

-
国际生物计划(International Biological Programme,IBP)将物候学定义为研究周期性生物事件的时间、这一时间变化的生物和非生物驱动因素及相同或不同物种阶段之间相互关系的科学[1],如植物开花和展叶的开始时间、结束时间等,对生态系统功能的各个方面都有重要影响[2-3]。叶物候持续时间对生态系统功能和树木生产力也有很大影响[4]。对于落叶树种,春季展叶和秋季叶衰老的时间决定了树冠的持续时间,从而决定了其年际营养生长速率和繁殖成功率[5]。植物碳汇潜力和植被对气候系统的反馈也依赖于叶物候的持续时间,而叶物候持续时间的变化是森林气候变化的一项指标[6-7]。在过去的40 a里,树木的生长期平均每10 a延长了2~3 d[8-9]。由于森林在陆地碳储存方面发挥着重要作用,其叶物候持续时间的变化不但将对森林产品和生物多样性,而且对全球气候本身都有重要影响[10],因此,研究植物叶物候持续时间及其驱动机制对于了解气候变化对森林生态系统的影响具有重要意义[7],但以往的叶物候研究对叶物候持续时间关注较少[11]。植物叶物候在种间和种内存在显著差异,且沿着环境梯度呈现明显的变化。研究大多使用平均性状值来研究物种尺度的功能性状,隐藏了许多内部差异,包括种内不同种群间的差异[12],特别是广布种[13]。因此,为了预测广布树种的命运,往往需要评估物种对当地条件的适应机制及其应对环境变化的适应潜力。气候变化对植物的叶物候有着显著的影响,其中温度是控制植物叶物候最主要的因素[14]。纬度代表的是一个更加复杂的环境条件综合体,大量非生物环境因子存在明显的交互作用,形成了强烈的自然选择压力,驱动广布种的快速适应进化过程[15]。但是目前尚缺对广布种沿纬度梯度的叶物候变化的研究,阻碍了对其未来可能受气候变化影响的评估与分析。
构树(Broussonetia papyrifera)是一种属于桑科、构属的乔木树种,分布于热带、亚热带地区以及我国绝大部分地区[16]。构树是一种先锋植物,分布广、适应性强、抗性强,是适合我国三北地区种植的防护林和山区大力推广种植的树种[17]。由于叶片本质上是获取能量的器官,因此季节性和非季节性环境中叶子在时间(叶物候)和空间(冠层结构)上的排列可被视为植物碳获取策略的核心要素[18]。构树不同种源的叶表现出了明显的地理变异趋势,其叶形态变异的主要环境因素可能是与采种点的经纬度相关的气候差异[19],如构树各种源叶片的叶面积、叶长、叶宽和绒毛密度与采种点纬度呈极显著的负相关,这表明低纬度种源比高纬度种源叶片更大[19]。目前关于几个地理上分布较远的种群的展叶物候、特定物种种群之间的物候差异及其驱动机制方面的研究较少[20]。这些问题不仅限制了评估气候变化对构树生产力的影响,也将影响预测构树植物的入侵风险和碳汇能力变化。本研究基于中国物候监测网多个城市站点记载的1963—2008年的物候数据拟研究以下3个问题:(1)构树叶物候持续时间是否有明显的纬度格局?(2)构树叶物候持续时间的延长是否是由展叶和落叶期共同引起的?(3)驱动构树叶物候性状纬度差异的关键环境因子是什么?以期从纬度地理差异角度研究广布种不同种群的物候分化,为评估物种未来的分布范围变化及灭绝风险提供参考。
-
选取中国暖温带和亚热带地区,该区域包含了半湿润和半干旱2个气候区。多为大陆性季风气候,该区域四季分明,降雨集中在夏季,冬季干燥,水热条件在季节与空间上存在差异,植物物候也表现出显著的时空变化特征,对气候变化响应较为敏感[21],因此,适合作为研究植物物候变化的区域。
选用的物候观测资料来自中国物候观测网(Chinese Phenology Observation Network,CPON),由于人为观测和虫害等因素导致物候数据部分缺失[22],依据观测数据较丰富和连续性较好的原则,选择含观测期内(1963—1968年,1973—1978年,1983—1991年,2003—2007年)的展叶期、叶变色期和落叶期的叶物候观测记录[23],为了满足统计分析的最小样本量,排除了数据少于10 a的站点记录[23],共有九个物候观测站点有充足的构树叶物候数据符合筛选要求,分别是:北京、西安、洛阳、芜湖、镇江、长沙、南昌、贵阳和桂林,纬度范围25°14′—39°9′N,每个站点的具体纬度见表1。采用儒略日期(Julian day)换算法(1月1日为第1天,2月1日为第32天,依此类推),将展叶期、落叶期转换为距离当年1月1日的天数[24]。然后计算每一个站点内每一叶物候期累年平均时间,作为该站点叶物候数据。
