

-
随着杀虫剂的大量使用,越来越多的害虫产生了相应的抗药性。抗药性的产生往往伴随着适合度代价(fitness cost)的出现。适合度代价是指在生物进化过程中,某种生物特性或行为所带来的对生存和繁殖能力的负面影响,也就是说,抗性种群为了适应杀虫剂的筛选压力可能从多种生理途径或在生态学特征上产生某些变化[1]。例如褐飞虱细胞色素 P450 基因的过量表达导致其产生抗药性,同时其对吡虫啉农药的代谢过程使昆虫体内活性氧(ROS)大量产生,产生适合度代价[2]。明确不同杀虫剂对害虫产生的抗性代价适合度差异,可以为杀虫剂的轮换使用提供科学依据。例如,某些杀虫剂可能会使害虫产生较高的抗性代价,在使用一段时间后,害虫种群的适合度下降,此时可以轮换使用另一种对害虫抗性代价影响较小的杀虫剂,以延缓害虫抗药性的产生,提高防治效果。白纹伊蚊是中国常见的蚊种之一,调查发现,由于长期使用多类杀虫剂,白纹伊蚊已对上述杀虫剂产生了不同程度的抗药性,并且白纹伊蚊幼虫产生的抗药性比成虫更加严重[3]。有研究表明,抗性伊蚊产生的适合度代价主要表现在发育时间延长、寿命缩短和雌性繁殖力下降等方面,从而导致抗性伊蚊种群的生存力降低[4 − 5]。目前已有很多有关拟除虫菊酯类杀虫剂产生抗药性的媒介蚊虫的研究,但对四氟甲醚菊酯产生抗药性的白纹伊蚊所伴随的适合度代价当前尚不清晰。本研究通过种群生命表比较法分析白纹伊蚊对四氟甲醚菊酯产生抗药性的适合度代价,明确了四氟甲醚菊酯抗性的发生及发展对白纹伊蚊种群的发育和生存的影响,并以此为依据合理指导抗性白纹伊蚊治理与防控。
-
敏感品系白纹伊蚊(SS)由广东省中山市疾病预防控制中心提供。抗药品系白纹伊蚊(RR)来自广东省中山市野外采集,在华南农业大学饲养,将幼虫暴露于四氟甲醚菊酯溶液中,筛选出具有抗性的白纹伊蚊(2014年6月至2024−09−30)。在此过程中,由人工饲养多代,浓度对其幼虫保持在半致死剂量,通过连续筛选获得稳定的遗传抗药种群(抗性倍数为27.02),其饲养环境保持在光照周期为白天∶夜晚=14 h∶10 h,温度为(27±1)℃,相对湿度为(75±5)%的室内环境。实验室采用标准化昆虫饲养[5]。幼虫饲养于22 cm×14 cm×6.3 cm的饲养盆中,加入盆容量3/5的去氯自来水,将小鼠粮切成小块,每日投喂定量小鼠粮,每2 d更换1次去氯自来水,保证水质干净清澈。幼虫化蛹后,使用一次性吸管将蛹吸出,置于装有去氯自来水的100 mL小烧杯中,将烧杯置于23 cm×23 cm×23 cm的养虫笼中,蚊蛹羽化为成蚊后,喂食10%葡萄糖水(将棉花芯浸渍在葡萄糖水中)。成蚊在羽化后的1~2 d后,每日16:00~18:00喂食小白鼠血供雌蚊产卵。
-
参考赵紫华[6]的生命表方法组建SS和RR品系白纹伊蚊的试验种群生命表,以羽化为发育期起始点,计算种群净增殖率(R0)、世代平均周期(T)、内禀增长率(rm)、周限增长率(λ)和种群加倍时间(D)等参数,计算公式如下:
$${R}_{ \mathrm{0}} \mathrm{=\sum l}_{ {x}} {m}_{ {x}}, $$ (1) $$ {T=\sum {\rm{l}}}_{ {x}}{m}_{ {x}} x\mathrm{/\sum l}_{ {x}} {m}_{ {x}}, $$ (2) $${r}_{ \mathrm{m}}{=(\ln R}_{ \mathrm{0}} {)/T}, $$ (3) $$ {\lambda ={\rm{e}}}^{ \mathrm{rm}}, $$ (4) $$ {D=\ln 2/r}_{ \mathrm{m}}, $$ (5) 式中,x为以d为单位的时间间隔,平均后代数mx是指成虫羽化后第x天,平均每头雌虫后代数[7]。
-
将两种品系卵块分别泡在去氯水中,孵化出幼虫后,用一次性吸管各取50只SS组和RR组幼虫,相同环境下采用标准化昆虫饲养。从一龄幼虫期开始每天记录,内容包括幼虫的数量,蛹的数量,幼虫的发育时长,蛹的发育时长。至所有幼虫羽化成成蚊,停止记录。
-
SS品系和RR品系的白纹伊蚊幼虫在相同的条件下饲养,羽化为成蚊后,用吸蚊器各取50头24 h内羽化出的成蚊,放入养虫笼中观察。养虫笼中每天添加干净的10%葡萄糖水,并记录两种品系死亡的成蚊数量,直至所有成蚊死亡。
-
用Excel记录数据,SPSS 19.0分析数据,GraphPad Prism 9绘制图表。一龄期幼虫发育至蛹的平均时间、蛹羽化的平均时间、一龄期幼虫发育至羽化的平均时间用t检验进行判定,p<0.05被认为有统计学差异;幼虫化蛹率和羽化率之间的差异用t检验进行判定,p<0.05被认为有统计学差异;成蚊生存时间之间的差异采用Kaplan-Meier生存分析和log-rank检验进行判定,生存时间中位数之间的差异用Mann-Whitney U检验判定,p<0.05被认为有统计学差异[8]。
-
敏感品系种群净增长率(R0)、世代平均周期(T)、内禀增长率(rm)、周限增长率(λ)、种群加倍时间(D)分别为41.13、11.77 d、0.32 d−1、1.38 d−1、2.17 d;抗性种群分别为26.48、14.27 d、0.23 d−1、1.26 d−1、3.01 d(表1)。敏感品系白纹伊蚊较抗性品系的种群加倍时间短,表明其繁殖速度较快。
表 1 敏感品系白纹伊蚊和抗性品系白纹伊蚊种群数量增长参数计算结果
Table 1. Population growth of the susceptible and resistant lines of Aedes albopictus
种类
Line种群净增殖率
Net reproduction rate世代平均周期/d
Mean generation time内禀增长率/d−1
Intrinsic rate of increase周限增长率/d−1
Finite rate of increase种群加倍时间/d
Population doubling timeSS 41.13 11.77 0.32 1.38 2.17 RR 26.48 14.27 0.23 1.26 3.01 -
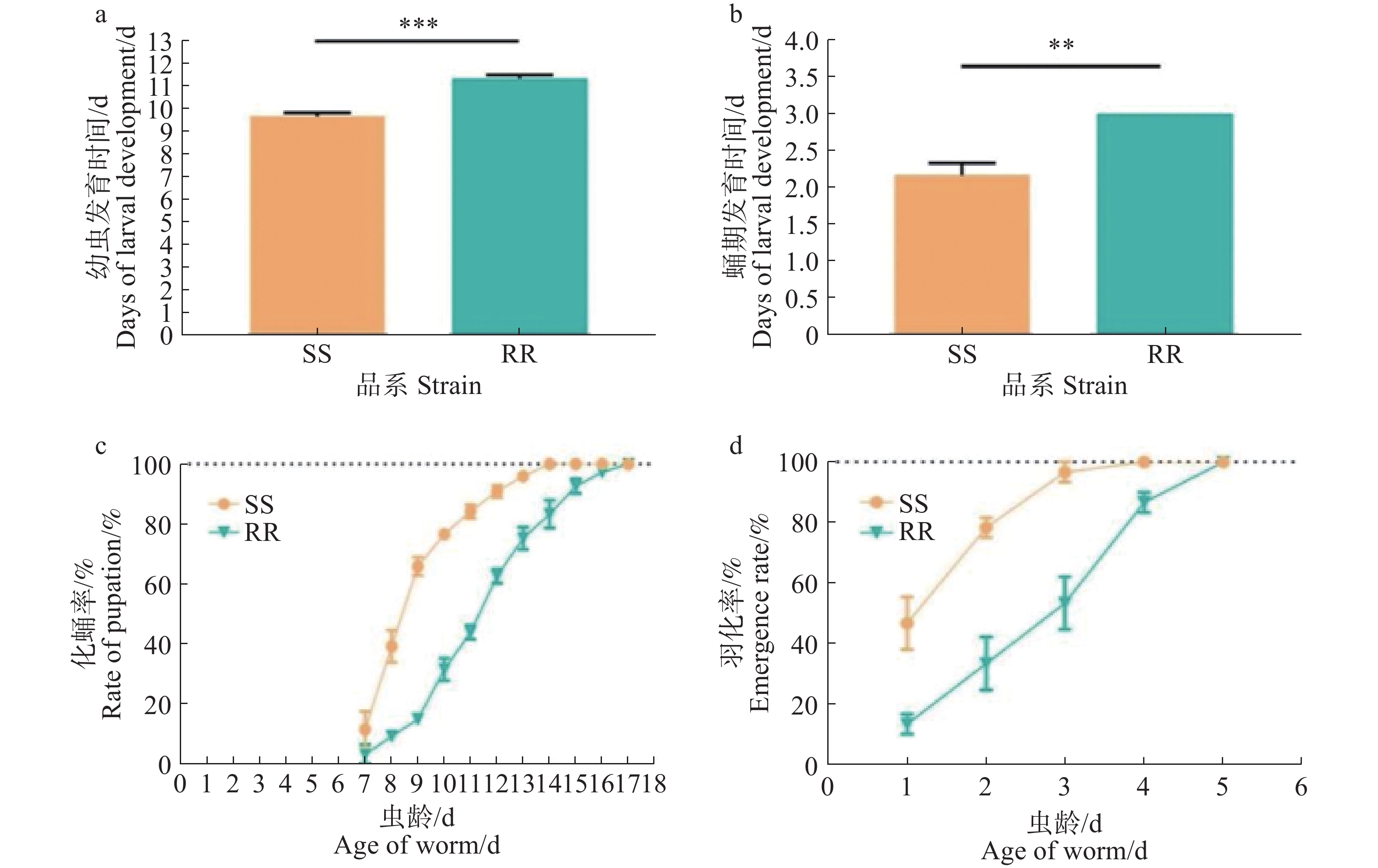
SS组和RR组的化蛹率和羽化率均为100%,两个品系的化蛹率和羽化率均无显著差异(p>0.05)。SS和RR从一龄幼虫发育至蛹的平均时间分别(9.66±0.17)和(11.31±0.17)d,蛹的平均羽化时间分别为(2.15±0.04)和(2.90±0.03)d(图1),结果显示,RR组一龄幼虫发育至蛹所需时间比SS组长,差异有统计学意义(p<0.005),且蛹发育至成蚊所需时间也具有统计学差异(p<0.005)。总的来说,RR组从一龄幼虫发育至成蚊的时间比SS组长,分别为(14.27±0.14 )和(11.77±0.16)d,说明四氟甲醚菊酯抗性的产生对白纹伊蚊的发育产生影响,并延长了白纹伊蚊幼虫发育至成蚊的时间。
