

 下载:
下载:
-
类受体激酶(Receptor Like Kinase)是拟南芥中最大的蛋白质家族,共有600多种,其中,凝集素类受体激酶(Lectin receptor-like kinases, LecRLKs)是拟南芥中占比第二大RLKs亚家族,用于识别和传递环境信号,在植物胁迫和发育途径中发挥重要作用[1-3]。LecRLKs由胞内激酶结构域、跨膜结构域和胞外凝集素结构域组成[4]。根据细胞外凝集素结构域的不同,通常将LecRLKs分为L、G和C 3种不同类型[5]。到目前为止,在拟南芥中已报道有42个L型LecRLKs,1个C型LecRLKs,32个G型LecRLKs[6]。LecRLKs不同结构域的功能不同,胞内激酶结构域C末端尾部包含1个保守xGxxx(V/I/L)P起始端和GR双重端,对激酶结构域催化活性和该结构域与下游信号分子相互作用发挥重要功能[7-8]。跨膜结构域是信号传输工具,由22~28个氨基酸组成;跨膜结构域连接细胞外和细胞内结构域,将信号从细胞外传输到细胞内[5]。细胞外凝集素结构域是信号组合的中心,典型的L型LecRLKs包含1个豆科植物样细胞外凝集素结构域,具有β折叠结构和单糖结合特异性[9-10]。
有研究表明,LecRLKs在植物生长、发育及抗病信号传导过程中发挥重要作用。大多数L型LecRLKs定位于细胞膜,可以转运细胞外刺激信号至细胞内激发信号转导通路,例如感知PAMPs(Pathogen-associated molecular patterns)信号后,触发PTI信号通路[11]。拟南芥中LecRK-lX.1和LecRK-lX.2作为PAMPs触发免疫的正调节因子,促进拟南芥对疫霉菌(Phytophthora)的抗性[12-13]。L型凝集素受体LecRK-VI.2正调控PTI信号通路,促进植物对丁香假单胞菌和胡萝卜软腐果胶杆菌的抗性[14]。LecRK-I.9是拟南芥抗番茄枯萎病菌的基因,并影响茉莉酸酯信号传导[15]。据报道细胞外ATP是L型凝集素受体的主要配体之一[16]。Wang等[17]研究表明LecRK-V是来自二倍体小麦亲缘种(Haynaldia villosa)的L型凝集素受体激酶,在转基因小麦植株中赋予对白粉病的广谱抗性,这表明L型凝集素受体激酶在植物与病原微生物互作过程中发挥重要作用。但L型LecRLK在拟南芥中抗白粉病机理仍不清楚,也未见LecRLKs基因在白粉菌抗性中的功能研究。课题组(海南大学植物白粉病抗性分子育种研究室)前期研究结果发现橡胶树白粉菌(Oidium heveae,O. heveae)与拟南芥野生型Col-0具有非亲和相互关系,TIR-NB-LRR(Toll/Interleukin-1 Receptor-Nucleotide-Binding-Leucine-Rich Repeat)类抗病基因WRR4C(White Rust Resistance 4 C)正调控拟南芥Col-0对橡胶树白粉菌的抗病性[18]。然而,PTI抗病信号通路是否参与对O. heveae的抗病性,目前仍然是未知的。本研究利用O. heveae接种野生型Col-0,通过RNA-seq数据分析,发现凝集素类受体基因LecRK-I.7在O. heveae侵染12 h发生上调表达,推测LecRK-I.7可能发挥对白粉菌的抗病功能。本研究通过鉴定LecRK-I.7基因T-DNA插入突变体纯合系,接种O. heveae进行表型分析,结果表明与野生型Col-0对比,O. heveae在lecrk-1.7突变体形成浓密的菌丝网络和少数的分生孢子,并且在lecrk-1.7突变体激发的细胞死亡和活性氧爆发也大大降低,过表达LecRK-I.7基因后可以回复突变体的抗病表型,暗示着LecRK-I.7正调控拟南芥对O. heveae的抗病性。同时发现LecRK-I.7基因定位于植物细胞膜,并且受白粉菌的诱导上调表达。这为进一步研究拟南芥PTI信号通路抗O. heveae作用机制奠定了基础。
-
