










-
水资源是地球−生物−化学−生态系统的核心载体,水资源空间差异是形成生态系统植被区域特征、影响总初级生产力分异的关键因素。水分利用效率(WUE)指的是植物光合作用中消耗单位质量水分所固定的光合产物的总量,通常以总初级生产力(GPP)与对应时期蒸散(ET)水分损失的比率[1-2]来表示,在生态系统水平上,WUE能够反映植物的水分利用策略[3-4],以及植物在光合碳同化过程中获取的碳与消耗的水之间的权衡,即所谓的碳水耦合特征[5-7]。WUE的高低主要由植被的生产力和蒸散量决定,即表示碳封存和水资源消耗之间的关系,是生态系统碳和水循环耦合的重要生态指标[8-9]。生态系统WUE的时间动态主要受自然地理、植物功能类型、干扰历史、气候和环境因素以及植物控制气孔的生理能力等因素的影响[10-11]。环境因素、生物因素及二者的交互作用以不同方式影响GPP和ET,进而导致生态系统WUE发生变化[12]。鉴于持续的气候变化,对生态系统WUE的深入了解将提高预测碳循环和水循环的能力,为区域水碳管理和生态环境的可持续发展提供参考[13-14]。
作为热带地区的“水泵”,橡胶林的蒸散量高于附近天然林的蒸散量,甚至会导致种植区雨季的土壤储水量不足以维持高蒸散量而引起橡胶园旱季缺水,有研究认为,种植区域在旱季面临的水资源短缺问题主要是橡胶树的“水泵”功能引起的[15-18]。因此,橡胶种植园的碳、水循环之间的耦合过程越来越受到人们的关注。为增加对橡胶园WUE的年度格局变化、年际趋势以及WUE调节因素的认识,Lin等[19]对云南省橡胶园进行研究后发现,年际尺度上WUE与光合有效辐射呈负相关,并且WUE 更多地受温度、净辐射和胸径增长率的控制,其中胸径增长率更能解释WUE的变化状况,说明橡胶园碳、水循环耦合过程的主要影响因素是碳封存而非蒸散耗水量;在Tan等[20]的研究中,温度是橡胶园WUE季节性变化的主要控制因素,太阳辐射次之,叶面积指数随温度升高而增加,也对WUE具有重要影响,饱和水汽压差(VPD)在短时间内对WUE的影响较为明显。目前,我国关于橡胶林WUE的研究较少[19-20],且研究多聚焦于WUE与某些影响因素的变化规律及相关性、多基于定性的方式分析WUE的驱动机制。例如,Li等[21]分析不同尺度下WUE与环境因素的关系后发现,在生态系统层面,叶面积指数是WUE变化的主要驱动因素,温度通过影响叶片蒸腾和光合作用,对WUE有很强的控制作用,即温度的升高导致蒸腾作用增强,叶片固有WUE下降,生态系统WUE增加;除此之外,土壤水分也是WUE的重要影响因素,Nie等[17]的研究表明,生态系统WUE与土壤含水量之间存在显著负相关的关系,且VPD越高负相关越明显。主要是由于在植被生长高峰期,随着土壤水分的增加,ET增长速度快于GPP,从而导致WUE随土壤含水量的增加而降低。上述研究结果表明,植被冠层活动和VPD在调控WUE中具有重要作用,如Zhao等[12]的研究结果表明, VPD和冠层导度是调控生态系统WUE最重要的因素,并在对北半球不同植被类型进行研究后发现,WUE对气温、降水和VPD具有负响应,对冠层导度和生态系统呼吸则呈正响应 。然而,各因素对WUE季节性变化的具体贡献尚不清楚,因此笔者拟在参考TRM的分析框架和基于EC-LUE模型、Penman-Monteith模型的基础上,量化关键的大气和地表生物物理因素在WUE季节性变化中的作用,以期为热带地区橡胶林生态系统保护、水资源合理利用提供理论依据。
-
本研究数据来源于2010年1月至2019年12月的涡度协方差测量数据,数据采集点位于海南省的农业农村部儋州热带作物科学观测实验站(19°32′16″N,109°28′06″E),平均海拔114 m。研究区域位于海南省西北部,属于典型热带海岛季风性气候,具有明显的雨旱季[22]。根据前人研究,笔者将该地区每年5—9月定义为雨季,10月至翌年4月为旱季[23];2010—2019年平均温度21.5~28.5 ℃;年平均降水量1476.3 mm,且主要集中在7—9月份,占全年降水的70%以上。研究区域橡胶林植物群落组成简单,主要为橡胶林乔木层和林下草本层,冠层平均高度为20 m[24]。
