


-
篦齿苏铁(Cycas pectinata)作为国家一级保护植物,对研究植物演化与生态适应具有重要价值[1]。雌雄异株植物的性别差异常体现在形态、生理及资源分配等方面[2-5],其中,茎干特征差异是其生存策略的直观体现[6-7]。南宁植物园篦齿苏铁迁地古树群是中国最大的篦齿苏铁迁地保护种群[8],探究其雌雄株茎干特征,对优化苏铁植物种植配置、提升迁地保护成效具有现实意义。目前,苏铁植物性别差异研究多集中于野生种群[9-11],对迁地保护古树群的系统性分析较少。本研究通过解析该古树群雌雄株茎干形态差异,为篦齿苏铁科学养护与种群复壮提供理论支撑。
-
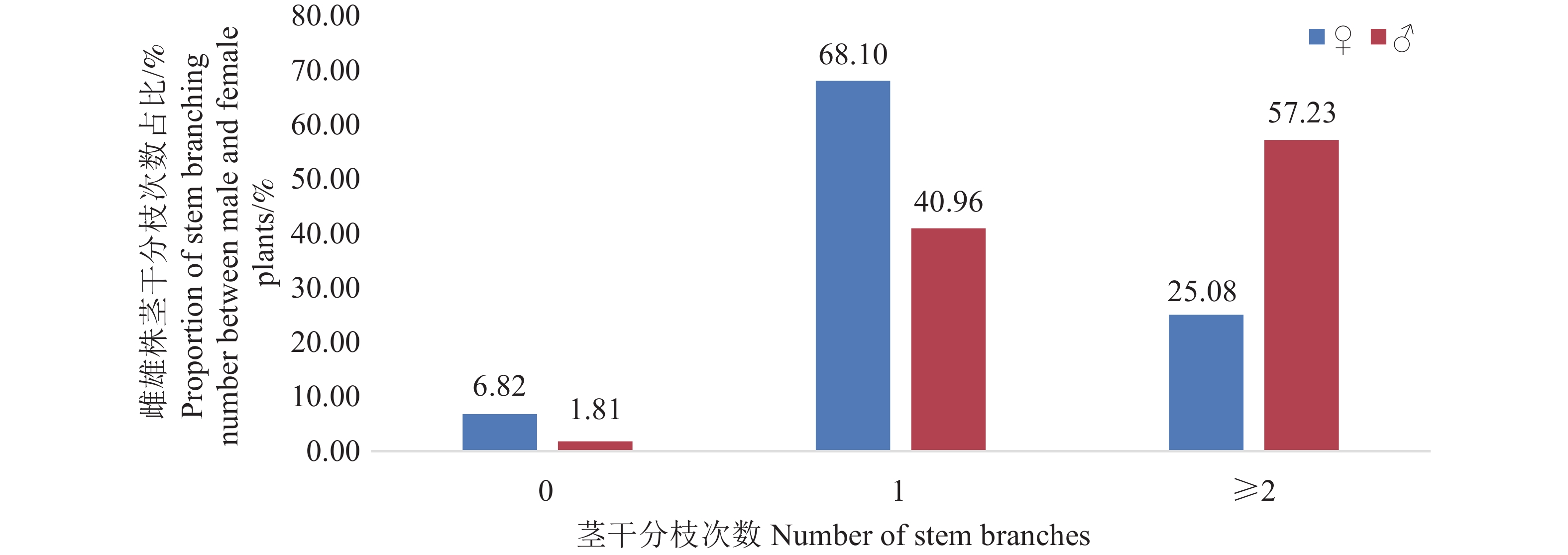
研究区域位于南宁植物园(青秀山)苏铁园(22° 79′ N,108° 39′ E),属亚热带季风气候,年均温21.6 ℃,雨量充沛。该园始建于1997年,以抢救性迁地保护散落于民间的苏铁类植物为目标,后经过不断补充、提升和完善,形成集科学研究、迁地保育、科普教育、旅游观光等多功能于一体的植物专类园,现保存有210株篦齿苏铁古树,依据《广西第二次古树名木资源普查结果》,该种群涵盖特级古树(≥1 000年,13株),一级古树(500~999年,104株),二级古树(300~499年,71株),三级古树(100~299年,22株)。由于苏铁类植物无木质部年轮结构,树龄主要通过叶痕生长节律观测、生长速率模拟及实践经验综合推断[12],难以获得准确生物学年龄,树龄分级受主观因素影响,因此,本研究将种群视为树龄≥100年的整体开展分析,暂不区分年龄级别讨论形态差异。
古树群分布面积2.8 hm2,以树龄最老的“苏铁王”为核心,植株密度最高;向四周道路沿线逐渐稀疏,形成 “中心密集、边缘离散”的集群模式(图1)。密集区植株间距2.5~8.0 m,边缘区植株间距15~20 m,生长环境(光照、土壤肥力、水分条件等)基本保持一致。

图 1 研究区域篦齿苏铁古树群分布图
Figure 1. Distribution of ancient plant Cycas pectinata
-
参照离心式方法,将茎干整体从下至上,主干定为1级,第一次分叉之后的茎干定为2级枝,第二次分叉之后的茎干定为3级枝(图2)[13-15]。

图 2 分枝解析图
Figure 2. Branch diagram
-
1)基础形态指标测定。使用胸径钢卷尺(精度0.1 cm)测量植株高度(茎顶至地面垂直距离)、地茎(距地面5 cm处茎干直径)、胸径(距地面1.3 m处茎干直径)及平均冠幅(东西与南北冠幅的算术平均值)。
2)分枝特征指标测定。
分枝长度:测量各级分枝基部至梢端的直线距离。
枝径:使用胸径尺(精度0.1 cm)测量各级分枝基部直径。
枝级数量:统计每株1~3级分枝的数量。
生长点与吸芽:记录茎顶末级生长点数量及茎干的吸芽数量。
-
基于上述数据,计算以下分枝特征参数。
$$ \mathrm{OBR=(} {Nt} - {Ns} )/( {Nt} -{N} _{1} ), $$ 式中,OBR 为总体分枝率(overall bifurcation ratio);Nt=∑Ni,表示所有枝级中的枝条总数;Ns为最高级分枝条数;N1为第1级的枝条数。
$$ \mathrm{SBR}_{{i:i} {+1}} = {N} _{i} /N _{i+1}, $$ 式中,SBR为逐步分枝率(stepwise bifurcation ratio);Ni和Ni+1分别是第i和第i+1级分枝的枝条总数。
$$ \mathrm{RBD=} \mathit{D} _{ \mathit{i} \mathrm{+1}} \mathrm{/} \mathit{D} _{ \mathit{i} } \mathrm{。} $$ 式中,RBD为枝径比(ratio of branch diameter); Di和Di+1分别是第i和第i+1级分枝的枝条直径(cm)。
-
采用SPSS26.0进行统计分析:卡方检验雌雄比例显著性;Kolmogorov-Smirnov检验数据正态性,正态分布数据用独立样本t检验,非正态分布用Mann-Whitney U检验;Pearson(正态)或Spearman(非正态)分析相关性。
-
南宁植物园篦齿苏铁古树群共210株,2018~2024年通过连续花期观测明确各株性别:雌株44株(20.95%),雄株166株(79.05%),雌雄比1:3.77,种群极显著偏雄(χ2=70.876,P<0.01)。
-
雄株(n=166)在茎高、平均冠幅、地径及胸径的均值表现上均优于雌株(n=44)。其中,雄株茎高平均值为452.92 cm,显著高于雌株的371.36 cm,且雄株茎高最大值达
1030.00 cm,最小值为220.00 cm,变异范围明显大于雌株(最大值510.00 cm,最小值250.00 cm)。地径与胸径指标中,雄株均值分别为71.02 cm和49.77 cm,均高于雌株的67.82 cm、48.55 cm,且雄株地径最大值(127.00 cm)与胸径最大值(95.00 cm)亦大于雌株对应值(105.00 cm、84.10 cm),显示雄株基部及主干生长更为粗壮(表1)。表 1 南宁植物园篦齿苏铁古树群雌雄株基本形态概况