供试菌株为橡胶树白粉菌Oidium heveae HN1106。利用橡胶树热研‘73397’古铜期叶片活体(离体?)传代培养橡胶树白粉菌Oidium heveae HN1106。温室培养条件为温度28℃,相对湿度85%,光照16 h/黑暗8 h。
-
供试拟南芥为野生型拟南芥Col-0。拟南芥凝集素类受体激酶突变体lecrk-1.7为实验室保存。拟南芥温室培养条件为温度23℃,相对湿度85%,光照10 h/黑暗14 h。
-
根据T-DNA插入位点,在插入位点上下游各取500 bp分别设计正反向引物,利用3'引物法进行PCR扩增,根据结果鉴定是否为突变体纯合系。引物序列:
LecRK-I.7 F:5'-CCCTGGTGGAGAAGCCTTGC-3'
LecRK-I.7 R:5'-TTCCAAAATAGCATAACCAT-3'
-
利用活体接种的方法,剪取接菌后8~10天带有橡胶树白粉菌生理小种O. heveae HN1106成熟孢子的橡胶树叶片,用毛刷将白粉菌从叶片均匀刷到孔径为50 μm的尼龙膜接种箱,最终使白粉菌均匀落下至4周龄拟南芥叶片上,每片橡胶树叶片产生的成熟孢子接种大约24株拟南芥。
-
针对LecRK-I.7基因设计上下游引物,PCR后,胶回收,采用诺唯赞同源重组试剂盒连接至pER8载体。测序验证后,转化农杆菌感受态细胞GV3101,利用浸花法进行转基因操作。收种后,利用1/2 MS培养基进行筛选,培养条件为温度23℃,相对湿度85%,光照10 h/黑暗14 h。
引物序列如下:
LecRK-I.7 cDNA F:5'-GACGAGCTCGGTACCATGATTCGAGGATTGCTTTT-3'
LecRK-I.7 cDNA R:5'-TCTTTGTGATCTTCGAATCGCCCACTCCCGTAGAGGA-3'
-
采用biosahrp公司(中国北京,兰杰柯科技有限公司)植物样本总RNA提取试剂盒提取拟南芥Col-0和突变体lecrk-1.7幼苗叶片总RNA,经过biosahrp公司反转录试剂盒反转录为cDNA。使用SYBR Green qPCR Mix试剂盒进行荧光定量PCR,每个反应体系共20 μL,2XSYBR MIX(TAKARA)酶10 μL,Rox dye 0.4 μL,正反向引物各0.4 μL,cDNA模板5 μL,水3.8 μL。
引物序列如下:
ACTIN:5'-TGGTGGAAGCACAGAAGTTG-3';
5'-GATCCATGTTTGGCTCCTTC-3';
LecRK-I.7 RT:5'-GGGACATCGCATCGACTCTTA-3';
5'-CCGGGTTCCACAGGTCTCC-3'。
-
设计GFP基因上下游引物,克隆GFP基因,测序验证后,插入构建完成pER8-LecRK-I.7载体。引物序列如下:
LecRK-I.7 GFP F:5'- GACTCTAGCCTCGAGATGATTCGAGGATTGCTTTT-3';
LecRK-I.7 GFP R:5'- GATAGCCTCGAGATGAGTAAAGGAGAAGAACT-3'。
-
拟南芥转基因植株至5周,使用雌激素处理,36 h后,剪取叶片,加入蛋白提取液,使用研磨棒磨碎,离心(4℃,12 000 r·min−1,10 min)。吸取上清液,加入loading buffer,于100℃金属浴加热10 min。
利用SDS-PAGE凝胶电泳分离蛋白,湿法转印至PVDF膜,5%脱脂奶封闭2 h,一抗4℃孵育2 h,TBST洗膜3次后,二抗室温孵育0.5 h,再次洗膜,加入ELC化学发光液,曝光成像。
-
白粉菌接种后10 d,取8片接菌叶片置于50 mL离心管中,加入5 mL叶片脱色液,待叶片完全脱色,使用考马斯亮蓝染色液对叶片染色,使用显微镜观察[18]。显微镜下找到可以产生分生孢子梗的孢子,并对其产生的分生孢子梗数量进行统计,通过3次独立的接种实验完成统计,每次实验每个基因型植物统计超过20个孢子形成的分生孢子梗数量。
-