于2009年11月起对该通量站点进行近地面通量观测,从地下1 m至观测塔顶端50 m,测量数据包含地上和地下两部分,具体测量仪器及安装见表1。
表 1 海南省儋州市的农业农村部儋州热带作物科学观测实验站涡度通量系统数据及测量仪器安装
数据类型 仪器 安装高度 风速 3杯风速计(Met 010C-1, Met One, USA) 1.5、6、10、15、33、41、50 m 温度、湿度 温湿计(HMP45C; Vaisala, Finland) 1.5、6、10、15、33、41、50 m 风向 风向仪(Met 020C-1, Met One, USA 50 m 降雨量 雨量筒(TE525MM, texas instruments, USA)
50 m光合有效辐射 PAR 传感器(LI-190SB, Li-cor , USA) 2、4、8、12、16、30 m 入射长、短波辐射、
出射长、短波辐射辐射观测传感器 (CNR-1, Kipp & Zonen, Holland)
25 m土壤湿度 土壤湿度传感器(CS616-L, Campbell Scientific Inc., USA)
5、20、50 cm土壤温度 土壤温度的观测仪(109, Campbell Scientific Inc., USA)
2、5、20、50、100 cm土壤热通量 土壤热通量板(HFP01, Hukseflux, Holland) 5、20、50 cm -
生态系统WUE在一定程度上反映了生态系统碳循环和水循环的耦合关系,并将光合作用和蒸散紧密联系在一起[25],通常采用GPP与ETET的比值进行计算[10,12,25-26]:
$$ WUE = \frac{{GPP}}{{ET}} \;,$$ 式中,WUE为水分利用效率,单位为 g·kg−1,GPP为生态系统总初级生产力,单位为g·m−2,ET为生态系统蒸散(橡胶林生态系统整体蒸散,包括冠层蒸腾量、裸地蒸发量和冠层截留水量),单位为kg·m−2。
GPP、ET及其中变量计算参考相关文献[26-34],将WUE公式计算解析为由不同影响因素组成的函数:
$$ WUE = f(Ta,\Pr ess,VPD,Rn,G,ra,rs,fPAR,{{S} _{in}}) \; 。$$ -
在本研究中,主要探讨2个问题:(1)WUE雨季至旱季的变化状况;(2)各大气因素和生物物理因素在WUE变化中起到的作用及具体贡献。在进行分析时,将WUE雨季至旱季的变化视为由多个自变量变化所导致的结果,泰勒公式可以将复杂函数使用多项式来近似表达,因此利用多元函数的泰勒公式拟合出近似值(接近实测ET计算所得WUE变化值)。结合拉格朗日中值定理可知函数增加量、自变量增加量及导数之间的关系为:
$ f(x + \Delta x) - f(x) = {f^{'}}(x + \theta \Delta x)\Delta x $ ,在此基础上,参考TRM方法的分析框架[5,35],量化关键大气和地表生物物理因素在WUE变化中的作用,对其公式进行一阶求导可得:$$ \begin{split} \Delta {{WUE}} = & \frac{{\partial {{WUE}} }}{{\partial Ta}}\Delta Ta + \frac{{\partial {{WUE}} }}{{\partial \Pr ess}}\Delta \Pr ess + \\& \frac{{\partial {{WUE}} }}{{\partial VPD}}\Delta VPD + \frac{{\partial {{WUE}} }}{{\partial {R_n}}}\Delta {R_n} + \frac{{\partial {{WUE}} }}{{\partial G}}\Delta G + \\&\frac{{\partial {{WUE}} }}{{\partial ra}}\Delta ra + \frac{{\partial {{WUE}} }}{{\partial rs}}\Delta rs + \frac{{\partial {{WUE}} }}{{\partial fPAR}}\Delta fPAR +\\& \frac{{\partial {{WUE}} }}{{\partial {{S} _{in}}}}\Delta {{S} _{in}} \;, \\[-5pt] \end{split}$$ 式中,Δ表示雨旱季差值(例如,
