

-
遗传信息的表达遵循中心法则:DNA经转录生成RNA,RNA再经翻译合成蛋白质以执行各类生命活动。该过程在转录、RNA加工与翻译等多个环节均受到多层级、精细化的调控。转运RNA(tRNA)是直接参与蛋白质合成的关键功能分子,成熟过程中常在多个位点发生转录后修饰(tRNA modification)。RNA修饰是表观转录组学(epitranscriptomics)的核心研究内容。自二十世纪以来,研究者已在古菌、真细菌和真核生物中鉴定出160余种RNA修饰类型,涵盖甲基化、假尿嘧啶化、甲硫醇化等多种形式,其中约70%的修饰被发现富集于tRNA及tRNA衍生片段(tRF,tRNA-derived fragments)上[1-5]。tRNA修饰通过影响tRNA的热稳定性,翻译效率,密码子识别等方式参与基因表达调控[6-9]。在酿酒酵母(Saccharomyces cerevisiae)中,第58位的m1A(1-methyladenosine)对于tRNAiMet的稳定性至关重要,该修饰在tRNA形成的早期引入,不依赖于其他修饰,但缺乏m1A修饰的tRNA会出现折叠错误,加速tRNA进入降解途径[10-11]。在嗜热栖热菌(Thermus thermophilus)中,第54位的m5U(5-methyluridine)存在动态调控机制,该修饰核苷可以进一步硫代为m5s2U(5-methyl-2-thiouridine),修饰丰度与生长温度呈正相关,硫代基团增加tRNA热稳定性,维持高温下的代谢水平,是经tRNA修饰介导的温度相关代谢调控机制[12]。tRNA第34位的修饰通常与密码子—反密码子识别相关,I(Inosine)广泛存在于tRNAANN的34位(I34),在tRNA形成早期由腺嘌呤水解脱氨基编辑而成,未修饰的A34通常只能与U配对,而I34可以识别U、C、A三种核苷酸,从而拓展密码子识别,提高了密码子解码的灵活性和基因表达效率[13-14]。由此可见,tRNA修饰在生命体中发挥至关重要的作用,是蛋白翻译调控系统的重要组成部分。
巴氏醋杆菌(Acetobacter pasteurianus)是醋酸发酵的微生物之一,广泛用于传统食品发酵中。巴氏醋杆菌具有极强的乙酸和乙醇耐受性,能在10%乙醇,2%乙酸胁迫条件下保持代谢活性,是研究微生物耐酸机制的重要微生物[15-16]。巴氏醋杆菌适应酸胁迫的机制有待深入研究,特别是从转录后修饰层面对巴氏醋杆菌环境适应策略的相关研究仍较为缺乏,观察巴氏醋杆菌的tRNA修饰类型,修饰位点以及酸胁迫环境下的tRNA修饰动态变化,有助于从转录后修饰层面理解微生物的环境胁迫适应机制[17]。
质谱分析是检测与鉴定转录后修饰的标准方法,也在转录组、代谢物谱构建等研究领域广泛使用[18-21]。质谱分析凭借高灵敏度、高分辨率及对化学结构解析的能力,为小分子物质表征提供可信数据[22-25]。质谱法绘制tRNA修饰图谱通常基于核苷分析和寡核苷酸分析相结合的策略,核苷分析提供修饰种类信息,寡核苷酸分析提供序列定位与修饰组合状态信息,通过自下而上逐级定位修饰,从而实现对修饰种类与位点的系统解析,此外,通过比较不同培养条件或遗传背景下的质谱数据,还可进一步揭示修饰的动态变化特征,为后续代谢工程改造提供基础数据支持。
-
无水乙醇、葡萄糖购买自西陇科学股份有限公司;蛋白胨和酵母提取物购买自英国Oxoid公司;甲酰胺、EDTA、丙烯腈、三甲胺购买自上海麦克林生化科技股份有限公司;SDS购买自北京索莱宝科技有限公司;1-溴-3氯丙烷购买自上海迈瑞尔化学技术有限公司;碱性磷酸酶购买自北京宝日医生物技术有限公司;RNase T1 和RNase A购买自赛默飞世尔科技公司;核酸酶P1购买自北京纽英伦生物技术有限公司;蛇毒磷酸二酯酶购买自上海西格玛奥德里奇贸易有限公司;甲醇、乙腈等流动相成分购买自德国默克公司。
-
巴氏醋杆菌AS 1.41(Acetobacter pasteurianus AS 1.41)购买自湖南丰晖生物科技有限公司。菌株培养使用GYP培养基(葡萄糖2%,酵母提取物0.5%,蛋白胨0.5%,乙醇3%),在此基础上分别设置0%、1%、2%(V/V)乙酸胁迫组,每个胁迫组别设置3个重复,170 r·min-1,30℃培养96 h。
-
修饰核苷鉴定使用Thermo Scientific Orbitrap Exploris 120 质谱仪;核苷分析使用SCIEX Triple Quad
7500 质谱仪;寡核苷酸分析使用Thermo Scientific Orbitrap Exploris 480质谱仪;各型质谱仪均联用高效液相色谱仪。 -
