

-
松材线虫(Pinewood Nematode Disease,PWND)病被称为松树的“癌症”,是由松材线虫(Bursaphelenchus xylophilus)引起的一种极具破坏性的森林病害,对世界的松林生态系统构成了严重威胁[1-2]。国内该病害自1982年在江苏省南京市首次发现以来,迅速蔓延至多个省份和地区,给林业生产带来了重大的经济损失和生态破坏[3]。广西壮族自治区位于中国的南部,是重要的木材产地之一,拥有丰富的森林资源,尤其是松林资源[4]。然而,近年来松材线虫病在广西地区的发生和蔓延已经成为一个不容忽视的问题[5]。该病害不仅严重威胁着广西的松林资源,还对当地的森林生态环境和经济发展造成了重大的影响[6]。
遗传多样性和遗传结构的研究对于理解松材线虫的起源、传播途径及种群动态至关重要[7]。通过对松材线虫种群的遗传多样性进行分析,可以有效地追踪病原体的来源和传播路径,评估其适应性和进化潜力[8]。高遗传多样性的种群通常具有更强的适应能力,能够更好地应对环境变化和抗药性发展,这对于预测松材线虫病害的发展趋势和制定有效的防控策略至关重要[9]。线粒体细胞色素氧化酶Ⅰ亚基基因片段(mitochondrial cytochrome c oxidase subunitⅠ,mt COⅠ)基因作为线粒体DNA中的一个高度保守而又变异丰富的区域,在种群遗传学研究中被广泛用作标记物[10]。通过分析mt COⅠ序列,可以揭示松材线虫种群间的遗传关系和地理分布特征,从而为制定有效的防控策略提供科学依据。
本研究基于松材线虫mt COⅠ基因,分析了广西壮族自治区8个区县松材线虫的遗传多样性与遗传结构,并将其与国内外其他地区的松材线虫mt COⅠ基因进行了系统发育分析,初步探讨了广西松材线虫遗传分化程度与扩散途径,为进一步研究松材线虫谱系地理学保证了重要的数据支撑,同时为制定广西松材线虫针对性防控策略提供了一定的理论依据。
-
本研究所用的67个松材线虫的虫株样本分别于2023年取自广西壮族自治区8个区县的松材线虫病疫区(表1)。疫木取样时分别取上、中、下部3~5 cm厚的圆盘,用密封袋保存带回实验室。
表 1 广西67个松材线虫虫株采集地点
Table 1. 67 collection sites for B. xylophilus in Guangxi
区/县
District/
county地区代码
Area code采样点
Sampling site虫株数量/个
Number of
strains/ind桂平市 GP 麻垌镇、大湾镇 4 临桂区 LG 临桂镇、五通镇 6 八步区 BB 桂岭镇、信都镇 7 城中区 CZ 静兰街道、河东街道 8 兴宁区 XN 三塘镇、五塘镇、昆仑镇 16 全州县 QZ 全州镇、庙头镇 6 苍梧县 CW 六堡镇、木双镇 7 容县 RX 六王镇、容州镇 13 -
利用贝尔曼漏斗法分离疫木中的松材线虫后,将线虫液置于15 mL EP管中进行形态学鉴定。挑取鉴定后的松材线虫雌雄成虫30~50条,接入长满灰葡萄孢菌(Botrytis cinerea)的PDA平板中,25 ℃下进行恒温培养[11]。待灰葡萄孢全部被吃完(10 d左右)后,再次使用贝尔曼漏斗法收集松材线虫于15 mL EP管中。后续将EP管3 500 r·min−1离心3 min;去掉上清液,等体积加入0.1%(V/V)硫酸链霉素,摇匀,静置5~10 min,离心,去上清;用无菌水反复洗涤3次后得到纯净的虫株。
