

-
气孔是叶片表皮特殊器官,主要由2个保卫细胞组成,大多数情况下保卫细胞被特殊分化的表皮细胞包围,称为附属细胞;气孔及其相邻的保卫细胞和附属细胞,被称为气孔复合体或气孔装置[1]。气孔复合体的显著特征是一对作为液压操作阀的保护细胞。在光合中起到吸收CO2释放O2的作用,当光合作用需要CO2时,保卫细胞吸收水分并膨胀以打开孔隙,当不需要CO2或水分胁迫超过植物的光合需要时,保卫细胞失去水分以关闭孔隙[2]。植物中超过90%的CO2和水蒸气都通过气孔,气孔容易受外界环境变化的影响,其形态结构特征是对外界环境变化长期适应的结果[3]。气孔可协调自身大小和密度以适应不同生境,对短期内环境突变快速调节气孔开闭应对环境变化以减少或避免对植物自身不可逆的损伤[4]。光合作用是绿色植物生长的基础,是地球上生物生命的驱动力,是作物产量的主要来源,是提高作物产量的重要方式,也是人类绝大多数能量的来源[5]。为了使植物产生固碳反应,大气中CO2首先要进入到叶片内部,这一过程发生在植物表面的气孔[6],这些气孔则促进CO2气体通过表皮的角质层,进入叶片的细胞间隙,然后扩散到含有叶绿体的器官,在叶绿体中CO2被羧化酶RuBisCO固定[7]。植物可通过调节其叶片的微观结构来适应外部光照变化,从而维持自身的生理机能[8]。叶片是高等植物光合作用的主要器官,叶片上的气孔特性与光合作用特性密切相关,不仅促进CO2的吸收,还促进植物地上部分与大气之间的其他物质的交换。植物在受到生物或非生物胁迫会使叶片表型发生变化,叶片表型结构的变化则会致使生理和生态机能的变化,气孔效应对于干旱或其他胁迫的早期反应是减少叶面积和植物生长,使得蒸腾作用受到影响,从而提高水分利用率[9]。Xiao等[10]通过过表达控制水稻(Oryza sativa)叶片表皮因子的发育信号,能减少气孔产生和降低叶片表面气孔密度,从而显著降低植物的气孔导度,解决了在干旱和高温条件下较难保存种质的问题,同时也提高了水稻的抗旱能力。Wall等[11]在小麦(Triticum aestivum)研究中发现,叶片正面气孔的功能可以保持光合作用的叶片温度,叶片背面气孔气体交换对叶片光合作用约50%的作用。
盐生植物海雀稗(Paspalum vaginatum)是禾本科(Poaceae Barnhart)雀稗(Paspalu)属多年生植物,作为优良的草坪草,海雀稗分枝多为匍匐茎且根系较发达、侵占性强且短时间内扩展较快,可用于土壤盐分较高,淡水资源有限的地区[12 − 13],已有研究证实,C4植物的叶片解剖结构与其光合能力之间存在内在联系。高粱(Sorghum bicolor)的叶片越宽其水分利用率越大,光合作用越强[14]。利用光合和生理特性筛选出了干旱胁迫下的高光效的甘蔗(Saccharum)品种[5]。罗俊等[15]对152份甘蔗光合参数研究建立了7个判别能力较高的判别模型。狗牙根(Cynodon dactylon)与黑麦草(Lolium perenne)同为禾本科草坪草,狗牙根在不同生境下叶表皮细胞差异显著,在黑麦草中发现,气孔导度是影响其叶片光能利用率和水分利用率的主要生理因子[16]。海雀稗作为C4植物与高粱和玉米有着高度的同源性[17]。目前,国内外对于海雀稗的研究大多集中在品种选育[18]、QTL[19]、病害[13]、耐盐性[20]等方面。前人的研究已经证实,C4植物的叶片解剖结构与其光合作用能力之间存在内在关联。目前,作为C4盐生植物的海雀稗其光合特性与气孔特征的相关性研究尚未见报道,因此,本试验采用25份2倍体海雀稗不同种质为研究材料,调查气孔特征与光合作用特性的相关性,旨在揭示海雀稗光合作用与叶片解剖结构之间的关系,为今后筛选盐生植物高光效种质提供科学依据。
-
25份供试种质资源来自美国、泰国、中国等热带和亚热带地区(表1),现保存于海南大学海甸校区热带草坪草与牧草种质资源圃。本试验采用水培培养,将材料装在5 L的盆中,每盆4杯育苗杯,正式试验之前先统一修剪材料至5 cm。材料生长期间每4天浇1次水,每10天加入霍格兰营养液A、B、C液各50 mL,待材料生长1个月后,选择完全展开且没有受到损伤的倒三叶进行指标的测量,并记录数据和采集叶片。
表 1 25份海雀稗种质信息来源
Table 1. Information sources of 25 accessions of seashore paspalum germplasm
种质来源
Germplasm sources材料名称
Name of germplasm澳大利亚Australia PI 647902 (Adalayd)巴哈马Bahama Bahama 中国海南Hainan, China grin_UPG198, 17HN-23 泰国Thailand PI 647900 美国America TG Kona, HI 101, TYB 2, Spence, 17HN-62, PI 647902 , PI 647912, PI 647909(Temple 1), PI 647908, grin_UPG144, PI 647903(Taylor 1), SeaIsle 2000, TFP 7-4, TCR3, Salam, Kai Luna, TGCGC, HH, PI 647904(Taylor 2), KC9 -