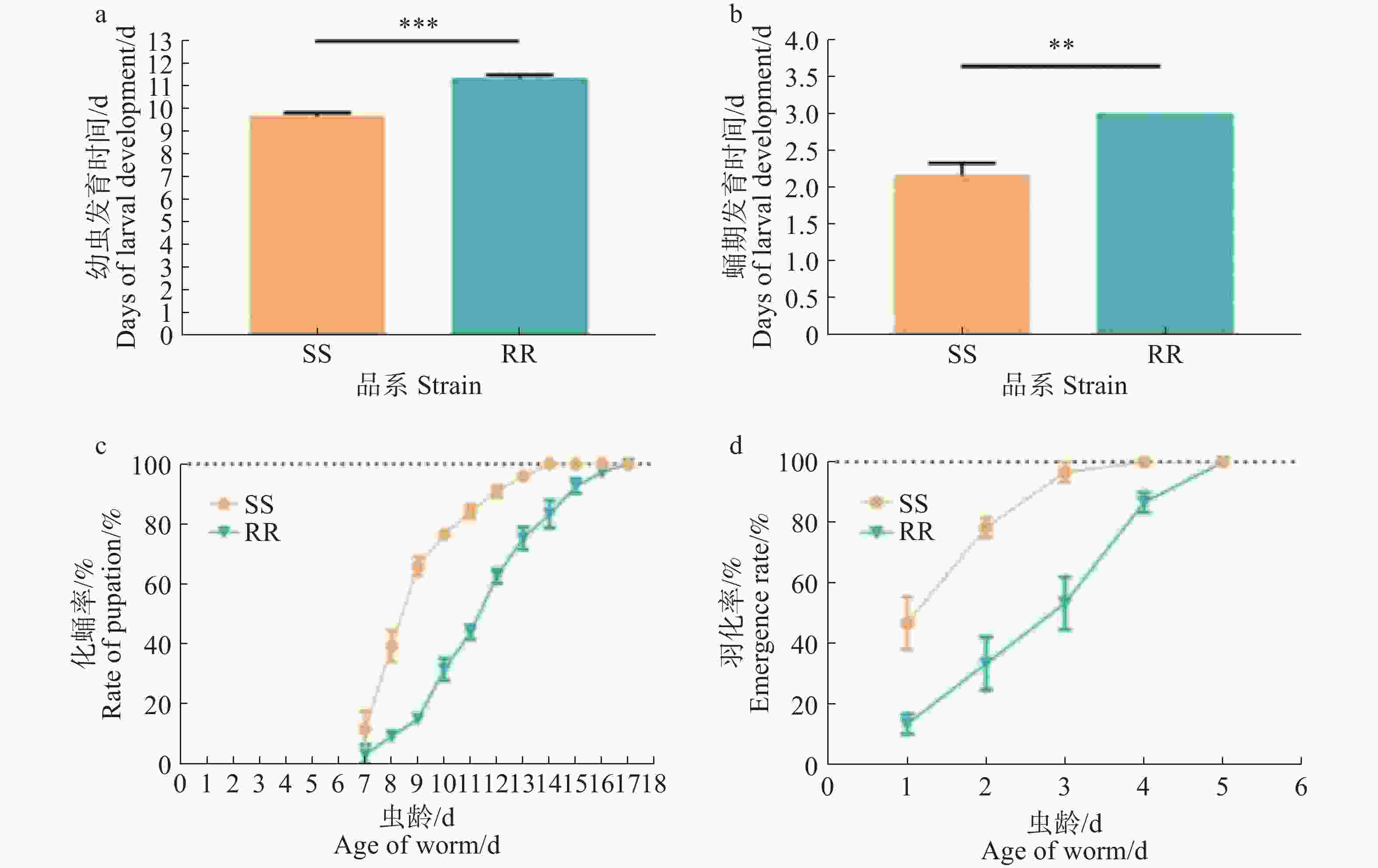
图 1 白纹伊蚊敏感品系和抗性品系一龄幼虫发育至成蚊的生命量表分析
Figure 1. Life table analysis for the susceptible and resistant lines of Aedes albopictus from larvae to adults
-
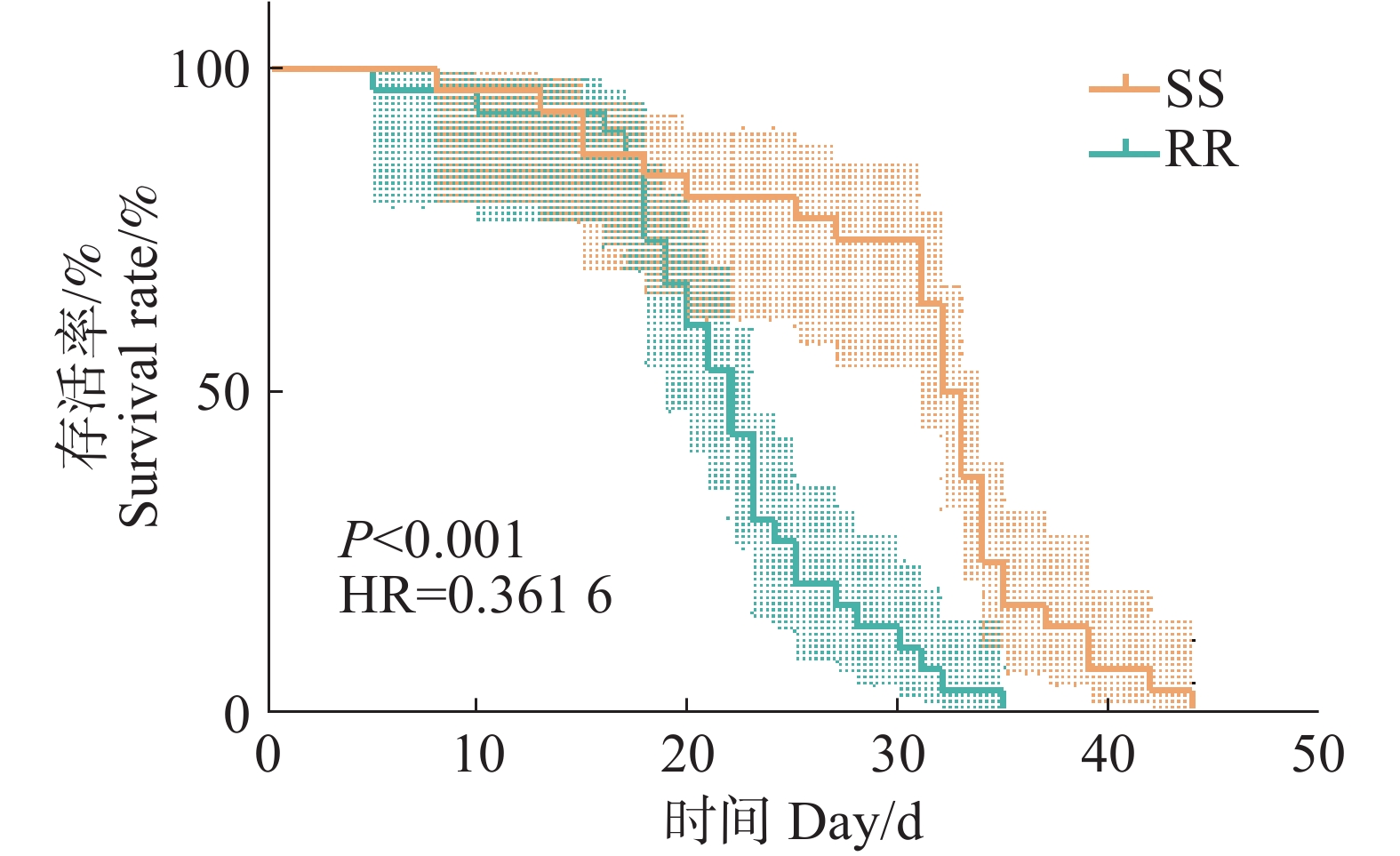
从SS组和RR组雌蚊的生存曲线来看,敏感品系和抗性品系雌蚊的平均生存时间分别为(30.03±1.60)和(21.83±1.12) d,生存时间中位数分别为(32.50±0.69)和(22.00±1.09) d(表2、3,图2),有统计学差异(c2=21.954, df=1, p<0.001, Mann-Whitney U test p<0.001)。结合成蚊的生存曲线,说明四氟甲醚菊酯抗性的产生对白纹伊蚊雌性成蚊的寿命产生了影响,缩短了雌性成蚊的寿命。
表 2 白纹伊蚊敏感品系和抗性品系幼虫发育至成蚊的生命量表
Table 2. Life table for susceptible and resistant lines of Aedes albopictus from larvae to adults
评价指标Evaluation index SS RR 显著性Significance 化蛹率/%
Rate of pupation/%100.00 100.00 ns 羽化率/%
Emergence rate/%100.00 100.00 ns 幼虫发育至蛹的时间/d
Days of larval development to pupaepa/d9.66±0.17 11.31±0.17 *** 蛹发育至成蚊的时间/d
Days of pupal development to adult mosquitoes/d2.15±0.04 2.90±0.03 ** 幼虫发育至成蚊的时间/d
Days of larval development to adult mosquito/d11.77±0.16 14.27±0.14 *** 注:用t检验进行判定,结果用平均值±标准误(SE)表示;ns为 p>0.05;**为 p<0.01;***为 p<0.005。
Note: A t-test is used for assessment, and the results are expressed as mean±standard error(SE); ns is p>0.05; ** is p<0.01; *** is p<0.005.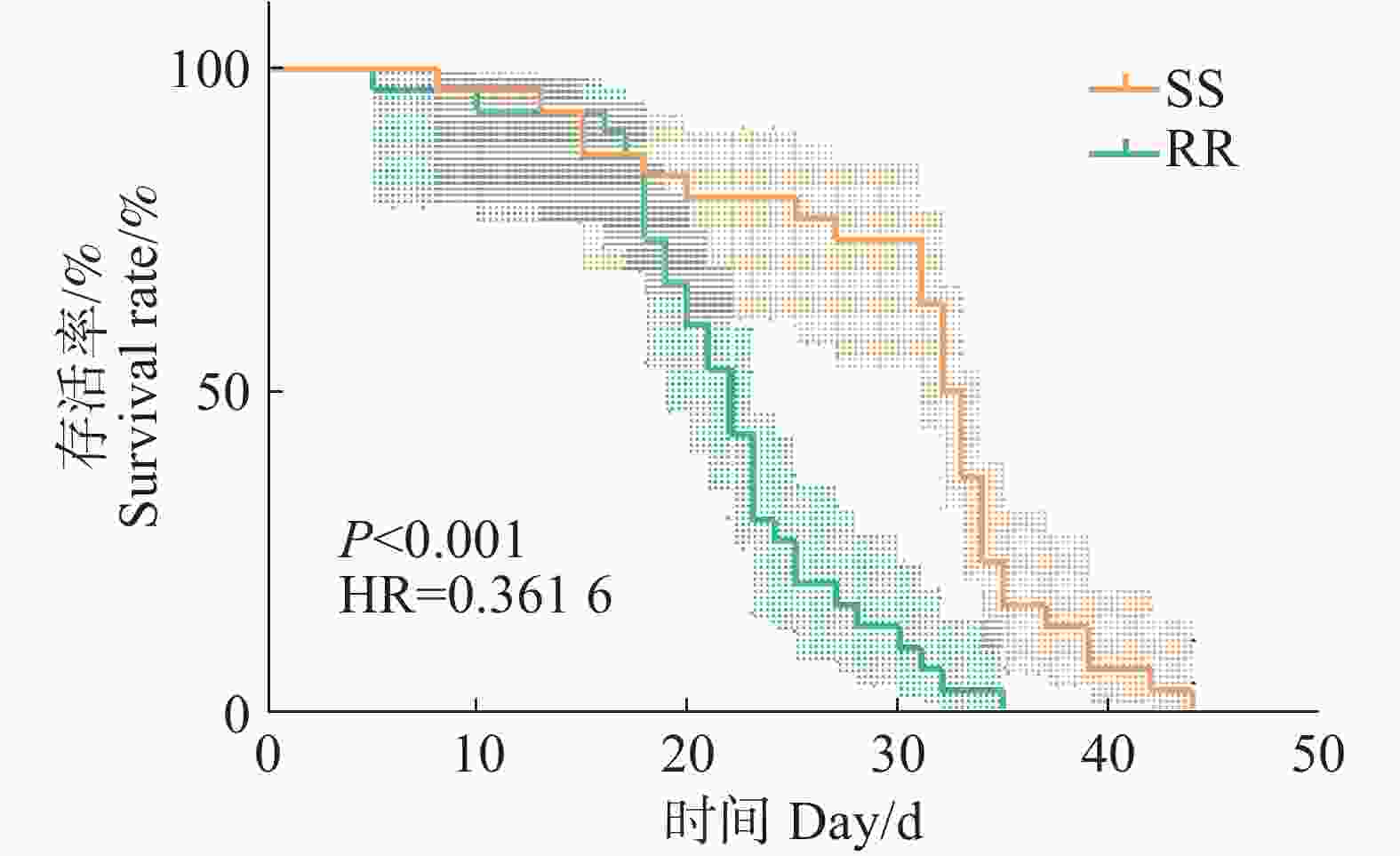
图 2 白纹伊蚊敏感品系和抗性品系成蚊Kaplan-Meier生存曲线