-
使用MolPure Tissue DNA Kit组织DNA提取试剂盒(上海翌圣生物科技股份有限公司)提取虫株DNA,使用超微量分光光度计UL−5000(上海美析仪器有限公司)检测DNA质量与浓度,DNA质量与浓度合格后进行PCR扩增。采用引物COⅠ-F1(5′-CCTACTATGATT GGTGGTTTTGGTAATTG-3′)与COⅠ-V2(5′-GTAGC AGCAGTAAAATAAGCACG-3′)对样本DNA进行扩增[12]。反应体系为25 μL,包含2×Hieff® PCR Master Mix(上海翌圣生物科技股份有限公司)12.5 μL、DNA模板1 μL、浓度10 μmol·L−1的上游和下游引物各1 μL,加ddH2O补足至25 μL。PCR反应条件为预变性94 ℃,5 min;变性94 ℃,30 s;退火51 ℃,30 s;延伸72 ℃,1 min;循环35次;72 ℃终延伸10 min。产物经1%(V/V)琼脂糖凝胶电泳检测合格后送至上海生工生物工程股份有限公司进行Sanger双向测序。
-
使用Chromas观察序列峰图,BioEdit拼接序列[13]。将拼接后的mt COⅠ序列在NCBI(ncbi.nlm.nih.gov)中使用Nucleotide BLAST功能进行序列比对,将比对结果大于99.0%的物种认定为松材线虫,用于后续分析[14]。使用MEGA(v11)的Alignment中的ClustalW功能对所有序列进行比对和修整[15]。使用DNAsp(v6.0)进行遗传多样性和遗传分化分析,使用IBM SPSS进行相关性分析[16]。使用MEGA 11(v11)基于邻接法(Neighbor Joining,NJ)绘制系统发育树[17]。使用Popart绘制单倍型网络图[18]。
-
根据遗传多样性系数可知,广西松材线虫基于mt COⅠ基因的变异程度低,表现出低水平的遗传多样性(表2)。广西8个区县67个虫株的松材线虫在647 bp内共存在3个多态性位点,1个简约位点;共发现4种不同的松材线虫单倍型,单倍型多样性总体为0.170;整体核苷酸多样性为0,核苷酸差异系数为0.174。各地区间的遗传距离均小于0.001,表明广西的松材线虫不存在明显的变异(表3)。使用SPSS对各地区的虫株数量与单倍型数量进行线性分析,结果表明随着虫株数的增加,单倍型数与虫株数量不存在相关性(P>0.05)。
表 2 基于mt COⅠ基因的广西松材线虫遗传多样性系数表
Table 2. Genetic diversity coefficients of B. xylophilus in Guangxi based on mt COⅠ gene
地区代码
Area
code虫株数量/个
Number of
insect strains/ind多态性位点
Polymorphic
loci简约位点
A minimalist
site单倍型数
Number of
haplotypes单倍型多样性
Haplotype
diversity核苷酸多样性
Nucleotide
diversity平均核苷酸差异系数
Average nucleotide
