

-
木薯(Manihot esculenta)是大戟科木薯属植物,广泛种植于热带与亚热带地区,于19世纪20年代引进中国栽培[1]。木薯因其适应性强、种植成本低和单产潜力高等优点,与红薯、马铃薯并称为3大薯类作物,是全球第6大粮食作物,8亿人赖以生存的粮食[2]。木薯的重要性不只在于其食用价值,还广泛应用于医药、造纸、纺织、化工等行业,尤其可以作为一种生物质燃料乙醇原料[3]。在石油、煤炭等不可再生能源日益枯竭的当下,新兴的可再生的绿色生物能源有力地保证了能源产业的发展。目前,中国木薯的供应主要依赖于进口,与天然橡胶(Hevea brasiliensis)、咖啡(Coffea arabica)等其他热带农产品相比,进口量居于首位[4]。随着对木薯加工技术与应用领域研究的进一步深入,木薯的需求量也逐步提升,国内木薯产量不足,因此过度依赖于进口,阻碍了木薯产业的发展[5]。中国要重视国内木薯产能,提高木薯单产及种植面积,来满足国内日益增长的需求。
植物的生长和生产离不开环境因素的影响。海水倒灌、过度灌溉、化肥的不合理使用加剧了土壤盐碱化程度,中国约有666.67万hm2盐碱地,严重制约了农业生产的发展[6]。因此,盐胁迫是保证作物产量所要应对的重要非生物胁迫之一。土壤中高浓度的盐离子会引发渗透胁迫,阻碍植物根部对水分的吸收,过量的Na+和Cl-使植物遭受离子毒害,引发氧化胁迫、营养胁迫等次生胁迫[7]。虽然在盐胁迫下木薯可以提高细胞内小分子有机物的含量来减少水分流失,但随时间延长木薯下层叶片发黄脱离,生长受到抑制从而影响木薯产量[8]。相关研究发现,MeSWEET15a/b(sugars will eventually be exported transporter 15a/b)抑制木薯植株叶片蔗糖含量增加,介导糖分分布来调节细胞内渗透压以应对盐胁迫的危害[9]。过表达MeRS40(Arginine/serine-rich 40)拟南芥(Arabidopsis thaliana)具有较高的耐盐性,转基因拟南芥中过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)和过氧化物酶(peroxidase,POD)活性显著高于对照,而在木薯中过表达MeRS40降低了耐盐性[10]。挖掘木薯盐胁迫响应基因,解析木薯耐盐分子机制对培育耐盐作物品种至关重要。盐胁迫下,对木薯植株进行转录组分析,共鉴定到36个与抗逆相关的差异表达基因,涉及植物激素信号转导、蛋氨酸代谢、植物昼夜节律等多条参与植物抗逆的信号通路[11]。
茉莉酸作为植物体内重要的胁迫激素,在受到盐胁迫时,通过调控植物的防御反应机制从而抵御侵害,起着至关重要的作用。外源茉莉酸喷施提高了小麦(Triticum aestivum)幼苗抗氧化化合物浓度和抗氧化酶活性,使其耐盐能力增强[12]。茉莉酸通过其信号转导途径发挥作用,当受到非生物胁迫时,JA-lle(茉莉酸异亮氨酸)含量增高,其受体F-box蛋白COI1(CORONATINE INSENSITIVE1)位于E3泛素连接酶SCFCOI1复合体中,相互结合进而使JAZ(Jasmonate ZIM-domain)阻遏蛋白泛素化,并通过26S蛋白酶体途径降解,从而释放转录因子,启动茉莉酸的胁迫响应基因[13]。JAZ蛋白作为阻遏蛋白在茉莉酸途径起到抑制因子的作用,除此之外,JAZs还可以同其他转录因子及共阻遏蛋白相结合,从而使茉莉酸信号通路与其他激素的信号通路相串联[14]。前期研究发现,JAZ2作为盐胁迫响应基因参与并调控生长代谢来应对盐胁迫。分析ScJAZ家族基因在盐胁迫下的表达,其中ScJAZ2响应盐胁迫的诱导最为强烈[15]。菠萝(Ananas comosus)[16]和薄荷(Mentha canadensis)[17]中JAZ2在盐处理下表达谱分析得出,AcJAZ2和McJAZ2同样受盐胁迫的诱导,表达量不同程度地上升。虽然许多研究都表明,JAZ2在盐胁迫条件下,其表达量均处于上升趋势,但是只有少量研究确定了JAZ2在植物应对盐胁迫中的功能。例如在拟南芥中,过表达GsJAZ2会导致NHX1(Na+/H+ antiporter 1)的表达水平上升,提高Na+跨膜转运蛋白活性来正调控植株耐盐性[18]。虽然当前研究表明JAZ正调控多种植物的耐盐性,但在木薯中的相关研究很少。在木薯中,有研究发现,大多数JAZ基因表达受盐胁迫的诱导显著上调,其中,Manes.03G042500和Manes.16G088300上调较为明显。但JAZ蛋白如何调控木薯耐盐性,以及其分子机制都尚未报道[19]。因此,本研究从木薯‘华南124’(‘SC124’)中克隆获得MeJAZ2.2基因,分析其在盐胁迫下表达量和表型变化,并通过酵母双杂交文库筛选实验获取候选互作基因,旨在为培育木薯抗逆新品种提供理论支持。
-
实验中选取的植物材料为木薯‘华南124’(‘SC124’),由海南大学儋州校区木薯种质资源圃保存。将成熟的木薯杆截取为15~20 cm的茎段,培养在温室内营养土和蛭石等比例混合的基质(V营养土:V蛭石 = 1:1)中,生长条件为12 h光照和12 h黑暗,温度控制在27~30 ℃,湿度为70%[20]。
DH5α大肠杆菌感受态细胞(DL1001S)购自上海唯地生物技术有限公司,GV3101农杆菌感受态细胞(ZC141)购自北京庄盟国际生物基因科技有限公司,逆转录试剂盒(K1622)购自赛默飞世尔科技公司,琼脂糖凝胶DNA回收试剂盒(DP209)和质粒小提试剂盒(DP104)购自天根生化科技公司,DO Supplement-Leu/-Trp(PM2220)与DO Supplement-Ade-His-Leu-Trp(PM2110)营养缺陷培养基购自北京酷来搏科技有限公司。木薯cDNA文库,AH109酵母感受态细胞,酵母双杂交表达载体pGADT7和pGBKT7等均来自实验室冻存。
引物合成和测序均由北京擎科生物股份有限公司完成。
-
从植物基因组网站Phytozome(
https://phytozome-next.jgi.doe.gov/ )上获取拟南芥JAZ2基因序列,以其为模式序列在木薯基因组文库中BLAST搜索,查找到两个同源性较高的基因MeJAZ2.1和MeJAZ2.2。从Phytozome上下载拟南芥、水稻(Oryza sativa)、番茄(Solanum lycopersicum)等多种植物JAZ2的氨基酸序列,数据整理完成后通过MEGA 11分子进化遗传分析软件进行序列比对和系统进化树的构建[21]。将MeJAZ2.1和MeJAZ2.2的氨基酸序列在NCBI的CDD(https://www.ncbi.nlm.nih.gov/cdd )数据库中分析比对,查找到的保守结构域数据通过ChiPlot(https://chiplot.online/ )绘图。 -
