


 下载:
下载:
-
后水湾网箱鱼类养殖区位于海南岛西北部,儋州市和临高县境内,以深水网箱养殖为主,主要养殖品种为卵形鲳鲹,年养殖产量达到5×104 t,为我国最大的深水网箱养殖基地[1]。然而,由于长期开展网箱养殖以及养殖规模不断扩大,养殖区的环境污染问题越来越突出。浮游细菌作为水体中各种有机物质的分解者和转化者,又是物质和能量的贮存者,在水体物质循环和能量流动中具有重要的作用[2-4]。同时,有些细菌,如弧菌、链球菌还是近海养殖的条件致病菌[1]。每当台风过后,养殖环境条件恶化时,这些有害细菌大量增殖,导致海水养殖鱼类发病[5]。已有研究结果[6]表明,养殖区微生物分布与鱼类疾病发生紧密相关。浙江舟山网箱养殖区网箱内细菌数量高于网箱外细菌数量,且细菌丰度的高峰期也是鱼类发病最严重的时期[7]。大亚湾网箱养殖水体中网箱内的细菌数量高于网箱外和对照区,且弧菌出现的高峰期与弧菌病害的流行期一致[8]。象山港网箱养殖区浮游细菌数量和弧菌数量均高于对照区,而多样性均低于对照区,且某些特定细菌富集较为严重[9-10]。因此,监测网箱养殖区域微生物群落结构和多样性的动态变化,探究浮游细菌群落,特别是潜在病原菌的分布规律,有助于在养殖生产过程中及早采取措施预防网箱养殖鱼类病害的爆发,推动深水网箱养殖产业健康可持续发展。本研究利用高通量测序技术分析临高后水湾深水网箱养殖区浮游细菌群落结构的动态变化,并探讨浮游细菌群落变化与鱼类细菌性病害的关系,研究结果将有助于揭示深水网箱养殖区不同养殖期浮游菌群变化规律,了解潜在病原菌对网箱养殖鱼类的危害,为深水网箱养殖的健康发展及养殖鱼类疾病的有效防控提供理论依据。
-
样品均采自海南临高后水湾深水网箱养殖基地。采样时间分别为2016−03−15、2016−05−16、2016−07−18、2016−09−16、2016−11−14的涨潮期。根据养殖过程,将其分为4个阶段,其中3月份为休养期,5月份为养殖初期,7月份和9月份为养殖中期,11月份为养殖后期。取样点共设置3个站位,分别为22号网箱内、网箱外以及对照区(离网箱养殖区1 km向海一侧的非养殖区),编号分别为03in,03out,05in,05out,05con,07in,07out,07con,09in,09out,09con,11in,11out,11con,每个站位3个重复。将采集到的水样低温运回实验室,充分摇匀后,重复组水样等量混合,先用3 μm无菌滤膜预过滤,再用0.22 μm无菌滤膜过滤1 L等量混合后的水样,将过滤后的滤膜4 ℃保存备用。同时,统计监测月份该网箱每天病死鱼数量。
-
利用OMEGA试剂盒(E.Z.N.ATM Mag-Bind Soil DNA Kit)提取宏基因组DNA。采用NanoDrop微量分光光度计和琼脂糖凝胶电泳检测DNA的纯度和完整性,用Qubit 2.0 DNA 检测试剂盒检测DNA浓度。将符合实验要求的DNA置于−20 ℃保存,备用。
-
DNA样品委托上海生工生物工程有限公司测序,采用16S rDNA V3-V4通用融合引物341F(5′-CCCTACACGACGCTCTTCCGATCTG(barcode)CCTACGGGNGGCWGCAG-3′)和805R(5′-GACTGGAGTTCCTTGGCACCCGAGAATTCCAGACTACHVGGGTATCTAATCC-3′)进行PCR扩增;扩增产物进行琼脂糖电泳检测,采用生工琼脂回收试剂盒(Cat:SK8131)纯化回收;利用Illumina Miseq 测序平台进行高通量测序。
-
所获得的序列分别利用Qiime(version1.7.0)和Mothur软件(version 1.31.2)进行过滤和去除嵌合体序列 [11],得到高质量的基因序列[12]。再运用Mothur软件对序列进行OTU(Operational Taxonomic Units)聚类。相似度在97%以上的OTU代表序列采用RDP classifier贝叶斯算法[13] 进行分类学分析,统计每个样品的群落组成。基于OTU聚类分析结果,对OTU进行Alpha多样性指数等分析、群落结构的统计分析以及unifrac等分析[14-15],最后进行群落结构和系统发育等的统计学和可视化分析。