表 1 9个站点构树叶物候数据和纬度坐标数据
持续
时间/d持续时间
标准差展叶
期/d落叶
期/d纬度 北京 202.38 10.64 113 317 39°90′ 西安 230.83 10.25 99 331 33°42′ 洛阳 229.69 8.26 99 328 34°35′ 芜湖 234.46 9.89 98 332 31°32′ 镇江 247.45 7.06 95 343 32°16′ 长沙 264.43 11.30 92 343 28°12′ 南昌 269.57 10.57 85 354 28°40′ 贵阳 271.43 13.95 85 352 26°11′ 桂林 278.50 23.36 83 361 25°14′ -
气候数据来源于中国气象数据网(
http://data.cma.cn/ )[25],包括9个物候站点对应的气象站点的年平均气温、累年月平均气温、累年月平均降水量(1981—2010年)[25]。之前的研究结果揭示展叶季前1~2个月的平均气温影响展叶期的年度变化,2~3个月的平均气温影响落叶期的年度变化[21]。在此基础上,笔者根据9个研究站点构树平均物候数据(平均展叶期为4月5日),将2—3月平均气温定义为展叶季前温度,10—11月平均气温定义为落叶季前温度(平均落叶期为12月7日),并用12月至翌年2月的平均气温作为冬季平均气温;降水量选取时间段与此相同。在选取气象站数据点时依据物候观测站点选取同一位置的气象站点,其中除镇江站外,其余物候观测站点均有与之地理位置相同的气象站点,则直接使用这些站点的气象数据[26]。镇江无与之地理位置相同的气象站点,以地理位置最近的南京站替代[26]。 -
所有统计分析用R语言4.2.2版(R Core Team,2022)。分别对3个构树叶物候进行统计分析,分别为:展叶期、落叶期和持续时间,计算每个站点内构树叶物候儒略日的平均值。叶物候持续时间计算为落叶期与展叶期的差值。对于秋季物候,主要关注的是叶片脱落,而不是叶片颜色的变化,因为任何剩余的绿叶都能够进行大量碳同化,支持生物量增长[27]。为了评价纬度对构树叶物候的影响,采用线性模型(LM)。笔者将3个叶物候性状数据与以往研究认为与展叶落叶物候相关的6个环境因素进行随机森林回归(RF)来检测与叶持续时间最相关的变量。6个环境因子分别是:展叶季前温度、冬季温度、落叶季前温度、展叶季前降水量、冬季降水量和落叶季前降水量[28-29];选择此方法的原因是它不需要严格的数据假设,可以更好地处理多重共线性和非线性关系[30-31]。生成变量指数MSE(均方误差)以证明解释变量对响应变量的重要性[32];根据MSE指数的大小选择最关键的因素,MSE越大变量越重要。
-
1963—2007年各站点构树种群平均展叶期分布在3月25日(桂林)至4月24日(北京)之间,平均日期为4月5日。落叶期分布在11月14日(北京)至12月28日(桂林)之间,平均日期为12月7日。叶物候持续时间分布在202.38 d(北京)至278.5 d(桂林)之间,平均为247.63 d(表1)。
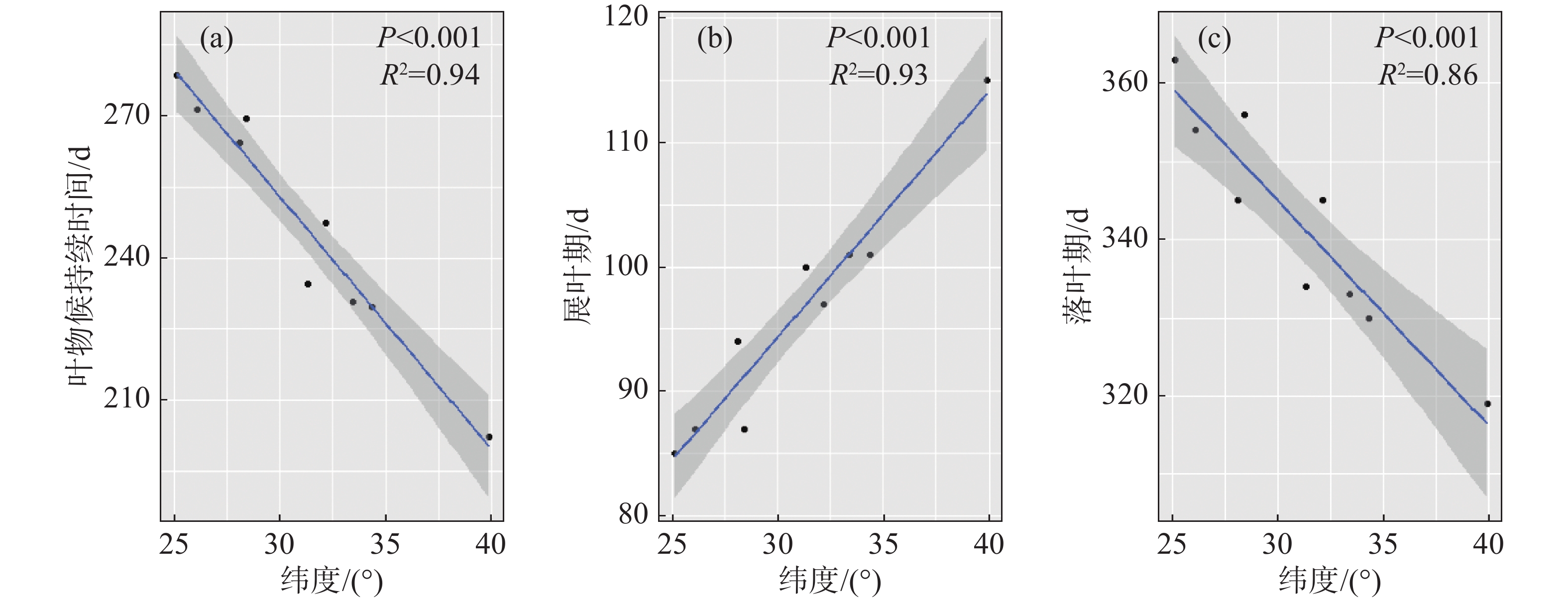
回归结果显示,构树的叶物候特征,无论是展叶期、落叶期和持续时间,都有明显的纬度梯度格局(展叶期:R2=0.93,P<0.001;落叶期:R2=0.86,P<0.001;叶持续时间:R2=0.94,P<0.001)(表2)。在种群水平,种群分布每降低1个纬度,构树展叶期平均提前1.97 d,落叶平均延迟2.88 d,持续时间增加5.32 d。展叶期有较落叶期更显著的纬度梯度格局,但是落叶期的斜率绝对值大于展叶(表2,图1)。
表 2 构树展叶期、落叶期和叶物候持续时间与纬度回归结果
