





-
大丽轮枝菌(Verticillium dahliae)寄主范围广,可危害660种植物,其中包括近200种作物[1 − 2]。在中国,大丽轮枝菌最常见的寄主有马铃薯(Solanum tuberosum)、辣椒(Capsicum annuum)、瓜类(Cucurbitaceae)、番茄(Lycopersicon esculentum)、豆类(Fabaceae)、茄子(Solanum melongena)[3]。大丽轮枝菌是引起中国棉花黄萎病的病原菌[4]。
中国是世界前三的棉花生产大国,形成了以新疆为主,包括长江流域、黄河流域的3大产棉区[5]。棉花黄萎病是棉花生产上危害最严重的病害之一,发病后可造成叶片失绿变黄、蕾铃脱落甚至整株死亡,严重影响棉花的产量和品质。由于棉花黄萎病症状表现多样,且其症状常因不同的环境条件、发病阶段、品种及生育期而呈现较大变化,当田间症状不够典型或诊断者经验不足时,只依据田间病害症状不易准确判别[6]。因此,为了准确地预测和有效地控制该病害,应用准确、快速、低成本的诊断方法进行早期诊断是控制棉花黄萎病的基础。
大丽轮枝菌可在土壤存活,是病害流行的重要侵染源。据报道,其产生的微菌核可以在土壤中存活数十年。棉籽带菌是大丽轮枝菌远距离传播的主要方式[7]。棉花黄萎病菌一旦在棉花维管束定殖,药剂防治困难。因此,及时地检测棉田土壤中大丽轮枝菌的菌量和棉籽带菌情况对棉花黄萎病的早期监测预警和防治具有重要意义。国内外已经开发了各种各样的分子检测技术检测病原菌,如普通PCR分子检测[8]、荧光定量PCR检测技术[9]、巢式PCR检测技术[10]、LAMP检测技术[11]及RPA-CRISPR/Cas12a技术[12]等,针对大丽轮枝菌也已开发多种分子检测技术和方法。包括PCR[8]、LAMP技术[6, 9]、微滴式数字PCR和实时定量PCR[10],以及RAPD-PCR[11]技术等。但这些技术大多针对土壤中是否携带大丽轮枝菌微菌核的检测。巢氏PCR技术在病原菌检测方面的应用已经极为广泛[12 − 13],且国内外也有巢氏PCR检测大丽轮枝菌的报道[14 − 15],但这技术只是用于土壤带菌检测,且不能区分大丽轮枝菌的近源种[16 − 19],针对棉籽带菌检测的巢氏PCR技术还未见开发。笔者筛选到一段大丽轮枝菌不同菌株中高度保守、种间有明显差异的基因序列(CP010981.1),依托其中一小段大丽轮枝菌特有的序列,设计了多对引物,通过筛选,找到了一组特异性强、灵敏度高的引物,并建立了检测棉籽是否携带大丽轮枝菌的巢氏PCR检测技术。
-
试验所用的健康棉籽及大丽轮枝菌菌株由新疆农业科学院刘海洋副研究员和徐建辉副研究员及江苏省农业科学院植物保护研究所林铃研究员馈赠,使用的发病田棉籽由石河子大学任毓忠副教授馈赠。立枯丝核菌(Rhizoctonia solani)由中国农业科学院棉花研究所魏锋副研究员馈赠,长孢轮枝菌(Verticillium longisporum)由甘肃省农业科学院植物保护研究所何苏琴副研究员馈赠,非苜蓿轮枝菌(Verticillium nonalfalfae)、黑白轮枝菌(Verticillium albo-atrum)、变黑轮枝菌(Verticillium nigrescens)由中国农业科学院苏晓峰副研究员馈赠,苜蓿轮枝菌(Verticillium alfalfae)、云状轮枝菌(Verticillium nubilum)由西北农林科技大学胡小平教授馈赠,哈茨木霉(Trichoderma harzianum)、禾谷镰刀菌(Fusarium graminearum)、辣椒疫霉(Phytophthora capsici)、棉花角斑病黄单胞菌(Xanthmonas citri subsp. malvacearum)、大豆疫霉(Phytophthora sojae)、棘孢木霉(Trichoderma asperellum)、尖孢镰刀菌(Fusarium oxysporum f. sp. cubense)、稻瘟菌(Magnaporthe oryzae)为海南大学植物保护学院保存(表1)。
表 1 供试菌株