光合指标测量于2022−11−13—2022−11−18。试验期间环境日平均温度28.5 ℃,日平均湿度77.4%。由于海雀稗2倍体叶片较其他植物小,测量光合参数之前需提前标记拍照并测量叶片面积以便测量光合参数时输入,转换数据结果。测量时间选择在天气晴朗的上午9:00—11:00,使用CIRAS3便携式光合测定系统(PP Systems, Amesbury, MA, USA)的狭长小型叶视(7 mm×25 mm),选取健康、长势一致的倒三片完全展开叶片进行光合气体交换特征的测量,包括以下参数:净光合速率(Pn)、水分利用率(iWUE)、气孔导度(Gs)。仪器环境参数设定:CO2 400 µmol·m−2;叶室光强1 200 µmol·(m2·s)−1。为减少环境CO2浓度波动对测定参数的影响,使用外接CO2小钢瓶,每个种质测量5个重复。然后,剪取当天所测种质的叶片放入装有体积分数70%的FAA固定液的小棕瓶(容量为3 mL)中保存,以便后续实验使用[9]。
-
自然界中大多数气孔主要分布于下表皮,海雀稗上表皮含有独有特殊突起叶乳突[3]几乎掩盖住上表皮所有气孔,种质不同遮盖量不同,故只印迹下表皮气孔作为统计内容。气孔临时装片采用指甲油印迹法[21],将保存装有FAA固定液的小棕瓶取出叶片待干后,在下表皮薄涂一层透明指甲油并用手持风扇吹干后使用高清透明胶带覆盖在涂有指甲油的面轻刮几下让胶带与指甲油贴合撕下置放载玻片制成临时装片并做编号,同时放置在光学显微镜(Olympus BX41,Olympus,Tokyo,Japan)对其进行图像采集。使用ImageJ软件进行单位面积的气孔形态数据测定,包括气孔密度(stomates·mm−2, SD)、气孔宽度(µm, SW)、保卫细胞长度和保卫宽度(µm, GL、GW)、气孔空隙长度和宽度(µm, PVL、PVW)。并将上述数据转化为气孔特征,包括气孔面积指数(SPI),气孔大小(µm2, SS=SW×GL)(单位面积内出现大小超过50%则算1个气孔)[14]。
-
采用Microsoft Excel 2019软件对各性状进行数据整理,在R(1.4.1)软件中使用corrplot包进行相关性分析[4]。作图使用IMB SPSS Statistics 25进行统计分析和显著性分析,利用主成分分析将计算结果进行聚类分析[22]。得到功能群类似的种质类群,图表中数据为平均值 ± 标准误。
-
从表2可知,光合性状在不同种质间存在极显著差异(P < 0.01)。其中,气孔导度变异系数较大,为40.5%,最小值和最大值分别为71.89 和378.41 mmol·(m2·s)−1。其次为净光合速率和水分利用率,变异系数分别为26.01%和27.4%,两者最大值为最小值的倍数分别是5.25和3.35倍。通过对叶片长宽及气孔结构测量观察发现(表2),叶片长度差异相较于其他结构性状变异系数最大达40.7%,其最小和最大值分别为19.81 µm和113.12 µm,叶宽次之变异系数CV为22.5%,气孔密度、气孔宽度、保卫细胞长度、保卫细胞宽度、气孔空隙(孔径)长度、气孔空隙(孔径)宽度、气孔面积指数和气孔大小的变异系数相对稳定(CV < 1%),对种质的选育分类作用不大。气孔指标中气孔密度、保卫细胞长度、气孔空隙长度、气孔空隙宽度、气孔大小呈极显著差(P < 0.01),保卫细胞宽度和气孔面积指数呈显著差异(P < 0.05)。
表 2 25种海雀稗叶片气孔特征与光合特征
Table 2. Stomatal characteristics and photosynthetic characteristics of leaves of 25 accessions of seashore paspalum
指标Parameter 最小值Min 最大值Max 均值Mean 标准差sd F CV/% Pn/[mol·(m2·s)−1] 10.01 35.38 22.42 5.83 9.21** 26.01 Gs/[mmol·(m2·s)−1] 71.89 378.41 139.50 56.52 317.87** 40.50 iWUE/[mmol·(m2·s)−1] 2.55 8.53 4.78 1.33 16.55** 27.40 LL/µm 19.81 113.12 54.76 22.26 109.30** 40.70 LW/µm 2.02 6.18 3.36 0.76 5.22** 22.50 SD/(stomates·mm−2) 24.11 88.02 47.00 12.91 4.27** 0.27 SW/µm 13.14 25.68 17.34 2.30 2.73** 0.13 GL/µm 20.08 43.52 29.58 4.05 2.14** 0.14 GW/µm 4.14 11.79 7.40 1.26 1.84* 0.17 PL/µm 7.01 26.41 16.05 3.72 2.09** 0.23 PW/µm 0.71 11.56 1.37 0.98 2.94** 0.71 SPI 11 169.43 90 737.51 41 472.50 14 098.62 1.69* 0.34 SS/(ind·µm−2) 323.51 817.30 544.01 110.83 2.92** 0.20 注:*表示在0.05水平上差异达到显著;**表示在0.01差异达到极显著。