Table 1. Basic information of ancient plant C. pectinata in Nanning Botanical Garden
指标Index /性别Sex ♂(n=166) ♀(n=44) 平均 Average 最大 Max. 最小 Min. 平均 Average 最大 Max. 最小 Min. 茎高 Stem height/cm 452.92±149.14* 1030.00 220.00 371.36±65.57* 510.00 250.00 平均冠幅 Average Crown Diameter/cm 432.29±75.25 650.00 200.00 447.27±67.85 575.00 320.00 地径 Ground Diameter/cm 71.02±16.81 127.00 41.00 67.82±14.34 105.00 46.00 胸径 Diameter at Breast Height/cm 49.77±15.13 95.00 28.00 48.55±14.17 84.10 23.50 注:*表示相关性显著在0.05级别;**表示相关性显著在0.01级别;下同。 Note: * indicates significant correlation at the 0.05 level; ** indicates significant correlation at the 0.01 level; Similarly hereinafter. -
对篦齿苏铁古树群雌雄株基本形态指标的两两相关性分析表明(表2),雄株中,茎高与平均冠幅呈显著正相关(r=0.166*),与胸径(r=0.383**)、地径(r=0.339**)均呈极显著正相关;平均冠幅与胸径(r=0.433**)、地径(r=0.505**),胸径与地径(r=0.743**)亦存在极显著正相关。雌株中,平均冠幅与茎高呈显著负相关(r=−0.320*),与胸径(r=0.660**)、地径(r=0.476**)呈极显著正相关;胸径与地径(r=0.741**)同样存在极显著正相关关系,表明雌雄株形态指标间的相关性存在差异,雄株整体形态指标协同生长特征更显著。
表 2 篦齿苏铁古树群雌雄株基本形态指标的两两相关性
Table 2. Correlation between basic morphological indicators of ancient plant C. pectinata
形态指标
Morphological indices茎高
Stem height平均冠幅
Average crown diameter胸径
Diameter at breasth height地径
Stem diameter at
the ground level♂ ♀ ♂ ♀ ♂ ♀ ♂ ♀ 茎高Stem height * * ** ** 平均冠幅
Average Crown Diameter0.166* −0.320* * ** ** ** 胸径
Diameter at Breast Height0.383** − 0.0188 0.433** 0.660** ** ** 地径Ground Diameter 0.339** −0.100 0.505** 0.476** 0.743** 0.741** -
从雌雄株茎干分枝次数占比(图3)可见,雌株1次分枝占比(68.10%)显著高于雄株(40.96%),而雄株≥2次分枝占比(57.23%)明显高于雌株(25.00%),0次分枝占比雌雄株均较低(雌株6.82%,雄株1.81%)。该结果表明,篦齿苏铁古树群中雌株更倾向于较少次数的分枝模式,而雄株更易发生多次分枝,雌雄株在分枝习性上存在性别差异。
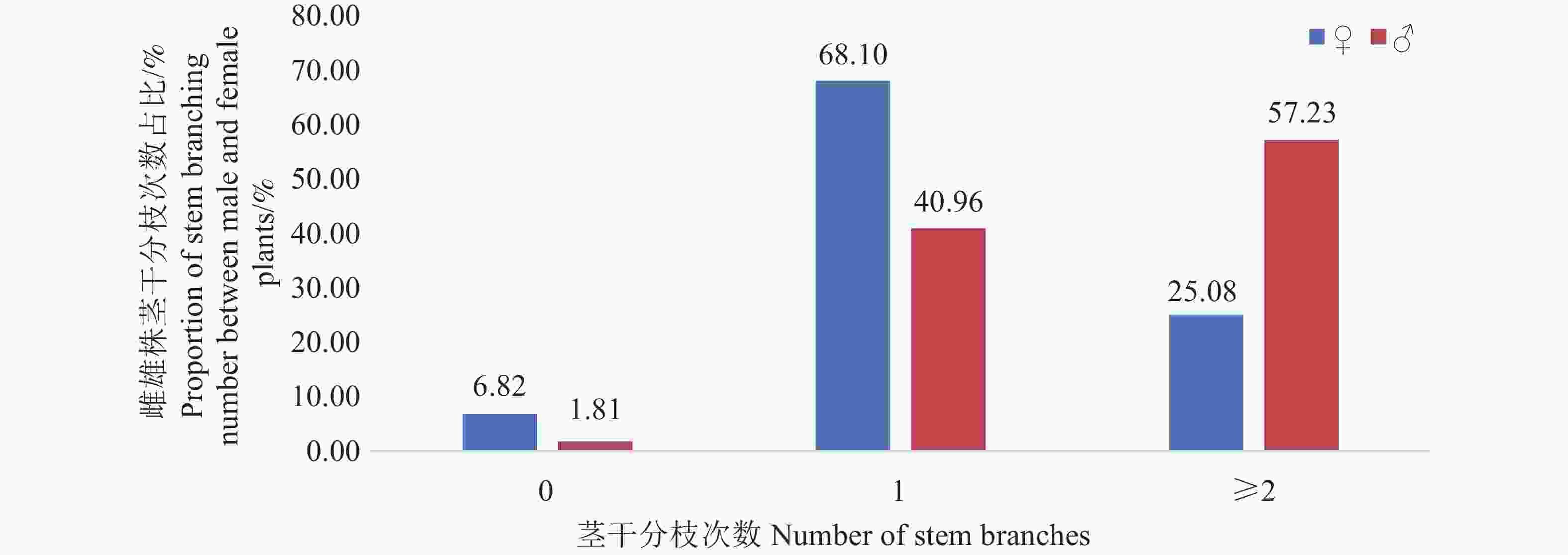
图 3 篦齿苏铁古树雌雄株茎干分枝次数差异
Figure 3. Differences in stem branching times between male and female of ancient plant C. pectinata
-
雌雄株的分枝长度均表现为1级分枝>2级分枝>3级分枝,且雄株各级分枝长度均显著大于雌株(表3)。且雄株各级分枝长度显著大于雌株。例如,1级分枝长度雄株为242.20 cm,雌株为194.34 cm;3级分枝长度雄株(61.31 cm)与雌株(17.72 cm)差异极显著。
表 3 分枝式样特征统计
Table 3. The branching characteristics of ancient plant C. pectinata in male and female plants