选取培养3~4周且生长状况良好表型一致的木薯‘SC124’植株,配置200 mmol·L−1的NaCl溶液处理木薯根部,设置3次重复。分别在0、3、6、12、24 h取木薯上层叶片,样品剪下后迅速放入液氮中,后置于−80 ℃冰箱冻存。将木薯叶片研磨,通过CTAB法[22]提取木薯RNA,并使用逆转录试剂盒将RNA反转为cDNA。使用SGN VIGS Tool筛选出基因中特异性较高的片段,利用Primer5设计定量引物,内参引物为MeEF1a-F/R(表1)。通过实时荧光定量逆转录PCR(Real-time RT-PCR, RT-qPCR)获得数据,分析在盐胁迫下MeJAZ2.1和MeJAZ2.2表达量变化。
表 1 引物序列信息
Table 1. Primer sequence information
引物名称
Primer引物序列(5′−3′)
Primer sequence (5′−3′)pGADT7-MeJAZ2.2-F gccatggaggccagtgaattcATGGCTGGTTCGCCGGAA pGADT7-MeJAZ2.2-R atgcccacccgggtggaattcCTACTGGCCAAGATTGAGCAGC pGBKT7-F GTAATACGACTCACTATAGGGCGA pGBKT7-R TTTTCGTTTTAAAACCTAAGAGTC qMeJAZ2.1-F ATGGCTGGCTCGCCGGAATTCGTTG qMeJAZ2.1-R ATTTCCTTCACCGCTG qMeJAZ2.2-F CGCGGATCCAAGGAGGTTATGCTCTTGGC qMeJAZ2.2-R TCCCCCGGGGGCTTGAGTGGGAGGTTGC MeEF1a-F TGAACCACCCTGGTCAGATTGGAA MeEF1a-R AACTTGGGCTCCTTCTCAAGCTCT MeJAZ2.2-pTRV-F CGCGGATCCAAGGAGGTTATGCTCTTGGC MeJAZ2.2-pTRV-R TCCCCCGGGGGCTTGAGTGGGAGGTTGC -
以木薯cDNA为模板,用特异性引物MeJAZ2.2-pTRV F/R(表1)扩取目的片段,在T4 DNA连接酶催化下与pTRV2载体连接,并转化到DH5α大肠杆菌中。挑选阳性单克隆37 ℃过夜培养,小提试剂盒提取质粒并酶切和测序检测,转入GV3101农杆菌感受态中,以备后续侵染使用。将连接pTRV2载体的MeJAZ2.2的农杆菌菌株和pTRV1农杆菌菌株放入28 ℃摇床过夜培养后,转入20 mL LB培养基中活化5~6 h,3 500 r·min−1离心10 min弃上清液。收集菌体用配置好的渗透液(10 mmol·L−1 MgCl2,50 mmol·L−1 MES,150 µmol·L−1 AS)悬浮,稀释至OD600=0.6,等体积混合。用一次性注射器将农杆菌菌液注射到木薯下层叶片中,12~14 d后取木薯上层叶片定量分析MeJAZ2.2基因抑制水平[23]。用200 mmol·L−1的NaCl溶液处理抑制株和对照株,观察0 d和4 d的木薯表型变化。
-
从木薯‘SC124’叶片中提取总RNA并反转为cDNA。在Phytozome木薯数据库中查找到MeJAZ2.2的编码区序列(csoding sequence,CDS),用Primer5设计引物pGADT7-MeJAZ2.2-F/R(表1)。以cDNA为模板扩增MeJAZ2.2的CDS,凝胶电泳检测片段长度。将PCR产物利用胶回收试剂盒回收,同EcoR Ⅰ单酶切的pGADT7空载质粒同源重组连接,并转化到DH5α大肠杆菌中。菌落PCR验证条带大小同目的基因一致,用氨苄抗性液体培养基培养过夜。质粒小提试剂盒提取质粒,最后通过EcoR Ⅰ单酶切验证和测序验证载体构建成功。
-
将pGADT7-MeJAZ2.2质粒和pGBKT7空载质粒等量混匀,转入制备好的AH109感受态细胞中,以pGADT7空载和pGBKT7空载为负对照。转化后的酵母菌液均匀涂在SD/-Leu/-Trp固体培养基上,28 ℃培养箱放置3~4 d。挑选SD/-Leu/-Trp固体培养基上生长的酵母单菌落,接种到SD/-Leu/-Trp液体培养基中,摇起后稀释100、10−1、10−2、10−3 4个浓度梯度,分别点板于SD/-Leu/-Trp和SD/-Ade-His-Leu-Trp固体缺陷培养基上。检测标准:SD/-Leu/-Trp固体培养基上有酵母菌落生长,SD/-Ade-His-Leu-Trp固体培养基上无酵母菌落生长,可证明MeJAZ2.2蛋白无自激活活性;若2种固体培养基上均有酵母菌落生长,则认定MeJAZ2.2蛋白存在自激活活性。
-
将pGADT7-MeJAZ2.2质粒与木薯cDNA文库质粒等量混匀,每份质粒0.1 μg。加入1 μg预变性的Carrier DNA(鲑鱼精DNA)和600 μL的PEG/LiAc制成酵母转化的预混液。将200 μL制备好的AH109酵母感受态细胞加入预混液中,放入28 ℃培养箱1 h后加入70 μL的DMSO,42 ℃水浴30 min完成转化。12 000 r·min−1离心30 s弃上清,沉淀用400 μL现配制的1×TE溶液悬浮,分别涂于SD/-Leu/-Trp和SD/-Ade-His-Leu-Trp固体缺陷培养基上,置于28 ℃培养箱中3~4 d[24]。二缺培养基上酵母菌落生长状况良好,则证明质粒转化成功。选取四缺板上的酵母单菌落进行菌落PCR,产物由北京擎科生物公司测序,在Phytozome木薯数据库中分析对比基因序列,筛选出候选互作蛋白。
-
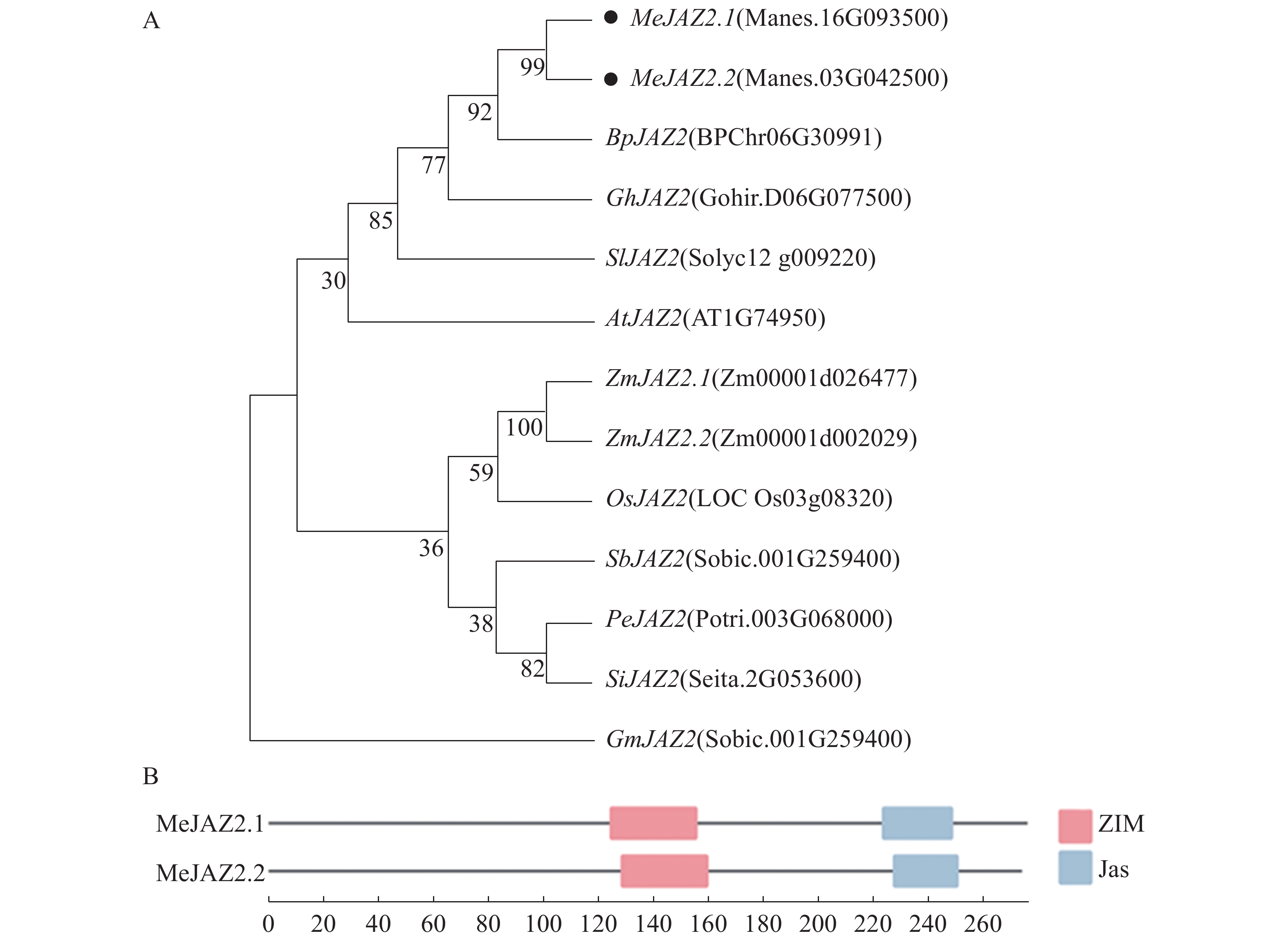
为了明确MeJAZ2基因与其他植物的进化关系。将木薯、拟南芥、水稻、番茄等10个植物的JAZ2基因同MeJAZ2基因采用最大似然法构建系统发育进化树(图1-A),Bootstrap method参数设定为1 000。发现木薯MeJAZ2.1、MeJAZ2.2同白桦BpJAZ2、棉花GhJAZ2及番茄SIJAZ2具有较近的亲缘关系。为进一步探索MeJAZ2.1和MeJAZ2.2的功能,通过保守结构分析可以发现,MeJAZ2.1位于氨基酸序列N端124~156为ZIM保守结构域,位于C端223~249为Jas保守结构域。MeJAZ2.2氨基酸序列在128~160和227~251位置同样含有ZIM和Jas保守结构域(图1-B)。ZIM和Jas作为JAZ家族特有的强保守结构域,从而证实MeJAZ2.1和MeJAZ2.2都属于JAZ基因家族。
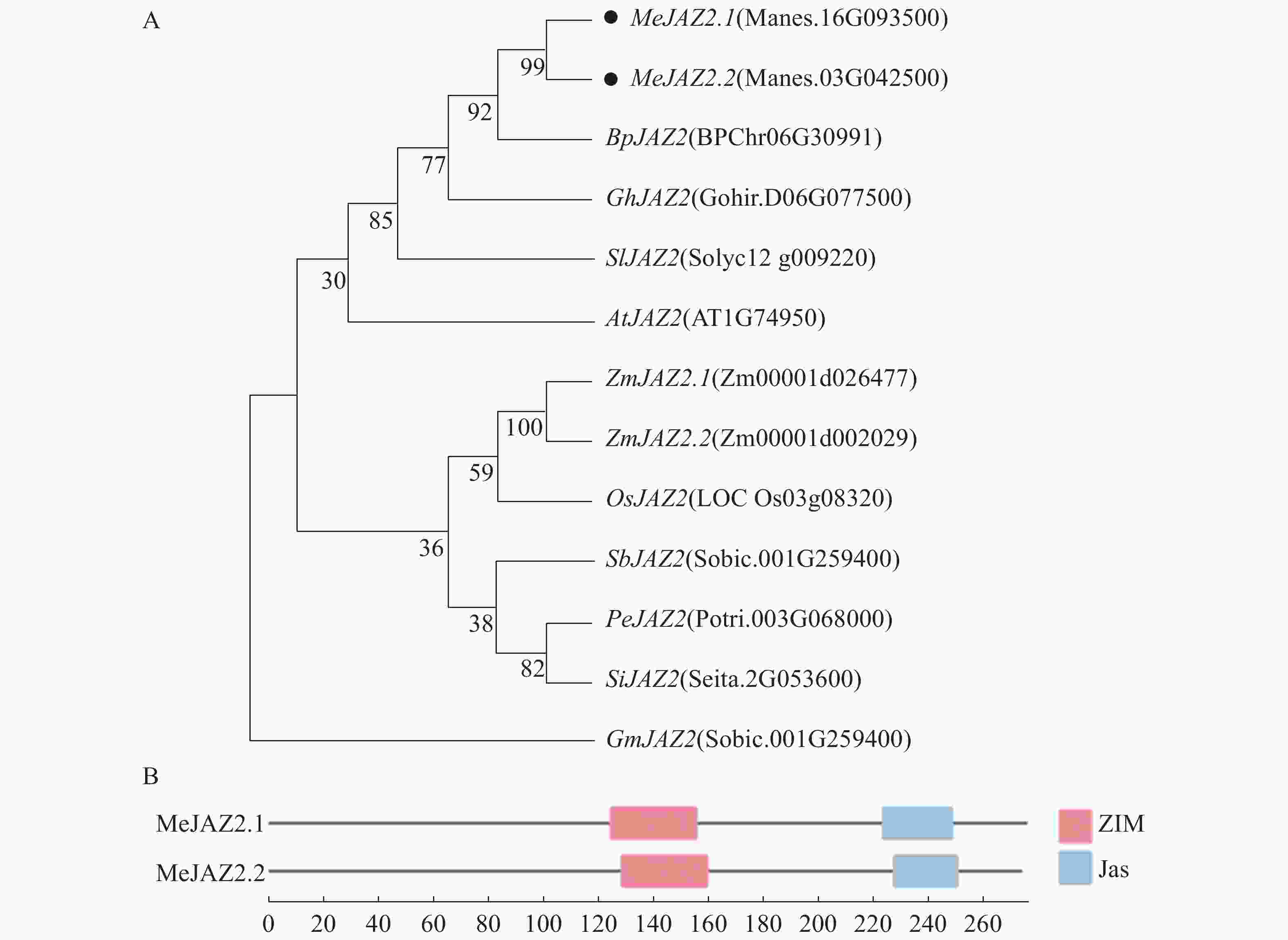
图 1 木薯与其他物种JAZ2的系统进化树(A)和蛋白质保守结构域分析(B)
Figure 1. Phylogenetic tree of JAZ2 (A) of cassava and other species and protein conserved domain analysis (B)
-
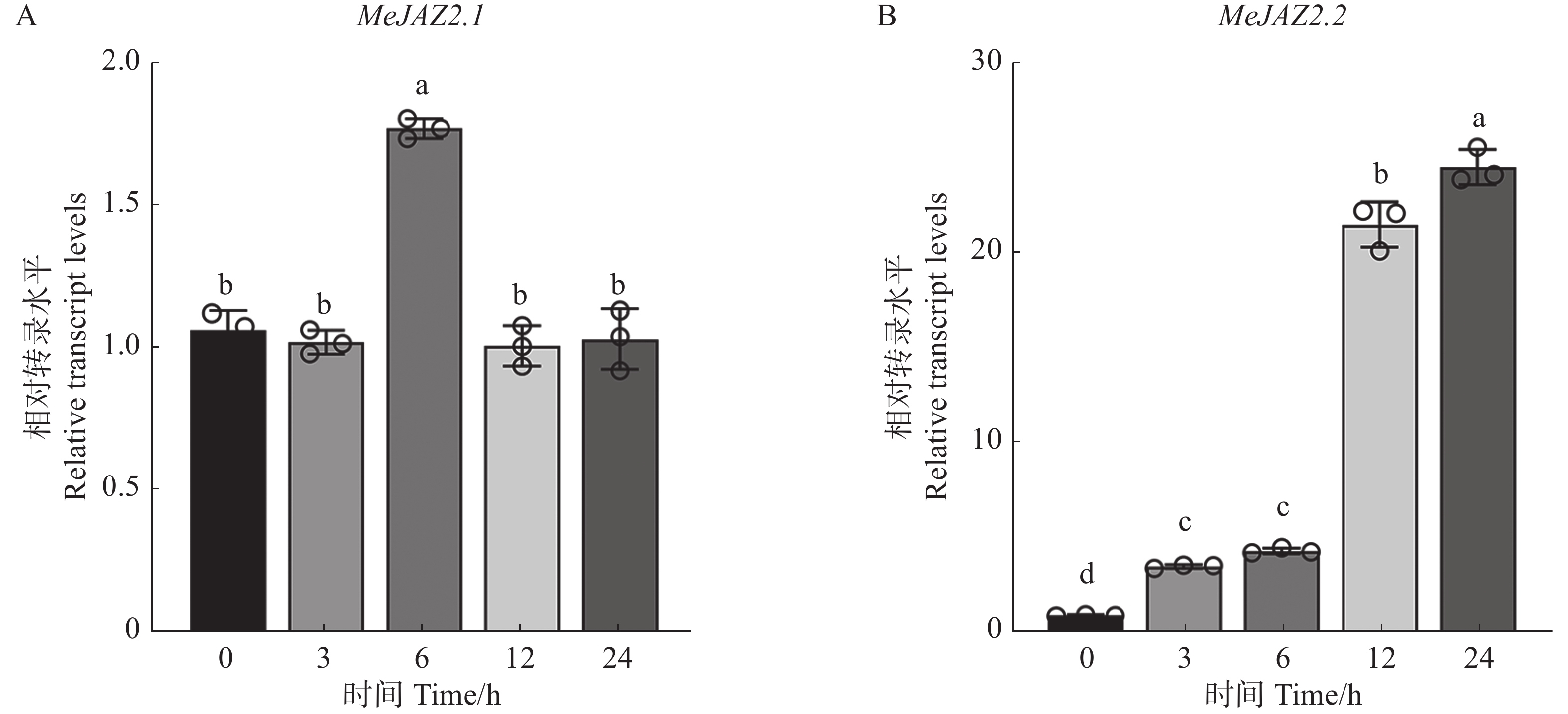