difference coefficientGP 4 − − 1 − − − LG 6 1 − 2 0.333 0.001 0.333 BB 7 − − 1 − − − CZ 8 − − 1 − − − XN 16 − − 1 − − − QZ 6 1 0 2 0.333 0.001 0.333 CW 7 1 1 2 0.571 0.001 0.571 RX 13 − − 1 − − − All 67 3 1 4 0.170 0 0.174 注:“−”表示不存在,当多样性系数小于0.000 5时,以“0”表示。 Note:‘−’ indicates that it does not exist. When the diversity coefficient is less than 0.000 5,it is expressed as ‘0’. 表 3 基于mt COⅠ基因的广西松材线虫遗传距离
Table 3. Genetic distances of B. xylophilus in Guangxi based on mt COⅠ gene
地区代码
Area codeGP LG BB CZ NN QZ CW LG 0.000 3 BB − 0.000 3 CZ − 0.000 3 − NN − 0.000 3 − − QZ 0.000 3 0.000 5 0.000 3 0.000 3 0.000 3 CW 0.000 9 0.001 1 0.000 9 0.000 9 0.000 9 0.001 1 RX − 0.000 3 − − − 0.000 3 0.000 9 注:“−”表示不存在。 Note:‘−’ indicates that it does not exist. -
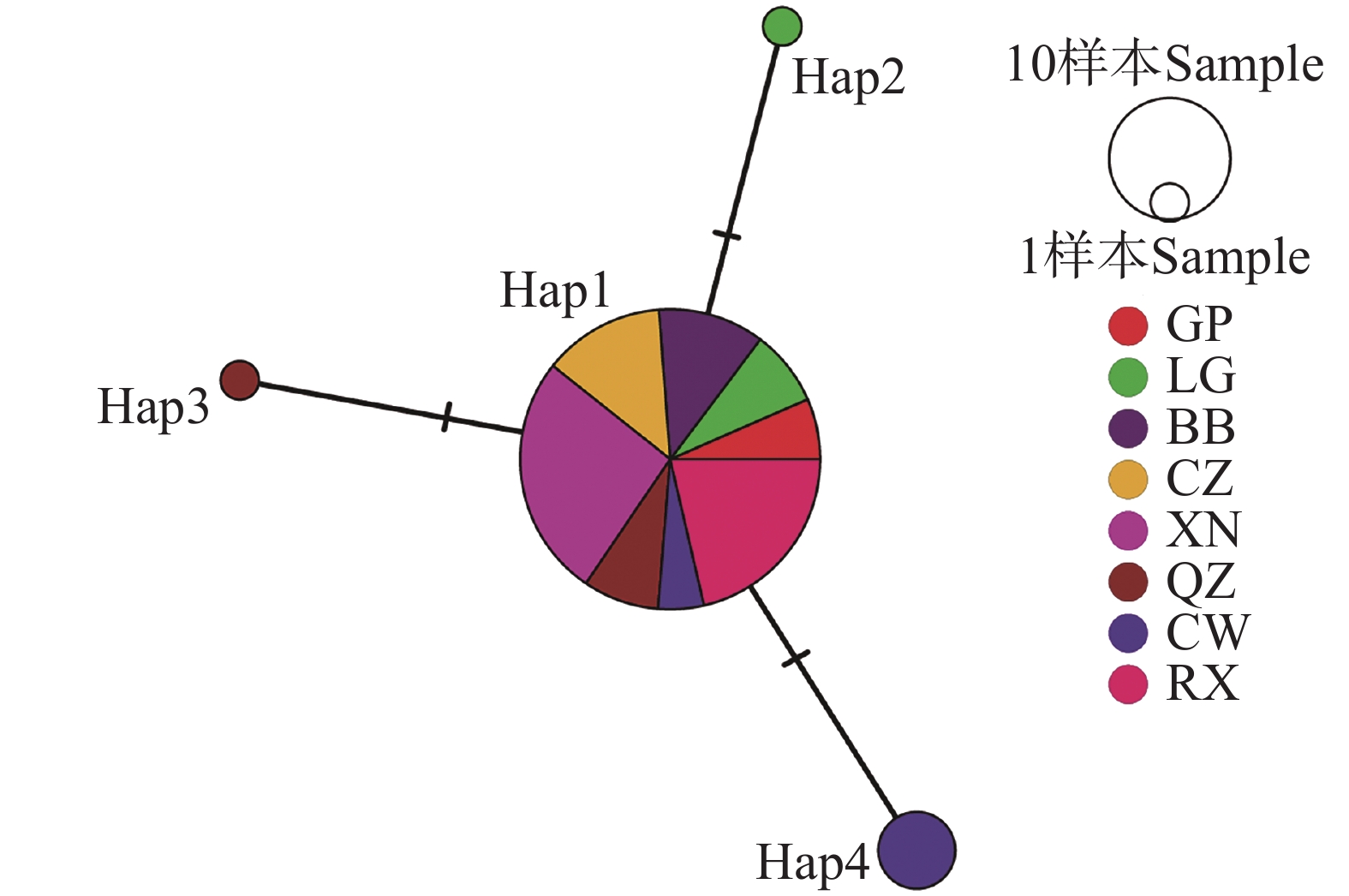
广西8个区县67个虫株的松材线虫共有4个单倍型(表4),其中Hap1出现的频率最高,占所有单倍型的91.0%。而Hap2、Hap3和Hap4出现的频率较低,Hap4占所有单倍型的6.0%,Hap2和Hap3均在本研究中出现1次。进一步地分析广西松材线虫单倍型网络图(图1),发现广西松材线虫以Hap1为中心节点,所有种群均包含了Hap1。同时,以Hap1分化出了3个进化分支,Hap2、Hap3和Hap4分别出现在临桂区、全州县和苍梧县内。
表 4 基于mt COⅠ基因的广西松材线虫单倍型在各个区县的分布
Table 4. Distribution of haplotypes of B. xylophilus based on mt COⅠ gene in various sampling sites in Guangxi
单倍型
Haplotype在各地区的分布 虫株数量/个
Number of strains/indGG GL HZ LZ NN QZ WZ YL Hap1 4 5 7 8 16 5 3 13 61 Hap2 − 1 − − − − − − 1 Hap3 − − − − − 1 − − 1 Hap4 − − − − − − 4 − 4 注:“−”表示不存在。 Note:‘−’ indicates that it does not exist. 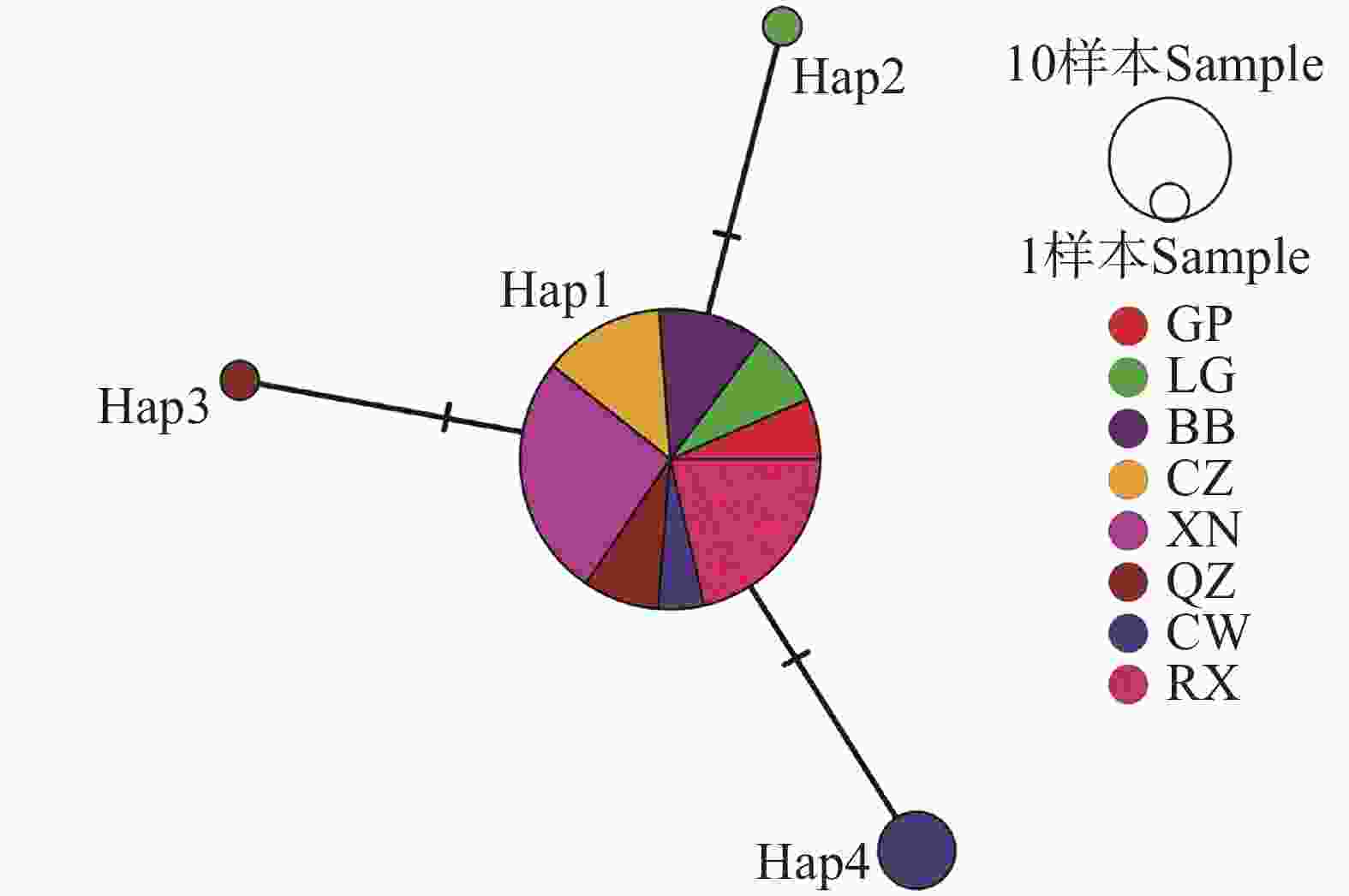
