

 下载:
下载:
-
木棉(Bombax ceiba)是锦葵科(Malvaceae)木棉亚科(Bombacaceae)木棉属(Bombax)植物[1],为亚洲热带地区森林生态系统中的优势种和“伞护种”[2]。然而,当前木棉自然种群的年龄结构呈衰老型,实生幼苗与幼树较少[3],表明其种群自然更新受限,但原因尚不明确。本研究团队在前期研究中观察发现,离斑棉红蝽(Dysdercus cingulatus)的若虫和成虫存在大量取食和搬运木棉种子的现象,这可能影响木棉种子的萌发与扩散。长期以来,昆虫对植物种子的取食行为多被简单归类为种间对抗关系,视为植物繁殖的限制因素之一[4]。然而,近年来越来越多的研究揭示,昆虫与植物种子的互作远比传统认知复杂,二者在长期进化过程中形成了多样的协同进化关系。例如,小檗科(Berberidaceae)[5]、紫堇属(Corydalis)[6]、百部属(Stemona)[7-8]的种子特化出适配昆虫口器的尖齿,或拟态为昆虫猎物的油质体作为报酬,吸引蚁科(Formicidae)、胡蜂科(Vespidae)等昆虫帮助其传播种子;球果假沙晶兰(Monotropastrum humile)、黄筒花(Phacellanthus tubiflorus)等则进化出肉质果实,且果实贴近地面,便于地面活动的两种驼螽(Tachycines elegantissima、Diestrammena japanica)取食,其种子加厚的木质化种皮能够保护种子通过驼螽的消化道而不被破坏[9]。被昆虫蛀食的卷果涩荠(Malcolmia scorpioides)种子能提前萌发,以避免种内生态位竞争[10]。研究发现,人为蚀刻部分种皮或经过昆虫捕食的种子萌发速率更高,其核心在于蚀刻或取食行为提高了种皮的吸水能力,从而促进种子萌发并提升幼苗的种内竞争优势[11-13]。
为探究离斑棉红蝽不同龄级对木棉种子的取食行为差异及其影响,本研究系统分析了各龄级离斑棉红蝽单独取食与共同取食木棉种子的比例,结合木棉种子解剖、显微计算机层析成像等方法,揭示了各龄级离斑棉红蝽的取食部位偏好与刺吸深度,量化了不同取食方式对木棉种子种皮、子叶及胚结构的影响程度,以期为植食性昆虫与寄主植物的协同进化研究及木棉天然林种质资源的保护提供新的视角与实证案例。
-
本研究的野外实验地点位于海南省昌江黎族自治县叉河镇排岸村(19°14′30″N,108°58′38″E),属热带季风气候,年平均气温25.5℃,年降水量1 676 mm(数据来源:海南省气象局,
http://www.hainanqx.cn ),该区域木棉分布广泛、种群规模大。 -
木棉种子通常于4月上旬至4月下旬集中成熟,因此,本研究于2025−04−11—17,每天5:30—19:30对离斑棉红蝽取食动态进行全周期连续监测,在排岸村3个被村庄、农田、公路、河流等阻隔的木棉树集中分布区,研究离斑棉红蝽的种群大小及其对木棉种子的取食行为。每个种群的数据记录为1个重复。其中,单只离斑棉红蝽取食单粒木棉种子的行为被视为单独取食;2只以上个体同时取食同1粒木棉种子的行为被视为共同取食。
-