Table 1. Pathogens under test
种名
Species分离材料
Separation materials来源
Source数量/株
Number/strain培养
Medium大丽轮枝菌(Verticillium dahliae) 棉花(Gossypium hirsutum) 新疆
江苏16 PDA 长孢轮枝菌(Verticillium longisporum) 萝卜(Raphanus sativus) 青海 1 PDA 非苜蓿轮枝菌(Verticillium nonalfalfae) — — 1 PDA 变黑轮枝菌(Verticillium nigrescens) — — 1 PDA 黑白轮枝菌(Verticillium albo-atrum) — — 1 PDA 苜蓿轮枝菌(Verticillium alfalfae) — — 1 PDA 云状轮枝菌(Verticillium nubilum) — — 1 PDA 立枯丝核菌(Rhizoctonia solani) — 河南 1 PDA 哈茨木霉(Trichoderma harzianum) 土壤 Soil — 1 PDA 禾谷镰刀菌(Fusarium graminearum) 小麦(Triticum aestivum) — 1 PDA 辣椒疫霉(Phytophthora capsici) 辣椒(Capsicum annuum) — 1 V8 棉花角斑病黄单胞菌(Xanthmonas citri subsp. malvacearum) 棉花(Gossypium hirsutum) — 1 NA 大豆疫霉(Phytophthora sojae) 大豆(Glycine max) — 1 V8 棘孢木霉(Trichoderma asperellum) 土壤 Soil — 1 PDA 尖孢镰刀菌(Fusarium oxysporum f. sp. cubense) 香蕉(Musa nana) — 1 PDA 稻瘟菌(Magnaporthe oryzae) 水稻(Oryza sativa) — 1 PDA 注:表中“—”表示分离材料或来源未知。 Note: In the table “—” indicates that the separation material or source is unknown. -
将各供试真菌接种在PDA培养基上培养15~20 d后刮取其菌丝,采用Fungal DNA Kit(OMEGA BIOTEK公司)提取总DNA。大豆疫霉和辣椒疫霉经V8液体培养基摇菌培养后获得菌体,用CTAB法提取基因组DNA。棉花细菌性角斑病菌通过NA液体培养基培养后获得菌体,用 TIANamp Bacteria DNA Kit(TIANGEN公司)提取基因组DNA。混菌棉花种子采用Fungal DNA Kit(OMEGA BIOTEK公司)提取其DNA。
-
将经PDA培养的大丽轮枝菌接种到液体查氏培养基中,25 ℃,180 r·min−1培养3~5 d,过滤获得孢子悬浮液。在显微镜下用血球计数板将浓度分别稀释成104、103、102和10 个·μL−1共4个梯度,用移液器分别各取其中100 μL滴入每颗切开的棉籽内部(胚芽上),并以接入无菌水的棉籽作为阴性对照,室内风干0.5~1.0 h,直至胚芽表面干燥。
-
通过序列比对确定候选序列(CP010981.1)利用NCBI (National Center of Biotechnology Information,
https://www.ncbi.nlm.nih.gov/ )和Primer 5共同设计6对引物(表2)。表 2 巢式PCR供试引物表
Table 2. Nested PCR primer
引物名称
Primer序列(5′-3′)
Sequence(5′-3′)PCR产物长度/bp
Length/bp引物名称
Primer序列(5′-3′)
Sequence(5′-3′)PCR产物长度/bp