图 1 基于mt COⅠ基因的广西松材线虫单倍型网络图
Figure 1. Haplotype network of B. xylophilus in Guangxi based on mt COⅠ gene
-
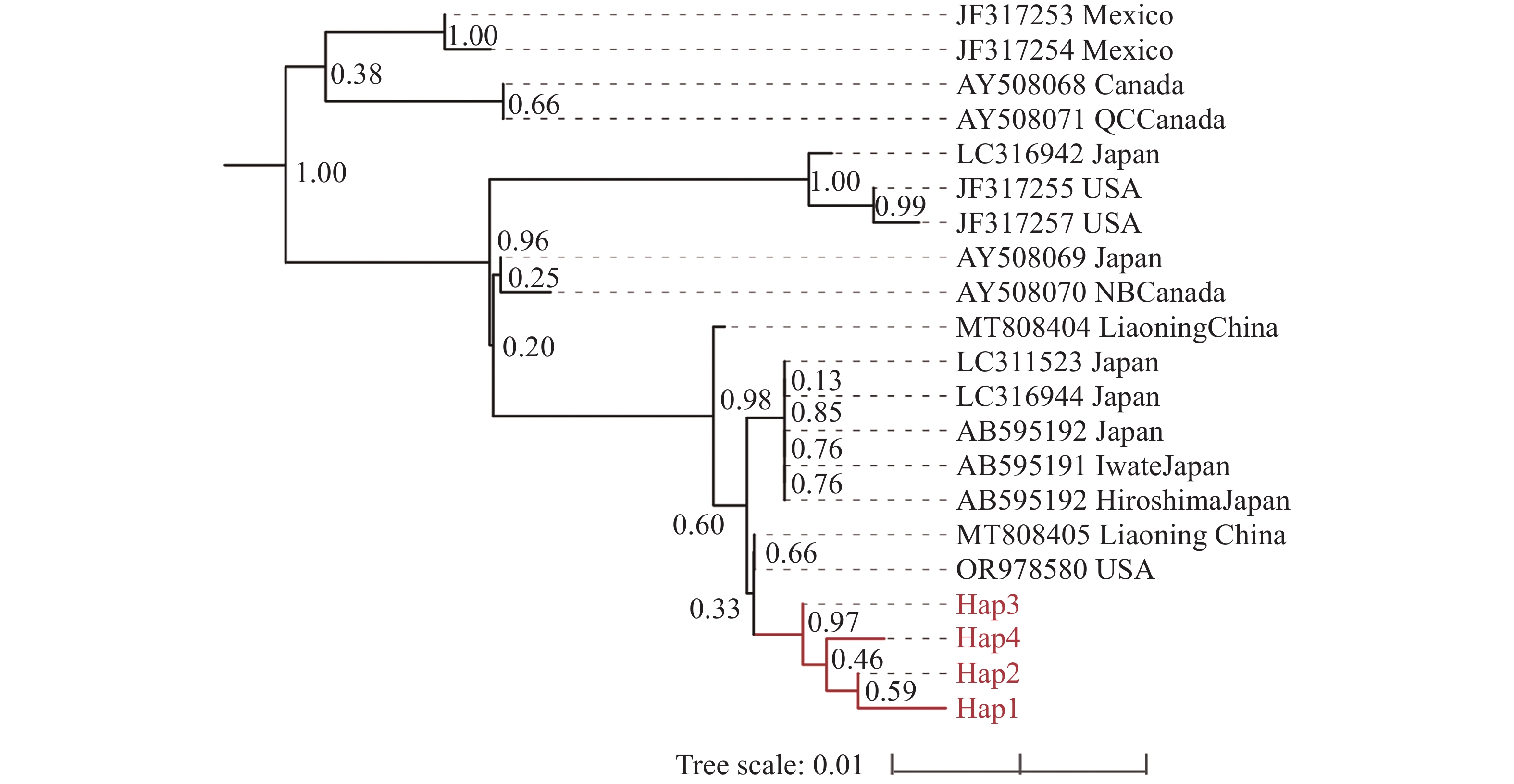
基于mt COⅠ基因构建的松材线虫系统发育树,21 条来自不同地区的松材线虫 mt COⅠ 基因分为两个大支(图2)。来自加拿大与墨西哥的松材线虫聚成一支;来自日本、加拿大、美国以及中国的松材线虫聚成一支,其中,中国广西的松材线虫独聚一支,与国内外其他地区的松材线虫明显区分开来。Hap3与其他3个单倍型具有明显区别(支持率 > 0.95)。
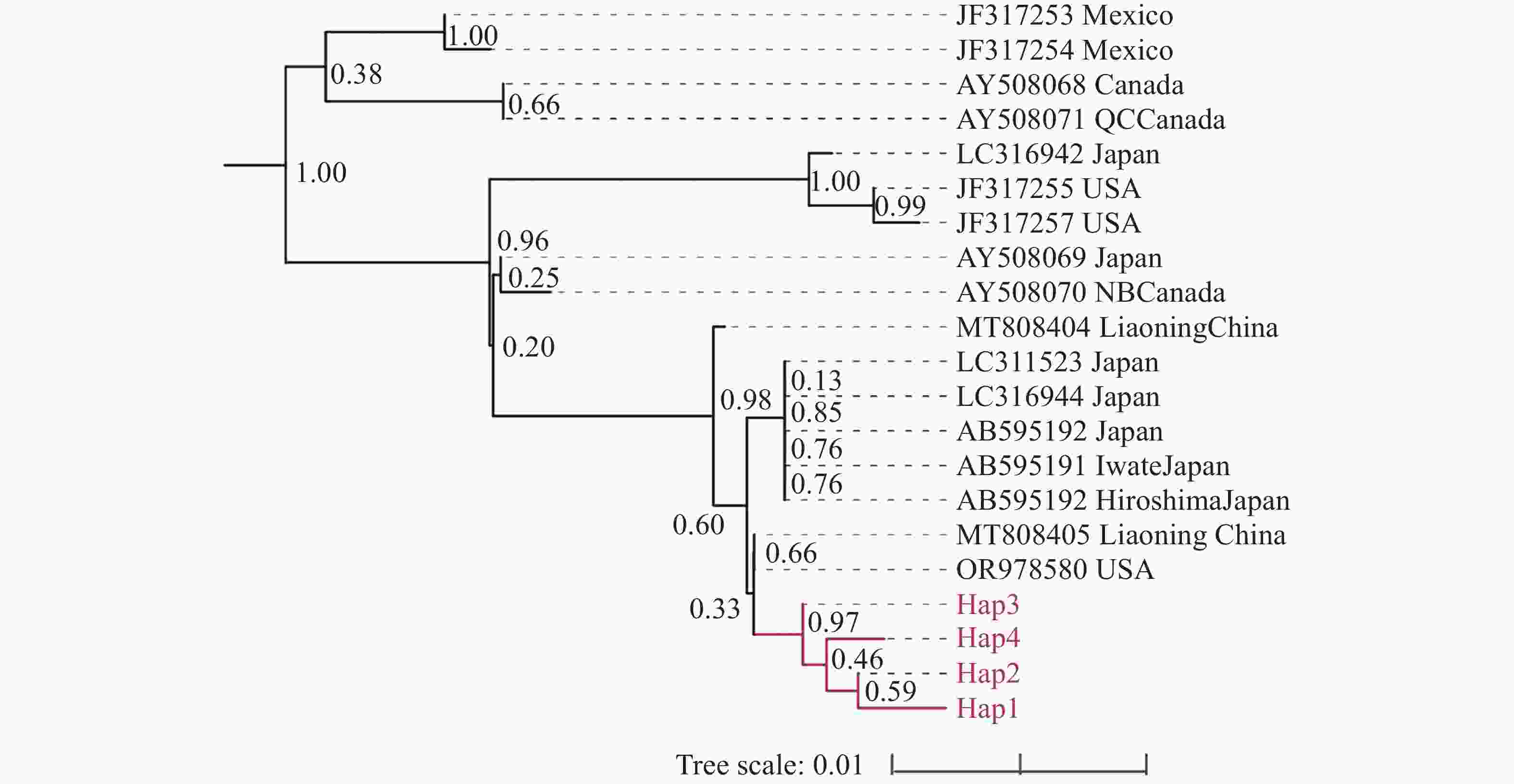
图 2 基于mt COⅠ基因的松材线虫系统发育树
Figure 2. Phylogenetic tree of B. xylophilus based on mt COⅠ gene
-
由于松材线虫病带来了巨大的生态危机和经济损失,关于松材线虫的研究在国内一直被高度重视。其中,关于松材线虫群体遗传的研究在1998年就有报道,在接下来的几年内出现了较多以RFLP和RAPD标记法来开展松材线虫种群遗传分化的研究[19]。Kanzaki等[20]于2002年采用了ITS标记和mt COⅠ标记对伞滑刃属的线虫进行了聚类分析,明确了mt COⅠ标记对伞滑刃属线虫的适用性。成飞雪等[21]于2005年开发了松材线虫AFLP标记体系,适用于大多数松材线虫种群的扩增。随着Illumina测序技术的开发,松材线虫种群遗传分化的研究方法得以更新。丁晓磊[22]通过Illumina全基因组重测序的方法,分析了来源不同的松材线虫毒力差异。虽然多数研究结果[23-26]表明mt COⅠ序列在种内表现出的遗传差异很低,但是依然有大量的研究人员将其用于种内遗传变异的相关研究。本研究基于前人结果,选取了较为简便的mt COⅠ对广西的松材线虫遗传多样性和遗传结构进行了分析,结果表明,广西地区来自8个区县67个松材线虫虫单倍型多样性和核苷酸多样性分别为0.170和0.174,未表现出明显的遗传多样性,仅存在着个别位点的变异。这说明广西的松材线虫遗传多样性较少,各个地区间的松材线虫基因交流频繁。
重大外来入侵物种特有的强大定殖能力和适应能力,会在其扩散地点产生始祖效应和遗传漂变,但此现象在松材线虫上并未有明显的表现[27-28]。由于松材线虫存在着较多不同的入侵来源,会存在多次入侵现象的发生,因此松材线虫会出现丰富的遗传多样性[29]。但本研究通过NJ法构建松材线虫mt COⅠ基因系统发育树,发现省域范围内的松材线虫遗传变异程度较小,汇聚成一个分支。与本研究相似,Watanabe和Matsunaga等也在日本不同地区发现了mt COⅠ基因单倍型一致的松材线虫(LC311523、LC316944、AB595191、AB595192),这进一步表明了松材线虫在一定范围内的遗传多样性较低,未发生明显的遗传分化现象。有趣的是,本研究得到的广西松材线虫mt COⅠ基因能够区分于国内外其他地区的松材线虫,特别是能够区分加拿大和墨西哥等北美地区的样本。