将5个不同龄级(2龄至成虫)共约250只离斑棉红蝽放入昆虫网箱中,再放入当日新鲜采收的自然凋落且去掉棉絮的木棉种子。观察并记录不同龄级离斑棉红蝽刺吸木棉种子部位。离斑棉红蝽的单粒种子取食时长通常为4~8 h,观察其稳定取食6 h后,用手术剪小心剪断其口器,得到带口器的木棉种子。
将带口器的种子固定于显微计算机层析成像仪(nanoVoxel-
3000 )样品台。通过XRM Companion软件调整X射线源、样品与探测器的间距,经SIDE CAM软件实时校准使样品居中;关闭舱门后启动扫描,全程保持环境安静无振动干扰。扫描原始数据导入GigaRecon软件完成三维重构,采用Avizo软件开展精细分析,明确口器刺吸路径、深度及对种皮、子叶、胚等种子结构的影响范围。 -
为明确离斑棉红蝽取食对种子内部结构的影响,制备种子硬组织切片。样品经体积分数70% 的FAA 固定液固定48 h,依次浸入体积分数60%、70%、80%、90%、95%乙醇(每个梯度处理48 h)、100% 乙醇(2 次,每次 12~24 h)完成脱水。脱水后用树脂(KULZER T7200)浸润 15~25 d,调整包埋面后,于光固化包埋机(EXAKT E520)固化12 h。采用硬组织磨片机(EXAKT E400CS),磨平组织面、制备“三明治”结构,经蓝光固化后,切取200 μm切片,经打磨抛光得到20~30 μm厚的硬组织磨片。使用苏木素−伊红高清恒染试剂盒(Servicebio G1076)完成染色,经脱水、二甲苯透明后中性树胶封片。最终用数字切片扫描仪(Pannoramic MIDI)明场扫描成像,通过CaseViewer 2.4软件实现切片1~400倍观察与目标图像截取。
-
使用R软件的lme4包构建了广义线性混合模型(GLMM)分析龄级对共同取食比例的影响,检验水准设定为α=0.05,P<0.05则认为差异有统计学意义。采用R软件进行数据可视化绘图并制作图表。
-
从图1可知,各个龄级的离斑棉红蝽之间,共同取食与单独取食偏好在不同龄级之间均无显著性差异,但随龄级增大,离斑棉红蝽单独取食比例呈上升趋势;同一龄级内,2龄、3龄和4龄若虫选择共同取食方式的比例极显著高于对应龄级选择单独取食方式的比例(P<0.01),5龄若虫和成虫的单独取食与共同取食比例接近且无显著差异(P>0.05),说明离斑棉红蝽的单独取食与共同取食行为随龄级变化表现出差异性。
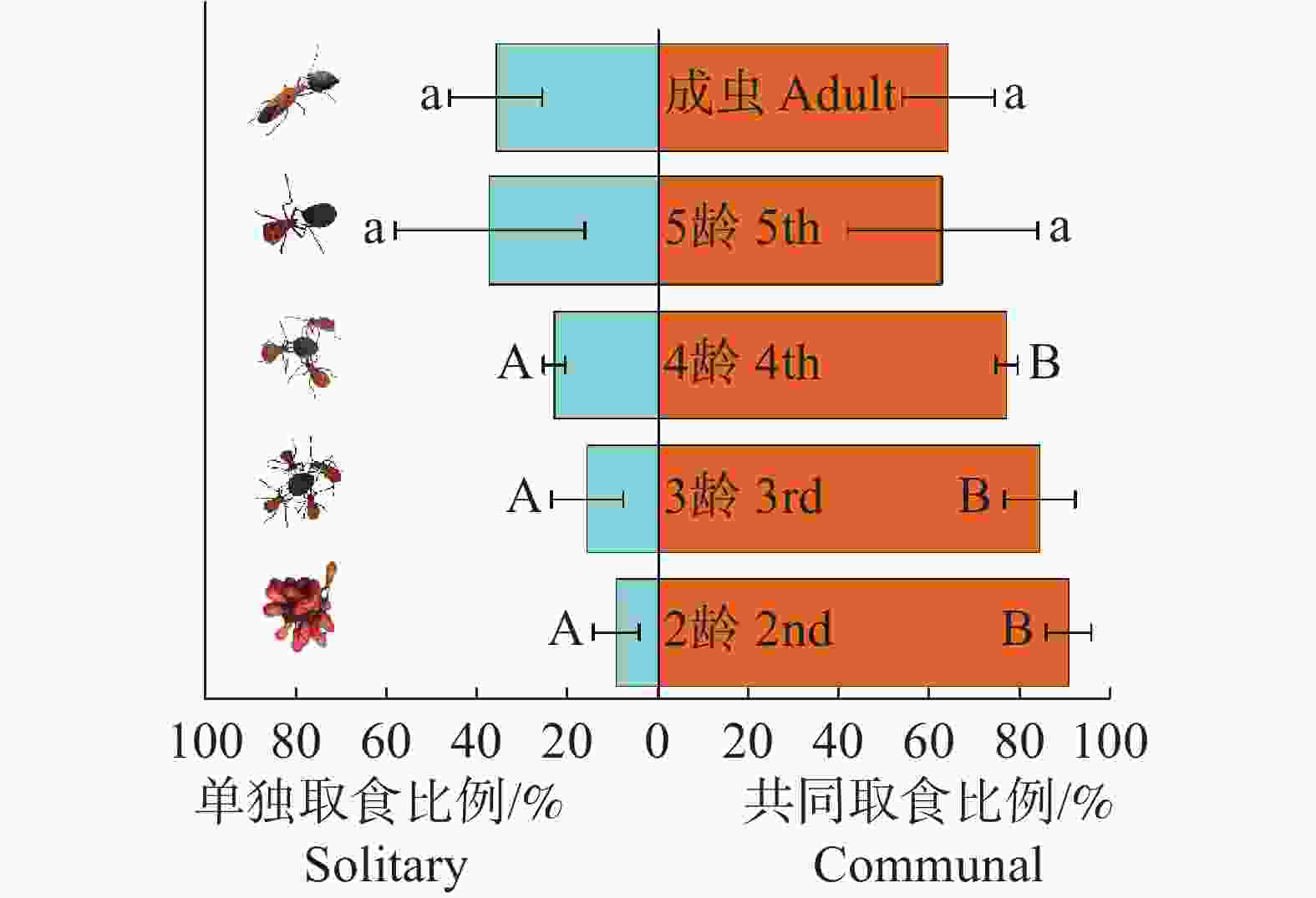
图 1 不同龄级离斑棉红蝽的取食方式
Figure 1. Predation modes of different instars of Dysdercus cingulatus
-
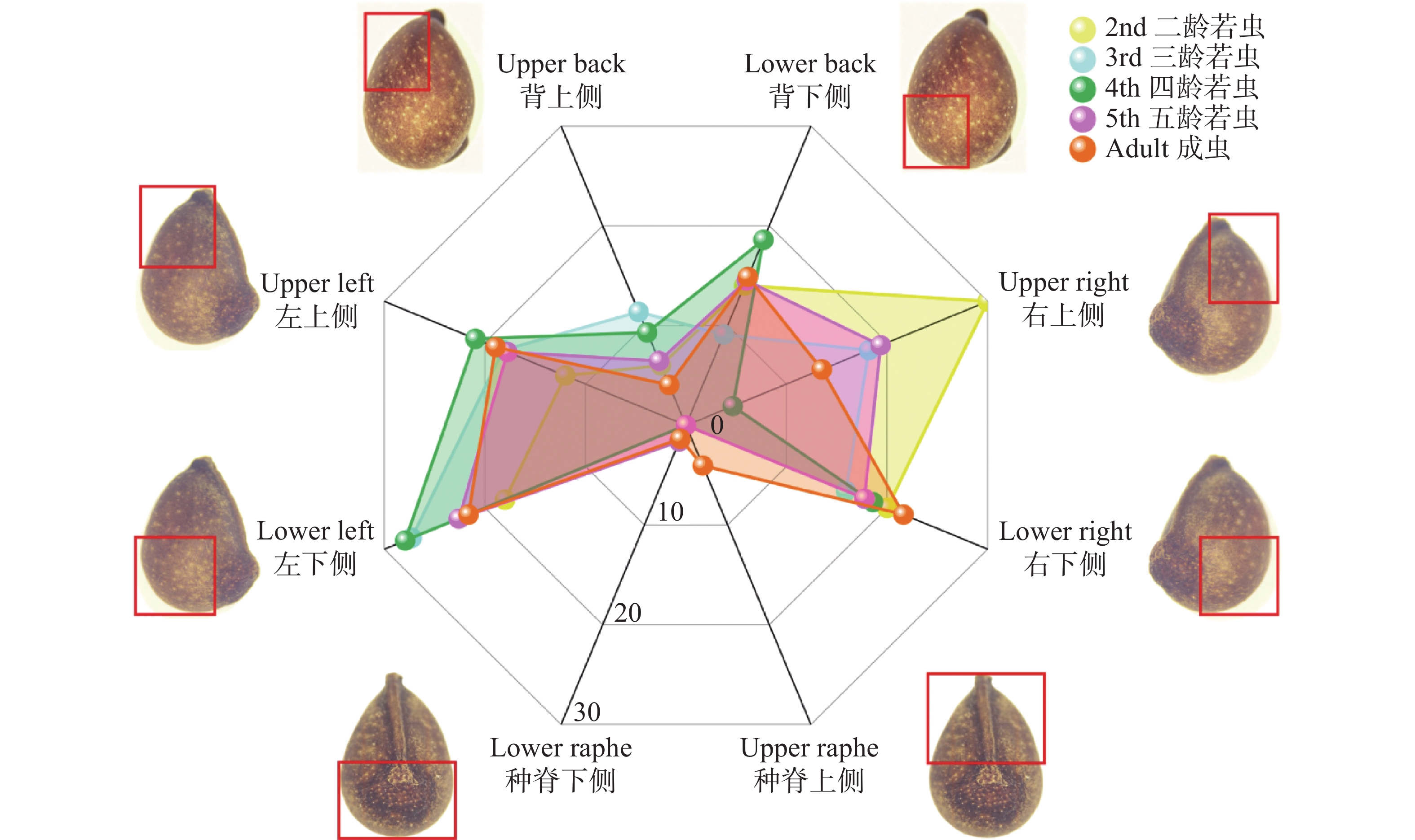
为统计不同龄级离斑棉红蝽取食木棉种子部位的差异,将木棉种子以种脊为正面、种孔为上端,将种子划分为八个区域(图2)。
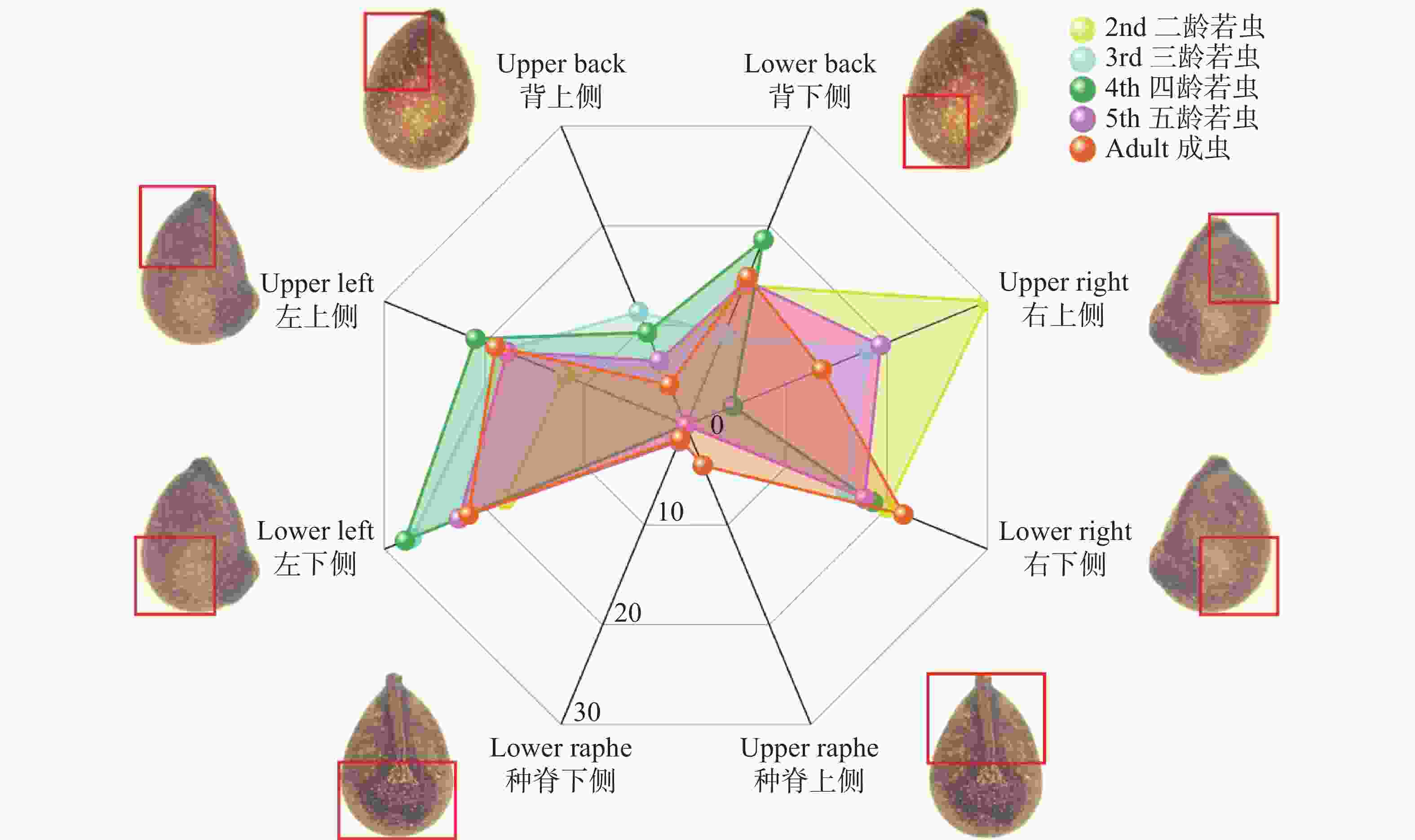
图 2 各龄级离斑棉红蝽取食木棉种子各部位比例
