

-
水稻(Oryza sativa)是全球半数以上人口的主粮作物,其产量的稳定与增长,是保障粮食安全、维系社会稳定的基石[1]。南繁是中国特有的育种体系,涵盖水稻等多种作物。该体系利用海南冬季特有的光热资源进行加代繁殖,实现一年内多代育种,从而极大缩短育种周期,加快优良品种的选育与推广进程。自20世纪50年代起,中国科学家便赴海南开展南繁育种工作。据统计,此后60余年间,经南繁审定推广的品种超过7 000个,占全国审定品种总数的70%[2]。可见,南繁已成为现代水稻育种不可或缺的“加速器”。
结实率,即成熟籽粒数占总颖花数的百分比,是决定水稻单位面积产量的核心因子,在南繁育种中其意义尤为突出。南繁季节的重要功能之一,是为全国不同生态区快速创制与筛选育种材料。若因遭遇气象灾害导致结实率不稳,不仅直接影响当季加代种子的产量与质量,还可能干扰对高产、稳产、耐逆等关键育种目标的准确评价,进而延误整个育种计划。水稻结实率的高低,取决于从穗分化到籽粒灌浆等一系列连续发育事件,包括穗原基分化、颖花分化、花粉母细胞减数分裂、花粉充实与成熟、开花授粉、籽粒灌浆与成熟等关键阶段[3]。在外界环境胁迫下,任一发育环节受阻,均可能导致颖花退化、受精失败或籽粒秕瘦,直接表现为结实率下降。在诸多环境因子中,气温是调控水稻各生殖发育阶段、进而影响结实率的最关键胁迫因子。研究表明,水稻生殖生长对温度胁迫存在明确的敏感窗口期与临界阈值。高温胁迫的危害主要集中在扬花期与灌浆初期:扬花期遭遇超过35°C的高温,会严重抑制花药开裂,降低花粉活力,导致授粉受精失败,空粒率急剧增加[4];灌浆初期的高温则会加速灌浆进程,但显著缩短灌浆持续期,同时削弱叶片光合功能与同化物转运效率,导致籽粒充实不良,秕粒增加。低温胁迫则主要威胁减数分裂期与扬花期,可诱导花粉母细胞异常,抑制花药开裂,同样严重阻碍受精过程[5]。
现有研究多阐明了高温与低温对水稻结实率的影响,但缺乏量化预测与描述。本研究拟系统解析不同水稻种质资源结实率对抽穗前后关键时段气温变化的量化响应模式。因此,本研究以涵盖主要遗传类型的水稻微核心种质为试验材料,于2009年11月至2010年4月的典型南繁季节,在海南三亚开展田间种植,考察其抽穗期与结实率;同时记录每日最高气温、最低气温及平均气温。通过整合气象数据与表型数据,分析气温影响南繁水稻结实率的关键发育时期,以期为提升南繁育种的防灾减灾能力与育种效率提供科学指导。
-
供试材料为水稻微核心种质[6]。为研究不同遗传背景类型对气温的响应模式,本研究采用两种分类方式对稻种资源进行分组。第一种方式参考水稻3k基因组分类[7],取每组至少5份材料的分组作为有效分组,共得到有效材料219份,划分为14个组,包括:华南籼(XI-SC)、云贵籼(XI-YG)、改良籼1群(XI-IMP1)、华中籼1群(XI-CC1)、东亚温带粳(GJ-Te-EA)、华中籼2群(XI-CC2)、南亚籼(XI-SA)、华北粳(GJ-NC)、改良籼2群(XI-IMP2)、混合群(admix)、温带粳(GJ-Te)、温带改良粳1群(GJ-Te-IMP1)和热带粳(GJ-Tr(3k))(表1)。第二种方式参考王学强等[8]的分类,共获得有效材料141份,划分为11个组,包括:典型地方籼(XI-LAN-T)、混合地方籼(XI-LAN-M)、地方及初级改良混合籼(XI-LAN-PIMP-M)、典型近期改良籼(XI-RIMP-T)、典型地方及初级改良籼(XI-LAN-PIMP-T)、混合近期改良籼(XI-RIMP-M)、地方温带非糯粳(GJ-Te-LAN-NG)、地方及初级改良温带粳(GJ-Te-LAN-PIMP)、地方温带糯粳(GJ-Te-LAN-GL)、混合近期改良温带粳(GJ-Te-RIMP-M)和热带粳(GJ-Tr)(表1)。
表 1 不同类型水稻品种抽穗期及结实率变异情况
Table 1. Variability of heading date and seed setting rate of different types of rice varieties
品种类型
Variety types份数
Num.抽穗期/d
Heading date结实率/%
Seed setting rate最小值
Min.平均值
Mean最大值
Max.最小值
Min.平均值
Mean最大值
Max.华南籼 XI-SC 24 66 88.4 125 61.5 86.4 95.4 云贵籼 XI-YG 21 70 98.2 123 72.7 87.3 93.6 改良籼1群 XI-IMP1 19 74 103.8 126 29.4 83.9 95.4 华中籼1群 XI-CC1 38 74 95.4 121 66.0 88.3 96.7 东亚温带粳
GJ-Te-EA21 63 88.3 114 50.7 88.2 96.8 华中籼2群 XI-CC2 23 80 97.0 112 71.8 86.6 94.9 南亚籼 XI-SA 6 82 92.2 98 67.5 81.3 91.7 华北粳 GJ-NC 19 65 88.9 106 62.4 83.0 96.7 改良籼2群 XI-IMP2 9 101 112.3 121 64.9 85.0 94.7 混合群 admix 9 77 100.2 122 72.5 85.6 93.0 温带粳 GJ-Te 17 76 97.8 109 72.4 87.9 96.3 温带改良粳1群
GJ-Te-IMP16 79 95.3 103 55.0 86.5 97.3 热带粳 GJ-Tr(3k) 7 108 115.4 127 68.5 81.8 91.7 典型地方籼
XI-LAN-T17 66 85.6 123 61.5 84.3 95.4 混合地方籼