Figure 2. Kaplan-Meier survival curves of susceptible and resistant strains of adult Aedes albopictus
表 3 白纹伊蚊敏感品系和抗性品系成蚊寿命表
Table 3. Life table of susceptible and resistant lines of Aedes albopictus
分组
Group性别
SexSS RR 显著性
Significance平均生存
时间/d
Mean days
of survival/d♀ 30.03±1.60 21.83±1.12 *** 生存时间
中位数/d
Median number
of days alive/d♀ 32.50±0.69 22.00±1.09 *** 注:采用Kaplan-Meier生存分析和log-rank检验,生存时间中位数之间的差异用Mann-Whitney U检验判定,结果用平均值±标准误(SE)表示;***为 p<0.005。
Note: Kaplan-Meier analysis and log-rank test are used, and Mann-Whitney U test is used to compare the difference between the median numbers of the survival time of the two lines. The results are expressed as mean ± standard error(SE); *** is p<0.005. -
长期、广泛使用杀虫剂会导致白纹伊蚊产生相应的抗药性,同时也往往伴随着适合度代价的出现[9]。昆虫产生的适合度代价通常与它们对某种环境压力或杀虫剂的抗性发展相关[10 − 11]。这种代价可能体现在昆虫滞育、寿命缩短、雌性或者雄性的生殖力的下降等,即昆虫在受到外界某种压力时,其生存和繁殖能力受到一定程度的影响[12]。昆虫产生适合度代价的现象和背后的机制是非常复杂的,不同品种的昆虫对不同杀虫剂产生抗性后所表现出的适合度代价也各不相同。例如小菜蛾对阿维菌素和多杀菌素产生抗性后,分别表现出繁殖力降低和耐低温能力下降等适合度代价[13]。烟粉虱对新烟碱类药剂产生抗性后,出现卵巢发育异常的生殖代价[14]。同时也存在现象相反情况,例如褐飞虱对吡虫啉产生抗性后,表现出了适合度下降的趋势[15]。不同的适合度代价意味着需要采取不同的治理策略来应对昆虫抗药性问题。目前已有很多关于对拟除虫菊酯类杀虫剂产生抗药性的媒介蚊虫的研究,而四氟甲醚菊酯作为一种常用的拟除虫菊酯类杀虫剂,对其产生抗药性的白纹伊蚊所伴随的适合度代价还尚不清晰。了解白纹伊蚊对四氟甲醚菊酯产生抗药性机理和伴随的适合度代价对今后抗药性蚊虫治理具有非常重要的意义。
有研究表明,溴氰菊酯抗性品系白纹伊蚊发育时间明显延长,成蚊的平均寿命明显缩短[16],证明了白纹伊蚊溴氰菊酯抗性的产生增加了其适合度代价。而白纹伊蚊对四氟甲醚菊酯产生抗药性所伴随的适合度代价尚不明确,本研究将实验室筛选的白纹伊蚊四氟甲醚菊酯抗性品系与敏感品系进行对比,结果显示,抗性品系从幼虫发育至成蚊的时间比敏感品系长,但二者的化蛹率和羽化率无显著差异,说明抗性品系明显滞育,但并不会导致它们在生长期的死亡。