-
根据16S rRNA基因高通量测定结果,在采集的14份水样中共得到758 641条原始序列,处理后得到686 191条高质量序列,每个样品序列数目为28 032~65 019条不等,序列长度平均值为421 bp。稀释曲线分析(图1-A)显示,随着测序数量的增加,稀释曲线逐渐趋于平缓,表明该测序量已经基本能反映该实验中细菌群落多样性。浮游细菌香农-威纳指数如图1-B所示,平均值变化范围为2.47~4.5,不同养殖期差异较大,总体上表现为养殖后期>休养期>养殖初期和养殖中期。
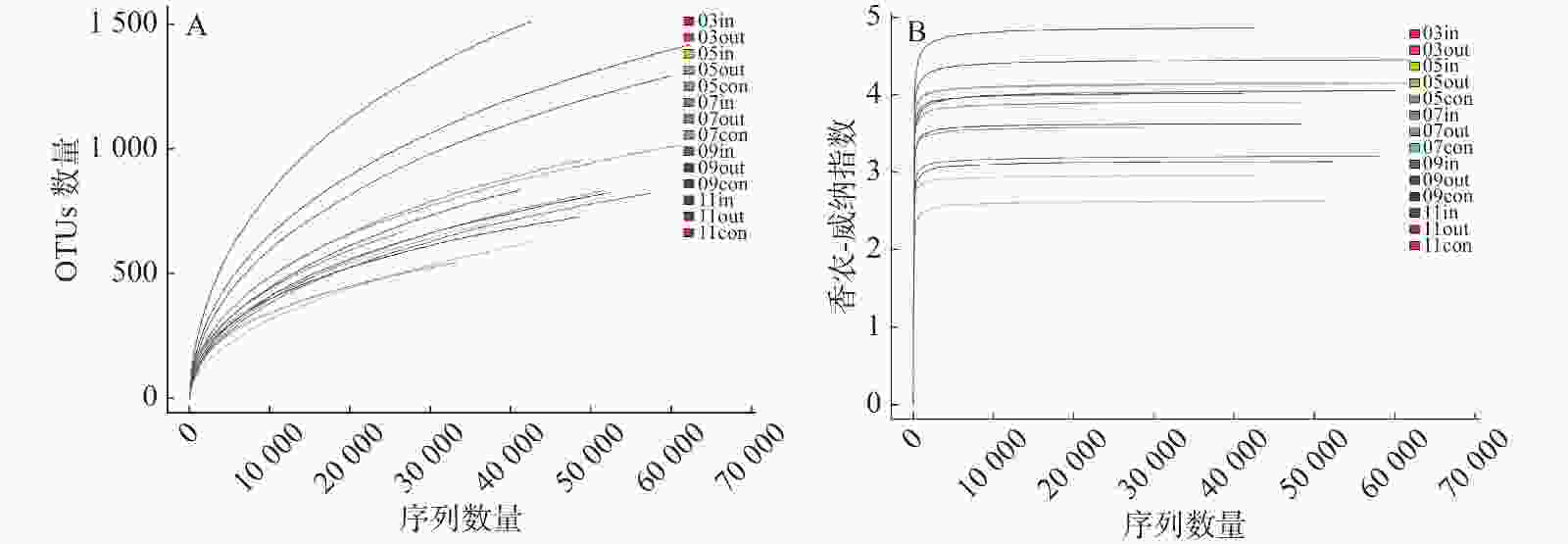
图 1 各样品的浮游细菌16S rRNA基因的α多样性分析
-
主成分分析(PCA)结果显示(图2-A),浮游细菌菌群可分为4组,分别为休养期、养殖初期、养殖中期、养殖后期。这表明养殖区水体中菌落组成与不同养殖期关系较大。基于Unifrac的聚类分析,可以分为3组,休养期、养殖后期、养殖初期和中期(图2-B)。科水平的细菌群落Heatmap分析结果基本一致,相同养殖期能较好地聚在一起(图2-C)。
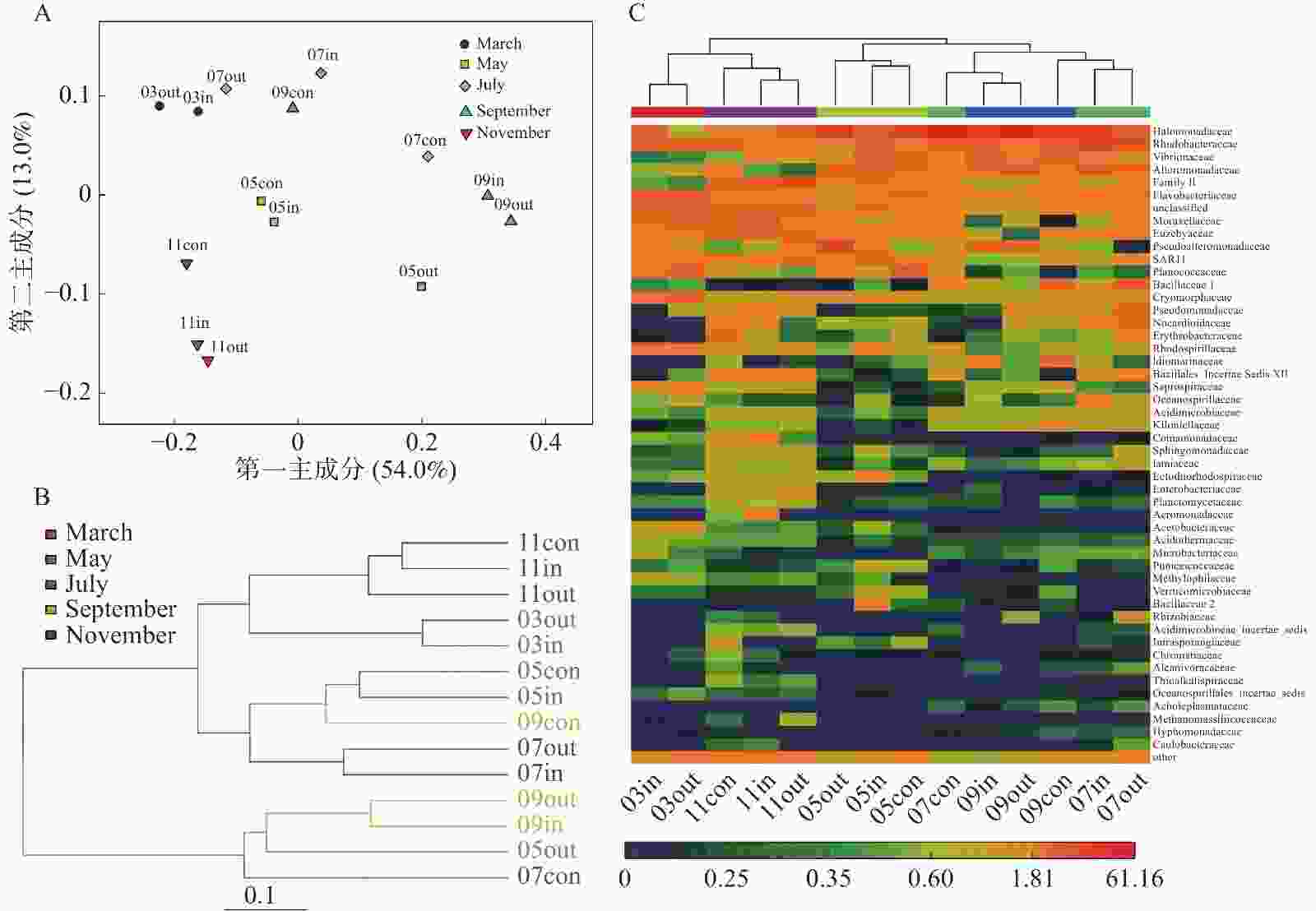
图 2 不同月份网箱内外及对照区样品浮游细菌群落相似性分析
-
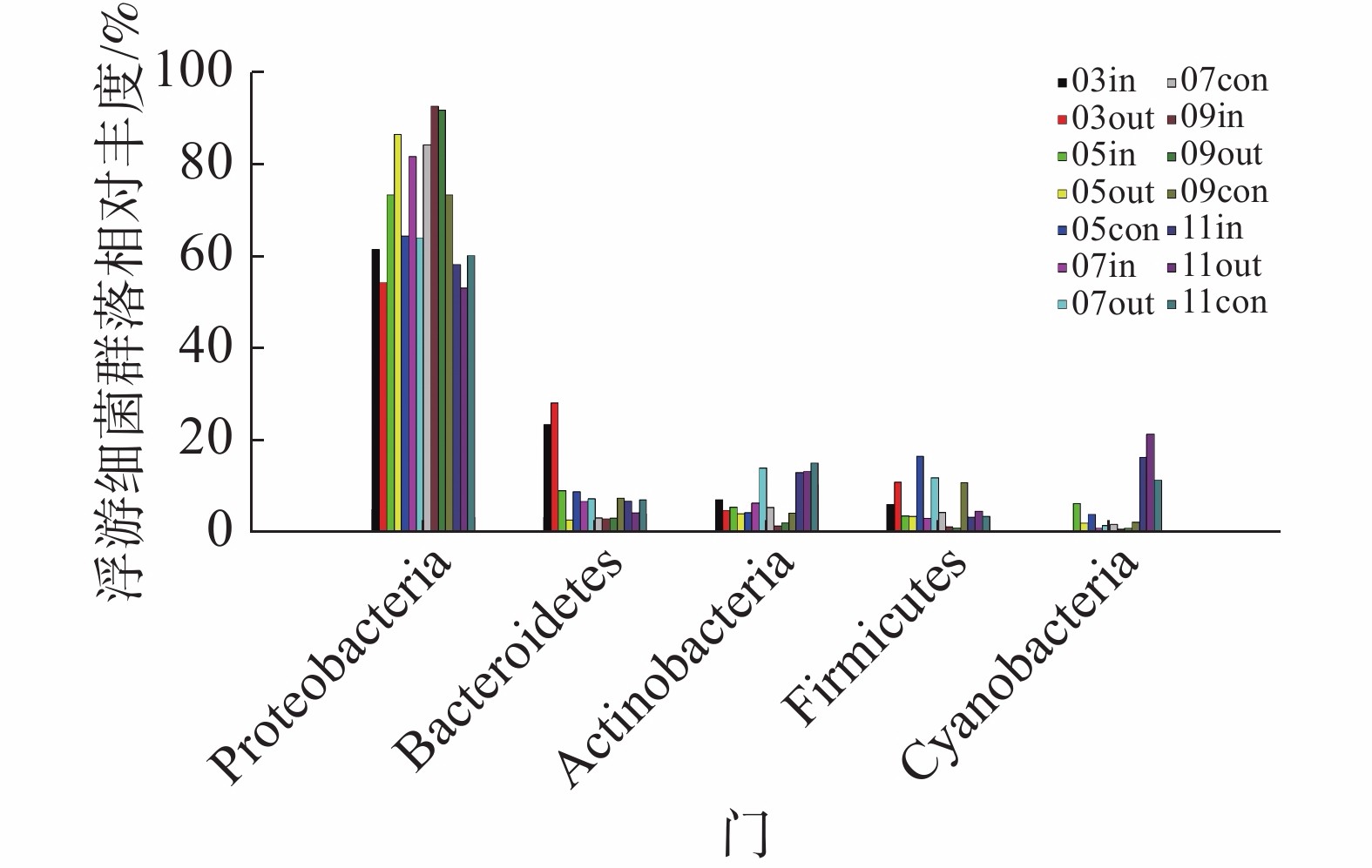
在采集到的14份样品中,共检测到36个门、56个纲、98个目、235个科、807个属的浮游细菌。未能鉴定的序列总数为41 404,占总序列数的6.21%,表明样本中还有大量未知的类群。浮游细菌群落分布结构(图3)相对较为稳定,以变形菌门(Proteobacteria)γ-变形菌纲为主要的优势类群;次优势类群为拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)和蓝细菌门(Cyanobacteria),以上5类菌门相对比例之和变化范围为97.14%~99.63%,其他类菌门所占比例之和不足3%。
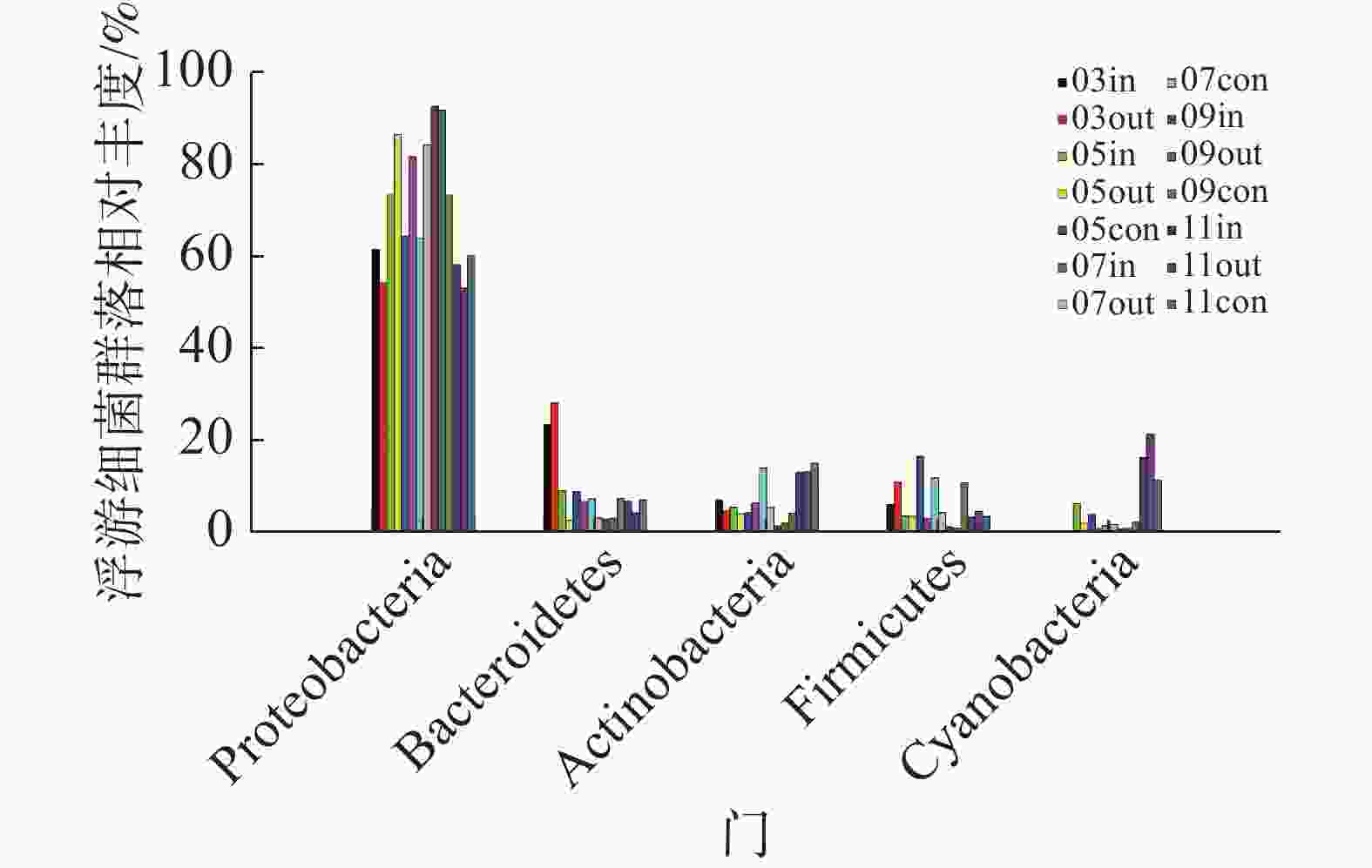
图 3 门水平浮游细菌群落组成
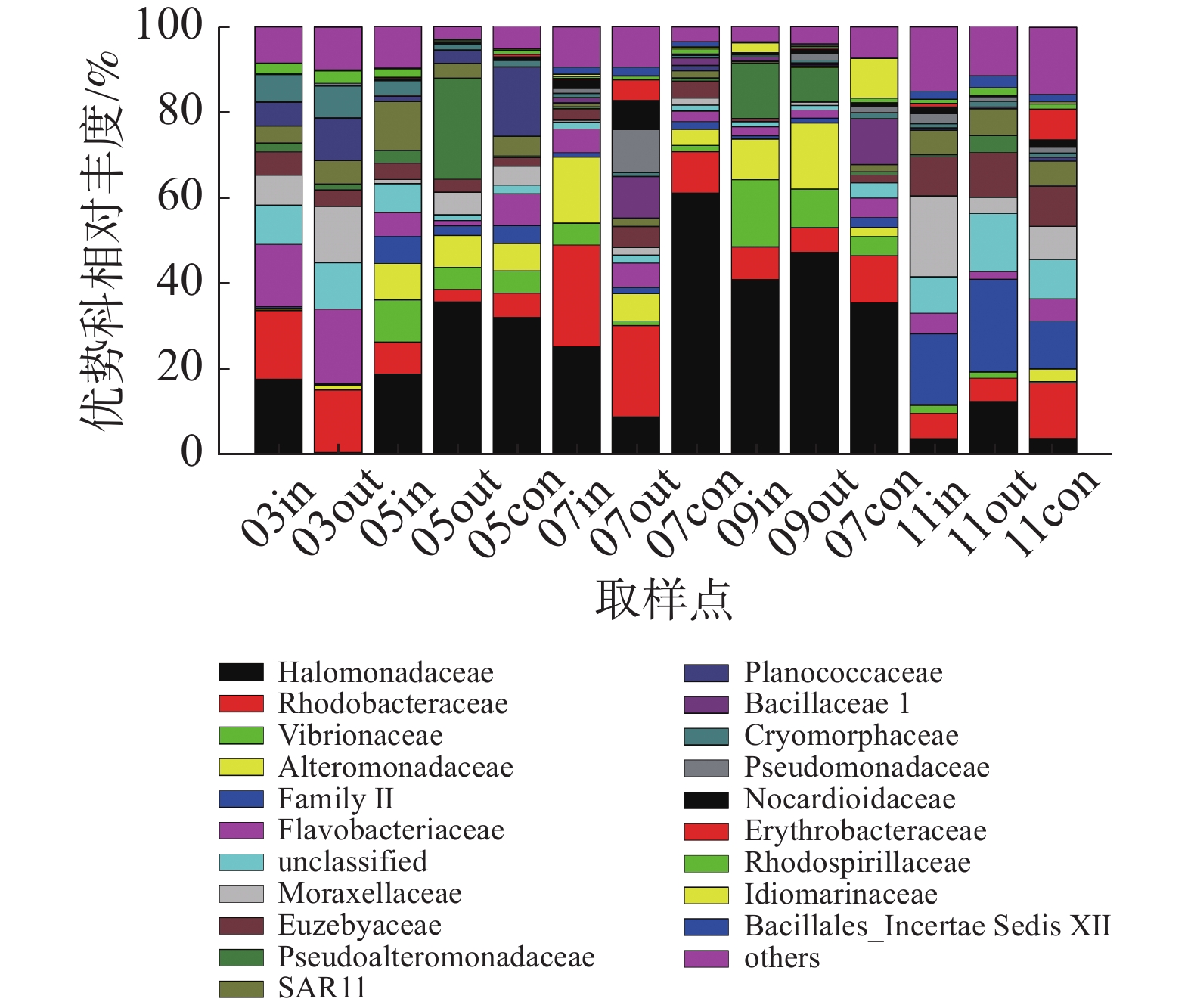
在科的水平上(图4),养殖区以盐单胞菌科(Halomonadaceae)、红杆菌科(Rhodobacteraceae)、弧菌科(Vibrionaceae)、交替单胞菌科(Alteromonadaceae)等为主要的优势类群,其中盐单胞菌科为养殖初期和养殖中期主要优势类群,其相对丰度最高可达61.16%;红杆菌科为休养期的主要优势类群;弧菌科细菌在各监测站位均有发现,以5月份和9月份网箱内丰度最高,相对丰度分别为9.91%和15.85%;蓝细菌门的Family II科为养殖后期的优势类群。
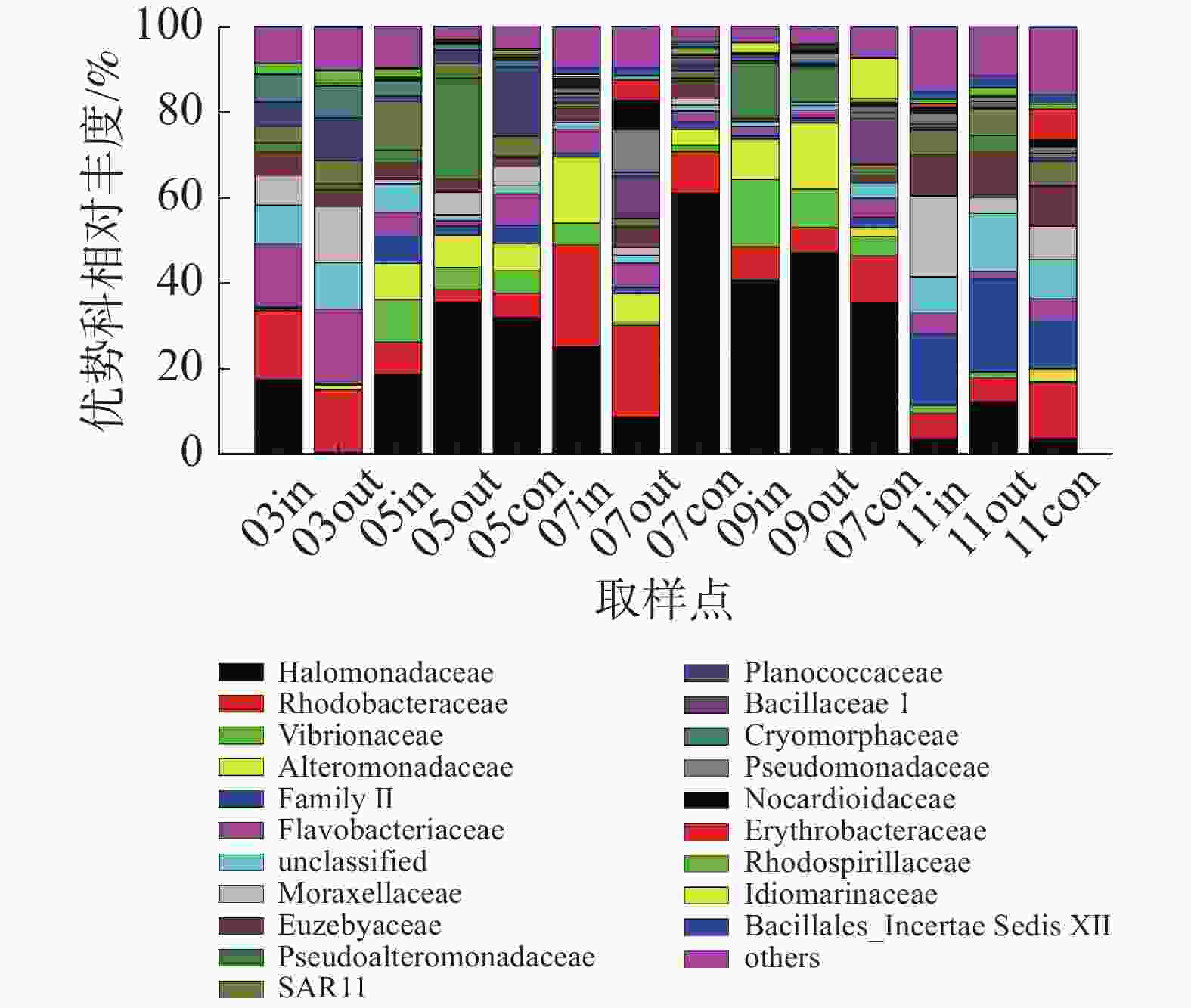
图 4 科水平浮游细菌群落组成
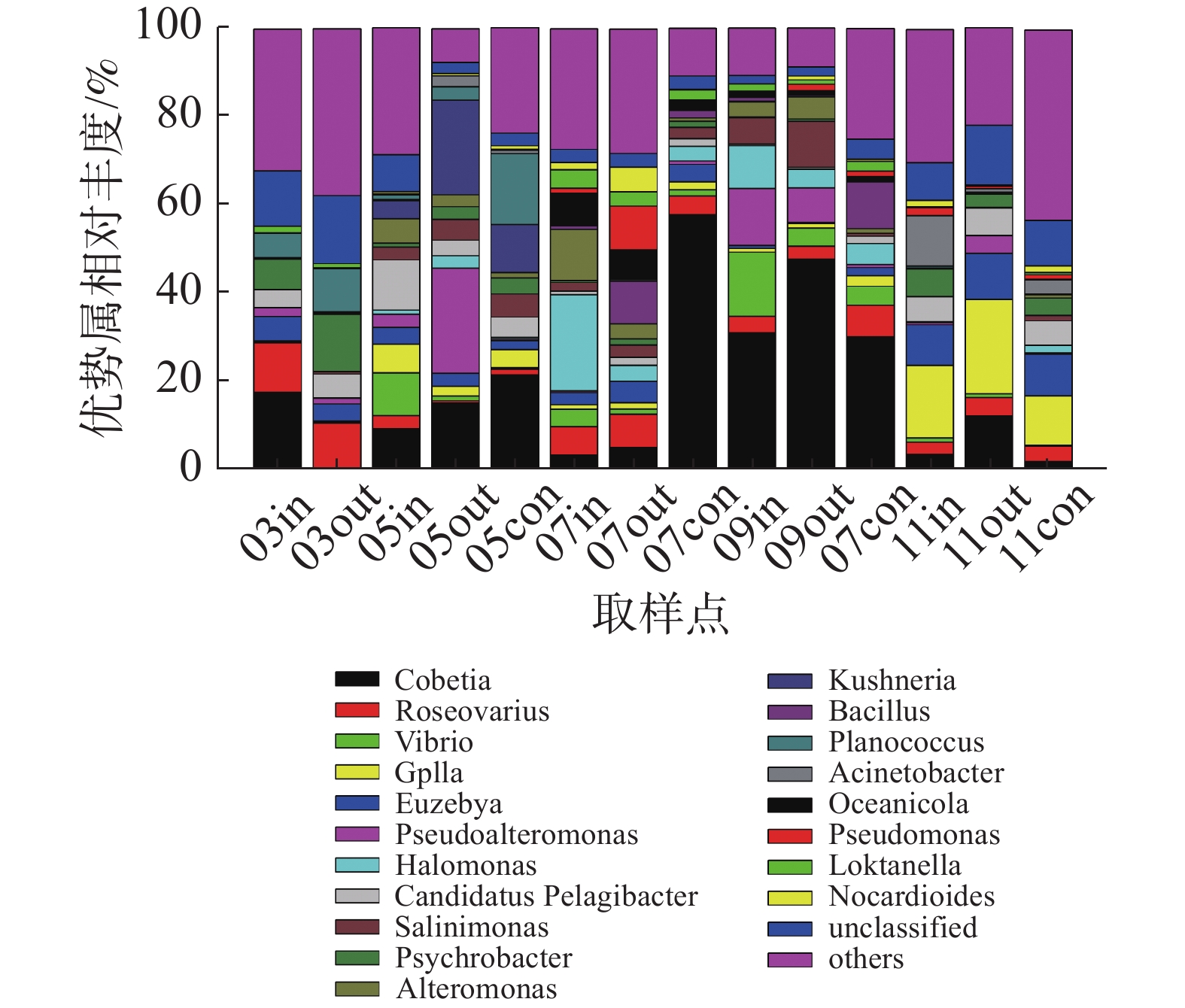
在属水平上(图5),浮游细菌群落主要优势类群包括Cobetia属、弧菌属(Vibrio)、假交替单胞菌属(Pseudoalteromonas)、盐单胞菌属(Halomonas)以及蓝细菌门的GpIIa属等。其中,休养期的优势类群为Cobetia属、玫瑰变色菌属(Roseovarius)和嗜冷杆菌属(Psychrobacter);养殖初期以Cobetia属、假交替单胞菌属、Kushneria属和弧菌属为主,其中网箱内外弧菌相对丰度分别为9.61%和1.06%;养殖中期以Cobetia属、盐单胞菌属、交替单胞菌属(Alteromonas)和弧菌属为主,其中9月份网箱内外弧菌属相对丰度分别为14.52%和4.11%;养殖后期优势类群为GpIIa属和Cobetia属。
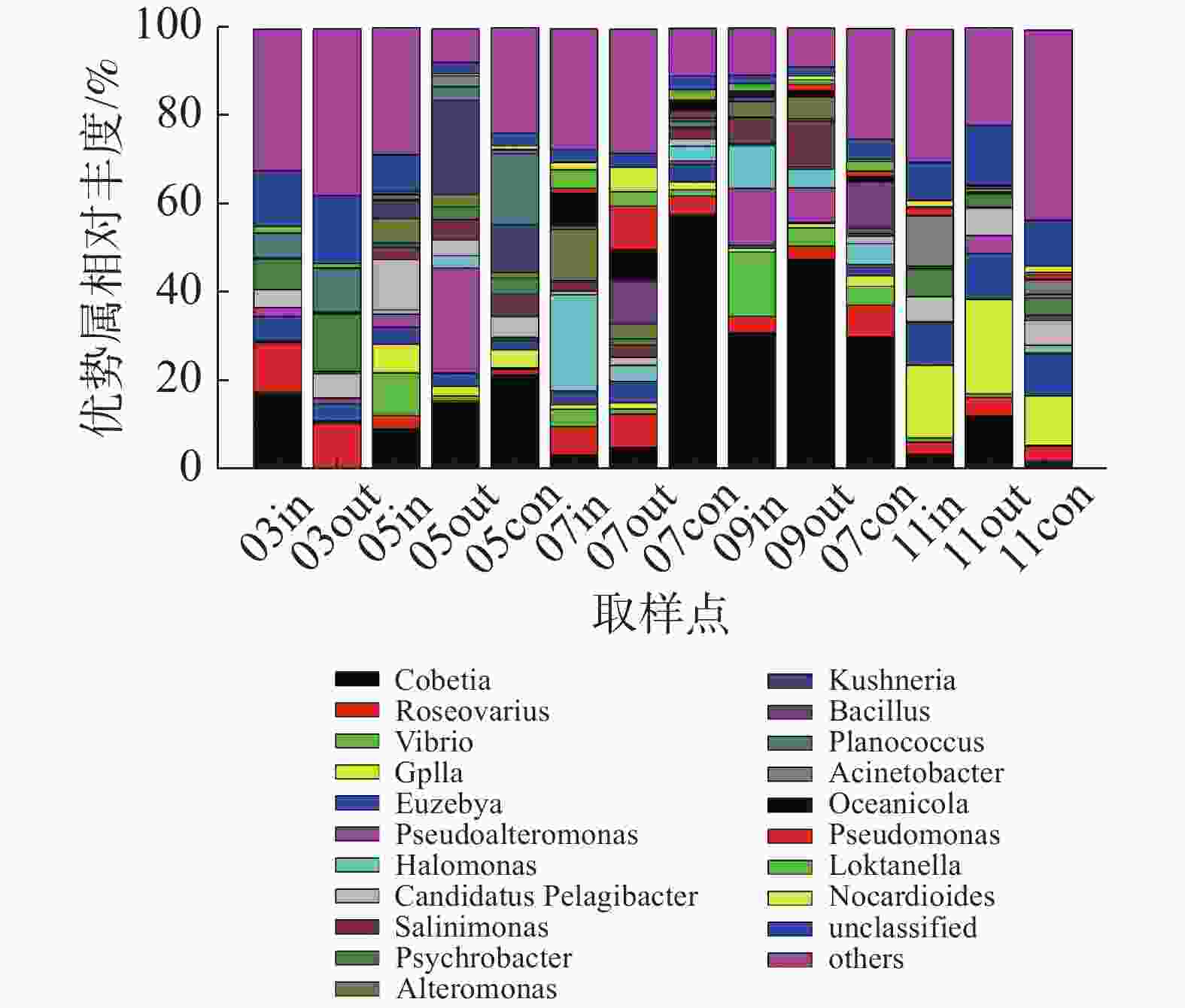