Pn,净光合速率; Gs,气孔导度; iWUE,水分利用率; LL,叶片长度; LW,叶片宽度; SD,气孔密度; SW,气孔宽度; GL,保卫细胞长度;GW,保卫细胞宽度; PVL,气孔空隙长度; PVW,气孔空隙宽度; SPI,气孔面积指数; SS,气孔大小,下同。
Note: * means significant difference at 0.05 level; ** means significant difference at 0.01 level; Pn. Net photosynthetic rate; Gs. Stomatal conductance; iWUE.Water use efficiency; LL. Leaf length; Lw. Leaf width; SD. Stomatal density; SW. Stomatal width; GL. Guard cell length; GW. Guard cell width; PVL. Pore void length; PW. Pore width; SPI. Stomatal pore index; SS. Stomatal size; similarly hereinafter. -
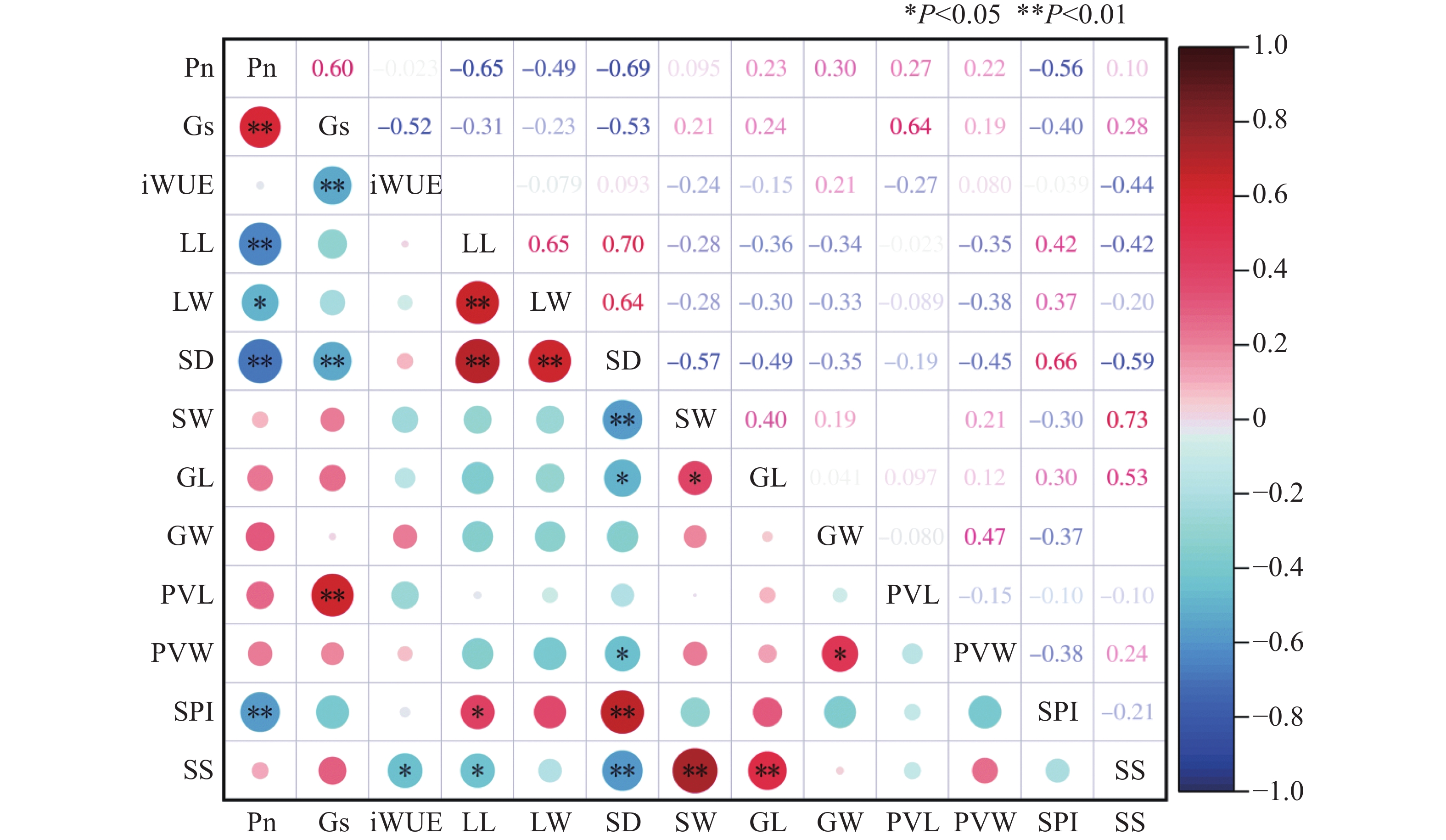
利用Pearson模型对光合作用特征和气孔特征相关性分析,结果表明(图1),净光合速率与叶宽呈显著负相关(P < 0.05),与叶长、气孔密度、气孔面积指数呈极显著负相关(P < 0.01),相关系数分别为−0.65、−0.69、−0.56,与Gs呈极显著正相关(P < 0.01)。气孔导度与水分利用率呈极显著负相关(P < 0.01),与气孔空隙长度呈极显著正相关(P < 0.01),相关系数为0.64。在叶片表型和气孔特征中,叶长和叶宽与气孔密度呈极显著正相关(P < 0.01),相关系数分别为0.70和0.64。气孔密度与保卫细胞长度和气孔大小呈极显著负相关性(P < 0.01),与气孔宽度和气孔空隙宽度呈显著负相关(P < 0.05),与气孔面积指数呈极显著正相关(P < 0.01)。气孔宽度与保卫细胞长度呈显著正相关(P < 0.05),与气孔大小呈极显著正相关(P < 0.01),保卫细胞宽度与气孔空隙宽度呈显著正相关(P < 0.05)。
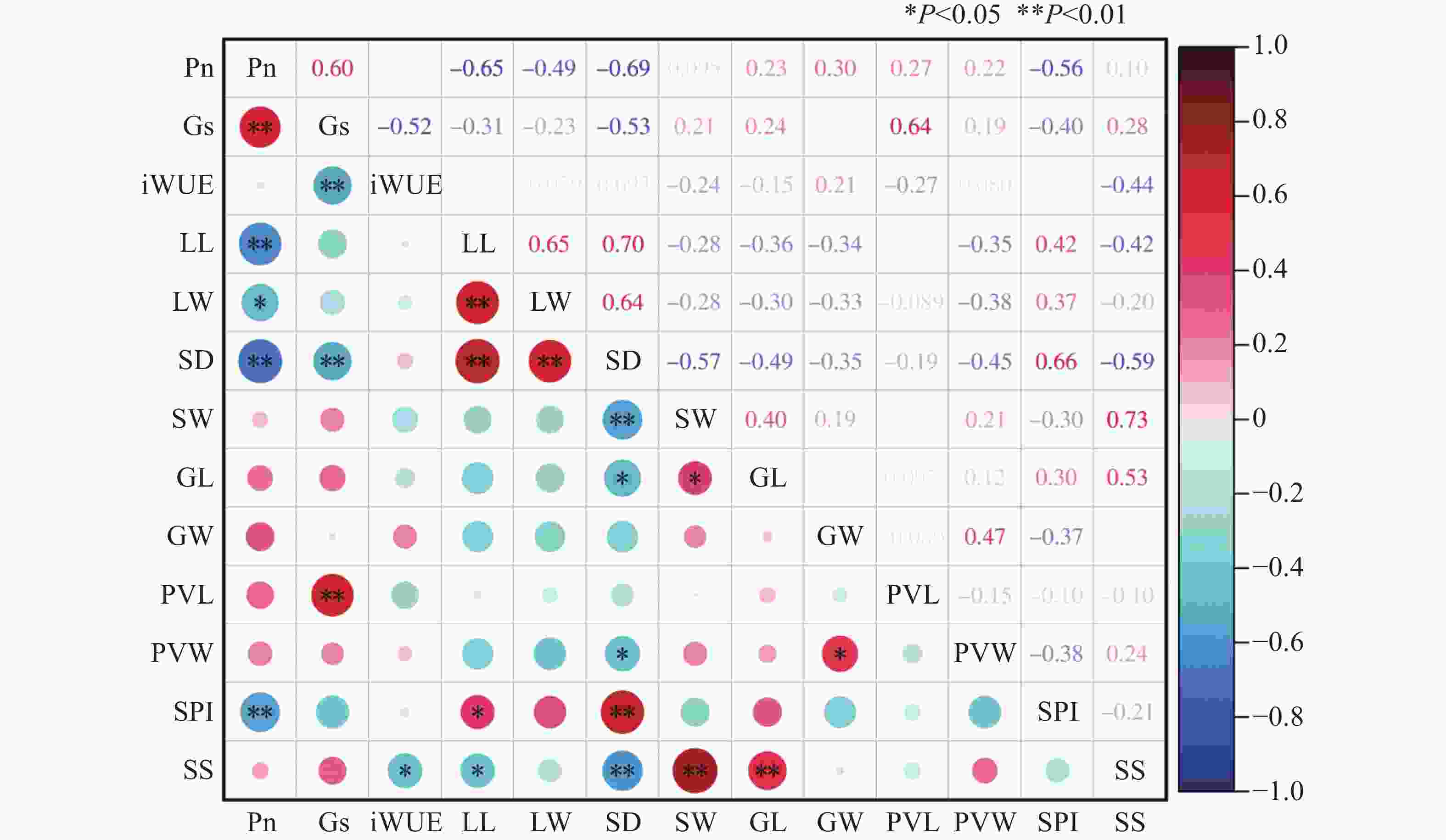
图 1 不同种质叶片气孔特征与光合特性相关性
Figure 1. There was a correlation between leaf stomatal characteristics and photosynthetic characteristics in different accessions of the germplasm
-
对材料叶片气孔特征参数进行主成分分析,前4个主成分特征值均大于1,每个成分累计贡献率分别为36.8%、16.27%、13.88%、8.85%。累计贡献率达75.80%(表3),该累计贡献率符合主成分分析要求可代表绝大部分叶片性状类型。第1主成分特征向量中载荷较大的因子有净光合速率、叶片长度、气孔密度,其中气孔密度特征向量最大为−0.95,其次是净光合速率和叶片长度分别为0.73和−0.78,这些性状主要与光合作用和叶片气孔密度有关。第2主成分中水分利用率负向特征量最大,为−0.76,其次气孔导度正特征向量最大为0.45。第3主成分在气孔孔径长特征向量最大,为−0.68,其次是气孔大小为0.56。第4主成分主要由保卫细胞长和气孔面积指数构成特征向量分别为0.59和0.53,主要与气孔密度和保卫细胞长度有关。净光合速率、气孔导度、气孔大小、保卫细胞长是4个主成分的主导因子,对海雀稗光合特性与气孔特征划分起主要作用。
表 3 海雀稗种质光合参数与叶片性状主成分分析
Table 3. Principal component analysis of phenotypic traits of leaves of seashore Paspalum germplasm
