

-
水稻(Oryza sativa)是全球最重要的粮食作物之一,其高产与稳产对保障世界粮食安全具有至关重要的意义。近年来,受自然环境演变及不合理灌溉等因素影响,土壤盐渍化问题日益突出 [1-2]。据统计,中国盐渍化土地总面积约 9 913 万hm2,大面积盐渍化土壤严重制约了中国农业生产与粮食供给 [3]。水稻属于盐敏感作物,在盐渍化环境中易遭受离子胁迫、氧化胁迫、渗透胁迫及pH胁迫等,造成生长发育不可逆损伤,进而严重影响产量,甚至导致绝收 [4-6]。因此,挖掘水稻耐盐基因、培育耐盐水稻新品种对保障我国粮食安全具有重要意义。
当前水稻育种面临遗传背景日趋狭窄的瓶颈,仅依靠现有栽培稻种质资源,难以实现耐盐性的系统性遗传改良。普通野生稻(Oryza rufipogon)作为栽培稻的近缘祖先种,具有极强的环境适应性与丰富的遗传多样性,多项研究已证实其蕴藏大量包括耐盐碱在内的优异抗逆等位基因 [7-8]。在长期的驯化过程中,人工定向选择使其形态发生明显的改变,如株型由匍匐生长转为直立生长 [9]、穗型由散穗变为紧穗 [10],落粒性 [11-12]、芒性 [13-14]及种子休眠性 [15]等性状减弱或消失,从而演变成现今的栽培稻。在这一过程中,除控制目标性状的关键基因外,大量天然等位基因因“搭车效应”被被一同保留,导致栽培稻遗传多样性显著下降。研究显示,栽培稻保留的等位基因数量仅为野生稻的 60% [16],在丢失的等位变异中,很可能包含大量与耐盐等抗逆性状相关的优异基因。因此,从野生稻中挖掘并利用优异耐盐等位基因开展水稻遗传改良,可有效拓宽栽培稻遗传背景,突破现有遗传资源狭窄的瓶颈,为培育耐盐水稻新品种提供重要基因资源。
水稻在应对盐胁迫时,维持细胞及体内适宜的 Na+/K+平衡至关重要。高亲和性钾离子转运体家族(HAK/KUP/KT)在该过程中发挥关键作用,广泛参与根系 K+吸收、向地
上部长距离运输及胞内离子区隔化等过程,进而影响渗透调节、酶活性维持及细胞信号转导等一系列生理代谢过程。系统进化分析表明,水稻基因组中包含27个HAK家族基因,可分为4个亚家族、6个进化分支(Clade Ⅰa、Ⅰb、Ⅱa、Ⅱb、Ⅲ、Ⅳ) [17]。不同亚族的HAK基因存在明显功能分化,这也是水稻适应复杂环境的重要分子基础。已有研究显示,OsHAK1参与维持 K+介导的正常生长,并在盐胁迫下调控体内 K+平衡 [18];OsHAK18主要在维管组织中表达,介导 K+的装载与再分配及糖转运,过表达OsHAK18可增加韧皮部汁液与根系 K+浓度,促进 Na+由地上部向根系回流,增强光合产物源库转运效率,从而提升水稻耐盐性 [19-20];OsHAK7主要在茎和根中表达,推测参与 K+运输过程 [21],盐胁迫下其表达量上调可显著增强水稻耐盐性 [22];OsHAK9通过调控 K+外排与赤霉素代谢途径,介导水稻种子萌发期的耐盐性 [23];OsHAK13在开花期盐胁迫早期表达显著上调,胁迫后期则表达下降,可能通过维持离子稳态在盐胁迫初期发挥重要作用 [24]。
鉴于HAK家族基因在水稻耐盐性调控中的重要作用,本研究基于水稻超级泛基因组信息资源数据库中的基因组数据,对HAK家族序列变异进行了系统分析,并以‘9311’及其与云南元江普通野生稻所构建的渗入系为试验材料,鉴定了OsHAK7、OsHAK9和OsHAK13的耐盐优异单倍型。
-
本研究以籼稻‘9311’为受体与云南元江普通野生稻为供体亲本构建的178份BC3F8渗入系群体为基础材料,并以‘9311’作为对照。试验材料均在人工气候室进行水培种植,环境参数设定为:14 h光照(28℃)/10 h黑暗(25℃),光照强度约500 µmol·(m2·s)−1,空气相对湿度维持在70%。正常培养阶段采用基于经典Yoshida配方的水稻标准干粉培养基配制的营养液。营养液每3 d彻底更换1次,并调节pH至5.5~5.8,以维持根系最佳的离子吸收微环境。
-
为探究HAK家族基因在水稻驯化中的受选择情况,本研究从水稻超级泛基因组信息资源数据库(
http://www.ricesuperpir.com/ )中检索并下载了27个HAK家族基因的基因组序列。该数据库共涵盖了251份具备高度遗传代表性的栽培稻与野生稻参考基因组 [25]。利用MEGA 6.0软件对基因组序列和氨基酸序列进行多重比对,并采用DnaSP v5软件分别计算野生稻、籼稻和粳稻群体中各基因的核苷酸多样性(π)和中性检验统计量(Tajima’s D值),据此筛选出在驯化中受强烈选择的候选靶标基因。序列分析所用材料主要是OsHAK7、OsHAK9和OsHAK13基因。用于分析OsHAK7基因序列的材料共211份,包括野生稻材料21份,籼稻材料132份,粳稻材料58份;用于分析OsHAK9基因序列的材料共211份,包括野生稻材料24份,籼稻材料129份,粳稻材料58份;用于分析OsHAK13基因序列的材料共214份,包括野生稻材料24份,籼稻材料133份,粳稻材料57份。表 1 HAK基因序列分析所用种质资源构成
Table 1. Composition of germplasm for HAK gene sequence analysis
基因名称
Gene name野生稻/份
Wild籼稻/份
Indica粳稻/份
Japonica总计/份
TotalOsHAK7 21 132 58 211 OsHAK9 24 129 58 211 OsHAK13 24 133 57 214 -
为验证前期泛基因组数据锁定的候选HAK基因的功能,本研究对178份BC3F8渗入系群体开展苗期盐胁迫表型初筛。试验采用完全随机设计,每份材料设3组生物学重复,每组重复24株。种子经消毒催芽后,播种于96孔PCR板中水培。待幼苗生长至二叶一心期时,将其划分为2组:对照组持续使用正常营养液培养,处理组则转入含有175 mmol·L−1 NaCl的营养液中进行盐胁迫处理。连续胁迫处理9 d后,处理组与对照组间呈现出显著的耐盐表型差异,将处理组转移至正常培养液中恢复生长7 d。通过统计幼苗存活率,筛选出耐盐性显著优于对照的候选材料。将初筛获得的高抗材料表型与全基因组重测序及渗入片段进行分析,结合前期泛基因组数据分析结果,锁定了3个在驯化中受强烈选择的候选基因(OsHAK7、OsHAK9和OsHAK13),最终从178份渗入系群体中筛选出分别携带云南元江普通野生稻来源OsHAK7、OsHAK9和OsHAK13基因区段且耐盐表型较为稳定的材料9YJ-12、9YJ-97和9YJ-100,作为渗入系材料。
对上述3份核心渗入系材料及对照品种‘9311’开展苗期耐盐性复筛试验。当幼苗二叶一心期时,将处理组幼苗转入含150 mM NaCl的营养液中持续胁迫14 d,对照组则正常培养。胁迫结束后将所有幼苗转移至正常营养液中恢复7 d。详细记录植株的受害表型及恢复情况并统计最终存活率。所得数据采用SPSS 25.0软件进行单因素方差分析与显著性检验。
-
取复筛验证后的渗入系9YJ-12、9YJ-97、9YJ-100和对照材料‘9311’的新鲜叶片1 g,液氮速冻后充分研磨,使用Solarbio公司的2×CTAB提取缓冲液并参照其说明书进行基因组DNA的提取与纯化。
-
利用Phytozome数据库(