Figure 2. Predation sites of Bombax ceiba seeds by Dysdercus cingulatus at different instar stages
在各龄级取食种子占各龄级总取食数占比中,取食种子左侧的比例分别为:2龄30.00%、3龄45.45%、4龄48.84%、5龄40.32%、成虫40.54%;取食种子右侧比例分别为:2龄50.00%、3龄34.09%、4龄23.25%、5龄37.09%、成虫35.13%;取食种背下侧比例分别为:2龄14.00%、3龄9.09%、4龄18.60%、5龄14.52%、成虫14.86%,所有龄级均表现出对这3个部位的取食偏好性;距离种子胚结构最近的种背上侧,基本符合随龄级增大,取食此部位比例更小的趋势,比例分别为:2龄6.00%、3龄11.36%、4龄9.30%、5龄6.45%、成虫4.05%;种脊部位只有5龄若虫和成虫取食,分别为2.96%和4.05%。
在对所有龄级的离斑棉红蝽取食数量进行研究时,离斑棉红蝽取食种子左侧、右侧、背部下侧分别占总取食量的40.65%、36.26%和14.28%,这3个部位的取食行为加起来,占总取食行为的91.21%。距离胚最近的背部上侧占总取食数量的6.96%。种脊部位仅占总取食量的1.83%。不同龄级离斑棉红蝽对木棉种子的取食部位具有高度一致的选择性,均集中于远离胚结构的子叶分布区,最大程度规避了对种子萌发核心结构的破坏,且高龄个体对胚区的规避性更强。
-
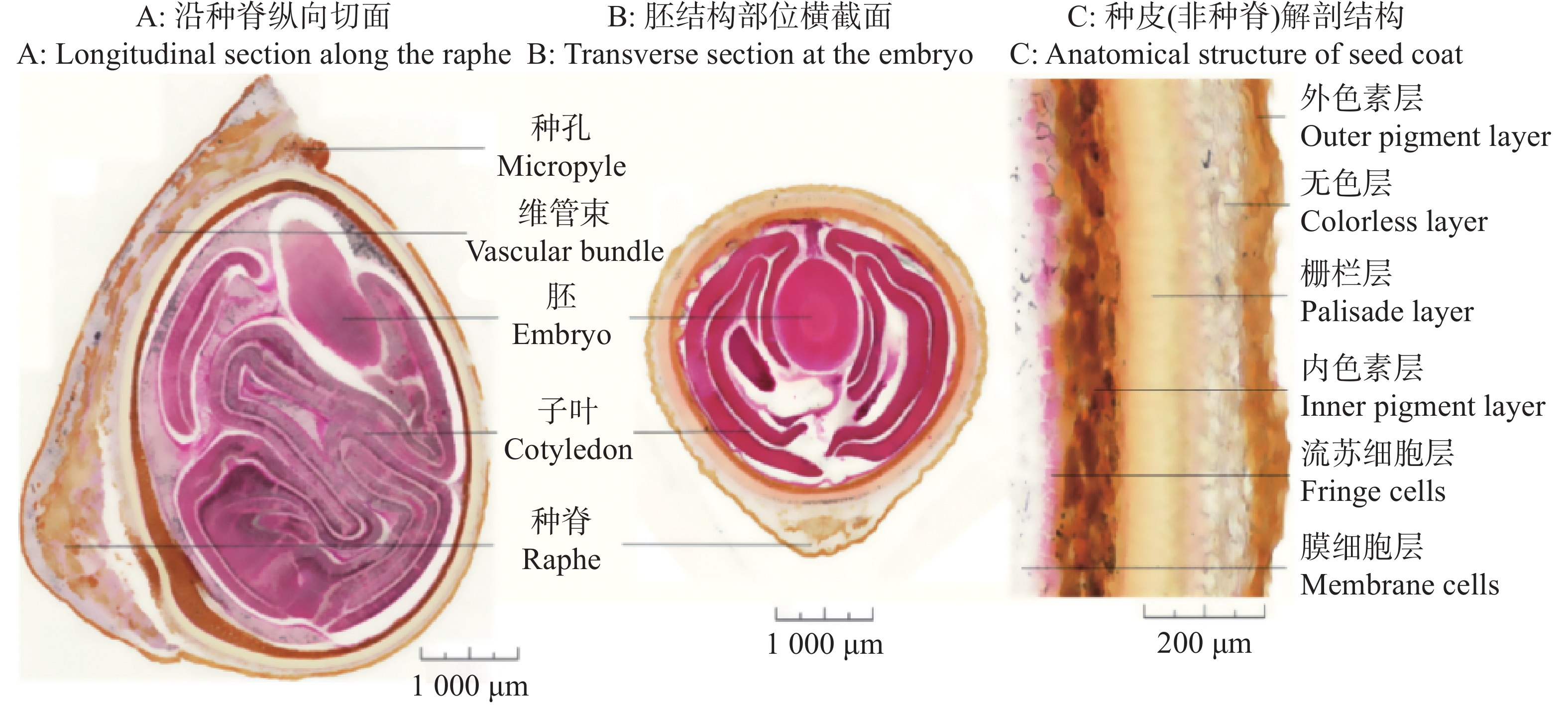
种脊是种子腹面的纵棱,为珠柄延伸形成的结构,维管束主要分布于种脊对应的种皮组织中,负责种子发育过程中养分的运输,但被栅栏层与内色素层隔绝于种子内部结构之外;种脊部分是木棉种子种皮最厚也是最坚硬的部分。胚结构位于种子顶端到种子上方约1/3处。子叶位于种子中下部分,高度折叠,并部分延伸至胚芽、胚轴所在区域,但未延伸至胚根区域(图3-A)。
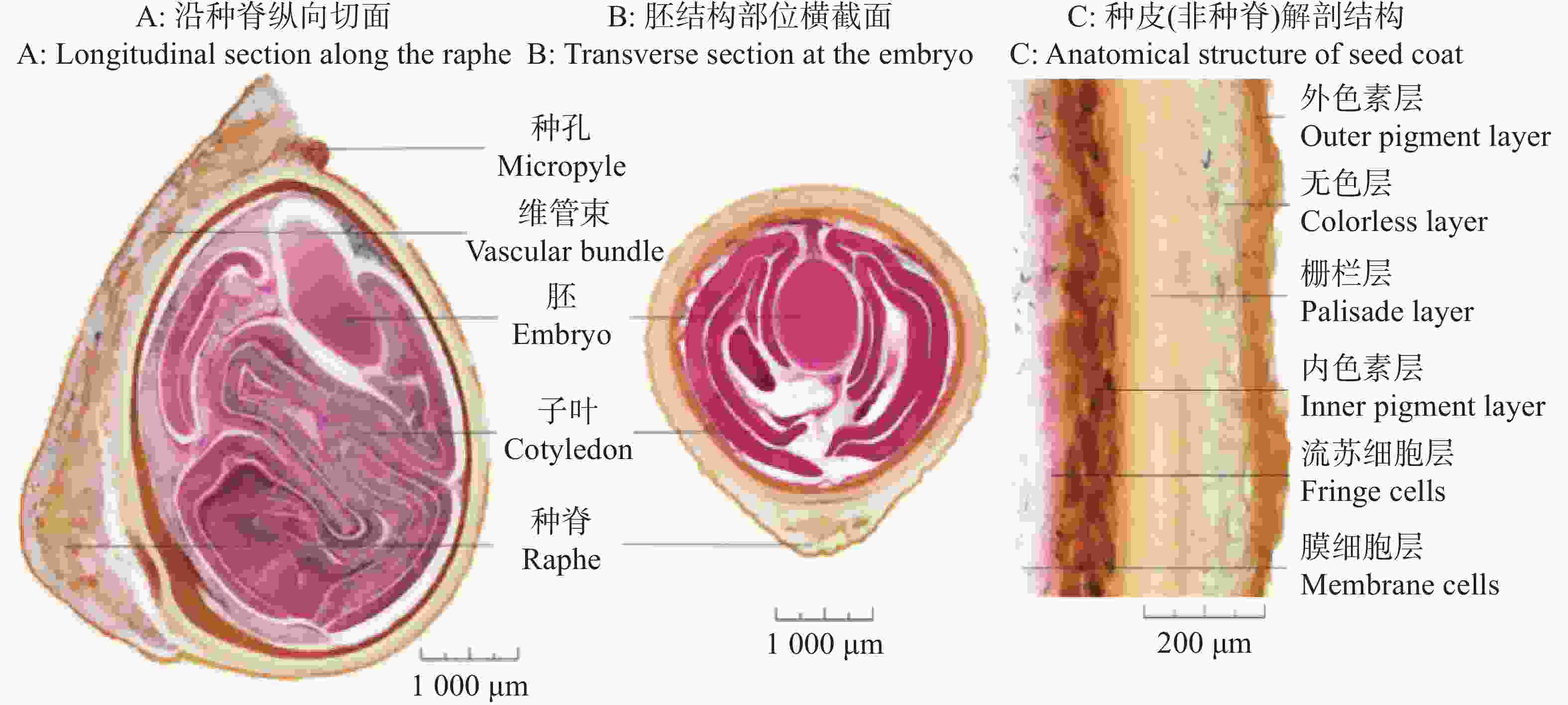
图 3 木棉种子解剖结构
Figure 3. Anatomical structure of Bombax ceiba seeds
木棉种子胚结构两侧被高度折叠的子叶紧密包裹,子叶肉质且折叠紧密,呈多层折叠状,将胚轴、胚芽等结构包裹其中,形成物理屏障,同时子叶储存的营养物质为胚萌发提供能量,兼具保护与营养双重功能(图3-B)。
木棉种子的种皮由多层特化组织构成,核心为栅栏层,由厚壁细胞组成,细胞壁木质化,硬度极高,可有效抵御机械损伤和生物侵害;但坚硬且木质化的种皮透水性和透气性较差。非种脊部位,种皮结构中维管束不明显,仅在种脊区域可见明显的维管束痕迹(图3-C)。
-
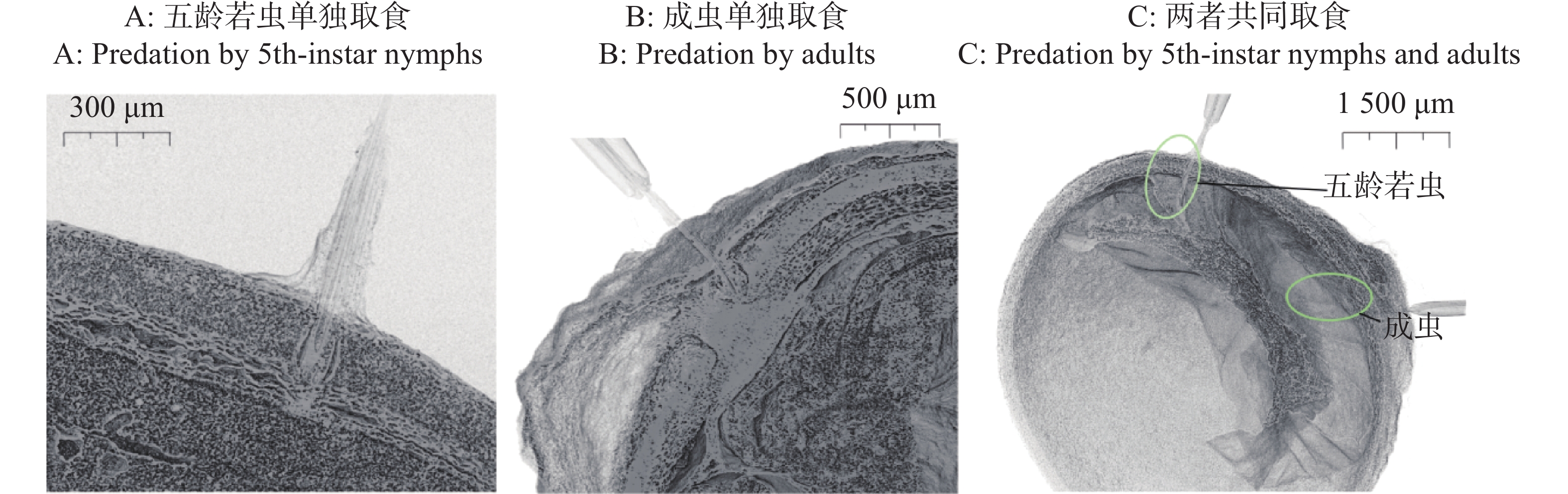
离斑棉红蝽的5龄若虫与成虫口器发育比低龄级若虫更为完善,刺吸深度更深,故选择成虫与5龄若虫进行观察。分析表明,离斑棉红蝽5龄若虫与成虫单独取食6 h时,均刺吸到木棉种子种皮栅栏层,未穿透内色素层,5龄若虫取食深度略大于200 μm与成虫取食深度略小于400 μm,对木棉种子内部结构无明显影响(图4-A—B)。