在玻璃管中加入4 mL台盼蓝染色液,取4片预接了白粉菌的叶片于玻璃管中,将玻璃管放入水中加热,水沸后计时10 min,关火,待叶片在水中自然放凉,6~8 h后将台盼蓝染色液换为水合氯醛,放置12 h后更换水合氯醛,重复更换3~4次,至叶片完全脱色,使用显微镜观察[18]。并计算每0.25 cm2叶片的细胞死亡面积占比。每个基因型植物观察8~12片接种叶片,分别通过3次独立的接种实验完成统计。
-
将含有目的基因农杆菌GV3101进行划线活化,置于5 mL LB液体培养基中培养。28℃,180 r·min−1振荡培养48 h后,吸取2 mL农杆菌菌液,5 000 ×g,离心10 min,去上清液。用缓冲液(10 mmol·L−1 MgCl2)漂洗3次,调至 OD 值0.5~0.6,进行注射。
-
剪取单个穴盘中生长4周龄拟南芥植株的地上部分,清除附着的泥土后立即称量,记录湿质量。将样本置于65℃恒温烘箱,每24h称量至质量恒定,记录质量,计算每64 cm2生物量。每个基因型植物测量6个穴盘。
-
称取 0.1 g拟南芥叶片鲜样,剪成0.5 cm左右的碎片,放入干燥的50 mL试管,向试管中加入8 mL 95%乙醇,确保叶片碎片完全浸没在溶液中,室温避光静置浸泡36 h至叶片完全变白。分光光度计分别测定提取液在665 nm和649 nm波长下的吸光度值,最终计算叶绿素含量。
-
采用Graphpad Prism 10(美国波士顿,GraphPad Software公司)软件对qPCR数据结果进行统计分析及作图。利用Smart软件(
https://smart.embl.de/ )对LecRK-I.7蛋白结构进行预测。 -
提取野生型拟南芥Col-0和凝集素类受体激酶LecRK-I.7基因T-DNA插入突变体lecrk-1.7(SAIL-717-B11)基因组DNA,LecRK-I.7基因由1个外显子编码,T-DNA插入位置为1 792 bp,插入方向为正向插入(图1-A),并通过三引物法验证纯合突变体植株。结果表明,利用T-DNA插入位点上下游引物进行PCR扩增,只有Col-0扩增出目的条带(图1-F),突变体不能扩增出特异性条带(图1-F);因T-DNA插入方向为正向插入,利用T-DNA引物和插入位点下游引物进行扩增,lecrk-1.7突变体1~10均扩增出目的条带(图1-G),而野生型Col-0未扩增出目的条带(图1-G)。这说明10株lecrk-1.7均为突变植株纯合系。纯合系突变体植株种植过程中,对生长4周龄的植株大小进行观察(图1-B),并且统计了地上生物量(图1-D)和叶绿素含量(图1-E)及抽苔后的株高(图1-C)。结果表明,突变体与野生型Col-0植株相比,差异性不显著;SAIL_717_B11突变体未表现出明显生长性状差异。
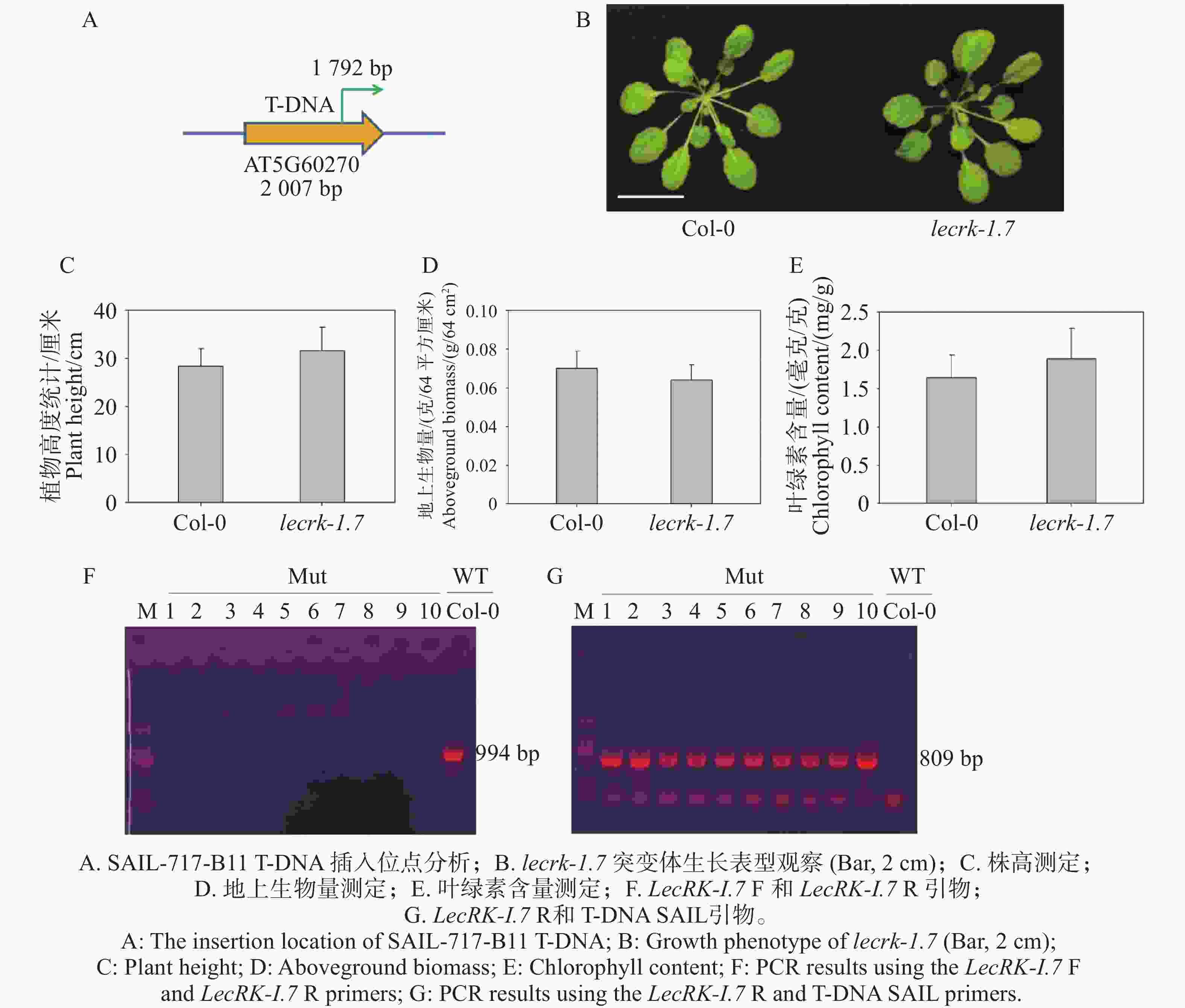
图 1 lecrk-1.7突变体纯合系鉴定结果
Figure 1. The characterization of homozygous T-DNA insertions of lecrk-1.7 mutants
-
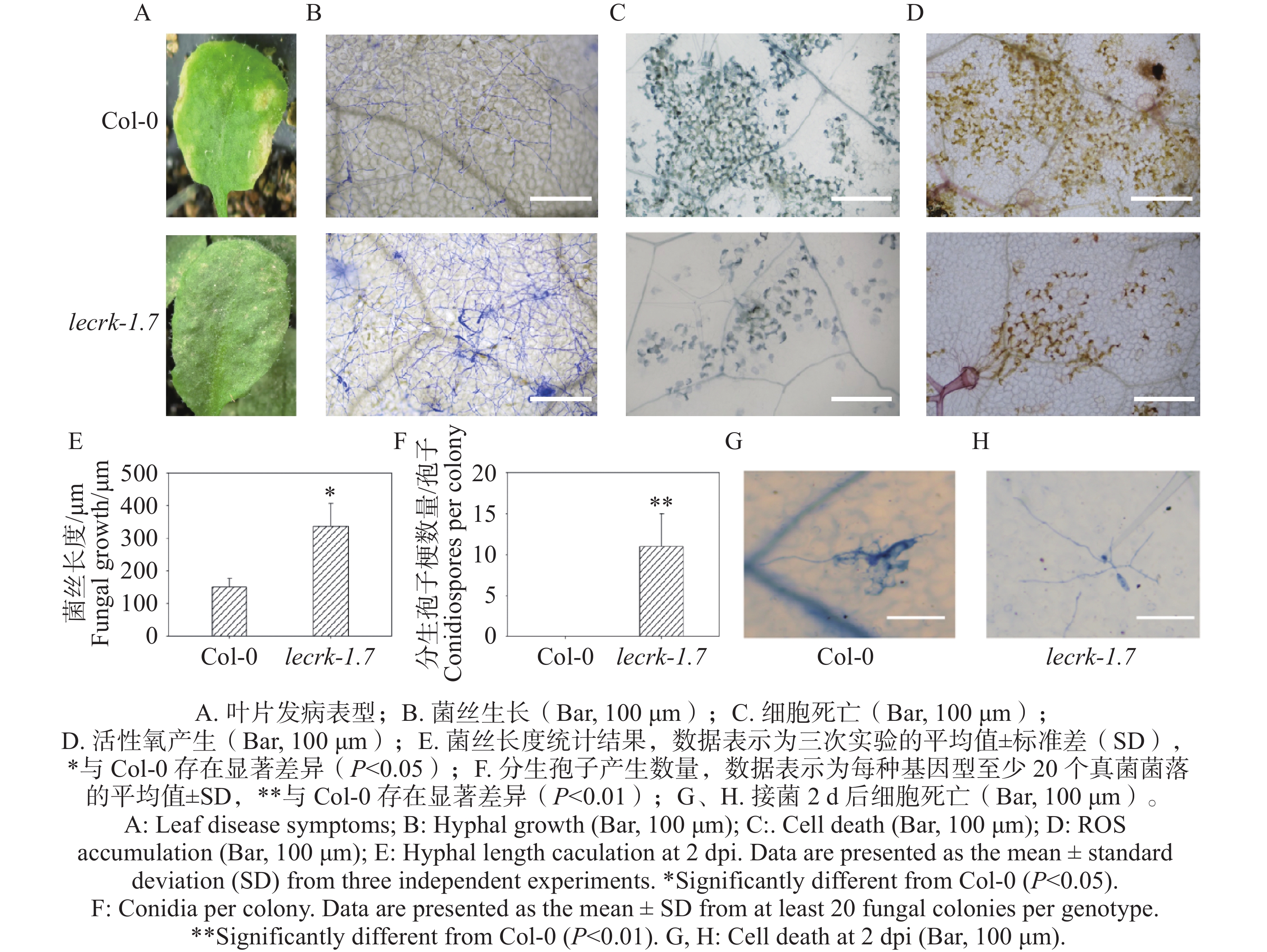
选择生长期为4周龄拟南芥Col-0和lecrk-1.7植株同时接种O. heveae,接种10 d,lecrk-1.7叶片出现点状白粉病病斑(图2-A),而Col-0叶片表现黄化的症状(图2-A),可以初步确定LecRK-I.7基因参与了拟南芥抗对白粉菌的抗病性。进一步通过对接菌10 d的Col-0和lecrk-1.7叶片进行考马斯亮蓝染色。显微镜观察显示,与野生型Col-0相比,突变体lecrk-1.7叶片上形成了更为浓密的菌丝网络,且有少量分生孢子的产生,而Col-0叶片上仅形成稀疏的菌丝网络,未观察到分生孢子的产生(图2-B,F)。同时, O. heveae在lecrk-1.7激发的细胞死亡和活性氧比在野生型Col-0都显著降低(图2-C,D),这进一步说明了凝集素受体激酶基因LecRK-I.7对O. heveae发挥抗病功能。