Length/bpVD-1F ATGAAACGTAAGGTAGGTAGC 601 VD-4F GCCAATCGTGACTGCCTCTT 465 VD-1R CCAGAGCAGCGCCTCATTCGG VD-4R GCGCAAACTTGACCAACAGC VD-2F GCGGAAGGGGCACTAGTAAT 512 VD-5F CTCGGCCAAGTCGGAACTTT 375 VD-2R CGCGCAAACTTGACCAACAG VD-5R CGCGCAAACTTGACCAACA VD-3F GTAGGGACGCAACAATGAGC 410 VD-6F CGCCTATGGTCGTGGTCTTT 170 VD-3R) TGACCAACAGCAGCCAGATC VD-6R GTCGGTGAGCTATGCCAGAA -
PCR体系(20 μL):上下游引物(10 μmol·L−1)各0.2 μL,2×Taq Plus Master Mix Ⅱ(Dye Plus)10 μL,DNA 1.0 μL,ddH2O补足至20 μL。
PCR程序:95 ℃ 5 min;95 ℃ 30 s;58 ℃ 30 s;72 ℃ 1 min,35个循环;72 ℃ 10 min,产物通过2%的凝胶电泳检测。
-
将第1轮PCR扩增产物稀释1 000倍,稀释产物作为第2轮巢式PCR扩增模板。巢式PCR第2轮体系(20 μL):巢式PCR第2轮上下游引物(10 μmol·L−1)各0.2 μL,2×Taq Plus Master Mix Ⅱ(Dye Plus)10 μL,第1轮稀释后的PCR扩增产物2 μL,ddH2O补足至20 μL。PCR程序同1.4.2,PCR产物通过2%的凝胶电泳检测。
-
从田间带病棉籽中随机抽取2份,每份400颗棉籽,一份使用建立的巢式PCR方法检测,另一份使用NP-10培养基培养,棉籽先浸泡在75%的乙醇中消毒1 min,然后用无菌水泡洗3次,每次30 s,用灭菌的滤纸吸干棉籽表面的水分,最后在每个NP-10培养基上放置5颗棉籽,28 ℃培养7~10 d,与阳性对照对比观察形态,从而确定培养基培养出大丽轮枝菌的棉籽颗粒数。最后计算两种方法的棉籽带菌率并比较。NP-10培养基由两部分组成。第一部分:PGA(P-3889)5 g,0.025 mol·L−1的氢氧化钠,蒸馏水定容至500 mL,121 ℃下高压灭菌15 min。第二部分:乳脂15 g,硝酸钾1 g,氯化钾1 g,七水合硫酸镁0.5 g,壬基酚聚氧乙烯醚(NP-10)0.5 mL,蒸馏水定容至500 mL,121 ℃下高压灭菌15 min,冷却至50 ℃时,加入50 mg·L−1的氯霉素、硫酸链霉素、盐酸金霉素。待第一部分也冷却至50 ℃时,将两部分混合。
-
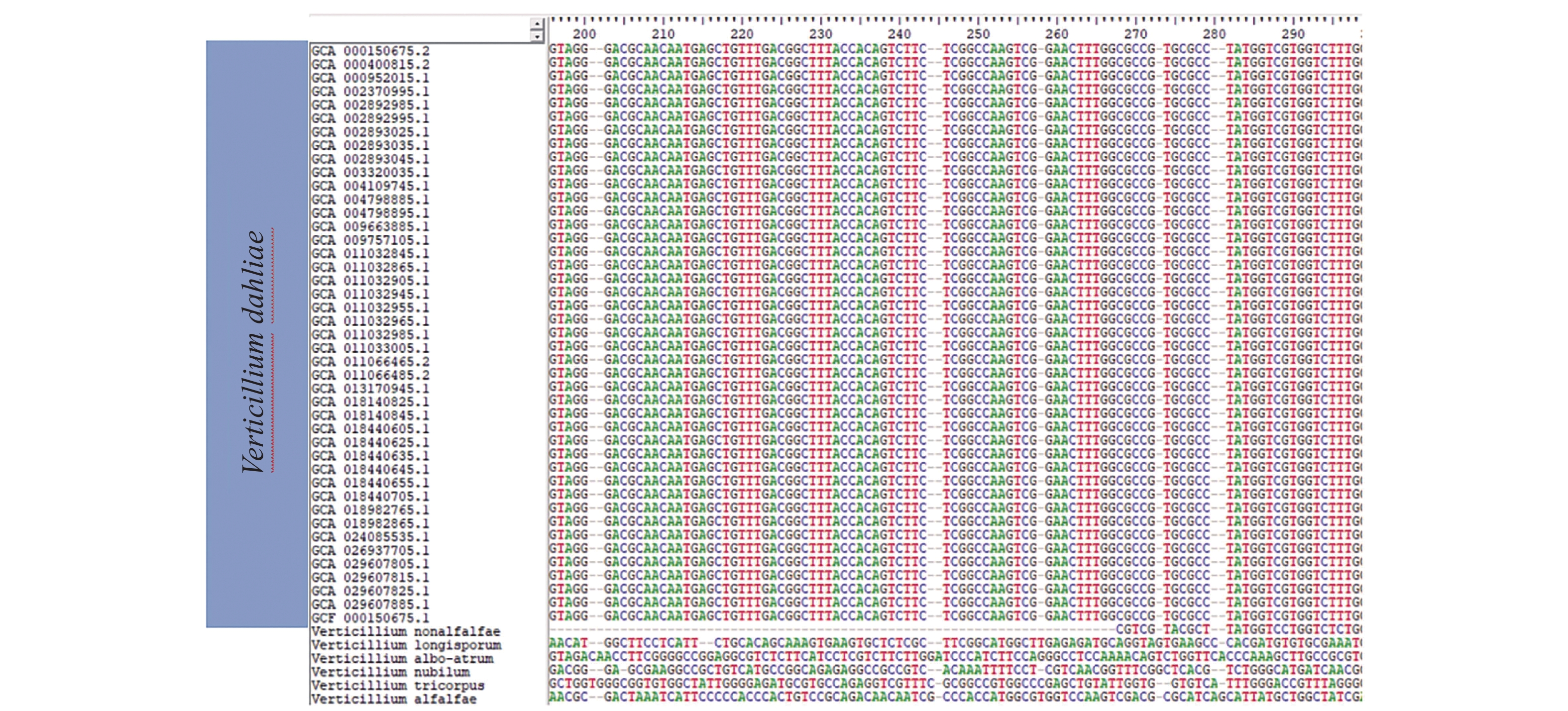
目前国内外用于大丽轮枝菌快速检测的靶标序列主要为ITS、IGS、Gpd基因和ef-1α基因等序列,笔者将这些靶标序列在NCBI数据库进行BLAST比对,发现与部分大丽轮枝菌的亲缘种有高度相似性,说明这些序列的特异性不足以区分大丽轮枝菌近源种。笔者通过大量筛选发现了一段916 bp的序列,利用BIO EDIT与NCBI数据库中已上传的43个大丽轮枝菌菌株全基因组序列进行比对,发现该序列在大丽轮枝菌不同菌株间高度保守,使用NCBI BLAST进行分析,发现该序列种间高度特异,且其中178 bp(CP010981.1:12 621~12 798)序列为大丽轮枝菌特有(图1)。
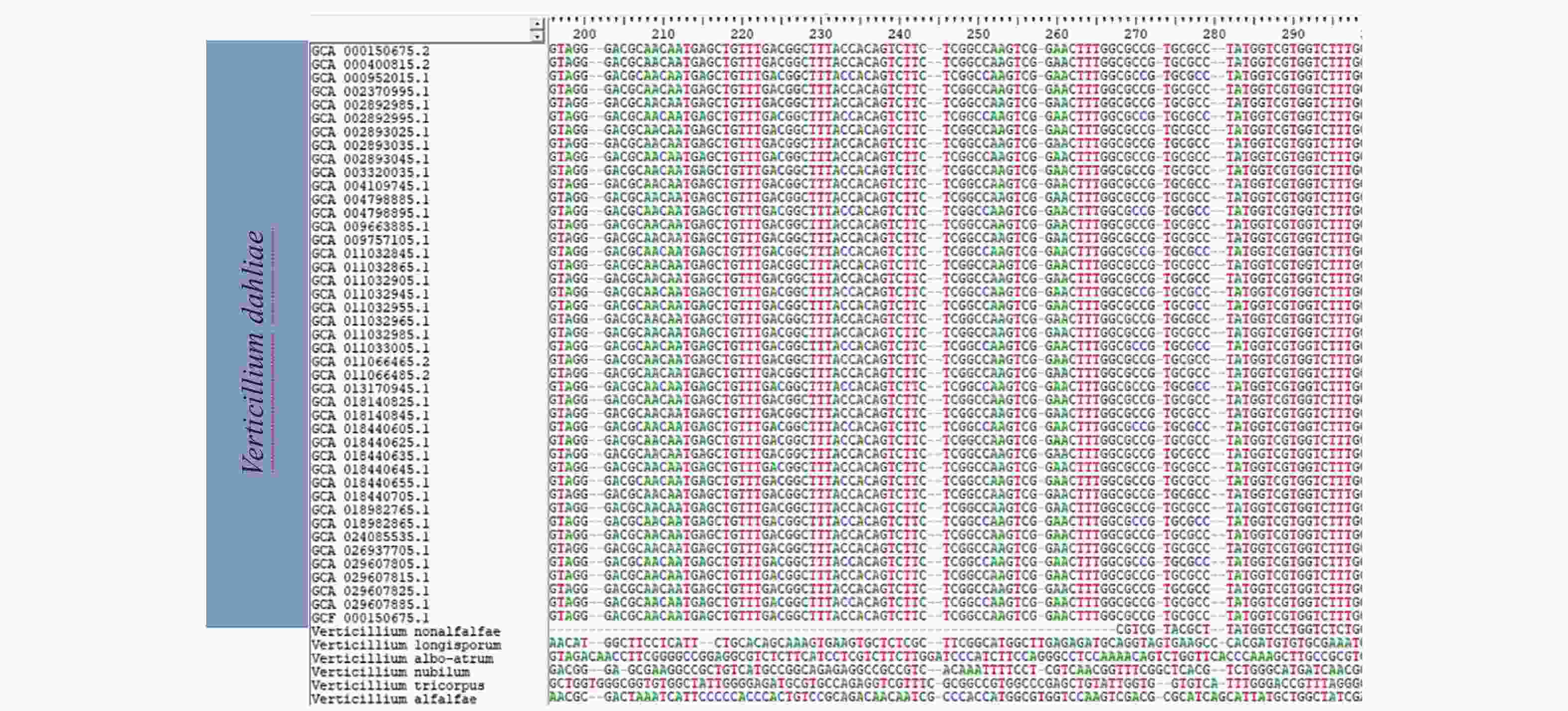
图 1 大丽轮枝菌候选DNA序列比对(仅展示部分)
Figure 1. Candidate DNA sequence alignment of Verticillium dahliae (only show part)
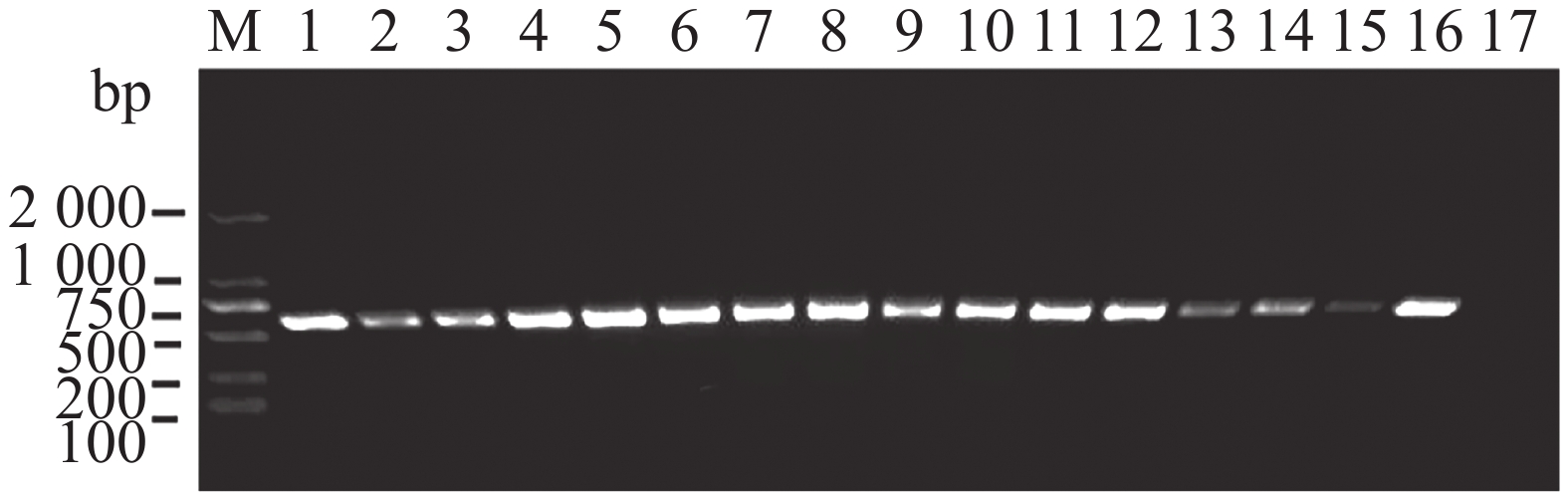
分别以来自新疆和江苏的16株大丽轮枝菌菌株基因组DNA作为模板,使用设计的引物VD-1进行扩增,PCR产物通过2%的凝胶电泳检测,结果表明16株大丽轮枝菌菌株均可以扩增出清晰的目的条带(图2),证明该基因片段在大丽轮枝菌种内具有保守性。

图 2 PCR检测16株大丽轮枝菌