测定指标 Testing indices ♂(n=166) ♀(n=44) Z P 1级平均分枝长度 Average length of 1st-order branches/cm 242.20±131.74 194.34±110.37 −2.247 0.025* 2级平均分枝长度 Average length of 2nd-order branches/cm 176.28±139.89 168.49±93.14 −0.391 0.696 3级平均分枝长度 Average length of 3rd-order branches/cm 61.31±86.73 17.72±38.22 −3.294 0.001** 总分枝率 Overall bifurcation ratio 0.57±0.21 0.51±0.21 −1.156 0.248 逐步分枝率2:1 Stepwise bifurcation ratio 2:1 2.29±0.95 1.91±0.72 −1.991 0.046* 逐步分枝率3:2 Stepwise bifurcation ratio 3:2 0.86±0.98 0.32±0.60 −3.107 0.002** 枝径比2:1 Ratio of branch diameter (RBD2:1) 0.66±0.19 0.63±0.22 −0.877 0.381 枝径比3:2 Ratio of branch diameter (RBD3:2) 0.33±0.31 0.15±0.28 −2.872 0.004** 2级数量/个 Number of 2nd-order branches/unit 2.29±0.95 1.91±0.72 −1.991 0.046* 3级数量/个 Number of 3rd-order branches/unit 1.89±2.13 0.55±1.03 −3.512 0.000** 吸芽数量/个 Number of suckers/unit 0.78±1.48 1.52±2.43 −2.004 0.045* 末端生长点/个 Number of terminal growing points/unit 2.92±1.58 2.06±0.86 −3.140 0.002* 总生长点数量/个 Total number of growing points/unit 3.69±0.23 2.03±0.13 −4.576 0.000** -
雄株逐步分枝率(SBR2:1=2.29,SBR3:2=0.86)显著高于雌株(SBR2:1=1.91,SBR3:2=0.32);雄株枝径比(RBD3:2=0.33)显著高于雌株(RBD3:2=0.15),表明雄株分枝结构的承载能力更具优势。
-
雄株在2级枝数量、3级枝数量、末端生长点及总生长点数量上均显著多于雌株;而雌株吸芽数量显著高于雄株,体现雌雄株在分枝组成在营养繁殖特性上的分化(表3)。
-
自然种群中雌雄异株植物的雌雄个体的性别比例通常为1:1[16],但环境压力的增大会导致种群中雌雄比例出现明显偏差[17-19]。南宁植物园篦齿苏铁迁地古树群的性别比例表现出极显著的雄性偏倚(1:3.77),χ2检验表明该比例严重偏离理论值(P<0.01),与曾辉等[1]报道的野生篦齿苏铁种群(雌雄比4:1)、黄应峰等[20]研究的仙湖苏铁种群(雌雄比6.6:1)存在显著差异。值得关注的是,本研究种群为迁地保护的古树群,其性别比例显著偏雄的主要原因可能是引种时的偶然性,即早期收集和移植过程中,无意中引入了更多雄株。尽管存在这种人为引入造成的性别比例偏倚,篦齿苏铁仍表现出在自然条件下恢复性别平衡的潜力。本研究基于深圳市仙湖植物园[21]发明专利技术,对5株雌株的100株F1小苗进行分子性别鉴定,发现后代雌雄比遵循1:1(P>0.05)。这表明篦齿苏铁在繁殖过程中性别决定机制并未受到根本影响,自然界具备通过有性繁殖实现性别比例自我纠正的能力。
除引种偶然性外,历史生境胁迫也可能对性别比例产生一定影响。迁地保护前的野生生境中,雌株会将更多的生物量分配到根系和果实中,而雄性植物可能更多地分配到茎和叶的生长上,进而抗逆性弱于雄性[22],在生存竞争或环境胁迫(如郁闭度大、光照不足等)中雌性更易被淘汰,导致老龄种群性别比例失衡。这与Clark等[23]对Zamia skinneri的研究一致,其雌株因繁殖代价更高而开花延迟,导致种群中雄株比例随龄级增长逐渐升高。本研究对象均为古树(≥100年),其性别比例是长期历史选择和环境胁迫累积的结果,雌株平均茎高(371.36 cm)显著低于雄株(452.92 cm),可能反映其资源分配策略向繁殖倾斜,牺牲部分营养生长以保障种子发育。此外,古树群个体已进入特定生长阶段(树龄≥100年),性别比例偏倚成为历史遗留问题。而后代小苗性别比例平衡,表明在人工养护的迁地环境中,篦齿苏铁有性繁殖过程得以正常开展,性别表达未受显著干扰,这为种群复壮提供了积极信号。后续可通过追踪后代个体生长,探究性别比例随龄级变化的规律,结合解剖学特征分析雌雄株抗逆性差异,进一步明晰篦齿苏铁性别偏倚的发生阶段与驱动机制。