为探究MeJAZ2.1和MeJAZ2.2在木薯盐胁迫过程中表达量变化。用200 mmol·L−1浓度氯化钠溶液处理样品根部,通过RT-qPCR技术进一步分析MeJAZ2.1和MeJAZ2.2在盐胁迫下基因表达水平的变化。结果如图2所示,MeJAZ2.1在盐胁迫条件下其表达量除在6 h略有上调外,均稳定在1倍左右(图2-A)。MeJAZ2.2表达量在受胁迫后显著上调,处理12 h时上调了21倍,在24 h达到峰值,上调至24倍(图2-B)。上述结果表明,MeJAZ2.1和MeJAZ2.2参与了对盐胁迫的响应。MeJAZ2.2表达量变化更为明显,选取其进行下一步研究。
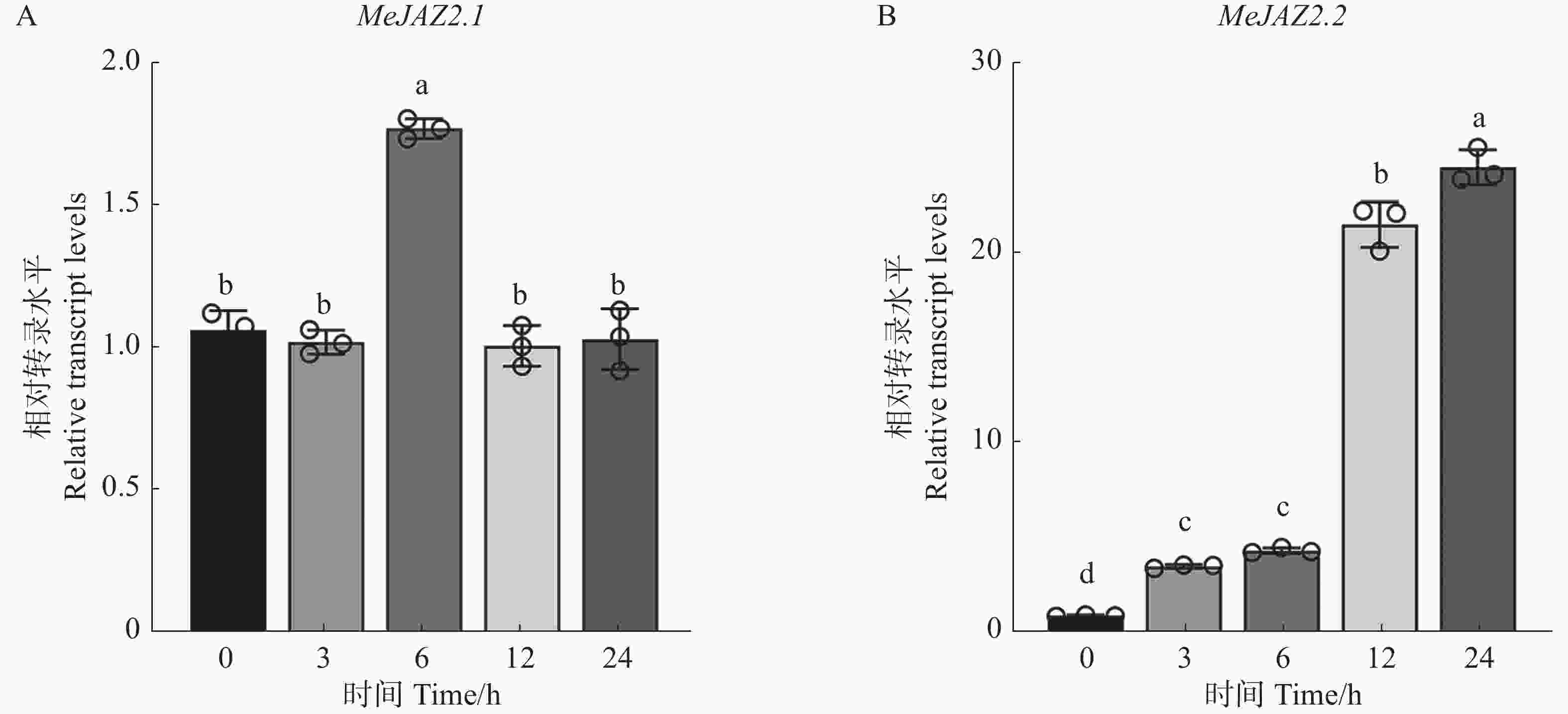
图 2 MeJAZ2.1和MeJAZ2.2在盐胁迫下表达量变化
Figure 2. Analysis of expression levels of MeJAZ2.1 and MeJAZ2.2 under salt stress
-
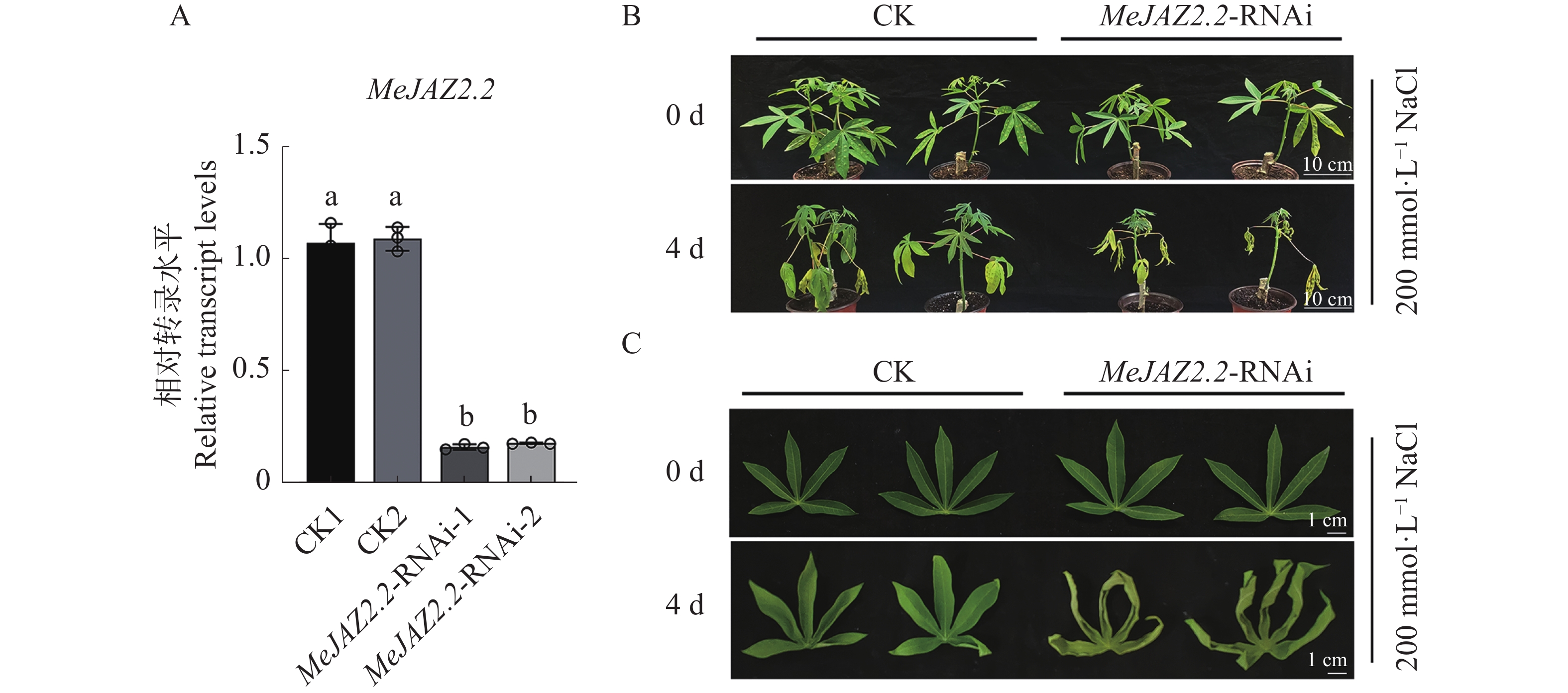
为鉴定盐胁迫下MeJAZ2.2-RNAi植株木薯幼苗的表型,构建了MeJAZ2.2抑制植株。与对照相比,抑制株MeJAZ2.2-RNAi-1和MeJAZ2.2-RNAi-2中MeJAZ2.2表达量下调至0.16和0.18(图3-A),已成功抑制。对野生型和MeJAZ2.2-RNAi植株木薯幼苗用200 mmol·L−1的氯化钠处理4 d后进行拍照分析。结果如图3-B所示,处理4 d后,MeJAZ2.2-RNAi植株木薯幼苗和野生型木薯幼苗生长均受抑制,但MeJAZ2.2-RNAi植株损伤更为严重。进一步观察叶片表型,与对照相比MeJAZ2.2-RNAi植株叶片变黄程度更明显,叶片卷曲更为严重(图3-C)。通过表型分析得出,MeJAZ2.2可能正调控木薯对盐胁迫的抗性。
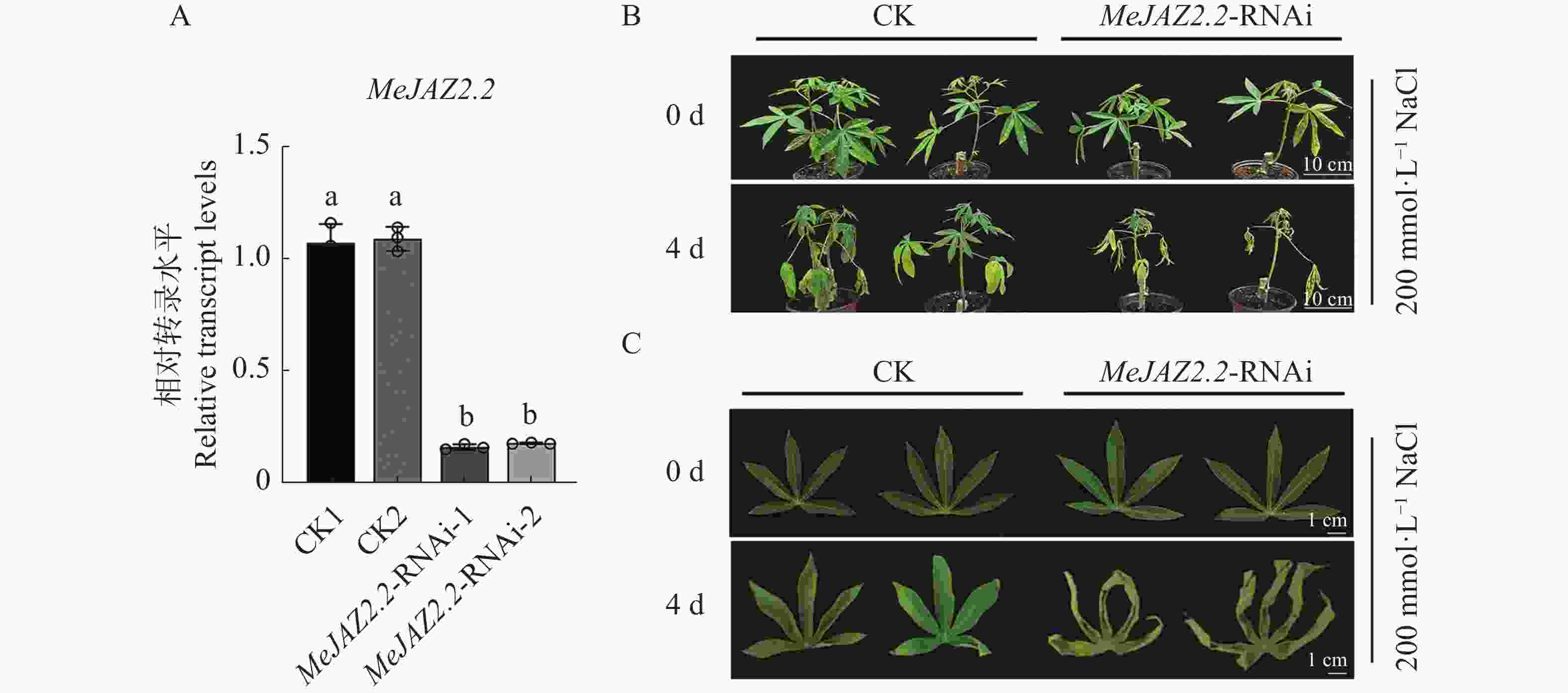
图 3 MeJAZ2.2抑制株构建及表型鉴定
Figure 3. MeJAZ2.2 inhibited plant construction and phenotypic identification
-
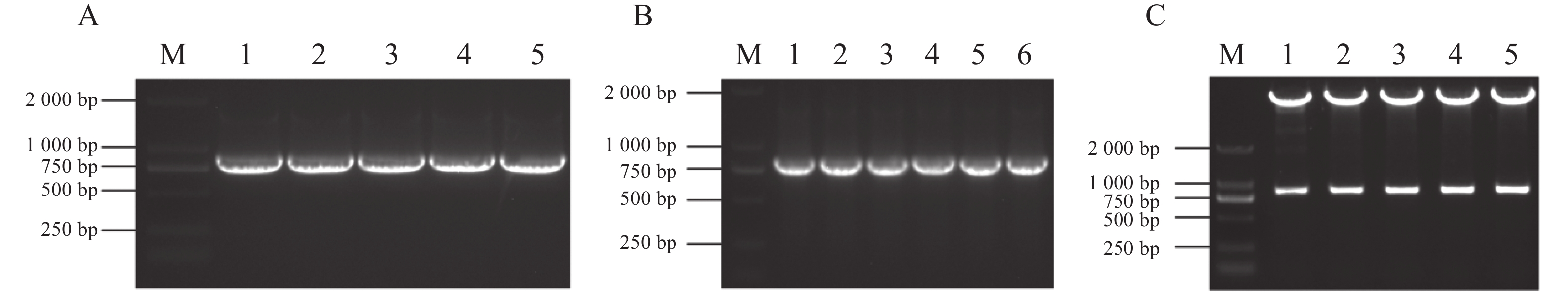
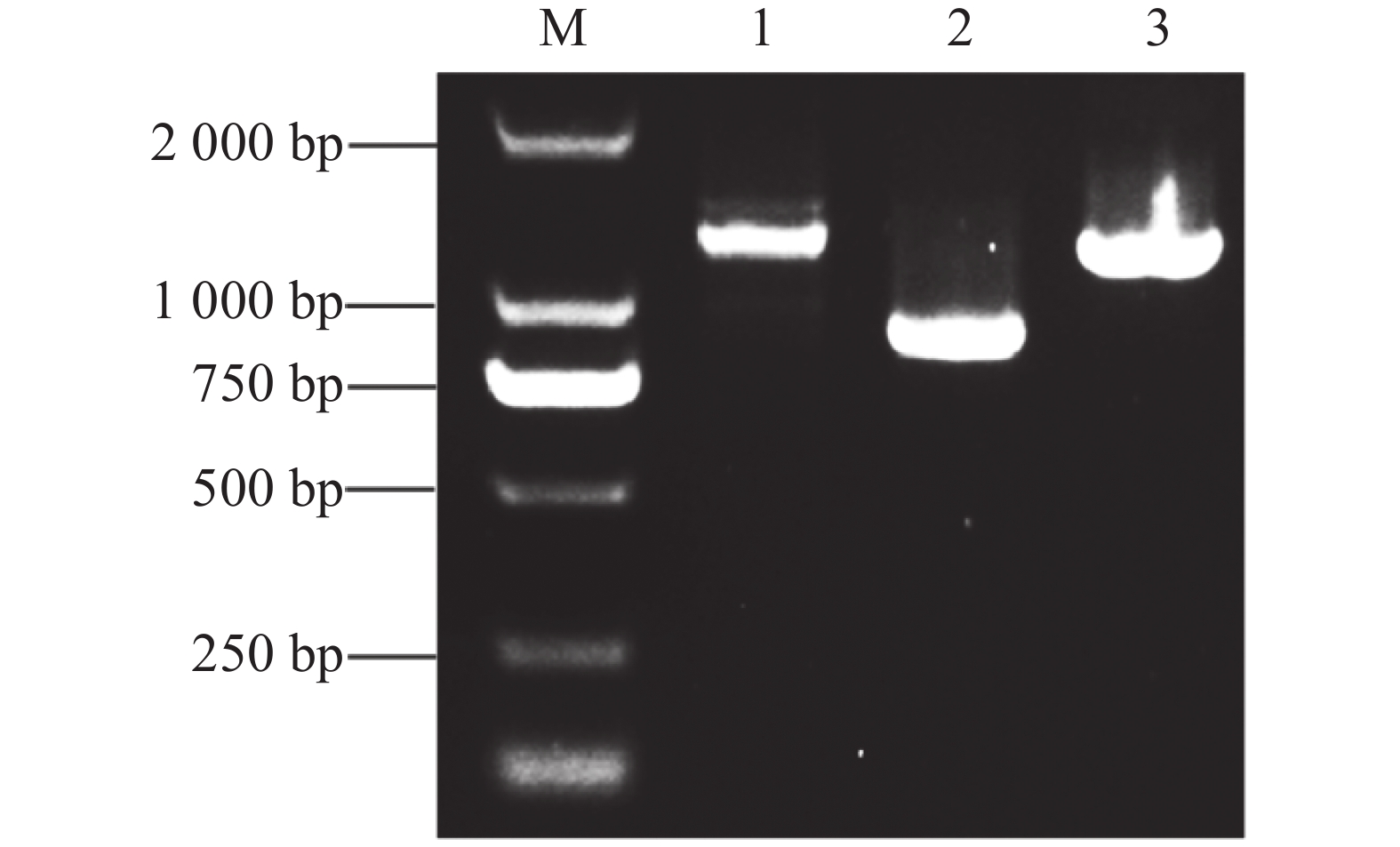
为了进一步解析MeJAZ2.2调控木薯耐盐性的分子机制,尝试筛选和鉴定MeJAZ2.2的互作蛋白。通过木薯数据库查询获得MeJAZ2.2基因序列,发现其CDS为825 bp,共编码274个氨基酸。以木薯cDNA为模板,用引物pGADT7-MeJAZ2.2-F/R进行PCR扩增,凝胶电泳检测到片段大小约800 bp,与目的基因实际长度一致(图4-A)。利用同源重组技术将目的片段连接到EcoR Ⅰ单酶切pGADT7载体上。连接产物转化到DH5α大肠杆菌感受细胞中,挑选单菌落作为模板并PCR验证。结果显示,扩增片段同目的基因克隆长度相同(图4-B)。挑选5个阳性克隆,对其质粒酶切验证(图4-C)并测序检验。保留测序结果与目的基因一致的克隆用于后续酵母双杂交实验。
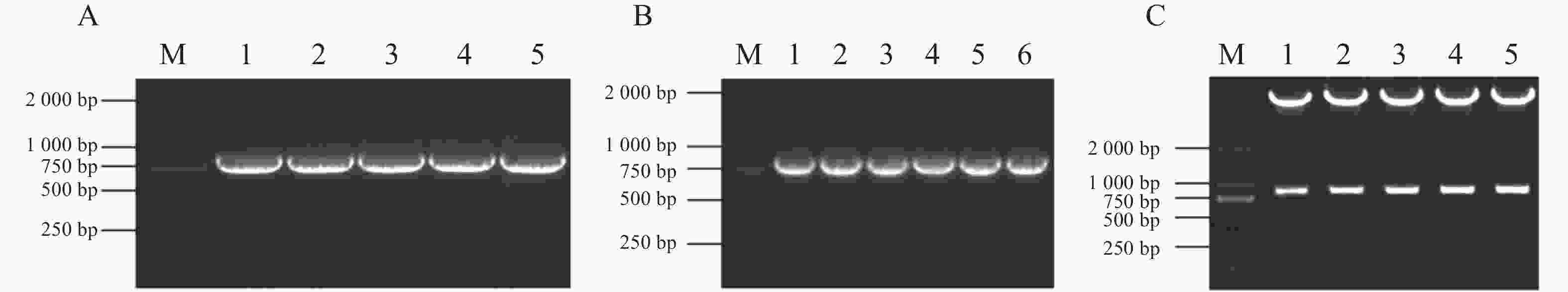
图 4 载体的构建
Figure 4. Vector construction
-
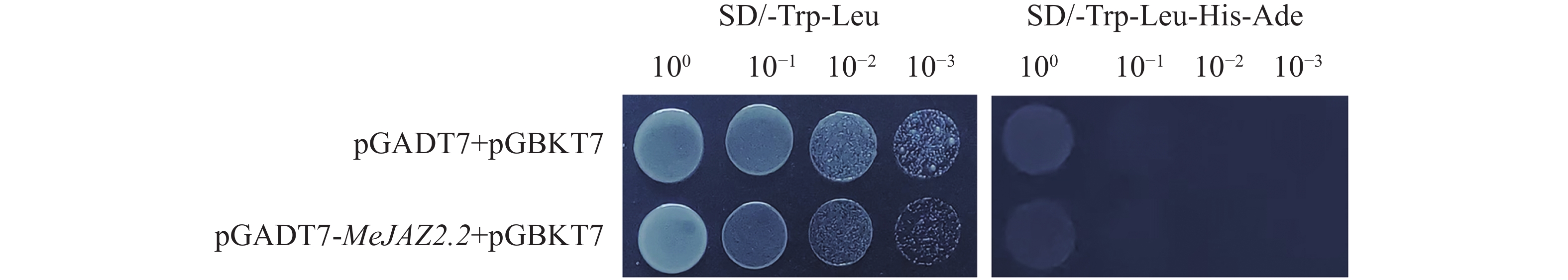
为避免酵母双杂交实验中存在假阳性转化子,首先验证了pGADT7-MeJAZ2.2是否存在自激活活性。将pGADT7-MeJAZ2.2和pGBKT7空载质粒共同转化到AH109酵母菌株中,并设置阴性对照(pGADT7空载+pGBKT7空载)。酵母菌液稀释3个梯度分别点于二缺(SD/-Leu/-Trp)和四缺(SD/-Ade-His-Leu-Trp)固体培养基上,观察酵母生长状况。结果显示,在四缺固体培养基上均无酵母菌落生长,而在二缺固体培养基上酵母正常生长,且数量随酵母稀释梯度的增长而降低(图5)。以上实验结果证明,MeJAZ2.2蛋白不能单独激活报告基因,不存在自激活活性,因此可作为诱饵蛋白来进行蛋白互作筛选。
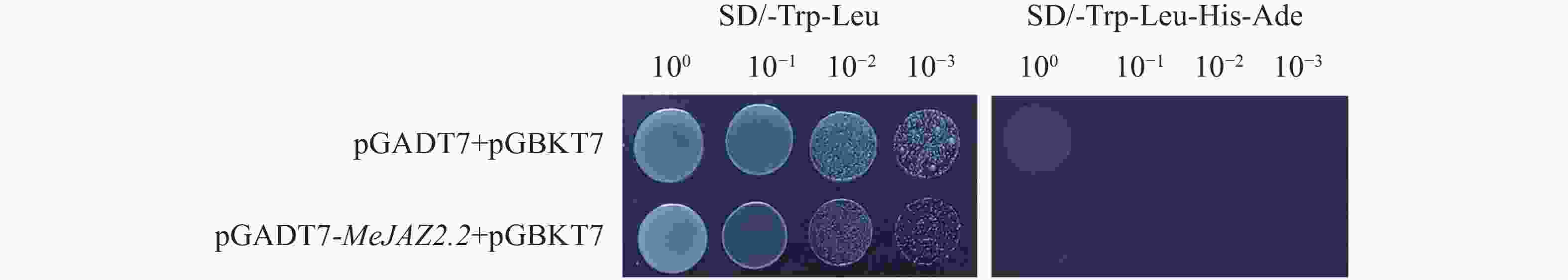
图 5 pGADT7-MeJAZ2.2自激活验证
Figure 5. Verification of pGADT7-MeJAZ2.2 self-activation
-
提取质粒pGADT7-MeJAZ2.2与木薯SC124的cDNA文库菌株质粒共转化到制备好的AH109酵母感受态细胞中,分别涂布于二缺(SD/-Leu/-Trp)和四缺(SD/-Ade-His-Leu-Trp)固体培养基上。二缺平板上酵母菌落正常生长,即质粒转入成功,选取四缺平板上单菌落阳性克隆作为模板,利用pGBKT7空载通用引物进行PCR扩增,凝胶电泳检测到3条大小不同的条带(图6)。
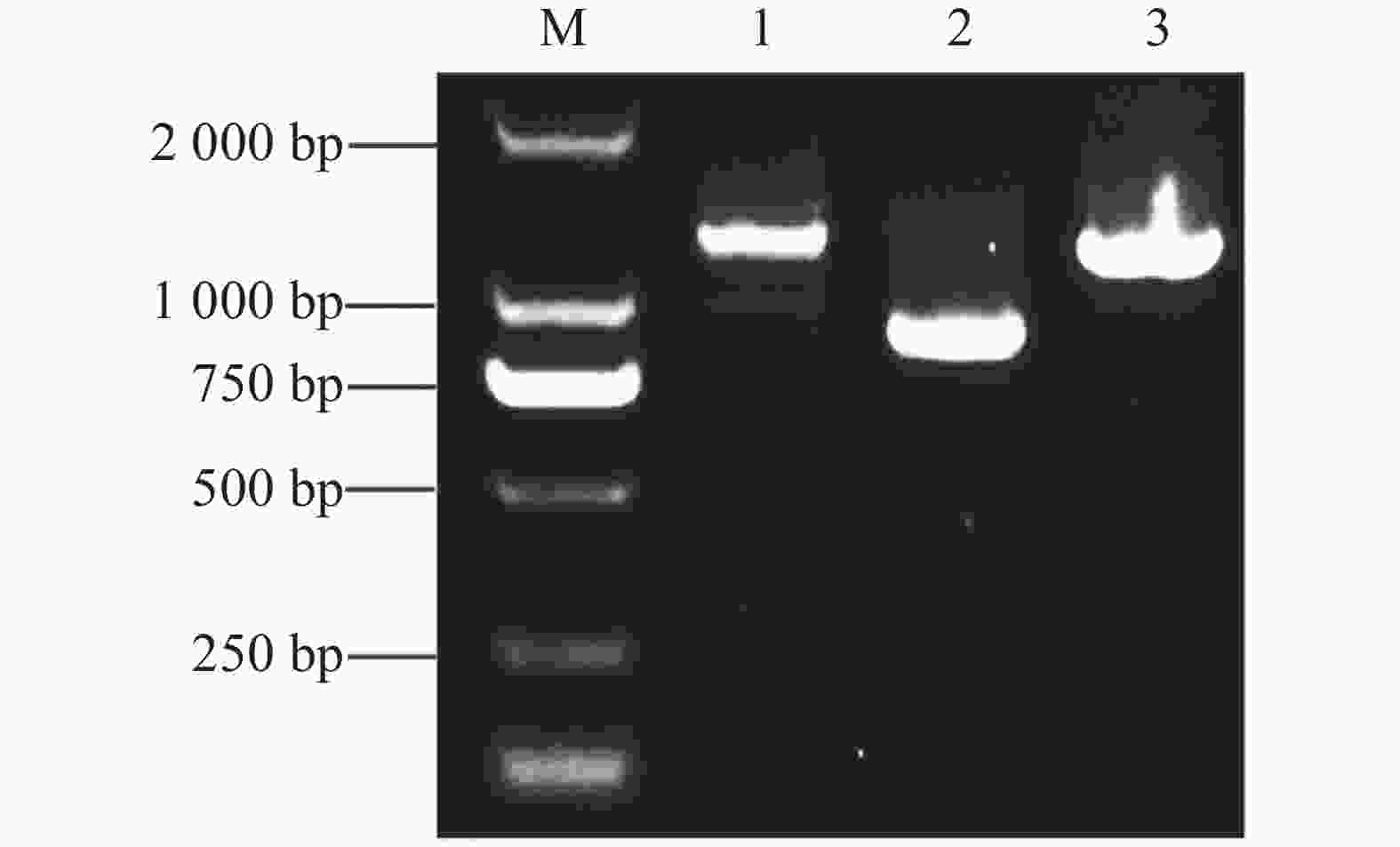
图 6 酵母菌落PCR验证
Figure 6. PCR verification of yeast colonies
将PCR产物回收后送测序,均成功获得序列信息。在Phytozome木薯数据库中比对共获得3个候选互作蛋白(表2),分别是谷氨酰胺合成酶(GLUTAMINE SYNTHETASE)、泛素3(UBIQUITIN 3)和FRIGIDA-LIKE蛋白(FRIGIDA-LIKE PROTEIN)。
表 2 MeJAZ2.2候选互作蛋白信息
Table 2. Information of MeJAZ2.2 candidate interacting proteins
编号
Code基因注释
Gene annotation蛋白名
Protein1 谷氨酰胺合成酶
GLUTAMINE SYNTHETASEGS 2 泛素 3
UBIQUITIN 3Ub3 3 FRIGIDA-LIKE蛋白
FRIGIDA-LIKE PROTEINFRI-L -
茉莉酸作为重要的内源信号分子,在拟南芥[25]、大豆(Glycine max)[26]、大麦(Hordeum vulgare)[27]中调控细胞的伸长、气孔的关闭、光合作用效率来影响耐盐性。JAZ2作为茉莉酸信号途径的抑制因子,包含与COI1和MYC2(myelocytomatosis proteins 2)相互作用所必需的2个强保守结构域ZIM和Jas,其中Jas结构域介导JAZ2蛋白与MYC2互作,抑制其转录活性,从而抑制茉莉酸胁迫响应基因的表达[28]。在拟南芥中JAZ2通过茉莉酸信号转导途径介导植株对盐胁迫的抗性,但其在热带作物木薯中调控盐胁迫的功能以及机制未知。因此,本研究通过分离鉴定木薯中MeJAZ2对其表达模式进行分析,并进行功能验证,进一步筛选互作蛋白以对其调控机制进行解析。