结合成蚊的生存曲线来看,雌蚊敏感品系平均寿命要显著长于抗性品系,因此四氟甲醚菊酯抗性的产生缩短了白纹伊蚊成蚊的寿命,导致成蚊过早死亡,表明抗药性种群产生了生长发育和生存的适合度代价,这与溴氰菊酯抗性品系白纹伊蚊适合度代价的报道结果相一致。此外本研究还通过分析白纹伊蚊四氟甲醚菊酯抗性品系的种群数量增长,发现抗性品系白纹伊蚊较敏感品系的种群加倍时间长,证明了抗性种群数量增长比敏感种群慢,表明抗药性种群产生了生殖适合度代价。本研究团队推测可能是靶标基因位点的改变导致耗能增加,如昆虫的抗药性是由于靶标基因位点敏感性降低引起的,那么昆虫可能会投入额外的能量来维持这种改变后的靶标状态。但是昆虫的生长同样需要大量能量维持细胞分裂、组织构建和器官发育等生理过程。充足的能量供应能促使昆虫快速生长,使其体型增大、体质量增加,而当昆虫调动过多能量投入到维持改变后的靶标状态,就会导致能量匮乏,出现昆虫生长缓慢,生长停滞,发育不良甚至生殖障碍的情况。所以靶标位点改变耗能对昆虫的生存和繁殖会产生多方面的影响。长期使用传统杀虫剂导致许多昆虫产生了抗药性,使得杀虫效果逐渐降低,而通过作用于新的靶标位点使白纹伊蚊体内能量失衡,以此研发新型杀虫剂,能够为解决日益严重的昆虫抗药性问题提供新的思路和方法。
-
本研究通过种群生命表比较法分析了抗药性白纹伊蚊适合度代价,发现抗性白纹伊蚊幼虫期和蛹期发育时间延长、成蚊雌蚊寿命缩短和繁殖力下降,表明白纹伊蚊对四氟甲醚菊酯产生抗性后存在一定适合度代价。适合度代价的研究对于探讨抗性发展规律以及完善抗性治理策略具有重要意义。通过了解白纹伊蚊种群生命适合度代价的产生机制和影响因素,本研究团队可以更有效地预测和管理害虫的抗性进化,从而延长杀虫剂的使用寿命,减少化学农药的使用量,保护生态环境和人类健康。
白纹伊蚊对四氟甲醚菊酯抗性适合度代价分析
DOI: 10.15886/j.cnki.rdswxb.20240152
 CSTR: 32425.14.j.cnki.rdswxb.20240152
CSTR: 32425.14.j.cnki.rdswxb.20240152
Fitness cost analysis of resistance to dimefluthrin in Aedes albopictus
-
摘要: 以室内筛选的白纹伊蚊抗性品系RR为研究对象,敏感品系SS为对照,采用种群生命表比较法,研究白纹伊蚊对四氟甲醚菊酯产生抗性后的适合度代价变化。结果表明,净增长率(R0)、世代平均周期(T)、内禀增长率(rm)、周限增长率(λ)、种群加倍时间(D)在敏感品系种群中分别为41.13、11.77 d、0.32 d−1、1.38 d−1、2.17 d;在抗性种群中分别为26.48、14.27 d、0.23 d−1、1.26 d−1、3.01 d。抗性品系白纹伊蚊较敏感品系的种群加倍时间长,表明其繁殖速度减慢。敏感品系和抗性品系的化蛹率和羽化率均为100%,两者无显著差异;一龄幼虫发育至蛹的平均时间分别为(9.66±0.17)和(11.31±0.17)d;蛹的平均羽化时间分别为(2.15±0.04)和(2.90±0.03)d;雌蚊的平均生存时间分别为(30.03±1.60)和(21.83±1.12)d,均存在显著差异。抗性白纹伊蚊幼虫期和蛹期发育时间延长,成蚊寿命缩短,表明白纹伊蚊对四氟甲醚菊酯产生抗性后存在一定适合度代价。