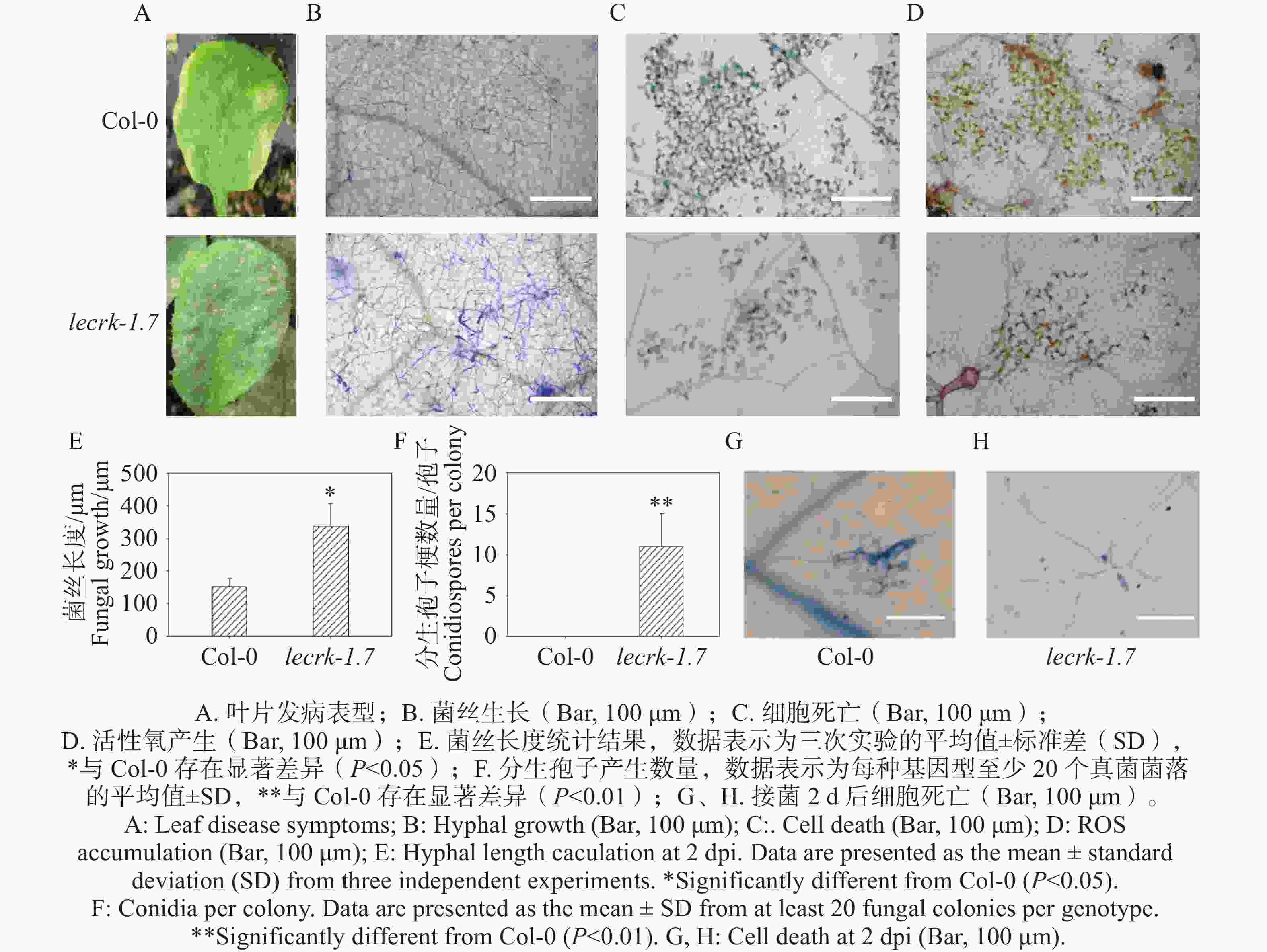
图 2 LecRK-I.7基因正调控拟南芥对O. heveae的抗病性
Figure 2. The LecRK-I.7 gene positively regulates Arabidopsis resistance to O. heveae
在Col-0和lecrk-1.7接种O. heveae 2 d时,剪取叶片进行考马斯亮蓝染色。显微镜观察发现,突变体lecrk-1.7叶片上形成树枝状菌丝网络,菌丝长度约340 μm,而野生型仅形成2根菌丝,长度约150 μm,差异显著(图2-E)。接种O. heveae 2 d的Col-0和lecrk-1.7经台盼蓝染色后,经显微观察发现,Col-0叶片在O. heveae菌丝生长部位引发明显细胞死亡,而在突变体O. heveae菌丝生长部位未观察到细胞死亡(图2-G,H),这暗示着LecRK-I.7同时参与对O. heveae的早期抗病性。
-
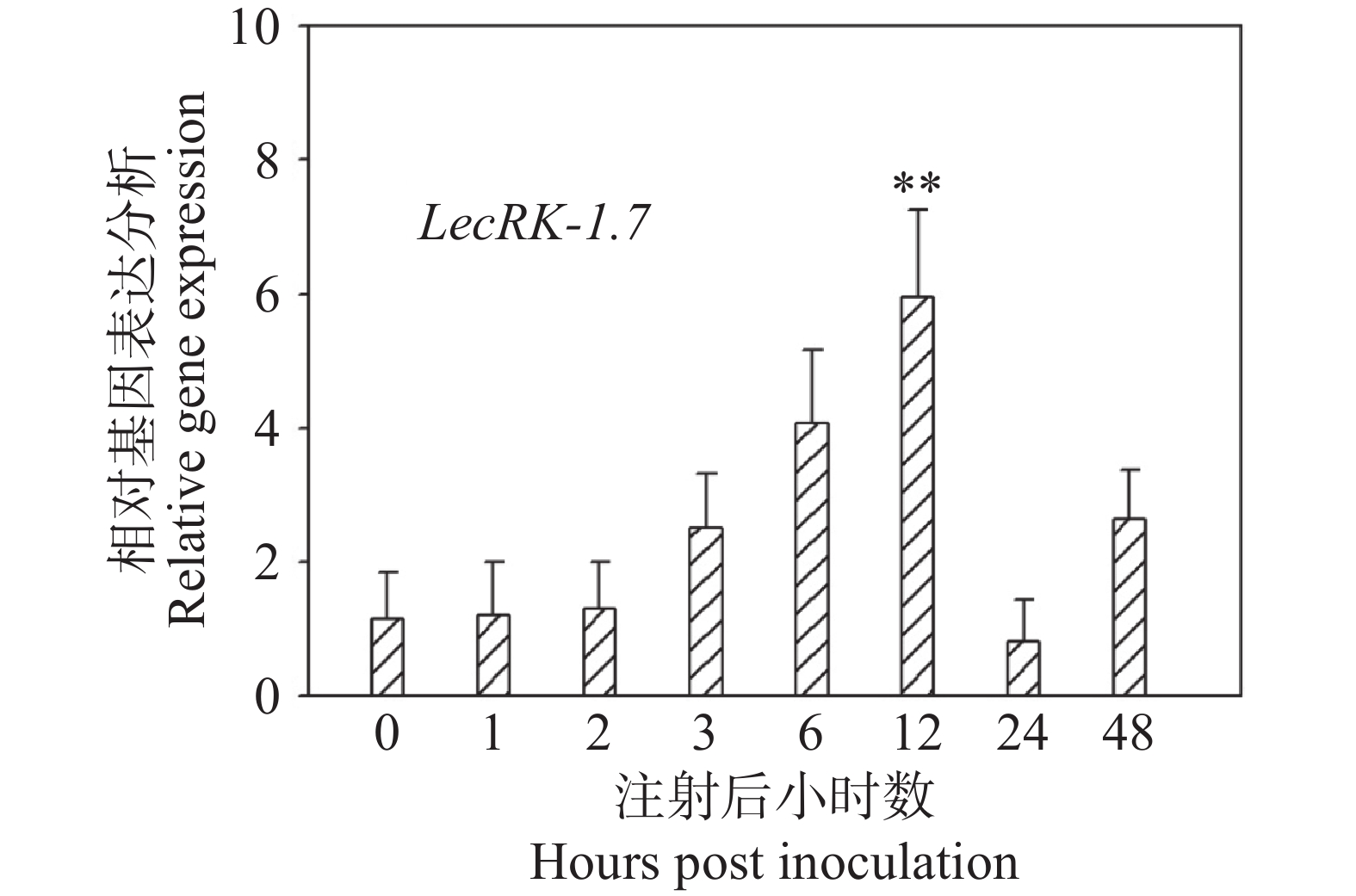
实验室(海南大学植物白粉病抗性分子育种研究室)通过前期RNA-seq数据发现LecRK-I.7基因在O. heveae侵染12 h时发生了上调表达,但为了进一步探索LecRK-I.7在白粉菌侵染过程中的表达模式,本研究团队对接菌后0 、1 、2 、3 、6 、12 、24 和48 h的Col-0取样,提取总RNA并进行反转录后,利用荧光定量PCR检测LecRK-I.7的表达水平,结果发现,与0 h相比,LecRK-I.7基因在白粉菌接种后3、6、12 h均发生了不同程度的上调表达,接种12 h后LecRK-I.7的表达量达到最高,是接种0 h的6倍左右。接种24 h后,基本回复到0 h正常水平(图3)。O. heveae在拟南芥侵染后12 h是第一根芽管形成附着孢刺入植物细胞形成初生吸器的关键阶段,因此推测LecRK-I.7基因可能在白粉菌进入植物细胞阶段表达发挥抗性功能。
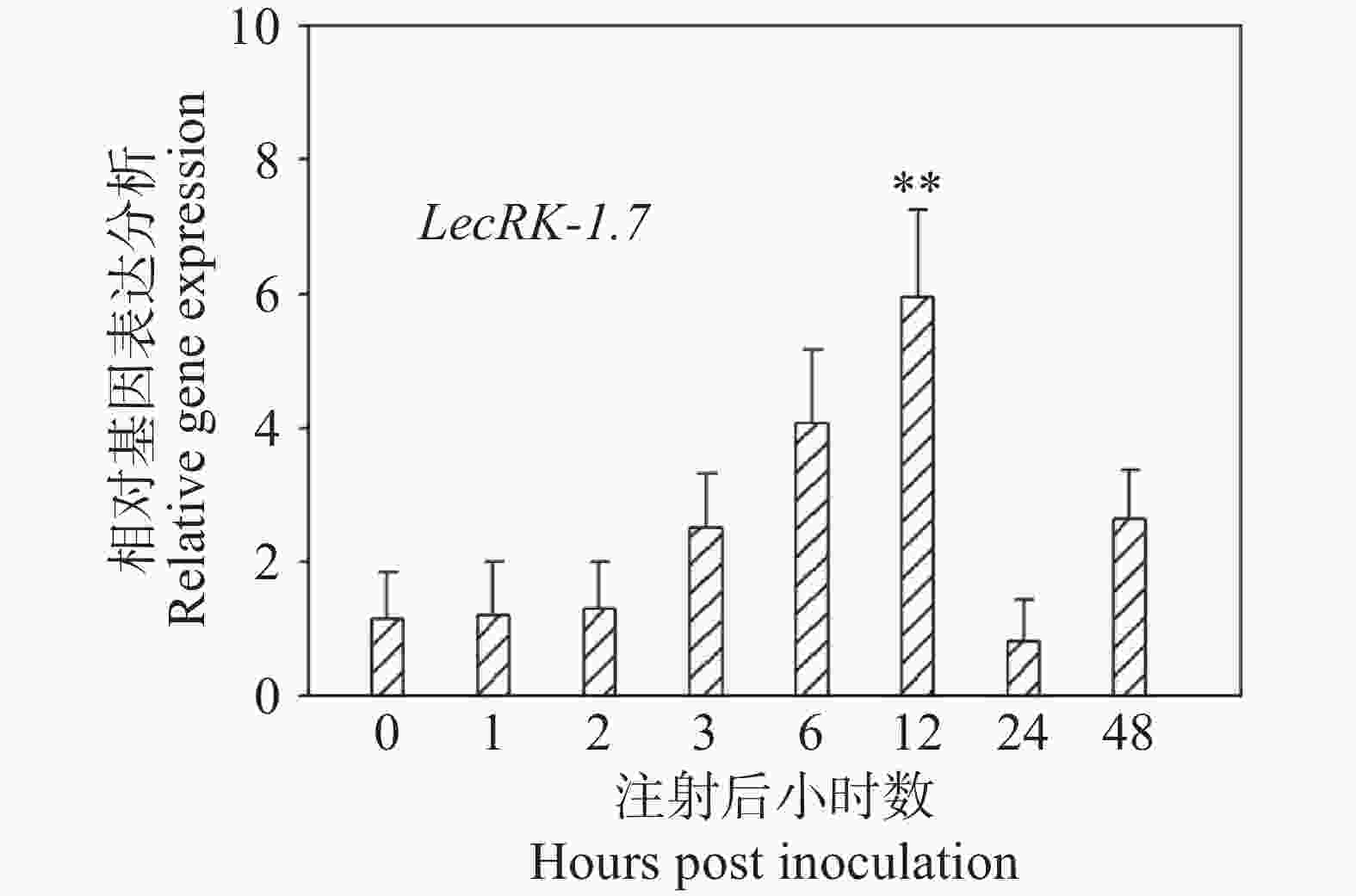
图 3 LecRK-I.7基因受O. heveae诱导上调表达
Figure 3. The expression of LecRK-I.7 gene is induced and upregulated by O. heveae
-
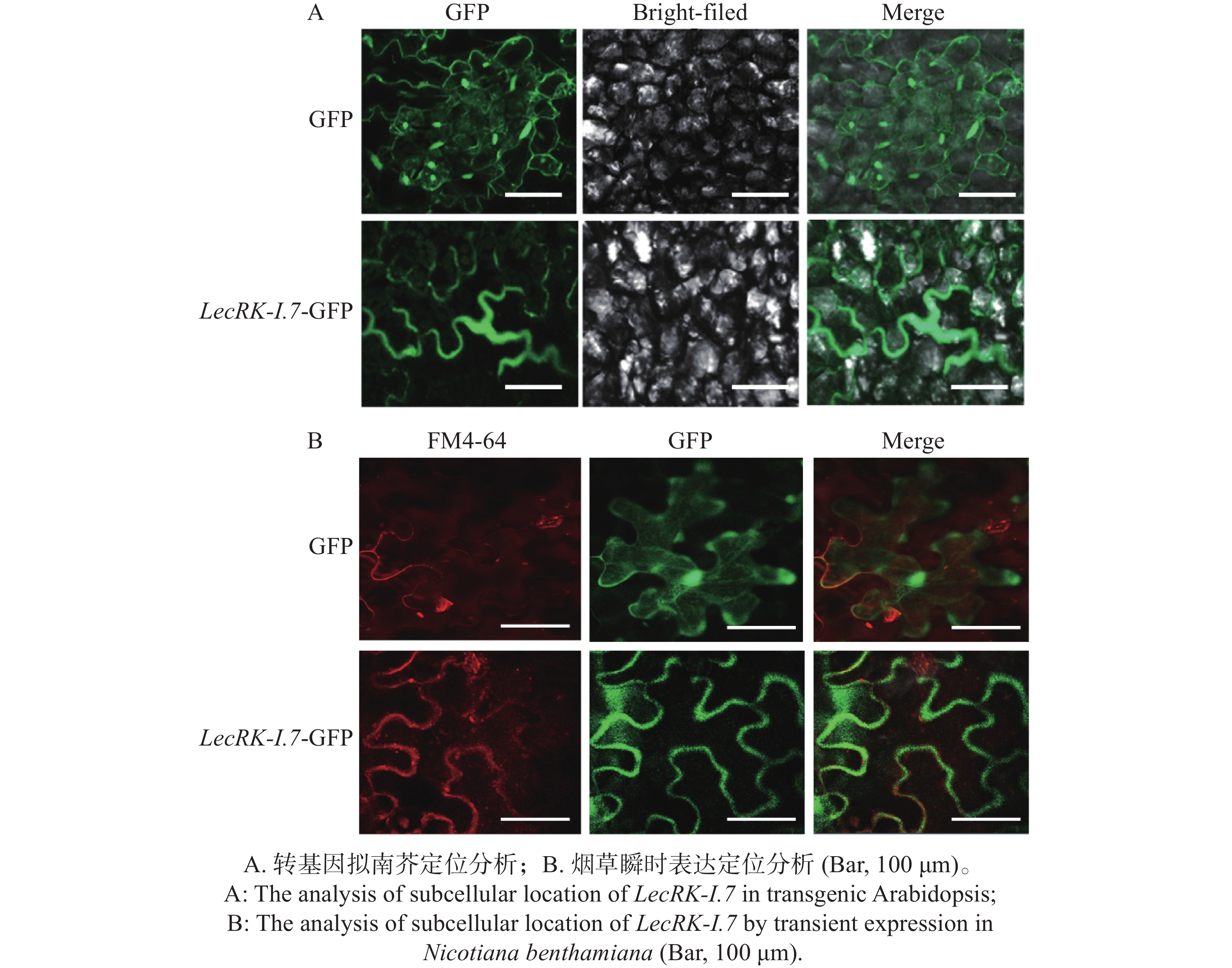
凝集素类受体激酶定位于植物细胞膜发挥功能[9]。通过蛋白结构预测发现LecRK-I.7蛋白具有胞内激酶结构域、胞外凝集素结构域和2个跨膜结构域,跨膜结构域分别位于233-255 AA和286-308 AA,因此推测LecRK-I.7基因定位于植物细胞膜发挥功能。为了确定LecRK-I.7基因是否定位于植物细胞膜,本研究构建了35S组成型启动子驱动的LecRK-I.7-GFP融合表达载体,利用农杆菌转化,将LecRK-I.7-GFP基因转入拟南芥Col-0,并筛选出阳性转基因植株,通过激光共聚焦显微镜对其叶片进行观察。结果发现,在488 nm激发光下,LecRK-I.7-GFP融合蛋白的绿色荧光信号主要分布于细胞膜(图4-A),对照GFP在细胞膜、细胞核和细胞质均有表达(图4-A)。利用本生烟瞬时表达系统,分别注射含有GFP空载体和LecRK-I.7-GFP载体的农杆菌,并用细胞膜染料FM4-64处理注射叶片,结果与拟南芥转基因相似,细胞膜、细胞核和细胞质均可以观察到对照GFP的存在,而LecRK-I.7-GFP融合蛋白的绿色荧光信号主要分布于细胞膜并与FM4-64信号相融合(图4-B),暗示着LecRK-I.7基因主要定位于细胞膜发挥功能。
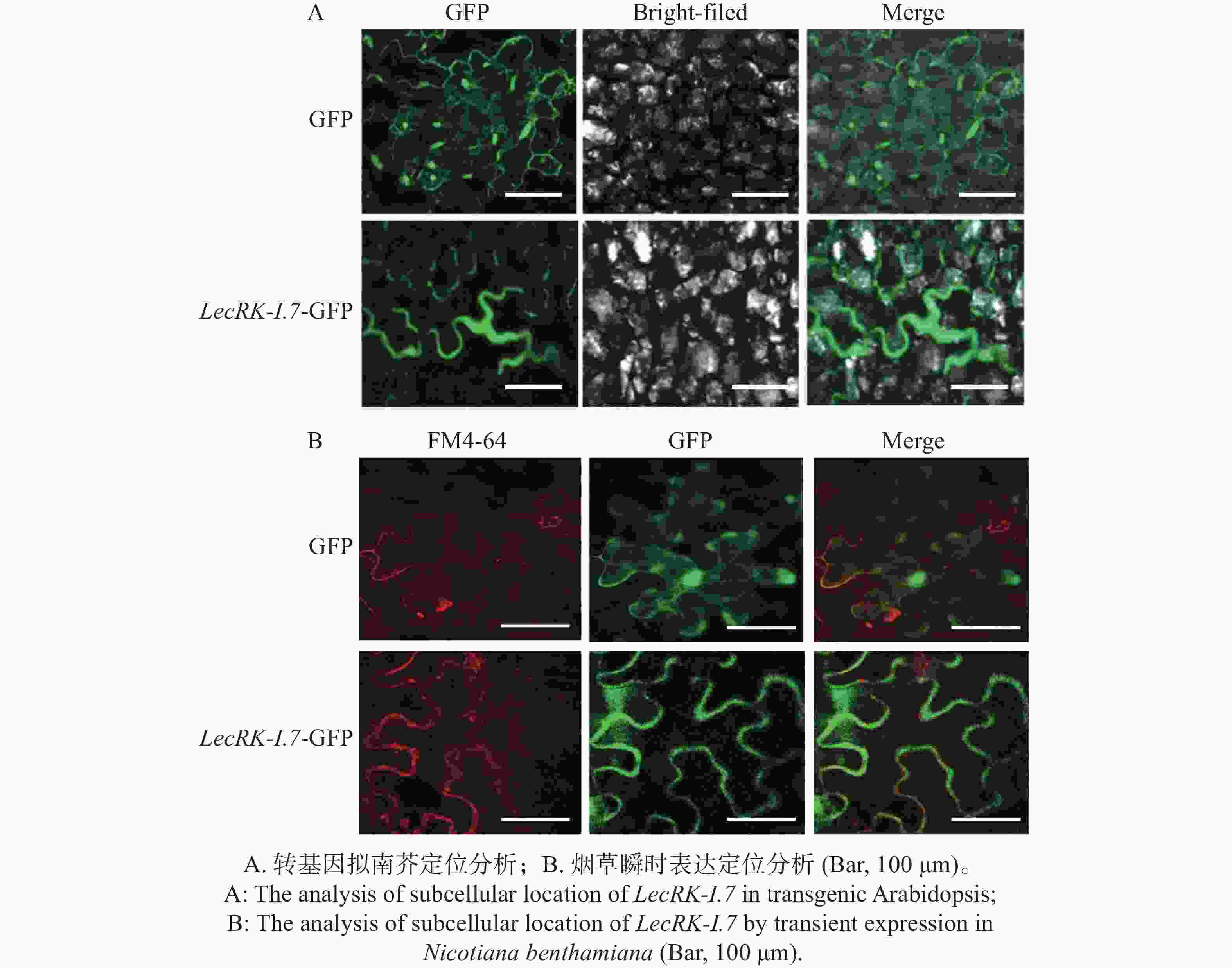
图 4 LecRK-I.7基因主要定位于植物细胞膜