XI-LAN-M7 70 98.6 110 72.7 83.7 89.3 地方及初级改良混合籼
XI-LAN-PIMP-M20 70 99.3 122 73.0 88.3 95.2 典型近期改良籼
XI-RIMP-T10 74 101.7 121 73.7 89.0 95.4 典型地方及初级改良籼
XI-LAN-PIMP-T32 74 93.8 116 71.8 88.3 96.7 混合近期改良籼
XI-RIMP-M5 93 109.2 119 84.6 88.9 93.4 地方温带非糯粳
GJ-Te-LAN-NG9 72 86.8 104 65.9 81.6 94.9 地方及初级改良温带粳
GJ-Te-LAN-PIMP15 74 90.2 114 67.6 89.6 96.8 地方温带糯粳
GJ-Te-LAN-GL11 76 97.8 109 76.7 87.4 94.4 混合近期改良温带粳
GJ-Te-RIMP-M9 77 93.9 106 84.4 89.1 94.3 热带粳 GJ-Tr 6 96 114.0 127 73.0 85.3 91.7 试验于2009年11月至2010年4月的典型南繁季进行,供试水稻微核心种质种植于海南三亚崖州南滨农场。田间试验采用随机区组设计,设2次重复,三行区,每行10株,种植密度为29.5 cm × 16.7 cm,田间管理与一般大田生产相同。播种日期为2009年11月20日,每行取中间5个健康单株,晾干后室内取单株主穗考察结实率,以10株(5株×2个重复)的平均值作为统计单位。
-
水稻在穗发育及扬花期遭遇高温或低温,易导致结实率降低。抽穗前40 d水稻进入穗发育阶段,主要发育阶段包括:抽穗前40~25 d为穗发育起始及枝梗分化与发育期;抽穗前25~12d为雌蕊发育及花粉母细胞形成与减数分裂期;抽穗前12~7 d为花粉内容物充实期;抽穗前7 d至抽穗为花粉稳定期;抽穗后约一周为扬花期;抽穗后20 d内为籽粒胚胎发育与快速灌浆期。
三亚南繁季节正值气温较低时期,水稻从穗发育至抽穗前常因低温导致结实率下降。为此,统计了每个品种在抽穗前40 d至抽穗后20 d的气温数据,包括日最低气温、日平均气温和日最高气温,数据由海南省气象局提供并进行分析。以每个品种的抽穗当日为0 d,抽穗前1 d记为−1,依此类推至抽穗前40 d记为−40;抽穗后1 d记为+1,依此类推至抽穗后20 d记为+20。因此,−40至+20表示抽穗前40 d至抽穗后20 d内的各个发育阶段。提取每个品种对应日期的单日温度(记为M1)、连续3日温度平均值(记为M3)、连续5日温度平均值(记为M5),作为气温的3种统计口径。
据此,抽穗当日的单日温度记为M1-0,抽穗前1至40 d的单日温度依次记为M1−1至M1−40,抽穗后1至20 d的单日温度依次记为M1+1至M1+20。抽穗当天及其前后各1 d的3日温度平均值记为M3−0,抽穗前1 d、前2 d及抽穗当天的3日温度平均值记为M3−1,其余依此类推,记为M3−40至M3+20。连续5日温度平均值的标记方式依此类推,记为M5−40至M5+20。
-
计算每种统计口径气温与结实率的相关系数,并通过Excel的CORREL函数计算其相关的显著性。显著正相关表示结实率随温度降低而降低,表明对冷胁迫敏感,不耐低温;显著负相关则表示结实率随温度升高而降低,表明对热胁迫敏感,不耐高温。统计各个发育阶段不耐低温和不耐高温的品种类型数量,统计各个品种类型不同统计口径不耐低温或不耐高温的发育阶段数量,由此分析不同类型不同发育阶段对温度的敏感性,为南繁水稻防灾提供参考。
-
本研究所分析的水稻品种中,从播种到抽穗的最短抽穗期仅63 d,最长抽穗期达127 d,平均抽穗期为96.7 d。同一类型内不同品种间的抽穗期最少相差16 d,最多相差64 d,平均差异为40.2 d(表1)。在同一天播种的情况下,品种间及类型间抽穗期的巨大差异,使得不同品种在不同发育阶段能够经历丰富多样的温度条件。
各水稻品种在抽穗前40 d至抽穗后20 d的不同生长发育阶段所遇到的气温情况显示,同一发育阶段不同品种经历的最低气温和最高气温均存在较大差异(图1)。254个水稻品种在同一发育阶段的最低气温差异幅度为10.1~12.0℃,平均气温差异幅度为6.1~11.7℃,最高气温差异幅度为7.5~12.3℃。各品种抽穗当天的最低气温为15.3~25.8℃,差异幅度为10.5℃;平均气温为17.0~27.4℃,差异幅度为10.4℃;最高气温为19.6~31.6℃,差异幅度为12.0℃。各品种在抽穗前不同发育阶段所经历的显著温度差异,为解析品种对温度响应机制提供了良好条件。
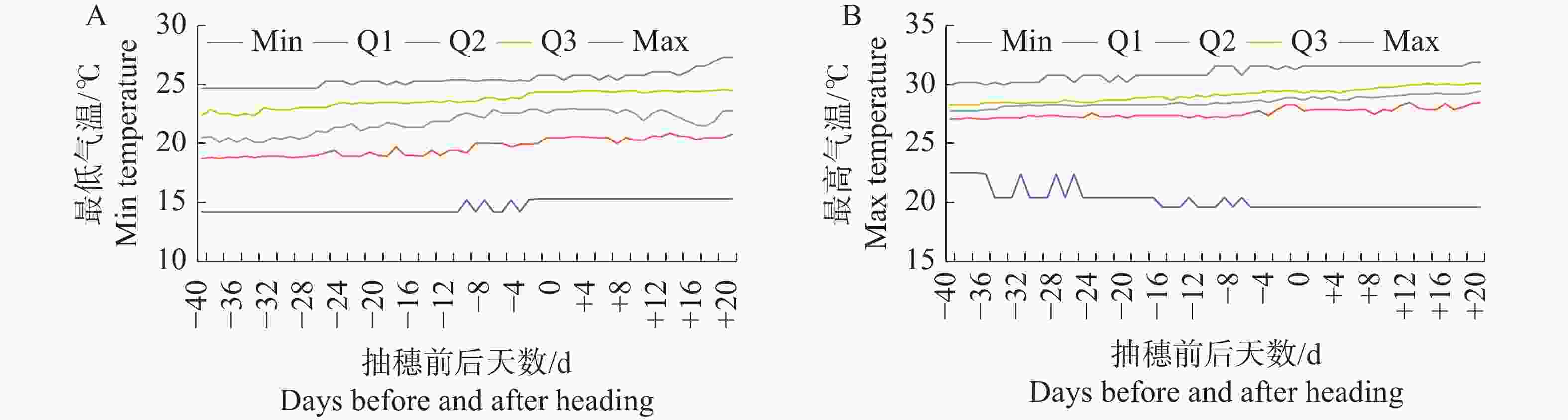
图 1 水稻254个品种抽穗前40 d至抽穗后20 d的最低(A)和最高气温(B)分布图
Figure 1. Distribution of the minimum (A)and maximum temperatures (B)of 254 rice varieties from 40th day before heading to 20th day after heading
各水稻品种的结实率数据显示,最低结实率仅为29.4%,最高结实率达97.3%,平均结实率为86.1%。同一类型内不同品种间的结实率最小相差8.8%,最大相差67.3%,3/4类型中品种间结实率差异高于21.1%,1/4类型中高于33.9%(表1)。上述结果表明,品种间及类型间在抽穗期、不同发育阶段所遇温度条件及结实率方面均存在显著差异,为研究不同品种在不同发育阶段所遇温度条件对结实率的影响奠定了重要基础。
-