Figure 2. PCR detection of 16 strains of Verticillium dahliae
-
将大丽轮枝菌起始质量浓度为1 ng·μL−1的基因组DNA梯度稀释后作为模板,用设计的4对第1轮PCR引物(VD-1、VD-2、VD-3、VD-4)进行扩增,结果发现4对引物中VD-2引物对灵敏度最高,当1 ng·μL−1的大丽轮枝菌基因组DNA作为模板时,目的条带清晰,模板浓度为1×10−4 ng·μL−1时,目的条带消失,表明第1轮PCR引物对VD-2的检测灵敏度为1×10−3 ng·μL−1的大丽轮枝菌基因组DNA(图3),其余引物对灵敏度稍低,其中引物对VD-1在DNA浓度为1 ng·μL−1时也无法检测到;引物VD-3与VD-4的检测灵敏度均为1×10−1 ng·μL−1。

图 3 常规PCR检测大丽轮枝菌的灵敏度
Figure 3. Sensitivity of conventional PCR for detection of Verticillium dahliae
-
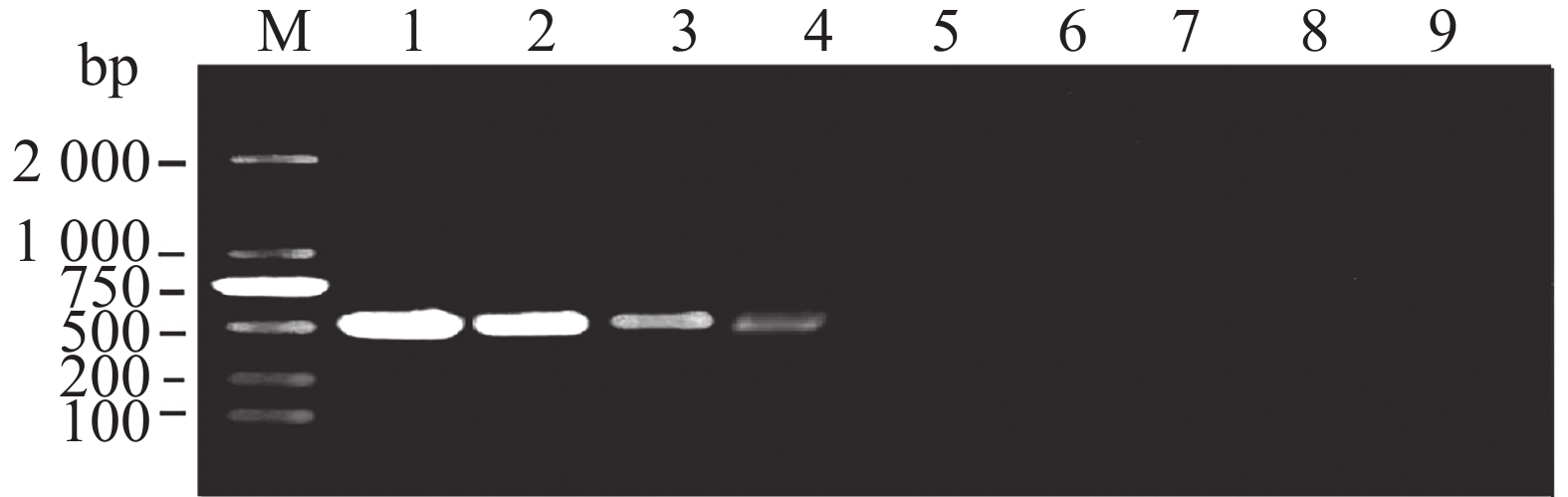
以大丽轮枝菌基因组DNA作为阳性对照,分别以包括长孢轮枝菌在内的6种大丽轮枝菌亲缘种基因组DNA以及立枯丝核菌、哈茨木霉、禾谷镰刀菌等9种植物病原菌基因组DNA以及ddH2O作为模板,利用设计的巢氏PCR第1轮引物进行扩增,验证第1轮引物的特异性,优先使用灵敏度最高的VD-2引物扩增。结果表明只有以大丽轮枝菌基因组DNA为模板的样品在512 bp位置扩增出单一条带,说明该巢氏PCR第1轮引物(VD-2)已具有高特异性(图4、5)。

图 4 常规PCR检测大丽轮枝菌的特异性
Figure 4. The specificity of conventional PCR for detection of Verticillium dahliae

图 5 常规PCR检测大丽轮枝菌的特异性
Figure 5. Specificity of Verticillium dahliae detected by conventional PCR
-
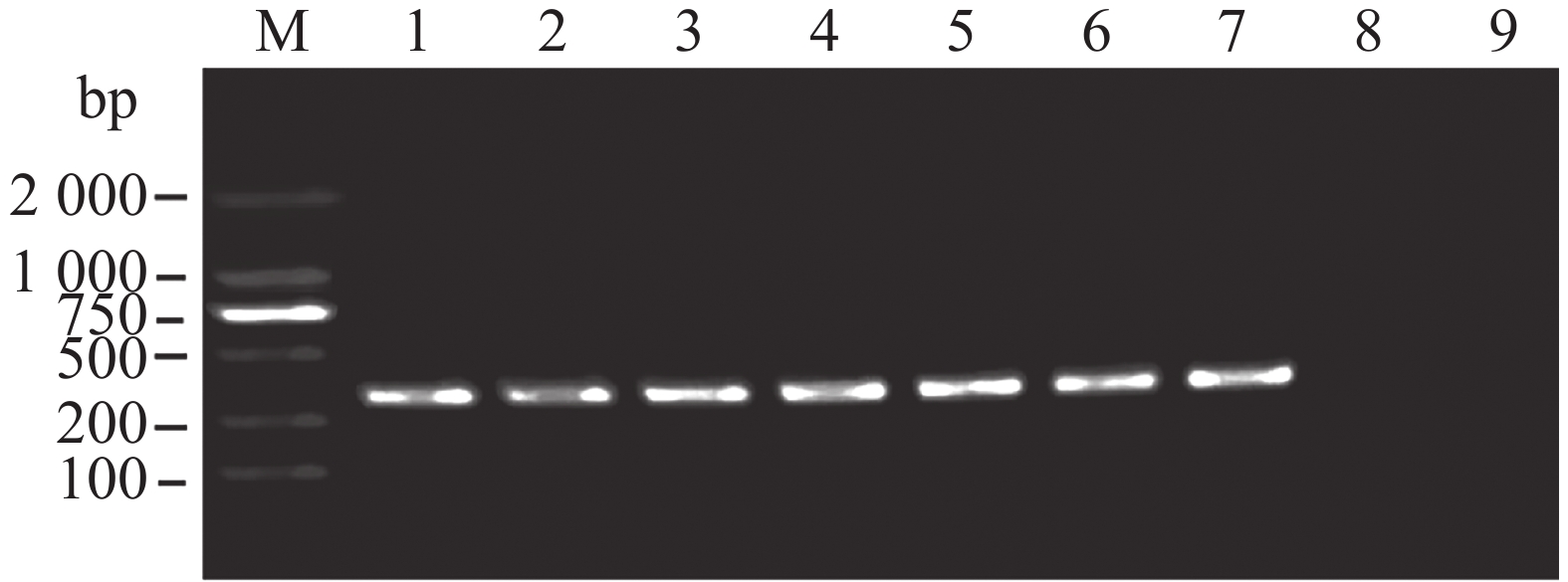
仍然以提取的大丽轮枝菌及其他15种供试菌的基因组DNA的巢氏PCR第1轮扩增产物稀释1 000倍后作为模板,利用设计的2对巢氏PCR第2轮引物(VD-5、VD-6)进行扩增,验证引物组的特异性。发现结合引物对VD-6时,虽然只有阳性对照有目的条带,但也出现了较多杂带,而结合引物对VD-5时,结果表明,仅大丽轮枝菌基因组DNA在375 bp位置扩增出单一条带(图6、7),证明经巢氏PCR的两轮扩增后,仍具有高度特异性,可以满足大丽轮枝菌的检测要求。

图 6 巢式PCR检测大丽轮枝菌的特异性
Figure 6. The specificity of nested PCR for detection of Verticillium dahliae

图 7 巢式PCR检测大丽轮枝菌的特异性
Figure 7. The specificity of nested PCR for detection of Verticillium dahliae
-
将第1轮巢氏PCR产物稀释1 000倍后作为模板,用第2轮引物进行巢氏PCR扩增。结果表明1×10−6~1 ng·μL−1的基因组DNA作为模板时,目的条带明显,模板浓度稀释到1×10−7 ng·μL−1时,目的条带消失,表明经巢氏PCR扩增后的检测灵敏度为1×10−6 ng·μL−1的大丽轮枝菌基因组DNA(图8),比只使用第1轮引物扩增的灵敏度提高了1 000倍,证明本研究的巢氏PCR引物具有高灵敏度。
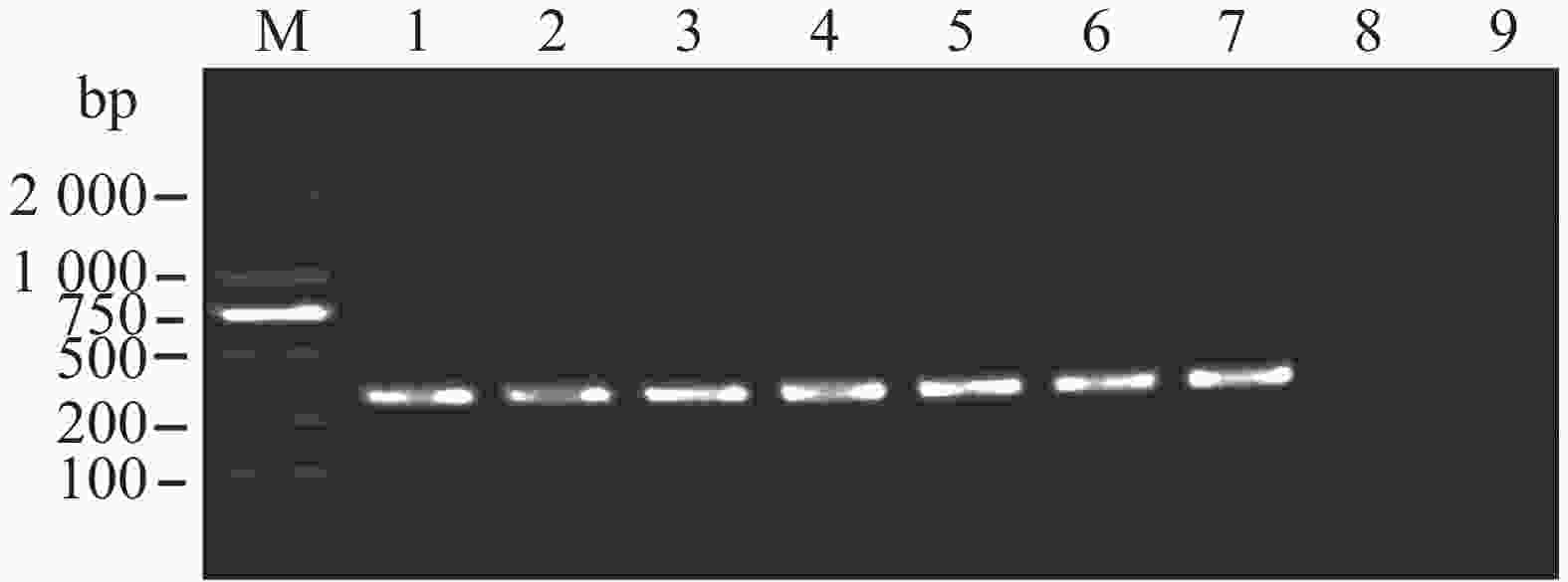
图 8 巢氏PCR检测大丽轮枝菌的灵敏度
Figure 8. Sensitivity of nested PCR for detection of Verticillium dahliae
-