Figure 4. The LecRK-I.7 gene is mainly localized to the plant cell membrane
-
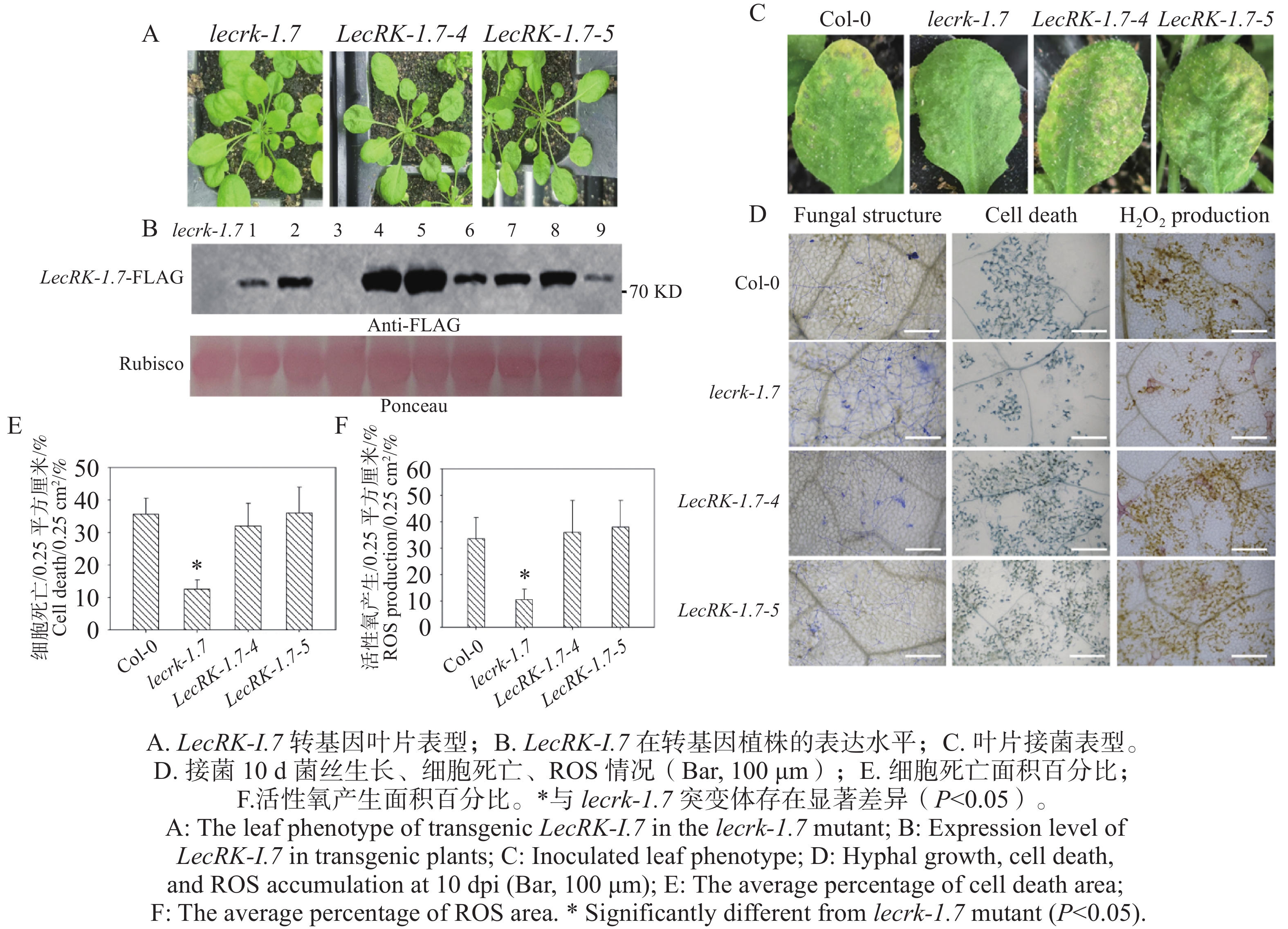
本研究构建了35S组成型启动子驱动的LecRK-I.7-FLAG融合表达载体,导入农杆菌,将LecRK-I.7-FLAG载体转化拟南芥lecrk-I.7突变体,利用免疫印迹筛选出高表达阳性转基因植株Line 4和Line 5(图5-B),通过表型观察,2个高表达阳性转基因植株Line 4和Line 5与突变体lecrk-1.7相比,未表现出明显抗性表型(图5-A)。分别对Col-0、lecrk-1.7突变体和2个LecRK-I.7转基因植株接种O. heveae,接种后10 d,野生型Col-0和2个LecRK-I.7转基因系叶片均表现典型黄化症状(图5-C),而lecrk-1.7叶片出现白粉病点状病斑(图5-C),未表现出发黄表型。进一步通过考马斯亮蓝染色,显微观察发现O. heveae在Col-0和2个LecRK-I.7转基因系形成相似的菌丝网络,未检测到分生孢子的产生,而在突变体lecrk-1.7形成浓密的菌丝网络和少量的分生孢子(图5-D)。利用台盼蓝和DAB染色,经显微观察发现在接菌后转基因叶片与野生型Col-0叶片均表现出明显细胞死亡与活性氧产生(图5-D)。进一步统计活性氧产生(图5-F)和细胞死亡(图5-E)面积百分比,野生型Col-0和2个转基因系与lecrk-1.7突变体存在显著差异,说明LecRK-I.7回复了突变体lecrk-1.7对O. heveae的抗病性。
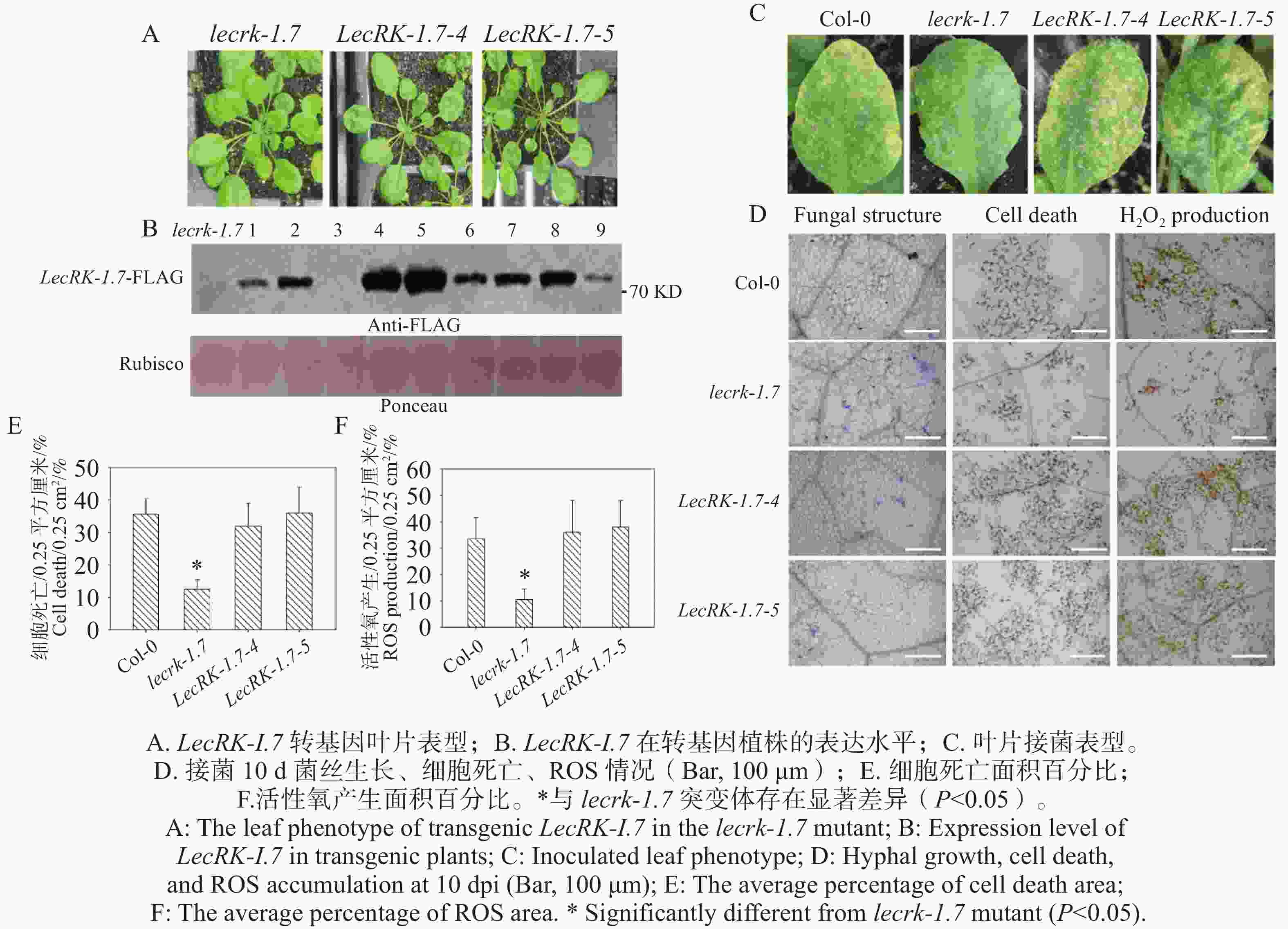
图 5 过表达LecRK-I.7回复突变体lecrk-1.7对O. heveae的抗病性
Figure 5. Overexpression of LecRK-I.7 rescues the resistance of the lecrk-1.7 mutant to O. heveae
-
橡胶树白粉病是橡胶树上危害严重的叶部病害,橡胶树白粉菌是一种专性寄生真菌,然而其与寄主互作的分子机制研究相对滞后[19]。G型凝集素受体激酶VqLecRK1基因具有增强葡萄抗白粉病的功能[20]。L型凝集素受体激酶LecRK-V在转基因小麦植株中赋予对白粉病的广谱抗性[17]。然而,凝集素受体激酶是否参与对O. heveae的抗病性,目前仍然是未知的。本研究前期(数据尚未发表)通过RNA-seq数据分析发现凝集素类受体激酶LecRK-I.7受橡胶树白粉菌诱导上调表达,推测LecRK-I.7对O. heveae可能发挥抗病功能。通过鉴定LecRK-I.7基因T-DNA插入突变体SAIL-717-B11纯合系,与野生型Col-0相比,突变体未表现出地上生物量、叶绿素和株高等生长性状显著差异。进一步在lecrk-1.7突变体接种O. heveae,发现O. heveae可以形成比Col-0更为浓密的菌丝网络和少量的分生孢子,以及激发的细胞死亡和活性氧也大大降低,这暗示着LecRK-I.7参与了拟南芥对O. heveae的抗病性。同时,利用回补实验,本研究构建了LecRK-I.7在lecrk-1.7突变体背景下的过表达转基因植物株。接种O. heveae,转基因植株可以回复突变体对白粉菌的抗病表型,从遗传学上进一步证实了LecRK-I.7抗病功能。然而,未接菌时,过表达转基因植物与lecrk-1.7突变体无明显生长表型差异,推测LecRK-I.7识别白粉菌并与相应PAMPs结合后才能发挥抗病功能。