持续天数 展叶期 落叶期 R² 0.94 0.93 0.86 P <0.001 <0.001 <0.001 斜率 −5.32 1.97 −2.88 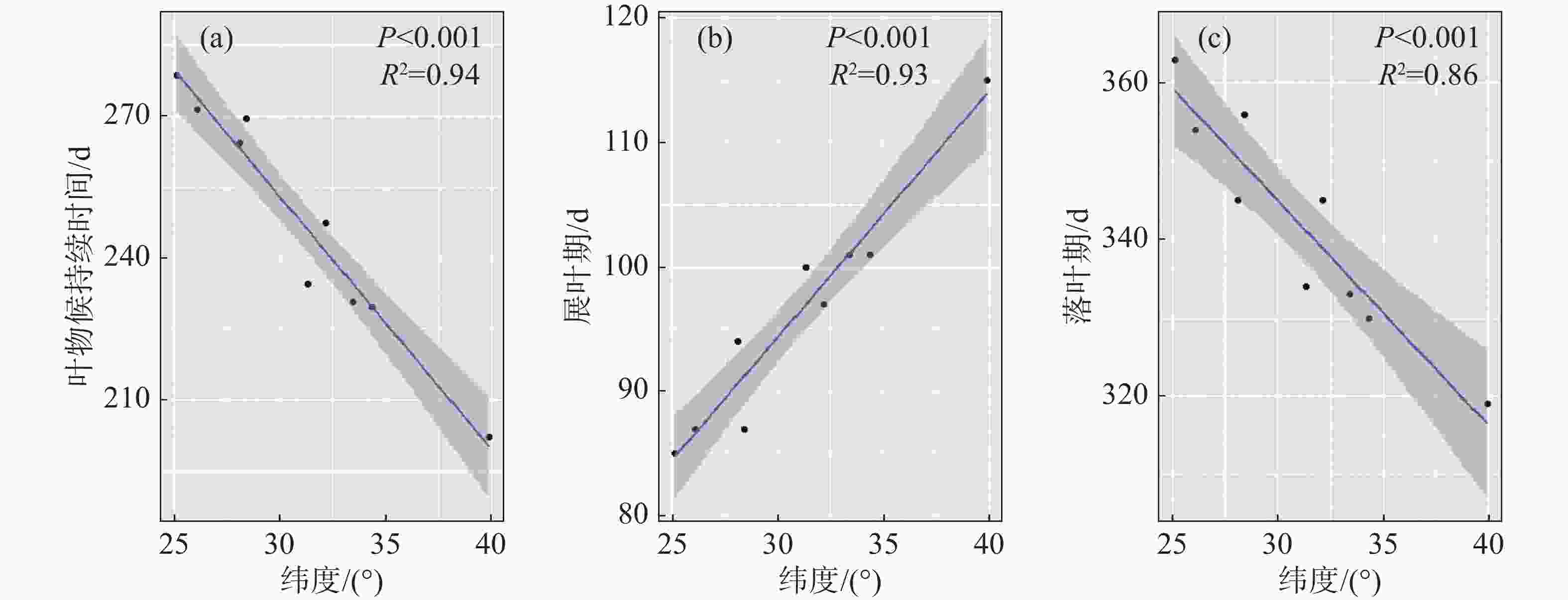
图 1 构树叶物候纬度梯度格局
-
如表3所示,对于纬度梯度上的构树种群叶物候,所选取的6个环境因子都有一定的相关性。造成构树3个叶物候性状纬度差异的环境因子较为一致,其最重要的前3个环境因子均是:展叶季前温度、落叶季前降水量和冬季温度。其余3个环境变量也对构树叶物候性状有影响,但相对不重要。同时,同一环境因子与不同叶物候特征的重要程度大小有一定差异,展叶季前温度是持续时间和展叶期最重要的环境因子(MSE值分别为:11.01、10.30),而落叶季前降水是落叶期最重要的环境因子(MSE值:9.51)。
表 3 随机森林回归环境因子对构树叶物候持续时间、展叶期和落叶期的重要程度分析结果
叶持续时间 展叶期 落叶期 展叶季前温度/℃ 11.01 10.30 8.29 冬季温度/℃ 8.63 8.89 7.61 落叶季前温度/℃ 4.95 4.43 4.22 冬季降水量/mm 5.44 3.20 5.25 展叶季前降水量/mm 4.54 4.87 4.97 落叶季前降水量/mm 10.75 9.51 9.51 注:数值为每一环境变量的MSE值,该值代表了当此变量被剔除时原模型对植物叶物候预测能力的下降程度,因此MSE值越大变量越重要。 -
本研究结果表明,构树叶物候性状沿纬度梯度有显著的变化规律;展叶期与纬度呈正相关,高纬度地区种群明显有更晚展叶的趋势;落叶期与纬度呈负相关,高纬度地区种群有明显更早落叶的趋势;叶物候的持续时间与纬度呈负相关关系,高纬度地区种群有着更短的叶持续时间。因此来自单个位置种群的叶物候数据不能用于预测其他地方同一物种的叶物候变化[33]。木本植物的物候通常被认为是适应当地气候的,许多使用起源测试的研究强调了种群之间的物候差异[34]。对于同一树种不同纬度的种群来说,预计植物物候的可塑性将有利于种群的持续存在,因为物候事件的时间决定了竞争(由较长的生长季节促进)和避免霜冻损害(由生长期较短)[35]。但是,以前的相关纬度格局研究大多单独关注于展叶期或叶变色期,对叶物候持续时间的空间变异研究尚属首次[27, 36]。
树木物候的种内变异性可能会严重限制研究人员预测物候对全球气候变化的一般反应的能力[36]。本研究结果表明了秋季落叶相比春季展叶对低纬度叶物候持续时间延长有更大的贡献,从而使低纬度地区种群获得更长的年际生长时间和更多的生物量增长。之前的研究也发现了展叶与落叶期对变暖响应的不一致,并认为落叶期主导植物的叶物候持续时间:对于平均叶持续时间延长的物种,落叶期延迟,而那些植被周期平均缩短的物种的落叶日期提前[37]。但是,并没有发现不同地理位置同一种群的秋季物候及叶持续时间差异。研究结果表明,同一物种的高纬度地区种群相较于低纬度地区种群具有较高的光饱和CO2同化率和较低的比叶面积[38-39],推测这是对较短生长季[40]的适应。此种情况下,尽管两者相关,但叶物候持续时间的差异所导致潜在生产力和碳固存能力的差异小于预期差异[41-42]。
-
在对环境因子的分析中,展叶季前的温度是展叶期和叶物候持续时间最关键的环境因子。尽管大多数植物控制长叶的确切生理机制还不清楚,但可以明确的是,大多数物种的叶子发育对温度极其敏感。不同纬度种群长叶时间的转变是由于温度在植物生长发育中所起的主导作用,此结果验证了植物物候对气候变化高度敏感这一猜测,并成为气候变化的绝佳指标[43]。构树叶物候纬度差异可以用气候因子的变化及其与叶物候的相关性来解释,但不同叶物候性状的主要气候驱动因素有所差异。本研究结果中,落叶物候最相关的环境因子是落叶季前的降水量。这种正相关可能与水分胁迫对秋季植物生长的影响有关。有限的水势会抑制植物生长和光合作用活动[44],增加叶绿素退化和植物死亡的风险,并提前叶片衰老的时间[45]。因此,季前期间降水增加可以缓解水分胁迫并延长秋季物候。目前对于植物秋季物候调控机制的研究尚存许多争议,对于给定的纬度,落叶树种的抗寒性发生在生长停止之后和叶片衰老之前,并且生长停止、叶片衰老和年内及年际之间的抗寒性之间也存在很强的相关性[46]。这解释了秋季物候对冬季温度和展叶季前温度敏感的可能原因,也强调了更深入地研究秋叶衰老及其潜在过程作为叶持续时间和全球变化驱动因素的重要性。