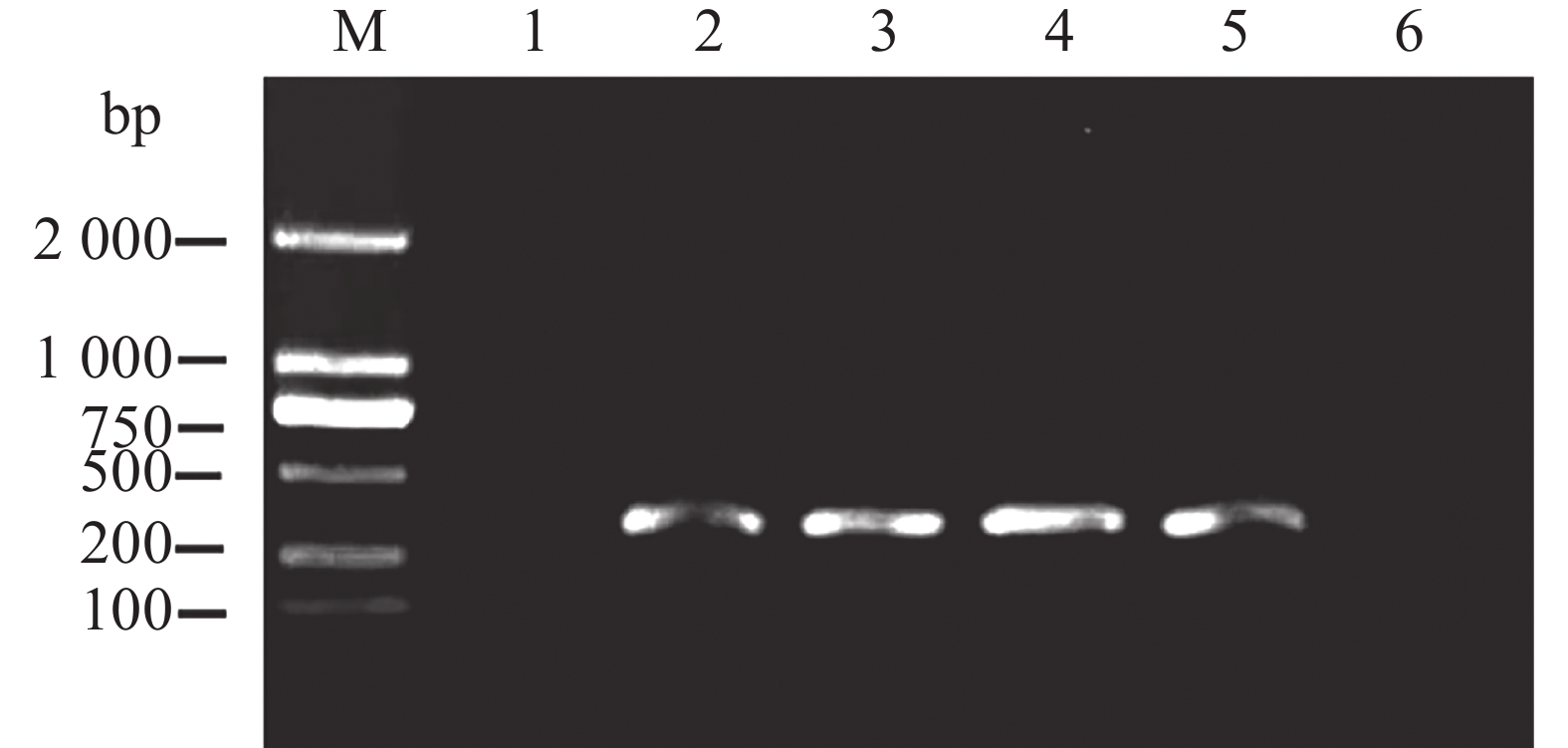
为了验证本方法能否用于检测带菌棉籽,从未发生棉花黄萎病的棉田中采集一批棉籽,约3 000 颗。按照GB15569中对于种子抽样的标准,随机抽取200个棉籽,提取总DNA后,使用上述巢氏PCR引物组扩增,产物经凝胶电泳观察后,均未产生目的条带,说明该批棉籽均不携带棉花黄萎病病原菌大丽轮枝菌。用不同浓度大丽轮枝菌孢子与棉籽充分混匀,提取带菌棉籽DNA。用上述巢氏PCR进行检测,结果表明,经第2轮巢氏PCR扩增后,扩增条带单一且检测灵敏度为每颗棉籽里混入10个孢子(图9),可以满足棉籽携带大丽轮枝菌的检测要求。
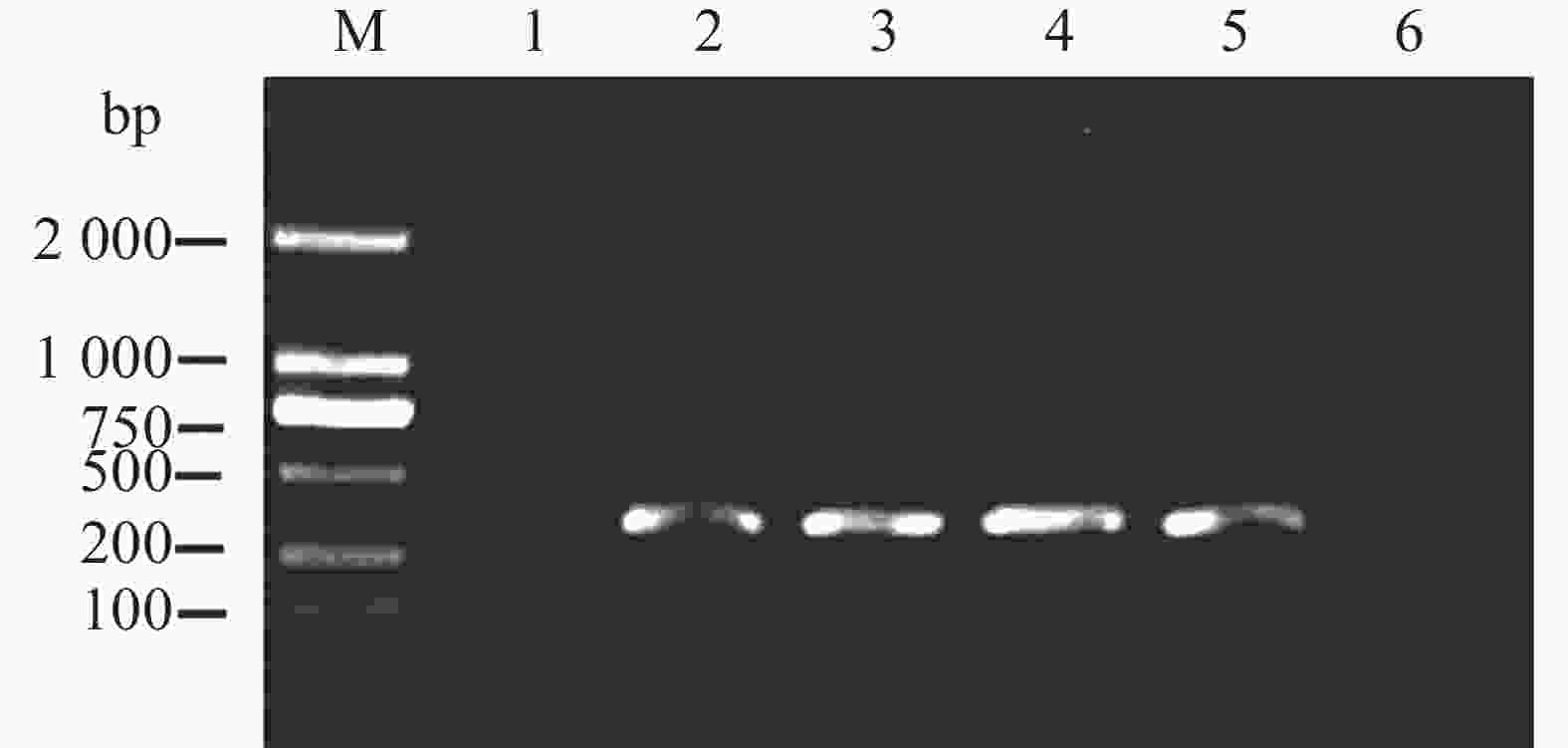
图 9 巢氏PCR检测混入不同孢子数量的棉籽DNA
Figure 9. Detection of cotton seed DNA mixed with different number of spores by nested PCR
-
从田间有疑似黄萎病的棉花植株上摘取一批棉籽作为检测样本,从这批棉籽中随机抽取400颗棉籽,提取总DNA后,用建立的巢式PCR检测方法检测,结果从该400颗棉籽中检测到共246颗棉籽携带棉花黄萎病大丽轮枝菌,检出率为61.5%。另外随机抽取的400颗棉籽放在NP-10培养基上培养,培养出大丽轮枝菌的棉籽共163颗,检出率为40.75%。
-
目前在中国,检测棉花黄萎病带菌棉籽,主要依据中华人民共和国国家标准GB/T 28084-2011及中华人民共和国出入境检验检疫行业标准SN/T 5138—2019,两种标准中检测带菌棉籽的方法都需要先分离培养,然后进行形态学及分子学鉴定,这些方法耗时长(需要10 d以上),检测灵敏度低,操作复杂且需要操作人员具有一定的专业知识。为了能够缩短时间、简化步骤以及节约成本,需要开创一种能够快速、微量检测棉籽是否携带大丽轮枝菌的方法,从而快速预防和阻遏棉花黄萎病菌的远距离传播,增加棉花作物产量,推动经济发展。巢氏PCR因为成本低、操作简单、特异性强及灵敏度高等优点,适合用于棉籽带菌的分子检测。现已有大量研究者以大丽轮枝菌的基因内间隔区(intergenic spacer)、Gpd(glyceraldehyde-3-phosphate dehydrogenase,甘油醛-3-磷酸脱氢酶)基因、β-微管蛋白基因和转录内间隔区(internal scribed spacer)等设计引物检测土壤中大丽轮枝菌,但这些引物基于的片段序列,均与大丽轮枝菌的部分亲缘种有不同程度的重叠。
笔者找到1条916 bp的基因序列,通过大量比对后在差异位点设计了1组特异性巢氏PCR引物,该方法特异性强,比常规PCR灵敏度提高了1 000倍且成本低、操作简单,适合用于棉籽携带棉花黄萎病大丽轮枝菌的快速检测,成功从田间发病棉籽中检测到了大丽轮枝菌,且灵敏度高于常规使用培养基培养的方法。最终建立了针对棉籽棉花黄萎病菌大丽轮枝菌的快速分子检测技术。
大丽轮枝菌巢氏PCR检测方法的建立及在棉籽上的应用
DOI: 10.15886/j.cnki.rdswxb.20240022
 CSTR: 32425.14.j.cnki.rdswxb.20240022
CSTR: 32425.14.j.cnki.rdswxb.20240022
Establishment of nested PCR detection method for Verticillium dahliae and its application in pathogen detection of cotton seeds
-