https://phytozome-next.jgi.doe.gov/ )获取OsHAK7、OsHAK9和OsHAK13的参考序列,分段设计覆盖各基因编码序列(coding sequence,CDS)的特异性引物。所有引物均由擎科生物技术有限公司合成,引物序列详见表2。以9YJ-12、9YJ-97和9YJ-100的DNA为模板分段进行扩增,扩增体系为DNA模板1 µL(200 ng)、100 µmol·L−1 Primer F/R各1 µL、ApexHF HS DNA Polymerase CL 1 µL、2×ApexHF CL Buffer 10 µL、H2O 6 µL,总体积20 µL。扩增程序:94℃预变性2 min,98℃变性10 s、60℃退火15 s、68℃延伸40 s(延伸时间根据产物长度按1 kb·min−1调整),35个循环,最后68℃终延伸10 min。取3µL PCR产物,使用1%的琼脂糖凝胶(含GelRed染料)进行电泳检测,将目的条带清晰、无杂带的扩增产物送至有康生物有限公司进行双向测序。表 2 OsHAK7、OsHAK9、OsHAK13基因分段设计覆盖各基因CDS的特异性引物
Table 2. Specific primers designed in segments to cover the entire CDS of OsHAK7, OsHAK9 and OsHAK13 genes
引物名称
Primer正向引物序列(5′−3′)
Forward primer sequence(5′−3′)反向引物序列(3′−5′)
Reverse primer sequence (3′−5′)OsHAK7-1F/R AAATTTGCGTGATCTATAGT TAACTAGACAGAACAATGCA OsHAK7-2F/R TGCATTGTTCTGTCTAGTTA TAGTAATGGCTGCTGGAATA OsHAK7-3F/R TATTCCAGCAGCCATTACTA CCGCTGCAAGAGTTGCAATG OsHAK7-4F/R GTAATCTTCATAACTCCAAG CAGATAAGAGAGCTATCATC OsHAK7-5F/R GATGATAGCTCTCTTATCTG GCACCGGTACATACGGTATT OsHAK7-6F/R TGCCTTCCATCTTCTCGCAC GGCAACTACATTACATGTAT OsHAK9-1F/R TTCCCAGGTGCGTCCTCTAT TGTAAGCAACTCTTTTCTCC OsHAK9-2F/R ATTGGAACCTGTATGGTCAT GCTAACTTCATCCAATAATC OsHAK9-3F/R TGCGTATGCTGAGATAAAGT TCACATTCTAAGCAACGCAA OsHAK9-4F/R CCATTACCATCCTGTCAGCT TACCGCACGATGCACCGGTA OsHAK9-5F/R TCTACACCGACCTCACCTCC TCGCCGTCGCCGGTTTCATC OsHAK13-1F/R AAGGCAGCAAGAATTCAGTA AGAATCCGCAGTAACACTGC OsHAK13-2F/R GAATACTATCTTGTTGCCTA CCAATATATCAGCTGCTTAC OsHAK13-3F/R CGTATATGATAGAGTAAGGA CTATCACCAATTCACCACTG OsHAK13-4F/R CAGTGGTGAATTGGTGATAG AATCAATGTACTCTCTGTTG OsHAK13-5F/R CTAATGCAGCAATTGAAGTT ATATCCGGTATCAGAAGGTA OsHAK13-6F/R TGGTCCGTAAGATATATCAA GACCGACATGGCCTCGTCCT OsHAK13-7F/R GCCTGTTCCGGTGCATCGTC CTACACTCTCTTCAGCGATG -
根据OsHAK7、OsHAK9和OsHAK13基因组序列比对结果,以CDS变异为多态性标记构建单倍型,并使用NETWORK v10软件通过中介网络法构建单倍型网络图来分析单倍型间的进化关系,ε参数设置为0(最简约网络)。
-
为了探索HAK家族基因在水稻驯化过程中是否受到人工选择,本研究从水稻超级泛基因组信息资源数据库中下载了27个HAK家族基因的基因组序列,并对其进行核苷酸多态性(π)及Tajima’s D值分析。
全家族的群体遗传学分析结果显示,HAK家族成员在水稻演化过程中经历了显著的亚种分化与选择性清除(表3~7)。具体而言,在整个栽培稻群体中,仅OsHAK7基因的核苷酸多态性相较于野生稻显著下降,且Tajima’s D值为−2.795 89(P < 0.001),表明该基因在早期驯化中受到了强烈的定向选择(表3)。在籼稻群体中,OsHAK5、OsHAK7、OsHAK8、OsHAK14、OsHAK15、OsHAK18、OsHAK20、OsHAK23基因均受到不同程度的选择,但仅OsHAK7基因受到强烈的选择(表3~7)。在粳稻群体中,OsHAK3、OsHAK5、OsHAK6、OsHAK7、OsHAK8、OsHAK9、OsHAK13、OsHAK15、OsHAK17、OsHAK18、OsHAK19、OsHAK20、OsHAK21、OsHAK22、OsHAK23、OsHAK27均受到不同程度的选择(Tajima’s D均小于−2.0,P < 0.01),其中,OsHAK13、OsHAK21、OsHAK27、OsHAK18、OsHAK9基因受选择程度最为强烈(表3~7)。
表 3 HAK家族核心候选基因OsHAK7、OsHAK9、OsHAK13的核苷酸多样性
Table 3. Nucleotide diversity of the core candidate genes OsHAK7, OsHAK9, and OsHAK13 in the HAK family
Locus
基因号Gene
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻π
核苷酸
多态性Tajima’s D π
核苷酸
多态性π ratio
(Cultivar/Wild)π比值(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_
Os07g47350OsHAK7 0.004 5 −0.510 59 P > 0.10 0.000 2 0.042 0 −2.795 89 ***, P < 0.001 0.000 2 0.033 2 −2.586 5 ***, P < 0.001 0.000 3 0.059 7 2.79826 ***,
P < 0.001LOC_
Os07g48130OsHAK9 0.003 8 −1.304 49 P > 0.10 0.001 9 0.502 7 −0.540 56
P > 0.100.002 4 0.633 0 0.408 81
P > 0.100.000 3 0.071 8 − 2.46569 **,
P < 0.01LOC_
Os06g45940OsHAK13 0.007 5 −0.769 11 P > 0.10 0.000 9 0.118 1 − 1.70986 0.10 > P > 0.050.001 2 0.162 4 −1.355 43
P > 0.100.000 1 0.018 8 − 2.43112 **,
P < 0.01表 7 HAK亚家族Ⅳ的核苷酸多态性
Table 7. Nucleotide polymorphism of HAK subfamily Ⅳ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os07g01214 OsHAK22 0.004 3 − 1.24745
P > 0.100.004 6 1.070 1 2.64627 *,
P < 0.050.003 1 0.731 3 0.7771
P > 0.100.002 7 0.621 5 3.16076 **,
P < 0.01Clade IV LOC_Os09g21000 OsHAK23 0.004 8 − 0.48915