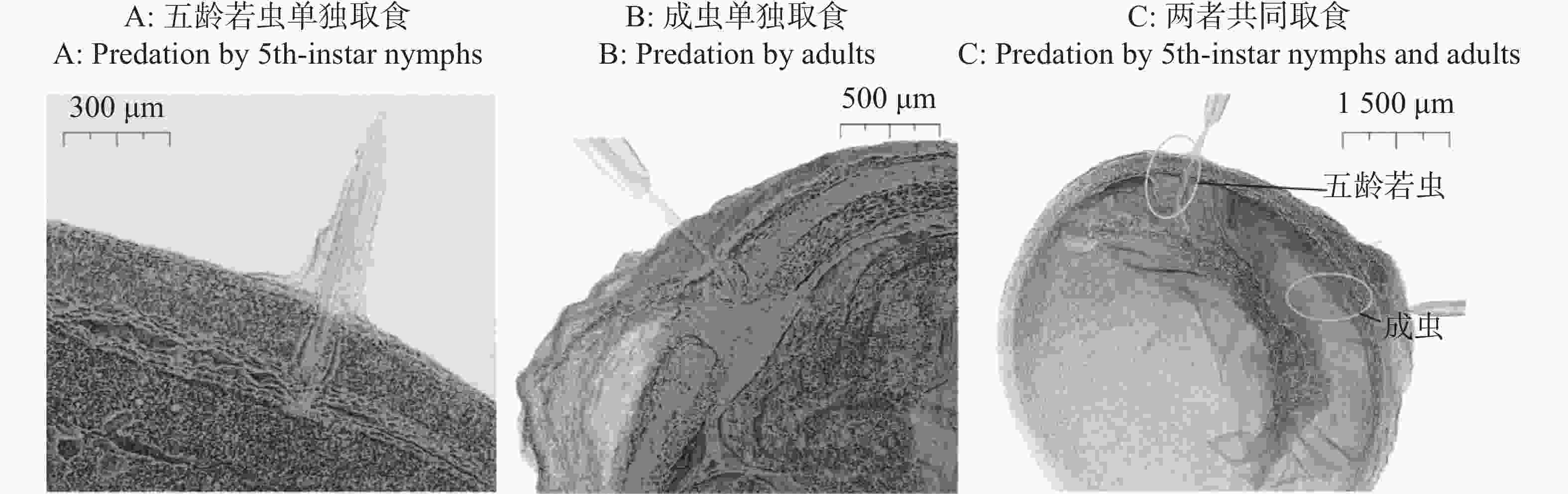
图 4 离斑棉红蝽取食木棉种子的显微计算机层析成像
Figure 4. Micro-computed tomography of Bombax ceiba seed predated by Dysdercus cingulatus
5龄若虫和成虫共同取食1粒木棉种子6 h时,口器均完全刺穿种皮,5龄若虫刺入深度略大于600 μm与成虫刺入深度略大于800 μm,能取食到木棉种子子叶等内部结构(图4-C)。
-
在昆虫与植物种子的取食互作中,取食部位的选择性是决定互作性质(对抗/互惠)的核心因素。前人研究证实,多种植食性昆虫在长期进化中形成了“胚规避”取食策略:优先消耗子叶或胚乳等营养组织,尽量避免损伤胚结构。例如,象甲属(Curculio)主要取食橡种子叶[14];蚕豆象(Bruchus rufimanus)幼虫取食豆科(Fabaceae)种子时仅消耗胚乳而不损伤胚组织[15];当1粒橡子中取食的象鼻虫(Curculio glandium)幼虫不超过5只时,它们只损伤子叶,而8~10只共同取食导致食物严重短缺时,才会取食胚组织[16]。
本研究结果与上述胚规避行为类似。离斑棉红蝽对木棉种子的取食部位表现出极强的偏好性:对离胚结构最近的种背上部区域,总取食占比仅为6.96%,且随龄级增大,该部位取食比例占该龄级总取食数的比例更小,从而使胚结构被破坏的概率更低。木棉种子的胚结构位于种子上方1/3处,且胚轴、胚芽等核心生命结构被多层折叠的肉质子叶紧密包裹,形成了天然的物理保护屏障。低龄级若虫因口器未发育完全,即便刺穿种皮也难以触及胚组织;5龄若虫与成虫取食胚所在部位的比例分别仅为6.45%和4.05%,低于2、3、4龄若虫。这可能与高龄虫体搬运种子能力更强、抢夺时体型优势更大、更易“独享”种子有关;而低龄级若虫因体型较小,搬运和抢夺能力不足,常与其他个体共同取食。离斑棉红蝽这种高度特化的取食部位选择策略,与木棉种子独特的解剖结构形成了完美的协同适应,最大程度地规避了对种子萌发关键结构的损伤。
-
Takakura等[17]发现,刺槐(Gleditsia japonica)硬种子被豆象鼻虫(Bruchidius dorsalis)取食,造成了物理损伤,但发芽率却更高,原因在于豆象鼻虫对刺槐种子刺吸形成的小孔,成为水分与氧气进入种子的通道。离斑棉红蝽单独取食时,5龄若虫单独取食口器刺入深度约为200 μm,成虫单独取食口器刺入深度约为400 μm,它们的口器均仅刺入木棉种子种皮栅栏层,未刺穿种皮;而木棉种子的种皮核心结构为木质化的栅栏层,该结构由厚壁细胞构成、硬度极高,可有效抵御生物侵害与机械损伤,但透水性与透气性差,是导致木棉种子产生休眠的关键因素[18],取食形成的种皮微损伤,恰好弥补了木棉种子非种脊部位维管束稀少、透水性差的缺陷,为种子打破物理休眠奠定了基础。同时,木棉种子的维管束主要分布于种脊部位,其他部位维管束痕迹极不明显,自然状态下水分与氧气难以通过这些部位进入种子内部。离斑棉红蝽的取食行为与豆象鼻虫类似,暗示了离斑棉红蝽取食行为可能对木棉种子萌发有积极作用。