除了性别比例的偏倚,雌雄株在茎干形态与分枝特征上的显著差异,同样反映了其生态适应策略的分化。雄株平均茎高(452.92 cm)显著高于雌株(371.36 cm),这一特征与其风媒传粉策略契合。杨泉光等[24]研究说明风和象鼻虫都是越南篦齿苏铁的有效传粉媒介,且风传粉距离通常≤3 m。雄株更高的茎干可突破冠层限制,扩大花粉扩散范围,提升繁殖成功率。此外,雄株逐步分枝率(SBR2:1=2.29,SBR3:2=0.86)显著高于雌株(SBR2:1=1.91,SBR3:2=0.32),雄株多级分枝(57.23%,分枝≥2次)也明显高于雌株(25.00%,分枝≥2次),雄株分枝系统呈现“多级分枝+强承载力”的结构特征,这一模式具有重要生态意义,暗示着雄株通过比雌株更高频的分枝,形成更复杂的垂直结构以增加“开花”(小孢子叶球)位点,为花粉的生产提供更多物理空间,促进传粉扩散范围和效率,弥补雌株不足对繁殖的制约[25]。这种“数量补偿”策略在风媒传粉植物中普遍存在[26]。在横向生长策略上,雌雄株表现出不同的“茎干—冠幅”权衡模式。雌株胸径(48.55 cm)与地径(67.82 cm)略小于雄株(49.77 cm、71.02 cm),但雌株平均冠幅(447.27 cm )略大于雄株(432.29 cm),出现雌株比雄株茎干略细但冠幅略大的现象。根据基本形态相关分析结果,雌雄株形态指标的协同模式存在差异:
1、雄株:茎高与冠幅(r=0.166*)、胸径(r=0.383**)、地径(r=0.339**)均呈显著正相关,冠幅与胸径(r=0.433**)、地径(r=0.505**)亦高度相关,表明其形态发育高度协调,冠幅与茎干增粗、增高同步,增强机械支撑能力,适应多级分枝带来的结构复杂性。符合“垂直生长—横向扩展”同步优化的策略。
2、雌株:冠幅与茎高呈显著负相关(r=−0.320*),但与胸径(r=0.660**)、地径(r=0.476**)显著正相关。冠幅体现林木间的竞争能力[27],在一定范围内,树冠体积越大,受光部位越多,光能利用率就越高[28]。一般认为雌雄异株植物中,雌株往往在生殖活动中投入了比雄株更多的资源[29]。考虑到本古树群处于人工养护环境中,人为保持了相对充足的生长空间,所以雌株可能将资源优先分配至冠幅扩展与茎干增粗,限制了茎高增长,以更大的冠幅来提升光能利用率,保障种子发育的高能耗需求(单株种子可达百粒以上)。尽管茎高受限,但胸径与冠幅显著正相关,表明其通过适度增粗茎干增强机械支撑,以承载种子重量。另外雌株吸芽数量(1.52个)显著多于雄株(0.78个),暗示雌株其能通过无性繁殖补偿有性繁殖的局限性。
上述研究结果对篦齿苏铁的迁地保护实践具有重要启示。基于雌雄株茎干特征差异,建议在迁地保护中:1)通过茎高、分枝次数、生长点数量等性状辅助性别鉴定;2)在现有种植密度或新定植时,为雌株预留充足的冠幅扩展空间,并在开花结种期对高大雌株或分枝较多的雌株增设支撑结构,以降低因种子重量或冠幅较大导致的分枝断裂风险;3)强化雌株养护,开花结种期增加养分供给;4)鉴于雌株数量稀少,且其平均单株吸芽数量(1.52 个)显著多于雄株(0.78 个),应重点保护和利用雌株吸芽进行人工繁殖,扩大雌株数量,改善种群性别结构,促进种群自然更新与复壮。
-
南宁植物园篦齿苏铁迁地古树群中,雄株茎高、生长点数量及分枝能力显著优于雌株,雌株则表现出更强的吸芽萌生能力。研究结果为篦齿苏铁性别鉴定、科学养护及迁地保护策略优化提供了参考,后续需进一步扩大样本量,深入解析性别差异的分子机制与环境响应模式。
Analysis of stem differences between male and female plants in ancient plant Cycas pectinata
-
摘要: 以南宁植物园篦齿苏铁(Cycas pectinata)迁地古树群为研究对象,统计雌雄株比例,分析茎高、胸径、冠幅、分枝及生长点等形态特征,通过探究篦齿苏铁古树群雌雄株茎干表型差异,为迁地保护与科学养护提供理论依据。统计发现,该古树群雌雄比为1:3.77,种群极显著偏雄;雄株平均茎高(452.92 cm)显著高于雌株(371.36 cm);雌株胸径、冠幅略大于雄株,但差异不显著;93.18%雌株与98.19%雄株发生分枝,其中,57.23%雄株分枝超2次;雄株平均单株生长点(3.69个)显著多于雌株(2.03个)。结果表明,篦齿苏铁古树雄株茎高、生长点数量及分枝能力均优于雌株,迁地栽培中可通过茎干性状初步判定性别,养护时应重点关注雌株生长,强化雌株保护措施。Abstract: An survey was made of the ex-situ population of ancient plant Cycas pectinata at Nanning Botanical Garden for analysis of the sex ratio and morphological characteristics of this ancient plant, including stem height, diameter at breast height (DBH), crown width, branching, and growth points in an attempt to explore the phenotypic