图 5 属水平浮游细菌群落组成
-
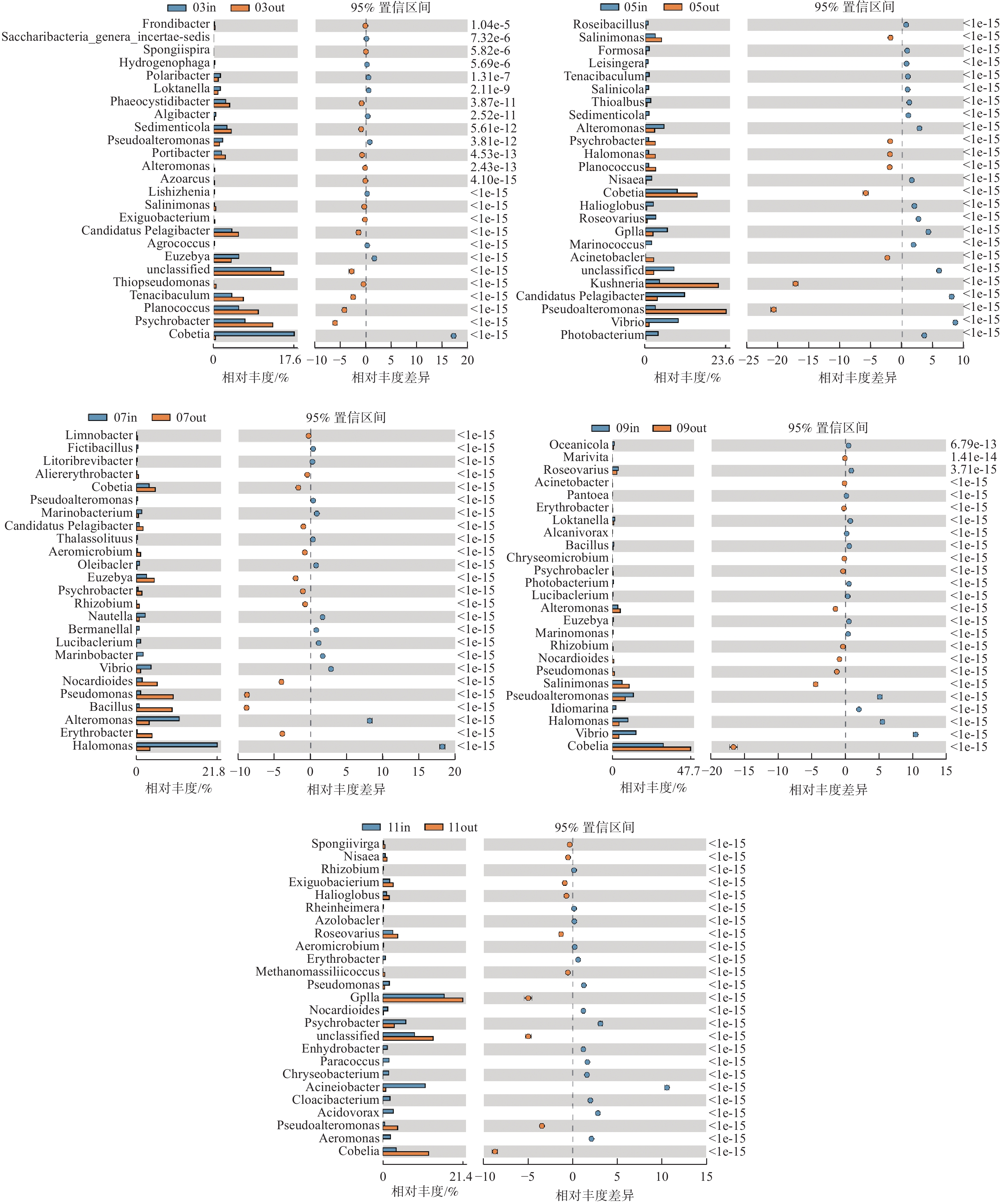
根据监测月份菌落丰度误差线(图6)可知,各养殖阶段差异菌群不同,休养期网箱内外主要差异菌群为Cobetia属、嗜冷杆菌以及黄杆菌属等(图6-A);养殖初期主要差异菌群为Cobetia属、Kushneria属、假交替单胞菌属、弧菌等(图6-B),其中网箱内弧菌、发光杆菌和交替单胞菌相对丰度高于网箱外;养殖中期主要差异菌类群为Cobetia属、芽孢杆菌属、弧菌属等(图6-C、D);养殖后期主要差异菌类群为GpIIa属、Cobetia属等(图6-E)。
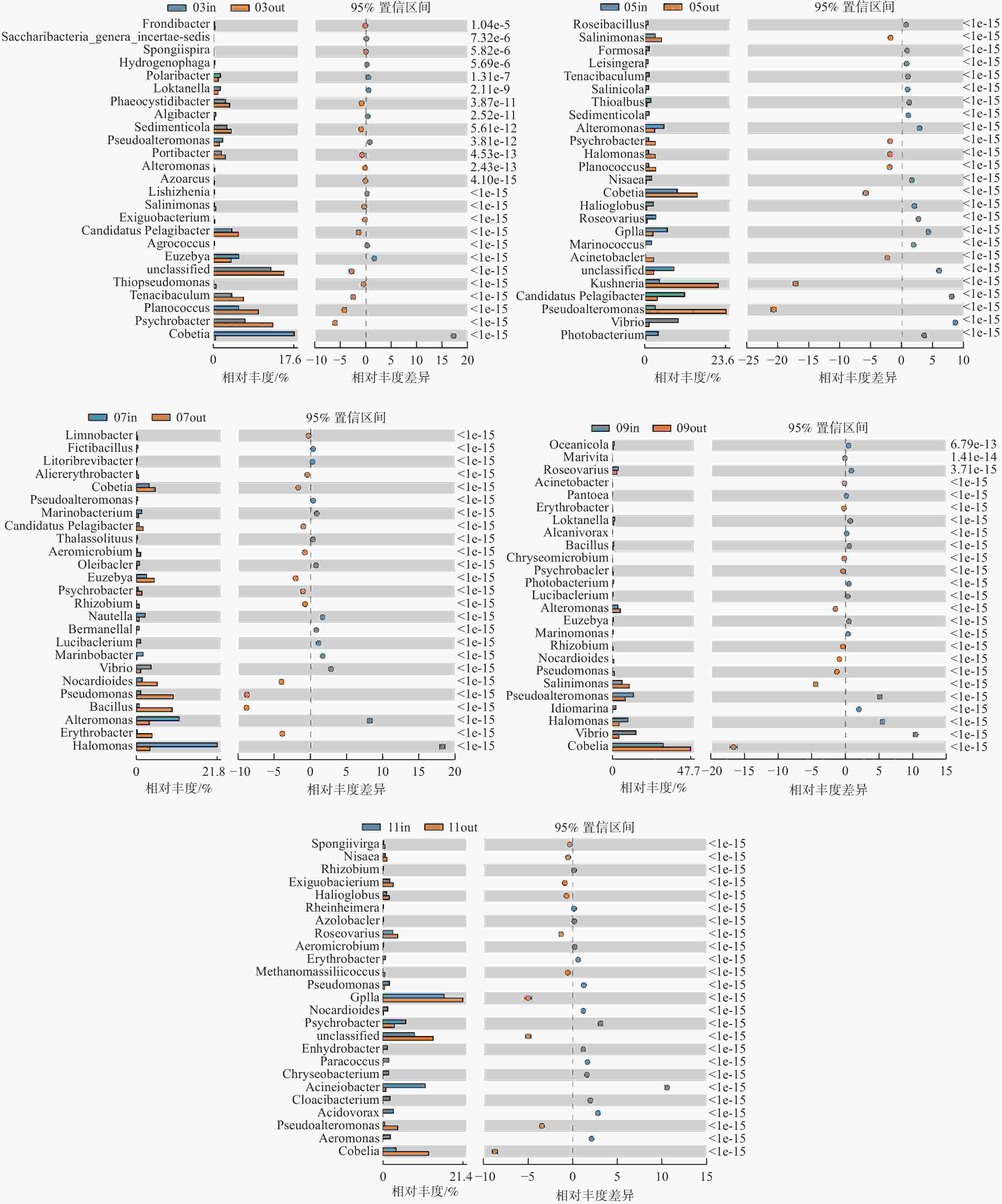
图 6 各监测月份属水平网箱内外浮游细菌菌落丰度误差线图
-
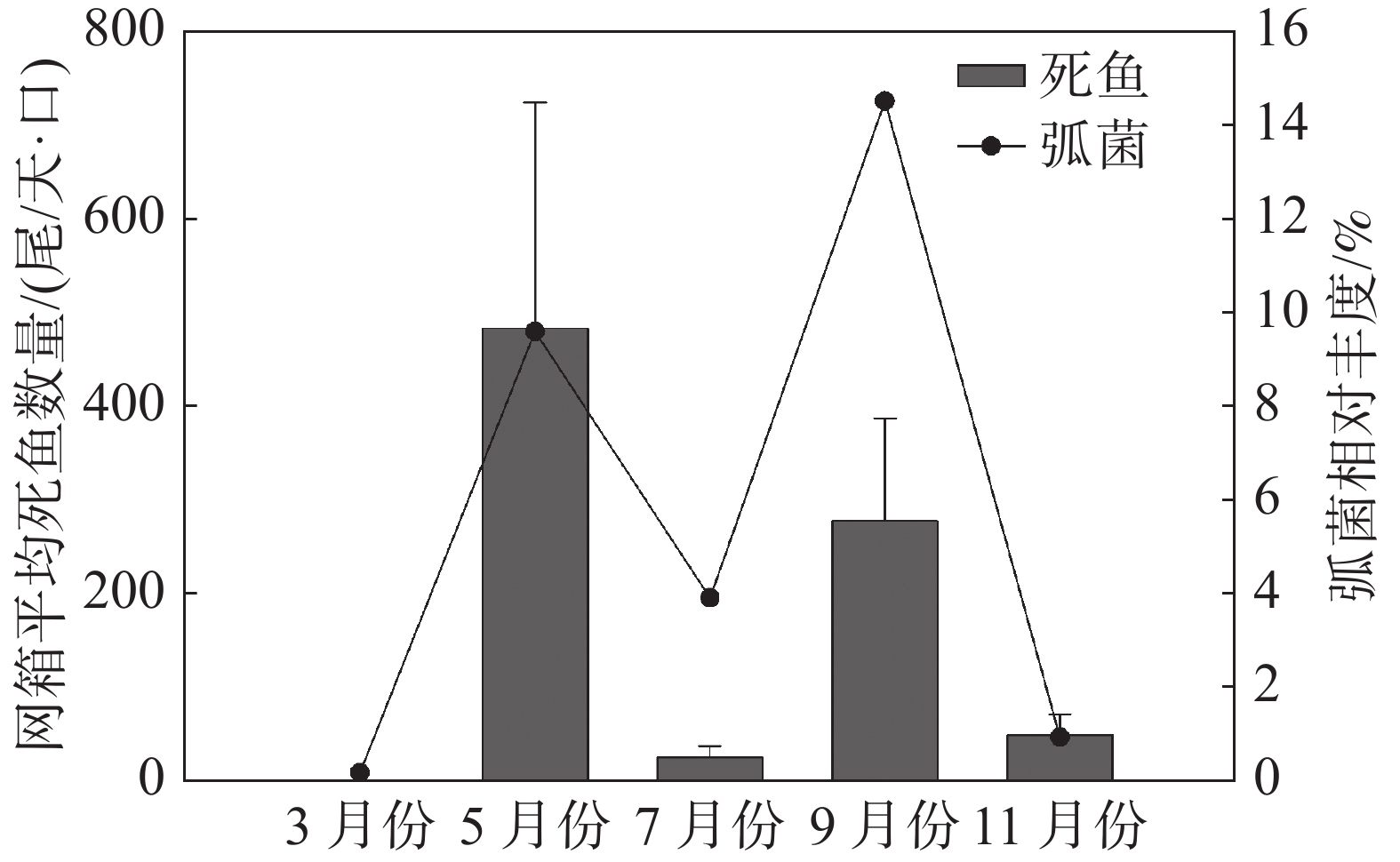
监测月份网箱内弧菌相对丰度与网箱内单日平均死鱼数量关系(图7)表明,在弧菌相对丰度含量较高的5、9月份,每天每口网箱的病死鱼数量分别达到(484±241)尾和(278±110)尾;而弧菌相对丰度较低的7月份和11月份,每天每口网箱病死鱼数量分别为(26±12)尾和(50±22)尾。
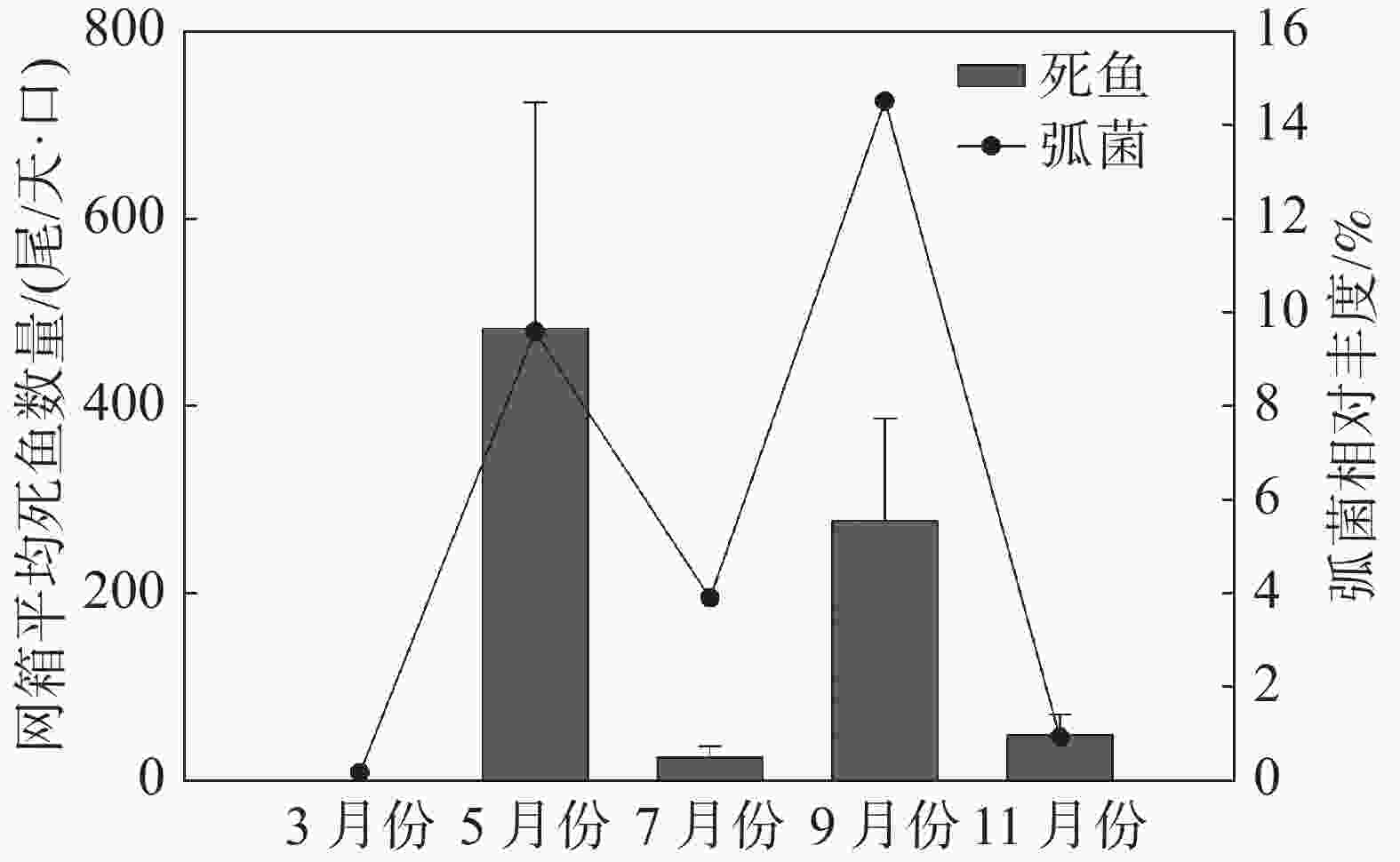
图 7 不同月份弧菌相对丰度与监测区域每日每口网箱平均死鱼数量的关系
由此可见,养殖初期和养殖中期9月份网箱内弧菌含量最高,且均显著高于网箱外弧菌相对丰度,而休养期、养殖中期7月份以及养殖后期网箱内外弧菌属相对丰度较低,表明调查区域水体中弧菌出现的高峰期在养殖初期和养殖中期9月份。根据监测月份网箱内单日平均死鱼数量可知,细菌性疾病引发鱼类死亡的高峰期也为每年5月份和9月份,表明弧菌引起的细菌病害为引起后水湾深水网箱养殖区域鱼类死亡的主要病害类型。后水湾每年从3月底至4月初开始投放卵形鲳鲹鱼苗进行标粗,至5月份鱼苗规格可以达到每尾10~15 cm,周长80 m的网箱放养密度每口高达40万~60万尾。高密度养殖会引起网箱养殖区域残饵及粪便增多,水体中营养盐富集,适宜弧菌等异养菌的生长。因此,5月份是养殖卵形鲳鲹弧菌病害发生的第一个高峰期。7月份,经过标粗的卵形鲳鲹苗被分装到其他网箱,网箱养殖密度随之下降,养殖区水体环境得到有效改善,短期内养殖鱼类病害明显减少。8月中下旬,随着海水平均温度下降到28~30 ℃,网箱养殖鱼类开始感染刺激隐核虫,随后继发性感染细菌,引起鱼类出现烂身、烂尾等病害,严重时鱼类会出现大量死亡。因此,9月份是后水湾网箱养殖鱼类弧菌病害发生的第二个高峰期。每年3月份和11月份海水温度较低,水体中以Cobetia和GpIIa属等细菌为主,弧菌含量较低,网箱养殖鱼类病害也较少。
-
传统的研究方法只能探测到环境中极少部分的微生物群落信息(0.01%~10.0%),难以客观地揭示微生物类群及多样性[16-17]。本研究利用16S rRNA基因高通量测序技术对深水网箱养殖区浮游细菌群落结构进行分析,结果表明,所获得测序样品文库的覆盖率均在99%以上,能够较高程度地反映水体中浮游细菌群落的多样性。调查区域浮游细菌Shannon-Wiener多样性指数平均值为3.65,表明浮游细菌群落比较丰富,多样性水平均较高。究其原因可能是调查期间海水平均温度均高于27 ℃,总体上适宜细菌繁殖与生长;此外,由于监测区域深水网箱养殖区位于离岸2 km,水深为15 m以上水域,水质良好,水体悬浮物浓度为0.5~7 mg·L−1,透光性好,初级生产力强,能为浮游细菌生长提供源源不断的营养物质。不同养殖期差异较大,总体上表现为养殖后期>休养期>养殖初期和养殖中期,这可能是养殖初期和养殖中期,水体富含营养盐,一些异氧菌,如盐单胞菌属、交替单胞菌属、假交替单胞和弧菌属等异氧菌含量较高,浮游细菌多样性下降;而在休养期和养殖后期,营养盐含量较低,不适宜异氧菌生长,其他菌属,如玫瑰变色菌属、嗜冷杆菌属和GpIIa属等成为优势菌属,浮游细菌多样性升高。
-