LecRLKs定位于植物细胞膜,利用胞外凝集素结构域识别病原菌相关分子模式(PAMPs, Pathogen-Associated Molecular Patterns)或寄主细胞表面的损伤相关分子模式(DAMPs, Damage-Associated Molecular Patterns),进而启动下游免疫信号传导[9, 21] 。因此,膜定位对于LecRLKs功能的正常发挥至关重要。蛋白结构预测分析,LecRK-I.7具有胞内激酶结构域、胞外凝集素结构域和2个跨膜结构域,跨膜结构域分别位于233-255 AA和286-308 AA,这与本研究得出的LecRK-I.7膜定位结论相一致,确定LecRK-I.7作为膜受体感知O. heveae的入侵。PTI信号通路通常在病原微生物与植物互作的早期被激活,荧光定量PCR发现,在O. heveae侵染的过程中,LecRK-I.7在侵染后的3~12 h均表现上调表达,12 h表达量达到最高,24 h后回复到正常水平。白粉菌侵染后的6~12 h是白粉菌入侵进入植物细胞的关键阶段,暗示着LecRK-I.7可能在该时间段阻碍了白粉菌的进入。进一步的台盼蓝染色分析发现,接种后2 d,O. heveae在Col-0进入植物细胞的位置形成明显可见的细胞死亡,而在lecrk-1.7突变体未发现细胞死亡,并且形成了显著长于Col-0的菌丝长度,这表明LecRK-I.7识别O. heveae后激发了细胞死亡从而抑制了菌丝的生长。而后期O. heveae在lecrk-1.7突变体诱导的细胞死亡可能是LecRK-I.7突变后导致大量白粉菌进入植物细胞从而激发的ETI抗性。不同类型凝集素类受体激酶识别的PAMPs不同,拟南芥G型凝集素受体激酶LORE可以感知细菌中链3-羟基脂肪酸(mc-3-OH-FA)[22-24],拟南芥L型凝集素受体激酶LecRK-IX.2特异性识别细菌鞭毛蛋白保守肽段flg22[12],关于LecRK-I.7识别的O. heveae PAMPs,在未来的研究中可以通过LecRK-I.7转基因植物接种白粉菌后,利用免疫沉淀质谱的方法进行分离。
植物与病原微生物长期互作过程中,植物进化出2层次的免疫系统PTI和ETI识别病原微生物的入侵,激发抗病反应。PTI和ETI信号通路既独立存在也相互交叉,协同抵御病原微生物的侵染。有研究报道表明O. heveae在拟南芥激发依赖于EDS1的抗病反应,并且TNL类蛋白家族WRR4可以识别O. heveae,激发ETI信号通路[25- 26]。本研究发现拟南芥凝集素受体激酶LecRK-I.7激发的PTI信号通路也参与对O. heveae的抗病性,这说明拟南芥PTI和ETI对O. heveae共同发挥抗病功能。PTI和ETI信号通路存在相互交叉,WRR4B通过正调控EDS1(Enhanced Disease Susceptibility 1)正反馈调控环相关基因表达,对白粉菌发挥广谱抗病功能;鉴于EDS1同时也参与PTI信号通路,并且eds1突变体对O. heveae表现完全感病性[27],故推测LecRK-I.7激发的抗病通路可能也依赖于EDS1,这一推测仍需要进一步实验证明。另外,拟南芥其他凝集素类受体基因是否参与对O. heveae的抗病性,及LecRK-I.7与抗性蛋白WRR4B是否在O. heveae抗病信号通路存在遗传上下游关系,也需要进一步验证。
Arabidopsis lectin-like receptor kinase LecRK-I.7 positively regulates the disease resistance against Oidium heveae
-
摘要: 凝集素受体激酶LecRLKs (Lectin receptor-like kinases )是一类关键的免疫相关受体,在植物识别病原信号和启动防御反应中扮演重要角色。橡胶树白粉菌 (Oidium heveae)在拟南芥激发抗病性,然而,凝集素受体激酶是否参与拟南芥对白粉菌的抗病性,目前仍然是未知的。本研究利用O. heveae接种野生型Col-0,通过RNA-seq数据分析,发现凝集素类受体基因LecRK-I.7 (Lectin receptor gene)在O. heveae侵染12 h发生上调表达,推测LecRK-I.7可能发挥对白粉菌的抗病功能。通过鉴定LecRK-I.7基因T-DNA插入突变体纯合系,接种O. heveae进行表型分析。