differences in stems between male and female plants, providing a theoretical basis for ex-situ conservation and scientific management. The results revealed a highly significant male-biased sex ratio (1:3.77) in this population. The average stem height of the male plants (452.92 cm) was significantly greater than that of the females (371.36 cm). The female plants had slightly higher DBH and crown width, but the differences were not statistically significant. Branching occurred in 93.18% of the female plants and 98.19% of the males, and 57.23% of the male plants branched more than twice. The male plants had a significantly higher average number of growing points per male plant (3.69) than the female plants (2.03). In conclusion, the male plants of the ancient plant C. pectinata exhibit superiority in stem height, number of growing points, and branching capacity compared to the female plants. In ex situ cultivation, stem traits can be used for preliminary sex determination, and protective measures should be strengthened to prioritize the growth of female plants in the plant management.
-
Key words:
- Cycas pectinata /
- dioecious plant /
- stem traits /
- ex-situ conservation
-
图 3 篦齿苏铁古树雌雄株茎干分枝次数差异
Fig. 3 Differences in stem branching times between male and female of ancient plant C. pectinata
表 1 南宁植物园篦齿苏铁古树群雌雄株基本形态概况
Table 1 Basic information of ancient plant C. pectinata in Nanning Botanical Garden
指标Index /性别Sex ♂(n=166) ♀(n=44) 平均 Average 最大 Max. 最小 Min. 平均 Average 最大 Max. 最小 Min. 茎高 Stem height/cm 452.92±149.14* 1030.00 220.00 371.36±65.57* 510.00 250.00 平均冠幅 Average Crown Diameter/cm 432.29±75.25 650.00 200.00 447.27±67.85 575.00 320.00 地径 Ground Diameter/cm 71.02±16.81 127.00 41.00 67.82±14.34 105.00 46.00 胸径 Diameter at Breast Height/cm 49.77±15.13 95.00 28.00 48.55±14.17 84.10 23.50 注:*表示相关性显著在0.05级别;**表示相关性显著在0.01级别;下同。 Note: * indicates significant correlation at the 0.05 level; ** indicates significant correlation at the 0.01 level; Similarly hereinafter.  下载: 导出CSV
下载: 导出CSV
表 2 篦齿苏铁古树群雌雄株基本形态指标的两两相关性
Table 2 Correlation between basic morphological indicators of ancient plant C. pectinata
形态指标
Morphological indices茎高
Stem height平均冠幅