不同纬度站点相同气候因子的变化导致当地树木的展叶和落叶物候都发生了变化,而秋季叶物候与主要环境因子的相关程度较小。这与之前的研究结果相一致,即与春季物候相比,秋季物候更多地受到遗传变异的影响[47]。全球温度升高可能会促进春季叶片提前展叶,并在秋季促进生长停止和叶片衰老[48-49]。已有研究结果表明,过去几十年的气温升高导致树木的春季物候发生了变化,而秋季叶物候的变化较小且不太一致[50-51]。因此笔者推断,构树可能通过适应性实现更早的叶片展叶和更长的叶物候持续时间。然而,后期生长停止和落叶期的发展可能将涉及种群差异和自然选择。未来的几十年里,世界上大部分温带地区的植物预计将继续提前落叶[52-53]。在较长一段时间内,一些树种可能会在局部范围内被消灭,并根据气候变化改变它们的活动范围[53]。因此对构树不同种群叶物候持续时间的环境调控机制和空间分布变化分析将有助于评估植物物种栖息地的变化[54],可能对生态系统动态产生重大影响,如碳和水的循环及植物-动物相互作用[55-56],以及对依赖树种的行业,如苹果、桃子和构树生产可能的经济影响。
-
本研究发现了构树种群叶物候持续时间沿纬度增加逐渐缩短的地理格局,并且发现这一格局是由低纬度种群展叶期提前和落叶期延迟共同导致。此外,本研究还发现构树展叶期和叶物候持续时间的主要调控因子是展叶季前温度。相比展叶期,构树的落叶期受季前降水的影响程度大于季前温度,这为进一步理解构树秋季物候的调控机制提供了参考。但是要清楚理解这一机制及不同纬度种群物候差异原因还需要其他相关工作,如系统发育差异的研究以及基因调控机制的研究等。
构树叶物候持续时间的纬度格局
DOI: 10.15886/j.cnki.rdswxb.2023.04.008
 CSTR: 32425.14.j.cnki.rdswxb.2023.04.008
CSTR: 32425.14.j.cnki.rdswxb.2023.04.008
Latitudinal patterns of leaf phenology duration in Broussonetia papyrifera
-
摘要: 利用中国物候观测网9个站点记录的构树(Broussonetia papyrifera)叶物候近半个世纪的数据,使用线性回归模型分析构树叶物候持续时间的纬度差异。结果显示,构树叶物候持续时间随纬度增加有明显逐渐缩短的趋势。具体表现为:随着纬度增加,展叶期逐渐提前,落叶期逐渐延迟。对环境因子的重要性进行随机森林回归分析,结果表明,展叶季前的温度是控制构树展叶期和叶持续时间最关键的环境因子,而落叶季前的降水量是落叶期最关键的环境因子,这表明叶物候持续时间的纬度格局是由温度和降水量共同塑造的。结果表明,广布种植物物候存在明显的纬度梯度, 以此适应不同的当地环境条件, 提高了自身适应性。从地理格局视角,评估纬度对同一物种不同种群叶物候的影响, 有助于评估物种未来的分布范围变化及灭绝风险。Abstract: In the Northern Hemisphere, leaf phenology is often reported to have a high spatial variability due to environmental differences at different latitudes. The duration of plant leaf phenology has an important impact not only on the carbon sequestration capacity of plants, but also on their reproductive success and ability to adapt to climate change. However, previous studies on population