总体上,水稻抽穗前40 d至抽穗后20 d这段时间有多个发育阶段受低温或高温胁迫会显著降低结实率;低温胁迫对结实率的影响略高于高温胁迫,前者对较低的最高气温更敏感,而后者对较高的最低气温和平均气温更敏感(图2~图5)。
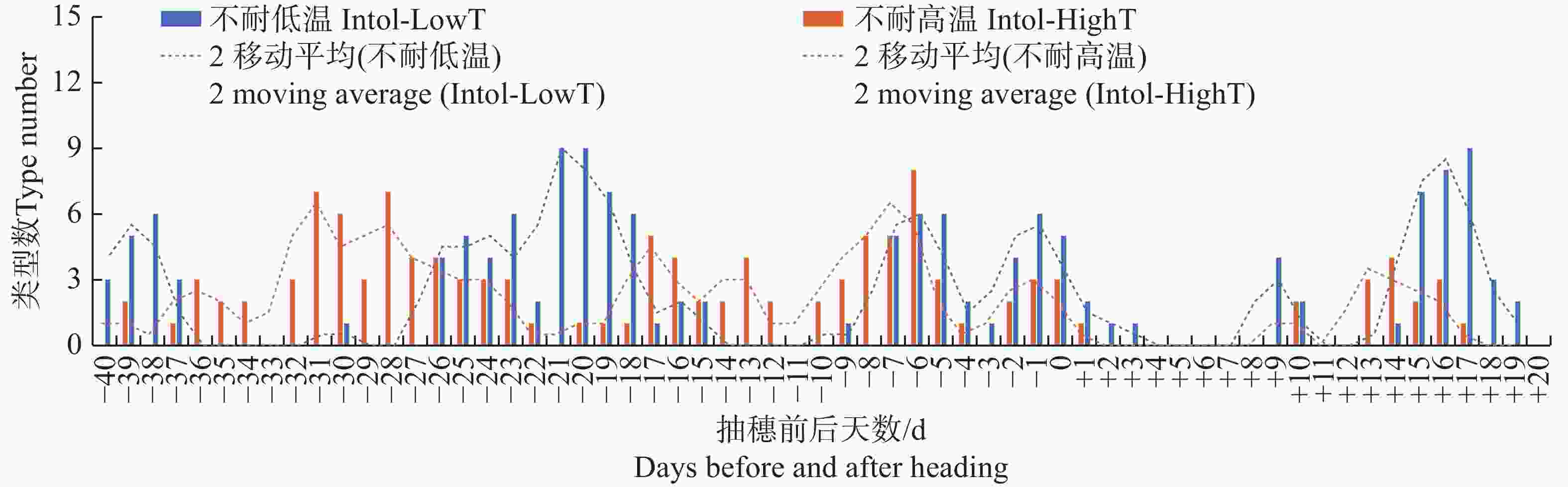
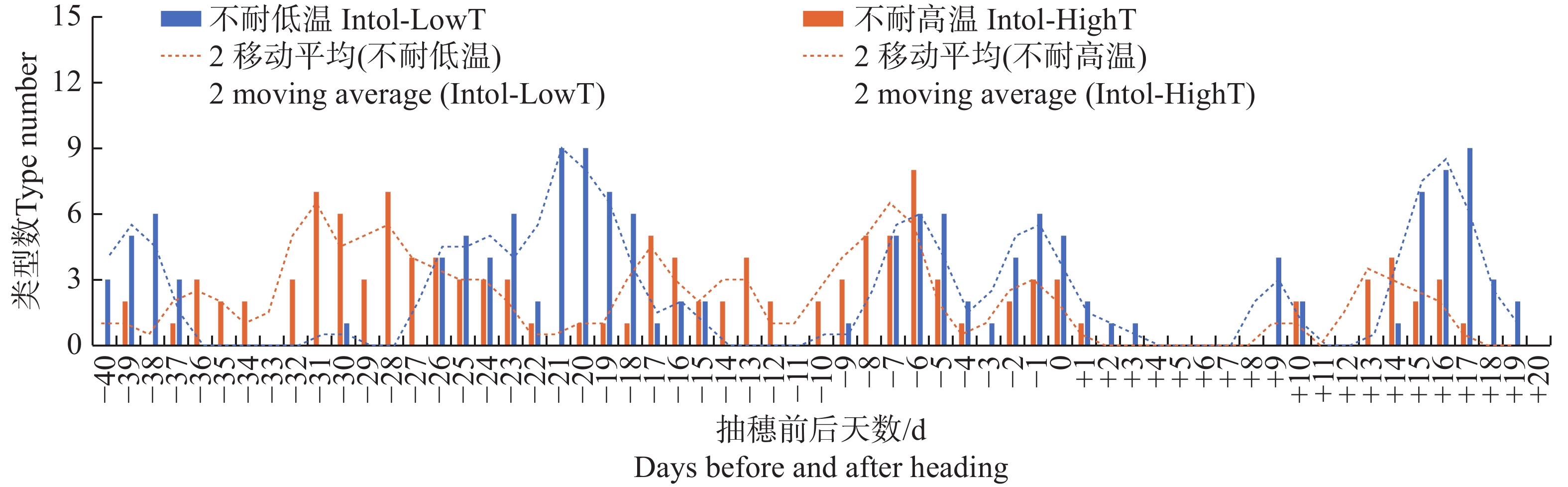
图 2 按抽穗前后各日期的最低气温统计不同日期不耐低温及不耐高温品种类型数分布
Figure 2. Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the minimum temperature recorded on each date before and after heading
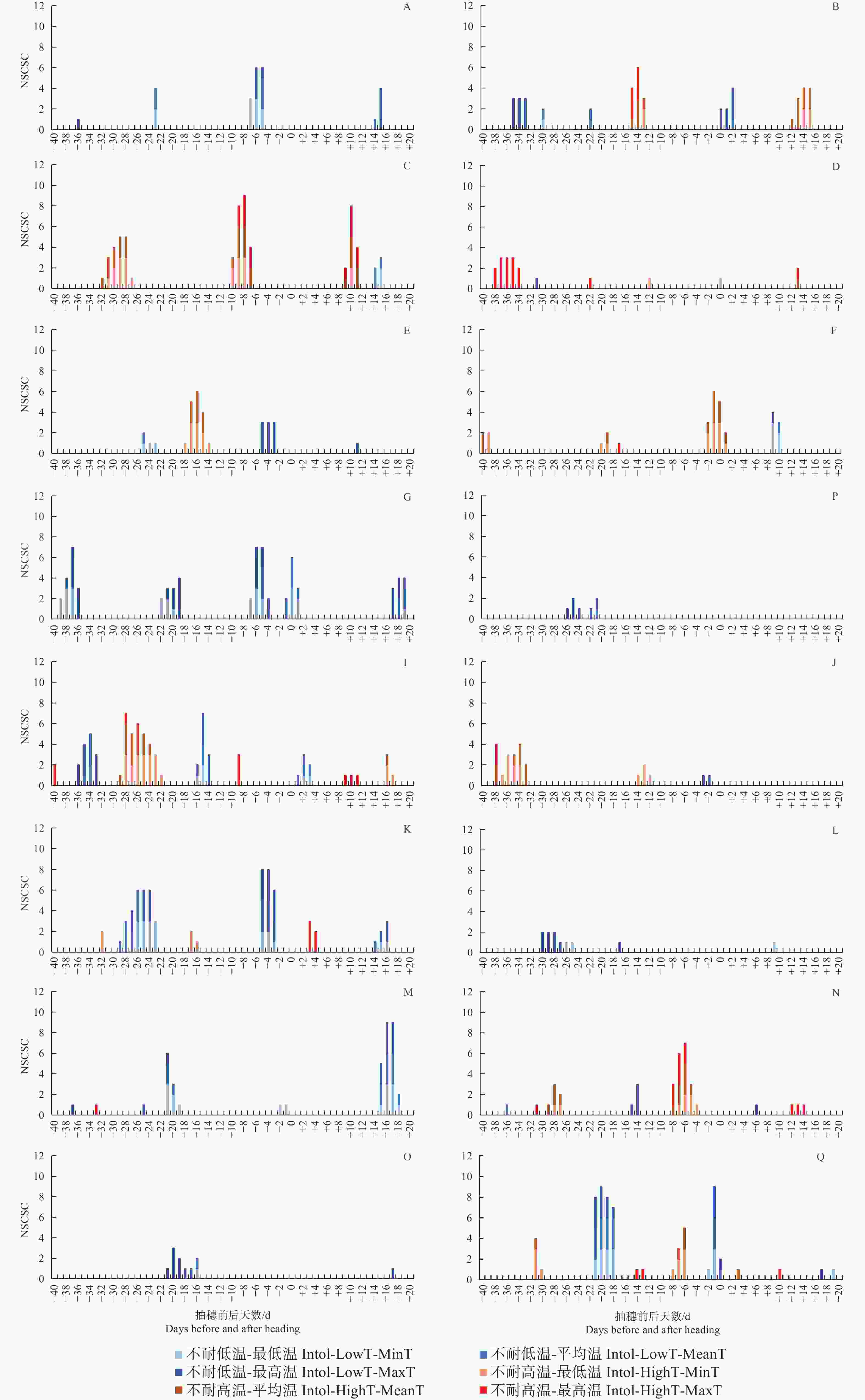
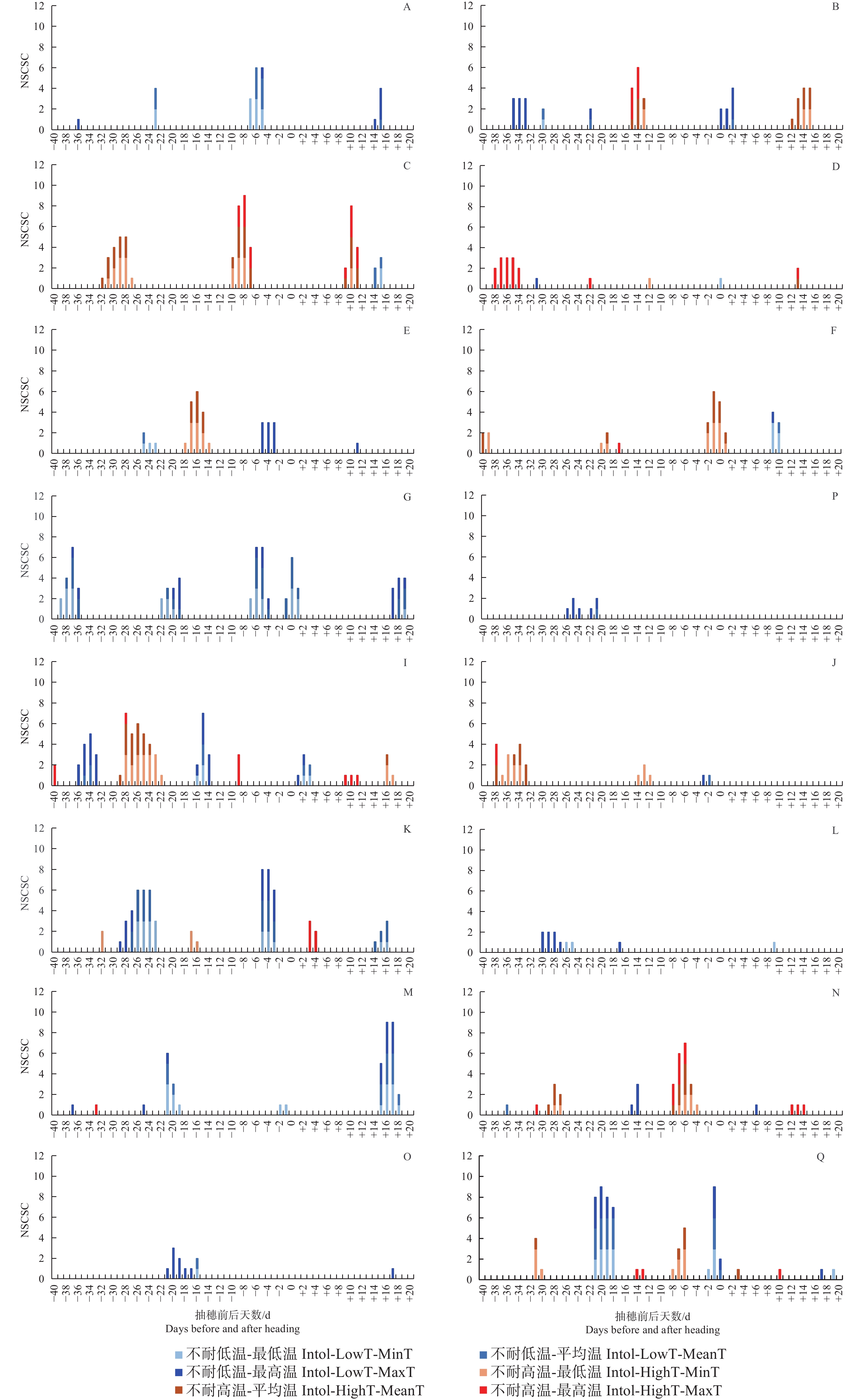
图 5 不同品种类型在不同发育阶段中不耐低温及不耐高温的统计口径数
Figure 5. Number of Statistical criteria for the intolerance to low and high temperatures in different varieties at different growth stages
不管按最低气温、最高气温还是平均气温统计,有3种品种类型在抽穗前21~18 d对低温敏感,该发育阶段很多品种类型处于花粉母细胞形成期,遇到低温会导致最终的结实率显著下降;受影响较明显的品种类型包括GJ-Te-LAN-GL、XI-SC和GJ-Tr。在抽穗当天至抽穗前7 d至抽穗后2 d左右,遇到较低的最低气温、最高气温及平均气温均会降低多种品种类型结实率,该阶段可能与花粉的活性有关系;受影响较明显的品种类型包括XI-LAN-T、XI-SC、XI-IMP1、XI-CC2、GJ-Te-RIMP-M和GJ-Tr。在抽随后14~18 d期间,水稻处于扬花期,GJ-Te-LAN-GL和XI-CC2等品种类型遇到低温也会导致结实率降低。另外,有些品种类型在不同发育阶段对不同气温类型响应不同。其中,XI-LAN-M在抽穗前35~33 d及抽穗当天至后2 d,XI-LAN-PIMP-T在抽穗前5至3天,XI-YG在抽穗当天至后2 d,XI-IMP1在抽穗前36~33 d,XI-CC2在抽穗前29~27 d,GJ-Te-LAN-PIMP在抽穗前30~25 d,GJ-Te-EA在抽穗前23~17 d,GJ-Te在抽穗前26~21 d,主要受较低的最高温度影响;而XI-SC在抽穗前39~36 d,XI-CC2在抽穗前26~23 d,主要受较低的最低气温和平均气温影响;XI-SC在抽穗前22~19 d,XI-IMP1在抽穗前16~14 d,则同时受较低的最低气温、平均气温和最高气温影响。
就高温胁迫而言,不管按最低气温、最高气温还是平均气温统计,在抽穗前9~5 d如遇高温,XI-LAN-PIMP-M和GJ-Te-RIMP-M等品种类型的结实率会显著降低。同时受到较高的最低气温、最高气温及平均气温影响的品种类型及发育阶段还包括抽穗前15~13 d的XI-LAN-M,抽随后9~11 d的XI-LAN-PIMPM,抽穗前8~4 d的GJ-Te-RIMP-M。另外,有些发育阶段和品种类型表现出对最低气温、最高气温及平均气温的不同响应。其中,XI-LAN-M在抽随后12~15 d,XI-LAN-PIMP-M在抽穗前32~27 d,XI-LAN-PIMP-T在抽穗前18~14 d,XI-RIMP-M在抽穗前后2 d,XI-IMP1在抽穗前28~22 d,XI-CC1在抽穗前37~33 d,GJ-Tr抽穗前8~6 d,在遇到较高的最低气温和平均气温时,结实率会显著降低;而XI-RIMP-T在抽穗前38~34 d,GJ-Te-RIMP-M在抽随后12~14 d主要受较高的最高温度胁迫。
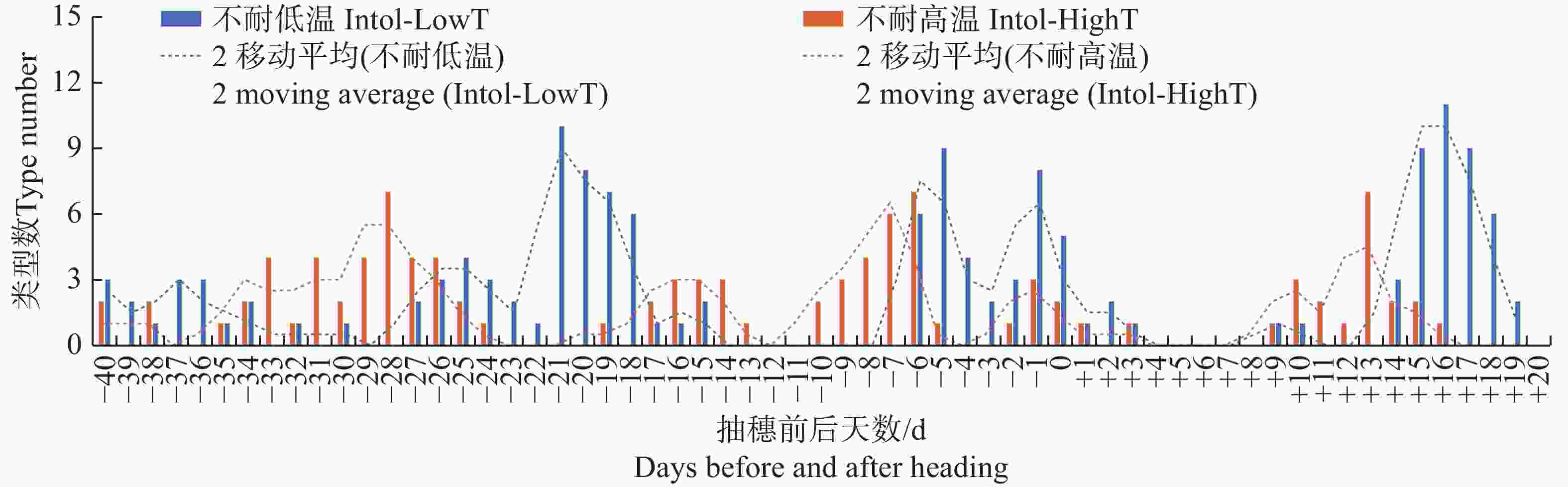
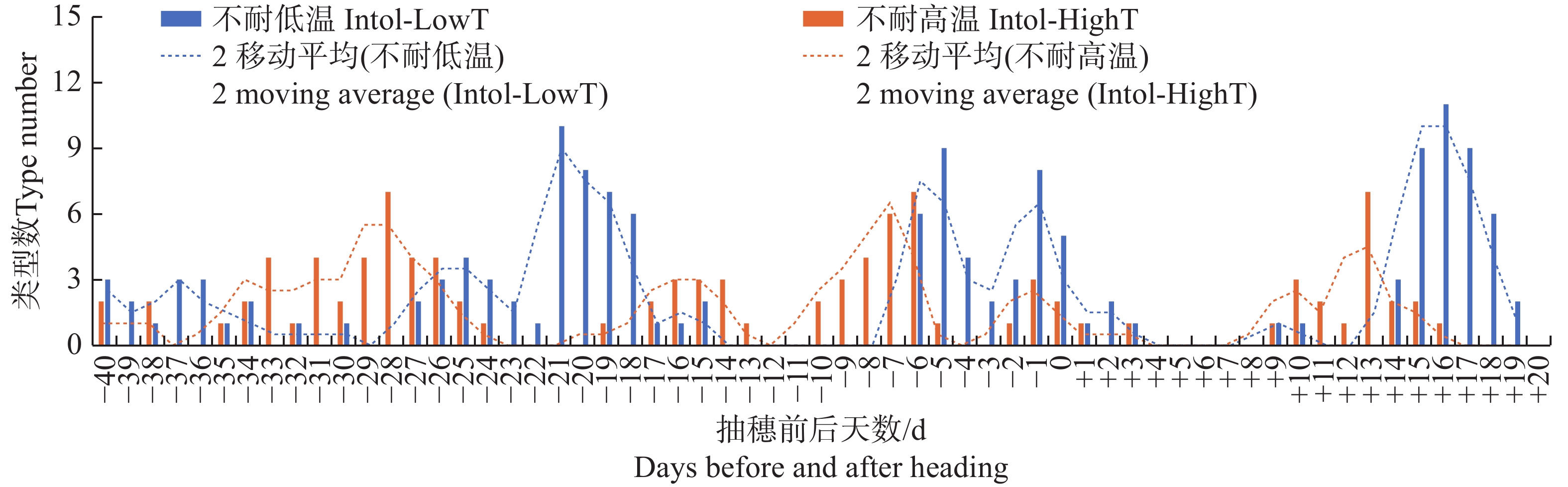
图 3 按抽穗前后各日期的平均气温统计不同日期不耐低温及不耐高温品种类型数分布
Figure 3. Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the average temperature recorded on each date before and after heading
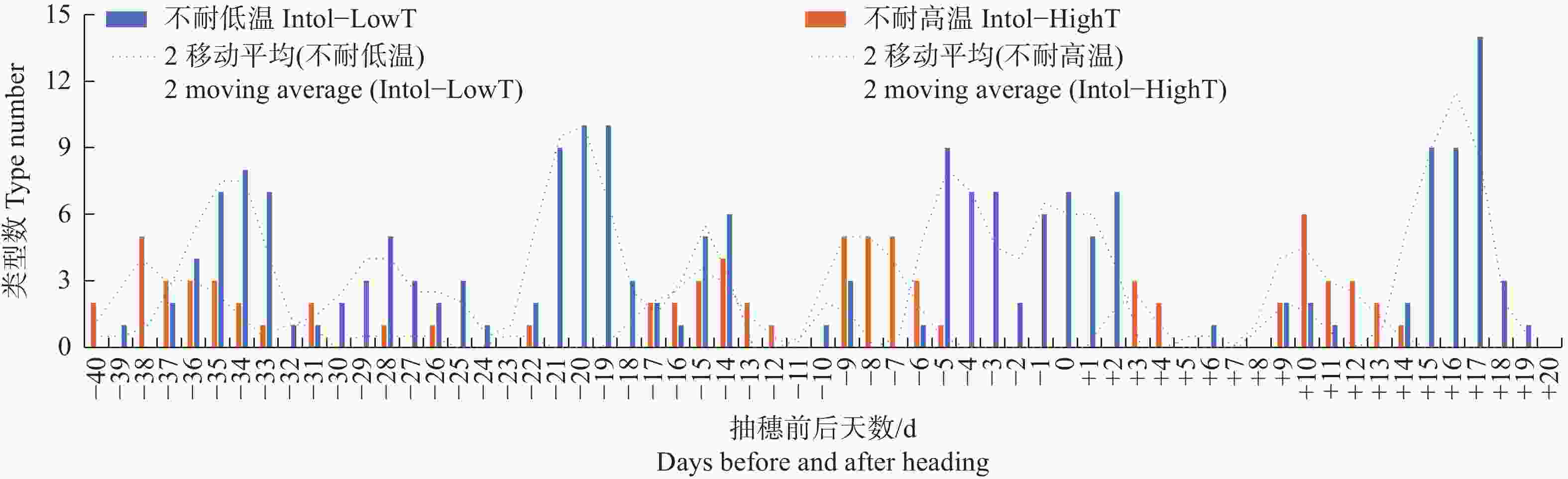
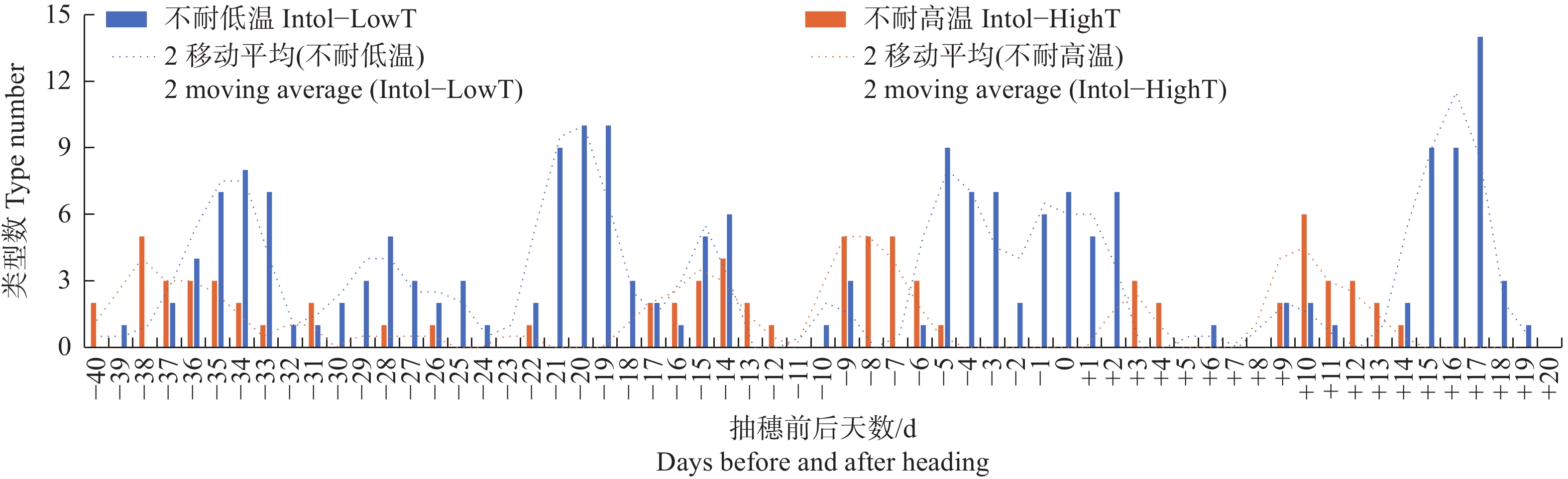
图 4 按抽穗前后各日期的最高气温统计不同日期不耐低温及不耐高温品种类型数分布
Figure 4. Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the maximum temperature recorded on each date before and after heading
-