Average crown diameter胸径
Diameter at breasth height地径
Stem diameter at
the ground level♂ ♀ ♂ ♀ ♂ ♀ ♂ ♀ 茎高Stem height * * ** ** 平均冠幅
Average Crown Diameter0.166* −0.320* * ** ** ** 胸径
Diameter at Breast Height0.383** − 0.0188 0.433** 0.660** ** ** 地径Ground Diameter 0.339** −0.100 0.505** 0.476** 0.743** 0.741**
下载: 导出CSV
表 3 分枝式样特征统计
Table 3 The branching characteristics of ancient plant C. pectinata in male and female plants
测定指标 Testing indices ♂(n=166) ♀(n=44) Z P 1级平均分枝长度 Average length of 1st-order branches/cm 242.20±131.74 194.34±110.37 −2.247 0.025* 2级平均分枝长度 Average length of 2nd-order branches/cm 176.28±139.89 168.49±93.14 −0.391 0.696 3级平均分枝长度 Average length of 3rd-order branches/cm 61.31±86.73 17.72±38.22 −3.294 0.001** 总分枝率 Overall bifurcation ratio 0.57±0.21 0.51±0.21 −1.156 0.248 逐步分枝率2:1 Stepwise bifurcation ratio 2:1 2.29±0.95 1.91±0.72 −1.991 0.046* 逐步分枝率3:2 Stepwise bifurcation ratio 3:2 0.86±0.98 0.32±0.60 −3.107 0.002** 枝径比2:1 Ratio of branch diameter (RBD2:1) 0.66±0.19 0.63±0.22 −0.877 0.381 枝径比3:2 Ratio of branch diameter (RBD3:2) 0.33±0.31 0.15±0.28 −2.872 0.004** 2级数量/个 Number of 2nd-order branches/unit 2.29±0.95 1.91±0.72 −1.991 0.046* 3级数量/个 Number of 3rd-order branches/unit 1.89±2.13 0.55±1.03 −3.512 0.000** 吸芽数量/个 Number of suckers/unit 0.78±1.48 1.52±2.43 −2.004 0.045* 末端生长点/个 Number of terminal growing points/unit 2.92±1.58 2.06±0.86 −3.140 0.002* 总生长点数量/个 Total number of growing points/unit 3.69±0.23 2.03±0.13 −4.576 0.000**
下载: 导出CSV
-
[1] 曾辉, 陈勇, 杜凡. 澜沧江中下游篦齿苏铁种群结构及其保护对策[J]. 安徽农业科学, 2011, 39(25), 15416−15419. https://doi.org/10.3969/j.issn.0517-6611.2011.25.088 doi: 10.3969/j.issn.0517-6611.2011.25.088 [2] Dawson T E, Ehleringer J R. Gender-specific physiology, carbon isotope discrimination, and habitat distribution in boxelder, Acer negundo [J]. Ecology, 1993, 74(3), 798−815. https://doi.org/10.2307/1940807 doi: 10.2307/1940807 [3] 尹春英, 李春阳. 雌雄异株植物与性别比例有关的性别差异研究现状与展望[J]. 应用与环境生物学报, 2007, 13(3), 419−425. https://doi.org/10.3321/j.issn:1006-687X.2007.03.028 doi: 10.3321/j.issn:1006-687X.2007.03.028 [4] Chen L H, Zhang S, Zhao H X, et al. Sex-related adaptive responses to interaction of drought and salinity in Populus yunnanensis [J]. Plant, Cell & Environment, 2010, 33(10), 1767−1778. https://doi.org/10.1111/j.1365-3040.2010.02182.x doi: 10.1111/j.1365-3040.2010.02182.x [5] 胥晓, 杨帆, 尹春英, 等. 