采用甲酰胺—苯酚法提取巴氏醋杆菌总RNA。菌株培养96 h后,7 000 r·min−1 离心5 min收集菌体,加入10%发酵液体积的甲酰胺抽提剂(EDTA 18 mmol·L−1,SDS 0.025%,β−巯基乙醇1%,甲酰胺定容至对应体积),涡旋混匀,95℃水浴振荡5 min,然后9 000 r·min−1 离心10 min取上清液,分别加入等体积的水饱和苯酚和1−溴−3氯丙烷,振荡混匀,9 000 r·min−1离心10 min取上清液,经乙醇沉淀回收获得总RNA,再经10%尿素聚丙烯酰胺凝胶电泳实现成分分离,切胶回收总tRNA。
-
将10 μg tRNA溶解于3 μL水中,加入30 μL 41%(V/V)乙醇/三甲胺乙酸盐和10 μL丙烯腈,72℃孵育2 h,乙醇沉淀回收[26]。
-
核苷分析:使用0.1 U核酸酶P1,在pH5.3环境下37℃消化1 h,然后使用500 mmol·L−1 Tris-HCl调整pH至9.0,加入5 U碱性磷酸酶和0.5 U蛇毒磷酸二酯酶,37℃酶切1 h,真空干燥。寡核苷酸分析:使用20 U的RNase T1和20 ng的RNase A在37℃消化1 h,缓冲液分别为0.25 mol·L−1 乙酸铵(pH 5.3)和0.25 mol·L−1 乙酸铵(pH 7.0)。
-
核苷分析采用电喷雾电离正离子模式,流动相A为0.1%(V/V)甲酸水,流动相B为0.1%(V/V)甲酸95%(V/V)乙腈水,采用Waters Acquity UPLC BEH amide色谱柱(2.1 mm×150 mm, 1.7 μm)在36℃条件下分离,流速为100 μL·min−1,梯度洗脱程序为:0~5 min,90% B相;5~35 min,由90% B相线性降至40% B相;35~40 min,维持40% B相。寡核苷酸分析采用电喷雾电离负离子模式,消化产物等体积混合0.1 mol·L-1三乙胺醋酸盐缓冲液(pH 7.0),流动相A为水,流动相B为乙腈。两相均含有1.0%(V/V)的六氟异丙醇和0.24%(V/V)的三乙胺,色谱柱为Oligonucleotide BEH C18柱(2.1 mm×100 mm, 1.7 μm),色谱分离条件参照此前报导的方法设置[27]。
-
tRNA修饰的类型、分子结构、分子质量、离子对、修饰酶的蛋白序列等信息来自Modomics数据库(
https://genesilico.pl/modomics/ )[1]和Uniprot数据库(https://www.uniprot.org/ )。Blastp工具来自NCBI(https://www.ncbi.nlm.nih.gov/ )。质谱数据使用Thermo Xcalibur Qual Browser和SCIEX OS软件进行分析。 -
通过尿素聚丙烯酰胺凝胶电泳分离总tRNA和其他RNA组分,切胶回收后,经尿素聚丙烯酰胺凝胶电泳验证。电泳图显示,巴氏醋杆菌总tRNA成分完整,无降解,可以用作后续分析(图1)。

图 1 巴氏醋杆菌的总 tRNA 组分
-
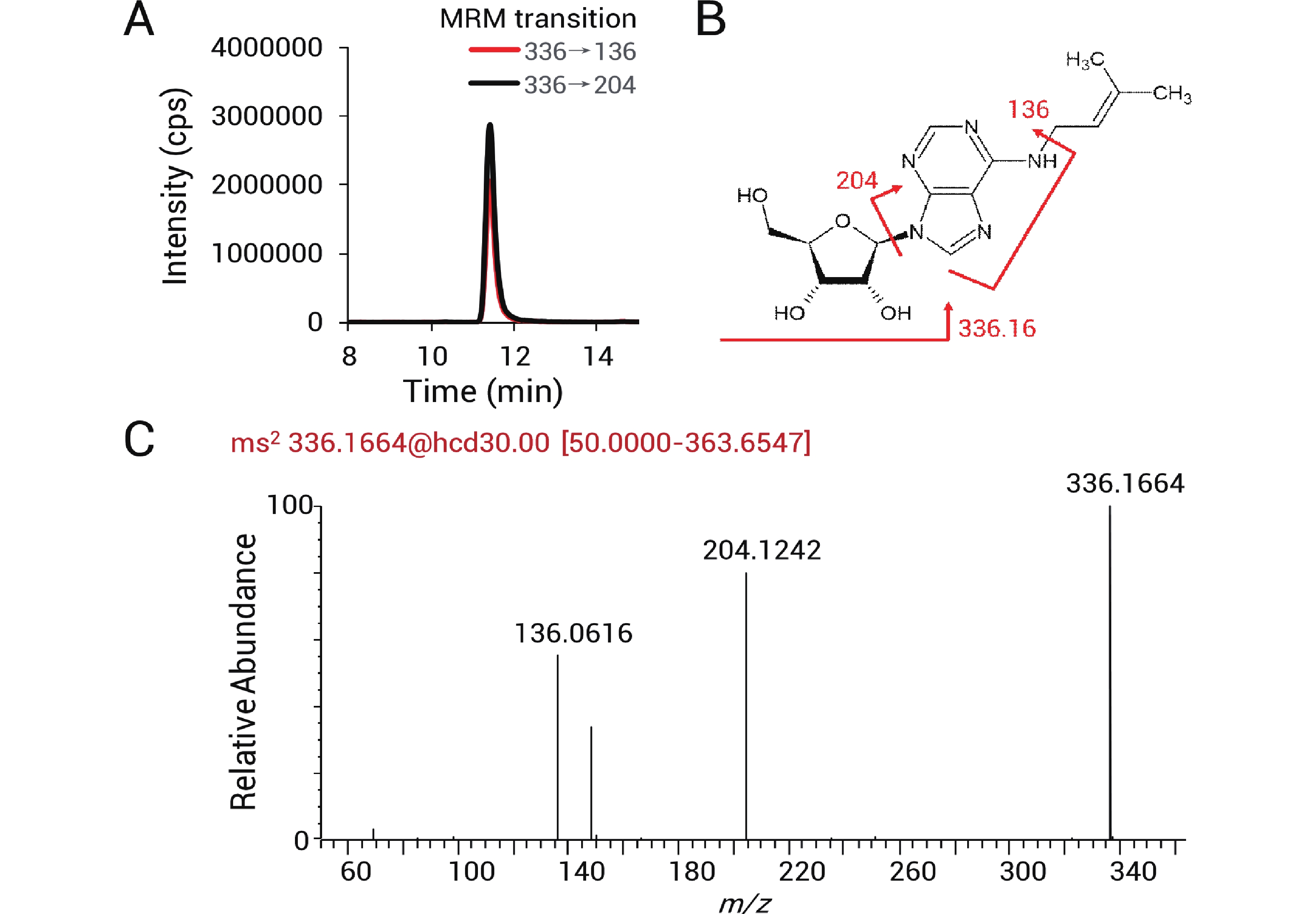
采用三重四极杆质谱和高分辨质谱对从巴氏醋杆菌tRNA上消化下来的修饰核苷进行鉴定和相对定量追踪。如图2-A所示,336→136和336→204离子对可用于检测修饰核苷i6A(N6-isopentenyladenosine)。m/z 136和204的产物离子分别对应i6A裂解得到的腺苷碎片及异戊烯基修饰腺苷碎片(图2-B)。同时可以在高分辨质谱上得到i6A正电离子(z=+1)的精确质荷比
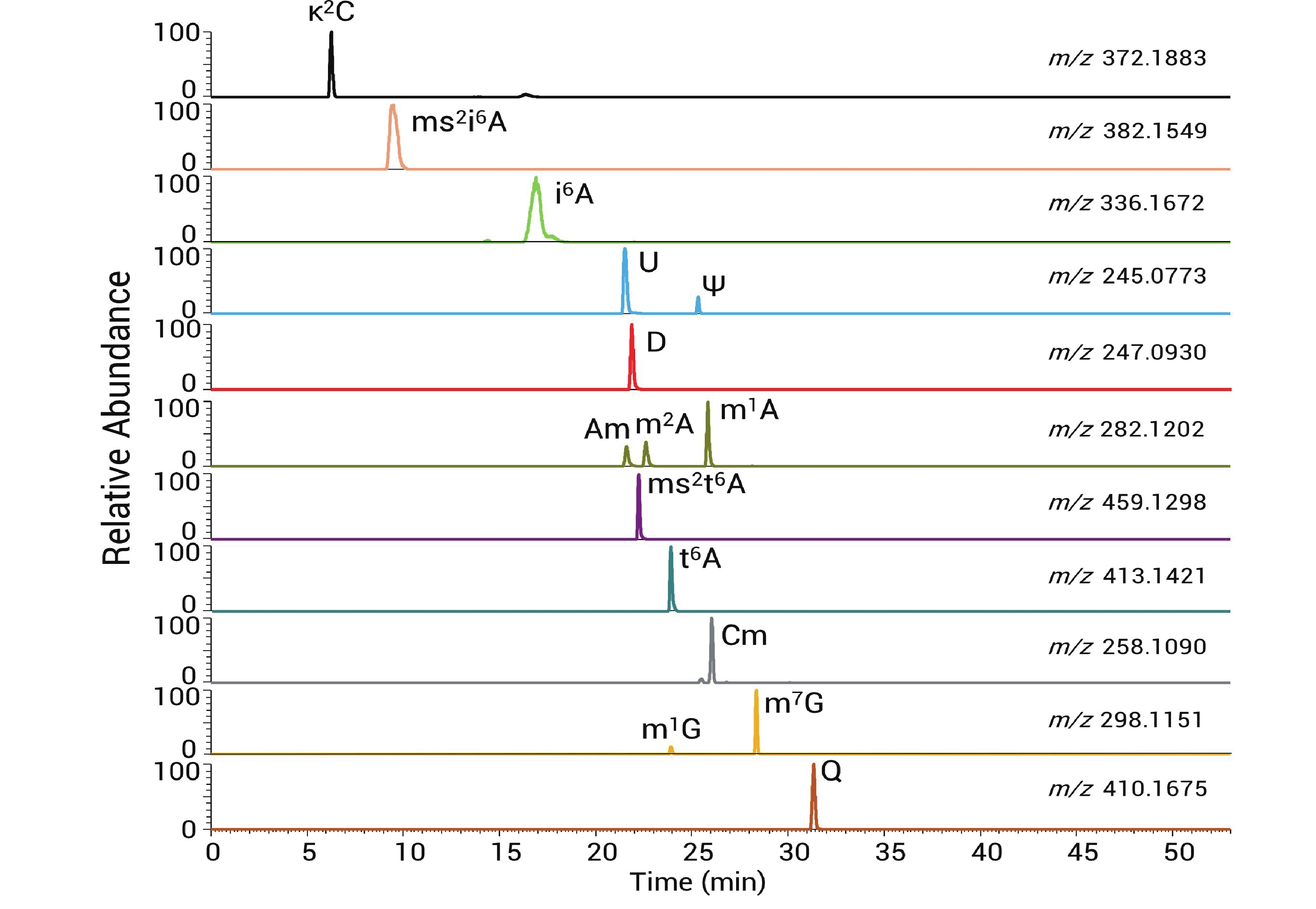
336.1664 (与理论值336.1672 质量差为-0.0008 Da)。二级谱显示对应清晰的136和204产物离子(图2-C)。通过这样的方法在质谱上检测出14种tRNA修饰核苷,包括κ2C(2-lysidine), ms2i6A(2-methylthio-N6-isopentenyladenosine), i6A(N6-isopentenyladenosine), Ψ(pseudouridine), D(dihydrouridine), Am(2'-O-methyladenosine), m2A(2-methyladenosine), m1A(1-methyladenosine), ms2t6A(2-methylthio-N6-threonylcarbamoyladenosine), t6A(N6-threonylcarbamoyladenosine), Cm(2'-O-methylcytidine), m1G(1-methylguanosine), m7G(7-methylguanosine)以及Q(queuosine),同分异构体通过保留时间区分,各修饰核苷的提取离子色谱峰如图3所示。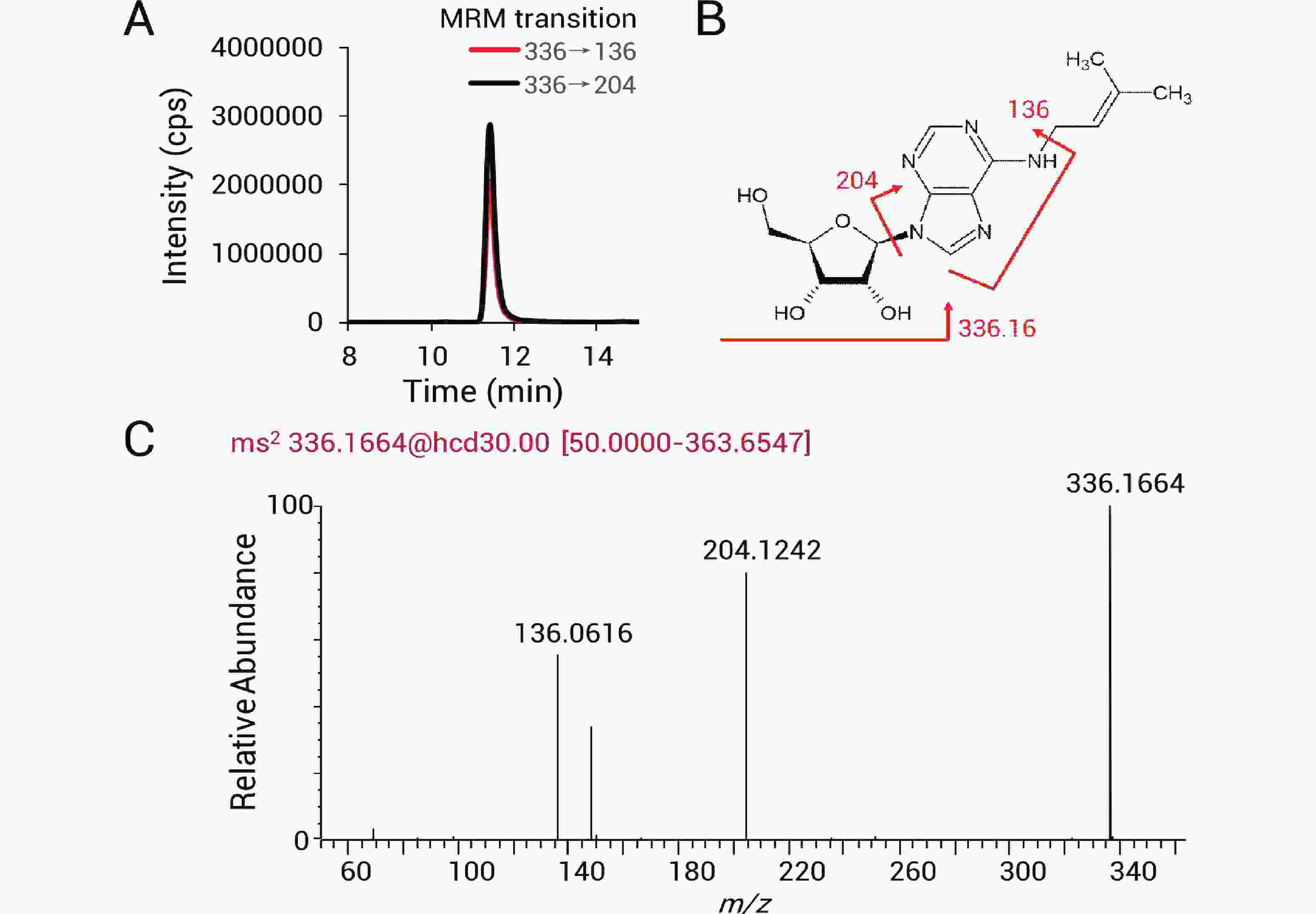