性状Traits 主成分1 Comp.1 主成分2 Comp.2 主成分3 Comp.3 主成分4 Comp.4 Pn/[µmol·(m2·s)−1] 0.73 −0.14 −0.44 0.21 Gs/[mmol·(m2·s)−1] 0.62 0.45 −0.53 −0.09 iWUE/[mmol·(m2·s)−1] −0.20 −0.76 0.07 0.31 LL/µm −0.78 0.18 −0.05 −0.26 LW/µm −0.69 0.28 −0.01 −0.31 SD/(stomates. mm−2) −0.95 0.02 −0.03 −0.01 SW/µm 0.58 0.26 0.49 −0.30 GL/µm 0.47 0.40 0.42 0.59 GW/µm 0.43 −0.57 0.06 −0.08 PVL/µm 0.22 0.43 −0.68 0.13 PVW/µm 0.52 −0.40 0.18 −0.21 SPI −0.63 0.35 0.31 0.53 SS/(ind·µm−2) 0.60 0.43 0.56 −0.22 特征值 Characteristic value 4.05 1.69 1.20 1.01 贡献率/% Contribution rate 36.80 16.27 13.88 8.85 累计贡献率/% Cumulative
contribution rate36.80 53.07 66.95 75.80 -
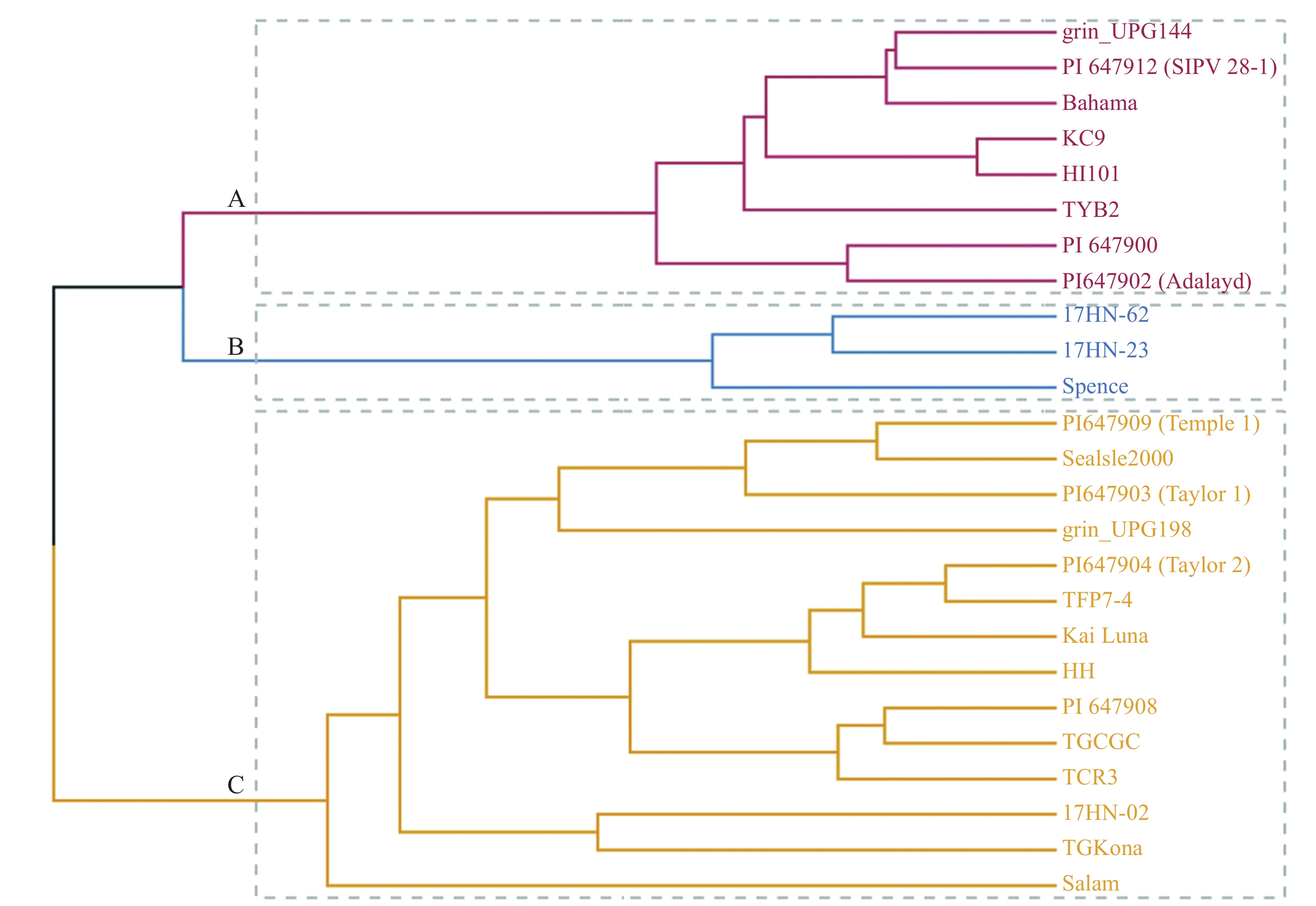
以供试材料叶长、叶宽及气孔性状特征值将25份种质进行聚类划分(图2),将其分为3类(A、B、C)。其中,A类包括grin_UPG144、PI647912、Bahama、KC9、HI101、TYB2、PI647900、PI647902(Adalayd) 8个种质;B类包括17HN-62、17HN-23、Spence 3个种质;C类种质包括PI647909(Temple1)、SeaIsle2000、PI647903 (Taylor1)、grin_UPG198、PI
647904 (Taylor2)、TFP7-4、KaiLua、HH、PI647908、TGCGC、TCR3、17HN-02、TG Kona、Salam,14个种质。从表4中可知,气孔导度、水分利用率、气孔宽度、气孔空隙长度在3个类群间差异均不显著。A类种质净光合速率显著高于B类群,但气孔密度显著低于B类群;B类群净光合速率显著低于A和B类群,但叶片长度、叶片宽度、气孔密度、气孔面积指数显著高于A和C类群;C类群气孔宽度、保卫细胞长度、气孔空隙宽度、气孔大小显著高于A和B类群,净光合速率显著高于B类群,高于A类群但差异不显著。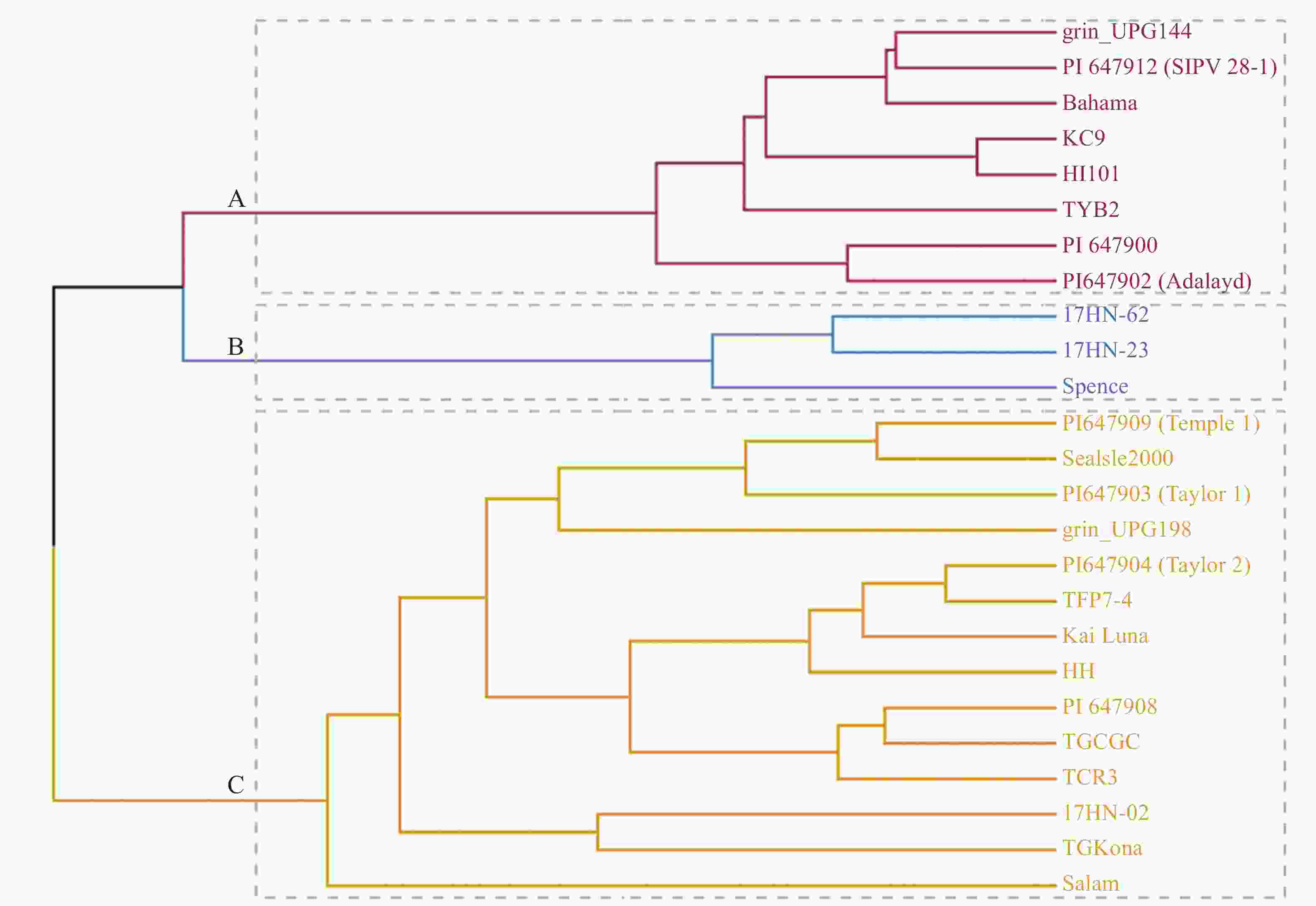
图 2 25份海雀稗主成分综合得分聚类分析
Figure 2. Clustering analysis of the composite scores of principal components of 25 accessions of seashore Paspalum germplasm
表 4 25份海雀稗各性状不同类群差异分析
Table 4. Analysis of differences of traits among different groups of populations of 25 accessions of seashore Paspalum germplasm
性状 Traits A类群 Group A B类群 Group B C类群 Group C Pn/[mol·(m2·s)−1] 23.37 ± 1.12a 13.48 ± 0.43b 23.96 ± 1.19a Gs/[mmol·(m2·s)−1] 127.43 ± 10.13a 101.91 ± 4.76a 154.45 ± 18.03a iWUE/[mmol·(m2·s)−1] 5.11 ± 0.3a 4.54 ± 0.67a 4.63 ± 0.36a LL/µm 53.56 ± 5.61b 99.11 ± 3.16a 45.96 ± 3.86b LW/µm 3.25 ± 0.13b 4.68 ± 0.25a 3.13 ± 0.07b SD/(stomates·mm−2) 51.87 ± 1.6b 70.13 ± 3a 39.7 ± 1.29c SW/µm 16.16 ± 0.29b 16.47 ± 0.84b 18.19 ± 0.3a GL/µm 28.31 ± 0.51ab 27.43 ± 0.69b 30.76 ± 0.64a GW/µm 6.95 ± 0.17a 7.07 ± 0.06a 7.71 ± 0.17a PL/µm 16.33 ± 0.5a 15.61 ± 1.51a 15.98 ± 0.73a PW/µm 1.14 ± 0.04ab 1.03 ± 0.06b 1.57 ± 0.11a SPI 42 180.51 ± 1 808.8b 52 897.75 ± 881.53a 38 664.19 ± 2 240.81b 注:不同小写字母表示差异达显著水平(P<0.05)。
Note: Different lowercaseletters indicate significant difference level (P<0.05). -