为评估不同发育阶段不同气温类型对各品种类型最终结实率的影响程度,以每种品种类型最终结实率作为因变量,分别以其不同发育阶段的最低气温、平均气温和最高气温为自变量,开展回归分析。表2所列为至少连续2 d显著相关的品种类型及其发育阶段的回归系数,以及利用回归方程所估算的结实率分别降至80%和90%时的气温。结果显示,总体上南繁季水稻结实率主要受低温胁迫,即回归系数大于0,重要类型阶段有43个;高温胁迫(即回归系数小于0)对结实率的影响情况较少,仅15个类型阶段。抽穗前40 d至抽穗后20 d的某些阶段,最低气温高于23.3~29.9℃会导致某些类型结实率降至80%,而低于21.0~14.4℃会导致多数品种类型结实率降至80%,最高温度低于19.1-27.0℃也会导致很多品种类型结实率降低至80%(表2)。就不同品种类型看,籼稻中XI-IMP1耐冷性较弱,最低温度19.5℃、最高温度26.8℃就能导致其结实率降低至80%;XI-CC2耐冷性相对较强,最高温度21.8℃才造成结实率降低至80%;粳稻中GJ-Te-LAN-GL及GJ-YG耐冷性较强,GJ-Tr耐冷性较弱。
表 2 不同品种结实率对不同发育阶段最低气温、平均气温和最高气温冷胁迫的回归
Table 2. Regression of the seed setting rate of different varieties to the cold stress of minimum, average and maximum temperature at different developmental stages
气温类型
Types of temperature品种类型
Variety types发育阶段/d
Development stages温度效应/%
Effect of temperatureT-SSR80/℃ T-SSR90/℃ 最低气温
Minimum temperatureGJ-Tr(3K) −31~−30 −5.86 23.3~23.3 21.6-21.6 XI-RIMP-M −20~−19 −1.37 28.0~28.5 20.9-21.0 XI-CC2 −17~−16 −1.30 26.3~27.3 18.8-19.2 XI-LAN-PIMP-M −29~−28 −1.10 28.1~28.7 18.8-19.7 XI-LAN-PIMP-M −10~−8 −1.02 29.3~30.7 18.7-20.9 XI-RIMP-M −2~0 −0.91 29.8~33.2 19.9~20.0 XI-LAN-PIMP-T −17~−16 −0.87 29.9~30.5 18.5~18.8 XI-CC2 −4~−3 1.02 15.9~16.2 25.7~26.1 XI-CC2 −26~−23 1.32 14.4~17.9 23.7~24.8 GJ-Te-LAN-GL +16~+17 1.34 15.0~16.3 22.9~23.6 XI-SC −38~−37 1.38 15.3~15.7 22.7~22.9 XI-SC 0~+1 1.38 16.4~17.7 24.2~24.5 GJ-Tr(3K) −2~−1 1.64 19.1~19.3 25.1~25.5 GJ-Tr −2~−1 1.71 16.9~17.7 23.1~23.2 GJ-Te-LAN-GL −2~−1 1.79 18.3~19.5 24.5~24.6 XI-SC −7~−6 1.90 16.4~19.1 23.0~24.5 XI-LAN-T −7~−6 2.04 18.6~19.8 23.6~25.7 XI-IMP1 −16~−15 2.14 19.5~19.8 24.2~24.5 GJ-Tr −21~−18 2.39 17.5~21.6 23.2~24.6 GJ-Te-LAN-NG +16~+17 2.53 18.4~19.3 22.6~23.0 GJ-Tr(3K) −20~−18 2.60 19.7~21.5 23.6~25.1 GJ-Tr(3K) +14~+15 2.90 21.0~21.4 24.0~25.9 GJ-Tr(3K) −40~−38 3.62 20.3~20.5 22.7~23.6 平均气温
Average temperatureXI-IMP1 −28~−26 −3.23 24.0~24.7 20.7~21.7 XI-LAN-PIMP-M −9~−8 −1.31 30.6~30.8 22.7~23.4 XI-CC2 −5~−3 1.27 19.0~20.0 27.2~27.7 GJ-Te-LAN-GL +16~+17 1.59 18.6~20.7 26.0~26.3 XI-CC2 −26~−24 1.77 19.9~20.6 25.7~26.3 XI-SC 0~+1 2.25 21.3~21.9 26.0~26.1 GJ-Te-LAN-NG +16~+17 2.50 20.7~22.0 25.2~25.6 GJ-Tr(3K) −21~−18 2.67 21.9~23.6 26.0~26.6 XI-SC −6~−5 3.52 21.9~22.5 25.0~25.3 XI-LAN-T −6~−5 3.55 22.6~22.9 25.3~26.0 GJ-Tr(3K) +15~+18 3.97 23.7~25.7 27.3~27.9 XI-IMP1 −35~−34 3.98 22.5~23.1 25.3~25.5 GJ-Tr(3K) −40~−39 6.97 23.6~23.7 25.0~25.2 最高气温