图 2 修饰核苷鉴定示意图
Figure 2. Schematic illustration of modified nucleoside identification
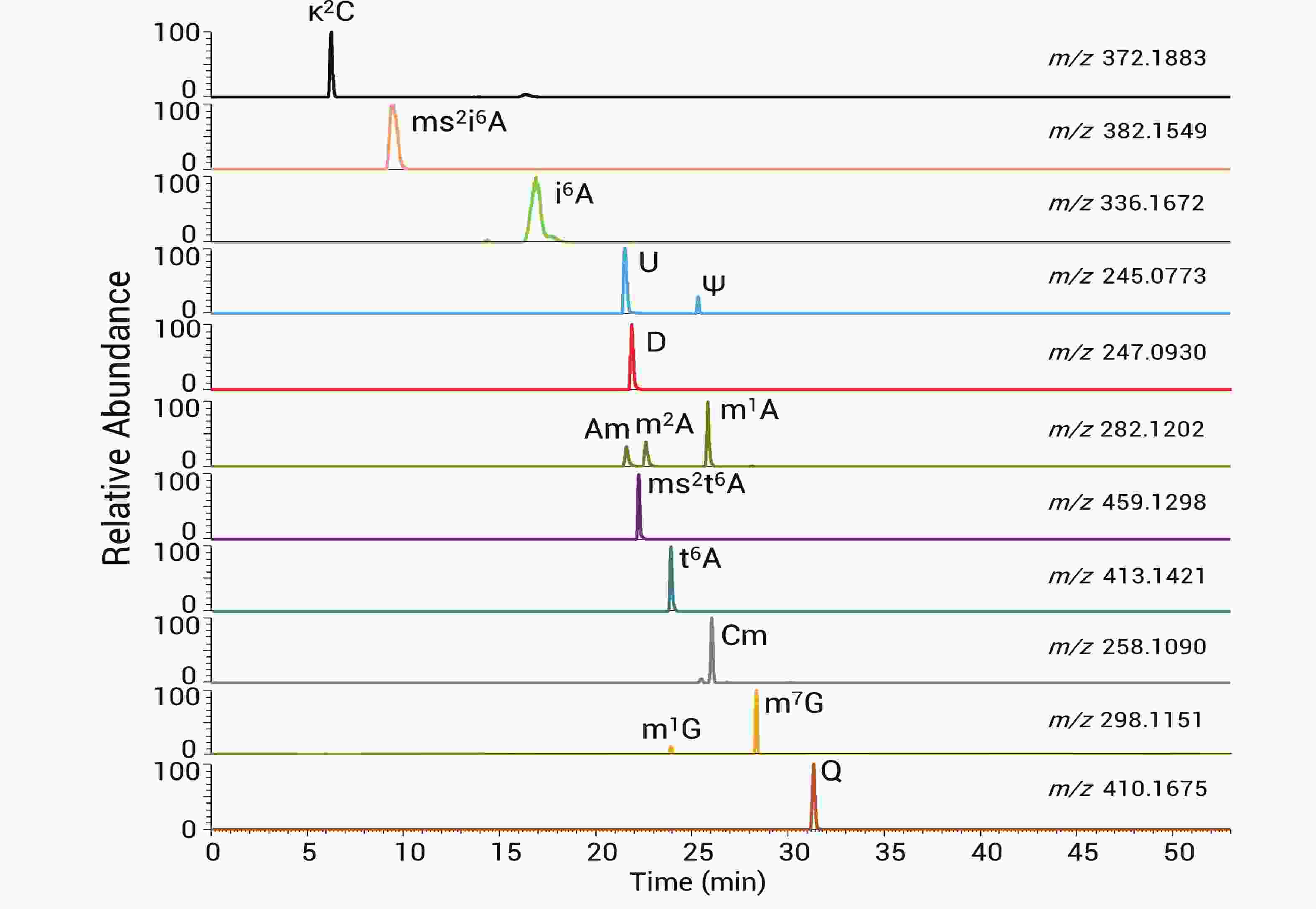
图 3 14 种修饰核苷提取色谱离子峰
-
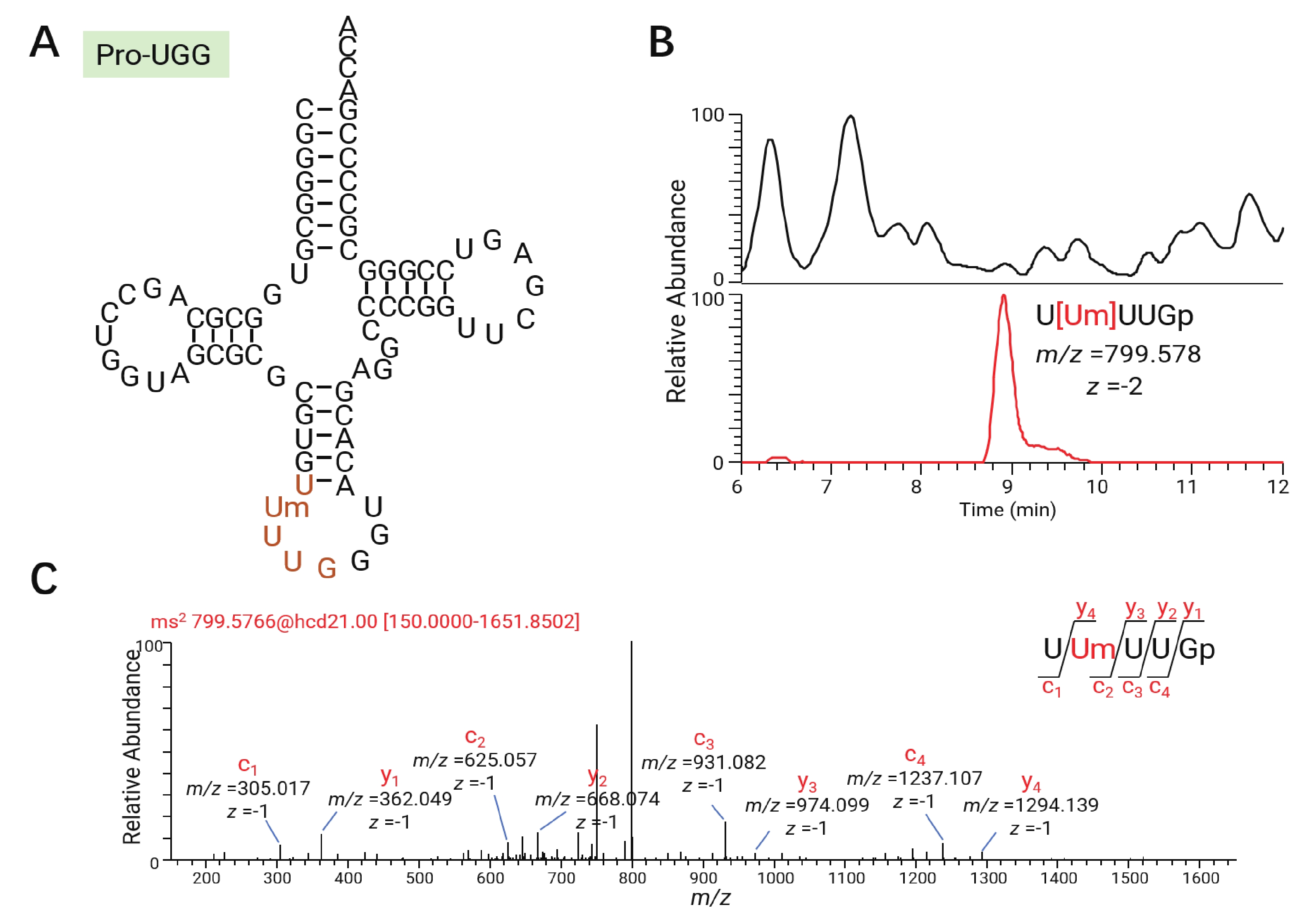
tRNA修饰所在的位点与它的功能密切相关。例如前面提到的Inosine在tRNA 第34位上参与wobble摆动拓宽了tRNA解读密码子的范围。将巴氏醋杆菌的总tRNA进行限制性核苷酸酶T1和限制性核苷酸酶A的消化,可以通过寡核苷酸分析定位tRNA修饰的具体位点。以巴氏醋杆菌tRNAPro-UGG为例,该tRNA经RNase T1消化切割后产生特征片段U[Um]UUGp,位于反密码子环,且携带一个甲基化修饰(图4-A)。通过观察该片段的一级精确质荷比(图4-B)和对碰撞诱导碎裂的特定产物离子的鉴定,得到其中的修饰Um的位置(图4-C)。在这里仅采用具有序列独特性的片段进行鉴定,即某片段的序列在巴氏醋杆菌所有tRNA中仅能在特定某一个tRNA上找到时,才认为其中的修饰位点是可信的。通过寡核苷酸分析方法,在核苷分析鉴定的14种tRNA修饰的基础上还鉴定出了Um,cmnm5Um,mnm5U,cmnm5U,cmnm5s2U五种修饰。这五种修饰都是正电模式下难以电离的尿苷衍生物,在负电模式的寡核苷酸分析中得以检出。序列独特性片段及修饰位点的汇总结果如表1所示。
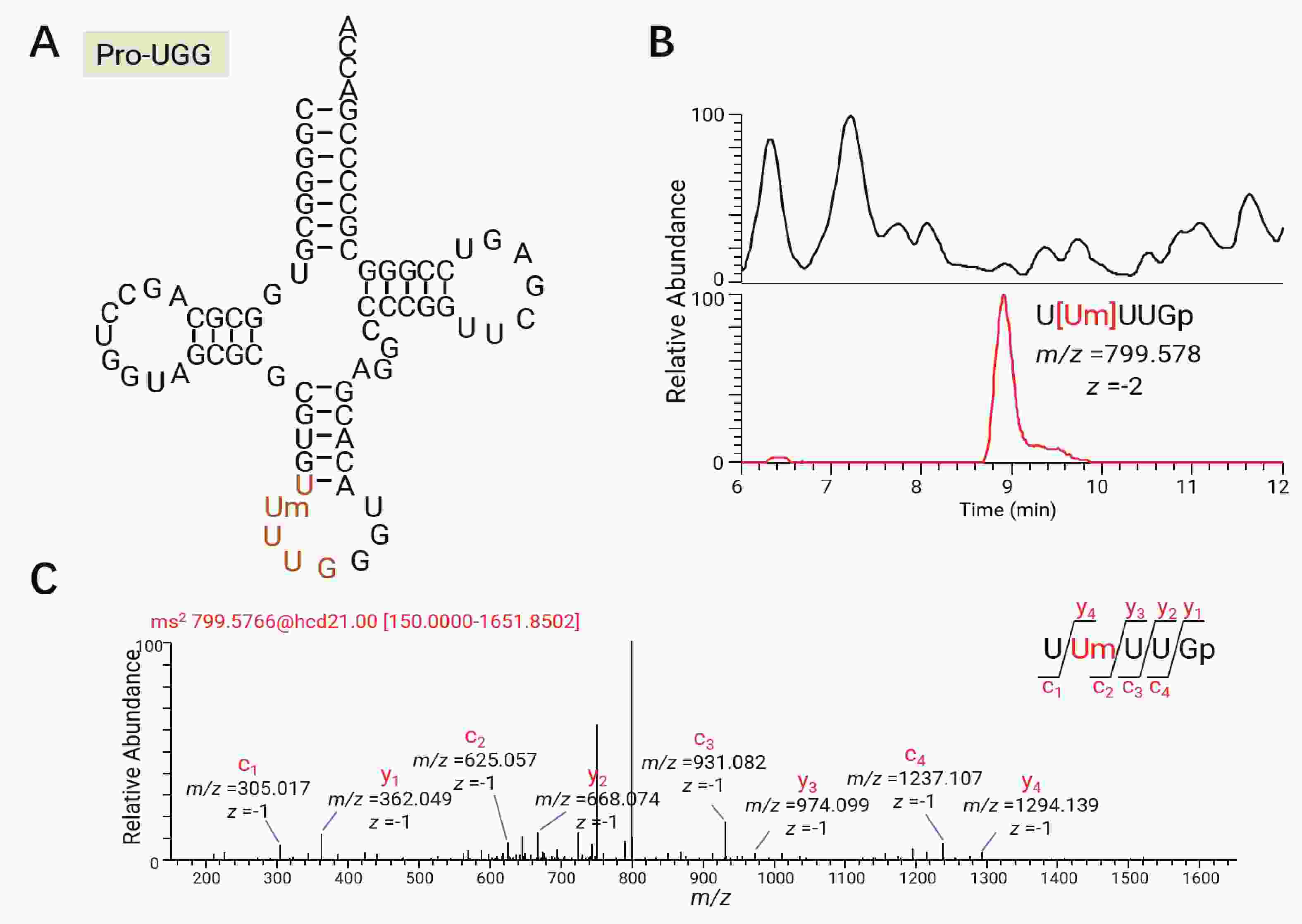
图 4 寡核苷酸分析及修饰核苷定位示意图
Figure 4. Schematic diagram of oligonucleotide analysis and modified nucleoside localization
表 1 巴氏醋杆菌tRNA序列独特性片段
Table 1. Unique tRNA fragments identified from Acetobacter pasteurianus
转运RNA
tRNA序列独特性片段
Unique fragments质荷比
m/z电荷
zMet-CAU AA[Um]CAU[t6A]AUCCCUUGp 1638.212 −3 UUCAAAUCCUUCCGp 1467.506 −3 Tyr-GUA U[i6A]AAUCCGp 1319.196 −2 U[ms2i6A]AAUCCGp 1342.190 −2 pGGAGGGAUp 1392.662 −2 AAAGGGGGCp 1516.714 −2 His-GUG UUUQUGp 1028.128 −2 Ala-CGC CAAUGp 815.106 −2 AUUCCCCUCAGp 1731.213 −2 CUCCACCA 1212.681 −2 GAGGAG[m7G]Up 1359.687 −2 Asp-GUC CCUQUC[m2A]CGp 1503.719 −2 Trp-CCA U[Cm]UCCA[i6A]AACCGp 1948.291 −2 U[Cm]UCCA[ms2i6A]AACCGp 1971.285 −2 ACCCUCCACCCCUGp 1458.519 −3 Pro-CGG AAUCUUCUCACUCCGp 1569.187 −3 AGGG[m7G]Cp 1022.146 −2 Lys-CUU A[Cm]UCUU[ms2t6A]AUCAGp 1340.168 −3 [m7G]UCCAAGp 1147.157 −2 Gly-GCC CACAACCUUGp 1590.206 −2 AAUCCCAUCGp 1590.206 −2 Ile-GAU AUAAGp 827.111 −2 UCCUCCCUGp 1402.160 −2 Ala-UGC UCUCCACCA 1365.693 −2 AACCCGp 967.134 −2 CUU[mnm5U]Gp 813.599 −2 AGGGG[m7G]Up 1195.161 −2 fMet-CAU [Cm]UCAU[m2A]ACCUGp 1757.234 −2 UUCAAAUCCUGp 1743.710 −2 Lys-UUU ACU[cmnm5s2U]UU[ms2t6A]AUCGp 1891.219 −2 Ser-GGA UUAAUAGp 1133.136 −2 Arg-CCG CC[Cm]UCCGp 1103.143 −2 Asn-GUU ACUQUU[t6A]AUCGp 1899.266 −2 UCCUACUUCGp 1567.178 −2 Ser-CGA A[ms2i6A]AACCGp 1200.691 −2 Ser-UGA UDDAAGp 970.626 −2 AAGGAACp 1163.669 −2 UCU[mnm5U]Gp 813.599 −2 A[ms2i6A]AACCAGp 1365.217 −2 AAUCCCACCCCAUCCGp 1677.887 −3 Pro-UGG [m1G]UACACGp 1147.157 −2 U[Um]UUGp 799.578 −2 Arg-UCU CCU[cmnm5U]CU[t6A]AGp 1542.206 −2 CCU[cmnm5Um]CU[t6A]AGp 1549.214 −2 Pro-GGG GGG[m1G]GACp 1194.669 −2 Leu-UAG AUUUAGp 968.610 −2 [m1G]UUCUGp 971.610 −2 UCCCUCCACCCGp 1871.235 −2 Val-UAC UUUACACCGp 1426.172 −2 Gln-CUG AACUACCGp 1284.673 −2 [Cm]UCUGp 798.594 −2 Cys-GCA CA[i6A]AUCCGp 1318.704 −2 CA[ms2i6A]AUCCGp 1341.698 −2 AGAAUp 827.111 −2 Val-CAC CC[Um]UCACACGp 1585.208 −2 CACCCACCA 1376.715 −2 Leu-CAG [m1G]UACCUGp 1135.644 −2 UUUCAGp 956.604 −2 UCUUCUUCUGp 1556.157 −2 Phe-GAA AA[ms2i6A]AUCGp 1201.183 −2 Glu-UUC CCDAGp 804.108 −2 CCCU[cmnm5s2U]UC[m2A]CGp 1625.199 −2 Arg-ACG ACUACGp 967.626 −2 [m2A]AUCUGp 975.126 −2 Thr-CGU UCAUUCGp 1109.125 −2 Ser-GCU AAUCCCUUCCCCUCCGp 1662.535 −3 CU[t6A]AACCGp 1357.191 −2 Thr-GGU CACUCCCUUGp 1566.686 −2 UUCAAUCCUGp 1579.184 −2 Thr-UGU UUCAAAUCCUACUGp 1475.510 −3 Gln-UUG AUU[cmnm5s2U]UGp 1008.601 −2 AAUCCUCCCGp 1578.200 −2 Ala-GGC CAUUCAGp 1120.639 −2 Leu-CAA UCCCCUCUCUCGp 1860.214 −2 ACUCAA[ms2i6A]AUCGp 1823.763 −2 Gly-UCC AUUCCCUCUGp 1567.178 −2 CCU[cmnm5U]CC[m2A]AGp 1476.203 −2 GAGGAGGGUp 1525.203 −2 Ile2-CAU CU[κ2C]AU[m2A]ACGp 1508.741 −2 Val-GAC CACUAUCUUGp 1579.184 −2 Gly-CCC C[Um]UCCCAAGp 1432.687 −2 UUCAAUGp 1121.131 −2 Leu-GAG AAUACCCCGp 1437.193 −2 UCUUCUCGp 1250.132 −2 -
假尿苷($ \Psi $)与尿苷(U)互为同分异构体,分子质量相同,因此难以在常规质谱分析中仅凭质量差异直接区分与鉴定$ \Psi $。为解决这一问题,本研究采用氰乙基化衍生化策略,使$\Psi $的N1位发生加成反应,生成人工衍生物“$\Psi $-CE”。寡核苷酸片段中每引入一个$\Psi $-CE位点,其分子质量将增加53Da,从而可在二级质谱(MS/MS)中实现$ \Psi$位点的判定。基于“化学加成—质谱检测”的方法,在巴氏醋杆菌总tRNA中共鉴定到7个$\Psi $位点,分别位于第31、32、38、39、40、55和70位,汇总见表2 。
表 2 巴氏醋杆菌tRNA中Ψ的定位
Table 2. Mapping of Ψ in Acetobacter pasteurianus tRNAs
转运RNA
tRNA序列独特性片段
Unique fragments质荷比
m/z电荷
z位置