摘要: 棉花黄萎病是棉花上的重要病害,严重威胁棉花产业。中国棉花黄萎病病原为大丽轮枝菌(Verticillium dahliae)。棉花黄萎病防治困难,建立快速准确的分子检测方法对于该病的早期诊断、预警和防治具有重要意义。通过NCBI BLAST进行序列比对,找到大丽轮枝菌种内具有高度种间特异性和种内保守性的物种特异性序列,根据序列设计6对引物,筛选出特异性强、灵敏度高可用于检测大丽轮枝菌的巢氏PCR引物,并摸索最佳扩增条件。筛选到了两组巢氏PCR引物分别是VD-2和VD-5,扩增条件为:95 ℃ 5 min;95 ℃ 30 s;58 ℃ 30 s;72 ℃ 1 min,35个循环;72 ℃ 10 min,条带大小分别为512 和375 bp。该方法可检测到的病菌DNA下限为1×10−6 ng·μL−1,比常规PCR灵敏度提高了3个数量级,使用包括长孢轮枝菌在内的15种菌对引物特异性进行了验证,检测混有大丽轮枝菌孢子的棉籽发现该方法的检测下限是每颗棉籽可达10个孢子。Abstract: Cotton verticillium wilt is a destructive disease that seriously affects the cotton industry. Verticillium dahliae is the main causing agent of cotton verticillium wilt in Chin. It is difficult to effectively controll this disease through conventional methods, and it hence is of great significance to establish a rapid and efficient detection method for early monitoring, diagnosis, and controlling of this disease. A highly conserved DNA sequence among strains of V. dahliae and a high divergence among different species were discovered by using NCBI BLAST. Based on this DNA segment, six pairs of PCR primers were designed and one set of primers that are highly specific and sensitive were eventually selected for detecting V. dahliae through nested PCR. And the optimal amplification conditions for these primers were determined. A set of nested PCR primers were identified, namely V.d-2 and V.d-5. The amplification conditions are as follows: 95 ℃ for 5 minutes, 95 ℃ for 30 seconds, 58 ℃ for 30 seconds, 72 ℃ for 1 minute, performed 35 cycles. The resulting bands are of sizes 512 bp and 375 bp, respectively. Resulting nested PCR can be used to detect the lowest amount of pathogenic DNA at 1×10−6 ng·μL−1, which is 3 orders of magnitude higher in sensitivity compared to the conventional PCR. The primer specifity was verified by using 15 species of pathogens such as Verticillium longisporum, etc. The lowest number of 10 spores each conton seed bearing pathogen of V. dahliae can be detected by using this method.