Maximum temperatureGJ-Tr(3K) −13~−12 −13.92 29.2~29.7 28.2~29.1 XI-RIMP-T −37~−35 −5.02 29.4~30.4 27.4~28.0 GJ-Te-RIMP-M −8~−6 −1.56 32.4~38.1 27.1~28.7 XI-LAN-PIMP-M −8~−7 −1.35 34.1~35.0 27.0~27.2 XI-LAN-M −15~−14 −1.27 28.2-28.2 19.4-21.0 XI-LAN-PIMP-M +10~+11 −0.96 36.1-38.1 26.1-26.8 XI-LAN-PIMP-T −5~−3 1.01 19.1-20.9 29.7-30.0 XI-CC2 −5~−3 1.15 21.8-23.5 30.7-32.0 GJ-Te-LAN-GL +16~+17 1.24 20.5-24.5 30.8-31.5 XI-YG −34~−33 1.56 22.3-23.2 29.1-29.4 GJ-Tr −21~−20 1.95 24.2-24.4 29.4-29.4 XI-CC2 −28~−27 1.99 24.5-24.9 29.5-29.9 GJ-Tr(3K) −21~−19 2.11 25.3-26.0 30.2-30.6 GJ-Te-LAN-NG +16~+17 2.19 24.3-25.9 29.7-30.0 XI-LAN-M −35~−33 2.60 25.6-26.8 29.6-30.5 GJ-Te-EA −21~−20 2.81 23.8-24.6 27.7-27.9 GJ-Te-LAN-PIMP −30~−29 3.00 24.3~24.5 27.7~27.8 GJ-Te-IMP1 −10~−9 4.01 24.5~26.1 27.9~28.1 GJ-Te-IMP1 +9~+10 5.11 24.5~26.9 27.1~28.5 GJ-Tr(3K) +15~+17 5.12 27.0~29.2 30.4~31.1 XI-IMP1 −36~−33 5.13 26.8~27.1 28.7~29.5 XI-IMP1 −15~−14 5.60 26.7~27.6 29.0~29.3 注:GJ-Tr(3k)表示3k基因分析中的GJ-Tr类型;发育阶段所示为抽穗前的天数;温度效应的值为结实率对气温的回归系数,表示最低气温每升高1℃结实率的增加量,也指气温每降低1℃导致结实率的降低量;T-SSR80和T-SSR90分别表示结实率达80%和90%时相应阶段的气温范围。 Note: GJ-Tr (3k) represents the GJ-Tr type in the 3k genome analysis[7]; the developmental stage shown is the number of days before heading; the value of the temperature effect is the regression coefficient of the seed setting rate to temperature, indicating the increase in seed setting rate for every 1℃ increase in the temperature, and also indicating the decrease in seed setting rate for every 1℃ decrease in the temperature; T-SSR80 and T-SSR90 respectively represent the temperature ranges for the 80% and 90% seed setting rate within the corresponding stage. -
本研究以微核心种质为研究对象,系统分析了其在2009年11月—2010年4月三亚南繁季结实率对抽穗前后气温变化的响应模式。结果表明,南繁季水稻的结实率主要受低温胁迫影响,其影响程度与敏感期因品种类型而异。研究实现了对温度—结实率关系的定量化描述,为南繁育种的精准防灾与耐逆品种选育提供了重要依据。
本研究利用自然条件下遗传变异丰富的微核心种质群体,系统量化了不同类型水稻品种在南繁季不同生殖阶段气温与结实率之间的关系,并明确了关键的低温敏感阶段。结果发现,抽穗前21~18 d(约花粉母细胞形成期)是群体中对低温胁迫响应最集中的时期,多个籼稻和粳稻类型的结实率在此阶段与低温呈显著正相关,这与早期研究指出的减数分裂期为冷害主要窗口的结论一致[9]。此外,抽穗前后一周(尤其是抽穗当天至后2 d)以及抽穗后14~18 d(扬花期)也是重要的低温敏感期,表明花粉活性与授粉过程是低温影响的另一关键环节。
更为重要的是,本研究揭示了不同亚群间耐冷性存在显著差异,并首次给出了具体的低温阈值范围。例如,籼稻中的改良籼1群(XI-IMP1)耐冷性最弱,其结实率降至80%时所对应的最低和最高气温分别为19.5℃和26.8℃;而温带粳稻(如GJ-Te-LAN-GL)则表现出更强的耐受性。这一发现将传统“粳稻比籼稻耐冷”的定性认识推进到可量化的品种群/亚群水平。造成这种差异的内在机制,可能与不同亚群在长期适应与人工选择过程中形成了各自特有的温度应答基因有关,如SLG1、HSP101等基因赋予籼稻与粳稻不同的耐热性[10-11];而粳稻在适应高纬度、高海拔冷凉环境的过程中,则富集了更多耐冷基因,如CTB4a、CTB3和CTB5等[12-14]。因此,在南繁育种实践中,应针对不同来源的育种材料,依据其亚群归属及对应的温度阈值,采取差异化的播期调整或田间保温措施,并有针对性地开展各发育阶段温度等气候因子的监控,从而有效规避低温风险。
相较于低温,本研究在南繁季条件下观察到的高温胁迫相对较弱,主要发生在抽穗前9~5 d(花粉充实期)以及某些特定类型的抽穗前后,这与前人认为扬花期是主要高温敏感期的观点有所差异[15-17]。这种差异可能源于三亚冬季极端高温事件较少,使得对高温更敏感的品种类型仅在特定发育阶段遇到相对“偏高”温度时才表现出胁迫响应。此外,由于不同品种类型发育过程的复杂性,以及自然气温变化的多样性,难以保证所有品种类型在关键发育阶段均遭受有效的低温或高温胁迫。因此,未来需要设计更为精细的实验,以评估南繁季可能存在的热害风险。
-