植物通过调控基因表达水平以适应外界环境的变化,如豌豆(Pisum sativum)在受到盐胁迫下,PsTPSs(Trehalose-6-phosphate synthase)基因表达模式发生显著变化,其中PsTPS4、PsTPS6 和 PsTPS19上调,通过渗透调节以应对胁迫[29]。同样,盐胁迫也诱导JAZ基因表达水平发生变化,如从苜蓿基因组中鉴定出11个JAZ基因,在盐胁迫条件下所有MsJAZ基因均有响应,并随时间推移而不同程度上升[30]。NaCl处理下高粱JAZ2基因表达量显著升高,在12 h时达到最大值[31]。本研究从木薯中分离出MeJAZ2.2,结构域分析发现含有ZIM和Jas两个强保守结构域,并在24 h内其表达量受盐胁迫时间延长而升高,与前人研究相一致。但在盐处理12 h时,MeJAZ2.2表达量上升更为明显,并在24 h时达到峰值。结果表明,该基因的表达受盐胁迫的诱导,可能参与木薯对盐胁迫的响应。表型鉴定对探究植物基因功能至关重要,观察表型变化从而解析目的基因在植物生长发育和应对非生物胁迫中的作用。过表达GsJAZ2表现出更快的抽薹率,增强了耐盐性,但植株形态在盐胁迫下的变化是不确定的[18]。在JAZ基因家族其他成员中,盐胁迫下的形态变化受基因抑制的影响,如棉花GaJAZ1沉默植株在盐处理下,植株萎蔫,叶片卷曲和发黄程度更为严重,导致耐盐性降低[32]。在木薯幼苗中抑制MeJAZ2.2,盐胁迫加快了叶片黄化和蜷曲,这与棉花叶片中表型相似。但不同的是,木薯JAZ2.2抑制株萎蔫和上层叶片黄化程度较轻,这可能是JAZ在不同植物中参与盐胁迫途径差异所导致的。通过表型分析推测MeJAZ2.2 可能正调控木薯对盐胁迫的抗性。
JAZs作为茉莉酸信号途径的调控枢纽与其他调节因子互作,协同调控植物对盐胁迫的抗性。在番茄中,JAZ蛋白同SIWRKY57-SIVQ21/SIVQ16模块互作,增强茉莉酸诱导的盐胁迫相关基因的表达[33]。OsJAZ9通过与OsNINJA和OsbHLH互作,形成转录调控复合物来介导水稻中参与盐胁迫的基因的表达[34]。在确定MeJAZ2.2正调控木薯耐盐性的基础上,本研究通过酵母双杂交实验筛选到3个MeJAZ2.2候选互作蛋白,分别是GS、Ub3和FRI-L蛋白。其中GS普遍存在于高等植物体内,消耗ATP从而催化谷氨酸和铵离子合成谷氨酰胺,是植物体内氮代谢途径的重要组成部分[35]。近年来,有研究表明,GS与植物的耐盐性密切相关。水稻转基因植株G39-2中GS2表达量上升,光呼吸作用增强,从而提高了水稻的耐盐性[36]。在盐胁迫条件下,过表达水稻GS1;2基因提高了植株代谢水平,并导致GS活性和可溶性蛋白浓度增加,使植株对盐胁迫具有更高的敏感性[37]。植株幼苗在茉莉酸甲酯(MeJA)处理后,体内GS、谷氨酸合成酶(Glutamate synthase,GOGAT)活性升高,改善氮代谢以提高植株渗透调节能力,从而应对盐胁迫的侵害[38]。因此推测,茉莉酸抑制因子MeJAZ2.2有可能通过与MeGS的互作,参与木薯对盐胁迫的响应。后续实验将通过Pull-down和双分子荧光互补(bimolecular fluorescence complementation,BiFC)实验进一步明确MeJAZ2.2和MeGS的互作关系。下一步通过生理生化测定实验,解析其调控木薯抗盐胁迫的分子机制,为木薯抗逆品种培育提供候选基因。
MeJAZ2.2基因的克隆及互作蛋白筛选
DOI: 10.15886/j.cnki.rdswxb.20240169
 CSTR: 32425.14.j.cnki.rdswxb.20240169
CSTR: 32425.14.j.cnki.rdswxb.20240169
Cloning and interaction protein screening of MeJAZ2.2 gene
-
摘要: 木薯(Manihot esculenta)作为热区重要的粮食作物,其产量受盐胁迫影响,粮食安全无法得到保障。JAZ(jasmonate ZIM-domain, JAZ)蛋白作为茉莉酸信号通路中的重要组成部分,参与调控多种作物对盐胁迫的抗性。为探究JAZ蛋白是否调控木薯抗盐胁迫及其中的调控机制。本研究通过生物信息学筛选到2个同源基因MeJAZ2.1和MeJAZ2.2,进化树和保守结构域分析发现均含有ZIM和Jas两个保守结构域,都属于JAZ基因家族。进一步研究发现,MeJAZ2.2在响应木薯盐胁迫中表达量变化更为明显。与野生型相比,MeJAZ2.2抑制木薯植株对盐胁迫的敏感性升高,这表明MeJAZ2.2可能正调控木薯对盐胁迫的抗性。本研究还构建了pGADT7-MeJAZ2.2诱饵载体并通过酵母双杂交实验发现其不存在自激活活性,并进一步筛选到3个MeJAZ2.2的候选互作蛋白,包括谷氨酰胺合成酶(GLUTAMINE SYNTHETASE)、泛素 3(UBIQUITIN 3)和FRIGIDA-LIKE蛋白(FRIGIDA-LIKE PROTEIN)。为进一步解析JAZ蛋白在木薯抗盐胁迫中的功能及其分子机制提供前期基础。Abstract: Cassava is an important food crop in tropical regions, but the yield of cassava is affected by salt stress, which endangers food security. JAZ (jasmonate ZIM-domain) proteins, as essential components in the jasmonate signaling pathway, are involved in regulating the tolerance to salt stress in a variety of crops. In order to investigate response of JAZ proteins in cassava to salt stress, as well as the underlying regulatory mechanisms two homologous genes MeJAZ2.1 and MeJAZ2.2 were identified from cassava variety SC124 through bioinformatics. The evolutionary tree and conserved domain analysis indicated that they both contain two conserved domains, ZIM and Jas, which belong to the JAZ gene family. Further research discovered that the expression level of MeJAZ2.2 changed more significantly in response