Abstract: The fitness cost of resistance to dimefluthrin in Aedes albopictus was analyzed by population life table comparison method using the resistant line (RR) of Ae. albopictus for experiment with the susceptible line SS as control. The results showed that the net reproduction rate (R0), mean generation time (T), intrinsic rate of increase (rm), finite rate of increase (λ), and population doubling time (D) were 41.13, 11.77 d, 0.32 d−1, 1.38 d−1, and 2.17 d, respectively, for the susceptible line population, and. 26.48, 14.27 d, 0.23 d−1, 1.26 d−1, and 3.01 d, respectively, for the resistant line. The resistant line of Ae. albopictus was longer in population doubling time than the susceptible line, indicating that its reproductive rate has slowed down. The pupation rate and emergence rate of the susceptible and resistant lines were all 100%, indicating there was no significant difference between them. The average development time from first instar larvae to pupae was (9.66±0.17)d for the susceptible line and (11.31±0.17)d for the resistant line; the average emergence time of pupae for both of the lines was (2.15±0.04) and (2.90±0.03)d, respectively; the average survival time of female mosquitoes was (30.03±1.60) and (21.83±1.12)d, respectively, for both of the lines, and there were all significant differences. The development time of larval and pupal stages and the life span of adult mosquitoes were prolonged, indicating that there is a certain fitness cost of resistance to dimefluthrin in Ae. albopictus.