PositionTrp-CCA [Ψ-CE][Cm]UCCA[i6A]AACCGp 1974.805 −2 31 Leu-UAG A[Ψ-CE]UUAGp 995.123 −2 32 Pro-UGG [m1G][Ψ-CE]ACACGp 1173.671 −2 38 Met-CAU AA[Um]CAU[t6A]A[Ψ-CE]CCCUUGp 1655.887 −3 39 Tyr-GUA U[i6A]AA[Ψ-CE]CCGp 1345.709 −2 40 Met-CAU U[Ψ-CE]CAAAUCCUUCCGp 1485.182 −3 55 Ala-UGC UC[Ψ-CE]CCACCA 1392.207 −2 70 -
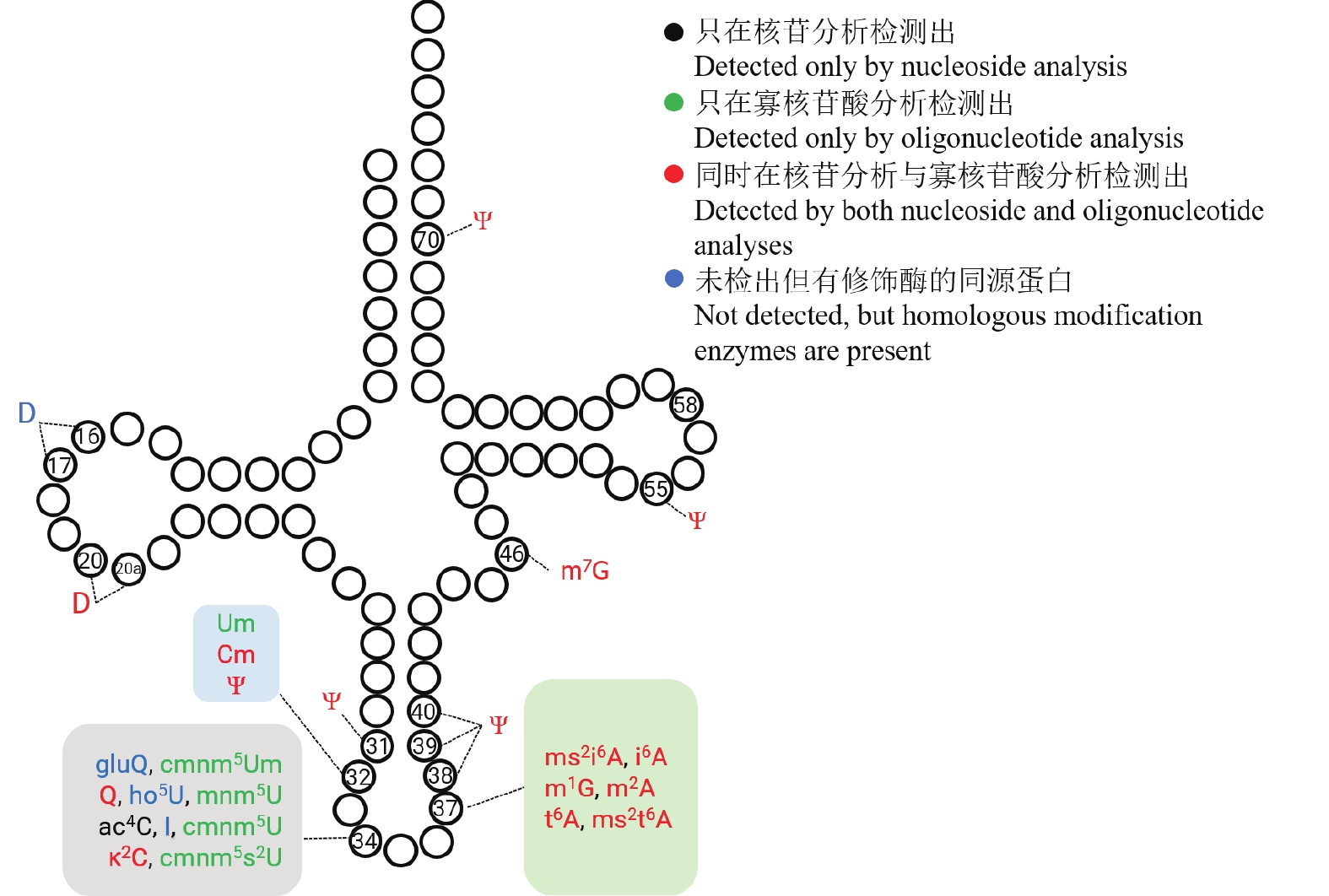
通过在巴氏醋杆菌蛋白组中检索并比对已报道的大肠杆菌/枯草芽孢杆菌tRNA修饰酶同源蛋白发现,尽管本研究的质谱数据尚未检出或定位到D16/17、gluQ34(glutamyl-queuosine)以及ho5U34(5-hydroxyuridine)等修饰,但巴氏醋杆菌基因组/蛋白组中存在对应的同源修饰酶DusB、GluQRS和TrhO。鉴于LC–MS检测存在一定检出限,同时细菌可能在不同生长条件下动态调节tRNA修饰丰度,综合酶学同源性证据与技术因素,初步推测巴氏醋杆菌中D16/17、I34、gluQ34及ho5U34等修饰具有潜在存在的可能。
-
综合核苷分析、寡核苷酸分析及同源比对结果,初步绘制了巴氏醋杆菌的tRNA修饰图谱(图5),系统呈现了21种修饰核苷的类型及其在tRNA上的位点分布特征。与多数真细菌一致,巴氏醋杆菌tRNA反密码子环上第34位和第37位包含的修饰类型最丰富,该区域修饰已被证明与翻译效率、翻译保真性和tRNA氨基酸酰化密切相关[28-29]。此外,巴氏醋杆菌在tRNA第31、32、38、39和40位存在相对密集的假尿苷(Ψ)分布,并在第46位检测到m7G修饰。
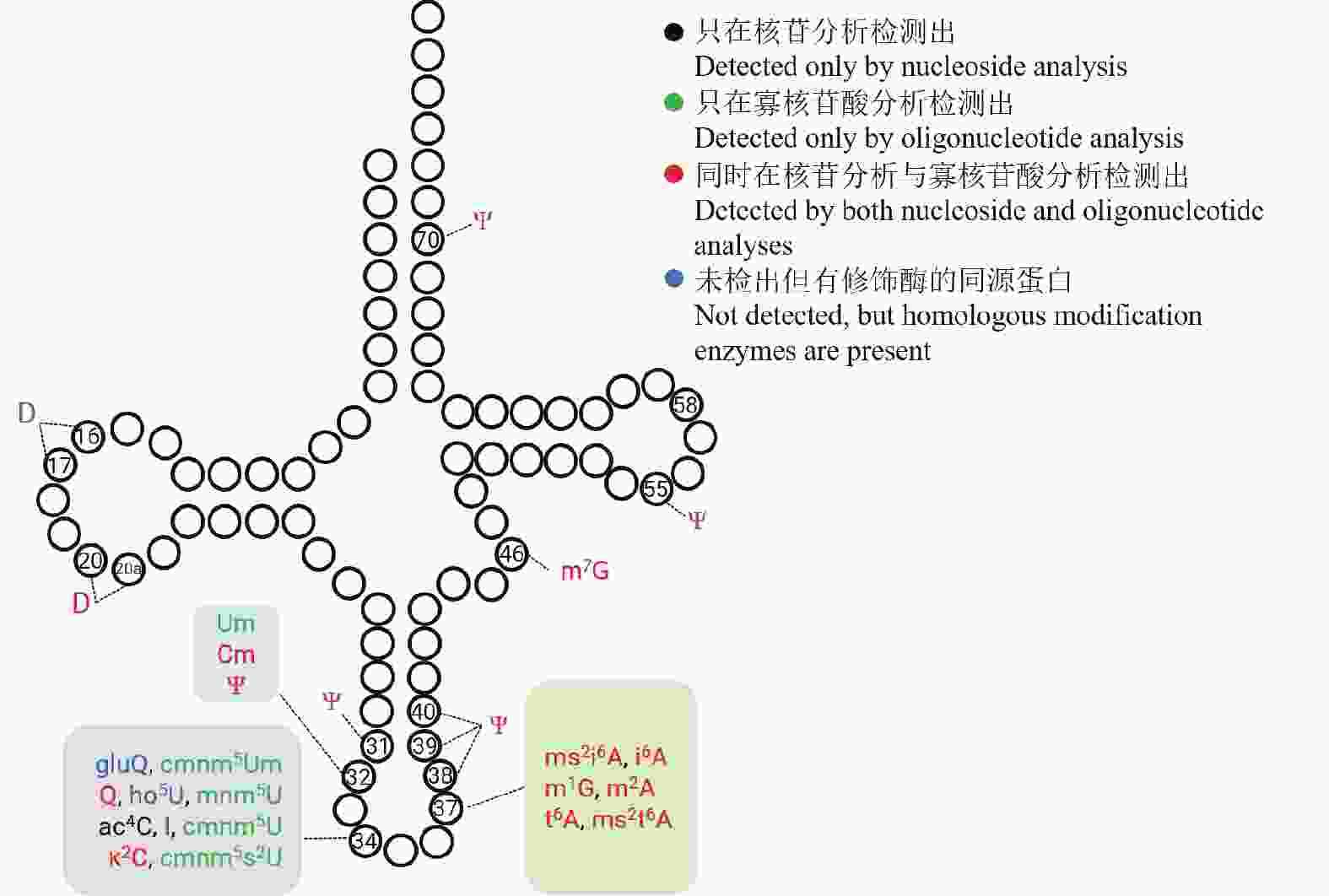
图 5 巴氏醋杆菌 tRNA 修饰图谱
Figure 5. Landscape of tRNA modifications in Acetobacter
-
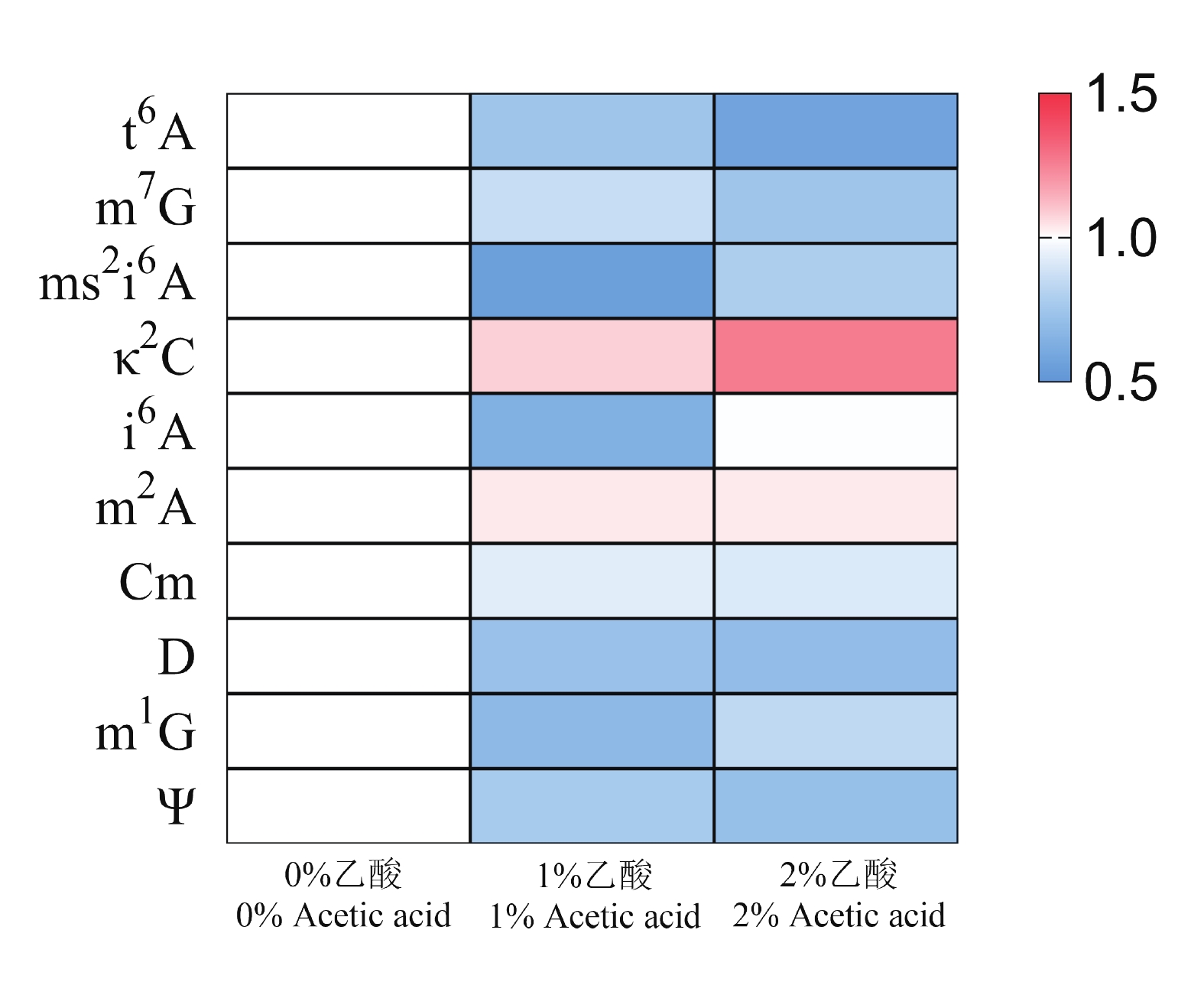
巴氏醋杆菌耐酸性能突出,即使在2%乙酸环境下仍可维持代谢活性与生长。尿苷(U)是RNA分子中的基础核苷之一,含量相对稳定,且在质谱分析中信号响应可靠,其峰面积可在一定程度上反映RNA总量,因此常被用作修饰核苷相对定量的归一化参照[30-31]。基于此,本研究采用三重四极杆质谱追踪1%与2%乙酸胁迫条件下巴氏醋杆菌tRNA中t6A、m7G、ms2i6A、κ2C、i6A、m2A、Cm、D、m1G及Ψ等修饰的信号强度,并以尿苷峰面积进行归一化处理,绘制获得tRNA修饰动态变化热图(图6)。热图结果显示,乙酸胁迫下κ2C水平呈剂量依赖性升高,而t6A、m7G、ms2i6A、D、m1G及Ψ等修饰则出现不同程度降低。上述下调可能与胁迫条件下整体代谢与能量供给受限有关:菌体通过重新分配资源,在保证生存与基本翻译需求的前提下降低部分修饰水平,仅维持满足tRNA功能与氨酰化效率所需的最低修饰率;而变化不明显的修饰则提示满足tRNA功能与氨酰化效率在胁迫状态下仍被优先维持,可能具有“house-keeping”性质。值得注意的是,κ2C的核心作用在于保障tRNAIle2(CAU)对AUA密码子的解码并且抑制对AUG的错误解码,而κ2C缺失会导致密码子误读并降低翻译效率[32]。因此,κ2C在酸胁迫下的上调提示巴氏醋杆菌可能通过提升AUA相关解码能力参与耐酸适应,甚至可能涉及AUA富集基因的表达调控。这一猜测需进一步实验验证。
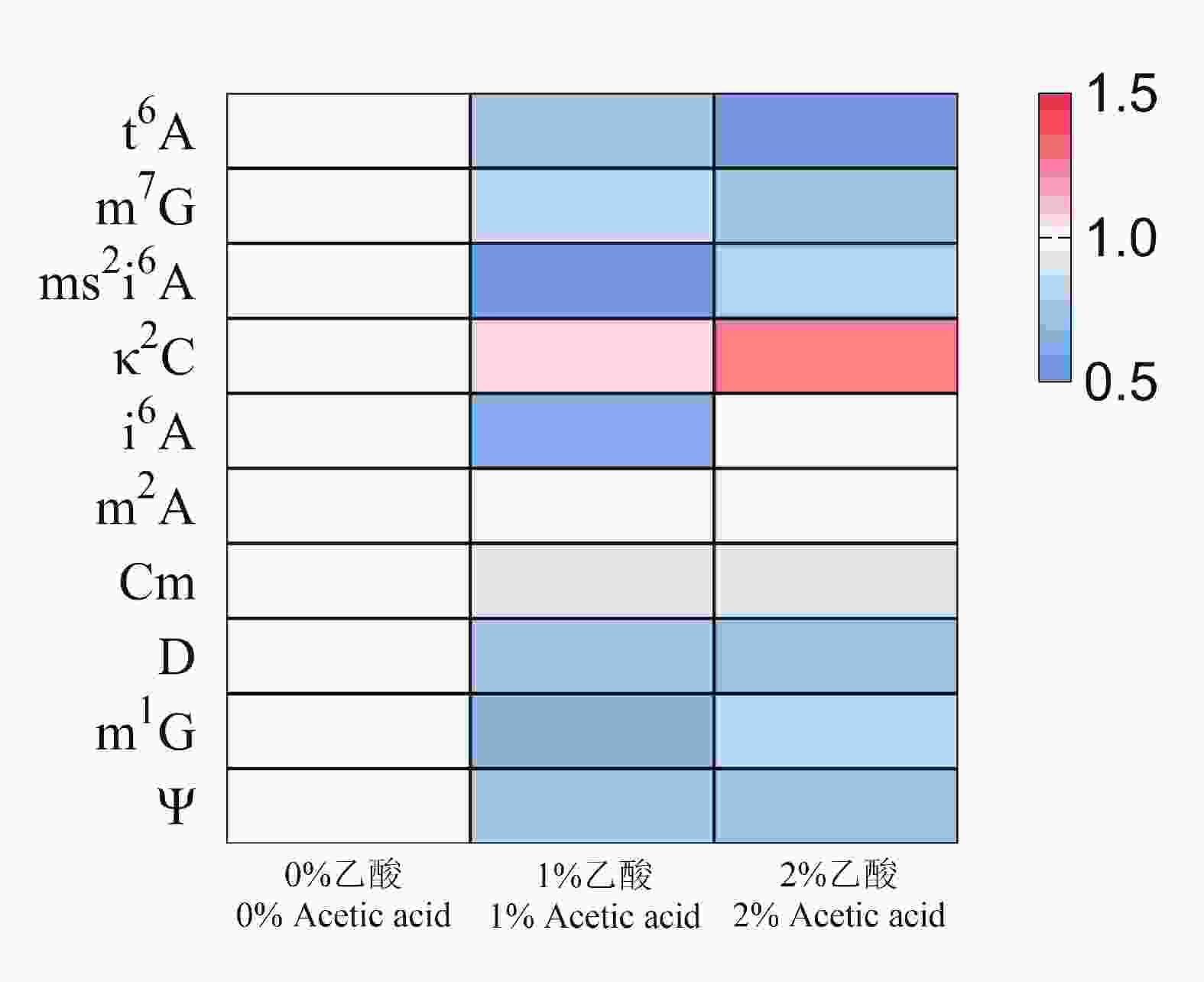
图 6 酸胁迫下tRNA修饰动态变化热图
Figure 6. Heatmap showing the dynamic response of tRNA modifications to acid stress
-
巴氏醋杆菌是醋酸发酵工业的核心菌种,其卓越的耐酸能力构成高效生产的关键基础。尽管已有研究从基因组特征、代谢重编程及细胞膜适应性等方面对巴氏醋杆菌耐酸机制进行了探讨,但从转录后修饰层面,尤其是tRNA修饰角度来解析巴氏醋杆菌环境适应策略的研究仍较为缺乏[33-35]。本研究首次对巴氏醋杆菌tRNA修饰图谱进行了初步绘制,鉴定获得21种修饰核苷并定位其位点分布,同时揭示了部分修饰在酸胁迫条件下的动态变化,为从表观转录组学视角理解巴氏醋杆菌耐酸生理机制提供了新的数据基础与研究线索。