P > 0.100.005 3 1.100 2 − 2.65601 ***,
P < 0.0010.003 8 0.782 9 − 2.91589 ***,
P < 0.0010.000 4 0.089 8 − 2.7639 ***,
P < 0.001Clade IV LOC_Os06g15910 OsHAK24 0.005 4 − 1.87643 *,
P < 0.050.003 7 0.686 9 0.94237
P > 0.100.003 2 0.594 8 1.58062
P > 0.100.002 5 0.464 1 − 0.04634
P > 0.10Clade IV LOC_Os02g49760 OsHAK25 0.004 0 − 1.29599
P > 0.100.003 0 0.738 8 1.46783
P > 0.100.002 4 0.589 6 0.66543
P > 0.100.000 2 0.047 3 − 1.02112
P > 0.10Clade IV LOC_Os08g39950 OsHAK26 0.010 9 0.28084
P > 0.100.001 6 0.145 1 − 0.46248
P > 0.100.001 6 0.150 6 − 0.46345
P > 0.100.000 2 0.013 8 − 1.64706 0.10 > P > 0.05Clade IV 表 4 HAK 亚家族Ⅰ的核苷酸多态性
Table 4. Nucleotide Polymorphism of HAK Subfamily Ⅰ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化
分支π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Cultivar/
Wild)
π比值
(栽培稻/
野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/
Wild)
π比值
(籼稻/
野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/
Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_
Os07g01214OsHAK22 0.004 3 − 1.24745 P > 0.100.004 6 1.070 1 2.64627 *,
P < 0.050.003 1 0.731 3 0.7771
P > 0.100.002 7 0.621 5 3.16076 **,
P < 0.01Clade IV LOC_
Os09g21000OsHAK23 0.004 8 − 0.48915 P > 0.100.005 3 1.100 2 − 2.65601 ***,
P < 0.0010.003 8 0.782 9 − 2.91589 ***,
P < 0.0010.000 4 0.089 8 − 2.7639 ***, P < 0.001Clade IV LOC_
Os06g15910OsHAK24 0.005 4 − 1.87643 *,
P < 0.050.003 7 0.686 9 0.94237
P > 0.100.003 2 0.594 8 1.58062
P > 0.100.002 5 0.464 1 − 0.04634
P > 0.10Clade IV LOC_
Os02g49760OsHAK25 0.004 0 − 1.29599 P > 0.100.003 0 0.738 8 1.46783
P > 0.100.002 4 0.589 6 0.66543
P > 0.100.000 2 0.047 3 − 1.02112 P > 0.10Clade IV LOC_
Os08g39950OsHAK26 0.010 9 0.28084 P > 0.100.001 6 0.145 1 − 0.46248
P > 0.100.001 6 0.150 6 − 0.46345
P > 0.100.000 2 0.013 8 − 1.64706 0.10 > P > 0.05Clade IV 表 5 HAK 亚家族Ⅱ的核苷酸多态性
Table 5. Nucleotide Polymorphism of HAK Subfamily Ⅱ
Locus
基因号Gene
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D π
核苷酸
多态性π ratio
(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os04g32920 OsHAK1 0.018 4 0.4915
P > 0.100.003 1 0.170 2 0.63282
P > 0.100.001 5 0.083 2 − 1.33417
P > 0.100.001 7 0.089 7 1.16604
P > 0.10Clade Ia LOC_Os08g36340 OsHAK4 0.002 8 − 1.41127
P > 0.100.003 3 1.187 1 − 0.33119
P > 0.100.003 3 1.197 8 − 0.27482
P > 0.100.003 4 1.233 8 0.11114
P > 0.10Clade Ia LOC_Os01g70940 OsHAK2 0.003 2 − 1.48245
P > 0.100.001 9 0.583 9 1.22065
P > 0.100.000 9 0.291 9 − 0.9904
P > 0.100.000 1 0.043 5 − 0.90566
P > 0.10Clade Ib LOC_Os01g27170 OsHAK3 0.005 2 − 1.06352
P > 0.100.005 8 1.112 2 − 0.24798
P > 0.100.006 0 1.150 9 0.50134
P > 0.100.001 1 0.210 8 − 2.53964 ***,
P < 0.001Clade Ib LOC_Os01g70490 OsHAK5 0.002 2 − 1.24384
P > 0.100.001 5 0.701 8 0.71477
P > 0.100.000 4 0.192 7 − 1.84369 *, P < 0.050.000 5 0.211 0 − 1.95666 *,
P < 0.05Clade Ib LOC_Os01g70660 OsHAK6 0.008 0 − 0.64962
P > 0.100.006 6 0.818 8 3.13463 **,P< 0.010.002 3 0.288 8 − 0.83886
P > 0.100.000 7 0.087 5 − 2.81216 ***,
P < 0.001Clade Ib LOC_Os03g37830 OsHAK27 0.004 9 − 1.28224
P > 0.100.003 8 0.765 3 1.03719
P > 0.100.001 7 0.353 1 − 0.86236
P > 0.100.000 2 0.046 9 − 2.8289 ***,
P < 0.001Clade Ib 表 6 HAK 亚家族Ⅲ的核苷酸多态性
Table 6. Nucleotide Polymorphism of HAK Subfamily Ⅲ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os03g37840 OsHAK16 0.004 5 − 1.55764 P>0.100.002 9 0.634 4 1.65425