本研究显示,南繁季低温是导致水稻结实率波动的主要温度胁迫因子,其影响具有明显的品种类型依赖性与发育阶段特异性。为提升南繁育种的防灾能力,建议构建基于“品种(亚群)−播期−温度敏感窗口期”三位一体的精准监控与管理体系。针对耐冷性较弱的材料(如部分改良籼稻),应通过调整播期,确保其关键敏感期避开当地低温时段。此外,本研究发现,地方非糯粳稻及来自云贵地区的粳稻类型表现出较强的耐冷性,籼稻中的华中类群2也具备较好的耐冷能力,未来可用于耐冷基因发掘、机制解析及耐冷性改良中的供体亲本。
Analysis of the response pattern of rice seed setting rate to temperature during the Nanfan season
-
摘要: 温度胁迫导致结实率降低是水稻南繁育种面临的主要风险之一。为系统解析不同水稻种质资源结实率对抽穗前后关键时段气温变化的量化响应模式,本研究以具有高度代表性的水稻微核心种质为对象,在典型南繁季节开展三亚田间试验,系统量化了抽穗前后关键发育阶段气温对不同遗传背景水稻结实率的影响。结果表明,南繁季水稻结实率主要受低温胁迫影响,且其作用显著高于高温胁迫。研究识别出2个关键的低温敏感窗口期,分别为抽穗前21~18 d(相当于花粉母细胞减数分裂期)和抽穗后14~18 d(扬花期)。通过回归分析,本研究首次量化了不同水稻亚群耐冷性的临界温度阈值,如改良籼1群(XI-IMP1)在最低气温降至19.5℃、最高气温降至26.8℃时,结实率可能下降至80%,而地方温带糯粳(GJ-Te-LAN-GL)等材料表现出更强的耐冷性。上述结果为南繁育种实践中基于品种遗传背景开展精准播期调整、耐冷亲本选择及关键生育期防灾提供了重要的理论依据与量化指标。Abstract: Temperature stress resulting in a decrease in seed setting rate is one of the greatest risks in rice during winter season in Hainan. Field experiments of a representative set of rice mini core collection were conducted in Sanya, Hainan during the typical winter season to systematically quantify the influence of temperature at different critical developmental stages before and after heading on the seed setting rate of rice with different genetic backgrounds. The results showed that the seed setting rate of rice during the winter season is mainly affected by low-temperature stress, with its impact being significantly greater than that of high-temperature stress. There are two key low-temperature sensitive phases, including those from 21st to 18th day before heading (equivalent to the meiotic division period of pollen mother cells) and those from 14th to 18th day after heading (representing pollination period). Through regression analysis, the critical temperature thresholds were quantified for cold tolerance of different rice subgroups for the first time. For example, the seed setting rate of the improved xian/indica 1 group (XI-IMP1) may drop to 80% when the minimum temperature drops to 19.5℃ and the maximum temperature drops to 26.8℃, while materials such as the landrace of temperate glutinous geng/japonica (GJ-Te-LAN-GL) exhibit higher cold tolerance. These findings provide important theoretical basis and quantitative indicators for precise adjustment of sowing date based on the genetic background of rice varieties, selection of cold-tolerant parents, and disaster prevention during critical growth periods in practices during winter season in Hainan.
