剂对稻纵卷叶螟成虫的诱杀效果研究[J]. 现代农业科技, 2021(23): 84−85. https://doi.org/10.3969/j.issn.1007-5739.2021.23.034 doi: 10.3969/j.issn.1007-5739.2021.23.034 [11] 易松望, 李晓维, 陈利民, 等. 番茄潜叶蛾食物源引诱剂筛选及引诱效果评估[J]. 昆虫学报, 2024, 67(12): 1671−1683. [12] 张博军, 李依然, 韦肖红, 等. 桃园中梨小食心虫的引诱剂及诱捕器改进[J]. 中国生物防治学报, 2025, 41(1): 71−79. [13] Karlsson M F, Proffit M, Birgersson G. Host-plant location by the Guatemalan potato moth Tecia solanivora is assisted by floral volatiles [J]. Chemoecology, 2017, 27(5): 187−198. https://doi.org/10.1007/s00049-017-0244-2 doi: 10.1007/s00049-017-0244-2 [14] Bai P H, Wang H M, Liu B S, et al. Botanical volatiles selection in mediating electrophysiological responses and reproductive behaviors for the fall webworm moth Hyphantria cunea [J]. Frontiers in Physiology, 2020, 11: 486. https://doi.org/10.3389/fphys.2020.00486 doi: 10.3389/fphys.2020.00486 [15] Wei H S, Qin J H, Cao Y Z, et al. Two classic OBPs modulate the responses of female to three major ester host plant volatiles [J]. Insect Molecular Biology, 2021, 30(4): 390−399. https://doi.org/10.1111/imb.12703 doi: 10.1111/imb.12703 [16] Arimura G, Matsui K, Takabayashi J. Chemical and molecular ecology of herbivore-induced plant volatiles: proximate factors and their ultimate functions [J]. Plant and Cell Physiology, 2009, 50(5): 911−923. https://doi.org/10.1093/pcp/pcp030 doi: 10.1093/pcp/pcp030 [17] Leal W S. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes [J]. Annual Review of Entomology, 2013, 58(1): 373−391. https://doi.org/10.1146/annurev-ento-120811-153635 doi: 10.1146/annurev-ento-120811-153635 [18] Kaupp U B. Olfactory signalling in vertebrates and insects: differences and commonalities [J]. Nature Reviews Neuroscience, 2010, 11(3): 188−200. https://doi.org/10.1038/nrn2789 doi: 10.1038/nrn2789 [19] Zhou J J, Zhang G A, Huang W, et al. Revisiting the odorant-binding protein LUSH of Drosophila melanogaster: evidence for odour recognition and discrimination [J]. Febs Letters, 2004, 558(1−3): 23−26. https://doi.org/10.1016/S0014-5793(03)01521-7 doi: 10.1016/S0014-5793(03)01521-7 [20] 张林雅. 美洲斑潜蝇气味结合蛋白鉴定及功能研究[D]. 北京: 中国农业科学院, 2017. [21] 常向前. 东方粘虫性信息素结合蛋白(MsepPBPs)鉴定, 表达及功能分析[D]. 武汉: 华中农业大学, 2020. [22] 吴少英, 王桂荣, 吴孔明, 等. 烟青虫气味结合蛋白基因的克隆与序列分析[J]. 中国农业科学, 2005, 38(9): 8. https://doi.org/10.3321/j.issn:0578-1752.2005.09.016 doi: 10.3321/j.issn:0578-1752.2005.09.016 [23] Liu X L, Wu Z R, Liao W, et al. The binding affinity of two general odorant binding proteins in Spodoptera frugiperda to general volatiles and insecticides [J]. International Journal of Biological Macromolecules, 2023, 252: 126338. https://doi.org/10.1016/j.ijbiomac.2023.126338 doi: 10.1016/j.ijbiomac.2023.126338 [24] Li L L, Xu B Q, Li C Q, et al. Different binding affinities of three general odorant-binding proteins in Grapholita funebrana (Treitscheke) (Lepidoptera: Tortricidae) to sex pheromones, host plant volatiles, and insecticides [J]. Journal of Economic Entomology, 2022, 115(4): 1129−1145. https://doi.org/10.1093/jee/toac063 doi: 10.1093/jee/toac063 [25] Zhang X Q, Purba E R, Sun J, et al. Functional differentiation of two general-odorant binding proteins in Hyphantria cunea (Drury) (Lepidoptera: Erebidae) [J]. Pest Management Science, 2023, 79(9): 3312−3325. https://doi.org/10.1002/ps.7515 doi: 10.1002/ps.7515 [26] Ma Y, Li Y, Wei Z Q, et al. Identification and functional characterization of general odorant binding proteins in Orthaga achatina [J]. Insects, 2023, 14(3): 216. https://doi.org/10.3390/insects14030216 doi: 10.3390/insects14030216 [27] Zhang X Q, Yan Q, Li L L, et al. Different binding properties of two general-odorant binding proteins in Athetis lepigone with sex pheromones, host plant volatiles and insecticides [J]. Pesticide Biochemistry and Physiology, 2020, 164: 173−182. https://doi.org/10.1016/j.pestbp.2020.01.012 doi: 10.1016/j.pestbp.2020.01.012 [28] Yin N N, Yang A J, Wu C, et al. Genome-wide analysis of odorant-binding proteins in Papilio xuthus with focus on the perception of two PxutGOBPs to host odorants and insecticides [J]. Journal of Agricultural and Food Chemistry, 2022, 70(35): 10747−10761. https://doi.org/10.1021/acs.jafc.2c03396 doi: 10.1021/acs.jafc.2c03396 [29] Jing D P, Zhang T T, Prabu S, et al. Molecular characterization and volatile binding properties of pheromone binding proteins and general odorant binding proteins in Conogethes pinicolalis (Lepidoptera: Crambidae) [J]. International Journal of Biological Macromolecules, 2020, 146: 263−272. https://doi.org/10.1016/j.ijbiomac.2019.12.248 doi: 10.1016/j.ijbiomac.2019.12.248 [30] 王正. 红脉穗螟性信息素识别机制研究与应用初步[D]. 海口: 海南大学, 2022. [31] Zhou X, Wang Z, Cui G C, et al. Binding properties of odorant-binding protein 4 of Tirathaba rufivena to Areca catechu volatiles [J]. Plants-Basel, 2022, 11(2): 167. https://doi.org/10.3390/plants11020167 doi: 10.3390/plants11020167 [32] 白春秀, 周祥, 郭技星, 等. 中对长小蠹PparOBP1基因的序列分析与原核表达[J]. 基因组学与应用生物学, 2021, 40(Z4): 3396−3403. [33] 杨岭, 田晓丽, 桂连友, 等. 柑橘大实蝇气味结合蛋白BminOBP6与其气味配体的互作机制[J]. 中国农业科学, 2023, 56(7): 1311−1321. https://doi.org/10.3864/j.issn.0578-1752.2023.07.009 doi: 10.3864/j.issn.0578-1752.2023.07.009 [34] 许冲. 班氏跳小蜂气味结合蛋白嗅觉功能分析[D]. 武汉: 华中农业大学, 2020. [35] 严玉婷, 吴帆, 张亚丽, 等. 茶尺蠖触角高丰度气味结合蛋白EoblOBP9、EoblOBP11的配基结合功能和模式差异研究[J]. 茶叶科学, 2021, 41(5): 643−653. https://doi.org/10.3969/j.issn.1000-369X.2021.05.005 doi: 10.3969/j.issn.1000-369X.2021.05.005 [36] 唐浩宇, 刘靖涛, 谢佼昕, 等. 多异瓢虫气味结合蛋白HvarOBP2的基因克隆及配体结合特性分析[J]. 昆虫学报, 2022, 65(8): 977−985. [37] Zhan Y D, Liu Y J, Liu J H, et al. Pheromones emitted by both female and male moths regulate coordination between the sexes for Agriphila aeneociliella (Lepidoptera: Crambidae) [J]. Insect Science, 2023, 30(5): 1481−1492. https://doi.org/10.1111/1744-7917.13171 doi: 10.1111/1744-7917.13171 [38] Wang C, Zhang S, Guo M B, et al. Optimization of a pheromone lure by analyzing the peripheral coding of sex pheromones of Spodoptera frugiperda in China [J]. Pest Management Science, 2022, 78(7): 2995−3004. https://doi.org/10.1002/ps.6924 doi: 10.1002/ps.6924 [39] 吴健, 张贺贺, 杨燕川, 等. 