$ \Delta G={G}_{旱季}-{G}_{雨季} $ ,表示G从雨季至旱季的变化量),偏微分项表示WUE对每个变量变化的敏感性(例如,$ \dfrac{{\partial WUE}}{{\partial G}} $ 表示WUE对G变化的敏感性)。归因方法是基于一阶泰勒公式展开的,并未将高阶项和交叉项考虑在内,通常只有当各归因变量的变化很小时,这种方法引起的误差才可以忽略不计,但本研究中部分影响因素的季节变化较大(表2)。为减少误差,笔者参考加权平均方法来优化结果[36]:
表 2 雨季至旱季各影响因素的变化量
因素 Ta/℃ Press/kPa VPD/kPa Rn/(W·m−2) Sin/(W·m−2) fPAR G/(W·m−2) rs/(s·m−1) ra/(s·m−1) Δ −6.23 0.83 −0.60 −100.93 −112.89 −0.03 −2.56 110.88 −7.70 $$ \begin{split} \\ X=\frac{\left({x}_{雨季}+m{x}_{旱季}\right)}{1+m} \;, \end{split}$$ 式中,X为模型中使用的最优偏微分项,m为平均权重因子(取−200至200,以0.01为步长,本研究m取值为0.04),
$ {x}_{雨季} $ 和$ {x}_{旱季} $ 分别为仅使用雨季和仅使用旱季数据计算所得偏微分项。 -
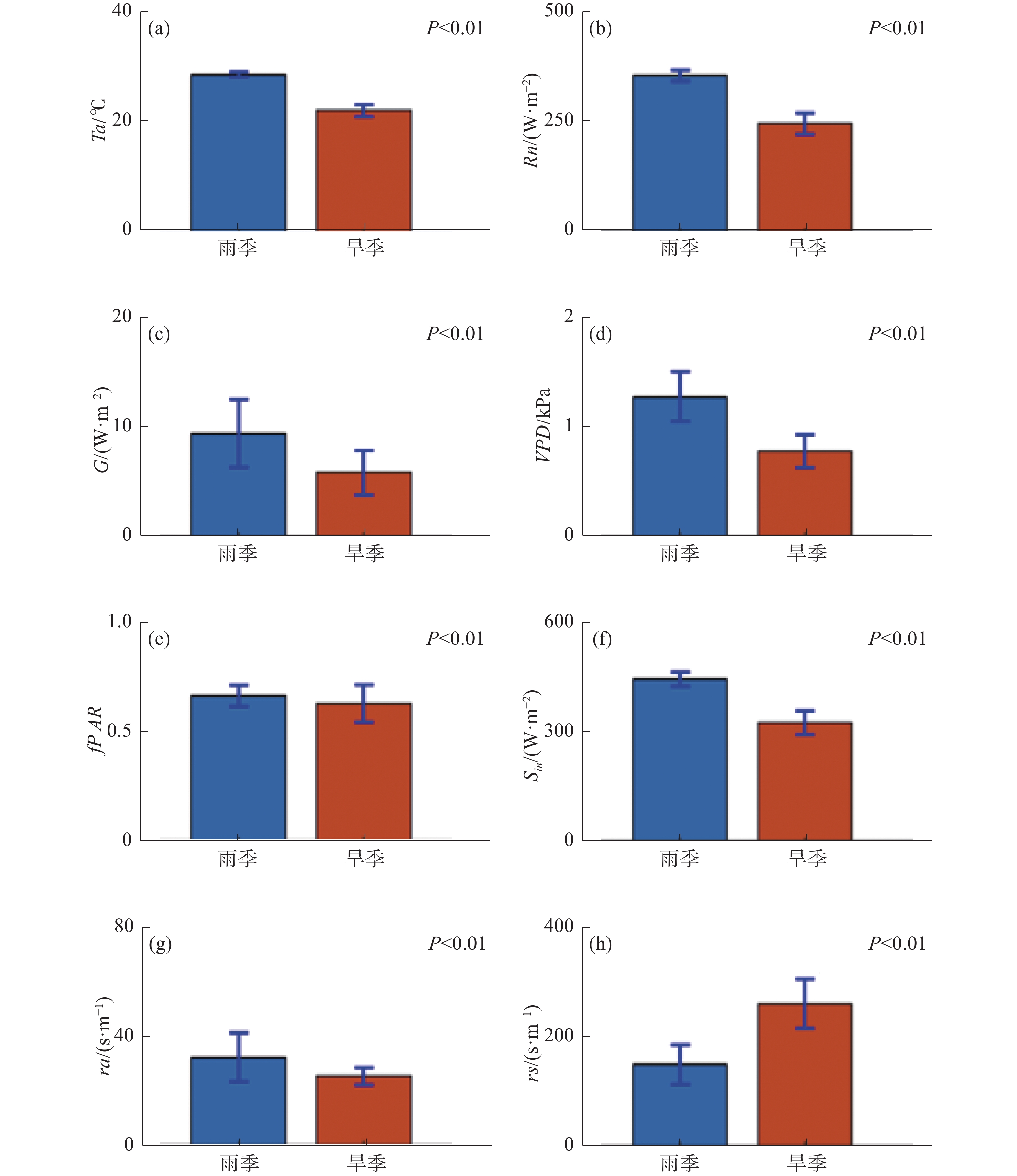
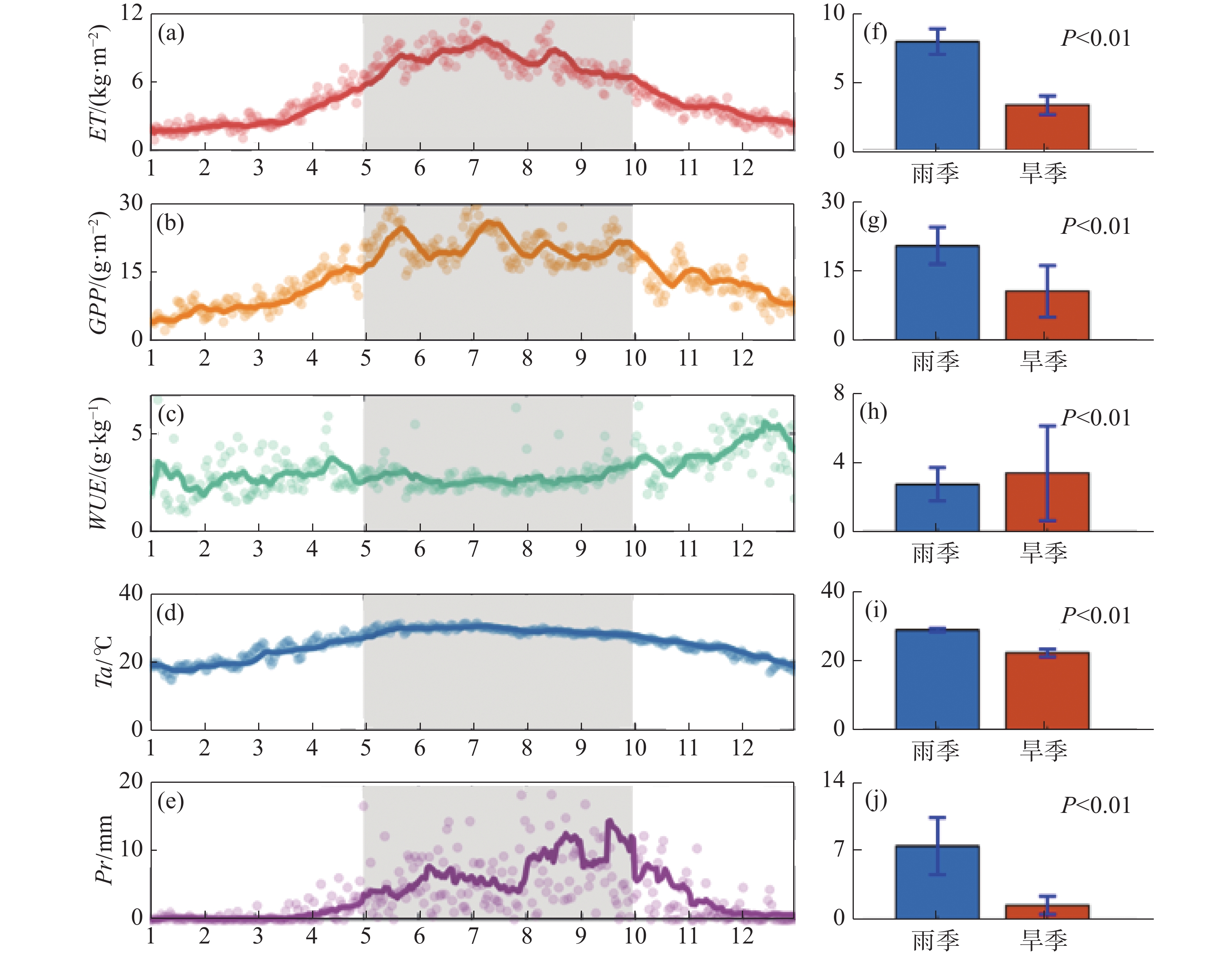
从图1可知,同温度(峰值位于雨季,且雨季平均温度高于旱季6.67 ℃)和降水(多年雨季平均日降水高达7.57 mm,远高于旱季平均日降水1.50 mm)的年内变化趋势类似,ET和GPP具有明显的季节性差异(P<0.01),整体呈“倒U”型曲线,峰值通常位于雨季中期,雨季ET日均为7.93 kg·m−2,旱季日均3.31 kg·m−2,相差4.62 kg·m−2(图1-a,图1-f);GPP雨季均值为20.89 g·m−2,旱季均值为10.83 g·m−2,相差10.06 g·m−2(图1- b,图1-g);整体而言,WUE旱季的波动幅度较大,平均值约为4.73 g·kg−1,雨季均值约为3.52 g·kg−1,雨旱季差异显著(P<0.01),旱季WUE平均高于雨季1.21 g·kg−1(图1-c,图1-h)。
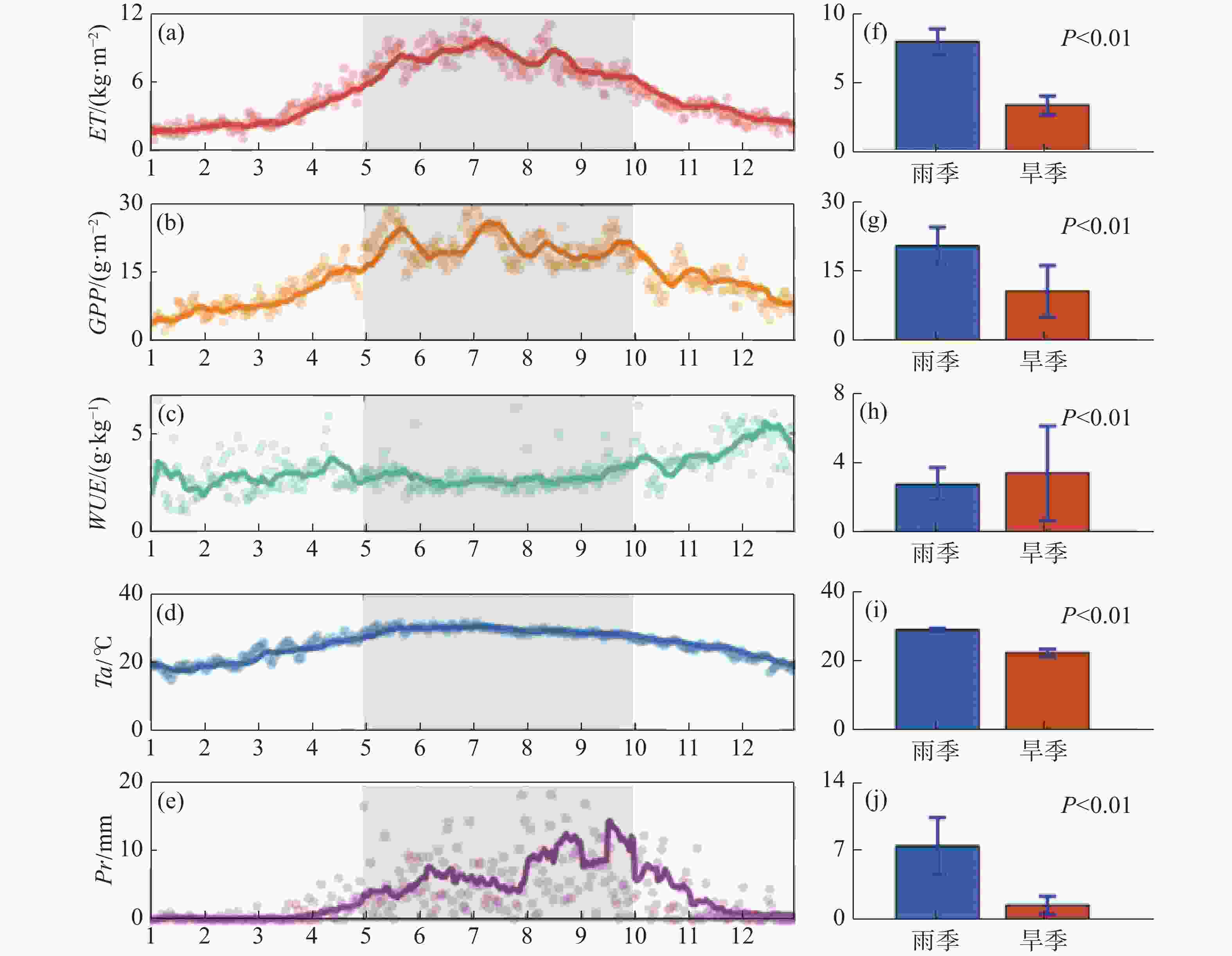
图 1 橡胶林蒸散(ET)、生态系统总初级生产力(GPP)、水分利用效率(WUE)、温度(Ta)、降水(Pr)年内变化及雨旱季对比
-
由各环境因素雨旱季的对比及显著性差异分析结果(图2)可知,各因素均具有显著性雨旱季差异(P<0.01),其中,温度、净辐射、土壤热通量、VPD、冠层光合有效辐射吸收比例、空气动力学阻力、太阳辐射的雨季平均值高于旱季,地表阻抗的雨季平均值低于旱季;2010—2019 年雨季温度平均值为29.11 ℃,旱季为22.44 ℃,相差6.67 ℃;净辐射雨季均值为359.13 W·m−2,旱季为249.45 W·m−2,相差109.68 W·m−2;雨季平均土壤热通量为9.48 W·m−2,旱季平均土壤热通量为5.86 W·m−2,相差3.62 W·m−2;VPD的雨季平均值1.28 kPa,旱季平均值0.78 kPa, 差值为0.50 kPa;雨季植被冠层光合有效辐射吸收比例均值为0.67,旱季为0.63,雨季高于旱季0.04;太阳辐射雨季均值为445.61 W·m−2,旱季为326.01 W·m−2,相差119.6 W·m−2;空气动力学阻力雨季均值32.14 s·m−1,旱季均值25.19 s·m−1,雨季高于旱季7.14 s·m−1;地表阻抗雨季均值为149.03 s·m−1,旱季均值260.46 s·m−1,雨季低于旱季111.43 s·m−1。