在技术层面,本研究所采用的RNA修饰鉴定流程仍存在一定局限性,主要体现:①对非序列特异性寡核苷酸片段中检测到的修饰,难以准确归属到具体tRNA分子;②部分同分异构体(如Am与m1A)在寡核苷酸层面的质谱行为相近、区分困难,因而未纳入最终图谱;③对于丰度较低但基因组中存在同源修饰酶的修饰类型,受检出限影响可能未能在本研究条件下被检出。为进一步提升图谱的分辨率与准确性,后续可结合单体tRNA分离纯化策略,或引入具备修饰识别能力的tRNA测序方法(如m1A-seq、ICE-seq等)开展更深入解析,这也将是后续工作的重点方向。
-
本研究结合液相色谱–质谱联用(LC–MS)的核苷分析与寡核苷酸分析,并辅以同源比对策略,对巴氏醋杆菌tRNA修饰开展初步绘制与解析,共鉴定获得21种修饰核苷并实现其在tRNA上的位点定位。具体而言,D、Q、κ2C、Cm、Ψ、ms2i6A、i6A、m1G、m2A、t6A、ms2t6A及m7G在核苷分析与寡核苷酸分析中均被检出;ac4C仅在核苷分析中检出;而Um、cmnm5Um、mnm5U、cmnm5U及cmnm5s2U仅在寡核苷酸分析中检出。此外,基于同源比对结果推测I、gluQ与ho5U可能存在。进一步的酸胁迫实验显示,κ2C、t6A、m7G、ms2i6A、D、m1G与Ψ等修饰水平随环境pH降低发生特异性变化。上述结果提示,tRNA修饰可能通过影响翻译效率和/或翻译保真度参与巴氏醋杆菌对酸性环境的适应性应激调控。
Identification of modified nucleosides in tRNA of Acetobacter pasteurianus and the characteristics of response to acid stress
-
摘要: 巴氏醋杆菌(Acetobacter pasteurianus)是一类耐酸性强的工业微生物,广泛用于食醋、果醋和酸奶等产品的发酵。深入解析巴氏醋杆菌的生理生化机制,是开展高效合成生物学改造的重要基础。本研究基于液相色谱-质谱联用(LC-MS)技术的核苷分析与寡核苷酸分析,结合同源比对方法,对巴氏醋杆菌的tRNA修饰进行了初步绘制与解析,鉴定获得21种修饰核苷,并实现修饰核苷在tRNA分子上的位置定位。酸胁迫实验结果表明,随环境pH降低,κ2C、t6A、m7G、ms2i6A、D、m1G及Ψ等修饰水平发生特异性变化。上述结果提示,tRNA修饰可能通过影响翻译效率和/或翻译保真度参与巴氏醋杆菌对酸性环境的适应性应激调控,为阐明巴氏醋杆菌耐酸机制及后续理性改造提供了新的线索。Abstract: Acetobacter pasteurianus is an acid-tolerant industrial microorganism widely used in the fermentation of vinegar, fruit vinegar, and yogurt. A deeper understanding of its physiological and biochemical basis is essential for rational strain engineering in synthetic biology. A preliminary map of tRNA modifications in A. pasteurianus was generated by using LC-MS-based nucleoside and oligonucleotide analyses, complemented by homology searches for putative tRNA-modifying enzymes. There were 21 modified nucleosides identified, and their positions within tRNA molecules were mapped. Acid-stress experiments further showed that κ2C, t6A, m7G, ms2i6A, D, m1G, and Ψ exhibit specific, pH-dependent changes as environmental pH decreases. These results suggest that tRNA modifications may contribute to acid adaptation in A. pasteurianus, potentially by tuning translational efficiency and/or fidelity.