一种桔小实蝇Minus-C气味结合蛋白的分子克隆和结合特征分析[J]. 江西农业大学学报, 2018, 40(2): 389−398. [40] 李亚伟, 张小菊, 王科珂, 等. 枣实蝇气味结合蛋白基因的克隆及表达谱分析[J]. 新疆农业科学, 2017, 54(2): 313−319. https://doi.org/10.6048/j.issn.1001-4330.2017.02.014 doi: 10.6048/j.issn.1001-4330.2017.02.014 [41] Zheng Z C, Li D Z, Zhou A, et al. Predicted structure of a Minus-C OBP from Batocera horsfieldi (Hope) suggests an intermediate structure in evolution of OBPs [J]. Scientific Reports, 2016, 6(1): 33981. https://doi.org/10.1038/srep33981 doi: 10.1038/srep33981 [42] Spinelli S, Lagarde A, Iovinella I, et al. Crystal structure of Apis mellifera OBP14, a C-minus odorant-binding protein, and its complexes with odorant molecules [J]. Insect Biochemistry and Molecular Biology, 2012, 42(1): 41−50. https://doi.org/10.1016/j.ibmb.2011.10.005 doi: 10.1016/j.ibmb.2011.10.005 [43] Li Z Q, Zhang S, Luo J Y, et al. Two Minus-C odorant binding proteins from Helicoverpa armigera display higher ligand binding affinity at acidic pH than neutral pH [J]. Journal of Insect Physiology, 2013, 59(3): 263−272. https://doi.org/10.1016/j.jinsphys.2012.12.004 doi: 10.1016/j.jinsphys.2012.12.004 [44] Vieira F G, Rozas J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the arthropoda: origin and evolutionary history of the chemosensory system [J]. Genome Biology and Evolution, 2011, 3: 476−490. https://doi.org/10.1093/gbe/evr033 doi: 10.1093/gbe/evr033 [45] 魏丹, 叶占峰, 高建清, 等. 二化螟Minus-C气味结合蛋白的分子克隆及功能鉴定[J]. 昆虫学报, 2013, 56(7): 754−764. [46] 陈秀琳, 苏丽, 陈丽慧, 等. 梨小食心虫Minus-C气味结合蛋白的分子克隆、表达谱及结合特性分析[J]. 昆虫学报, 2018, 61(7): 771−783. [47] Qu Y, Liu X, Zhao X, et al. Evidence of the involvement of a Plus-C odorant-binding protein HparOBP14 in host plant selection and oviposition of the scarab beetle Holotrichia parallela [J]. Insects, 2021, 12(5): 430. https://doi.org/10.3390/insects12050430 doi: 10.3390/insects12050430 [48] 都梓萌. 红脉穗螟气味结合蛋白TrufOBP1和TrufOBP7基因克隆及功能初探[D]. 海口: 海南大学, 2023. [49] Sasaerila Y H, Gries R, Gries G, et al. Sex pheromone components of male Tirathaba mundella (Lepidoptera: Pyralidae) [J]. Chemoecology, 2003, 13: 89−93. https://doi.org/10.1007/s00049-003-0233-5 doi: 10.1007/s00049-003-0233-5 [50] Lavogez O, Pinier C, Nibouche S, et al. Volatile compounds release by the hair pencils in male Prophantis smaragdina (Lepidoptera: Crambidae: Spilomelinae) [J]. Journal of Insect Science, 2017, 17(6): 171. [51] Leal W S, Barbosa R M R, Xu W, et al. Reverse and conventional chemical ecology approaches for the development of oviposition attractants for Culex Mosquitoes [J]. PLOS ONE, 2008, 3(8): e3045. https://doi.org/10.1371/journal.pone.0003045 doi: 10.1371/journal.pone.0003045 [52] Zhu J, Ban L, Song L M, et al. General odorant-binding proteins and sex pheromone guide larvae of Plutella xylostella to better food [J]. Insect Biochemistry and Molecular Biology, 2016, 72: 10−19. https://doi.org/10.1016/j.ibmb.2016.03.005 doi: 10.1016/j.ibmb.2016.03.005 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 920
- HTML全文浏览量: 803
- PDF下载量: 64
- 被引次数: 0