differences in leaf phenology often were only focused on the differences in leaf unfolding or leaf discoloration stages among different populations, and less attention was paid to the duration of leaf phenology and its driving mechanism. In this study, linear regression models were established by using the phenological data of the leaves of paper mulberry (Broussonetia papyrifera). recorded by China Phenology Observation Network to analyze the latitudinal differences in the duration of phenology of the leaves. The analysis showed that with the increase of latitude, the leaf-unfolding stage for paper mulberry gradually advances and the leaf-falling period gradually delays, indicating that the leaf phenology duration for paper mulberry tended to be shortened with the increase in latitude. The analysis of the importance of environmental factors by using random forest regression showed that the temperature before the leaf-unfolding stage was the most critical environmental factor controlling the leaf-unfolding stage and leaf duration of paper mulberry, and that the precipitation prior to leaf fall was the most critical environmental factor at the leaf fall season, indicating that the latitudinal patterns in leaf phenology duration are jointly shaped by temperature and precipitation. All these results show that the phenology of widely distributed plants has obvious latitudinal gradients in different populations, so as to adapt to different local environmental conditions and improve their own adaptability. Studying the phenological differentiation of different populations of the same species from the perspective of latitudinal differences is helpful to assess the changes in the distribution range and extinction risk of species in the future.