to salt stress in cassava. MeJAZ2.2-silenced cassava plants were more susceptibility to salt stress compared to the wild type, indicating that MeJAZ2.2 may positively regulate cassava resistance to salt stress. The pGADT7-MeJAZ2.2 bait vector was constructed and no self-activating activity was found by yeast two-hybrid experiment. Moreover, three candidate interacting proteins of MeJAZ2.2 were screened, including glutamine synthetase (GS), ubiquitin 3 (Ub3), and FRIGIDA-LIKE PROTEIN (FRI-L), which provides a preliminary framework for analyzing the function and molecular mechanism of JAZ proteins to salt stress in cassava.
-
Key words:
- cassava /
- salt stress /
- MeJAZ2.2 /
- interacting protein
-
图 2 MeJAZ2.1和MeJAZ2.2在盐胁迫下表达量变化
注:小写字母表示存在显著性差异(P<0.05)。
Fig. 2 Analysis of expression levels of MeJAZ2.1 and MeJAZ2.2 under salt stress
Note: Lowercase letters indicate significant differences (P < 0.05).
图 3 MeJAZ2.2抑制株构建及表型鉴定
A. 应为MeJAZ2.2相对转录水平。小写字母表示存在显著性差异(P<0.05);B. 应为盐胁迫条件下木薯整株生长情况(标尺为10 cm);C.应为盐胁迫处理条件下木薯叶片表型(标尺为1 cm)。
Fig. 3 MeJAZ2.2 inhibited plant construction and phenotypic identification
A. Relative transcription level of MeJAZ2.2. Lowercase letters indicate significant differences (P < 0.05); B. Growth of cassava plant under salt stress (scale: 10 cm); C. Leaf morphology of cassava under salt stress treatment (scale: 1 cm).
图 4 载体的构建
注:M,DL2000 DNA Maker;A1~A5,MeJAZ2.2基因的克隆;B1~B6,大肠杆菌菌落PCR;C1~C5,酶切验证。
Fig. 4 Vector construction
Note: M, DL2000 DNA Maker; A1−A5, Cloning of MeJAZ2.2 gene; B1−B6, Escherichia coli colony PCR; C1−C5, Enzyme digestion validation.
图 5 pGADT7-MeJAZ2.2自激活验证
注:100、10−1、10−2、10−3为酵母菌液稀释倍数。SD/-Trp/-Leu为色氨酸(Trp)和亮氨酸(Leu)酵母缺陷型培养基;SD/-Trp-Lcu-His-Ade为色氨酸(Trp)、亮氨酸(Leu)、组氨酸(His)和腺嘌呤(Ade)酵母缺陷型培养基。
Fig. 5 Verification of pGADT7-MeJAZ2.2 self-activation
Note: 100, 10−1, 10−2 and 10−3 represent the yeast cell dilution fold. SD/-Trp/-Leu is a tryptophan (Trp) and leucine (Leu) yeast defective medium; SD/-Trp-Lcu-His-Ade is a Trp, Leu, histidine (His), and adenine (Ade) yeast defective medium.
图 6 酵母菌落PCR验证
注:M,DL2000 DNA Maker;1~3,酵母菌落PCR。
Fig. 6 PCR verification of yeast colonies
Note: M, DL2000 DNA Maker; 1−3, PCR of yeast colonies.
表 1 引物序列信息
Table 1 Primer sequence information
引物名称
Primer引物序列(5′−3′)
Primer sequence (5′−3′)pGADT7-MeJAZ2.2-F gccatggaggccagtgaattcATGGCTGGTTCGCCGGAA pGADT7-MeJAZ2.2-R atgcccacccgggtggaattcCTACTGGCCAAGATTGAGCAGC pGBKT7-F GTAATACGACTCACTATAGGGCGA pGBKT7-R TTTTCGTTTTAAAACCTAAGAGTC qMeJAZ2.1-F ATGGCTGGCTCGCCGGAATTCGTTG qMeJAZ2.1-R ATTTCCTTCACCGCTG qMeJAZ2.2-F CGCGGATCCAAGGAGGTTATGCTCTTGGC qMeJAZ2.2-R TCCCCCGGGGGCTTGAGTGGGAGGTTGC MeEF1a-F TGAACCACCCTGGTCAGATTGGAA MeEF1a-R AACTTGGGCTCCTTCTCAAGCTCT MeJAZ2.2-pTRV-F CGCGGATCCAAGGAGGTTATGCTCTTGGC MeJAZ2.2-pTRV-R TCCCCCGGGGGCTTGAGTGGGAGGTTGC  下载: 导出CSV
下载: 导出CSV
表 2 MeJAZ2.2候选互作蛋白信息
Table 2 Information of MeJAZ2.2 candidate interacting proteins
编号
Code基因注释
Gene annotation蛋白名
Protein1 谷氨酰胺合成酶
GLUTAMINE SYNTHETASEGS 2 泛素 3