结合刺吸部位来看,离斑棉红蝽绝大多数共同取食行为可能刺吸到木棉种子的子叶部位,有研究证实,莲玉蕊(Gustavia superba)种子不仅能耐受昆虫侵染,且在移除一半子叶储备后仍可成功萌发[19],五台山栎(Quercus wutaishanica)种子在切除1/4子叶后,发芽指数甚至显著高于对照,仅在切除1/2以上的子叶时才对发芽产生负面影响[20]。这些结果都说明,子叶少量营养流失不会直接导致萌发能力丧失。对于共同取食的离斑棉红蝽而言,其口器虽可以刺穿种皮,但木棉种子的胚轴、胚芽等核心结构被折叠紧密的子叶完全包裹,形成了致密的物理阻隔层,胚与种皮最近处约为800 μm,只有成虫刺吸深度虽略大于这个距离,但结合成虫刺吸此部概率仅占成虫取食数量的4.05%,其对木棉胚结构完整性构成的威胁微乎其微。有研究显示,同为红蝽科(Pyrrhocoridae)与锦葵科的取食案例中,始红蝽(Pyrrhocoris apterus)幼虫阶段的口针穿刺能力不足,无法突破同为锦葵科的大叶椴(Tilia platyphyllos)种子[21],结合5龄若虫刺吸深度略大于600 μm,更低龄级若虫口器即使能刺穿种皮,也仅能到达子叶,更无法接触到胚结构。木棉种子的子叶保护结构与低龄若虫的口器发育不完全,共同构成了胚结构的双重保护屏障,这一特征使得离斑棉红蝽低龄级若虫即使在口器穿透种皮的情况下,也无法对木棉种子造成较大损伤。
这种非致命性取食的取食策略并非偶然,而是植食性昆虫与寄主种子在长期协同进化过程中形成的适应性特征。在蝽类与种子植物的互作中,取食行为通过打破种子休眠、促进种子萌发,对植物种群更新产生正向调控作用,例如褐曲跗缘蝽(Hyalymenus tarsatus)对德拉蒙田菁(Sesbania drummondii)的取食行为可以提高种子萌发率[12];澳洲野生棉(Gossypium sturtianum)和瑟伯氏棉(Gossypium thurberi)被尖长蝽属的Oxycarenus luctuosus、盾蝽科(Scutelleridae)的Sphyrocoris punctellus,以及棉铃象甲(Anthonomus grandis)取食后,澳洲野生棉种子萌发率是未被捕食种子的2~3倍;瑟伯氏棉被昆虫取食后种子萌发率约为未被捕食种子的2倍[11]。对离斑棉红蝽而言,不同龄级与不同取食方式形成的取食行为与取食深度差异,在满足自身营养需求的同时最大程度保留了木棉种子的结构完整性,而这一特征印证了刺吸式种子植食昆虫与寄主植物在长期协同演化中可形成非致死性的互惠互作关系。
-