多数研究认为北美洲是松材线虫的原产地,随着木材的流通扩散至世界各地[30]。而系统发育树结果表明,4个来自墨西哥和加拿大的松材线虫样本(JF317253、JF317254、AY508068、AY508071)与其他所有松材线虫样本存在着一定的遗传变异。这说明这4个松材线虫样本可能是源于北美的松材线虫本土种群,未发生远距离迁移,从而导致与其他地区的松材线虫存在着明显的遗传分化现象。以上结果均说明了使用mt COⅠ基因对松材线虫进行遗传标记可区分松材线虫的种内变异。随着中国“十三五”和“十四五”规划的有效推进,松材线虫病防控得以落实。积极政策的实施有效地抑制了松材线虫病的远距离传播,因此出现了奠基者效应导致当地的遗传多样性锐减。
广西松材线虫的主要传播媒介松墨天牛(Monochamus alternatus)在野外具有极强的自然扩散能力,成虫最大飞行距离可以达到3.2 km[31]。因此,松墨天牛的扩散会引起某一种群的松材线虫在一定范围内大规模扩散。基于mt COⅠ基因的单倍型网络图表明,单倍型Hap1在广西67个松材线虫虫株中出现的频率最大,占所有虫株的91%,即Hap1为广西松材线虫的主要单倍型。Hap3与Hap4分别存在于两省交界处的全州县和苍梧县,其中Hap4为苍梧县的主要单倍型,说明苍梧县的松材线虫存在着一定的遗传分化现象。由于松材线虫特有的远距离传播特点,其基因交流往往会无视地理隔离[32]。但是由于变异程度较小,与广西主要单倍型Hap1仅在一个多态性位点上存在差异,因此本研究排除苍梧县存在着较为严重的远距传播现象。因此可推测Hap3与Hap4两种单倍型的松材线虫可能在两省交界处随着自然寄主的繁衍而出现基因交流的现象。后续本研究可以参考丁晓磊[22]的Illumina全基因组重测序技术,对广西4种松材线虫单倍型进行深入分析,进而得出单倍型之间是否存在明显的遗传分化现象。
-
本研究基于mt COⅠ序列,初步探讨了广西松材线虫的遗传多样性和遗传分化。结果表明广西松材线虫整体遗传多样性较低,并未出现明显的遗传分化现象。通过与国内外其他地区的松材线虫的mt COⅠ序列构建系统发育树,发现广西松材线虫与其他地区明显区分,独成一支。这表明了松材线虫在近年来在广西主要以自然扩散的形式进行传播,未发现明显的外来种群定殖的现象,也侧面反映了广西松材线虫病防控工作和检疫措施的有效性。
基于mt COⅠ序列的广西松材线虫遗传多样性和遗传分化的初探
DOI: 10.15886/j.cnki.rdswxb.20240135
 CSTR: 32425.14.j.cnki.rdswxb.20240135
CSTR: 32425.14.j.cnki.rdswxb.20240135
Genetic diversity and genetic differentiation of Bursaphelenchus xylophilus in Guangxi based on mt COⅠ sequences
-