P>0.100.000 9 0.198 2 − 1.73911 0.10>P>0.050 0.008 8 − 1.2672 ,
P>0.10Clade III LOC_Os09g27580 OsHAK17 0.004 9 − 1.03831 P>0.100.003 7 0.745 4 0.06718
P>0.100.002 3 0.468 4 − 1.08931 P>0.100.001 1 0.224 0 − 2.32309 **, P < 0.01Clade III LOC_Os09g38960 OsHAK18 0.006 3 − 1.17953 P>0.100.004 5 0.715 0 − 1.08768 P>0.100.002 2 0.355 1 − 2.2962 **,
P < 0.010.000 4 0.065 3 − 2.74023 ***,
P < 0.001Clade III LOC_Os02g31910 OsHAK19 0.003 1 − 0.81272 P>0.100.002 8 0.891 4 1.58937
P>0.100.001 5 0.476 0 − 0.35248 P>0.100.000 6 0.204 5 − 2.04205 *,
P < 0.05Clade III LOC_Os02g31940 OsHAK20 0.006 1 − 1.59181 0.10>P>0.050.003 3 0.544 4 − 1.88162 *, P < 0.050.002 4 0.399 7 − 2.22657 **,
P < 0.010.001 7 0.281 3 − 2.17062 *,
P < 0.05Clade III LOC_Os03g37930 OsHAK21 0.018 4 0.89146 P>0.100.004 6 0.249 7 0.52287
P>0.100.003 2 0.171 0 − 0.61199 P>0.100.000 3 0.017 4 − 2.33498 **, P < 0.01Clade III -
群体序列分析从基因组学层面揭示了HAK家族基因在水稻驯化过程中的演化特征。然而,单纯的生物信息学推断不足以明确这些自然变异的实际生物学效应。为了进一步验证上述HAK家族基因的功能,本研究以‘9311’为对照,对178份BC3F8渗入系进行175 mmol·L−1 NaCl极端盐胁迫表型鉴定。幼苗长至二叶一心期时进行盐处理,持续胁迫9 d后,转入正常营养液恢复7 d并统计存活率。图1结果显示,‘9311’对胁迫高度敏感,恢复期出现大面积枯死,最终存活率仅为3.5%。而渗入系群体的耐盐表型出现显著遗传分离。以20.0%为存活率为阈值,初筛共鉴定出17份耐盐表现突出的候选材料。将这17份材料的表型数据与全基因组重测序及渗入片段进行分析。结果显示,渗入系9YJ-12、9YJ-97和9YJ-100的染色体导入区段精准覆盖了OsHAK7、OsHAK9和OsHAK13所在的野生稻单倍型物理区间。基于此,确定这3份材料为后续机制研究的核心实验材料。
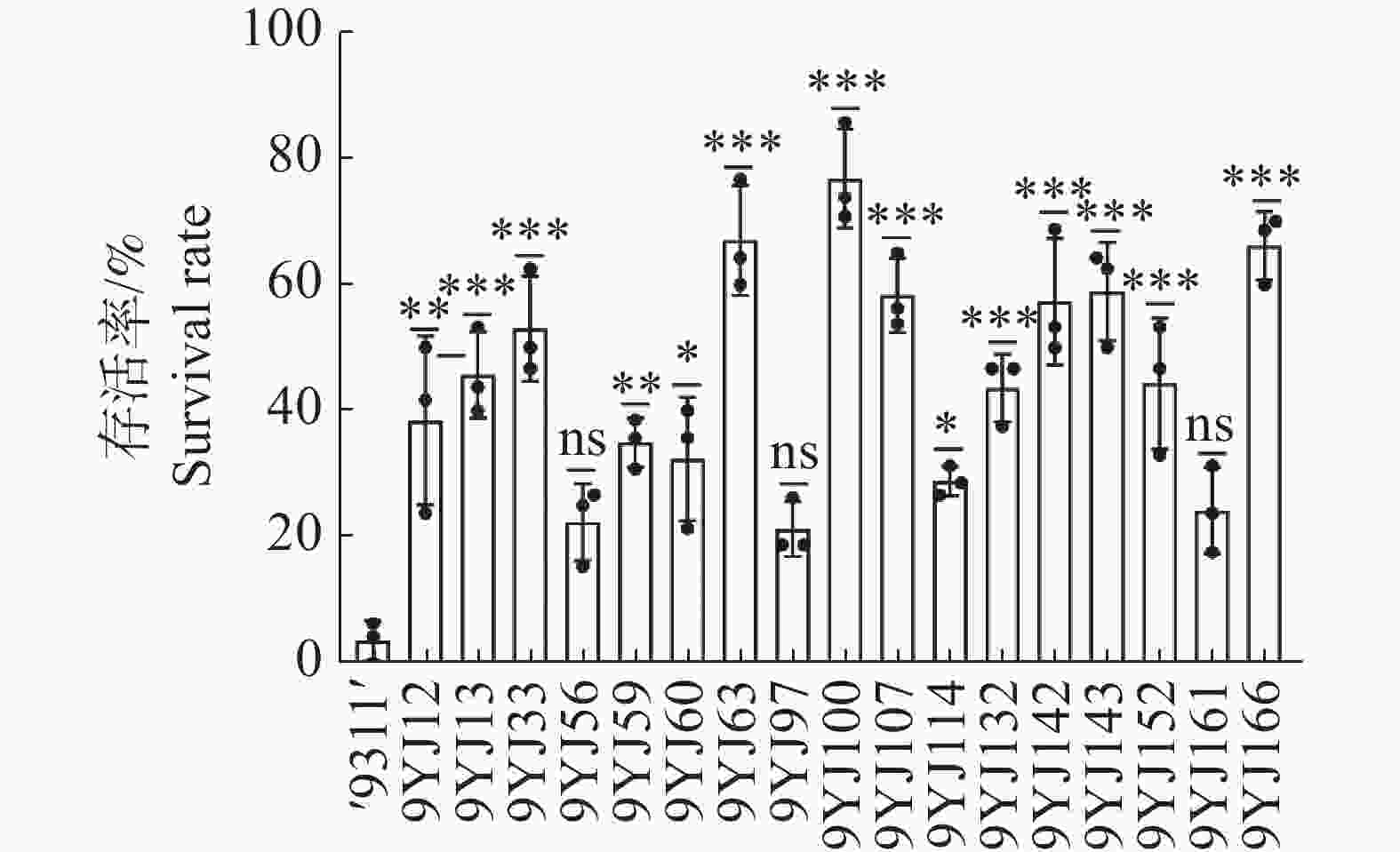
图 1 野生稻渗入系苗期耐盐性初筛与候选材料存活率分析
Figure 1. Initial screening of seedling salt tolerance in introgression lines and survival rate analysis of candidate lines
-
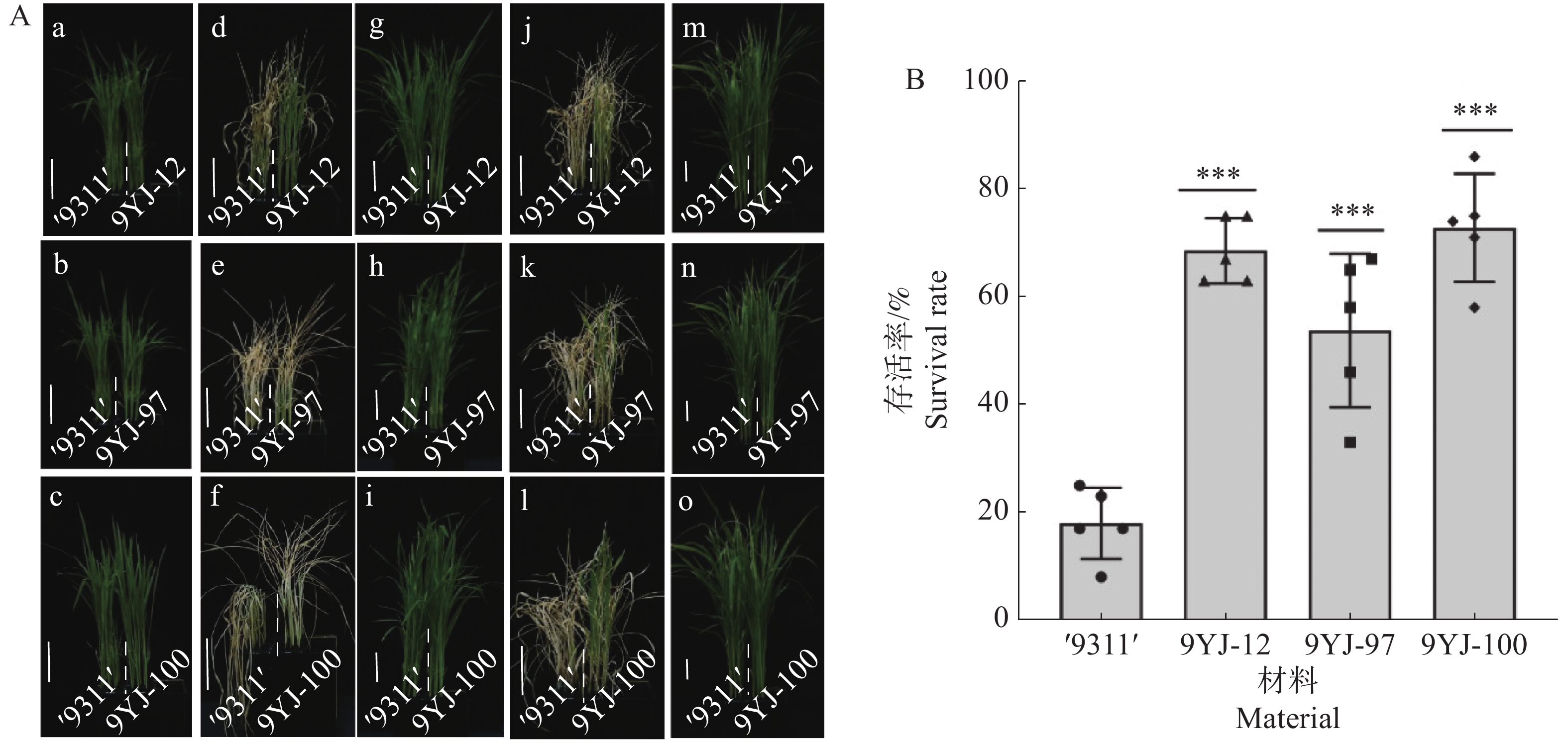
为系统验证野生稻靶基因导入区段的耐盐稳定性,本研究对携有云南元江普通野生稻OsHAK7、OsHAK9和OsHAK13基因的3份渗入系9YJ-12、9YJ-97和9YJ-100以及对照‘9311’开展150 mmol·L−1 NaCl盐胁迫复筛。在两叶一心期对4份材料用150 mmol·L−1 NaCl营养液处理14 d,28℃条件下恢复生长7 d,统计存活率。结果显示,9YJ-12、9YJ-97、9YJ-100的存活率分别为69%、54%、73%,相较于对照‘9311’(18%)分别显著提高了51%、36%、55%(图2)。研究结果表明,携有云南元江普通野生稻OsHAK7、OsHAK9、OsHAK13基因的渗入系能显著增强水稻耐盐性。
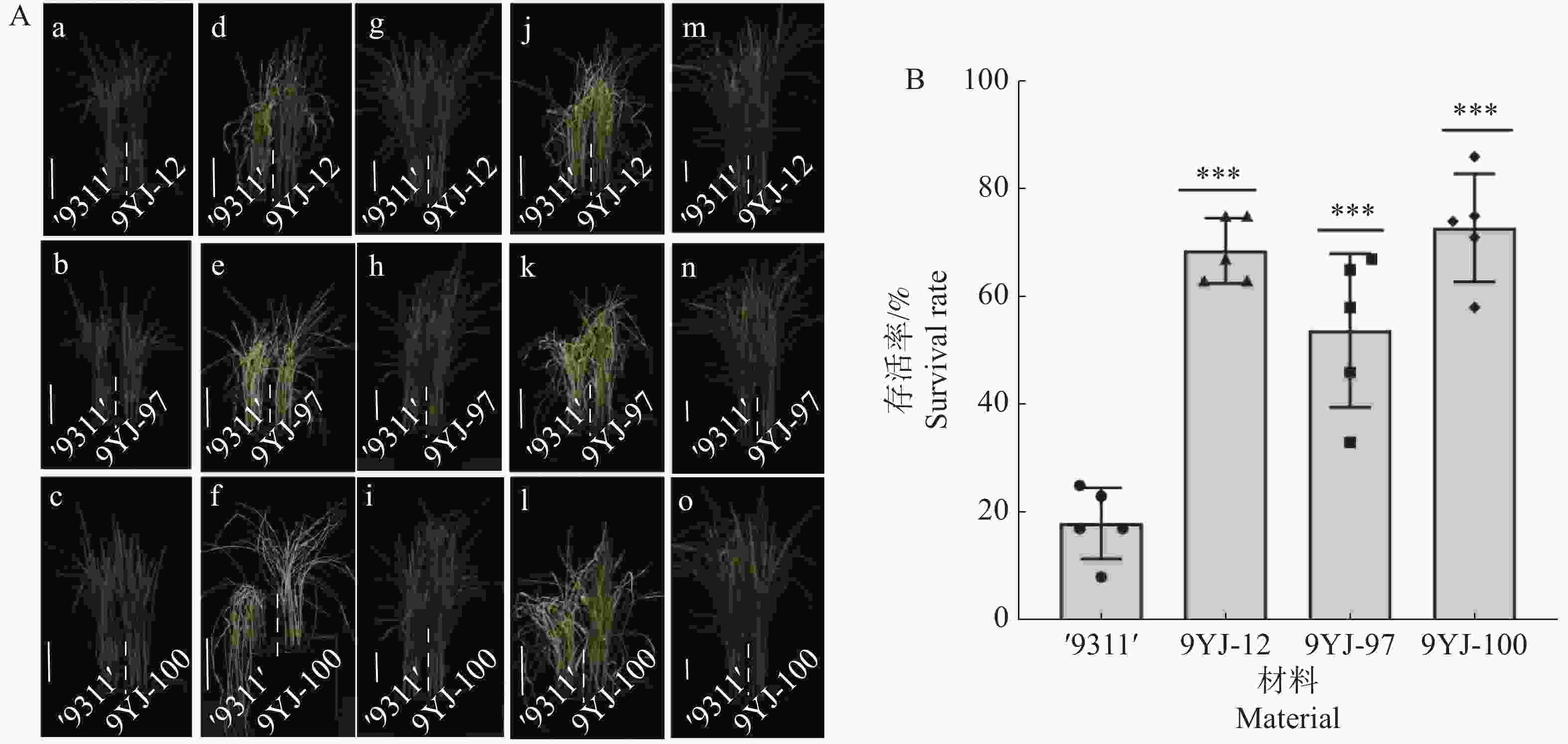
图 2 ‘9311’与核心渗入系耐盐性表型分析
Figure 2. Phenotypic analysis of salt tolerance of ‘9311’ and the core introgression lines
-
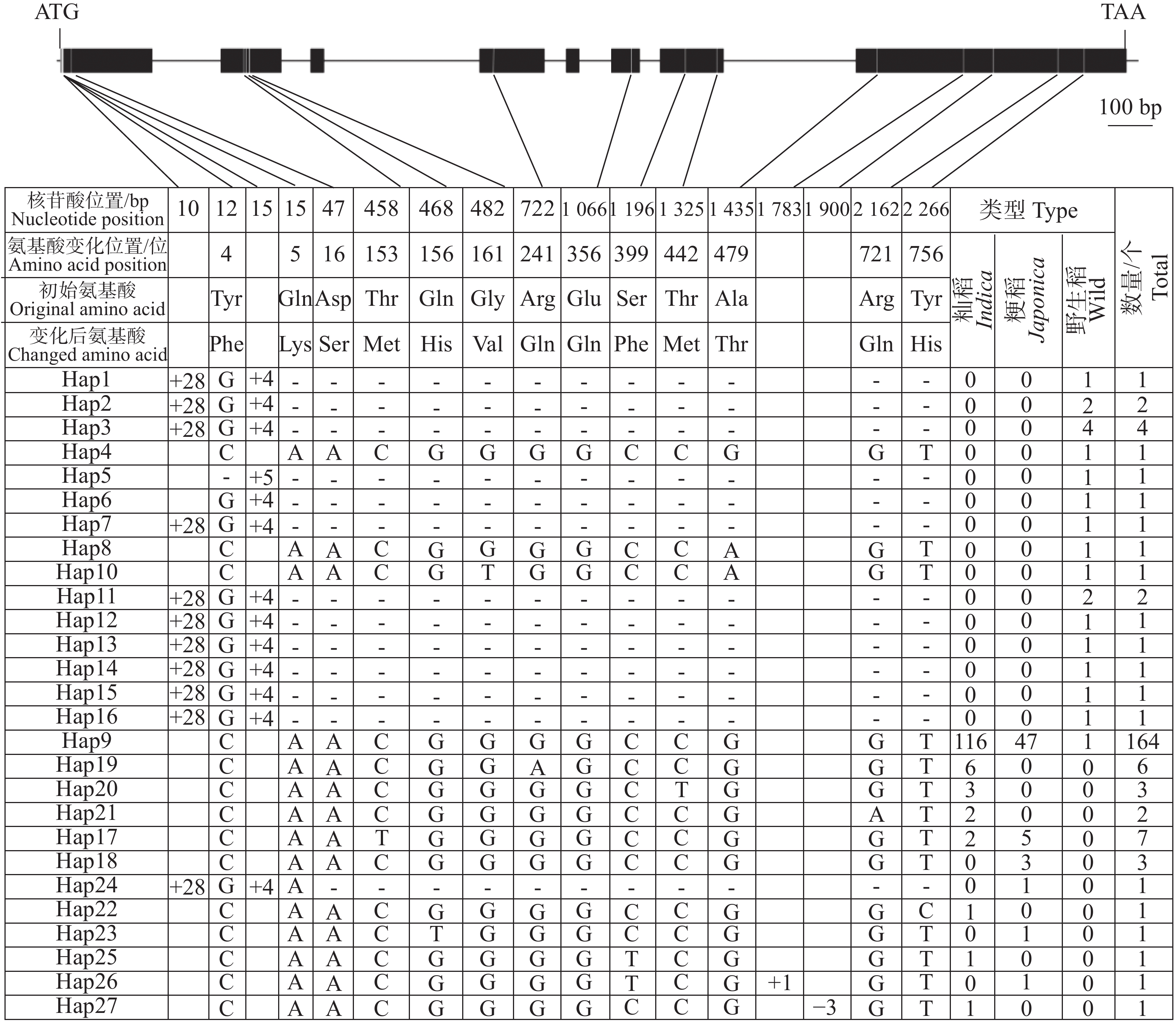
鉴于云南元江普通野生稻中的OsHAK7、OsHAK9和OsHAK13等位基因可能具有耐盐功能,本研究对这3个基因在野生稻和栽培稻中的序列变异进行了系统分析。OsHAK7(LOC_Os07g47350)基因全长5 563 bp,编码区长度为2 436 bp。对211份水稻种质基因组序列分析结果显示,OsHAK7基因的编码区存在17处核苷酸变异,包括4个插入/缺失变异(Insertion/Deletion,InDel)和13个单核苷酸多态性(Single Nucleotide Polymorphism,SNP)变异(图3)。其中,4个InDel分别于编码区第10 bp和第15 bp插入28个和4个碱基、第
1783 bp插入1个碱基以及第1900 bp缺失3个碱基;13个SNP位点均引起了氨基酸发生变化,这13个氨基酸变化分别位于以下位置:第4位(Tyr/Phe)、第5位(Gln/Lys)、第16位(Asp/Ser)、第153位(Thr/Met)、第156位(Gln/His)、第161位(Gly/Val)、第241位(Arg/Gln)、第356位(Glu/Gln)、第399位(Ser/Phe)、第442位(Thr/Met)、第479位(Ala/Thr)、第721位(Arg/Gln)和第756位(Tyr/His)(图3)。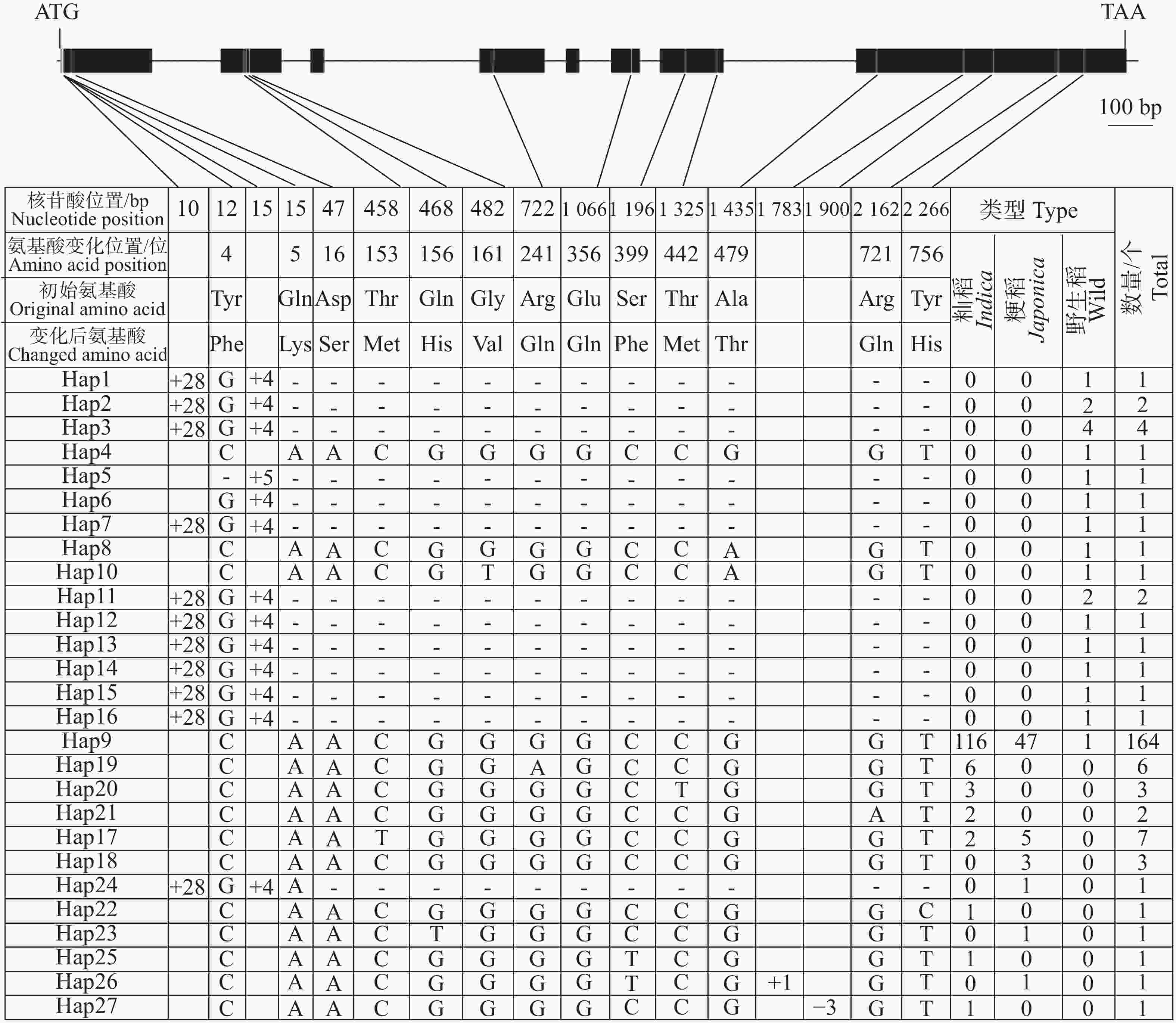
图 3 OsHAK7基因编码区核苷酸多样性
Figure 3. Nucleotide diversity of the OsHAK7 gene coding sequence
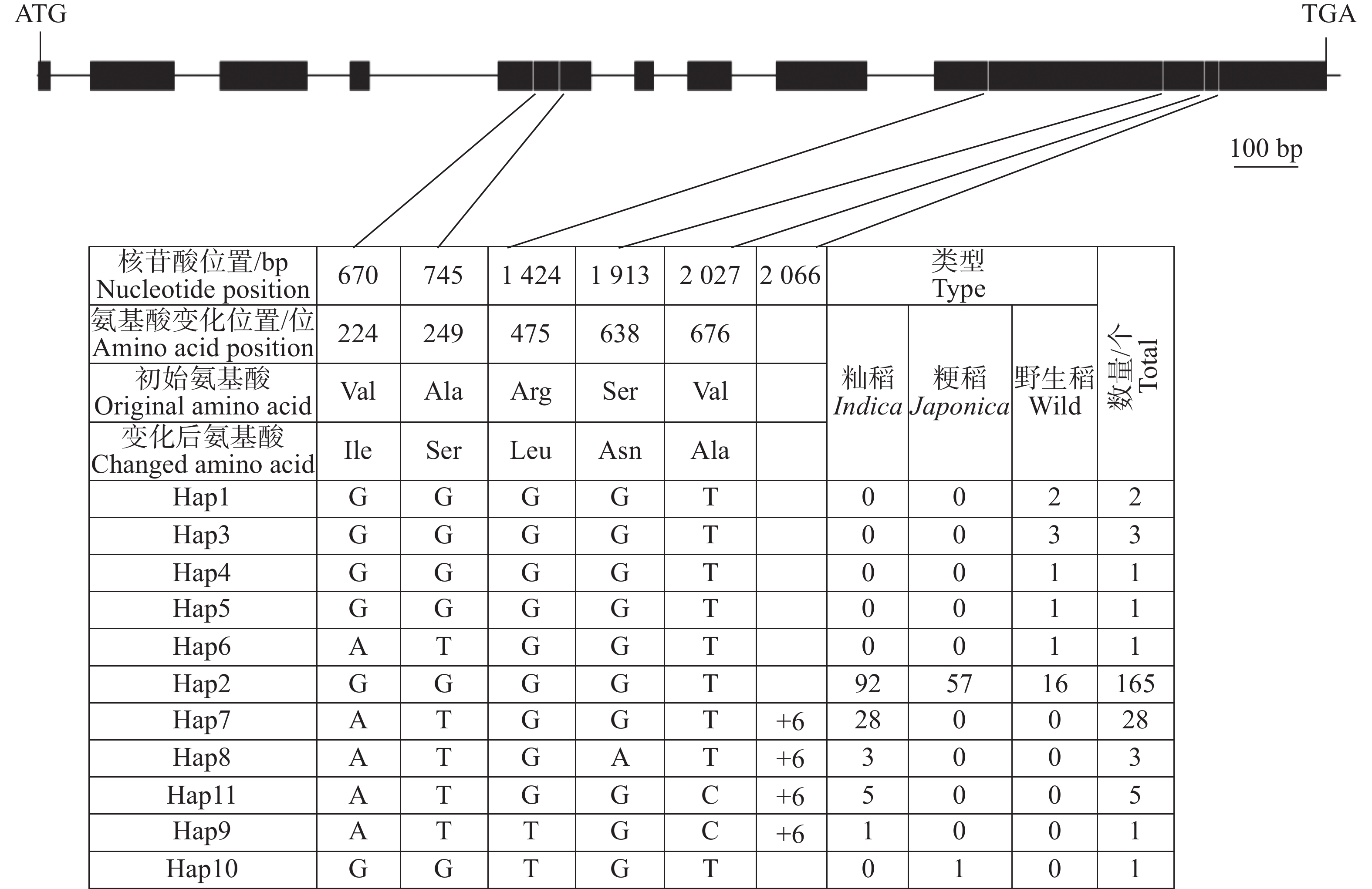
OsHAK9(LOC_Os07g48130)基因全长5 542 bp,编码区长度为2 367 bp。对211份水稻种质基因组序列分析表明,在OsHAK9基因的编码区共存在6处核苷酸变异,包括1个InDel和5个SNP变异。其中,1个InDel于编码区第206 6 bp插入6个碱基;5个SNP位点均引起了氨基酸的变化,分别位于编码区第670位、第745位、第1 424位、第1 913位和第2 027位各存在1个单碱基替换,均引起相应位置的氨基酸发生改变(图4)。
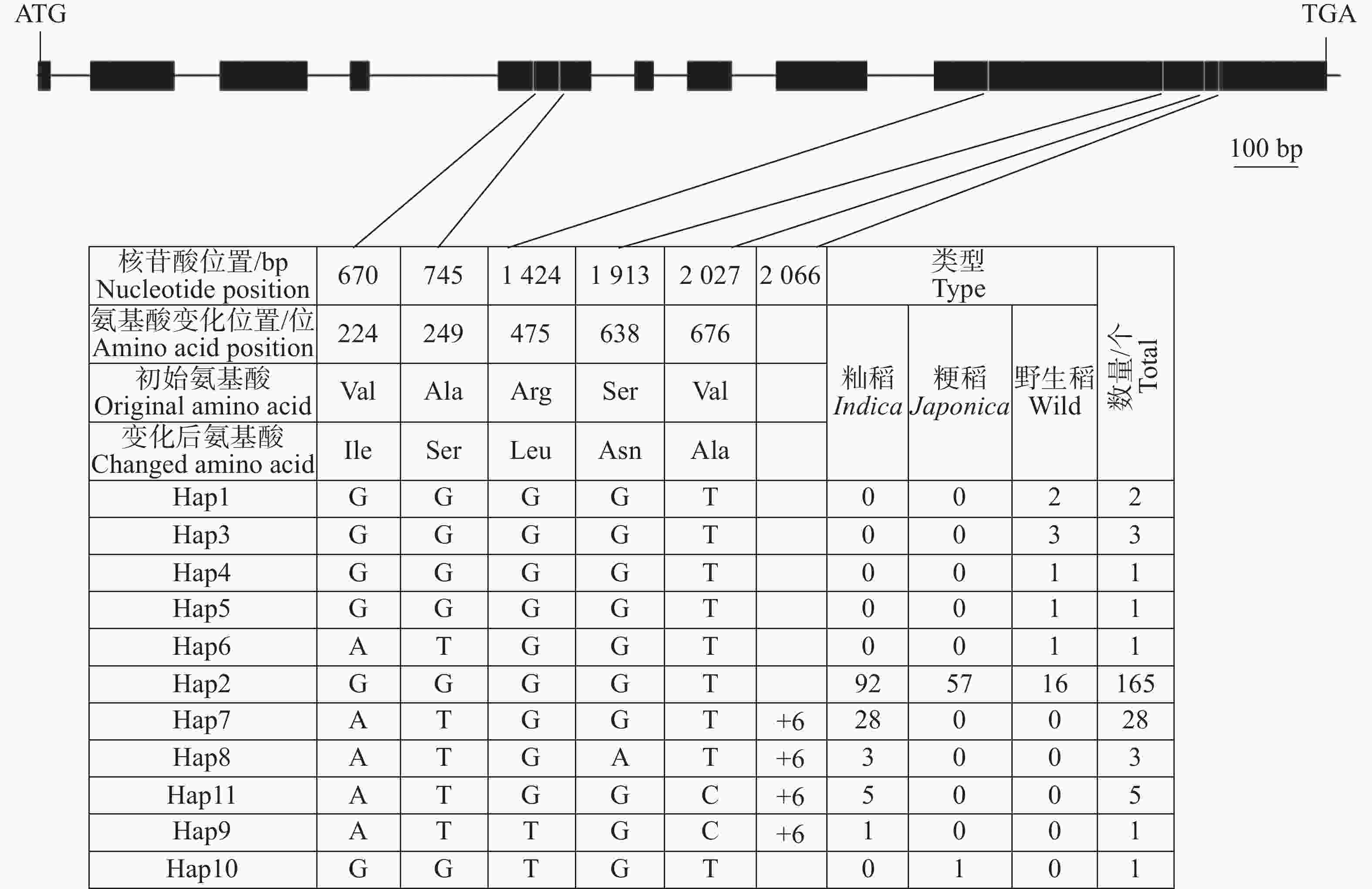
图 4 OsHAK9 基因编码区核苷酸多样性
Figure 4. Nucleotide diversity of the OsHAK9 gene coding sequence
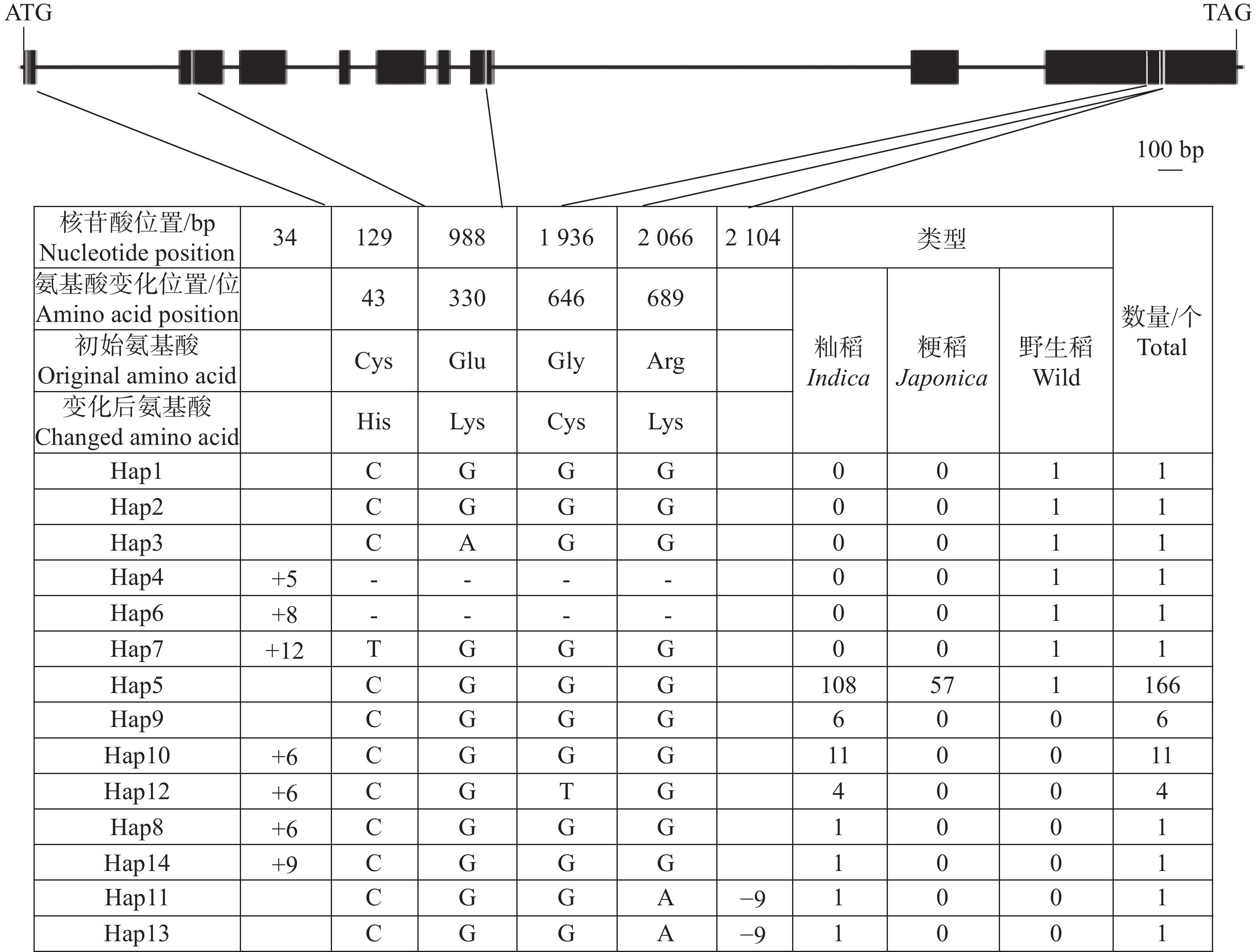