不同海域浮游细菌群落组成存在差异。洋山港水域细菌主要由变形菌、拟杆菌这两大类组成,其中γ-变形菌为优势类群[18];山东近海海水样品中α-变形菌为优势类群[19];北黄海近海海水样品中γ-变形菌和黄杆菌为优势类群[20];南海南部海域浮游细菌群落优势类群为变形菌门γ-变形菌纲和α-变形菌纲、蓝藻门蓝藻菌纲和拟杆菌门黄杆菌纲[6]。临高后水湾深水网箱养殖区主要的优势类群为变形菌门的γ-变形菌。尽管如此,这些类群均为海水常见细菌类群,与国内外海洋细菌组成的相关研究结果一致[21-23]。
不同养殖期属水平优势类群差异较大,可能与温度和营养盐变化有关。Cobetia属因为具有较强的适应能力,对温度、盐度和溶解氧的适应范围广[24],成为各养殖期的主要优势类群。弧菌等异养菌的分布受温度、总有机碳、总磷和活性磷酸盐等环境因子驱动[25],在温度以及总氮或者总磷含量较高的养殖初期和中期,弧菌等异养菌的相对丰度也较高。蓝细菌丰度仅在养殖后期最高,可能的原因为养殖后期大量浮游植物繁殖,导致秋季营养盐低于春季[26],营养盐降低不利于异氧菌生长,而蓝细菌可以通过光合作用成为该时段的优势类群。嗜冷杆菌为一种革兰氏阴性菌,目前已知的嗜冷杆菌种类主要来自南极冰层、永久冻土以及海洋等环境 [27],嗜冷杆菌的分布主要与温度有关,温度越低,嗜冷杆菌相对丰度越高,也与本次研究结果一致。
-
网箱养殖卵形鲳鲹受细菌性病害和寄生虫病害危害较大,特别是由于刺激隐核虫病害发生后继发性细菌感染越来越严重。目前已报道的卵形鲳鲹细菌性病原菌包括哈维氏弧菌(Vibrio harveyi)、嗜麦芽假单胞菌(Pseudomonas maltophilia)、美人鱼发光杆菌杀鱼亚种(Photobacterium damselae subsp. piscicida)、海豚链球菌(Streptococcus iniae)和鰤鱼诺卡氏菌(Nocardia seriolae) [5,28-31],尤其以弧菌引起的病害最为严重,且哈维氏弧菌已经成为影响海南海水养殖鱼类弧菌病的主要病原菌 [32]。
网箱养殖初期和中期由于养殖密度较大,网箱内鱼类残饵及粪便增多,易引起水体中营养盐富集,为弧菌快速繁殖提供了有利条件,从而引发网箱养殖鱼类弧菌病害。湛江港网箱养殖海区弧菌丰度出现在营养盐丰富、水温较高、水体污染严重的夏、秋季,且优势弧菌种类以哈氏弧菌、溶藻弧菌、副溶血弧菌、创伤弧菌为主[10]。基于16S rDNA宏基因组构建的系统进化树分析结果显示,后水湾监测月份优势弧菌属分别与哈维氏弧菌、溶藻弧菌、副溶血弧菌、塔氏弧菌、鳗弧菌、锡那罗州弧菌等致病性弧菌聚类,进一步表明弧菌可能为引起后水湾深水网箱养殖区鱼类发病的主要潜在病原菌。
-
后水湾深水网箱养殖区浮游细菌群落具有高度丰富的多样性,变形菌门是养殖区主要优势门类,其次为拟杆菌门、放线菌门、厚壁菌门和蓝细菌门;主要类群为Cobetia属、弧菌属(Vibrio)、假交替单胞菌属(Pseudoalteromonas)、盐单胞菌属(Halomonas)以及蓝细菌门的GpIIa属等,不同养殖期优势类群差异较大。网箱内外主要差异类群为Cobetia属、假交替单胞菌属和弧菌属等。潜在致病菌弧菌属在养殖初期、中期含量较高,而在休养期和养殖后期含量较低,与网箱养殖卵形鲳鲹细菌性病害发病规律相吻合,表明弧菌病为该养殖区鱼类的主要病害类型。
浮游细菌群落结构及其与网箱养殖鱼病害的关系
DOI: 10.15886/j.cnki.rdswxb.2023.01.015
 CSTR: 32425.14.j.cnki.rdswxb.2023.01.015
CSTR: 32425.14.j.cnki.rdswxb.2023.01.015
Structure of bacterioplankton community and its relationship with bacterial diseases of fish maricultured in cage
-