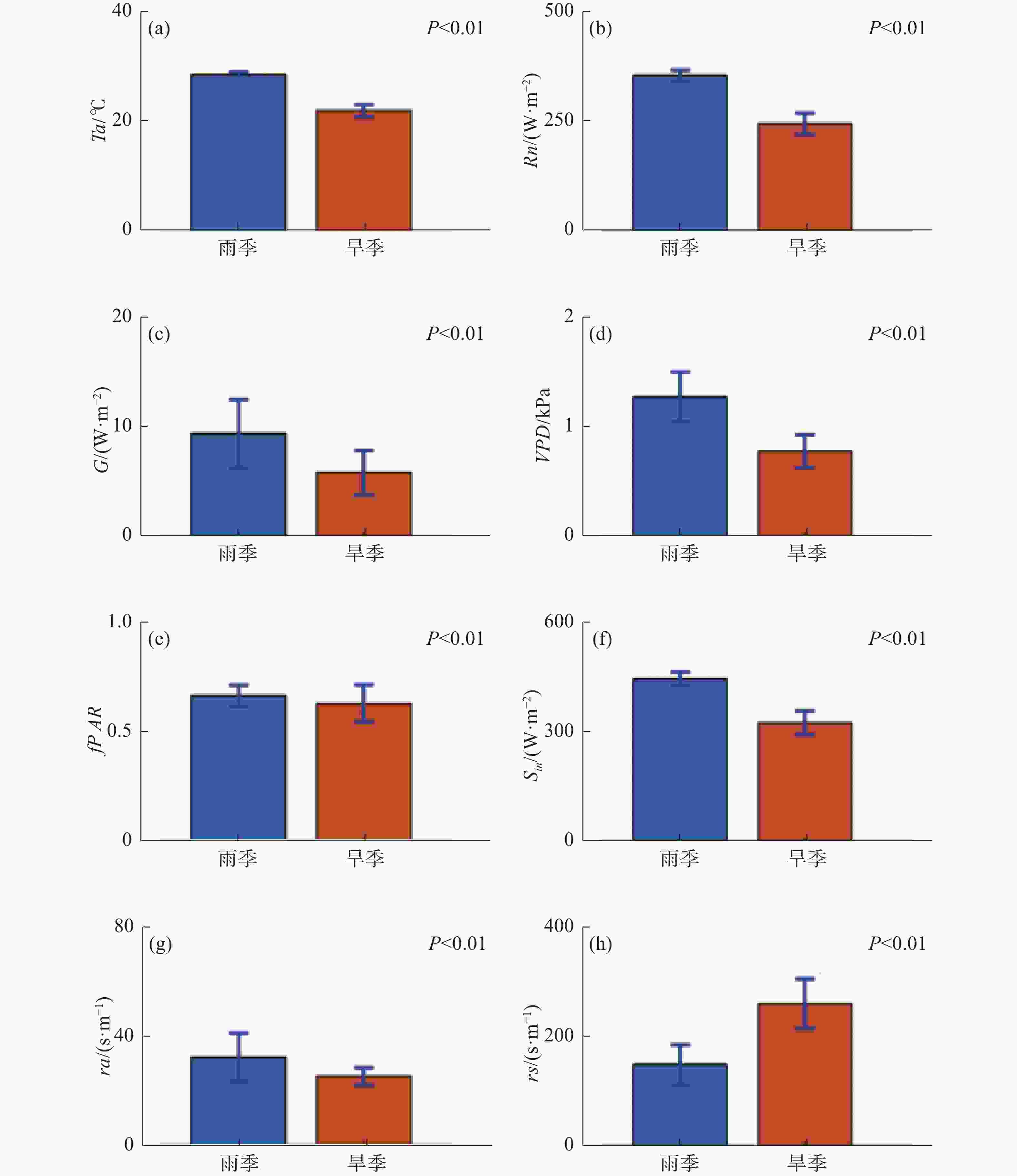
图 2 温度(Ta)、净辐射(Rn)、土壤热通量(G)、饱和水汽压差(VPD)、冠层光合有效辐射吸收比例(fPAR)、太阳辐射(Sin)、空气动力学阻力(ra)、地表阻抗(rs)的雨旱季对比
-
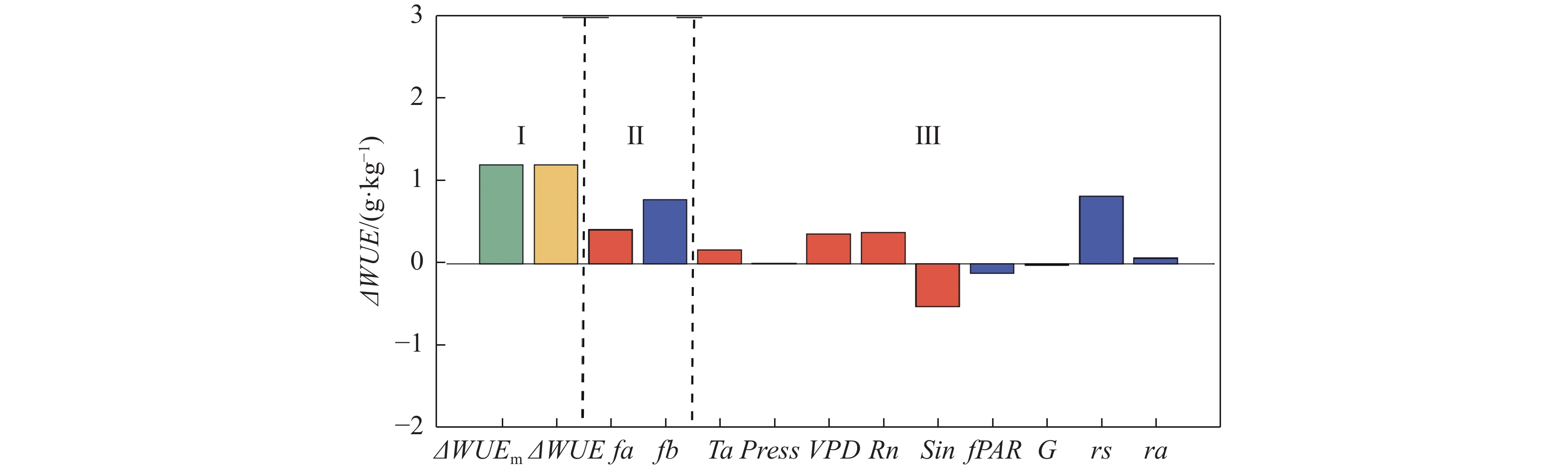
各影响因素对WUE变化的贡献可见图3。将从雨季至旱季的WUE变化分解为5个大气因素(温度、大气压、饱和水汽压差、净辐射、太阳辐射)和4个生物物理因素(植被冠层光合有效辐射吸收比例、土壤热通量、空气动力学阻力、地表阻抗)的贡献,如图3所示,第Ⅰ部分显示模型估算GPP和模型估算ET计算所得ΔWUE与模型估算GPP和实测ET计算所得ΔWUE是相似的,WUE在旱季增加。第Ⅱ部分可知,大气因素总贡献和生物物理因素总贡献皆为正向,这说明,进入旱季后,大气因素和生物物理因素都倾向于增加WUE,并且生物物理因素的贡献高于大气因素的贡献。
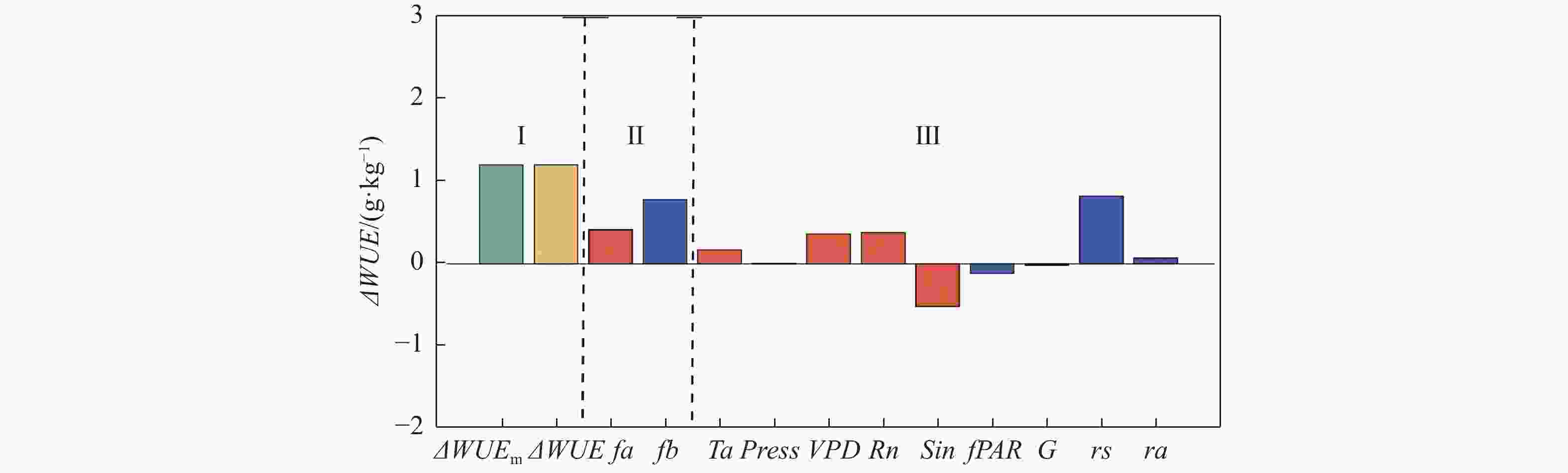
图 3 各影响因素对WUE变化量的贡献量化图
在大气因素中(图3中第Ⅲ部分,橙色条),温度、VPD、净辐射产生正增益(即三者的变化倾向于增加WUE),而太阳辐射的变化倾向于减少旱季WUE(表3),并且被温度、VPD和净辐射的正增益所抵消,大气压的影响可忽略不计。
表 3 WUE对大气和生物物理因素变化的敏感性
$\dfrac{{\partial WUE}}{{\partial Ta}}$/
℃$\dfrac{{\partial WUE}}{{\partial \Pr ess}}$/
kPa$\dfrac{{\partial WUE}}{{\partial VPD}}$/
kPa$\dfrac{{\partial WUE}}{{\partial Rn}}$/
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial fPAR}}$ $\dfrac{{\partial WUE}}{{\partial {{S} _{in}}}}$/
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial G}}$/
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial rs}}$/
(m·s−1)$\dfrac{{\partial WUE}}{{\partial ra}}$/
(m·s−1)−2.76×10−2 1.40×10−2 −0.62 −3.84×10−3 3.39 4.60×10−3 3.83×10−3 7.44×10−3 −9.80×10−3 在生物物理因素中(图3中的第Ⅲ部分,蓝色条),植被冠层光合有效辐射吸收比例的雨旱季差别不大(图2-e, 表2),但WUE对其变化的敏感性较大,故产生负增益,倾向于减少旱季WUE(表3);空气动力学阻力旱季的减少导致WUE(表3)的增加;地表阻抗的变化倾向于增加旱季WUE(表3),并抵消了植被冠层光合有效辐射吸收比例的负增益。土壤热通量对WUE的贡献则小到可以忽略不计。
第I部分中ΔWUEm代表GPP与模型ET计算所得WUE雨旱季差值;ΔWUE代表GPP与实测ET计算所得WUE雨旱季差值)。第II部分中,fa为大气因素的总贡献量,fs为生物物理因素的总贡献量;第III部分中,橙色代表各大气因素的贡献,蓝色代表各生物物理因素的贡献。
-