-
Key words:
- Acetobacter pasteurianus /
- tRNA modification landscape /
- RNA-MS /
- acid stress
-
图 4 寡核苷酸分析及修饰核苷定位示意图
Fig. 4 Schematic diagram of oligonucleotide analysis and modified nucleoside localization
图 6 酸胁迫下tRNA修饰动态变化热图
Fig. 6 Heatmap showing the dynamic response of tRNA modifications to acid stress
表 1 巴氏醋杆菌tRNA序列独特性片段
Table 1 Unique tRNA fragments identified from Acetobacter pasteurianus
转运RNA
tRNA序列独特性片段
Unique fragments质荷比
m/z电荷
zMet-CAU AA[Um]CAU[t6A]AUCCCUUGp 1638.212 −3 UUCAAAUCCUUCCGp 1467.506 −3 Tyr-GUA U[i6A]AAUCCGp 1319.196 −2 U[ms2i6A]AAUCCGp 1342.190 −2 pGGAGGGAUp 1392.662 −2 AAAGGGGGCp 1516.714 −2 His-GUG UUUQUGp 1028.128 −2 Ala-CGC CAAUGp 815.106 −2 AUUCCCCUCAGp 1731.213 −2 CUCCACCA 1212.681 −2 GAGGAG[m7G]Up 1359.687 −2 Asp-GUC CCUQUC[m2A]CGp 1503.719 −2 Trp-CCA U[Cm]UCCA[i6A]AACCGp 1948.291 −2 U[Cm]UCCA[ms2i6A]AACCGp 1971.285 −2 ACCCUCCACCCCUGp 1458.519 −3 Pro-CGG AAUCUUCUCACUCCGp 1569.187 −3 AGGG[m7G]Cp 1022.146 −2 Lys-CUU A[Cm]UCUU[ms2t6A]AUCAGp 1340.168 −3 [m7G]UCCAAGp 1147.157 −2 Gly-GCC CACAACCUUGp 1590.206 −2 AAUCCCAUCGp 1590.206 −2 Ile-GAU AUAAGp 827.111 −2 UCCUCCCUGp 1402.160 −2 Ala-UGC UCUCCACCA 1365.693 −2 AACCCGp 967.134 −2 CUU[mnm5U]Gp 813.599 −2 AGGGG[m7G]Up 1195.161 −2 fMet-CAU [Cm]UCAU[m2A]ACCUGp 1757.234 −2 UUCAAAUCCUGp 1743.710 −2 Lys-UUU ACU[cmnm5s2U]UU[ms2t6A]AUCGp 1891.219 −2 Ser-GGA UUAAUAGp 1133.136 −2 Arg-CCG CC[Cm]UCCGp 1103.143 −2 Asn-GUU ACUQUU[t6A]AUCGp 1899.266 −2 UCCUACUUCGp 1567.178 −2 Ser-CGA A[ms2i6A]AACCGp 1200.691 −2 Ser-UGA UDDAAGp 970.626 −2 AAGGAACp 1163.669 −2 UCU[mnm5U]Gp 813.599 −2 A[ms2i6A]AACCAGp 1365.217 −2 AAUCCCACCCCAUCCGp 1677.887 −3 Pro-UGG [m1G]UACACGp 1147.157 −2 U[Um]UUGp 799.578 −2 Arg-UCU CCU[cmnm5U]CU[t6A]AGp 1542.206 −2 CCU[cmnm5Um]CU[t6A]AGp 1549.214 −2 Pro-GGG GGG[m1G]GACp 1194.669 −2 Leu-UAG AUUUAGp 968.610 −2 [m1G]UUCUGp 971.610 −2 UCCCUCCACCCGp 1871.235 −2 Val-UAC UUUACACCGp 1426.172 −2 Gln-CUG AACUACCGp 1284.673 −2 [Cm]UCUGp 798.594 −2 Cys-GCA CA[i6A]AUCCGp 1318.704 −2 CA[ms2i6A]AUCCGp 1341.698 −2 AGAAUp 827.111 −2 Val-CAC CC[Um]UCACACGp 1585.208 −2 CACCCACCA 1376.715 −2 Leu-CAG [m1G]UACCUGp 1135.644 −2 UUUCAGp 956.604 −2 UCUUCUUCUGp 1556.157 −2 Phe-GAA AA[ms2i6A]AUCGp 1201.183 −2 Glu-UUC CCDAGp 804.108 −2 CCCU[cmnm5s2U]UC[m2A]CGp 1625.199 −2 Arg-ACG ACUACGp 967.626 −2 [m2A]AUCUGp 975.126 −2 Thr-CGU UCAUUCGp 1109.125 −2 Ser-GCU AAUCCCUUCCCCUCCGp 1662.535 −3 CU[t6A]AACCGp 1357.191 −2 Thr-GGU CACUCCCUUGp 1566.686 −2 UUCAAUCCUGp 1579.184 −2 Thr-UGU UUCAAAUCCUACUGp 1475.510 −3 Gln-UUG AUU[cmnm5s2U]UGp 1008.601 −2 AAUCCUCCCGp 1578.200 −2 Ala-GGC CAUUCAGp 1120.639 −2 Leu-CAA UCCCCUCUCUCGp 1860.214 −2 ACUCAA[ms2i6A]AUCGp 1823.763 −2 Gly-UCC AUUCCCUCUGp 1567.178 −2 CCU[cmnm5U]CC[m2A]AGp 1476.203 −2 GAGGAGGGUp 1525.203 −2 Ile2-CAU CU[κ2C]AU[m2A]ACGp 1508.741 −2 Val-GAC CACUAUCUUGp 1579.184 −2 Gly-CCC C[Um]UCCCAAGp 1432.687 −2 UUCAAUGp 1121.131 −2 Leu-GAG AAUACCCCGp 1437.193 −2 UCUUCUCGp 1250.132 −2  下载: 导出CSV
下载: 导出CSV
表 2 巴氏醋杆菌tRNA中Ψ的定位
Table 2 Mapping of Ψ in Acetobacter pasteurianus tRNAs
转运RNA
tRNA序列独特性片段
Unique fragments质荷比
m/z电荷
z位置
PositionTrp-CCA [Ψ-CE][Cm]UCCA[i6A]AACCGp 1974.805 −2 31 Leu-UAG A[Ψ-CE]UUAGp 995.123 −2 32 Pro-UGG [m1G][Ψ-CE]ACACGp 1173.671 −2 38 Met-CAU AA[Um]CAU[t6A]A[Ψ-CE]CCCUUGp 1655.887 −3 39 Tyr-GUA U[i6A]AA[Ψ-CE]CCGp 1345.709 −2 40 Met-CAU U[Ψ-CE]CAAAUCCUUCCGp 1485.182 −3 55 Ala-UGC UC[Ψ-CE]CCACCA 1392.207 −2 70
下载: 导出CSV
-
[1] Cappannini A, Ray A, Purta E, et al. MODOMICS: a database of RNA modifications and related information [J]. Nucleic Acids Research, 2024, 52(D1): D239−D244. https://doi.org/10.1093/nar/gkad1083 doi: 10.1093/nar/gkad1083 [2] Zhang W, Foo M, Eren A M, et al. tRNA modification dynamics from individual organisms to metaepitranscriptomics of microbiomes [J]. Molecular Cell, 2022, 82(5): 891−906. https://doi.org/10.1016/j.molcel.2021.12.007 doi: 10.1016/j.molcel.2021.12.007 [3] Suzuki T. The expanding world of tRNA modifications and their disease relevance [J]. Nature Reviews Molecular Cell Biology, 2021, 22(6): 375−392. https://doi.org/10.1038/s41580-021-00342-0 doi: 10.1038/s41580-021-00342-0 [4] Roundtree I A, Evans M E, Pan T, et al. Dynamic RNA modifications in gene expression regulation [J]. Cell, 2017, 169(7): 1187−1200. https://doi.org/10.1016/j.cell.2017.05.045 doi: 10.1016/j.cell.2017.05.045 [5] Motorin Y, Helm M. RNA nucleotide methylation: 2021 update [J]. WIREs RNA, 2022, 13(1): e1691. https://doi.org/10.1002/wrna.1691 doi: 10.1002/wrna.1691 [6] Lorenz C, Lünse C E, Mörl M. tRNA modifications: impact on structure and thermal adaptation [J]. Biomolecules, 2017, 7(2): 35. https://doi.org/10.3390/biom7020035 doi: 10.3390/biom7020035 [7] Zhang Y, Zhou J B, Yin Y, et al. Multifaceted roles of t6A biogenesis in efficiency and fidelity of mitochondrial gene expression [J]. Nucleic Acids Research, 2024, 52(6): 3213−3233. https://doi.org/10.1093/nar/gkae013 doi: 10.1093/nar/gkae013 [8] Vinayak M, Pathak C. Queuosine modification of tRNA: its divergent role in cellular machinery [J]. Bioscience Reports, 2010, 30(2): 135. https://doi.org/10.1042/BSR20090057 doi: 10.1042/BSR20090057 [9] Gupta R, Laxman S. tRNA wobble-uridine modifications as amino acid sensors and regulators of cellular metabolic state [J]. Current Genetics, 2020, 66(3): 475−480. https://doi.org/10.1007/s00294-019-01045-y doi: 10.1007/s00294-019-01045-y [10] Basavappa R, Sigler P B. The 3 A crystal structure of yeast initiator tRNA: functional implications in initiator/elongator discrimination [J]. The EMBO Journal, 1991, 10(10): 3105−3111. https://doi.org/10.1002/j.1460-2075.1991.tb07864.x doi: 10.1002/j.1460-2075.1991.tb07864.x [11] Yared M J, Yoluç Y, Catala M, et al. Different modification pathways for m1A58 incorporation in yeast elongator and initiator tRNAs [J]. Nucleic Acids Research, 2023, 51(19): 10653−10667. https://doi.org/10.1093/nar/gkad722 doi: 10.1093/nar/gkad722 [12] Ohira T, Suzuki T. Transfer RNA modifications and cellular thermotolerance [J]. Molecular Cell, 2024, 84(1): 94−106. https://doi.org/10.1016/j.molcel.2023.11.041 doi: 10.1016/j.molcel.2023.11.041 [13] Torres A G, Piñeyro D, Filonava L, et al. A-to-I editing on tRNAs: biochemical, biological and evolutionary implications [J]. FEBS Letters, 2014, 588(23): 4279−4286. https://doi.org/10.1016/j.febslet.2014.09.025 doi: 10.1016/j.febslet.2014.09.025 [14] Torres A G, Piñeyro D, Rodríguez-Escribà M, et al. Inosine modifications in human tRNAs are incorporated at the precursor tRNA level [J]. Nucleic Acids Research, 2015, 43(10): 5145−5157. https://doi.org/10.1093/nar/gkv277 doi: 10.1093/nar/gkv277 [15] 王娜. 