本研究采用野外调查、木棉种子解剖及显微计算机层析成像技术,明确了不同龄级离斑棉红蝽对木棉种子的取食行为差异。结果表明:取食行为存在显著的龄级分化——2至4龄若虫以共同取食为主,5龄若虫和成虫的单独与共同取食比例无显著差异,且整体单独取食比例随龄级上升呈递增趋势。所有龄级的取食部位高度一致,均精准规避胚区,集中取食种子左侧、右侧及背侧下部(占总取食行为的91.21%),其中高龄个体对胚结构所在区域的规避性更强。取食深度受龄级与取食方式共同影响:单独取食时,5龄若虫和成虫仅刺入种皮栅栏层;共同取食时,二者可刺穿种皮取食子叶,但对胚结构的威胁较小。综上,离斑棉红蝽不同龄级分化的取食策略,在满足自身营养需求的同时最大程度保留了木棉种子的萌发潜力,为解析二者之间的协同进化关系提供了初步依据。
Predation behavior of Dysdercus cingulatus (Fabricius) at different instars on seeds of Bombax ceiba L.
-
摘要: 木棉(Bombax ceiba)是亚洲热带与亚热带地区森林生态系统的优势种和“伞护种”,其种子扩散及植食性昆虫取食是影响种群更新的关键。为探究主要取食者离斑棉红蝽(Dysdercus cingulatus)对木棉种子的取食行为差异、直接危害及其对种子萌发与扩散的潜在作用,本研究通过野外观察、显微计算机层析成像扫描和种子解剖等方法,分析了离斑棉红蝽取食行为随龄级的分化特征及其对种子的影响。结果表明:1)2至4龄若虫常聚集取食同一颗种子,而5龄若虫与成虫多单独取食,并将种子拖拉至较远距离;2)成虫与若虫主要从远离胚结构的种子左侧、右侧及背部下侧进行刺吸,这3个部位的刺吸次数占总数的91.21%;3)单独取食时,5龄若虫与成虫的刺吸深度分别约为200 μm和400 μm,均未刺穿种皮;共同取食时,二者刺吸深度分别超过600 μm和800 μm,均能刺穿种皮。本研究证实,不同龄级的离斑棉红蝽均选择远离胚的部位刺吸种子,并存在长距离拖拉种子的行为,这可能对木棉种子的萌发与扩散具有积极作用。Abstract: Bombax ceiba is a dominant umbrella species in forest ecosystems in tropical Asia, and seed dispersal and predation by insects are key factors affecting its population regeneration. Both adults and nymphs of Dysdercus cingulatus (Fabricius) are the main seed predators of B. ceiba. To investigate the differences in predation behavior of Dysdercus cingulatus (Fabricius), the main seed predators of B. ceiba, and their direct damage to B. ceiba seeds, and their effects on the germination and dispersal of B. ceiba seeds, we conducted field observations, micro-computed tomography scanning, and anatomical dissection of B. ceiba seeds to examine the instar-dependent differentiation of the predation behavior of D. cingulatus. The results showed that the 2nd to 4th instar nymphs usually aggregated to perform communal predation on the same B. ceiba seed, while 5th instar nymphs and adults tended to engage in solitary predation and drag the seeds to relatively long distances. Both adults and nymphs of D. cingulatus mainly pierced and sucked from the left, right, and lower dorsal regions of the seeds, far from the region housing the embryo, and predation events on these three regions accounted for 91.21% of the total. Under solitary predation, the penetration depth was approximately 200 μm for 5th instar nymphs and 400 μm for adults, and neither of the nymphs and adults pierced through the seed coat. However, under communal predation, the penetration depth exceeded 600 μm for 5th instar nymphs and was slightly greater than 800 μm for adults, both of which could fully pierce through the seed coat. These findings confirm that D. cingulatus at different instar stages exhibit the behavior of dragging B. ceiba seeds over long distances, and that all their predation sites are far from the embryo of B. ceiba seeds, which may play a positive role in the seed dispersal and germination of B. ceiba.