OsHAK13(LOC_Os06g45940)基因在17份野生稻材料中存在缺失。OsHAK13基因全长7 530 bp,编码区长度为2 337 bp。对197份水稻种质基因组序列分析结果显示,OsHAK13编码区存在6处核苷酸变异,包括2个InDel和4个SNP变异。其中,2个InDel分别为编码区第34 bp 存在5~12个碱基的片段插入以及第
2104 bp存在9个碱基的缺失;4个SNP位点引起了氨基酸的变化,分别位于以下位置:第43位(Cys/His)、第330位(Glu/Lys)、第646位(Gly/Cys)和第689位(Arg/Lys)(图5)。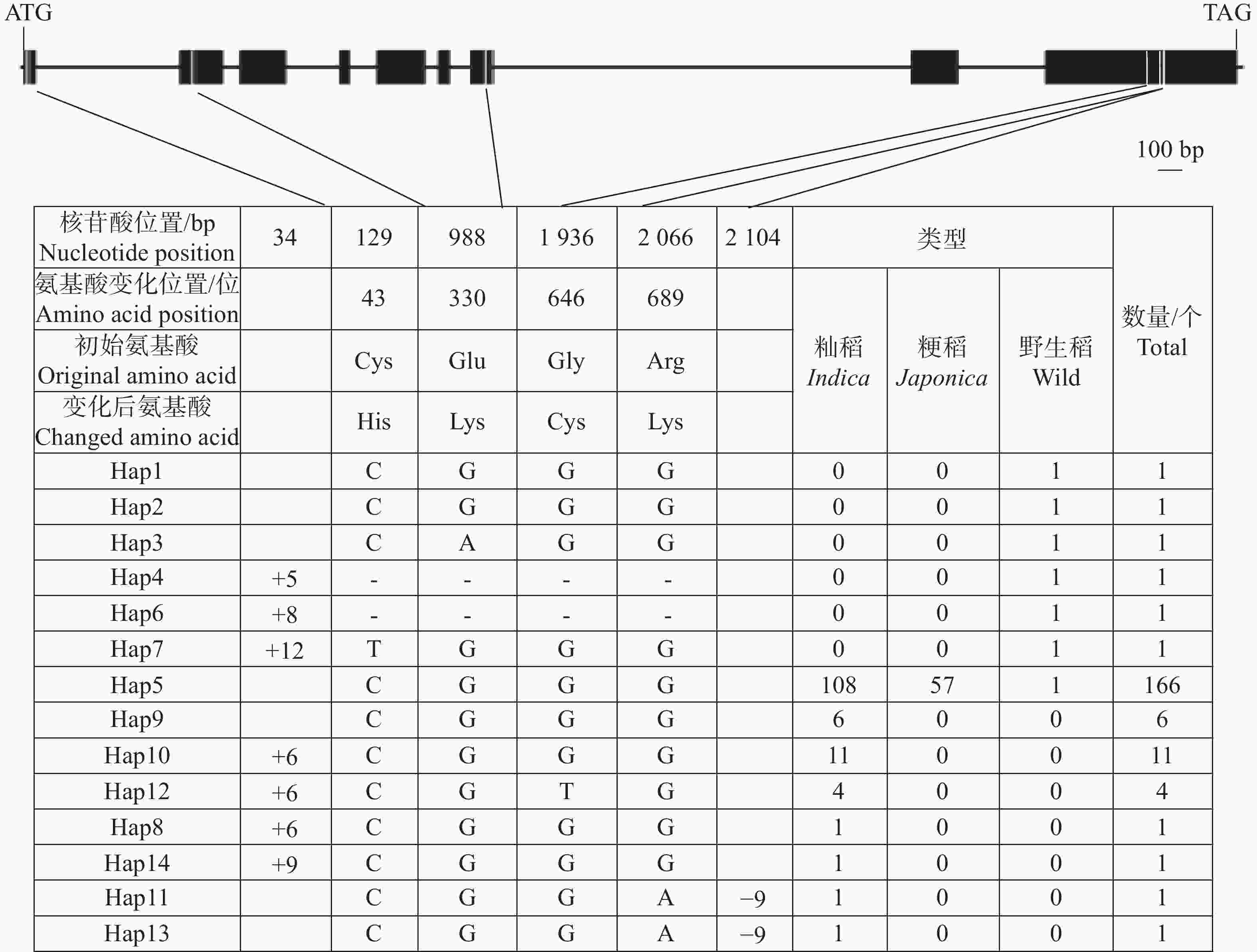
图 5 OsHAK13基因编码区核苷酸多样性
Figure 5. Nucleotide diversity of the OsHAK13 gene coding sequence
-
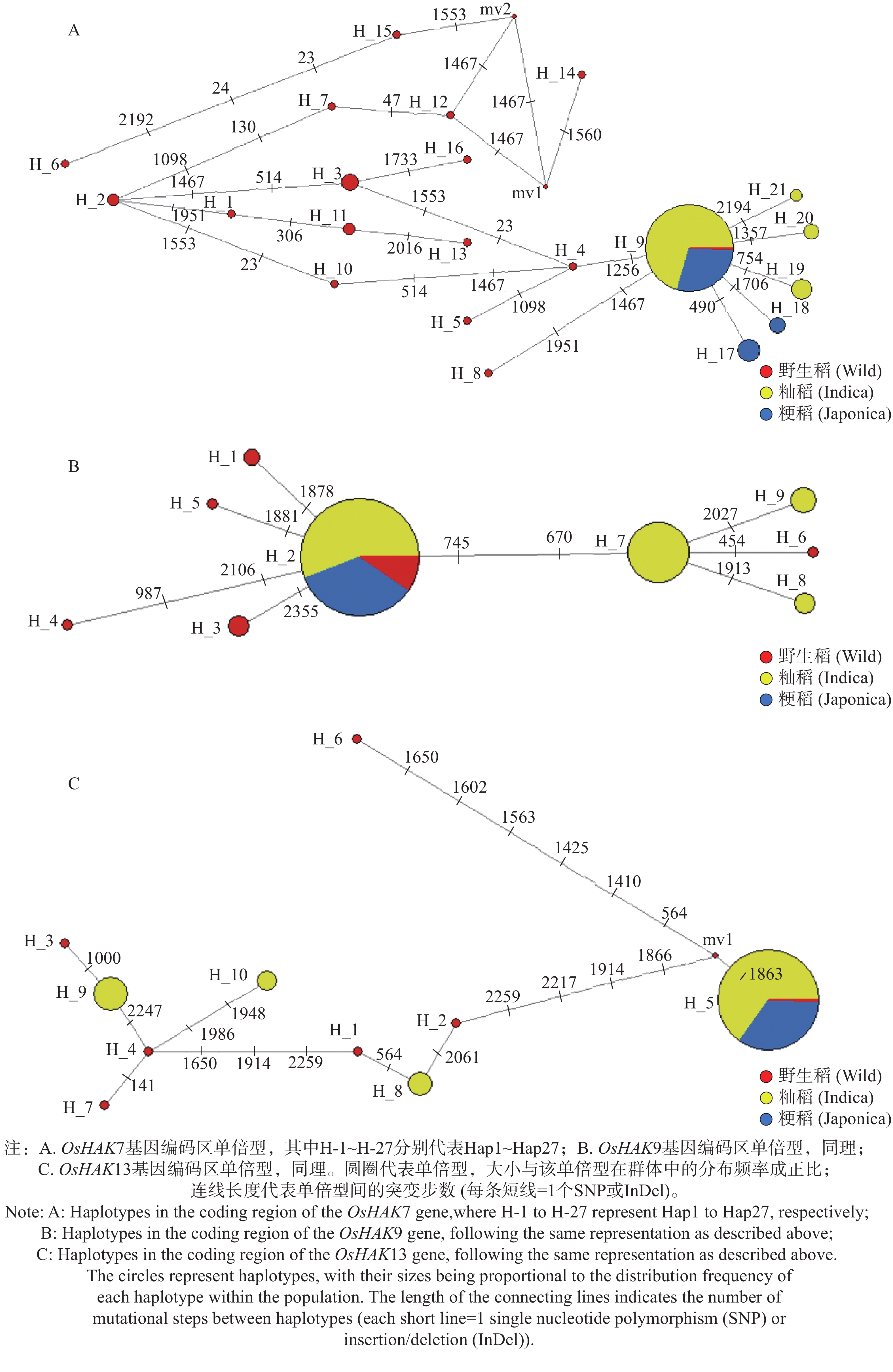
根据上述OsHAK7、OsHAK9和OsHAK13基因序列变异分析的结果,利用各基因编码区的序列变异作为多态性标记,分别构建3个基因的单倍型网络。为避免低频单倍型对网络拓扑结构的干扰,本研究仅保留样本出现频率≥2的主效单倍型用于单倍型网络构建,以便更清晰地解析核心单倍型间的进化关系与连锁特征。在21份野生稻中,OsHAK7基因存在16种单倍型(Hap1~Hap16);在132份籼稻和58份粳稻中,共有12种单倍型(Hap9,Hap17~Hap27),主要为Hap9,在籼稻和粳稻中分别占比87.9%、81.0%。单倍型分析结果显示栽培稻中的OsHAK7基因均来自野生稻中的Hap9或由野生稻中的Hap9突变而来(图6-A)。在24份野生稻中,OsHAK9基因存在6种单倍型(Hap1~Hap6);在129份籼稻和58份粳稻中,共有6种单倍型(Hap2,Hap7~Hap11),主要为Hap2,在籼稻和粳稻中分别占比71.3%、98.3%。单倍型分析结果显示粳稻中的OsHAK9基因均来自野生稻中的Hap2,籼稻中的OsHAK9基因源自野生稻中的Hap2单倍型或由其突变而来(图6-B)。在7份野生稻中,OsHAK13基因存在7种单倍型(Hap1~Hap7);在133份籼稻和57份粳稻中,共有8种单倍型(Hap5、Hap8~Hap14),主要为Hap5,在籼稻和粳稻中分别占比81.2%和100%。单倍型分析结果显示粳稻中的OsHAK13基因均为野生稻中的Hap5类型,籼稻中的OsHAK13基因源自野生稻中的Hap5类型或由其突变而来(图6-C)。
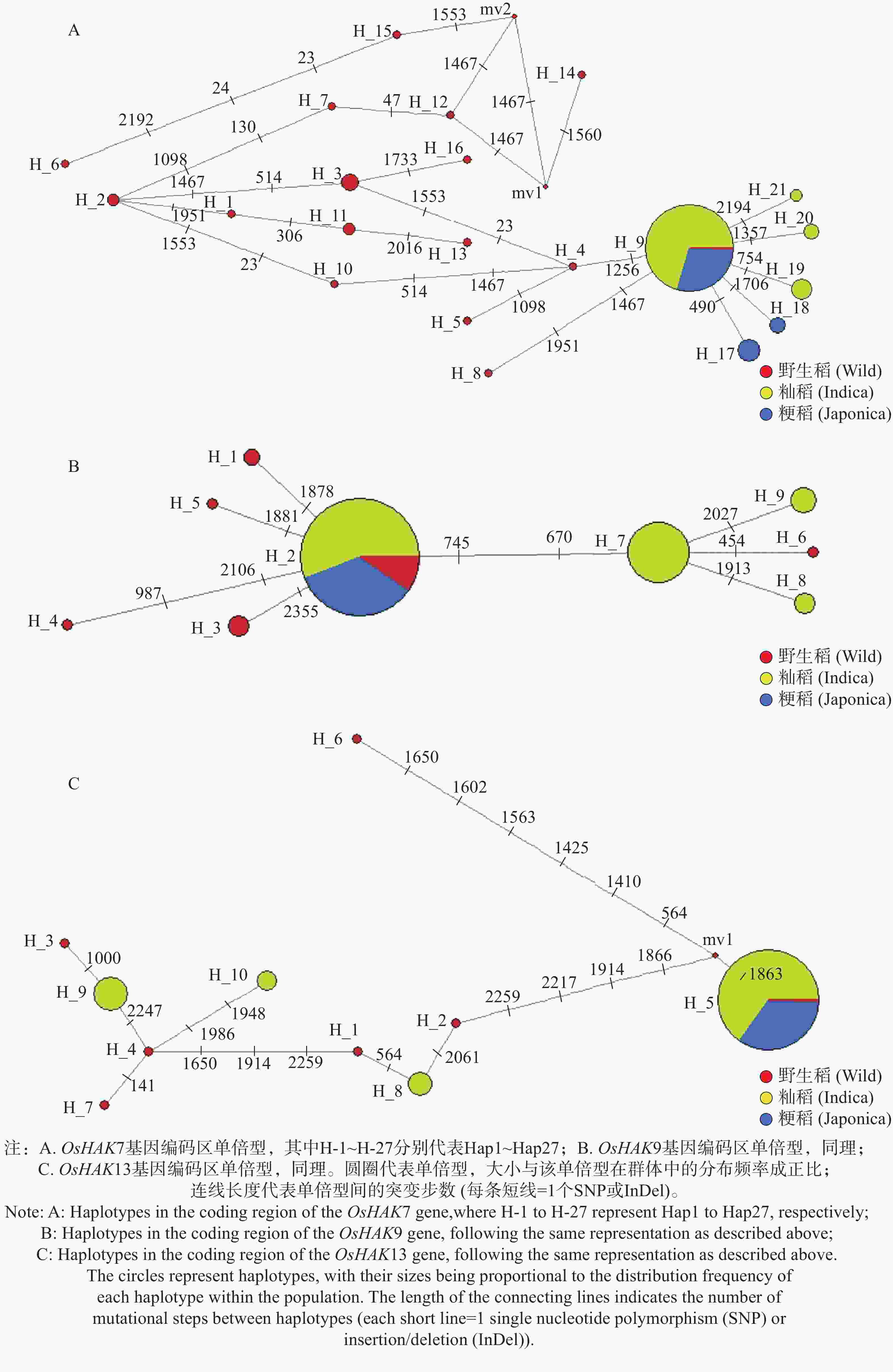
图 6 基因编码区单倍型
Figure 6. gene coding region haplotypes
-
上述结果表明OsHAK7、OsHAK9和OsHAK13在驯化过程中受到不同程度的定向选择。为了进一步探索这3个基因的分子演化规律,本研究对9YJ-12(OsHAK7)、9YJ-100(OsHAK9)、9YJ-97(OsHAK13)、‘9311’(OsHAK7、OsHAK9和OsHAK13)及Nipponbare(OsHAK7、OsHAK9和OsHAK13)中这3个基因的编码区进行了克隆测序。测序结果显示OsHAK7基因编码区在9YJ-12、‘9311’和Nipponbare中无差异;OsHAK9基因编码区在‘9311’和Nipponbare中无差异,9YJ-100中第474 bp插入1个碱基,导致移码突变;OsHAK13基因编码区在9YJ-97、‘9311’和Nipponbare中无差异。以上结果表明,野生稻的这3个基因在编码区检测到的自然变异,尚不足以全面解释其在驯化过程中的受选择信号及耐盐性差异。为了验证这一推测,本研究根据前人的报道查询水稻基因组选择性清除区域发现栽培稻在其第7号染色体物理位置28.8 Mb~29.0 Mb(国际水稻基因组测序计划第4版,IRGSP 4)区间、第6号染色体物理位置28.5 Mb~28.7 Mb(IRGSP 4)区间均存在选择性清除区域,与其关联的性状均为粒长/粒重(表8) [26]。而OsHAK7和OsHAK9基因位于第7号染色体物理位置28.8 Mb~29.0 Mb(IRGSP 4)选择性清除区域内或边缘,OsHAK13基因则位于6号染色体物理位置28.6 Mb~28.7 Mb(IRGSP 4)选择性清除区域内(表8)。鉴于上述结果,推测这3个基因可能位于基因组中的选择性清除区域,因为“搭车效应”而被定向选择。
表 8 OsHAK7、OsHAK9和OsHAK13基因所在选择性清除区域
Table 8. Genomic regions of selective sweep containing OsHAK7, OsHAK9 and OsHAK13 genes
基因名称
Gene基因符号
Gene IDMUS7物理位置/Mb
Physical position
of MUS7IRGSP4物理位置/Mb
Physical position
in IRGSP 4栽培稻选择性
清除区域/Mb
Selective sweep
areas in cultivated rice峰值位置/Mb
Peak position关联性状
Associated traitsOsHAK7 LOC_Os07g47350 Chr7: 28312107 ..28317670 Chr7: 28972267 ..28977790 28.8~29.0 28.9 粒长 OsHAK9 LOC_Os07g48130 Chr7: 28741517 ..28747059 Chr7: 29407218 ..29401677 28.8~29.0 28.9 粒长 OsHAK13 LOC_Os06g45940 Chr6: 27819963 ..27827493 Chr6: 28696966 ..28704495 28.5~28.7 28.5 粒质量/粒长 -
本研究揭示了HAK家族OsHAK7、OsHAK9与OsHAK13基因的序列变异特征及其与水稻耐盐性的关系,为解析这些钾转运蛋白编码基因在水稻驯化过程中的分子演化规律提供了新线索。群体遗传分析显示,OsHAK7、OsHAK9及OsHAK13基因在野生稻中保持了丰富的等位变异与较高的遗传多样性,而在栽培稻中,其序列多态性水平显著降低,π值下降达1~2个数量级。粳稻亚种中的核苷酸多样性下降尤为突出,其π值(0.000 14~0.000 27)不仅显著低于籼稻,也远低于也远低于水稻全基因组背景的平均多态性水平(0.000 50~0.001 70) [26]。这种遗传多样性的急剧下降与驯化过程中的定向选择密切相关 [27]。结合前文单倍型网络分析中栽培稻高度富集于单一或少数核心单倍型的分布特征,可以推断这3个基因在驯化过程中经历了选择性清除(selective sweep) [28]。
人们在水稻驯化过程中对高产(如粒长、粒宽、千粒重及穗粒数)等重要农艺性状的强烈选择,促使关键的驯化基因在群体中迅速固定。由于连锁不平衡(linkage disequilibrium,LD)的作用,位于这些驯化基因附近同一染色体区块的OsHAK7、OsHAK9和OsHAK13基因,可能通过“搭车效应”而被间接固定。这种伴随选择性清除的过程,降低了周边的核苷酸多样性,导致野生稻中一些与耐逆相关的有利等位基因在长期的育种过程中被无意丢失,加剧了栽培稻遗传基础的狭窄化。进一步的基因组分析表明,OsHAK7和OsHAK9基因分别位于已知驯化基因FZP上游约12.5 kb与441.9 kb处。FZP基因是穗枝梗发育的关键调控因子,在驯化过程中受到强烈选择 [29]。由于连锁不平衡的作用,对FZP基因的选择导致其周边大片段基因组区域的多样性被“拖带”固定。对于OsHAK13基因,序列分析表明其栽培稻优势单倍型(Hap5)与野生稻单倍型在编码区未出现导致蛋白质结构显著变化的非同义突变,结合该基因在盐胁迫下呈现的强烈转录响应特征,其在栽培稻中被高度保留的机制可能存在两种情况:其一是该基因在调控钾离子稳态或参与其他胁迫响应通路时,因基因多效性间接影响了产量等农艺性状,从而在驯化中被保留。其二是该基因恰好与某个尚未鉴定的受到强烈选择的驯化基因紧密连锁,从而在遗传重组中通过“搭车效应”被间接固定。