本研究结果显示,橡胶林WUE的年内变化与水分充足的中国东部森林保持一致,即在GPP和ET较高的季节,WUE较低[37]。在本研究中,橡胶林WUE的年内变化幅度较小,多年旱季WUE平均高于雨季1.21 g·kg−1,与同处于热带地区的鼎湖山针阔叶混交林雨旱季WUE差值相近[38],平均日均值为4.16 g·kg−1,略高于云南地区热带森林WUE 3.16 g·kg−1,旱季水分利用效率的增加则是ET比GPP更强烈的下降引起的[20], 结合本研究结果,从雨季过渡至旱季,ET降低幅度58.26%,GPP减少48.16%。橡胶树在雨季从上层土壤中提取水分,在旱季从深层土壤中提取水分。依次来响应土壤水分的变化[25]。对深层土壤水分的利用使其在旱季仍能提供一定水分以供光合作用固碳活动的正常进行,因此,旱季橡胶林生态系统的GPP下降趋势较缓。
另一方面,根据降水的年内分布可知,研究区域的降水集中分布在7—9月份,且雨热同期,年降水量达1 476.3 mm,雨季降水的增加、气温的上升导致ET明显升高,但GPP增加有限。在同处于热带的亚马逊雨林生态系统的研究中,已有结果表明ET 是森林WUE变化的主导因子[18],即相同增幅的降水量对GPP的促进作用较小,但大幅度增加蒸散用水量,使得该地区雨季的WUE低于旱季[39]。在同处于热带地区的研究中,鼎湖山针阔叶混交林WUE变化出现相似情况,即降雨量集中的湿季WUE低于干季[38]。
-
归因分析结果表明,WUE的变化是大气因素和生物物理因素共同作用的结果。随着旱季的到来,温度的降低对WUE的增加产生正增益,符合已有结论,在对西双版纳橡胶林的研究中,蒸散对温度降低更为敏感,且温度 和 WUE之间存在负相关的关系[20,40-41];旱季VPD的减少引起 WUE增加,主要原因是VPD和ET的正相关性,这一点不仅在Zhao等[12]对北半球不同森林类型的研究中得以体现,而且在亚马逊热带雨林的研究中也有类似结论[29];净辐射量的降低更倾向于减少蒸散,因而旱季净辐射的减少导致WUE的增加。旱季入射短波辐射的降低会导致植物光合有效辐射减少,光合作用活动减弱,从而引起WUE的减少(产生负增益)。
旱季植被冠层光合有效辐射吸收比例的减少意味着植被冠层固定能量减少,植被GPP降低,导致WUE的减少;但是,植被冠层光合有效辐射吸收比例带来的负增益完全被地表阻抗抵消。从归因分析中的生物物理要素来看,地表阻抗的贡献量最高。以往的研究结果表明,冠层导度和生态系统呼吸是影响森林生态系统WUE的2个关键生物因素[12]。气孔是冠层与大气进行气体交换的主要通道,并且可以有效控制水分流失与植物碳的固定及有机物生产之间的平衡[42-43]。在同样位于热带地区研究中,亚马逊热带雨林旱季气孔关闭通常会导致比光合作用更强烈的蒸腾作用下降[44]。本研究结果也与之相符,进入旱季后,气孔导度下降,地表阻抗增加,从而导致WUE增加。结合现有研究认为,气孔是控制植物蒸腾作用的最直接的因素[48],其开闭活动对植物蒸腾作用的直接影响远大于其对光合作用生产有机物的影响,橡胶林主要通过气孔调节来应对进入旱季后供水量的减少。
Seasonal differences and attribution analysis of water use efficiency in rubber plantations at the north fringe of the tropics
-
摘要: 为了深入了解橡胶树(Hevea brasiliensis)的生态水文和碳水循环耦合过程,依托涡度通量数据,利用EC-LUE模型和Penman-Monteith模型,在模拟橡胶林生态系统总初级生产力(GPP)和蒸散(ET)数据的基础上,分析了2010—2019年橡胶林生态系统WUE季节性差异。并在此基础上,定量评估了各因素对WUE变化的影响。结果表明:(1)从时序变化来看,2010—2019年间橡胶林生态系统雨季WUE(3.52 g·kg−1)小于旱季(4.73 g·kg−1),GPP和ET均表现出明显的“单峰型”特征,二者峰值出现在雨季;(2)从归因分析来看,大气因素中,太阳辐射倾向于在旱季抑制WUE的增加(呈现负贡献),但是这种抑制作用被大气因素中的其他因素抵消(呈现正贡献)。