UBIQUITIN 3Ub3 3 FRIGIDA-LIKE蛋白
FRIGIDA-LIKE PROTEINFRI-L
下载: 导出CSV
-
[1] 曹升, 陈江枫, 黄富宇, 等. 广西木薯产业现状分析及其发展建议[J]. 南方农业学报, 2021, 52(6): 1468−1476. https://doi.org/10.3969/j.issn.2095-1191.2021.06.005 doi: 10.3969/j.issn.2095-1191.2021.06.005 [2] Mcmahon J, Sayre R, Zidenga T. Cyanogenesis in cassava and its molecular manipulation for crop improvement [J]. Journal of Experimental Botany, 2022, 73(7): 1853−1867. https://doi.org/10.1093/jxb/erab545 doi: 10.1093/jxb/erab545 [3] 罗春芳, 杨龙, 欧珍贵, 等. 木薯食品加工的现状及前景展望[J]. 江西农业学报, 2019, 31(3): 107−112. [4] 王莉, 邓婷鹤. 2014年我国热作产品进出口贸易情况分析[J]. 中国热带农业, 2015(2): 4−7. https://doi.org/10.3969/j.issn.1673-0658.2015.02.001 doi: 10.3969/j.issn.1673-0658.2015.02.001 [5] 谭砚文, 李丛希, 曾华盛. 中国木薯生产和贸易发展分析[J]. 世界农业, 2018(10): 163−168. [6] 云雪雪, 陈雨生. 国际盐碱地开发动态及其对我国的启示[J]. 国土与自然资源研究, 2020(1): 84−87. [7] Yang Y, Guo Y. Elucidating the molecular mechanisms mediating plant salt-stress responses [J]. The New Phytologist, 2018, 217(2): 523−539. https://doi.org/10.1111/nph.14920 doi: 10.1111/nph.14920 [8] 庄盈婷. 木薯叶片和叶绿体应答盐胁迫的比较蛋白质组学研究[D]. 海口: 海南大学, 2014. [9] Fan X W, Sun J L, Cai Z, et al. MeSWEET15a/b genes play a role in the resistance of cassava (Manihot esculenta Crantz) to water and salt stress by modulating sugar distribution [J]. Plant Physiology and Biochemistry: PPB, 2023, 194: 394−405. https://doi.org/10.1016/j.plaphy.2022.11.027 doi: 10.1016/j.plaphy.2022.11.027 [10] Ma X W, Ma Q X, Ma M Q, et al. Cassava MeRS40 is required for the regulation of plant salt tolerance [J]. Journal of Integrative Agriculture, 2023, 22(5): 1396−1411. https://doi.org/10.1016/j.jia.2023.04.003 doi: 10.1016/j.jia.2023.04.003 [11] 吕瑞. 木薯耐盐种质筛选及盐种质SC8盐胁迫转录组分析[D]. 海口: 海南大学, 2019. [12] Qiu Z, Guo J, Zhu A, et al. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress [J]. Ecotoxicology and Environmental Safety, 2014, 104: 202−208. https://doi.org/10.1016/j.ecoenv.2014.03.014 doi: 10.1016/j.ecoenv.2014.03.014 [13] Miccono M L A, Yang H W, Demott L, et al. Review: losing JAZ4 for growth and defense [J]. Plant Science: an International Journal of Experimental Plant Biology, 2023, 335: 111816. [14] Wan S, Xin X F. Regulation and integration of plant jasmonate signaling: a comparative view of monocot and dicot [J]. Journal of Genetics and Genomics, 2022, 49(8): 704−714. https://doi.org/10.1016/j.jgg.2022.04.002 doi: 10.1016/j.jgg.2022.04.002 [15] Liu F, Sun T, Wang L, et al. Plant jasmonate ZIM domain genes: shedding light on structure and expression patterns of JAZ gene family in sugarcane [J]. BMC Genomics, 2017, 18(1): 771. https://doi.org/10.1186/s12864-017-4142-3 doi: 10.1186/s12864-017-4142-3 [16] Ye L, Cao L, Zhao X, et al. Investigation of the JASMONATE ZIM-DOMAIN gene family reveals the canonical JA-signaling pathway in pineapple [J]. Biology, 2022, 11(3): 445. https://doi.org/10.3390/biology11030445 doi: 10.3390/biology11030445 [17] Xu D B, Ma Y N, Qin T F, et al. Transcriptome-wide identification and characterization of