在云南元江普通野生稻中,本研究检测到OsHAK9基因编码区第474 bp处存在单碱基(1 bp)插入。而OsHAK7和OsHAK13在元江野生稻与栽培稻‘9311’之间仅检测到同义突变。虽然同义突变也可能通过改变mRNA稳定性、翻译效率或蛋白质空间结构间接影响耐盐性,但仅凭上述单一变异,难以完全解释携带这些野生稻来源区段的渗入系在盐胁迫下表现出的显著耐盐性提升。进一步印证了本研究利用染色体片段渗入系所评估的耐盐性,反映的是导入的大片段染色体区段的综合遗传效应,而非单一突变位点的直接遗传效应。受限于群体内的重组频率与连锁累赘(linkage drag),野生稻导入片段不可避免地包含了共渗入(co-introgression)区段内尚未鉴定的顺式调控元件,或是与该区段紧密连锁的其他未知耐盐相关基因的协同互作。基于钾转运蛋白保守功能与前期转录响应特征,本研究认为OsHAK7、OsHAK9和OsHAK13基因所在区间可能在水稻驯化过程中受到了定向选择,但仅凭渗入系表型无法排除旁侧连锁基因的干扰。未来亟需构建次级分离群体以缩减候选物理区间,并结合CRISPR/Cas9靶向敲除与转基因互补等反向遗传学手段,最终剔除紧密连锁基因的遗传干扰,实现对野生稻优异耐盐等位基因功能的精准验证。
为客观解析野生稻染色体片段的耐盐遗传效应,本研究选取水稻苗期存活率作为核心鉴定指标。苗期是水稻抵御渗透胁迫与离子毒害最敏感的生长阶段,该时期耐盐性鉴定结果,可直观反映植株维持体内离子稳态、构建早期抗氧化防御体系的综合能力 [30-31]。在 150 mmol·L-1 NaCl 盐胁迫条件下,渗入系 9YJ-12、9YJ-97 与 9YJ-100 的耐盐能力均显著提升。由此说明,野生稻导入的基因组片段携带优良耐盐遗传因子,可通过调控离子平衡、提升活性氧清除效率、强化渗透调节等生理途径,弥补栽培稻进化过程中丢失的耐盐等位基因功能,进而重塑植株盐胁迫应答的遗传调控通路。值得注意的是,作物盐胁迫应答机制贯穿整个生长周期。受试验周期限制,本研究仅开展了苗期耐盐性分析,尚未针对抽穗、开花等生殖生长阶段进行系统性耐盐评价。后续可通过设置生殖生长期盐胁迫处理试验,结合结实率、产量等农艺性状测定,进一步探究水稻不同生育时期耐盐调控机制的差异,明确 OsHAK13 基因在生殖生长阶段的具体生物学功能。
综合而言,本研究系统阐明了水稻 HAK 家族基因在驯化进程中的分子演化特征,提出了“搭车效应”可能导致抗逆基因多样性丧失的演化机制;同时借助野生稻渗入系表型鉴定,证实野生稻种质中含有可显著提升栽培稻耐盐能力的优良等位基因与关键遗传区段。该研究结果不仅丰富了水稻耐盐分子演化的理论认知,也为耐盐水稻分子设计育种、优异耐盐种质挖掘与新品种创制提供了重要理论支撑和基因资源。
水稻驯化中HAK家族基因的序列变异与耐盐单倍型挖掘
DOI: 10.15886/j.cnki.rdswxb.20260052
 CSTR: 32425.14.j.cnki.rdswxb.20260052
CSTR: 32425.14.j.cnki.rdswxb.20260052
Sequence variation and salt-tolerant haplotype mining of HAK family genes during rice domestication
-
摘要: 为了揭示钾离子转运蛋白(HAK)家族基因在水稻(Oryza sativa)驯化过程中的序列变异及其对耐盐性的影响,本研究基于水稻超级泛基因组信息资源数据库中的基因组数据,对HAK家族基因的序列变异进行了分析,并对栽培稻‘9311’及其与云南元江普通野生稻所构建的渗入系进行了耐盐性鉴定。结果表明,OsHAK7、OsHAK9和OsHAK13基因所在区间可能在水稻驯化过程中受到了定向选择;分别携带野生稻来源的OsHAK7、OsHAK9和OsHAK13基因的渗入系,其耐盐性均显著强于‘9311’。进一步分析发现,这3个基因可能因“搭车效应”在栽培稻中被固定。本研究为利用野生稻优异等位基因进行耐盐水稻品种的分子设计育种提供了新的遗传材料和基因资源,对盐碱地水稻高效生产具有较好的应用参考价值。Abstract: To uncover the sequence variations of high-affinity potassium (K+) transporter (HAK) family genes during rice (Oryza sativa) domestication and their impacts on salt tolerance, an analysis was made of the genomic data from the Rice Super Pan-genome Information Resource Database (Rice Super Pan-genome Information Resource Database). Additionally, salt tolerance evaluations were conducted on the cultivated rice variety '9311' and its introgression lines (ILs) developed with Yuanjiang common wild rice from Yunnan. The results indicated that the genomic intervals harboring OsHAK7, OsHAK9, and OsHAK13 genes were likely subjected to directional selection during rice domestication. Notably, the introgression lines carrying the wild rice-derived alleles of OsHAK7, OsHAK9, and OsHAK13 exhibited significantly enhanced salt tolerance compared to '9311'. Further analysis revealed that these three genes might have been fixed in cultivated rice through the "hitchhiking effect". These findings provide novel genetic materials and gene resources for the molecular design-based breeding of salt-tolerant rice varieties by leveraging elite alleles from wild rice, offering significant reference for the efficient production of rice in saline-alkali lands.