摘要: 为了揭示深水网箱养殖区养殖过程中浮游细菌群落结构及其与网箱养殖鱼类细菌性病害的关系,采用Illumina测序技术对海南后水湾深水网箱养殖水样进行16S rRNA基因高通量测序。结果表明,检测到浮游细菌种群归属于36个门、56个纲、98个目、235个科、807个属,香农-威纳指数范围为2.47~4.5,说明该养殖区水体中浮游细菌具有丰富的多样性。变形菌门是主要优势门类,其次为拟杆菌门、放线菌门、厚壁菌门和蓝细菌门;主要类群为Cobetia属、弧菌属(Vibrio)、假交替单胞菌属(Pseudoalteromonas)、盐单胞菌属(Halomonas)等,不同养殖期优势类群差异较大,其中休养期以Cobetia属、玫瑰变色菌属和嗜冷杆菌属为主;养殖初期以Cobetia属、假交替单胞菌属、Kushneria属和弧菌属为主;养殖中期优势类群为Cobetia属、盐单胞菌属、交替单胞菌属(Alteromonas)、弧菌属和假交替单胞菌属;养殖后期优势类群为GpIIa属和Cobetia属。潜在致病菌弧菌属相对丰度在不同养殖期差异较大,在养殖初期、中期含量较高,而在非养殖期和养殖后期含量较低,与网箱养殖卵形鲳鲹细菌性病害发病规律相吻合,表明由于弧菌引起的鱼类病害为养殖区的主要细菌性病害类型。
-
关键词:
- 网箱养殖 /
- 16S rRNA基因高通量测序 /
- 浮游细菌群落结构 /
- 水质因子 /
- 细菌性病害
Abstract: To explore the structure of bacterioplankton community and its relationship with bacterial diseases of marine fish aquacultured in cage, bacterial communities from sea water sampled in a deep-water cage mariculture area at Houshuiwan Bay, Lingao, Hainan were analyzed through Illumina Miseq sequencing based on 16S RNA gene. The results showed that bacterioplankton species detected in the sea water samples belonged to 36 phyla, 56 classes, 98 orders, 235 families and 807 genera, with their Shannon-Wiener index ranging from 2.47 to 4.5, which indicated a high diversity of bacterioplankton in the deep-sea cage mariculture area at Houshuiwan Bay. Proteobacteria were the main dominant phylum at all the sampling sites, followed by phyla Bacteroidetes, Actinobacteria, Firmicutes and Cyanobacteria. Dominant genera were Cobetia, Roseovarius and Psychrobacter in the non-mariculture period (March), Cobetia, Pseudoalteromonas, Kushneria and Vibrio in the early period of mariculture (May), Cobetia, Halomonas, Alteromonas, Vibrio and Pseudoalteromonas in the mid-period of mariculture (July and September), GpIIa and Cobetia in the late period of