巴氏醋杆菌对环境胁迫的生理响应机制及提高冻干存活率的研究[D]. 秦皇岛: 河北科技师范学院, 2015. doi: 10.7666/d.Y2858581 [16] Wang B, Shao Y C, Chen T, et al. Global insights into acetic acid resistance mechanisms and genetic stability of Acetobacter pasteurianus strains by comparative genomics [J]. Scientific Reports, 2015, 5: 18330. https://doi.org/10.1038/srep18330 doi: 10.1038/srep18330 [17] Fruchard L, Salinas C, Carvalho A, et al. tRNA-modifying enzymes in bacterial stress adaptation [J]. Open Biology, 2025, 15(10): 250194. https://doi.org/10.1098/rsob.250194 doi: 10.1098/rsob.250194 [18] 赖军, 张越冉, 周海鸿, 等. 基于代谢组学的重要热带植物的研究进展[J]. 热带生物学报, 2023, 14(3): 248−258. https://doi.org/10.15886/j.cnki.rdswxb.2023.03.002 doi: 10.15886/j.cnki.rdswxb.2023.03.002 [19] 张忠辉, 邓渊, 强奇, 等. 基于高效液相色谱-质谱联用技术解析山栏稻营养品质的代谢物谱[J]. 热带生物学报, 2021, 12(4): 419−427. https://doi.org/10.15886/j.cnki.rdswxb.2021.04.003 doi: 10.15886/j.cnki.rdswxb.2021.04.003 [20] 朱昆宇, 潘成才, 吉怡颖, 等. 盐胁迫下冰菜盐囊泡细胞的比较代谢组学分析[J]. 热带生物学报, 2024, 15(2): 224−231. https://doi.org/10.15886/j.cnki.rdswxb.20230020 doi: 10.15886/j.cnki.rdswxb.20230020 [21] 范煜, 何桢锐, 黄晓彤, 等. 基于多组学的真菌病毒与寄主真菌互作的研究进展[J]. 热带生物学报, 2023, 14(4): 399−404,440. https://doi.org/10.15886/j.cnki.rdswxb.2023.04.007 doi: 10.15886/j.cnki.rdswxb.2023.04.007 [22] Qin Y C, Zhong Q S, Zhang Y, et al. Micro-flow hydrophilic interaction liquid chromatography coupled with triple quadrupole mass spectrometry detects modified nucleosides in the transfer RNA pool of cyanobacteria [J]. Journal of Separation Science, 2021, 44(17): 3208−3218. https://doi.org/10.1002/jssc.202100417 doi: 10.1002/jssc.202100417 [23] Ross R, Cao X Y, Yu N X, et al. Sequence mapping of transfer RNA chemical modifications by liquid chromatography tandem mass spectrometry [J]. Methods, 2016, 107: 73−78. https://doi.org/10.1016/j.ymeth.2016.03.016 doi: 10.1016/j.ymeth.2016.03.016 [24] Douthwaite S, Kirpekar F. Identifying modifications in RNA by MALDI mass spectrometry [J]. Methods in Enzymology, 2007, 425: 1,3−20. https://doi.org/10.1016/S0076-6879(07)25001-3 doi: 10.1016/S0076-6879(07)25001-3 [25] 张耀元, 刘玲, 周秀娟, 等. 转录组和代谢组揭示独脚金内酯缺失影响水稻根和叶中的脂质和黄酮代谢[J]. 热带生物学报, 2020, 11(4): 415−425. https://doi.org/10.15886/j.cnki.rdswxb.2020.04.004 doi: 10.15886/j.cnki.rdswxb.2020.04.004 [26] Mengel-Jørgensen J, Kirpekar F. Detection of pseudouridine and other modifications in tRNA by cyanoethylation and MALDI mass spectrometry [J]. Nucleic Acids Research, 2002, 30(23): e135. https://doi.org/10.1093/nar/gnf135 doi: 10.1093/nar/gnf135 [27] Yang W Q, Xiong Q P, Ge J Y, et al. THUMPD3-TRMT112 is a m2G methyltransferase working on a broad range of tRNA substrates [J]. Nucleic Acids Research, 2021, 49(20): 11900−11919. https://doi.org/10.1093/nar/gkab927 doi: 10.1093/nar/gkab927 [28] Rapino F, Zhou Z L, Roncero Sanchez A M, et al. Wobble tRNA modification and hydrophilic amino acid patterns dictate protein fate [J]. Nature Communications, 2021, 12(1): 2170. https://doi.org/10.1038/s41467-021-22254-5 doi: 10.1038/s41467-021-22254-5 [29] Agris P F, Narendran A, Sarachan K, et al. The importance of being modified: the role of RNA modifications in translational fidelity [J]. Enzymes, 2017, 41: 1−50. https://doi.org/10.1016/bs.enz.2017.03.005 doi: 10.1016/bs.enz.2017.03.005 [30] Deng D H, Qin Y C, Lin X Y, et al. Unveiling transfer RNA modifications of oil palm and their dynamic changes during fruit ripening [J]. BMC Plant Biology, 2025, 25(1): 398. https://doi.org/10.1186/s12870-025-06426-9 doi: 10.1186/s12870-025-06426-9 [31] Yao C, Lv D Z, Zhou X Q, et al. Exploring urinary modified nucleosides as biomarkers for diabetic retinopathy: Development and validation of a ultra performance liquid chromatography-tandem mass spectrometry method [J]. Journal of Chromatography B, 2024, 1232: 123968. https://doi.org/10.1016/j.jchromb.2023.123968 doi: 10.1016/j.jchromb.2023.123968 [32] Soma A, Ikeuchi Y, Kanemasa S, et al. An RNA-modifying enzyme that governs both the codon and amino acid specificities of isoleucine tRNA [J]. Molecular Cell, 2003, 12(3): 689−698. https://doi.org/10.1016/s1097-2765(03)00346-0 doi: 10.1016/s1097-2765(03)00346-0 [33] Zheng Y, Zhang R K, Yin H S, et al. Acetobacter pasteurianus metabolic change induced by initial acetic acid to adapt to acetic acid fermentation conditions [J]. Applied Microbiology and Biotechnolog, 2017, 101(18): 7007−7016. https://doi.org/10.1007/s00253-017-8453-8 doi: 10.1007/s00253-017-8453-8 [34] Gao L, Shi W, Xia X L. Genomic plasticity of acid-tolerant phenotypic evolution in Acetobacter pasteurianus [J]. Applied Biochemistry and Biotechnology, 2023, 195(10): 6003−6019. https://doi.org/10.1007/s12010-023-04353-9 doi: 10.1007/s12010-023-04353-9 [35] Gao L, Wu X D, Li C Y, et al. Exploitation of strong constitutive and stress-driven promoters from Acetobacter pasteurianus for improving acetic acid tolerance [J]. Journal of Biotechnology, 2022, 350: 24−30. https://doi.org/10.1016/j.jbiotec.2022.03.013 doi: 10.1016/j.jbiotec.2022.03.013 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 230
- HTML全文浏览量: 137
- 被引次数: 0