-
Key words:
- cotton seed /
- cotton verticillium wilt /
- Verticillium dahliae /
- nested PCR /
- molecular detection
-
图 2 PCR检测16株大丽轮枝菌
M:DL2000 DNA marker;1~16:不同大丽轮枝菌菌株的DNA;17:ddH2O。
Fig. 2 PCR detection of 16 strains of Verticillium dahliae
M: DL2000 DNA marker;lanes 1-16: DNA of different V. dahliae strains;lane 17: ddH2O.
图 3 常规PCR检测大丽轮枝菌的灵敏度
M:DL2000;1:1 ng·μL−1;2:1×10−1 ng·μL−1;3:1×10−2 ng·μL−1;4:1×10−3 ng·μL−1;5:1×10−4 ng·μL−1;6:1×10−5 ng·μL−1;7:1×10−6 ng·μL−1;8:1×10−7 ng·μL−1;9:ddH2O。
Fig. 3 Sensitivity of conventional PCR for detection of Verticillium dahliae
M: DL2000;lane 1: 1 ng·μL−1;lane 2: 1×10−1 ng·μL−1;lane 3: 1×10−2 ng·μL−1;lane 4: 1×10−3 ng·μL−1;lane 5: 1×10−4 ng·μL−1;lane 6: 1×10−5 ng·μL−1;lane 7: 1×10−6 ng·μL−1;lane 8: 1×10−7 ng·μL−1;lane 9: ddH2O.
图 4 常规PCR检测大丽轮枝菌的特异性
M:DL2000;1:大丽轮枝菌基因组DNA;2:长孢轮枝菌基因组DNA;3:非苜蓿轮枝菌基因组DNA;4:变黑轮枝菌基因组DNA;5:黑白轮枝菌基因组DNA;6:苜蓿轮枝菌基因组DNA;7:云状轮枝菌基因组DNA;8:ddH2O。
Fig. 4 The specificity of conventional PCR for detection of Verticillium dahliae
M: DL2000;lane 1: Genomic DNA of V. dahliae;lane 2: Genomic DNA of V. longisporum;lane 3: Genomic DNA of V. nonalfalfae;lane 4: Genomic DNA of V. nigrescens;lane 5: Genomic DNA of V. albo-atrum;lane 6: Genomic DNA of V. alfalfae;lane 7: Genomic DNA of V. nubilum;lane 8: ddH2O.
图 5 常规PCR检测大丽轮枝菌的特异性
M:DL2000;1:大丽轮枝菌基因组DNA;2:立枯丝核菌基因组DNA;3:哈茨木霉基因组DNA;4:禾谷镰刀菌基因组DNA;5:辣椒疫霉基因组DNA;6:棉花角斑病菌基因组DNA;7:大豆疫霉基因组DNA;8:棘孢木霉基因组DNA;9:尖孢镰刀菌(FOC)基因组DNA;10:稻瘟菌基因组 DNA;11: ddH2O。
Fig. 5 Specificity of Verticillium dahliae detected by conventional PCR
M: DL2000;lane 1: Genomic DNA of V. dahliae ;lane 2: Genomic DNA of R. solani;lane 3: Genomic DNA of T. harzianum;lane 4: Genomic DNA of F. graminearum;lane 5: Genomic DNA of P. capsici;lane 6: Genomic DNA of XCM;lane 7: Genomic DNA of P. sojae;lane 8: Genomic DNA of T. asperellum;lane 9: Genomic DNA of FOC;lane 10: Genomic DNA of M. oryzae; lane 11: ddH2O.
图 6 巢式PCR检测大丽轮枝菌的特异性
M:DL2000;1:大丽轮枝菌基因组DNA;2:长孢轮枝菌基因组DNA;3:非苜蓿轮枝菌基因组DNA;4:变黑轮枝菌基因组DNA;5:黑白轮枝菌基因组DNA;6:苜蓿轮枝菌基因组DNA;7:云状轮枝菌基因组DNA;8:ddH2O。
Fig. 6 The specificity of nested PCR for detection of Verticillium dahliae
M: DL2000;lane 1: Genomic DNA of V. dahliae;lane 2: Genomic DNA of V. longisporum;lane 3: Genomic DNA of V. nonalfalfae;lane 4: Genomic DNA of V. nigrescens;lane 5: Genomic DNA of V. albo-atrum;lane 6: Genomic DNA of V. alfalfae;lane 7: Genomic DNA of V. nubilum;lane 8.ddH2O.
图 7 巢式PCR检测大丽轮枝菌的特异性
M:DL2000;1:大丽轮枝菌基因组DNA;2:立枯丝核菌基因组DNA;3:哈茨木霉基因组DNA;4:禾谷镰刀菌基因组DNA;5:辣椒疫霉基因组DNA;6:棉花角斑病菌基因组DNA;7:大豆疫霉基因组DNA;8:棘孢木霉基因组DNA;9:尖孢镰刀菌基因组DNA;10:稻瘟菌基因组DNA;11:ddH2O。
Fig. 7 The specificity of nested PCR for detection of Verticillium dahliae
M: DL2000;lane 1: Genomic DNA of V. dahliae;lane 2: Genomic DNA of R. solani;lane 3: Genomic DNA of T. harzianum;lane 4: Genomic DNA of F. graminearum;lane 5: Genomic DNA of P. capsici;lane 6: Genomic DNA of XCM;lane 7: Genomic DNA of P. sojae;lane 8: Genomic DNA of T. asperellum;lane 9: Genomic DNA of FO ;lane 10: Genomic DNA of M. oryzae: lane 11: ddH2O.
图 8 巢氏PCR检测大丽轮枝菌的灵敏度
M:DL2000;1:1 ng·μL−1;2:1×10−1 ng·μL−1;3:1×10−2 ng·μL−1;4:1×10−3 ng·μL−1;5:1×10−4 ng·μL−1;6:1×10−5 ng·μL−1;7:1×10−6 ng·μL−1;8:1×10−7 ng·μL−1;9:ddH2O。
Fig. 8 Sensitivity of nested PCR for detection of Verticillium dahliae
M: DL2000;lane 1: 1 ng·μL−1;lane 2: 1×10−1 ng·μL−1;lane 3: 1×10−2 ng·μL−1;lane 4: 1×10−3 ng·μL−1;lane 5: 1×10−4 ng·μL−1;lane 6: 1×10−5 ng·μL−1;lane 7: 1×10−6 ng·μL−1;lane 8: 1×10−7 ng·μL−1;lane 9: ddH2O.