mariculture. Vibrio, potential pathogens, varied greatly during the mariculture and its abundance was found higher in the early period and mid-period of mariculture, and the lower in the non- and late mariculture periods, which was consistent with the incidence of bacterial diseases of fish in the deep-water cage mariculture area, indicating that vibriosis was the main bacterial disease of fish in the mariculture area. -
图 2 不同月份网箱内外及对照区样品浮游细菌群落相似性分析
A. 细菌群落的PCA分析;B. 基于Unweighted UniFrac的聚类分析;C. 科水平的细菌群落heatmap。
图 6 各监测月份属水平网箱内外浮游细菌菌落丰度误差线图
注:图的左侧所示为不同物种分类在2个样品(组)中的丰度比例,中间所示为95%置信度区间内物种分类丰度的差异比例,最右边的值为P值;P值<0.05,表示差异显著,相应物种分类用红色标识,图中仅列出P值最低的25个物种。
-
[1] 涂志刚. 海南后水湾深水网箱养殖区微生物多样性及其养殖卵形鲳鲹“烂身病”研究[D]. 海口: 海南大学, 2017. [2] JING X, GOU H, GONG Y, et al. Seasonal dynamics of the coastal bacterioplankton at intensive fish-farming areas of the Yellow Sea, China revealed by high-throughput sequencing [J]. Marine Pollution Bulletin, 2019, 139: 366 − 375. doi: 10.1016/j.marpolbul.2018.12.052 [3] XIONG J, CHEN H, HU C, et al. Evidence of bacterioplankton community adaptation in response to long-term mariculture disturbance [J]. Scientific Reports, 2015, 5(1): 15274 − 15274. doi: 10.1038/srep15274 [4] COTNER J B, BIDDANDA B A, SMALL PLAYERS, et al. Microbial influence on biogeochemical processes in pelagic aquatic ecosystems [J]. Ecosystems, 2002, 5(2): 105 − 121. doi: 10.1007/s10021-001-0059-3 [5] 涂志刚, 崔婧, 严耿杰, 等. 网箱养殖卵形鲳鲹 “烂身病”病原分离鉴定及药敏分析[J]. 水产科学, 2018, 37(5): 634 − 639. [6] 白洁, 刘小沙, 侯瑞, 等. 南海南部海域浮游细菌群落特征及影响因素研究[J]. 中国环境科学, 2014, 34(11): 2950 − 2957. [7] 薛超波, 王建跃, 王世意, 等. 大黄鱼养殖网箱内外细菌的数量分布及区系组成[J]. 中国微生态学杂志, 2005, 17(5): 20 − 22. doi: 10.3969/j.issn.1005-376X.2005.05.009 [8] 吴建平, 蔡创华, 周毅频, 等. 大亚湾网箱养殖区异养细菌和弧菌的数量动态[J]. 