-
Key words:
- Winter breeding and multiplication in Hainan /
- temperature /
- rice /
- seed setting rate /
- pattern analysis
-
图 2 按抽穗前后各日期的最低气温统计不同日期不耐低温及不耐高温品种类型数分布
注:不耐低温 Intol-LowT指结实率与温度显著正相关;不耐高温 Intol-HighT指结实率与温度显著负相关;类型数为同一天3种统计口径(M1、M3及M5,即单日温度、连续3日温度平均值、连续5日温度平均值)中显著相关品种类型的累加。下同。
Fig. 2 Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the minimum temperature recorded on each date before and after heading
Note: Intol-LowT, intolerance to low temperature, means that the seed setting rate is significantly positively correlated with temperature; Intol-HighT, intolerance to high temperature, means that the seed setting rate is significantly negatively correlated with temperature; the type number is the cumulative total of the three statistical categories (M1, M3 and M5, namely single-day temperature, average temperature of three consecutive days, and average temperature of five consecutive days)of the varieties that show significant correlation on the same day. Similarly hereinafter.
图 5 不同品种类型在不同发育阶段中不耐低温及不耐高温的统计口径数
注:最低温度、平均温度、最高温度是指分别按3种温度类型做结实率与温度的相关性;统计口径是指单日温度、连续3日温度平均值、连续5日温度平均值;NSCSC指显著相关统计口径数,即某个温度类型下结实率与温度显著相关的统计口径数。
Fig. 5 Number of Statistical criteria for the intolerance to low and high temperatures in different varieties at different growth stages
Note: Minimum temperature, average temperature, and maximum temperature refer to the correlations between seed setting rate and temperature for the three temperature types, respectively; Statistical criteria refer to single-day temperature, average temperature of three consecutive days, and average temperature of five consecutive days; NSCSC is number of statisitical criteria with significant correlation, which refers to the correlation between seed setting rate and temperature under certain temperature types.
图 3 按抽穗前后各日期的平均气温统计不同日期不耐低温及不耐高温品种类型数分布
Fig. 3 Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the average temperature recorded on each date before and after heading
图 4 按抽穗前后各日期的最高气温统计不同日期不耐低温及不耐高温品种类型数分布
A: XI-LAN-T; B: XI-LAN-M; C: XI-LAN-PIMP-M; D: XI-RIMP-T; E: XI-LAN-PIMP-T; F: XI-RIMP-M; G: XI-SC; H: XI-YG;
Fig. 4 Distribution of the number of different types of varieties that are intolerant to low temperature or high temperature, based on the maximum temperature recorded on each date before and after heading
I: XI-IMP1; J: XI-CC1; K: XI-CC2; L: GJ-Te-LAN-PIMP; M: GJ-Te-LAN-GL; N: GJ-Te-RIMP-M; O: GJ-Te-EA; P: GJ-Te; Q: GJ-Tr.
表 1 不同类型水稻品种抽穗期及结实率变异情况
Table 1 Variability of heading date and seed setting rate of different types of rice varieties
品种类型
Variety types份数
Num.抽穗期/d
Heading date结实率/%
Seed setting rate最小值
Min.平均值
Mean最大值
Max.最小值
Min.平均值
Mean最大值
Max.华南籼 XI-SC 24 66 88.4 125 61.5 86.4 95.4 云贵籼 XI-YG 21 70 98.2 123 72.7 87.3 93.6 改良籼1群 XI-IMP1 19 74 103.8 126 29.4 83.9 95.4 华中籼1群 XI-CC1 38 74 95.4 121 66.0 88.3 96.7 东亚温带粳
GJ-Te-EA21 63 88.3 114 50.7 88.2 96.8 华中籼2群 XI-CC2 23 80 97.0 112 71.8 86.6 94.9 南亚籼 XI-SA 6 82 92.2 98 67.5 81.3 91.7 华北粳 GJ-NC 19 65 88.9 106 62.4 83.0 96.7 改良籼2群 XI-IMP2 9 101 112.3 121 64.9 85.0 94.7 混合群 admix 9 77 100.2 122 72.5 85.6 93.0 温带粳 GJ-Te 17 76 97.8 109 72.4 87.9 96.3 温带改良粳1群
GJ-Te-IMP16 79 95.3 103 55.0 86.5 97.3 热带粳 GJ-Tr(3k) 7 108 115.4 127 68.5 81.8 91.7 典型地方籼
XI-LAN-T17 66 85.6 123 61.5 84.3 95.4 混合地方籼
XI-LAN-M7 70 98.6 110 72.7 83.7 89.3 地方及初级改良混合籼
XI-LAN-PIMP-M20 70 99.3 122 73.0 88.3 95.2 典型近期改良籼
XI-RIMP-T10 74 101.7 121 73.7 89.0 95.4 典型地方及初级改良籼
XI-LAN-PIMP-T32 74 93.8 116 71.8 88.3 96.7 混合近期改良籼
XI-RIMP-M5 93 109.2 119 84.6 88.9 93.4 地方温带非糯粳
GJ-Te-LAN-NG9 72 86.8 104 65.9 81.6 94.9 地方及初级改良温带粳
GJ-Te-LAN-PIMP15 74 90.2 114 67.6 89.6 96.8 地方温带糯粳
GJ-Te-LAN-GL11 76 97.8 109 76.7 87.4 94.4 混合近期改良温带粳
GJ-Te-RIMP-M9 77 93.9 106 84.4 89.1 94.3 热带粳 GJ-Tr 6 96 114.0 127 73.0 85.3 91.7  下载: 导出CSV
下载: 导出CSV
表 2 不同品种结实率对不同发育阶段最低气温、平均气温和最高气温冷胁迫的回归
Table 2 Regression of the seed setting rate of different varieties to the cold stress of minimum, average and maximum temperature at different developmental stages
气温类型
Types of temperature品种类型
Variety types发育阶段/d
Development stages温度效应/%
Effect of temperatureT-SSR80/℃ T-SSR90/℃ 最低气温
Minimum temperatureGJ-Tr(3K) −31~−30 −5.86 23.3~23.3 21.6-21.6 XI-RIMP-M −20~−19 −1.37 28.0~28.5 20.9-21.0 XI-CC2 −17~−16 −1.30 26.3~27.3 18.8-19.2 XI-LAN-PIMP-M −29~−28 −1.10 28.1~28.7 18.8-19.7 XI-LAN-PIMP-M −10~−8 −1.02 29.3~30.7 18.7-20.9 XI-RIMP-M −2~0 −0.91 29.8~33.2 19.9~20.0 XI-LAN-PIMP-T −17~−16 −0.87 29.9~30.5 18.5~18.8 XI-CC2 −4~−3 1.02 15.9~16.2 25.7~26.1 XI-CC2 −26~−23 1.32 14.4~17.9 23.7~24.8 GJ-Te-LAN-GL +16~+17 1.34 15.0~16.3 22.9~23.6 XI-SC −38~−37 1.38 15.3~15.7 22.7~22.9 XI-SC 0~+1 1.38 16.4~17.7 24.2~24.5 GJ-Tr(3K) −2~−1 1.64 19.1~19.3 25.1~25.5 GJ-Tr −2~−1 1.71 16.9~17.7 23.1~23.2 GJ-Te-LAN-GL −2~−1 1.79 18.3~19.5 24.5~24.6 XI-SC −7~−6 1.90 16.4~19.1 23.0~24.5 XI-LAN-T −7~−6 2.04 18.6~19.8 23.6~25.7 XI-IMP1 −16~−15 2.14 19.5~19.8 24.2~24.5 GJ-Tr −21~−18 2.39 17.5~21.6 23.2~24.6 GJ-Te-LAN-NG +16~+17 2.53 18.4~19.3 22.6~23.0 GJ-Tr(3K) −20~−18 2.60 19.7~21.5 23.6~25.1 GJ-Tr(3K) +14~+15 2.90 21.0~21.4 24.0~25.9 GJ-Tr(3K) −40~−38 3.62 20.3~20.5 22.7~23.6 平均气温
Average temperatureXI-IMP1 −28~−26 −3.23 24.0~24.7 20.7~21.7 XI-LAN-PIMP-M −9~−8 −1.31 30.6~30.8 22.7~23.4 XI-CC2 −5~−3 1.27 19.0~20.0 27.2~27.7 GJ-Te-LAN-GL +16~+17 1.59 18.6~20.7 26.0~26.3 XI-CC2 −26~−24 1.77 19.9~20.6 25.7~26.3 XI-SC 0~+1 2.25 21.3~21.9 26.0~26.1 GJ-Te-LAN-NG +16~+17 2.50 20.7~22.0 25.2~25.6 GJ-Tr(3K) −21~−18 2.67 21.9~23.6 26.0~26.6 XI-SC −6~−5 3.52 21.9~22.5 25.0~25.3 XI-LAN-T −6~−5 3.55 22.6~22.9 25.3~26.0 GJ-Tr(3K) +15~+18 3.97 23.7~25.7 27.3~27.9 XI-IMP1 −35~−34 3.98 22.5~23.1 25.3~25.5 GJ-Tr(3K) −40~−39 6.97 23.6~23.7 25.0~25.2 最高气温