结果表明,与野生型Col-0对比,O. heveae在lecrk-1.7突变体形成浓密的菌丝网络和少数的分生孢子,并且在lecrk-1.7突变体激发的细胞死亡和活性氧爆发也大大降低。进一步过表达LecRK-I.7基因后可以回复突变体的抗病表型,从遗传上证实LecRK-I.7正调控拟南芥对O. heveae的抗病性。另外发现LecRK-I.7基因主要定位于植物细胞膜,并且受白粉菌的诱导上调表达,这为进一步研究拟南芥对O. heveae的抗病机制奠定了基础。Abstract: Lectin receptor-like kinases (LecRLKs) are a class of key immune-related receptors that play an important role in plant recognition of pathogen signals and the initiation of defense responses. The powdery mildew of rubber tree, Oidium heveae, triggers disease resistance in Arabidopsis. However, it remains unknown whether the LecRLKs are involved in Arabidopsis resistance against O. heveae. In this study, RNA-seq analysis revealed that the lectin receptor gene LecRK-I.7 was up-regulated by 12 hours after O. heveae infection, suggesting that LecRK-I.7 may play a role in resistance to O. heveae. The T-DNA homozygous insertion mutants of LecRK-I.7 were characterized and inoculated with O. heveae. The results showed that compared with wild-type Col-0, O. heveae formed dense hyphal networks and a few conidia in the lecrk-1.7 mutants, and that the cell death and reactive oxygen burst induced by O. heveae in lecrk-1.7 mutants were also greatly reduced than those in WT Col-0. Furthermore, the resistance phenotype of mutant lecrk-1.7 was restored by overexpression of LecRK-I.7 in the mutant, indicating that LecRK-I.7 positively regulates Arabidopsis resistance to O. heveae. It was also confirmed that LecRK-I.7 was mainly localized to the plant cell membrane to fulfill its function, and that the expression of LecRK-I.7 gene could be induced by O. heveae, which provides a basis for further study on the disease resistance mechanisms of Arabidopsis against O. heveae.