广东海洋大学学报, 2006, 26(3): 21 − 25. doi: 10.3969/j.issn.1673-9159.2006.03.006 [9] 刘晶晶, 曾江宁, 陈全震, 等. 象山港网箱养殖区水体和沉积物的细菌生态分布[J]. 生态学报, 2010, 30(2): 377 − 388. [10] 邝英朋. 湛江港网箱养殖海区水质因子与细菌数量的关系及弧菌属细菌的分布[D]. 广东: 广东海洋大学, 2011. [11] CAPORASO J G, KUCZYNSKI J, STOMBAUGH J, et al. QIIME allows analysis of high-throughput community sequencing data [J]. Nature Methods, 2010, 7(5): 335 − 336. doi: 10.1038/nmeth.f.303 [12] SCHLOSS PD, WESTCOTT SL, RYABIN T, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities [J]. Applied and Environmental Microbiology, 2009, 75(23): 7537 − 7541. doi: 10.1128/AEM.01541-09 [13] WANG Q, GARRITY G M, TIEDJE J M, et al. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy [J]. Applied and Environmental Microbiology, 2007, 73(16): 5261 − 5267. doi: 10.1128/AEM.00062-07 [14] SCHLOSS P D, GEVERS D, WESTCOTT S L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies [J]. PLOS ONE, 2011, 6(12): e27310. doi: 10.1371/journal.pone.0027310 [15] OBERAUNER L, ZACHOW C, LACKNER S, et al. The ignored diversity: complex bacterial communities in intensive care units revealed by 16S pyrosequencing [J]. Scientific Reports, 2013, 3(1): 1413 − 1413. doi: 10.1038/srep01413 [16] STREIT W R, SCHMITZ R A. Metagenomics–the key to the uncultured microbes [J]. Current Opinion in Microbiology, 2004, 7(5): 492 − 498. doi: 10.1016/j.mib.2004.08.002 [17] AMANN R I, LUDWIG W, SCHLEIFER K H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation [J]. Microbiol Mol Biol Rev, 1995, 59(1): 143 − 169. [18] 薛俊增, 肖南燕, 王琼, 等. 洋山港海域细菌群落多样性的季节变化[J]. 生态学报, 2016, 36(23): 7758 − 7767. [19] 张艳. 山东近岸海域水体细菌多样性研究[D]. 青岛: 中国海洋大学, 2010. [20] 赵顺. 北黄海微表层与次表层细菌丰度及群落结构的时空变化特征研究[D]. 大连: 大连海洋大学, 2014. [21] 白洁, 李海艳, 赵阳国. 黄海北部不同站位海洋细菌群落分布特征[J]. 