the JAZ gene family in Mentha canadensis L [J]. International Journal of Molecular Sciences, 2021, 22(16): 8859. https://doi.org/10.3390/ijms22168859 doi: 10.3390/ijms22168859 [18] Zhu D, Cai H, Luo X, et al. Over-expression of a novel JAZ family gene from Glycine soja, increases salt and alkali stress tolerance [J]. Biochemical and Biophysical Research Communications, 2012, 426(2): 273−279. https://doi.org/10.1016/j.bbrc.2012.08.086 doi: 10.1016/j.bbrc.2012.08.086 [19] Zheng L, Wan Q, Wang H, et al. Genome-wide identification and expression of TIFY family in cassava (Manihot esculenta Crantz) [J]. Frontiers in Plant Science, 2022, 13: 1017840. https://doi.org/10.3389/fpls.2022.1017840 doi: 10.3389/fpls.2022.1017840 [20] Zheng L, Gao S, Bai Y, et al. NF-YC15 transcription factor activates ethylene biosynthesis and improves cassava disease resistance [J]. Plant Biotechnology Journal, 2024, 22(9): 2424−2434. https://doi.org/10.1111/pbi.14355 doi: 10.1111/pbi.14355 [21] Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11 [J]. Molecular Biology and Evolution, 2021, 38(7): 3022−3027. https://doi.org/10.1093/molbev/msab120 doi: 10.1093/molbev/msab120 [22] Mushtaq S, Tayyeb A, Firdaus-E-Bareen. A comparison of total RNA extraction methods for RT-PCR based differential expression of genes from Trichoderma atrobrunneum [J]. Journal of Microbiological Methods, 2022, 200: 106535. https://doi.org/10.1016/j.mimet.2022.106535 doi: 10.1016/j.mimet.2022.106535 [23] Zeng H, Xie Y, Liu G, et al. Agrobacterium-mediated gene transient overexpression and Tobacco rattle virus (TRV)-based gene silencing in cassava [J]. International Journal of Molecular Sciences, 2019, 20(16): 3976. https://doi.org/10.3390/ijms20163976 doi: 10.3390/ijms20163976 [24] Jin G, Qi J, Zu H, et al. Jasmonate-mediated gibberellin catabolism constrains growth during herbivore attack in rice [J]. The Plant Cell, 2023, 35(10): 3828−3844. https://doi.org/10.1093/plcell/koad191 doi: 10.1093/plcell/koad191 [25] Valenzuela C E, Acevedo-Acevedo O, Miranda G S, et al. Salt stress response triggers activation of the jasmonate signaling pathway leading to inhibition of cell elongation in Arabidopsis primary root [J]. Journal of Experimental Botany, 2016, 67(14): 4209−4220. https://doi.org/10.1093/jxb/erw202 doi: 10.1093/jxb/erw202 [26] Sheteiwy M S, Shao H, Qi W, et al. Seed priming and foliar application with jasmonic acid enhance salinity stress tolerance of soybean (Glycine max L.) seedlings [J]. Journal of the Science of Food and Agriculture, 2021, 101(5): 2027−2041. https://doi.org/10.1002/jsfa.10822 doi: 10.1002/jsfa.10822 [27] Aliakbari M, Tahmasebi S, Sisakht J N. Jasmonic acid improves barley photosynthetic efficiency through a possible regulatory module, MYC2-RcaA, under combined drought and salinity stress [J]. Photosynthesis Research, 2024, 159(1): 69−78. https://doi.org/10.1007/s11120-023-01074-2 doi: 10.1007/s11120-023-01074-2 [28] Hou X, Singh S K, Werkman