植物光合性状的差异与其自身的遗传特性、生物学特性以及外界生理生态环境影响有关,不同品种间其光合能力存在较大差异[1]。本研究中25份海雀稗光合参数(净光合速率、气孔导度、水分利用率)和叶片性状(叶长、叶宽、气孔密度、气孔宽度、保卫细胞长度、保卫细胞宽度、气孔空隙长度、气孔空隙宽度、气孔面积指数、气孔大小)间均呈显著差异。植物光合性状的差异与其自身的遗传特性、生物学特性以及外界生理生态环境影响有关,马铃薯(Solanum tuberosum)不同品种间其光合能力存在较大差异[23]。枸杞(Lycium chinense)的净光合速率与气孔导度呈正相关[24],气孔导度与水分利用率呈负相关;龙明秀等[16]在黑麦草的光合与水分利用效率研究中发现,净光合速率与气孔导度呈显著正相关;高粱群体光合特征之间的关联性分析结果得到,净光合速率与气孔导度呈显著正相关;甘蔗的近缘种蔗茅,36份不同种质的光合特性的差异分析中净光合速率与气孔导度呈显著正相关[25],本研究结果表明,净光合速率与气孔导度呈极显著正相关性,此结果与上述作物研究结果一致。秦茜等[26]在甘蔗的光合与解剖结构研究中发现,净光合速率与气孔密度呈显著正相关,但海雀稗叶片净光合速率与气孔密度的相关性与甘蔗研究中结果相反,虽然甘蔗与海雀稗同为禾本科植物,其结果不同的原因可能是糖料作物与盐生植物的区别,也可能是环境不同所导致,因为植物器官大多都是为响应生存环境从而协调自身器官功能。
气孔特殊孔径的打开和关闭控制着叶片与大气进行气体交换的通道,也可以通过协调气孔大小甚至孔径大小适应不同环境[27]。在叶片较宽的作物中,较宽的叶片则需要蒸发更多的水分防止叶片在高温环境受到损害,从而提高植物蒸腾速率,进而降低了水分利用率[28]。对于田间生长的大豆(Glycine max),叶片较小的植株遇高温气候,蒸腾速率降低,水分利用率升高。本研究中,海雀稗各种质间变异系数为范围为0.13% ~ 40.7%,其中,净光合速率、水分利用率、叶长、叶宽变异系数都超过20%,叶长变异系数高达40.7%,叶长和叶宽呈极显著正相关,与卫尊征等[29]对小叶杨(Populus simonii)叶片性状研究结果一致。本研究发现,叶宽与水分利用率没有显著负相关,这一结果与上述前人的研究结果相左,可能是因为受环境地区或温度的影响所致,但水分利用率随着叶片宽度增加而呈降低趋势,而叶宽与净光合速率呈显著负相关,由于植物水分利用率是由净光合速率和蒸腾速率共同决定,结果从侧面反映了叶宽与植物水分利用率呈负相关性,该结果与糖料作物高粱基因型间水分利用率较高的预测指标的结果相同[14]。以上可为海雀稗的高光效良种选育提供参考。
主成分分析将海雀稗光合参数、叶长、叶宽以及气孔特征指标转化为4个主成分,4个主成分提供了原指标的75.80%的信息。其中,净光合速率、气孔导度、气孔大小,保卫细胞长是4个主成分的主导因子,将其聚类得到3个不同类群种质。A类属中等净光合速率的类群,叶长、叶宽气孔等其他指标属中等类群。B类光合效率显著低于A、C两类群,但叶长、叶宽,气孔密度等指标较高的类群。C类群属净光合速率较高类群,且气孔宽度、保卫细胞长度、气孔空隙宽度以及气孔大小显著高于其他2个类群。小气孔对大气孔而言,在短时间内环境变化,可以快速开闭气孔减少或避免对植物损害[5]。甜菜气孔小且密度大,利用气孔密度和气孔大小相互协调作用于净光合速率和气孔导度以此来优化叶片水分利用率[30]。针对没有盐腺的盐生植物海蓬子(Salicornia europae),在盐质量浓度为4%时光合能力显著下降,导致光合能力下降的主要原因是气孔关闭导致的气孔限制或是其他器官为响应胁迫而引起的非气孔限制,最终结果表明气孔限制值是抑制海蓬子光合作用的关键因素[31]。本研究中气孔密度与保卫细胞长度,气孔宽度、气孔孔径、气孔大小呈显著负性,说明海雀稗可调控叶片气孔形态特征对环境变化的响应,此结果与前人研究一致[4]。高光效亲本是对作物选育高光效育种的关键,筛选优良种质资源作为高光效育种亲本,结合光合速率的遗传规律,可实现培育出高光效优质的海雀稗新品种。
综上所述,海雀稗光合气体交换受气孔各个指标的调控,净光合速率与气孔密度呈极显著负相关,利用主成分分析将25份海雀稗进行聚类得到3个不同类群种质,3个类群净光合速率强弱顺序为C > A > B类群,C类群属净光合速率高的种质,其气孔特征为气孔密度低,而气孔大小、气孔宽度、气孔空隙宽度都显著高于其他净光合速率强的类群的特点。本研究成果为探索盐生植物海雀稗光合作用与气孔特征的关联性提供了新思路,同时也为后期开展盐生植物高光效育种提供了理论参考。
Stomatal and photosynthetic characteristics of halophyte plant, seashore Paspalum
-