摘要: 松材线虫病是一种由松材线虫(Bursaphelenchus xylophilus)介导引起的森林病害,严重威胁着广西壮族自治区的森林生态环境和经济发展。本研究通过线粒体细胞色素氧化酶Ⅰ亚基基因片段(mt COⅠ)分析了来自广西壮族自治区8个不同区县的67个松材线虫虫株,初步探究了松材线虫在广西地区的遗传多样性和遗传分化程度。结果表明,广西松材线虫在647 bp内共存在3个多态性位点,1个简约信息位点,遗传多样性系数处于极低水平。广西松材线虫共存在4种单倍型,单倍型多样性总体为0.170,其中Hap1为广西松材线虫的主要单倍型。广西松材线虫明显区分于国内外地区的松材线虫,形成独立的分支。松材线虫近年来在广西地区主要以自然扩散的形式进行传播。本研究结果为制定广西松材线虫针对性防控策略提供了一定的理论依据。Abstract: An attempt was made to investigate the genetic diversity and genetic differentiation of Bursaphelenchus xylophilus, the causative agent of pine wilt disease, which is posing a severe threat to the forest ecosystem and economic development in Guangxi Zhuang Autonomous Region. Sixty seven strains of B. xylophilus collected from eight different districts/counties within Guangxi were analyzed by using the mitochondrial cytochrome c oxidase subunit Ⅰ (mt COⅠ) gene fragment. The results indicated that there were three polymorphic sites and one parsimony-informative site within a 647 bp region, with genetic diversity indices at extremely low levels. Four haplotypes of B. xylophilus were identified in Guangxi, with an overall haplotype diversity of 0.170, where Hap1 was the predominant haplotype. The B. xylophilus strains in Guangxi were clearly distinct from those found in other regions both domestically and internationally, forming an independent branch. In recent years, the spread of B. xylophilus in Guangxi occurred primarily through natural dispersion. All these findings might provide a theoretical basis for developing targeted prevention and control strategies for B. xylophilus in Guangxi.