-
Key words:
- Rice domestication /
- OsHAK7 /
- OsHAK9 /
- OsHAK13 /
- sequence variation /
- salt-tolerant
-
图 2 ‘9311’与核心渗入系耐盐性表型分析
注:A(a~c).营养液正常生长12 d的幼苗表型;(d~f).150 mmol·L−1 NaCl胁迫处理14 d后的受害表型;(g~i).与盐处理同期的正常营养液对照组表型;(j~l).盐胁迫结束后恢复营养液生长7 d的表型;(m~o).与恢复期同期的正常营养液对照组表型。图中标尺为6 cm。
Fig. 2 Phenotypic analysis of salt tolerance of ‘9311’ and the core introgression lines
Note: A (a−c)represent the seedling phenotypes of plants growing normally in nutrient solution for 12 days; A (d−f)show the damage phenotypes after 14 days of treatment with 150 mM NaCl stress; A (g−i)display the phenotypes of the control group growing in the normal nutrient solution during the same period as the salt treatment; A (j−l)illustrate the phenotypes after 7 days of recovery growth in the nutrient solution following salt stress; A (m−o)depict the phenotypes of the control group growing in the normal nutrient solution during the same period as the recovery phase. The scale bar in the figure represents 6 cm.
图 5 OsHAK13基因编码区核苷酸多样性
Fig. 5 Nucleotide diversity of the OsHAK13 gene coding sequence
图 6 基因编码区单倍型
注:A. OsHAK7基因编码区单倍型,其中H-1~H-27分别代表Hap1~Hap27;B. OsHAK9基因编码区单倍型,同理;C. OsHAK13基因编码区单倍型,同理。圆圈代表单倍型,大小与该单倍型在群体中的分布频率成正比;连线长度代表单倍型间的突变步数(每条短线=1个SNP或InDel)。
Fig. 6 gene coding region haplotypes
Note: A: Haplotypes in the coding region of the OsHAK7 gene,where H-1 to H-27 represent Hap1 to Hap27, respectively; B: Haplotypes in the coding region of the OsHAK9 gene, following the same representation as described above; C: Haplotypes in the coding region of the OsHAK13 gene, following the same representation as described above. The circles represent haplotypes, with their sizes being proportional to the distribution frequency of each haplotype within the population. The length of the connecting lines indicates the number of mutational steps between haplotypes (each short line = 1 single nucleotide polymorphism (SNP)or insertion/deletion (InDel)).
表 1 HAK基因序列分析所用种质资源构成
Table 1 Composition of germplasm for HAK gene sequence analysis
基因名称
Gene name野生稻/份
Wild籼稻/份
Indica粳稻/份
Japonica总计/份
TotalOsHAK7 21 132 58 211 OsHAK9 24 129 58 211 OsHAK13 24 133 57 214  下载: 导出CSV
下载: 导出CSV
表 2 OsHAK7、OsHAK9、OsHAK13基因分段设计覆盖各基因CDS的特异性引物
Table 2 Specific primers designed in segments to cover the entire CDS of OsHAK7, OsHAK9 and OsHAK13 genes
引物名称
Primer正向引物序列(5′−3′)
Forward primer sequence(5′−3′)反向引物序列(3′−5′)
Reverse primer sequence (3′−5′)OsHAK7-1F/R AAATTTGCGTGATCTATAGT TAACTAGACAGAACAATGCA OsHAK7-2F/R TGCATTGTTCTGTCTAGTTA TAGTAATGGCTGCTGGAATA OsHAK7-3F/R TATTCCAGCAGCCATTACTA CCGCTGCAAGAGTTGCAATG OsHAK7-4F/R GTAATCTTCATAACTCCAAG CAGATAAGAGAGCTATCATC OsHAK7-5F/R GATGATAGCTCTCTTATCTG GCACCGGTACATACGGTATT OsHAK7-6F/R TGCCTTCCATCTTCTCGCAC GGCAACTACATTACATGTAT OsHAK9-1F/R TTCCCAGGTGCGTCCTCTAT TGTAAGCAACTCTTTTCTCC OsHAK9-2F/R ATTGGAACCTGTATGGTCAT GCTAACTTCATCCAATAATC OsHAK9-3F/R TGCGTATGCTGAGATAAAGT TCACATTCTAAGCAACGCAA OsHAK9-4F/R CCATTACCATCCTGTCAGCT TACCGCACGATGCACCGGTA OsHAK9-5F/R TCTACACCGACCTCACCTCC TCGCCGTCGCCGGTTTCATC OsHAK13-1F/R AAGGCAGCAAGAATTCAGTA AGAATCCGCAGTAACACTGC OsHAK13-2F/R GAATACTATCTTGTTGCCTA CCAATATATCAGCTGCTTAC OsHAK13-3F/R CGTATATGATAGAGTAAGGA CTATCACCAATTCACCACTG OsHAK13-4F/R CAGTGGTGAATTGGTGATAG AATCAATGTACTCTCTGTTG OsHAK13-5F/R CTAATGCAGCAATTGAAGTT ATATCCGGTATCAGAAGGTA OsHAK13-6F/R TGGTCCGTAAGATATATCAA GACCGACATGGCCTCGTCCT OsHAK13-7F/R GCCTGTTCCGGTGCATCGTC CTACACTCTCTTCAGCGATG
下载: 导出CSV
表 3 HAK家族核心候选基因OsHAK7、OsHAK9、OsHAK13的核苷酸多样性
Table 3 Nucleotide diversity of the core candidate genes OsHAK7, OsHAK9, and OsHAK13 in the HAK family
Locus
基因号Gene
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻π
核苷酸
多态性Tajima’s D π
核苷酸
多态性π ratio
(Cultivar/Wild)π比值(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_
Os07g47350OsHAK7 0.004 5 −0.510 59 P > 0.10 0.000 2 0.042 0 −2.795 89 ***, P < 0.001 0.000 2 0.033 2 −2.586 5 ***, P < 0.001 0.000 3 0.059 7 2.79826 ***,
P < 0.001LOC_
Os07g48130OsHAK9 0.003 8 −1.304 49 P > 0.10 0.001 9 0.502 7 −0.540 56
P > 0.100.002 4 0.633 0 0.408 81
P > 0.100.000 3 0.071 8 − 2.46569 **,
P < 0.01LOC_
Os06g45940OsHAK13 0.007 5 −0.769 11 P > 0.10 0.000 9 0.118 1 − 1.70986 0.10 > P > 0.050.001 2 0.162 4 −1.355 43
P > 0.100.000 1 0.018 8 − 2.43112 **,
P < 0.01
下载: 导出CSV
表 7 HAK亚家族Ⅳ的核苷酸多态性
Table 7 Nucleotide polymorphism of HAK subfamily Ⅳ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os07g01214 OsHAK22 0.004 3 − 1.24745
P > 0.100.004 6 1.070 1 2.64627 *,
P < 0.050.003 1 0.731 3 0.7771
P > 0.100.002 7 0.621 5 3.16076 **,
P < 0.01Clade IV LOC_Os09g21000 OsHAK23 0.004 8 − 0.48915
P > 0.100.005 3 1.100 2 − 2.65601 ***,
P < 0.0010.003 8 0.782 9 − 2.91589 ***,
P < 0.0010.000 4 0.089 8 − 2.7639 ***,
P < 0.001Clade IV LOC_Os06g15910 OsHAK24 0.005 4 − 1.87643 *,
P < 0.050.003 7 0.686 9 0.94237
P > 0.100.003 2 0.594 8 1.58062
P > 0.100.002 5 0.464 1 − 0.04634
P > 0.10Clade IV LOC_Os02g49760 OsHAK25 0.004 0 − 1.29599
P > 0.100.003 0 0.738 8 1.46783
P > 0.100.002 4 0.589 6 0.66543
P > 0.100.000 2 0.047 3 − 1.02112
P > 0.10Clade IV LOC_Os08g39950 OsHAK26 0.010 9 0.28084
P > 0.100.001 6 0.145 1 − 0.46248
P > 0.100.001 6 0.150 6 − 0.46345
P > 0.100.000 2 0.013 8 − 1.64706 0.10 > P > 0.05Clade IV
下载: 导出CSV
表 4 HAK 亚家族Ⅰ的核苷酸多态性
Table 4 Nucleotide Polymorphism of HAK Subfamily Ⅰ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化
分支π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Cultivar/
Wild)
π比值
(栽培稻/
野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/
Wild)
π比值
(籼稻/
野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/
Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_
Os07g01214OsHAK22 0.004 3 − 1.24745 P > 0.100.004 6 1.070 1 2.64627 *,
P < 0.050.003 1 0.731 3 0.7771
P > 0.100.002 7 0.621 5 3.16076 **,
P < 0.01Clade IV LOC_
Os09g21000OsHAK23 0.004 8 − 0.48915 P > 0.100.005 3 1.100 2 − 2.65601 ***,
P < 0.0010.003 8 0.782 9 − 2.91589 ***,
P < 0.0010.000 4 0.089 8 − 2.7639 ***, P < 0.001Clade IV LOC_
Os06g15910OsHAK24 0.005 4 − 1.87643 *,
P < 0.050.003 7 0.686 9 0.94237
P > 0.100.003 2 0.594 8 1.58062
P > 0.100.002 5 0.464 1 − 0.04634
P > 0.10Clade IV LOC_
Os02g49760OsHAK25 0.004 0 − 1.29599 P > 0.100.003 0 0.738 8 1.46783
P > 0.100.002 4 0.589 6 0.66543
P > 0.100.000 2 0.047 3 − 1.02112 P > 0.10Clade IV LOC_
Os08g39950OsHAK26 0.010 9 0.28084 P > 0.100.001 6 0.145 1 − 0.46248
P > 0.100.001 6 0.150 6 − 0.46345
P > 0.100.000 2 0.013 8 − 1.64706 0.10 > P > 0.05Clade IV
下载: 导出CSV
表 5 HAK 亚家族Ⅱ的核苷酸多态性
Table 5 Nucleotide Polymorphism of HAK Subfamily Ⅱ
Locus
基因号Gene
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D π
核苷酸
多态性π ratio
(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os04g32920 OsHAK1 0.018 4 0.4915
P > 0.100.003 1 0.170 2 0.63282
P > 0.100.001 5 0.083 2 − 1.33417
P > 0.100.001 7 0.089 7 1.16604
P > 0.10Clade Ia LOC_Os08g36340 OsHAK4 0.002 8 − 1.41127
P > 0.100.003 3 1.187 1 − 0.33119
P > 0.100.003 3 1.197 8 − 0.27482
P > 0.100.003 4 1.233 8 0.11114
P > 0.10Clade Ia LOC_Os01g70940 OsHAK2 0.003 2 − 1.48245
P > 0.100.001 9 0.583 9 1.22065
P > 0.100.000 9 0.291 9 − 0.9904
P > 0.100.000 1 0.043 5 − 0.90566
P > 0.10Clade Ib LOC_Os01g27170 OsHAK3 0.005 2 − 1.06352
P > 0.100.005 8 1.112 2 − 0.24798
P > 0.100.006 0 1.150 9 0.50134
P > 0.100.001 1 0.210 8 − 2.53964 ***,
P < 0.001Clade Ib LOC_Os01g70490 OsHAK5 0.002 2 − 1.24384
P > 0.100.001 5 0.701 8 0.71477
P > 0.100.000 4 0.192 7 − 1.84369 *, P < 0.050.000 5 0.211 0 − 1.95666 *,
P < 0.05Clade Ib LOC_Os01g70660 OsHAK6 0.008 0 − 0.64962
P > 0.100.006 6 0.818 8 3.13463 **,P< 0.010.002 3 0.288 8 − 0.83886
P > 0.100.000 7 0.087 5 − 2.81216 ***,
P < 0.001Clade Ib LOC_Os03g37830 OsHAK27 0.004 9 − 1.28224
P > 0.100.003 8 0.765 3 1.03719
P > 0.100.001 7 0.353 1 − 0.86236
P > 0.100.000 2 0.046 9 − 2.8289 ***,
P < 0.001Clade Ib
下载: 导出CSV
表 6 HAK 亚家族Ⅲ的核苷酸多态性
Table 6 Nucleotide Polymorphism of HAK Subfamily Ⅲ
Locus
基因号Gene Name
基因名称Wild
野生稻Cultivar
栽培稻Indica
籼稻Japonica
粳稻进化分支 π
核苷酸
多态性Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio(Cultivar/Wild)
π比值
(栽培稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Indica/Wild)
π比值
(籼稻/野生稻)Tajima’s D
中性检验
统计量π
核苷酸
多态性π ratio
(Japonica/Wild)
π比值
(粳稻/野生稻)Tajima’s D
中性检验
统计量LOC_Os03g37840 OsHAK16 0.004 5 − 1.55764 P>0.100.002 9 0.634 4 1.65425
P>0.100.000 9 0.198 2 − 1.73911 0.10>P>0.050 0.008 8 − 1.2672 ,