-
Key words:
- paper mulberry /
- leaf phenology /
- duration /
- latitudinal pattern /
- intraspecific variation
-
表 1 9个站点构树叶物候数据和纬度坐标数据
持续
时间/d持续时间
标准差展叶
期/d落叶
期/d纬度 北京 202.38 10.64 113 317 39°90′ 西安 230.83 10.25 99 331 33°42′ 洛阳 229.69 8.26 99 328 34°35′ 芜湖 234.46 9.89 98 332 31°32′ 镇江 247.45 7.06 95 343 32°16′ 长沙 264.43 11.30 92 343 28°12′ 南昌 269.57 10.57 85 354 28°40′ 贵阳 271.43 13.95 85 352 26°11′ 桂林 278.50 23.36 83 361 25°14′  下载: 导出CSV
下载: 导出CSV
表 3 随机森林回归环境因子对构树叶物候持续时间、展叶期和落叶期的重要程度分析结果
叶持续时间 展叶期 落叶期 展叶季前温度/℃ 11.01 10.30 8.29 冬季温度/℃ 8.63 8.89 7.61 落叶季前温度/℃ 4.95 4.43 4.22 冬季降水量/mm 5.44 3.20 5.25 展叶季前降水量/mm 4.54 4.87 4.97 落叶季前降水量/mm 10.75 9.51 9.51 注:数值为每一环境变量的MSE值,该值代表了当此变量被剔除时原模型对植物叶物候预测能力的下降程度,因此MSE值越大变量越重要。
下载: 导出CSV
-
[1] LIETH H. Purposes of a phenology book [M]. Berlin Heidelberg: Phenology and Seasonality Modeling. Springer. 1974: 3 − 19. [2] RICHARDSON A D, KEENAN T F, MIGLIAVACCA M, et al. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system [J]. Agricultural and Forest Meteorology, 2013, 169: 156 − 173. doi: 10.1016/j.agrformet.2012.09.012 [3] 杨乐. 浙江天童常绿阔叶林植物春季叶物候研究[D]. 上海: 华东师范大学, 2009. [4] 赵永宏, 赵维俊, 马剑. 祁连山青海云杉物候对水热驱动的响应[J]. 生态学报, 2020, 40(24): 9083 − 9093. [5] RATHCKE B, LACEY E P. Phenological patterns of terrestrial plants [J]. Annual Review of Ecology and Systematics, 1985: 179 − 214. [6] FU Y H, PIAO S, ZHAO H, et al. Unexpected role of winter precipitation in determining heat requirement for spring vegetation green‐up at northern middle and high latitudes [J]. Global Change Biology, 2014, 20(12): 3743 − 3455. doi: 10.1111/gcb.12610 [7] 景军. 亚热带常绿阔叶林食叶昆虫和寄主植物物候同步性及其对气候变化的响应[D]. 上海: 华东师范大学, 2015. [8] MYNENI R B, KEELING C, TUCKER C J, et al. Increased plant growth in the northern high latitudes from 1981 to 1991 [J]. Nature, 1997, 386(6626): 698 − 702. doi: 10.1038/386698a0 [9] PARMESAN C, YOHE G. A globally coherent fingerprint of climate change impacts across natural systems [J]. Nature, 2003, 421(6918): 37 − 42. doi: 10.1038/nature01286 [10] SITCH S, SMITH B, PRENTICE I C, et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model [J]. Global Change Biology, 2003, 9(2): 161 − 185. doi: 10.1046/j.1365-2486.2003.00569.x [11] 张静, 孙路. 浅谈植物功能性状对气候变化的响应[J]. 南方农业, 2019, 13(14): 150 − 151. doi: 10.19415/j.cnki.1673-890x.2019.14.077 [12] BOLNICK D I, AMARASEKARE P, ARAÚJO M S, et al. Why intraspecific trait variation matters in community ecology [J]. Trends in Ecology & Evolution, 2011, 26(4): 183 − 192. [13] FAJARDO A, PIPER F I. Intraspecific trait variation and covariation in a widespread tree species (Nothofagus pumilio) in southern Chile [J]. New Phytologist, 2011, 189(1): 259 − 271. doi: 10.1111/j.1469-8137.2010.03468.x [14] FU Y H, PIAO S, DELPIERRE N, et al. Larger temperature response of autumn leaf senescence than spring leaf‐out phenology [J]. Global Change Biology, 2018, 24(5): 2159 − 2168. doi: 10.1111/gcb.14021 [15] MARON J L, ELMENDORF S C, VILÀ M. Contrasting plant physiological adaptation to climate in the native and introduced range of Hypericum perforatum [J]. Evolution, 2007, 61(8): 1912 − 1924. doi: 10.1111/j.1558-5646.2007.00153.x [16] LIAO S, DENG Z, CUI K, et al. Genetic diversity of Broussonetia papyrifera populations in southwest China [J]. Genetics and Molecular Research, 2014, 13(3): 7553 − 7563. doi: 10.4238/2014.September.12.22 [17] 杨平, 朱学慧, 姬慧娟, 等, 温度和水分胁迫对构树种子萌发的影响[J]. 