-
Key words:
- Bursaphelenchus xylophilus /
- COⅠ gene /
- genetic diversity /
- genetic differentiation
-
图 2 基于mt COⅠ基因的松材线虫系统发育树
Fig. 2 Phylogenetic tree of B. xylophilus based on mt COⅠ gene
表 1 广西67个松材线虫虫株采集地点
Table 1 67 collection sites for B. xylophilus in Guangxi
区/县
District/
county地区代码
Area code采样点
Sampling site虫株数量/个
Number of
strains/ind桂平市 GP 麻垌镇、大湾镇 4 临桂区 LG 临桂镇、五通镇 6 八步区 BB 桂岭镇、信都镇 7 城中区 CZ 静兰街道、河东街道 8 兴宁区 XN 三塘镇、五塘镇、昆仑镇 16 全州县 QZ 全州镇、庙头镇 6 苍梧县 CW 六堡镇、木双镇 7 容县 RX 六王镇、容州镇 13  下载: 导出CSV
下载: 导出CSV
表 2 基于mt COⅠ基因的广西松材线虫遗传多样性系数表
Table 2 Genetic diversity coefficients of B. xylophilus in Guangxi based on mt COⅠ gene
地区代码
Area
code虫株数量/个
Number of
insect strains/ind多态性位点
Polymorphic
loci简约位点
A minimalist
site单倍型数
Number of
haplotypes单倍型多样性
Haplotype
diversity核苷酸多样性
Nucleotide
diversity平均核苷酸差异系数
Average nucleotide
difference coefficientGP 4 − − 1 − − − LG 6 1 − 2 0.333 0.001 0.333 BB 7 − − 1 − − − CZ 8 − − 1 − − − XN 16 − − 1 − − − QZ 6 1 0 2 0.333 0.001 0.333 CW 7 1 1 2 0.571 0.001 0.571 RX 13 − − 1 − − − All 67 3 1 4 0.170 0 0.174 注:“−”表示不存在,当多样性系数小于0.000 5时,以“0”表示。 Note:‘−’ indicates that it does not exist. When the diversity coefficient is less than 0.000 5,it is expressed as ‘0’.
下载: 导出CSV
表 3 基于mt COⅠ基因的广西松材线虫遗传距离
Table 3 Genetic distances of B. xylophilus in Guangxi based on mt COⅠ gene
地区代码
Area codeGP LG BB CZ NN QZ CW LG 0.000 3 BB − 0.000 3 CZ − 0.000 3 − NN − 0.000 3 − − QZ 0.000 3 0.000 5 0.000 3 0.000 3 0.000 3 CW 0.000 9 0.001 1 0.000 9 0.000 9 0.000 9 0.001 1 RX − 0.000 3 − − − 0.000 3 0.000 9 注:“−”表示不存在。 Note:‘−’ indicates that it does not exist.
下载: 导出CSV
表 4 基于mt COⅠ基因的广西松材线虫单倍型在各个区县的分布
Table 4 Distribution of haplotypes of B. xylophilus based on mt COⅠ gene in various sampling sites in Guangxi
单倍型
Haplotype在各地区的分布 虫株数量/个
Number of strains/indGG GL HZ LZ NN QZ WZ YL Hap1 4 5 7 8 16 5 3 13 61 Hap2 − 1 − − − − − − 1 Hap3 − − − − − 1 − − 1 Hap4 − − − − − − 4 − 4 注:“−”表示不存在。 Note:‘−’ indicates that it does not exist.