在各因素中,对WUE变化贡献最高的因素为地表阻抗,其在旱季倾向于促进WUE的增加,贡献占比为33.29%。橡胶林WUE在雨旱季转换的变化主要受生物物理因素调控。Abstract: Water use efficiency (WUE), a critical element of carbon and hydrological cycle, plays a key role in land-atmosphere feedback. The seasonal differences of rubber plantation ecosystems in WUE from 2010 to 2019 were analyzed by using the EC-LUE model and the Penman-Monteith model based on the data of gross primarily productivity (GPP) and evapotranspiration (ET). Furthermore, the impact of each factor on the change of WUE was quantitatively evaluated. The factors were then classified into two groups: atmospheric (temperature, atmospheric pressure, water vapor deficit, net radiation, and solar radiation) and biophysical (fraction of photosynthetically active radiation, soil heat flux, aerodynamic resistance, and canopy resistance). The results showed that the rubber plantation ecosystems had a lower WUE(3.52 g·kg−1) in the rainy season than in the dry season (4.73 g·kg−1) from 2010 to 2019, and that both GPP and ET curves were clearly unimodal, with their peaks all in the rainy season. Solar radiation tended to reduce the increase of WUE in the dry season, indicating a negative contribution, although this suppression was counteracted by other atmospheric factors, showing a positive contribution. Surface impedance, which tended to encourage the growth of WUE in the dry season, had the highest contribution to WUE change, accounting for 33.29% of the total. The WUE change in rubber plantations from rainy to drought seasons were primarily governed by biophysical parameters, which might serve as a reference for a further study of the ecohydrological and carbon-water cycle activities in rubber plantations.