J R, et al. Partial desensitization of MYC2 transcription factor alters the interaction with jasmonate signaling components and affects specialized metabolism [J]. International Journal of Biological Macromolecules, 2023, 252: 126472. https://doi.org/10.1016/j.ijbiomac.2023.126472 doi: 10.1016/j.ijbiomac.2023.126472 [29] Yuan H, Liu B, Zhang G, et al. Genome-wide identification and expression analysis of the PsTPS gene family in Pisum sativum [J]. Horticulturae, 2024, 10(10): 1104. https://doi.org/10.3390/horticulturae10101104 doi: 10.3390/horticulturae10101104 [30] Cui J, Jiang X, Li Y, et al. Genome-wide identification, phylogenetic, and expression analysis of jasmonate ZIM-domain gene family in Medicago sativa L. [J]. International Journal of Molecular Sciences, 2024, 25(19): 10589. https://doi.org/10.3390/ijms251910589 doi: 10.3390/ijms251910589 [31] Du Q L, Fang Y P, Jiang J M, et al. Genome-wide identification and characterization of the JAZ gene family and its expression patterns under various abiotic stresses in Sorghum bicolor [J]. Journal of Integrative Agriculture, 2022, 21(12): 3540−3555. https://doi.org/10.1016/j.jia.2022.08.104 doi: 10.1016/j.jia.2022.08.104 [32] Zhao G, Song Y, Wang Q, et al. Gossypium hirsutum salt tolerance is enhanced by overexpression of G. arboreum JAZ1 [J]. Frontiers in Bioengineering and Biotechnology, 2020, 8: 157. https://doi.org/10.3389/fbioe.2020.00157 doi: 10.3389/fbioe.2020.00157 [33] Ma J, Li C, Sun L, et al. The SlWRKY57-SlVQ21/SlVQ16 module regulates salt stress in tomato [J]. Journal of Integrative Plant Biology, 2023, 65(11): 2437−2455. https://doi.org/10.1111/jipb.13562 doi: 10.1111/jipb.13562 [34] Wu H, Ye H, Yao R, et al. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice [J]. Plant Science: an International Journal of Experimental Plant Biology, 2015, 232: 1−12. [35] Zhang W, Yuan S, Liu N, et al. Glutamine synthetase and glutamate synthase family perform diverse physiological functions in exogenous hormones and abiotic stress responses in Pyrus betulifolia bunge (P. be) [J]. Plants, 2024, 13(19): 2759. https://doi.org/10.3390/plants13192759 doi: 10.3390/plants13192759 [36] Hoshida H, Tanaka Y, Hibino T, et al. Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase [J]. Plant Molecular Biology, 2000, 43(1): 103−111. https://doi.org/10.1023/A:1006408712416 doi: 10.1023/A:1006408712416 [37] Cai H, Zhou Y, Xiao J, et al. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice [J]. Plant Cell Reports, 2009, 28(3): 527−537. https://doi.org/10.1007/s00299-008-0665-z doi: 10.1007/s00299-008-0665-z [38] Fortunato S, Nigro D, Lasorella C, et al. The role of glutamine synthetase (GS) and glutamate synthase (GOGAT) in the improvement of nitrogen use efficiency in cereals [J]. Biomolecules, 2023, 13(12): 1771. https://doi.org/10.3390/biom13121771 doi: 10.3390/biom13121771 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1159
- HTML全文浏览量: 1224
- PDF下载量: 59
- 被引次数: 0