摘要: 为探讨不同种质海雀稗(Paspalum vaginatum)叶片气孔特征与光合特性之间的潜在关系,以25份海雀稗种质为研究对象,对光合参数、叶长、叶宽及气孔特征指标进行差异分析。结果表明,不同种质海雀稗各性状均呈显著性(P < 0.05)且不同性状间均表现出显著正相关或负相关(P < 0.05)。净光合速率与叶长、气孔密度、气孔面积指数呈极显著负相关(P < 0.01),与叶宽呈显著负相关(P < 0.05)。气孔导度与气孔密度呈极显著负相关(P < 0.01),与气孔空隙长度呈极显著正相关(P < 0.01),而水分利用率除了气孔大小呈显著负相关性(P < 0.05),与其他气孔特性相关性均不显著(P > 0.05)。利用主成分分析得到净光合速率、气孔导度、气孔大小、保卫细胞长度4个为主导因子,累计贡献率达75.80%。利用这4个指标对25份海雀稗种质进行聚类,共分成3类(A、B和C)。C类包括14个种质,其光合特性表现优异,主要体现在净光合速率最高、气孔密度最低、气孔尺寸最大以及气孔开度最宽等特征。海雀稗C类种质能够通过调控气孔及其附属细胞的功能来优化光合能力,其中气孔相关指标是影响净光合速率的关键因素。Abstract: An attempt was made to analyze the relationship between stomatal and photosynthetic characteristics of different accessions of seashore paspalum (Paspalum vaginatum Sw.), and 25 seashore Paspalum germplasm were analyzed in terms of photosynthetic parameters, leaf length, leaf width and stomatal characteristics. The results showed that all traits of the paspalum germplasm were significantly different (P < 0.05), and were significantly (P < 0.05) positively or negatively correlated with each other. The net photosynthetic rate was significantly negatively correlated with the leaf length, stomatal density and stomatal area index (P < 0.01), and significantly negatively correlated with leaf width (P < 0.05). Stomatal conductance was significantly negatively correlated with stomatal density (P < 0.01), and positively with stomatal pore length (P < 0.01), while water utilization was negatively correlated with stomatal size (P < 0.05), but not with other stomatal characteristics (P > 0.05). The principal component analysis showed that the cumulative contribution rate of the four main components (net photosynthetic rate, stomatal conductance, stomatal size and guard cell length) was 75.80%. Twenty-five accessions of the paspalum germplasm were classified into three groups (A, B and C) by using four indexes. Group C includes 14 accessions that have the highest net photosynthetic rate with the largest stomatal size, the widest stomatal pore and low stomatal density. Group C are accessions of seashore Paspalum germplasm with better photosynthetic characteristics, with their photosynthetic capacity regulated through the function of stomatal accessory cells, and the stomatal traits are also key indicators for regulating net photosynthetic rate.