P>0.10Clade III LOC_Os09g27580 OsHAK17 0.004 9 − 1.03831 P>0.100.003 7 0.745 4 0.06718
P>0.100.002 3 0.468 4 − 1.08931 P>0.100.001 1 0.224 0 − 2.32309 **, P < 0.01Clade III LOC_Os09g38960 OsHAK18 0.006 3 − 1.17953 P>0.100.004 5 0.715 0 − 1.08768 P>0.100.002 2 0.355 1 − 2.2962 **,
P < 0.010.000 4 0.065 3 − 2.74023 ***,
P < 0.001Clade III LOC_Os02g31910 OsHAK19 0.003 1 − 0.81272 P>0.100.002 8 0.891 4 1.58937
P>0.100.001 5 0.476 0 − 0.35248 P>0.100.000 6 0.204 5 − 2.04205 *,
P < 0.05Clade III LOC_Os02g31940 OsHAK20 0.006 1 − 1.59181 0.10>P>0.050.003 3 0.544 4 − 1.88162 *, P < 0.050.002 4 0.399 7 − 2.22657 **,
P < 0.010.001 7 0.281 3 − 2.17062 *,
P < 0.05Clade III LOC_Os03g37930 OsHAK21 0.018 4 0.89146 P>0.100.004 6 0.249 7 0.52287
P>0.100.003 2 0.171 0 − 0.61199 P>0.100.000 3 0.017 4 − 2.33498 **, P < 0.01Clade III
下载: 导出CSV
表 8 OsHAK7、OsHAK9和OsHAK13基因所在选择性清除区域
Table 8 Genomic regions of selective sweep containing OsHAK7, OsHAK9 and OsHAK13 genes
基因名称
Gene基因符号
Gene IDMUS7物理位置/Mb
Physical position
of MUS7IRGSP4物理位置/Mb
Physical position
in IRGSP 4栽培稻选择性
清除区域/Mb
Selective sweep
areas in cultivated rice峰值位置/Mb
Peak position关联性状
Associated traitsOsHAK7 LOC_Os07g47350 Chr7: 28312107 ..28317670 Chr7: 28972267 ..28977790 28.8~29.0 28.9 粒长 OsHAK9 LOC_Os07g48130 Chr7: 28741517 ..28747059 Chr7: 29407218 ..29401677 28.8~29.0 28.9 粒长 OsHAK13 LOC_Os06g45940 Chr6: 27819963 ..27827493 Chr6: 28696966 ..28704495 28.5~28.7 28.5 粒质量/粒长
下载: 导出CSV
-
[1] 石荃. 水稻耐盐碱性生理和遗传研究进展分析[J]. 南方农机, 2020, 51(8): 39. https://doi.org/10.3969/j.issn.1672-3872.2020.08.036 doi: 10.3969/j.issn.1672-3872.2020.08.036 [2] Singh A. Soil salinization management for sustainable development: a review [J]. Journal of Environmental Management, 2021, 277: 111383. https://doi.org/10.1016/j.jenvman.2020.111383 doi: 10.1016/j.jenvman.2020.111383 [3] 杨劲松. 中国盐渍土研究的发展历程与展望[J]. 土壤学报, 2008, 45(5): 837−845. https://doi.org/10.3321/j.issn:0564-3929.2008.05.010 doi: 10.3321/j.issn:0564-3929.2008.05.010 [4] 王洋, 张瑞, 刘永昊, 等. 水稻对盐胁迫的响应及耐盐机理研究进展[J]. 中国水稻科学, 2022, 36(2): 105−117. https://doi.org/10.16819/j.1001-7216.2022.210609 doi: 10.16819/j.1001-7216.2022.210609 [5] 刘松瑞, 陈敏, 许昊, 等. 盐胁迫下不同亚种水稻种质资源的根系形态特征分析[J]. 南方农业学报, 2025, 56(11): 3320−3331. https://doi.org/10.3969/j.issn.2095-1191.2025.11.002 doi: 10.3969/j.issn.2095-1191.2025.11.002 [6] Liu C T, Mao B G, Yuan D Y, et al. Salt tolerance in rice: physiological responses and molecular mechanisms [J]. The Crop Journal, 2022, 10(1): 13−25. https://doi.org/10.1016/j.cj.2021.02.010 doi: 10.1016/j.cj.2021.02.010 [7] 姜浩, 杨和生, 王波, 等. 近40年云南普通野生稻优异特性及有利基因的发掘与利用[J]. 生物技术进展, 2024, 14(5): 724−737. https://doi.org/10.19586/j.2095-2341.2024.0119 doi: 10.19586/j.2095-2341.2024.0119 [8] Wang S S, Cao M, Ma X, et al. Integrated RNA sequencing and QTL mapping to identify candidate genes from Oryza rufipogon associated with salt tolerance at the seedling stage [J]. Frontiers in Plant Science, 2017, 8: 1427. https://doi.org/10.3389/fpls.2017.01427 doi: 10.3389/fpls.2017.01427 [9] Tan L B, Li X R, Liu F X, et al. Control of a key transition from prostrate to erect growth in rice domestication [J]. Nature Genetics, 2008, 40(11): 1360−1364. https://doi.org/10.1038/ng.197 doi: 10.1038/ng.197 [10] Zhu Z F, Tan L B, Fu Y C, et al. Genetic control of inflorescence architecture during rice domestication [J]. Nature Communications, 2013, 4: 2200. https://doi.org/10.1038/ncomms3200 doi: 10.1038/ncomms3200 [11] Lin Z W, Griffith M E, Li X R, et al. Origin of seed shattering in rice (Oryza sativa L. ) [J]. Planta, 2007, 226(1): 11−20. https://doi.org/10.1007/s00425-006-0460-4 doi: 10.1007/s00425-006-0460-4 [12] Li C B, Zhou A L, Sang T. Rice domestication by reducing shattering [J]. Science, 2006, 311(5769): 1936−1939. https://doi.org/10.1126/science.1123604 doi: 10.1126/science.1123604 [13] Luo J H, Liu H, Zhou T Y, et al. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice [J]. The Plant Cell, 2013, 25(9): 3360−3376. https://doi.org/10.1105/tpc.113.113589 doi: 10.1105/tpc.113.113589 [14] Hua L, Wang D R, Tan L B, et al. LABA1, a domestication gene associated with long, barbed awns in wild rice [J]. The Plant Cell, 2015, 27(7): 1875−1888. https://doi.org/10.1105/tpc.15.00260 doi: 10.1105/tpc.15.00260 [15] Sugimoto K, Takeuchi Y, Ebana K, et al. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(13): 5792−5797. https://doi.org/10.1073/pnas.0911965107 doi: 10.1073/pnas.0911965107 [16] Sun C Q, Wang X K, Li Z C, et al. Comparison of the genetic diversity of common wild rice (Oryza rufipogon Griff. ) and cultivated rice (O. sativa L. ) using RFLP markers [J]. Theoretical and Applied Genetics, 2001, 102(1): 157−162. https://doi.org/10.1007/s001220051631 doi: 10.1007/s001220051631 [17] Yang Z F, Gao Q S, Sun C S, et al. Molecular evolution and functional divergence of HAK potassium transporter gene family in rice (Oryza sativa L. ) [J]. Journal of Genetics and Genomics, 2009, 36(3): 161−172. https://doi.org/10.1016/S1673-8527(08)60103-4 doi: 10.1016/S1673-8527(08)60103-4 [18] Chen G, Hu Q D, Luo L, et al. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges [J]. Plant, Cell & Environment, 2015, 38(12): 2747−2765. https://doi.org/10.1111/pce.12585 doi: 10.1111/pce.12585 [19] Peng L R, Xiao H J, Li R, et al. Potassium transporter OsHAK18 mediates potassium and sodium circulation and sugar translocation in rice [J]. Plant Physiology, 2023, 193(3): 2003−2020. https://doi.org/10.1111/pce.12585 doi: 10.1111/pce.12585 [20] Shen L K, Fan W X, Li N, et al. Rice potassium transporter OsHAK18 mediates phloem K+ loading and redistribution [J]. The Plant Journal, 2023, 116(1): 201−216. https://doi.org/10.1111/tpj.16371 doi: 10.1111/tpj.16371 [21] Banuelos M A, Garciadeblas B, Cubero B, et al. Inventory and functional characterization of the HAK potassium transporters of rice [J]. Plant Physiology, 2002, 130(2): 784−795. https://doi.org/10.1104/pp.007781 doi: 10.1104/pp.007781 [22] Shen T, Xu F J, Chen D, et al. A B-box transcription factor OsBBX17 regulates saline-alkaline tolerance through the MAPK cascade pathway in rice [J]. New Phytologist, 2024, 241(5): 2158−2175. https://doi.org/10.1111/nph.19480 doi: 10.1111/nph.19480 [23] Zeng P, Xie T, Shen J X, et al. Potassium transporter OsHAK9 regulates seed germination under salt stress by preventing gibberellin degradation through mediating OsGA2ox7 in rice [J]. Journal of Integrative Plant Biology, 2024, 66(4): 731−748. https://doi.org/10.1111/jipb.13642 doi: 10.1111/jipb.13642 [24] Pruthi R, Chapagain S, Coronejo S, et al. Quantitative trait loci, candidate genes, and breeding lines to improve salt tolerance at the flowering and seedling stages in rice [J]. Food and Energy Security, 2023, 12(6): e433. https://doi.org/10.1002/fes3.433 doi: 10.1002/fes3.433 [25] Shang L G, Li X X, He H Y, et al. A super pan-genomic landscape of rice [J]. Cell Research, 2022, 32(10): 878−896. https://doi.org/10.1038/s41422-022-00685-z doi: 10.1038/s41422-022-00685-z [26] Huang X H, Kurata N, Wei X H, et al. A map of rice genome variation reveals the origin of cultivated rice [J]. Nature, 2012, 490(7421): 497−501. https://doi.org/10.1038/nature11532 doi: 10.1038/nature11532 [27] 常晶晶. 水稻驯化对根际微生物的选择及驱动机制研究[D]. 长春: 中国科学院大学(中国科学院东北地理与农业生态研究所), 2022. [28] 杨学勇, 苏汉东, 张梦卓, 等. 多倍化和驯化研究进展与展望[J]. 中国科学: 生命科学, 2021, 51(10): 1457−1466. https://doi.org/10.1360/SSV-2021-0220 doi: 10.1360/SSV-2021-0220 [29] Huang Y Y, Zhao S S, Fu Y C, et al. Variation in the regulatory region of FZP causes increases in secondary inflorescence branching and grain yield in rice domestication [J]. The Plant Journal, 2018, 96(4): 716−733. https://doi.org/10.1111/tpj.14062 doi: 10.1111/tpj.14062 [30] Ismail A M, Horie T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance [J]. Annual Review of Plant Biology, 2017, 68(1): 405−434. https://doi.org/10.1146/annurev-arplant-042916-040936 doi: 10.1146/annurev-arplant-042916-040936 [31] Zeng L H, Shannon M C. Salinity effects on seedling growth and yield components of rice [J]. Crop Science, 2000, 40(4): 996−1003. https://doi.org/10.2135/cropsci2000.404996x doi: 10.2135/cropsci2000.404996x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 228
- HTML全文浏览量: 157
- 被引次数: 0