-
Key words:
- lectin-like receptor kinase /
- LecRK-I.7 /
- Arabidopsis thaliana /
- Oidium heveae /
- disease resistance
-
图 2 LecRK-I.7基因正调控拟南芥对O. heveae的抗病性
Fig. 2 The LecRK-I.7 gene positively regulates Arabidopsis resistance to O. heveae
图 3 LecRK-I.7基因受O. heveae诱导上调表达
**与接菌后0 h存在显著差异(P<0.01)
Fig. 3 The expression of LecRK-I.7 gene is induced and upregulated by O. heveae
** Significantly different from 0 hours post inoculation (P < 0.01).
图 4 LecRK-I.7基因主要定位于植物细胞膜
Fig. 4 The LecRK-I.7 gene is mainly localized to the plant cell membrane
-
[1] Jose J, Ghantasala S, Roy Choudhury S. Arabidopsis transmembrane receptor-like kinases (RLKs): a bridge between extracellular signal and intracellular regulatory machinery [J]. International Journal of Molecular Sciences, 2020, 21(11): 4000. https://doi.org/10.3390/ijms21114000 doi: 10.3390/ijms21114000 [2] Shiu S H, Bleecker A B. Plant receptor-like kinase gene family: diversity, function, and signaling [J]. Science’s STKE, 2001, 2001(113): re22. https://doi.org/10.1126/stke.2001.113.re22 doi: 10.1126/stke.2001.113.re22 [3] He Y X, Zhou J G, Shan L B, et al. Plant cell surface receptor-mediated signaling – a common theme amid diversity [J]. Journal of Cell Science, 2018, 131(2): jcs209353. https://doi.org/10.1242/jcs.209353 doi: 10.1242/jcs.209353 [4] 彭小群, 邹雅琦, 骆素微, 等. 植物凝集素类受体激酶参与抗病的研究进展[J]. 植物科学学报, 2022, 40(1): 105−114. https://doi.org/10.11913/PSJ.2095-0837.2022.10105 doi: 10.11913/PSJ.2095-0837.2022.10105 [5] Ye Y Y, Ding Y F, Jiang Q, et al. The role of receptor-like protein kinases (RLKs) in abiotic stress response in plants [J]. Plant Cell Reports, 2017, 36(2): 235−242. https://doi.org/10.1007/s00299-016-2084-x doi: 10.1007/s00299-016-2084-x [6] Bellande K, Bono J J, Savelli B, et al. Plant lectins and lectin receptor-like kinases: how do they sense the outside? [J]. International Journal of Molecular Sciences, 2017, 18(6): 1164. https://doi.org/10.3390/ijms18061164 doi: 10.3390/ijms18061164 [7] Vaid N, Pandey P K, Tuteja N. Genome-wide analysis of lectin receptor-like kinase family from Arabidopsis and rice [J]. Plant Molecular Biology, 2012, 80(4): 365−388. https://doi.org/10.1007/s11103-012-9952-8 doi: 10.1007/s11103-012-9952-8 [8] Bhat A, Haney C H. The role of plant receptor-like kinases in sensing extrinsic and host-derived signals and shaping the microbiome [J]. Cell Host & Microbe, 2025, 33(8): 1233−1240. https://doi.org/10.1016/j.chom.2025.07.012 doi: 10.1016/j.chom.2025.07.012 [9] Sun Y L, Qiao Z Z, Muchero W, et al. Lectin receptor-like kinases: the sensor and mediator at the plant cell surface [J]. Frontiers in Plant Science, 2020, 11: 596301. https://doi.org/10.3389/fpls.2020.596301 doi: 10.3389/fpls.2020.596301 [10] Wang Y, Bouwmeester K. L-type lectin receptor kinases: new forces in plant immunity [J]. PLoS Pathogens, 2017, 13(8): e1006433. https://doi.org/10.1371/journal.ppat.1006433 doi: 10.1371/journal.ppat.1006433 [11] Liu L, Liu J, Xu N. Ligand recognition and signal transduction by lectin receptor-like kinases in plant immunity [J]. Frontiers in Plant Science, 2023, 14: 1201805. https://doi.org/10.3389/fpls.2023.1201805 doi: 10.3389/fpls.2023.1201805 [12] Luo X M, Xu N, Huang J K, et al. A lectin receptor-like kinase mediates pattern-triggered salicylic acid signaling [J]. Plant Physiology, 2017, 174(4): 2501−2514. https://doi.org/10.1104/pp.17.00404 doi: 10.1104/pp.17.00404 [13] Wang Y, Cordewener J H G, America A H P, et al. Arabidopsis lectin receptor kinases LecRK-IX. 1 and LecRK-IX. 2 are functional analogs in regulating Phytophthora resistance and plant cell death [J]. Molecular Plant-Microbe Interactions, 2015, 28(9): 1032−1048. https://doi.org/10.1094/MPMI-02-15-0025-R doi: 10.1094/MPMI-02-15-0025-R [14] Singh P, Kuo Y C, Mishra S, et al. The lectin receptor kinase-VI. 2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity [J]. The Plant Cell, 2012, 24(3): 1256−1270. https://doi.org/10.1105/tpc.112.095778 doi: 10.1105/tpc.112.095778 [15] Balagué C, Gouget A, Bouchez O, et al. The Arabidopsis thaliana lectin receptor kinase LecRK-I. 9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling [J]. Molecular Plant Pathology, 2017, 18(7): 937−948. https://doi.org/10.1111/mpp.12457 doi: 10.1111/mpp.12457 [16] Choi J, Tanaka K, Cao Y R, et al. Identification of a plant receptor for extracellular ATP [J]. Science, 2014, 343(6168): 290−294. https://doi.org/10.1126/science.343.6168.290 doi: 10.1126/science.343.6168.290 [17] Wang Z K, Cheng J Y, Fan A Q, et al. LecRK-V, an L-type lectin receptor kinase in Haynaldia villosa, plays positive role in resistance to wheat powdery mildew [J]. Plant Biotechnology Journal, 2018, 16(1): 50−62. https://doi.org/10.1111/pbi.12748 doi: 10.1111/pbi.12748 [18] 王昊, 戎伟. WRR4C基因正调控拟南芥对橡胶树白粉菌和豇豆白粉菌的抗病性[J]. 植物病理学报, 2024, 54(2): 355−363. https://doi.org/10.13926/j.cnki.apps.001026 doi: 10.13926/j.cnki.apps.001026 [19] Wu H, Pan Y W, Di R, et al. Molecular identification of the powdery mildew fungus infecting rubber trees in China [J]. Forest Pathology, 2019, 49(5): e12519. https://doi.org/10.1111/efp.12519 doi: 10.1111/efp.12519 [20] 魏振江. 中国野生毛葡萄VqLecRK1抗白粉病功能研究及互作蛋白筛选[D]. 杨凌: 西北农林科技大学, 2022. doi: 10.27409/d.cnki.gxbnu.2022.000872 [21] Bouwmeester K, De Sain M, Weide R, et al. The lectin receptor kinase LecRK-I. 9 is a novel Phytophthora resistance component and a potential host target for a RXLR effector [J]. PLoS Pathogens, 2011, 7(3): e1001327. https://doi.org/10.1371/journal.ppat.1001327 doi: 10.1371/journal.ppat.1001327 [22] Kutschera A, Dawid C, Gisch N, et al. Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants [J]. Science, 2019, 364(6436): 178−181. https://doi.org/10.1126/science.aau1279 doi: 10.1126/science.aau1279 [23] Luo X M, Wu W, Liang Y B, et al. Tyrosine phosphorylation of the lectin receptor-like kinase LORE regulates plant immunity [J]. The Embo Journal, 2020, 39(4): e102856. https://doi.org/10.15252/embj.2019102856 doi: 10.15252/embj.2019102856 [24] Wang Z, Gou X P. The first line of defense: receptor-like protein kinase-mediated stomatal immunity [J]. International Journal of Molecular Sciences, 2021, 23(1): 343. https://doi.org/10.3390/ijms23010343 doi: 10.3390/ijms23010343 [25] Mei S S, Hou S G, Cui H T, et al. Characterization of the interaction between Oidium heveae and Arabidopsis thaliana [J]. Molecular Plant Pathology, 2016, 17(9): 1331−1343. https://doi.org/10.1111/mpp.12363 doi: 10.1111/mpp.12363 [26] Mei S S, Song Y X, Zhang Z E, et al. WRR4B contributes to a broad-spectrum disease resistance against powdery mildew in Arabidopsis [J]. Molecular Plant Pathology, 2024, 25(1): e13415. https://doi.org/10.1111/mpp.13415 doi: 10.1111/mpp.13415 [27] Pruitt R N, Locci F, Wanke F, et al. The EDS1–PAD4–ADR1 node mediates Arabidopsis pattern-triggered immunity [J]. Nature, 2021, 598(7881): 495−499. https://doi.org/10.1038/s41586-021-03829-0 doi: 10.1038/s41586-021-03829-0 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 270
- HTML全文浏览量: 146
- 被引次数: 0