-
图 2 温度(Ta)、净辐射(Rn)、土壤热通量(G)、饱和水汽压差(VPD)、冠层光合有效辐射吸收比例(fPAR)、太阳辐射(Sin)、空气动力学阻力(ra)、地表阻抗(rs)的雨旱季对比
条形分别代表雨旱季多年日均值,误差棒分别代表变量雨旱季的标准偏差。
表 1 海南省儋州市的农业农村部儋州热带作物科学观测实验站涡度通量系统数据及测量仪器安装
数据类型 仪器 安装高度 风速 3杯风速计(Met 010C-1, Met One, USA) 1.5、6、10、15、33、41、50 m 温度、湿度 温湿计(HMP45C; Vaisala, Finland) 1.5、6、10、15、33、41、50 m 风向 风向仪(Met 020C-1, Met One, USA 50 m 降雨量 雨量筒(TE525MM, texas instruments, USA)
50 m光合有效辐射 PAR 传感器(LI-190SB, Li-cor , USA) 2、4、8、12、16、30 m 入射长、短波辐射、
出射长、短波辐射辐射观测传感器 (CNR-1, Kipp & Zonen, Holland)
25 m土壤湿度 土壤湿度传感器(CS616-L, Campbell Scientific Inc., USA)
5、20、50 cm土壤温度 土壤温度的观测仪(109, Campbell Scientific Inc., USA)
2、5、20、50、100 cm土壤热通量 土壤热通量板(HFP01, Hukseflux, Holland) 5、20、50 cm  下载: 导出CSV
下载: 导出CSV
表 2 雨季至旱季各影响因素的变化量
因素 Ta/℃ Press/kPa VPD/kPa Rn/(W·m−2) Sin/(W·m−2) fPAR G/(W·m−2) rs/(s·m−1) ra/(s·m−1) Δ −6.23 0.83 −0.60 −100.93 −112.89 −0.03 −2.56 110.88 −7.70
下载: 导出CSV
表 3 WUE对大气和生物物理因素变化的敏感性
$\dfrac{{\partial WUE}}{{\partial Ta}}$ /
℃$\dfrac{{\partial WUE}}{{\partial \Pr ess}}$ /
kPa$\dfrac{{\partial WUE}}{{\partial VPD}}$ /
kPa$\dfrac{{\partial WUE}}{{\partial Rn}}$ /
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial fPAR}}$ $\dfrac{{\partial WUE}}{{\partial {{S} _{in}}}}$ /
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial G}}$ /
( m2·W−1)$\dfrac{{\partial WUE}}{{\partial rs}}$ /
(m·s−1)$\dfrac{{\partial WUE}}{{\partial ra}}$ /
(m·s−1)−2.76×10−2 1.40×10−2 −0.62 −3.84×10−3 3.39 4.60×10−3 3.83×10−3 7.44×10−3 −9.80×10−3
下载: 导出CSV
-
[1] 董璐. 热带季节性落叶与常绿森林的生态系统—大气交换通量比较[D]. 太原: 山西大学, 2020. [2] 史晓亮, 吴梦月, 张娜. 中国典型陆地生态系统水分利用效率及其对气候的响应[J]. 农业工程学报, 2020, 36(9): 152 − 159. [3] 胡中民, 于贵瑞, 王秋凤等. 生态系统水分利用效率研究进展[J]. 生态学报, 2009, 29(3): 1498 − 1507. doi: 10.3321/j.issn:1000-0933.2009.03.048 [4] 凌祯. 西双版纳橡胶林蒸散量时空变异特征及其预报模型研究[D]. 昆明: 云南师范大学, 2021. [5] FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis [J]. Annual review of plant physiology, 1982, 33(1): 317 − 345. doi: 10.1146/annurev.pp.33.060182.001533 [6] 郑周涛, 张扬建. 1982—2018年青藏高原水分利用效率变化及归因分析[J/OL]. 植物生态学报. (2022-10-25)[2023-03-12]. http://kns.cnki.net/kcms/detail/11.3397.Q.20221024.1750.002.html. [7] ZHOU S, YU B F, HUANG Y F, WANG G Q. The effect of vapor pressure deficit on water use efficiency at the subdaily time scale [J]. Geophysical Research Letters, 2014, 41(14): 5005 − 5013. doi: 10.1002/2014GL060741 [8] 赵玮, 张一平, 宋清海, 等. 橡胶树蒸腾特征及其与环境因子的关系[J]. 生态学杂志, 2014, 33(7): 1803 − 1810. doi: 10.13292/j.1000-4890.20140422.034 [9] ZHU X J, YU G R, WANG Q F, et al. Spatial variability of water use efficiency in China's terrestrial ecosystems [J]. Global and Planetary Change, 2015, 129: 37 − 44. doi: 10.1016/j.gloplacha.2015.03.003 [10] TONG X, ZHANG J, MENG P, et al. Ecosystem water use efficiency in a warm-temperate mixed plantation in the North China [J]. Journal of Hydrology, 2014, 512: 221 − 228. doi: 10.1016/j.jhydrol.2014.02.042 [11] YU G, SONG X, WANG Q, et al. Water‐use efficiency of forest ecosystems in eastern China and its relations to climatic variables [J]. New Phytologist, 2008, 177(4): 927 − 937. doi: 10.1111/j.1469-8137.2007.02316.x [12] ZHAO J, FENG H, XU T, et al. Physiological and environmental control on ecosystem water use efficiency in response to drought across the northern hemisphere [J]. Science of the Total Environment, 2021, 758: 143599. doi: 10.1016/j.scitotenv.2020.143599 [13] MA J, JIA X, ZHA T, et al. Ecosystem water use efficiency in a young plantation in Northern China and its relationship to drought [J]. Agricultural and Forest Meteorology, 2019, 275: 1 − 10. doi: 10.1016/j.agrformet.2019.05.004 [14] PAN S, CHEN G, REN W, et al. Responses of global terrestrial water use efficiency to climate change and rising atmospheric CO2 concentration in the twenty-first century [J]. International Journal of Digital Earth, 2018, 11(6): 558 − 582. doi: 10.1080/17538947.2017.1337818 [15] 祁栋灵, 兰国玉, 陈帮乾, 等. 橡胶林生态系统生态功能述评[J]. 