-
图 2 25份海雀稗主成分综合得分聚类分析
Fig. 2 Clustering analysis of the composite scores of principal components of 25 accessions of seashore Paspalum germplasm
表 1 25份海雀稗种质信息来源
Table 1 Information sources of 25 accessions of seashore paspalum germplasm
种质来源
Germplasm sources材料名称
Name of germplasm澳大利亚Australia PI 647902 (Adalayd)巴哈马Bahama Bahama 中国海南Hainan, China grin_UPG198, 17HN-23 泰国Thailand PI 647900 美国America TG Kona, HI 101, TYB 2, Spence, 17HN-62, PI 647902 , PI 647912, PI 647909(Temple 1), PI 647908, grin_UPG144, PI 647903(Taylor 1), SeaIsle 2000, TFP 7-4, TCR3, Salam, Kai Luna, TGCGC, HH, PI 647904(Taylor 2), KC9  下载: 导出CSV
下载: 导出CSV
表 2 25种海雀稗叶片气孔特征与光合特征
Table 2 Stomatal characteristics and photosynthetic characteristics of leaves of 25 accessions of seashore paspalum
指标Parameter 最小值Min 最大值Max 均值Mean 标准差sd F CV/% Pn/[mol·(m2·s)−1] 10.01 35.38 22.42 5.83 9.21** 26.01 Gs/[mmol·(m2·s)−1] 71.89 378.41 139.50 56.52 317.87** 40.50 iWUE/[mmol·(m2·s)−1] 2.55 8.53 4.78 1.33 16.55** 27.40 LL/µm 19.81 113.12 54.76 22.26 109.30** 40.70 LW/µm 2.02 6.18 3.36 0.76 5.22** 22.50 SD/(stomates·mm−2) 24.11 88.02 47.00 12.91 4.27** 0.27 SW/µm 13.14 25.68 17.34 2.30 2.73** 0.13 GL/µm 20.08 43.52 29.58 4.05 2.14** 0.14 GW/µm 4.14 11.79 7.40 1.26 1.84* 0.17 PL/µm 7.01 26.41 16.05 3.72 2.09** 0.23 PW/µm 0.71 11.56 1.37 0.98 2.94** 0.71 SPI 11 169.43 90 737.51 41 472.50 14 098.62 1.69* 0.34 SS/(ind·µm−2) 323.51 817.30 544.01 110.83 2.92** 0.20 注:*表示在0.05水平上差异达到显著;**表示在0.01差异达到极显著。Pn,净光合速率; Gs,气孔导度; iWUE,水分利用率; LL,叶片长度; LW,叶片宽度; SD,气孔密度; SW,气孔宽度; GL,保卫细胞长度;GW,保卫细胞宽度; PVL,气孔空隙长度; PVW,气孔空隙宽度; SPI,气孔面积指数; SS,气孔大小,下同。
Note: * means significant difference at 0.05 level; ** means significant difference at 0.01 level; Pn. Net photosynthetic rate; Gs. Stomatal conductance; iWUE.Water use efficiency; LL. Leaf length; Lw. Leaf width; SD. Stomatal density; SW. Stomatal width; GL. Guard cell length; GW. Guard cell width; PVL. Pore void length; PW. Pore width; SPI. Stomatal pore index; SS. Stomatal size; similarly hereinafter.
下载: 导出CSV
表 3 海雀稗种质光合参数与叶片性状主成分分析
Table 3 Principal component analysis of phenotypic traits of leaves of seashore Paspalum germplasm
性状Traits 主成分1 Comp.1 主成分2 Comp.2 主成分3 Comp.3 主成分4 Comp.4 Pn/[µmol·(m2·s)−1] 0.73 −0.14 −0.44 0.21 Gs/[mmol·(m2·s)−1] 0.62 0.45 −0.53 −0.09 iWUE/[mmol·(m2·s)−1] −0.20 −0.76 0.07 0.31 LL/µm −0.78 0.18 −0.05 −0.26 LW/µm −0.69 0.28 −0.01 −0.31 SD/(stomates. mm−2) −0.95 0.02 −0.03 −0.01 SW/µm 0.58 0.26 0.49 −0.30 GL/µm 0.47 0.40 0.42 0.59 GW/µm 0.43 −0.57 0.06 −0.08 PVL/µm 0.22 0.43 −0.68 0.13 PVW/µm 0.52 −0.40 0.18 −0.21 SPI −0.63 0.35 0.31 0.53 SS/(ind·µm−2) 0.60 0.43 0.56 −0.22 特征值 Characteristic value 4.05 1.69 1.20 1.01 贡献率/% Contribution rate 36.80 16.27 13.88 8.85 累计贡献率/% Cumulative
contribution rate36.80 53.07 66.95 75.80
下载: 导出CSV
表 4 25份海雀稗各性状不同类群差异分析
Table 4 Analysis of differences of traits among different groups of populations of 25 accessions of seashore Paspalum germplasm
性状 Traits A类群 Group A B类群 Group B C类群 Group C Pn/[mol·(m2·s)−1] 23.37 ± 1.12a 13.48 ± 0.43b 23.96 ± 1.19a Gs/[mmol·(m2·s)−1] 127.43 ± 10.13a 101.91 ± 4.76a 154.45 ± 18.03a iWUE/[mmol·(m2·s)−1] 5.11 ± 0.3a 4.54 ± 0.67a 4.63 ± 0.36a LL/µm 53.56 ± 5.61b 99.11 ± 3.16a 45.96 ± 3.86b LW/µm 3.25 ± 0.13b 4.68 ± 0.25a 3.13 ± 0.07b SD/(stomates·mm−2) 51.87 ± 1.6b 70.13 ± 3a 39.7 ± 1.29c SW/µm 16.16 ± 0.29b 16.47 ± 0.84b 18.19 ± 0.3a GL/µm 28.31 ± 0.51ab 27.43 ± 0.69b 30.76 ± 0.64a GW/µm 6.95 ± 0.17a 7.07 ± 0.06a 7.71 ± 0.17a PL/µm 16.33 ± 0.5a 15.61 ± 1.51a 15.98 ± 0.73a PW/µm 1.14 ± 0.04ab 1.03 ± 0.06b 1.57 ± 0.11a SPI 42 180.51 ± 1 808.8b 52 897.75 ± 881.53a 38 664.19 ± 2 240.81b 注:不同小写字母表示差异达显著水平(P<0.05)。
Note: Different lowercaseletters indicate significant difference level (P<0.05).