-
Key words:
- Aedes albopictus /
- dimefluthrin /
- pesticide-resistant /
- fitness cost
-
图 2 白纹伊蚊敏感品系和抗性品系成蚊Kaplan-Meier生存曲线
Fig. 2 Kaplan-Meier survival curves of susceptible and resistant strains of adult Aedes albopictus
表 1 敏感品系白纹伊蚊和抗性品系白纹伊蚊种群数量增长参数计算结果
Table 1 Population growth of the susceptible and resistant lines of Aedes albopictus
种类
Line种群净增殖率
Net reproduction rate世代平均周期/d
Mean generation time内禀增长率/d−1
Intrinsic rate of increase周限增长率/d−1
Finite rate of increase种群加倍时间/d
Population doubling timeSS 41.13 11.77 0.32 1.38 2.17 RR 26.48 14.27 0.23 1.26 3.01  下载: 导出CSV
下载: 导出CSV
表 2 白纹伊蚊敏感品系和抗性品系幼虫发育至成蚊的生命量表
Table 2 Life table for susceptible and resistant lines of Aedes albopictus from larvae to adults
评价指标Evaluation index SS RR 显著性Significance 化蛹率/%
Rate of pupation/%100.00 100.00 ns 羽化率/%
Emergence rate/%100.00 100.00 ns 幼虫发育至蛹的时间/d
Days of larval development to pupaepa/d9.66±0.17 11.31±0.17 *** 蛹发育至成蚊的时间/d
Days of pupal development to adult mosquitoes/d2.15±0.04 2.90±0.03 ** 幼虫发育至成蚊的时间/d
Days of larval development to adult mosquito/d11.77±0.16 14.27±0.14 *** 注:用t检验进行判定,结果用平均值±标准误(SE)表示;ns为 p>0.05;**为 p<0.01;***为 p<0.005。
Note: A t-test is used for assessment, and the results are expressed as mean±standard error(SE); ns is p>0.05; ** is p<0.01; *** is p<0.005.