雌雄异株植物对环境胁迫响应的性别差异研究进展[J]. 应用生态学报, 2007, 18(11), 2626−2631. https://doi.org/10.13287/j.1001-9332.2007.0434 doi: 10.13287/j.1001-9332.2007.0434 [6] 张琦, 叶尔江·拜克吐尔汉, 王娟. 东北鼠李雌雄异株个体对种间竞争响应的性别差异[J]. 林业科学, 2023, 59(11), 33−41. https://doi.org/10.11707/j.1001-7488.LYKX20210989 doi: 10.11707/j.1001-7488.LYKX20210989 [7] 施梦娇, 李斌, 伊力塔, 等. 美洲黑杨幼苗生长和生理生态指标对干旱-复水响应的性别差异[J]. 植物生态学报, 2023, 47(8), 1159−1170. https://doi.org/10.17521/cjpe.2022.0173 doi: 10.17521/cjpe.2022.0173 [8] 袁琳, 许恬, 李德祥, 等. 苏铁迁居青山家族转危为安[N]. 广西日报, 2023-12-20(10). https://doi.org/10.28292/n.cnki.ngxrb.2023.007462 [9] 文云燕. 云南双柏恐龙河自然保护区滇南苏铁物候监测[J]. 林业调查规划, 2021, 46(5), 25−28. https://doi.org/10.3969/j.issn.1671-3168.2021.05.005 doi: 10.3969/j.issn.1671-3168.2021.05.005 [10] 李娟, 林建勇, 何应会, 等. 广西崇左叉叶苏铁种群结构与分布格局研究[J]. 广东农业科学, 2016, 43(12), 25−29. https://doi.org/10.16768/j.issn.1004-874X.2016.12.005 doi: 10.16768/j.issn.1004-874X.2016.12.005 [11] 何永华, 王乾, 石培礼. 攀枝花苏铁的生物学特性、树干解剖和生长模式[J]. 植物学报, 1995(6), 443−451. [12] 《广西古树名木概览》编辑委员会. 广西古树名木概览[M]. 南宁: 广西科学技术出版社, 2017. [13] West P W, Ratkowsky D A, Smith R G B. Factors controlling individual branch development during early growth of an experimental plantation of Eucalyptus pilularis in sub-tropical Australia [J]. Trees, 2021, 35 (2), 395−405. https://doi.org/10.1007/s00468-020-02040-4 doi: 10.1007/s00468-020-02040-4 [14] 郑玉, 张化永, 邹恒超, 等. 冀西北山地森林不同生境下白桦分枝结构特征[J]. 科学技术与工程, 2024, 24(3), 970−979. https://doi.org/10.12404/j.issn.1671-1815.2301657 doi: 10.12404/j.issn.1671-1815.2301657 [15] 张丹, 黄文娟, 李志军, 等. 濒危荒漠植物灰叶胡杨分枝格局及其分形特征[J]. 江苏农业科学, 2016, 44(7), 209−212. https://doi.org/10.15889/j.issn.1002-1302.2016.07.058 doi: 10.15889/j.issn.1002-1302.2016.07.058 [16] 赵亚洲, 辛雅芬, 马钦彦, 等. 雌雄异株树种黄连木种群性比及空间分布[J]. 生态学杂志, 2010, 29(6), 1087−1093. https://doi.org/10.13292/j.1000-4890.2010.0192 doi: 10.13292/j.1000-4890.2010.0192 [17] 彭丹, 武志强. 植物雌雄异株性别决定研究进展[J]. 