四川林业科技, 2015, 36(6): 90−92. [18] KIKUZAWA K. Leaf phenology as an optimal strategy for carbon gain in plants [J]. Canadian Journal of Botany, 1995, 73(2): 158 − 163. doi: 10.1139/b95-019 [19] 崔思明, 郝亚涵, 周玮, 等. 构树不同种源叶性状变异研究[J]. 中南林业科技大学学报, 2020, 40(5): 104 − 110. [20] CHUINE I, BELMONTE J, MIGNOT A. A modelling analysis of the genetic variation of phenology between tree populations [J]. Journal of Ecology, 2000: 561 − 570. [21] DAI J, WANG H, GE Q. The spatial pattern of leaf phenology and its response to climate change in China [J]. International journal of biometeorology, 2014, 58(4): 521 − 528. doi: 10.1007/s00484-013-0679-2 [22] 于裴洋, 同小娟, 李俊, 等, 中国暖温带木本植物物候模拟分析[J]. 北京林业大学学报, 2021, 43(11): 28 − 39. [23] 陈沁, 唐欣然, 薛乾怀, 等. 亚热带植物春季和秋季物候格局及其对气候变化的响应[J]. 广西植物, 42, 7: 1105 − 1115. [24] 崔雪娜, 杜彦君, 赵袁, 等. 南亚热带阔叶林展叶物候及其种内种间差异探讨[J]. 广西植物, 2017, 37(3): 322 − 328. [25] 陶泽兴, 葛全胜, 徐韵佳, 等. 西安和宝鸡木本植物花期物候变化及温度敏感度对比[J]. 生态学报, 2020, 40(11): 123 − 133. [26] 于裴洋, 同小娟, 李俊, 等, 中国东部暖温带刺槐物候模型比较[J]. 中国农业气象, 2020, 41(10): 609−621. [27] VICO G, KARACIC A, ADLER A, et al. Consistent poplar clone ranking based on leaf phenology and temperature along a latitudinal and climatic gradient in Northern Europe [J]. BioEnergy Research, 2021, 14(2): 445 − 459. doi: 10.1007/s12155-021-10249-5 [28] FU Y H, ZHAO H, PIAO S, et al. Declining global warming effects on the phenology of spring leaf unfolding [J]. Nature, 2015, 526(7571): 104 − 107. doi: 10.1038/nature15402 [29] LIU Q, FU Y H, ZENG Z, et al. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China [J]. Global Change Biology, 2016, 22(2): 644 − 655. doi: 10.1111/gcb.13081 [30] BREIMAN L. Random forests [J]. Machine learning, 2001, 45(1): 5 − 32. doi: 10.1023/A:1010933404324 [31] YUE J, YANG G, FENG H. Comparative of remote sensing estimation models of winter wheat biomass based on random forest algorithm [J]. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(18): 175 − 182. [32] NAM H-K, CHOI S-H, YOO J-C. Patterning waterbird assemblages on rice fields using self-organizing map and random forest [J]. Korean Journal of Environmental Agriculture, 2015, 34(3): 168 − 177. doi: 10.5338/KJEA.2015.34.3.26 [33] SCHABER J, BADECK F-W. Physiology-based phenology models for forest tree species in Germany [J]. International Journal of Biometeorology, 2003, 47(4): 193 − 201. doi: 10.1007/s00484-003-0171-5 [34] VON WUEHLISCH G, KRUSCHE D, MUHS H. Variation in temperature sum requirement for Flushing [J]. Silvae Genetica, 1995, 44: 5 − 6. [35] DE KORT H, VANDER MIJNSBRUGGE K, VANDEPITTE K, et al. Evolution, plasticity and evolving plasticity of phenology in the tree species Alnus glutinosa [J]. Journal of Evolutionary Biology, 2016, 29(2): 253 − 264. doi: 10.1111/jeb.12777 [36] VITASSE Y, DELZON S, DUFRÊNE E, et al. Leaf phenology sensitivity to temperature in European trees: Do within-species populations exhibit similar responses? [J]. Agricultural and Forest Meteorology, 2009, 149(5): 735 − 744. doi: 10.1016/j.agrformet.2008.10.019 [37] GARONNA I, DE JONG R, DE WIT A J, et al. Strong contribution of autumn phenology to changes in satellite‐derived growing season length estimates across Europe (1982–2011) [J]. Global Change Biology, 2014, 20(11): 3457 − 3470. doi: 10.1111/gcb.12625 [38] KALUTHOTA S, PEARCE D W, EVANS L M, et al. Higher photosynthetic capacity from higher latitude: foliar characteristics and gas exchange of southern, central and northern populations of Populus angustifolia [J]. Tree physiology, 2015, 35(9): 936 − 948. doi: 10.1093/treephys/tpv069 [39] SOOLANAYAKANAHALLY R Y, GUY R D, SILIM S N, et al. Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L. ) [J]. Plant, Cell & Environment, 2009, 32(12): 1821 − 1832. [40] GORNALL J L, GUY R D. Geographic variation in ecophysiological traits of black cottonwood (Populus trichocarpa) [J]. Botany, 2007, 85(12): 1202 − 1213. [41] MCKOWN A D, GUY R D, KLÁPŠTĚ J, et al. Geographical and environmental gradients shape phenotypic trait variation and genetic structure in Populus trichocarpa [J]. New Phytologist, 2014, 201(4): 1263 − 76. doi: 10.1111/nph.12601 [42] POLGAR C A, PRIMACK R B. Leaf‐out phenology of temperate woody plants: from trees to ecosystems [J]. New Phytologist, 2011, 191(4): 926 − 941. doi: 10.1111/j.1469-8137.2011.03803.x [43] TEZARA W, MITCHELL V, DRISCOLL S, et al. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP [J]. Nature, 1999, 401(6756): 914 − 917. doi: 10.1038/44842 [44] 董晓宇. 2000—2017年内蒙古荒漠草原植被物候和净初级生产力对气候变化的响应[D]. 西安: 长安大学, 2020. [45] REPO T, ZHANG G, RYYPPö A, et al. The relation between growth cessation and frost hardening in Scots pines of different origins [J]. Trees, 2000, 14(8): 456 − 464. doi: 10.1007/s004680000059 [46] FRIEDMAN J M, ROELLE J E, CADE B S. Genetic and environmental influences on leaf phenology and cold hardiness of native and introduced riparian trees [J]. International Journal of Biometeorology, 2011, 55(6): 775 − 787. doi: 10.1007/s00484-011-0494-6 [47] LINKOSALO T, LECHOWICZ M J. Twilight far-red treatment advances leaf bud burst of silver birch (Betula pendula) [J]. Tree physiology, 2006, 26(10): 1249 − 1256. doi: 10.1093/treephys/26.10.1249 [48] SAXE H, CANNELL M G, JOHNSEN ø, et al. Tree and forest functioning in response to global warming [J]. New phytologist, 2001, 149(3): 369 − 399. doi: 10.1046/j.1469-8137.2001.00057.x [49] CHMIELEWSKI F-M, RöTZER T. Response of tree phenology to climate change across Europe [J]. Agricultural and Forest Meteorology, 2001, 108(2): 101 − 112. doi: 10.1016/S0168-1923(01)00233-7 [50] MENZEL A, SPARKS T H, ESTRELLA N, et al. European phenological response to climate change matches the warming pattern [J]. Global change biology, 2006, 12(10): 1969 − 1976. doi: 10.1111/j.1365-2486.2006.01193.x [51] LEBOURGEOIS F, PIERRAT J-C, PEREZ V, et al. Simulating phenological shifts in French temperate forests under two climatic change scenarios and four driving global circulation models [J]. International journal of biometeorology, 2010, 54(5): 563 − 581. doi: 10.1007/s00484-010-0305-5 [52] MORIN X, LECHOWICZ M J, AUGSPURGER C, et al. Leaf phenology in 22 North American tree species during the 21st century [J]. Global Change Biology, 2009, 15(4): 961 − 975. doi: 10.1111/j.1365-2486.2008.01735.x [53] 回爽. 沈阳城-郊梯度下三种观赏树种的物候特征及影响因素研究[D]. 沈阳: 沈阳农业大学, 2020. [54] PIAO S, CIAIS P, FRIEDLINGSTEIN P, et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming [J]. Nature, 2008, 451(7174): 49 − 52. doi: 10.1038/nature06444 [55] WHITE A, CANNELL M G, FRIEND A D. Climate change impacts on ecosystems and the terrestrial carbon sink: a new assessment [J]. Global environmental change, 1999, 9: 21 − 30. doi: 10.1016/S0959-3780(99)00016-3 [56] CHMIELEWSKI F-M, MÜLLER A, BRUNS E. Climate changes and trends in phenology of fruit trees and field crops in Germany, 1961–2000 [J]. Agricultural and Forest Meteorology, 2004, 121(1/2): 69 − 78. -

 点击查看大图
点击查看大图
图(1) / 表 (3)
计量
- 文章访问数: 1856
- HTML全文浏览量: 471
- PDF下载量: 26
- 被引次数: 0