微生物学报, 2009, 49(3): 343 − 350. doi: 10.3321/j.issn:0001-6209.2009.03.010 [22] JING H, XIA X, SUZUKI K, et al. Vertical profiles of bacteria in the tropical and subarctic oceans revealed by pyrosequencing [J]. PLOS ONE, 2013, 8(11): e79423. doi: 10.1371/journal.pone.0079423 [23] ZENG Y X, ZHANG F, HE J F, et al. Bacterioplankton community structure in the Arctic waters as revealed by pyrosequencing of 16S rRNA genes [J]. Antonie van Leeuwenhoek, 2013, 103(6): 1309 − 1319. doi: 10.1007/s10482-013-9912-6 [24] ARAHAL D R, CASTILLO A M, LUDWIG W, et al. Proposal of cobetia marina gen. nov. comb. nov. within the family halomonadaceae, to include the species halomonas marina [J]. Systematic and Applied Microbiology, 2002, 25(2): 207 − 211. doi: 10.1078/0723-2020-00113 [25] 唐姝, 熊金波, 王凯, 等. 象山港海水网箱养殖区弧菌群落动态变化[J]. 生物学杂志, 2017, 34(4): 76 − 81. doi: 10.3969/j.issn.2095-1736.2017.04.076 [26] 陈丹丹, 庞巧珠, 陈晓慧, 等. 2018年春秋季海南后水湾浮游植物群落结构特征及其与环境因子的关系[J]. 海洋湖沼通报, 2020, 42(5): 113 − 120. [27] BOWMAN J P. The Genus Psychrobacter[M]. In: DWORKIN M, FALKOW S, ROSENBERG E, et at. The Prokaryotes: Volume 6: Proteobacteria: Gamma Subclass. New York: Springer, 2006. [28] 周永灿, 朱传华, 张本, 等. 卵形鲳鲹大规模死亡的病原及其防治[J]. 海洋科学, 2001, 25(4): 40 − 44. doi: 10.3969/j.issn.1000-3096.2001.04.013 [29] 王瑞旋, 冯娟, 苏友禄, 等. 卵形鲳鲹美人鱼发光杆菌杀鱼亚种的分离鉴定[J]. 中国水产科学, 2010, 17(5): 1020 − 1027. [30] 黄婷, 李莉萍, 王瑞, 等. 卵形鲳鲹感染无乳链球菌与海豚链球菌的研究[J]. 大连海洋大学学报, 2014, 29(2): 161 − 166. doi: 10.3969/J.ISSN.2095-1388.2014.02.012 [31] 王瑞旋, 刘广锋, 王江勇, 等. 养殖卵形鲳鲹诺卡氏菌病的研究[J]. 海洋湖沼通报, 2010, 32(1): 52 − 58. doi: 10.3969/j.issn.1003-6482.2010.01.008 [32] 崔婧, 范雪亭, 刘文竹, 等. 华南地区海水养殖鱼类主要弧菌病原的分离与鉴定[J]. 海南大学学报(自然科学版), 2014, 32(3): 244 − 251. -

 点击查看大图
点击查看大图

图(7)
计量
- 文章访问数: 1220
- HTML全文浏览量: 719
- PDF下载量: 23
- 被引次数: 0