下载: 导出CSV
-
[1] 张星耀, 骆有庆. 中国森林重大生物灾害[M]. 北京: 中国林业出版社, 2003: 35−45. [2] 理永霞, 张星耀. 松材线虫入侵扩张趋势分析[J]. 中国森林病虫, 2018, 37(5): 1−4. https://doi.org/10.3969/j.issn.1671-0886.2018.05.001 doi: 10.3969/j.issn.1671-0886.2018.05.001 [3] 叶建仁. 中国森林病虫害防治现状与展望[J]. 南京林业大学学报, 2000, 24(6): 1−5. [4] 吴强, PENG Y, 马恒运, 等. 森林生态系统服务价值及其补偿校准——以马尾松林为例[J]. 生态学报, 2019, 39(1): 117−130. [5] 郑光楠, 杨秀好, 韦曼丽, 等. 广西松褐天牛成虫种群动态规律及其与林分和气象因子相关性[J]. 林业科学, 2023, 59(1): 128−142. https://doi.org/10.11707/j.1001-7488.LYKX20210609 doi: 10.11707/j.1001-7488.LYKX20210609 [6] 郑光楠, 杨秀好, 董子舒, 等. 松褐天牛繁殖行为及其影响因素研究进展[J]. 广西林业科学, 2022, 51(1): 128−135. [7] 叶建仁, 黄麟. 松材线虫病病原学研究的几个问题[J]. 中国森林病虫, 2012, 31(5): 13−21. [8] 黄金思, 奚晓桐, 丁晓磊, 等. 基于SNP标记的广东省松材线虫种群分化研究[J]. 南京林业大学学报(自然科学版), 2019, 43(6): 25−31. [9] 陈婷. 景观特征对小菜蛾种群遗传结构及基因流的影响[D]. 福州: 福建农林大学, 2015. [10] Hebert P, Cywinska A, Ball S, et al. Biological identifications through DNA barcodes [J]. Biological sciences, 2003, 270(1512): 313−321. [11] 杨振德, 赵博光, 郭建. 松材线虫行为学研究进展[J]. 南京林业大学学报(自然科学版), 2003, 27(1): 87−92. [12] 薛清, 杜虹锐, 薛会英, 等. 苜蓿滑刃线虫线粒体基因组及其系统发育研究[J]. 生物技术通报, 2021, 37(7): 98−106. [13] 田月珍, 黄锡霞, 田可川, 等. 细毛羊KRT26基因多态性及其与羊毛细度的关联性分析[J]. 中国畜牧兽医, 2015, 42(1): 161−166. [14] Vasudevan K, Vera C, Gruissem W, et al. Large scale germplasm screening for identification of novel rice blast resistance sources [J]. Frontiers in plant science, 2014, 5: 505. [15] Sharma J, Dutta P, Khan S. Population genetic study of Plasmodium falciparum parasites pertaining to dhps gene sequence in malaria endemic areas of Assam [J]. Indian journal of medical microbiology, 2015, 33(3): 401−405. https://doi.org/10.4103/0255-0857.158565 doi: 10.4103/0255-0857.158565 [16] Rozas J, Ferrer-Mata A, Sánchez-Delbarrio J, et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets [J]. Molecular Biology and Evolution, 2017, 34(12): 3299−3302. https://doi.org/10.1093/molbev/msx248 doi: 10.1093/molbev/msx248 [17] 刘驰, 李家宝, 芮俊鹏, 等. 16S rRNA基因在微生物生态学中的应用[J]. 生态学报, 2015, 35(9): 2769−2788. [18] Jessica W L, David B. POPART: full-feature software for haplotype network construction [J]. Methods in Ecology and Evolution, 2015, 6(9): 1110−1116. https://doi.org/10.1111/2041-210X.12410 doi: 10.1111/2041-210X.12410 [19] 黄麟, 叶建仁, 刘雪莲. 松材线虫病病原种群分化研究现状[J]. 南京林业大学学报(自然科学版), 2009, 33(4): 135−139. [20] Kanzaki N, Futai K. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus group [J]. Nematology, 2002, 4(1): 35−41. https://doi.org/10.1163/156854102760082186 doi: 10.1163/156854102760082186 [21] 成飞雪, 成新跃, 谢丙炎, 等. 松材线虫种群遗传多样性AFLP标记的建立及其应用[J]. 植物病理学报, 2005, 1(5): 410−419. [22] 丁晓磊. 基于高通量测序的松材线虫致病机理与毒力差异研究[D]. 南京: 南京林业大学, 2016. [23] 彭居俐, 王绪桢, 何舜平. DNA条形码技术的研究进展及其应用[J]. 水生生物学报, 2008, 1(6): 916−919. [24] 彭士明, 施兆鸿, 侯俊利, 等. 银鲳3个野生群体线粒体COⅠ基因的序列差异分析[J]. 上海海洋大学学报, 2009, 18(4): 398−402. [25] 李玉婷, 陈茂华. 苹果蠹蛾种群遗传多样性研究进展[J]. 生物安全学报, 2015, 24(4): 287−293. [26] 杨喜书, 章群, 薛丹, 等. 中国近海细鳞鯻线粒体控制区的遗传多样性[J]. 生态学报, 2018, 38(5): 1852−1859. [27] Ye H, Li J, Cao L, et al. Genetic uniformity of Alternanthera philoxeroides in South China [J]. Weed Research, 2003, 43(4): 297−302. https://doi.org/10.1046/j.1365-3180.2003.00346.x doi: 10.1046/j.1365-3180.2003.00346.x [28] 万宣伍, 刘映红, 罗林明, 等. 基于分子生物学方法的外来入侵物种入侵历史重构[J]. 生态学报, 2015, 35(4): 1296−1309. [29] 谢丙炎, 成新跃, 石娟, 等. 松材线虫入侵种群形成与扩张机制——国家重点基础研究发展计划“农林危险生物入侵机理与控制基础研究”进展[J]. 中国科学(C辑: 生命科学), 2009, 39(4): 333−341. [30] 叶建仁, 吴小芹. 松材线虫病研究进展[J]. 中国森林病虫, 2022, 41(3): 1−10. [31] 来燕学. 松墨天牛的飞行特性与防治松材线虫病的指导思想[J]. 浙江林学院学报, 1998, 15(3): 98−101. [32] 叶建仁. 松材线虫病在中国的流行现状、防治技术与对策分析[J]. 林业科学, 2019, 55(9): 1−10. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 927
- HTML全文浏览量: 484
- PDF下载量: 61
- 被引次数: 0