生物学杂志, 2021, 38(1): 102 − 105. doi: 10.3969/j.issn.2095-1736.2021.01.102 [16] BEER C, CIAIS P, REICHSTEIN M, et al. Temporal and among‐site variability of inherent water use efficiency at the ecosystem level[J]. Global Biogeochemical Cycles, 2009, 23(2). D0I:10.1029/2008GB003233. [17] NIE C, HUANG Y, ZHANG S, et al. Effects of soil water content on forest ecosystem water use efficiency through changes in transpiration/evapotranspiration ratio [J]. Agricultural and Forest Meteorology, 2021, 308: 108605. [18] GIAMBELLUCA T W, MUDD R G, LIU W, et al. Evapotranspiration of rubber (Hevea brasiliensis) cultivated at two plantation sites in Southeast Asia [J]. Water Resources Research, 2016, 52(2): 660 − 679. doi: 10.1002/2015WR017755 [19] LIN Y, GRACE J, ZHAO W, et al. Water-use efficiency and its relationship with environmental and biological factors in a rubber plantation [J]. Journal of Hydrology, 2018, 563: 273 − 282. doi: 10.1016/j.jhydrol.2018.05.026 [20] TAN Z H, ZHANG Y P, DENG X B, et al. Interannual and seasonal variability of water use efficiency in a tropical rainforest: Results from a 9 year eddy flux time series [J]. Journal of Geophysical Research:Atmospheres, 2015, 120(2): 464 − 479. doi: 10.1002/2014JD022535 [21] LI H, WEI M, DONG L, et al. Leaf and ecosystem water use efficiencies differ in their global-scale patterns and drivers [J]. Agricultural and Forest Meteorology, 2022, 319: 108919. doi: 10.1016/j.agrformet.2022.108919 [22] 童俊儒, 安锋, 谢贵水. 基于CROPWAT模型的橡胶树需水量[J]. 热带生物学报, 2022, 13(1): 48 − 56. [23] LU H, LU F P, XU X L, et al. The control technique of rubber borers by pheromone traps in Hainan[C] Switzerland: Advanced Materials Research. Trans Tech Publications Ltd, 2014: 699 − 702. [24] 吴志祥. 海南岛橡胶林生态系统碳平衡研究[D]. 海口: 海南大学, 2013. [25] AGUILOS M, STAHL C, BURBAN B, et al. Interannual and seasonal variations in ecosystem transpiration and water use efficiency in a tropical rainforest [J]. Forests, 2018, 10(1): 14. doi: 10.3390/f10010014 [26] 黄卓, 曹银轩, 徐喜娟, 等. 基于多源 GPP 和 ET 产品的秦岭生态系统水分利用效率研究[J]. 水土保持学报, 2022, 36(5): 181 − 194. [27] 王大为, 赵成章, 方锋, 等. 石羊河流域水分利用效率特征及其对饱和水汽压差的响应[J/OL]. 生态学报, 2023(8): 1−13[2023−03−12]. http://kns.cnki.net/kcms/detail/11.2031.Q.20220826.1622.058.html. [28] MONTEITH J L. The state and movement of water in living organisms[C]//19th Symposia of the Society for Experimental Biology. London: Cambridge University Press, 1965: 205 − 234. [29] THOM A S. Momentum, mass and heat exchange of vegetation [J]. Quarterly Journal of the Royal Meteorological Society, 1972, 98(415): 124 − 134. doi: 10.1002/qj.49709841510 [30] KABAT P, DOLMAN A J, ELBERS J A. Evaporation, sensible heat and canopy conductance of fallow savannah and patterned woodland in the Sahel [J]. Journal of Hydrology, 1997, 188: 494 − 515. [31] YUAN W, LIU S, ZHOU G, et al. Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes [J]. Agricultural and Forest Meteorology, 2007, 143(3/4): 189 − 207. [32] RAICH J W, RASTETTER E B, MELILLO J M, et al. Potential net primary productivity in South America: application of a global model [J]. Ecological Applications, 1991, 1(4): 399 − 429. doi: 10.2307/1941899 [33] WANG L, LI M, WANG J, et al. An analytical reductionist framework to separate the effects of climate change and human activities on variation in water use efficiency [J]. Science of the Total Environment, 2020, 727: 138306. doi: 10.1016/j.scitotenv.2020.138306 [34] WANG Y, LI R, HU J, et al. Daily estimation of gross primary production under all sky using a light use efficiency model coupled with satellite passive microwave measurements [J]. Remote Sensing of Environment, 2021, 267: 112721. doi: 10.1016/j.rse.2021.112721 [35] WANG P, LI D, LIAO W, et al. Contrasting evaporative responses of ecosystems to heatwaves traced to the opposing roles of vapor pressure deficit and surface resistance [J]. Water Resources Research, 2019, 55(6): 4550 − 4563. doi: 10.1029/2019WR024771 [36] LIAO W, RIGDEN A J, LI D. Attribution of local temperature response to deforestation [J]. Journal of Geophysical Research: Biogeosciences, 2018, 123(5): 1572 − 1587. doi: 10.1029/2018JG004401 [37] 张良侠, 胡中民, 樊江文, 等. 区域尺度生态系统水分利用效率的时空变异特征研究进展[J]. 地球科学进展, 2014, 29(6): 691 − 699. doi: 10.11867/j.issn.1001-8166.2014.06.0691 [38] 黄健强, 邓永红, 曾小平, 等. 南亚热带针阔叶混交林生态系统水分利用效率[J]. 生态学杂志, 2020, 39(8): 2538 − 2545. doi: 10.13292/j.1000-4890.202008.015 [39] 杜晓铮, 赵祥, 王昊宇, 等. 陆地生态系统水分利用效率对气候变化的响应研究进展[J]. 生态学报, 2018, 38(23): 8296 − 8305. [40] LI S, KANG S, ZHANG L, et al. Ecosystem water use efficiency for a sparse vineyard in arid northwest China [J]. Agricultural Water Management, 2015, 148: 24 − 33. doi: 10.1016/j.agwat.2014.08.011 [41] 邓慧平, 丹利, 王倩, 等. 气候与植被覆盖变化对中国西南亚高山区流域碳水循环的影响模拟[J]. 气候与环境研究, 2020, 25(1): 19 − 31. [42] 宋春林, 孙向阳, 王根绪. 森林生态系统碳水关系及其影响因子研究进展[J]. 应用生态学报, 2015, 26(9): 2891 − 2902. doi: 10.13287/j.1001-9332.20150630.020 [43] WANG M, CHEN Y, WU X, et al. Forest‐type‐dependent water use efficiency trends across the Northern Hemisphere [J]. Geophysical Research Letters, 2018, 45(16): 8283 − 8293. doi: 10.1029/2018GL079093 [44] 周洁. 北京大兴杨树人工林生态系统水分利用效率研究[D]. 北京: 北京林业大学, 2013. -

 点击查看大图
点击查看大图
图(3) / 表 (3)
计量
- 文章访问数: 453
- HTML全文浏览量: 182
- PDF下载量: 15
- 被引次数: 0