图 9 巢氏PCR检测混入不同孢子数量的棉籽DNA
1:0个孢子;2:10个孢子;3:100个孢子;4:1 000个孢子;5:10 000个孢子;6:ddH2O。
Fig. 9 Detection of cotton seed DNA mixed with different number of spores by nested PCR
Lane 1: 0 spores;lane 2: 10 spores;lane 3: 100 spores;lane 4: 1 000 spores;lane 5: 10 000 spores;lane 6: ddH2O.
表 1 供试菌株
Table 1 Pathogens under test
种名
Species分离材料
Separation materials来源
Source数量/株
Number/strain培养
Medium大丽轮枝菌(Verticillium dahliae) 棉花(Gossypium hirsutum) 新疆
江苏16 PDA 长孢轮枝菌(Verticillium longisporum) 萝卜(Raphanus sativus) 青海 1 PDA 非苜蓿轮枝菌(Verticillium nonalfalfae) — — 1 PDA 变黑轮枝菌(Verticillium nigrescens) — — 1 PDA 黑白轮枝菌(Verticillium albo-atrum) — — 1 PDA 苜蓿轮枝菌(Verticillium alfalfae) — — 1 PDA 云状轮枝菌(Verticillium nubilum) — — 1 PDA 立枯丝核菌(Rhizoctonia solani) — 河南 1 PDA 哈茨木霉(Trichoderma harzianum) 土壤 Soil — 1 PDA 禾谷镰刀菌(Fusarium graminearum) 小麦(Triticum aestivum) — 1 PDA 辣椒疫霉(Phytophthora capsici) 辣椒(Capsicum annuum) — 1 V8 棉花角斑病黄单胞菌(Xanthmonas citri subsp. malvacearum) 棉花(Gossypium hirsutum) — 1 NA 大豆疫霉(Phytophthora sojae) 大豆(Glycine max) — 1 V8 棘孢木霉(Trichoderma asperellum) 土壤 Soil — 1 PDA 尖孢镰刀菌(Fusarium oxysporum f. sp. cubense) 香蕉(Musa nana) — 1 PDA 稻瘟菌(Magnaporthe oryzae) 水稻(Oryza sativa) — 1 PDA 注:表中“—”表示分离材料或来源未知。 Note: In the table “—” indicates that the separation material or source is unknown.  下载: 导出CSV
下载: 导出CSV
表 2 巢式PCR供试引物表
Table 2 Nested PCR primer
引物名称
Primer序列(5′-3′)
Sequence(5′-3′)PCR产物长度/bp
Length/bp引物名称
Primer序列(5′-3′)
Sequence(5′-3′)PCR产物长度/bp
Length/bpVD-1F ATGAAACGTAAGGTAGGTAGC 601 VD-4F GCCAATCGTGACTGCCTCTT 465 VD-1R CCAGAGCAGCGCCTCATTCGG VD-4R GCGCAAACTTGACCAACAGC VD-2F GCGGAAGGGGCACTAGTAAT 512 VD-5F CTCGGCCAAGTCGGAACTTT 375 VD-2R CGCGCAAACTTGACCAACAG VD-5R CGCGCAAACTTGACCAACA VD-3F GTAGGGACGCAACAATGAGC 410 VD-6F CGCCTATGGTCGTGGTCTTT 170 VD-3R) TGACCAACAGCAGCCAGATC VD-6R GTCGGTGAGCTATGCCAGAA
下载: 导出CSV
-
[1] BHAT R G, SUBBARAO K V. Host range specificity in Verticillium dahliae[J]. Phytopathology, 1999, 89(12): 1218 − 1225. doi: 10.1094/PHYTO.1999.89.12.1218 [2] 林玲, 张昕, 邓晟. 棉花黄萎病研究进展[J]. 棉花学报, 2014, 26(3): 260 − 267. doi: 10.3969/j.issn.1002-7807.2014.03.010 [3] 殷锡圣, 刘润进. 棉花黄萎病研究进展[J]. 中国棉花, 1996, 23(5): 2 − 6. [4] 李廷刚, 巩东营, 张倩倩. 棉花抗黄萎病遗传学研究进展[J]. 农学学报, 2022, 12(9): 31 − 36. doi: 10.11923/j.issn.2095-4050.2022.9.zgncxkkj202209006 [5] 喻树迅, 张雷, 冯文娟. 棉花生产规模化、机械化、信息化、智能化和社会服务化发展战略研究[J]. 中国工程科学, 2016, 18(1): 137 − 148. [6] 田擎, 张海峰, 曾丹丹, 等. 环介导等温扩增技术检测大丽轮枝菌[J]. 植物病理学报, 2016, 46(6): 721 − 729. [7] 朱荷琴, 李志芳, 冯自力, 等. 我国棉花黄萎病研究十年回顾及展望[J]. 棉花学报, 2017, 29(S1): 37 − 50. [8] 沈林波, 吴楠楠, 冯小艳, 等. 我国蔗区甘蔗宿根矮化病发生情况的分子检测[J]. 热带生物学报, 2019, 10(4): 314 − 318. [9] 杜艳楠, 王萌, 马建强, 等. 植物病原真菌早期检测技术及其在橡胶树炭疽病预测预报中的应用[J]. 热带生物学报, 2021, 12(1): 124 − 131. [10] BAO J, CHEN Y, XING Y, et al. Development of a nested PCR assay for specific detection of Metschnikowia bicuspidata infecting Eriocheir sinensis[J]. Frontiers in Cellular and Infection Microbiology, 2022, 12: 930585. doi: 10.3389/fcimb.2022.930585 [11] 许晓琳, 袁向芬, 孔玉方, 等. 猴痘病毒实时荧光LAMP检测方法的建立[J]. 黑龙江畜牧兽医, 2024(2): 72 − 78. [12] ZHANG K, SUN Z, SHI K, et al. RPA-CRISPR/Cas12a-based detection of Haemophilus parasuis[J]. Animals, 2023, 13(21): 3317. doi: 10.3390/ani13213317 [13] 李园. 土壤消毒及土传病原真菌的分子生物学鉴定 [D]. 北京: 中国农业科学院, 2006. [14] MORADI A, ALMASI M A, JAFARY H, et al. A novel and rapid loop-mediated isothermal amplification assay for the specific detection of Verticillium dahliae[J]. Journal of Applied Microbiology, 2014, 116(4): 942 − 954. doi: 10.1111/jam.12407 [15] WANG D, JIAO X, JIA H, et al. Detection and quantification of Verticillium dahliae and V. longisporum by droplet digital PCR versus quantitative real-time PCR[J]. Frontiers in Cellular and Infection Microbiology, 2022, 12: 995705. doi: 10.3389/fcimb.2022.995705 [16] PÉREZ-ARTÉS E, GARCÍA-PEDRAJAS M D, BEJARANO- ALCÁZAR J, et al. Differentiation of cottondefoliating and nondefoliating pathotypes of Verticillium dahliae by RAPD and specific PCR analyses[J]. European Journal of Plant Pathology, 2000, 106(6): 507 − 517. [17] PÉREZ-ARTÉS E, MERCADO-BLANCO J, RUZCARRILLO A R, et al. Detection of the defoliating and nondefoliating pathotypes of Verticillium dahliae in artificial and natural soils by nested PCR[J]. Plant and Soil, 2005, 268(1): 349 − 356. [18] 庞莉, 李梅, 孙青, 等. 棉花黄萎病病原菌大丽轮枝菌的快速分子检测[J]. 植物保护学报, 2016, 43(6): 892 − 899. [19] WANG D, ZHANG D D, USAMI T, et al. Functional genomics and comparative lineage-specific region analyses reveal novel insights into race divergence in Verticillium dahliae[J]. Microbiology Spectrum, 2021, 9(3): e0111821. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 982
- HTML全文浏览量: 475
- PDF下载量: 11
- 被引次数: 0