下载: 导出CSV
-
[1] LAWSON T, BLATT M R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency[J]. Plant Physiology, 2014, 164(4): 1556 − 1570. doi: 10.1104/pp.114.237107 [2] XU Z Z, ZHOU G S. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass[J]. Journal of Experimental Botany, 2008, 59(12): 3317 − 3325. doi: 10.1093/jxb/ern185 [3] THOLEN D, BOOM C, ZHU X G. Opinion: prospects for improving photosynthesis by altering leaf anatomy[J]. Plant Science, 2012, 197: 92 − 101. doi: 10.1016/j.plantsci.2012.09.005 [4] 杨克彤, 常海龙, 陈国鹏, 等. 兰州市主要绿化植物气孔性状特征[J]. 植物生态学报, 2021, 45(2): 187 − 196. doi: 10.17521/cjpe.2020.0257 [5] 刘硕, 樊仙, 全怡吉, 等. 干旱胁迫对甘蔗光合生理特性的影响[J]. 西南农业学报, 2022, 35(8): 1776 − 1785. doi: 10.16213/j.cnki.scjas.2022.8.007 [6] FRANKS P J, CASSON S. Connecting stomatal development and physiology[J]. New Phytologist, 2014, 201(4): 1079 − 1082. doi: 10.1111/nph.12673 [7] HARRISON E L, ARCE CUBAS L, GRAY J E, et al. The influence of stomatal morphology and distribution on photosynthetic gas exchange[J]. The Plant Journal, 2020, 101(4): 768 − 779. doi: 10.1111/tpj.14560 [8] 王玟玥, 赵丹丹, 张鸽香, 等. 毛茛科3种植物在遮光下叶片解剖结构比较及耐阴性评价[J]. 南方农业学报, 2024, 55(6): 1713 − 1723. doi: 10.3969/j.issn.2095-1191.2024.06.016 [9] 王孟珂, 田梦妮, 毕泉鑫, 等. 基于气孔性状的文冠果种质资源抗旱性评价及抗旱资源筛选[J]. 植物研究, 2021, 41(6): 957 − 964. doi: 10.7525/j.issn.1673-5102.2021.06.014 [10] XIAO Y, SLOAN J, HEPWORTH C, et al. Defining the scope for altering rice leaf anatomy to improve photosynthesis: a modelling approach[J]. New Phytologist, 2023, 237(2): 441 − 453. doi: 10.1111/nph.18564 [11] WALL S, VIALET-CHABRAND S, DAVEY P, et al. Stomata on the abaxial and adaxial leaf surfaces contribute differently to leaf gas exchange and photosynthesis in wheat[J]. New Phytologist, 2022, 235(5): 1743 − 1756. doi: 10.1111/nph.18257 [12] 刘刊, 耿士均, 商海燕, 等. 草坪草抗性研究进展[J]. 草业科学, 2012, 29(7): 1058 − 1064. [13] 许涛, 胡旭, 孙劝劝, 等. 海雀稗种质资源币斑病抗性鉴定[J]. 分子植物育种, 2024, 22(18): 6090 − 6100. doi: 10.13271/j.mpb.022.006090 [14] PAN L, GEORGE-JAEGGLI B, BORRELL A, et al. Coordination of stomata and vein patterns with leaf width underpins water-use efficiency in a C4 crop[J]. Plant, Cell & Environment, 2022, 45(6): 1612 − 1630. doi: 10.1111/pce.14225 [15] 罗俊, 张华, 徐良年, 等. 甘蔗不同品种光合特性比较及其聚类分析[J]. 中国农业科学, 2005, 38(8): 1562 − 1569. doi: 10.3321/j.issn:0578-1752.2005.08.009 [16] 龙明秀, 高景慧, 李波, 等. 黑麦草杂交F2代光合及水分利用效率研究[J]. 草地学报, 2009, 17(2): 151 − 156. doi: 10.3969/j.issn.1007-0435.2009.02.004 [17] SUN G C, WASE N, SHU S Q, et al. Genome of Paspalum vaginatum and the role of trehalose mediated autophagy in increasing maize biomass[J]. Nature Communications, 2022, 13(1): 7731. doi: 10.1038/s41467-022-35507-8 [18] LIU Y M, DU H M, WANG K, et al. Differential photosynthetic responses to salinity stress between two perennial grass species contrasting in salinity tolerance[J]. HortScience, 2011, 46(2): 311 − 316. doi: 10.21273/HORTSCI.46.2.311 [19] WU P P, COGILL S, QIU Y J, et al. Comparative transcriptome profiling provides insights into plant salt tolerance in seashore paspalum (Paspalum vaginatum)[J]. BMC Genomics, 2020, 21(1): 131. doi: 10.1186/s12864-020-6508-1 [20] 申晴, 韦海燕, 卞华, 等. 海雀稗种质资源的耐盐性评价[J]. 热带生物学报, 2020, 11(1): 11 − 19. doi: 10.15886/j.cnki.rdswxb.2020.01.003 [21] 张景润, 王彬存. 春季温室内两种竹类植物叶片解剖结构与光合特性研究[J]. 湖北农业科学, 2020, 59(20): 108 − 111. doi: 10.14088/j.cnki.issn0439-8114.2020.20.024 [22] 田英, 朱强, 尹蓉, 等. 基于光合特性的不同欧李种质资源评价[J]. 经济林研究, 2021, 39(2): 1 − 9. [23] 闫雷, 邹莹, 张等宏, 等. 马铃薯叶片光合效率遗传变异分析及高光效种质筛选[J]. 种子, 2022, 41(11): 25 − 30. doi: 10.16590/j.cnki.1001-4705.2022.11.025 [24] 何昕孺, 王玉静, 李妍颖, 等. 枸杞光合特性评价及高光效指标筛选[J]. 西北农业学报, 2022, 31(7): 893 − 901. doi: 10.7606/j.issn.1004-1389.2022.07.011 [25] 徐超华, 李纯佳, 陆鑫, 等. 甘蔗近缘种蔗茅(Erianthus fulvus)光合气体交换特性的差异分析[J]. 中国农业科学, 2016, 49(15): 2909 − 2920. doi: 10.3864/j.issn.0578-1752.2016.15.005 [26] 秦茜, 朱俊杰, 关心怡, 等. 七个甘蔗品种叶片解剖结构特征与光合能力和耐旱性的关联[J]. 植物生理学报, 2017, 53(4): 705 − 712. doi: 10.13592/j.cnki.ppj.2017.0038 [27] 倪榕蔚, 甘玉婷, 杨桂梅, 等. 热岛效应下亚热带城市植被叶气孔权衡特征及其与叶功能性状的关系[J]. 生态学报, 2023, 43(13): 5336 − 5346. doi: 10.5846/stxb202112243645 [28] JONES H G. Partitioning stomatal and non-stomatal limitations to photosynthesis[J]. Plant, Cell & Environment, 1985, 8(2): 95 − 104. doi: 10.1111/j.1365-3040.1985.tb01227.x [29] 卫尊征, 潘炜, 赵杏, 等. 我国东北及华北地区小叶杨形态及生理性状遗传多样性研究[J]. 北京林业大学学报, 2010, 32(5): 8 − 14. doi: 10.13332/j.1000-1522.2010.05.011 [30] BARRATT G E, SPARKES D L, MCAUSLAND L, et al. Anisohydric sugar beet rapidly responds to light to optimize leaf water use efficiency utilizing numerous small stomata[J]. AoB PLANTS, 2021, 13(1): plaa067. doi: 10.1093/aobpla/plaa067 [31] 刘伟成, 郑春芳, 陈琛, 等. 花期海蓬子对盐胁迫的生理响应[J]. 生态学报, 2013, 33(17): 5184 − 5193. doi: 10.5846/stxb201205250778 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 831
- HTML全文浏览量: 425
- PDF下载量: 32
- 被引次数: 0