生物多样性, 2022, 30(3), 21416. https://doi.org/10.17520/biods.2021416 doi: 10.17520/biods.2021416 [18] Bertiller M B, Sain C L, Carrera A L. Effect of fine-scale spatial variation of soil nitrogen on the performance of the sexes of Poa ligularis in patchy ecosystems of northern Patagonia [J]. International Journal of Plant Sciences, 2002, 163(3), 419−425. https://doi.org/10.1086/339515 doi: 10.1086/339515 [19] THOMAS S C, LAFRANKIE J V. Sex, size and interyear variation in flowering among dioecious trees of the Malayan rain forest [J]. Ecology, 1993, 74(5): 1529−1537. https://doi.org/10.2307/1940080 doi: 10.2307/1940080 [20] 黄应锋, 廖绍波, 陈勇, 等. 深圳梅林仙湖苏铁的种群特征与保护研究[J]. 林业科学研究, 2013, 26(5), 668−672. https://doi.org/10.13275/j.cnki.lykxyj.2013.05.020 doi: 10.13275/j.cnki.lykxyj.2013.05.020 [21] 深圳市仙湖植物园(深圳市园林研究中心), 深圳华大生命科学研究院. 分子标记、引物对及其应用和苏铁科植物性别的鉴定方法: 中国, 202210023091.5[P]. 2022−05−31. https://d.wanfangdata.com.cn/patent/ZL_CN202210023091.5_CN114561457A_20220531 [22] 张胜, 刘志斌, 胥晓, 等. 专栏评述: 雌雄异株植物的生理生态学[J]. 四川大学学报(自然科学版), 2025, 62(2), 257−258. https://doi.org/10.19907/j.0490-6756.250063 doi: 10.19907/j.0490-6756.250063 [23] Clark D A, Clark D B. Temporal and environmental patterns of reproduction in Zamia skinneri, a tropical rain forest Cycad [J]. Journal of Ecology, 1987, 75(1), 135−149. https://doi.org/10.2307/2260540 doi: 10.2307/2260540 [24] 杨泉光, 李楠, 李志刚, 等. 越南篦齿苏铁传粉媒介的研究[J]. 热带亚热带植物学报, 2010, 18(2), 129−132. https://doi.org/10.3969/j.issn.1005-3395.2010.02.003 doi: 10.3969/j.issn.1005-3395.2010.02.003 [25] Marler T E, Ferreras U F. Current status, threats and conservation needs of the endemic Cycas wadei merrill [J]. Journal of Biodiversity & Endangered Species, 2017, 5 (3), 1−8. https://doi.org/10.4172/2332-2543.1000193 doi: 10.4172/2332-2543.1000193 [26] 杜凤国, 王柏青. 森林植物学[M]. 长春: 吉林大学出版社, 2008. [27] 康义龙, 贾炜玮, 王帆, 等. 基于BLS数据的人工红松冠幅预测模型构建[J]. 农业工程学报, 2025, 41(3), 179−186. https://doi.org/10.11975/j.issn.1002-6819.202407002 doi: 10.11975/j.issn.1002-6819.202407002 [28] 朱更瑞, 方伟超, 王力荣, 等. 果树保护地栽培问答丛书-桃[M]. 太原: 山西科学技术出版社, 2003. [29] 黄云浩, 辛本花, 王娟. 雌雄异株植物鼠李生殖分配与生殖耗费补偿机制[J]. 北京林业大学学报, 2019, 41(11), 31−36. https://doi.org/10.13332/j.1000-1522.20180425 doi: 10.13332/j.1000-1522.20180425 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 480
- HTML全文浏览量: 291
- PDF下载量: 47
- 被引次数: 0