-
图 2 各龄级离斑棉红蝽取食木棉种子各部位比例
Fig. 2 Predation sites of Bombax ceiba seeds by Dysdercus cingulatus at different instar stages
-
[1] 向文倩, 王文娟, 任明迅. 木棉文化的生物多样性传统知识及其传承与利用[J]. 生物多样性, 2023, 31(3): 22524. https://doi.org/10.17520/biods.2022524 doi: 10.17520/biods.2022524 [2] Jain V, Verma S K, Katewa S S. Myths, traditions and fate of multipurpose Bombax ceiba L. - an appraisal [J]. Indian Journal of Traditional Knowledge, 2009, 8(4): 638−644. [3] 王健, 水庆艳, 石晶, 等. 海南木棉植物资源调查与分类初步研究[J]. 亚热带植物科学, 2010, 39(1): 53−56. https://doi.org/10.3969/j.issn.1009-7791.2010.01.013 doi: 10.3969/j.issn.1009-7791.2010.01.013 [4] 李宏俊, 张知彬. 动物与植物种子更新的关系Ⅱ. 动物对种子的捕食, 扩散, 贮藏及与幼苗建成的关系[J]. 生物多样性, 2001, 9(1): 2537. (查阅网上资料,不确定本条文献doi信息是否正确,请确认) https://doi.org/10.1088/0256-307X/18/11/313 [5] Lengyel S, Gove A D, Latimer A M, et al. Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey [J]. Perspectives in Plant Ecology, Evolution and Systematics, 2010, 12(1): 43−55. https://doi.org/10.1016/j.ppees.2009.08.001 doi: 10.1016/j.ppees.2009.08.001 [6] Zhu Y, Wang D. Leaf volatiles from two corydalis species lure a keystone seed-dispersing ant and enhance seed retrieval [J]. Sociobiology, 2018, 65(3): 370−374. https://doi.org/10.13102/sociobiology.v65i3.2726 doi: 10.13102/sociobiology.v65i3.2726 [7] Pellmyr O. Yellow jackets disperse Vancouveria seeds (Berberidaceae) [J]. Madroñ o, 1985, 32(1): 56. https://doi.org/10.2307/41424516 doi: 10.2307/41424516 [8] Qin R M, Wen P, Corlett R T, et al. Plant-defense mimicry facilitates rapid dispersal of short-lived seeds by hornets [J]. Current Biology, 2022, 32(15): 3429−3435. e5. https://doi.org/10.1016/j.cub.2022.06.034 doi: 10.1016/j.cub.2022.06.034 [9] Suetsugu K. Independent recruitment of a novel seed dispersal system by camel crickets in achlorophyllous plants [J]. New Phytologist, 2018, 217(2): 828−835. https://doi.org/10.1111/nph.14859 doi: 10.1111/nph.14859 [10] 文晓虎, 张玲卫, 刘会良, 等. 昆虫蛀食对卷果涩荠种子萌发特征的影响[J]. 种子, 2023, 42(5): 103−109. https://doi.org/10.16590/j.cnki.1001-4705.2023.05.103 doi: 10.16590/j.cnki.1001-4705.2023.05.103 [11] Karban R, Lowenberg G. Feeding by seed bugs and weevils enhances germination of wild Gossypium species [J]. Oecologia, 1992, 92(2): 196−200. https://doi.org/10.1007/BF00317364 doi: 10.1007/BF00317364 [12] Lundbye H, Johansson D K, Andersen M R, et al. The effect of a seed-sucking bug on seed germination of an arctic cushion plant [J]. É coscience, 2012, 19(3): 209−212. https://doi.org/10.2980/19-3-3510 doi: 10.2980/19-3-3510 [13] Rolston M P. Water impermeable seed dormancy [J]. The Botanical Review, 1978, 44(3): 365−396. https://doi.org/10.1007/BF02957854 doi: 10.1007/BF02957854 [14] Hou X G, Yi X F, Yang Y Q, et al. Acorn germination and seedling survival of Q. variabilis: effects of cotyledon excision [J]. Annals of Forest Science, 2010, 67(7): 711. https://doi.org/10.1051/forest/2010036 doi: 10.1051/forest/2010036 [15] 敏玉霞, 毛玉萍, 柳慧玲, 等. 蚕豆象综述[J]. 农学学报, 2024, 14(2): 30−35. https://doi.org/10.11923/j.issn.2095-4050.cjas2023-0069 doi: 10.11923/j.issn.2095-4050.cjas2023-0069 [16] Reut M, Bonal R, Chrabąszcz M, et al. Cannibalism as competition strategy in larvae of the acorn weevil Curculio glandium (Coleoptera: Curculionidae) [J]. Diversity, 2023, 15(2): 145. https://doi.org/10.3390/d15020145 doi: 10.3390/d15020145 [17] Takakura K. The specialist seed predator Bruchidius dorsalis (Coleoptera: Bruchidae) plays a crucial role in the seed germination of its host plant, Gleditsia japonica (Leguminosae) [J]. Functional Ecology, 2002, 16(2): 252−257. https://doi.org/10.1046/j.1365-2435.2002.00619.x doi: 10.1046/j.1365-2435.2002.00619.x [18] Gama-Arachchige N S, Baskin J M, Geneve R L, et al. Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes [J]. Annals of Botany, 2013, 112(1): 69−84. https://doi.org/10.1093/aob/mct094 doi: 10.1093/aob/mct094 [19] Dalling J W, Harms K E. Damage tolerance and cotyledonary resource use in the tropical tree Gustavia superba [J]. Oikos, 1999, 85(2): 257−264. https://doi.org/10.2307/3546491 doi: 10.2307/3546491 [20] 闫兴富, 方苏, 石淳, 等. 模拟动物取食子叶对辽东栎种子萌发和幼苗早期生长的影响[J]. 生态学杂志, 2014, 33(4): 973−981. https://doi.org/10.13292/j.1000-4890.2014.0098 doi: 10.13292/j.1000-4890.2014.0098 [21] Kristenová M, Exnerová A, Štys P. Seed preferences of Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae): are there specialized trophic populations? [J]. European Journal of Entomology, 2011, 108(4): 581−586. https://doi.org/10.14411/eje.2011.075 doi: 10.14411/eje.2011.075 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 124
- HTML全文浏览量: 76
- 被引次数: 0