下载: 导出CSV
表 3 白纹伊蚊敏感品系和抗性品系成蚊寿命表
Table 3 Life table of susceptible and resistant lines of Aedes albopictus
分组
Group性别
SexSS RR 显著性
Significance平均生存
时间/d
Mean days
of survival/d♀ 30.03±1.60 21.83±1.12 *** 生存时间
中位数/d
Median number
of days alive/d♀ 32.50±0.69 22.00±1.09 *** 注:采用Kaplan-Meier生存分析和log-rank检验,生存时间中位数之间的差异用Mann-Whitney U检验判定,结果用平均值±标准误(SE)表示;***为 p<0.005。
Note: Kaplan-Meier analysis and log-rank test are used, and Mann-Whitney U test is used to compare the difference between the median numbers of the survival time of the two lines. The results are expressed as mean ± standard error(SE); *** is p<0.005.
下载: 导出CSV
-
[1] 邓惠, 刘礼平, 蔡松武, 等. 广东省2007-2017年白纹伊蚊种群密度调查研究[J]. 中国媒介生物学及控制杂志, 2019, 30(1): 60 − 64. doi: 10.11853/j.issn.1003.8280.2019.01.013 [2] PANG R, XING K, YUAN L Y, et al. Peroxiredoxin alleviates the fitness costs of imidacloprid resistance in an insect pest of rice[J]. PLoS Biology, 2021, 19(4): e3001190. doi: 10.1371/journal.pbio.3001190 [3] SMITH L B, KASAI S, SCOTT J G. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: important mosquito vectors of human diseases[J]. Pesticide Biochemistry and Physiology, 2016, 133: 1 − 12. doi: 10.1016/j.pestbp.2016.03.005 [4] RIGBY L M, RAŠIĆ G, PEATEY C L, et al. Identifying the fitness costs of a pyrethroid-resistant genotype in the major arboviral vector Aedes aegypti[J]. Parasites & Vectors, 2020, 13(1): 358. doi: 10.1186/s13071-020-04238-4 [5] BOCK F, KUCH U, PFENNINGER M, et al. Standardized laboratory feeding of larval Aedes japonicus japonicus(Diptera: Culicidae)[J]. Journal of Insect Science, 2015, 15(1): 144. doi: 10.1093/jisesa/iev126 [6] 赵紫华. 虫口统计学的概念与应用[J]. 植物保护学报, 2020, 47(4): 904 − 911. doi: 10.13802/j.cnki.zwbhxb.2020.2020043 [7] 贾筱沄, 王祎丹, 赵紫华. 应用昆虫生命表定量评价外来实蝇入侵力[J]. 植物保护学报, 2023, 50(3): 839 − 840. doi: 10.13802/j.cnki.zwbhxb.2023.2021175 [8] 邓洁琳. 白纹伊蚊对溴氰菊酯的抗性对其适合度和媒介能力的影响[D]. 广州: 南方医科大学, 2021. doi: 10.27003/d.cnki.gojyu.2021.000666. [9] ANDERSON J B. Evolution of antifungal-drug resistance: mechanisms and pathogen fitness[J]. Nature Reviews Microbiology, 2005, 3(7): 547 − 556. doi: 10.1038/nrmicro1179 [10] GUL H, GADRATAGI B G, GÜNCAN A, et al. Fitness costs of resistance to insecticides in insects[J]. Frontiers in Physiology, 2023, 14: 1238111. doi: 10.3389/fphys.2023.1238111 [11] MBUJI A L, XUE Z X, GUO M C, et al. Resistance and fitness costs of the Helicoverpa armigera after selection with the tetraniliprole newly developed diamide insecticide[J]. Crop Protection, 2024, 179: 106622. doi: 10.1016/j.cropro.2024.106622 [12] SUN X, HUA W J, WANG K K, et al. A novel V263I mutation in the glutamate-gated chloride channel of Plutella xylostella (L.) confers a high level of resistance to abamectin[J]. International Journal of Biological Macromolecules, 2023, 230: 123389. doi: 10.1016/j.ijbiomac.2023.123389 [13] FU B L, LIANG J J, HU J Y, et al. GPCR-MAPK signaling pathways underpin fitness trade-offs in whitefly[J]. Proceedings of the National Academy of Sciences of the United States of America, 2024, 121(28): e2402407121. doi: 10.1073/pnas.2402407121 [14] 温玉丛, 范凯华, 师慈, 等. 褐飞虱对吡虫啉的田间抗性动态分析[J]. 南京农业大学学报, 2010, 33(4): 55 − 58. [15] CHEN L, ZHOU K M, SHI J, et al. Pyrethroid resistance status and co-occurrence of V1016G, F1534C and S989P mutations in the Aedes aegypti population from two dengue outbreak counties along the China-Myanmar border[J]. Parasites & Vectors, 2024, 17(1): 91. doi: 10.1186/s13071-024-06124-9 [16] JIANG H, MENG X K, ZHANG N, et al. The pleiotropic AMPK-CncC signaling pathway regulates the trade-off between detoxification and reproduction[J]. Proceedings of the National Academy of Sciences of the United States of America, 2023, 120(10): e2214038120. doi: 10.1073/PNAS.2214038120 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1049
- HTML全文浏览量: 641
- PDF下载量: 19
- 被引次数: 0