Maximum temperatureGJ-Tr(3K) −13~−12 −13.92 29.2~29.7 28.2~29.1 XI-RIMP-T −37~−35 −5.02 29.4~30.4 27.4~28.0 GJ-Te-RIMP-M −8~−6 −1.56 32.4~38.1 27.1~28.7 XI-LAN-PIMP-M −8~−7 −1.35 34.1~35.0 27.0~27.2 XI-LAN-M −15~−14 −1.27 28.2-28.2 19.4-21.0 XI-LAN-PIMP-M +10~+11 −0.96 36.1-38.1 26.1-26.8 XI-LAN-PIMP-T −5~−3 1.01 19.1-20.9 29.7-30.0 XI-CC2 −5~−3 1.15 21.8-23.5 30.7-32.0 GJ-Te-LAN-GL +16~+17 1.24 20.5-24.5 30.8-31.5 XI-YG −34~−33 1.56 22.3-23.2 29.1-29.4 GJ-Tr −21~−20 1.95 24.2-24.4 29.4-29.4 XI-CC2 −28~−27 1.99 24.5-24.9 29.5-29.9 GJ-Tr(3K) −21~−19 2.11 25.3-26.0 30.2-30.6 GJ-Te-LAN-NG +16~+17 2.19 24.3-25.9 29.7-30.0 XI-LAN-M −35~−33 2.60 25.6-26.8 29.6-30.5 GJ-Te-EA −21~−20 2.81 23.8-24.6 27.7-27.9 GJ-Te-LAN-PIMP −30~−29 3.00 24.3~24.5 27.7~27.8 GJ-Te-IMP1 −10~−9 4.01 24.5~26.1 27.9~28.1 GJ-Te-IMP1 +9~+10 5.11 24.5~26.9 27.1~28.5 GJ-Tr(3K) +15~+17 5.12 27.0~29.2 30.4~31.1 XI-IMP1 −36~−33 5.13 26.8~27.1 28.7~29.5 XI-IMP1 −15~−14 5.60 26.7~27.6 29.0~29.3 注:GJ-Tr(3k)表示3k基因分析中的GJ-Tr类型;发育阶段所示为抽穗前的天数;温度效应的值为结实率对气温的回归系数,表示最低气温每升高1℃结实率的增加量,也指气温每降低1℃导致结实率的降低量;T-SSR80和T-SSR90分别表示结实率达80%和90%时相应阶段的气温范围。 Note: GJ-Tr (3k) represents the GJ-Tr type in the 3k genome analysis[7]; the developmental stage shown is the number of days before heading; the value of the temperature effect is the regression coefficient of the seed setting rate to temperature, indicating the increase in seed setting rate for every 1℃ increase in the temperature, and also indicating the decrease in seed setting rate for every 1℃ decrease in the temperature; T-SSR80 and T-SSR90 respectively represent the temperature ranges for the 80% and 90% seed setting rate within the corresponding stage.
下载: 导出CSV
-
[1] Zong W B, Guo X T, Zhang K, et al. Photoperiod and temperature synergistically regulate heading date and regional adaptation in rice [J]. Journal of Experimental Botany, 2024, 75(13): 3762−3777. https://doi.org/10.1093/jxb/erae209 doi: 10.1093/jxb/erae209 [2] 陈冠铭, 李劲松, 林亚琼. 国家南繁功能价值与发展机遇研究分析[J]. 种子, 2012, 31(3): 69−71. https://doi.org/10.16590/j.cnki.1001-4705.2012.03.041 doi: 10.16590/j.cnki.1001-4705.2012.03.041 [3] Counce P A, Keisling T C, Mitchell A J. A uniform, objective, and adaptive system for expressing rice development [J]. Crop Science, 2000, 40(2): 436−443. https://doi.org/10.2135/cropsci2000.402436x doi: 10.2135/cropsci2000.402436x [4] 李文彬, 王贺, 张福锁. 高温胁迫条件下硅对水稻花药开裂及授粉量的影响[J]. 作物学报, 2005, 31(1): 134−136. https://doi.org/10.3321/j.issn:0496-3490.2005.01.025 doi: 10.3321/j.issn:0496-3490.2005.01.025 [5] 随晶晶, 赵桂龙, 金欣, 等. 水稻孕穗期耐冷调控的分子及生理机制研究进展[J]. 中国水稻科学, 2025, 39(1): 1−10. https://doi.org/10.16819/j.1001-7216.2025.231208 doi: 10.16819/j.1001-7216.2025.231208 [6] Zhang H L, Zhang D L, Wang M X, et al. A core collection and mini core collection of Oryza sativa L. in China [J]. Theoretical and Applied Genetics, 2011, 122(1): 49−61. https://doi.org/10.1007/s00122-010-1421-7 doi: 10.1007/s00122-010-1421-7 [7] Wang W S, Mauleon R, Hu Z Q, et al. Genomic variation in 3, 010 diverse accessions of Asian cultivated rice [J]. Nature, 2018, 557(7703): 43−49. https://doi.org/10.1038/s41586-018-0063-9 doi: 10.1038/s41586-018-0063-9 [8] Wang X Q, Wang W S, Tai S S, et al. Selective and comparative genome architecture of Asian cultivated rice (Oryza sativa L. ) attributed to domestication and modern breeding [J]. Journal of Advanced Research, 2022, 42: 1−16. https://doi.org/10.1016/j.jare.2022.08.004 doi: 10.1016/j.jare.2022.08.004 [9] González-Schain N, Roig-Villanova I, Kater M M. Early cold stress responses in post-meiotic anthers from tolerant and sensitive rice cultivars [J]. Rice, 2019, 12(1): 94. https://doi.org/10.1186/s12284-019-0350-6 doi: 10.1186/s12284-019-0350-6 [10] Xu Y F, Zhang L, Ou S J, et al. Natural variations of SLG1 confer high-temperature tolerance in indica rice [J]. Nature Communications, 2020, 11(1): 5441. https://doi.org/10.1038/s41467-020-19320-9 doi: 10.1038/s41467-020-19320-9 [11] Tripathi G, Biswal A K, Sarkar N K, et al. HSP101 alleles from indica and japonica subspecies of rice (Oryza sativa L. ) show differential response to heat stress [J]. Planta, 2025, 262(6): 140. https://doi.org/10.1007/s00425-025-04863-2 doi: 10.1007/s00425-025-04863-2 [12] Guo H F, Gao S L, Li H H, et al. Natural variation of CTB5 confers cold adaptation in plateau japonica rice [J]. Nature Communications, 2025, 16(1): 1032. https://doi.org/10.1038/s41467-025-56174-5 doi: 10.1038/s41467-025-56174-5 [13] Li J, Guo H F, Lou Q J, et al. Natural variation of indels in the CTB3 promoter confers cold tolerance in japonica rice [J]. Nature Communications, 2025, 16(1): 1613. https://doi.org/10.1038/s41467-025-56992-7 doi: 10.1038/s41467-025-56992-7 [14] Zhang Z Y, Li J J, Pan Y H, et al. Natural variation in CTB4a enhances rice adaptation to cold habitats [J]. Nature Communications, 2017, 8: 14788. https://doi.org/10.1038/ncomms14788 doi: 10.1038/ncomms14788 [15] Xing Y H, Lu H Y, Zhu X F, et al. How rice responds to temperature changes and defeats heat stress [J]. Rice, 2024, 17(1): 73. https://doi.org/10.1186/s12284-024-00748-2 doi: 10.1186/s12284-024-00748-2 [16] Matsui T, Omasa K. Rice (Oryza sativa L. ) cultivars tolerant to high temperature at flowering: anther characteristics [J]. Annals of Botany, 2002, 89(6): 683−687. https://doi.org/10.1093/aob/mcf112 doi: 10.1093/aob/mcf112 [17] Zhen F X, Wang W, Wang H Y, et al. Effects of short-term heat stress at booting stage on rice-grain quality [J]. Crop & Pasture Science, 2019, 70(6): 486−498. https://doi.org/10.1071/cp18260 doi: 10.1071/cp18260 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 163
- HTML全文浏览量: 126
- 被引次数: 0
