


-
2019年一项针对上海市学龄前儿童过敏性疾病的分析显示,哮喘、过敏性鼻炎、特应性皮炎的患病率依次为13.9%、18.2%、34.3%[1],而在2005年上述三类过敏性疾病的发生率仅为7.2%、12.9%、6.4%[2],由此可见,过敏反应已成为影响人们生活的主要健康问题。其中,过敏性哮喘是一种以气道炎症和气道高反应性为特征的慢性疾病,常与过敏性鼻炎共同存在,对过敏原的暴露会加重症状,严重影响患者的生活质量,并给社会带来沉重的经济负担[3]。尘螨是中国哮喘患者最常见的室内过敏原之一[4],50%以上的过敏患者和80%以上的哮喘儿童对螨虫过敏原过敏[5]。因此,由尘螨诱发的哮喘模型在模拟人类哮喘方面具有更高价值,能更有效地揭示过敏相关的免疫与应答机制。屋尘螨(Dermatophagoides pteronyssinus,Der p)过敏原Der p 1是与哮喘有关的最主要抗原[6],被称为Immunoglobulin E(IgE)抗体反应的强诱导剂[7],主要来源于屋尘螨的消化液和粪便,具有较强的抗原性,是最早在1988年通过分子克隆技术阐明其一级结构的过敏原之一[8]。屋尘螨过敏原进入呼吸道后会破坏上皮屏障,激活免疫细胞,导致大量促炎细胞因子的产生[9]。粉尘螨(Dermatophagoides farinae,Der f)也是常见螨种之一,用粉尘螨浸液与哮喘患者血清进行放射免疫吸附实验,实验结果表明约30余种成分具有特异性IgE结合力,粉尘螨主要变应原Der f 1被认为是引起变态反应性疾病最重要的变应原之一[10]。热带无爪螨(Blomia tropicalis,Blo t)又称热带螨,是热带和亚热带地区的优势螨种,其主要抗原是第5组过敏原,即Blo t 5,高于70%的热带螨过敏患者的血清IgE对其呈阳性反应[11-12]。上述三种螨类中,Der p 1、Der f 1同属Ⅰ类半胱氨酸蛋白酶家族,IgE交叉抑制实验显示二者重叠表位达42%,而Blo t 5是迄今为止研究最多的热带无爪螨过敏原,中国华南及热带地区60%~80%哮喘患儿的血清可同时识别Der p 1、Der f 1与Blo t 5,提示三者共覆盖>85%的尘螨过敏患者[13-14]。
随着尘螨过敏的临床负担日益加重,当前主流的防治手段主要包括:避免接触过敏原、药物对症控制以及过敏原特异性免疫治疗(Allergen-specific immunotherapy, AIT),但以上手段也存在诸多局限。避免接触过敏原是预防疾病发作的基础,但实际上尘螨广泛分布于各处,难以完全清除,因此该措施效果有限。抗组胺药、糖皮质激素等药物仅能暂时缓解症状,无法阻断疾病进展,长期使用还会带来副作用。AIT通过长期、规律地给予患者标准化的尘螨提取物,虽可诱导免疫耐受,但治疗周期长达3~5年[15],依从性差,且存在诱发全身过敏反应的风险,尤其在高度敏感个体中应用受限[16]。因此,迫切需要开发更安全、更高效、更精准的新型免疫治疗策略。
近年来,随着新型疫苗技术的发展,基于特定过敏原表位的疫苗设计为过敏性疾病的治疗开辟了新途径。表位疫苗设计最简单的方法是直接串联表位,但在连接处可能会形成新的拼接表位,从而掩盖对原表位的免疫反应。因此,可在表位之间添加间隔序列如连接子,使每个表位独立发挥其免疫作用,避免产生新表位。由Gly和Ser残基组成的GS linker是目前表位疫苗设计中最常用的柔性连接子之一,其免疫原性极低,不引入额外CD4+ T细胞表位[17]。PADRE(泛DR结合表位)是人工设计的通用Th细胞表位肽,能高亲和力结合多种 HLA-DR分子,增强抗原呈递,激活CD4+ Th细胞,并协助B细胞产生高滴度IgG,降低IgE,其序列无致敏性,现已用于肿瘤等多肽疫苗的设计[18]。DNA疫苗通过载体携带过敏原表位基因,可在宿主细胞内持续表达抗原,激活机体特异性细胞免疫,诱导Th1型免疫应答偏移,从源头调控免疫失衡状态[19],但单独应用时,DNA疫苗存在免疫原性较弱、抗体产生滞后等问题。蛋白疫苗则能直接提供成熟抗原,快速激发体液免疫,诱导中和抗体产生,但诱导细胞免疫和长效免疫耐受的能力可能不足[20]。复旦大学王宾教授课题组[21]利用卵清蛋白(Ovalbumin,OVA)诱导哮喘小鼠模型,发现使用OVA表位肽与编码相同表位的DNA疫苗共同免疫能够预防这种实验性哮喘,显著减少嗜酸性粒细胞对过敏原攻击部位的浸润,诱导调节性T细胞增多,缓解小鼠的气道高反应性及其他过敏症状。
基于此,本研究采用GS linker作为表位间的连接子,将PADRE置于融合表位N端,通过构建尘螨3种主要变应原融合表位的DNA疫苗和重组蛋白疫苗,以尘螨过敏小鼠模型为对象,探究DNA疫苗和蛋白疫苗联合免疫策略的治疗效果,有望为开发新型、高效的尘螨过敏性疾病免疫治疗方案提供重要的实验依据和研究思路。
-
使用4~6周龄,健康的雌性BALB/c小鼠,购自斯贝福(北京)生物技术有限公司,所有实验动物均在特定的无病原体的环境下进行饲养,饲料和垫料均为SPF(Specific Pathogen Free)级,适应性饲养1周后开始实验。所有动物实验均严格按照海南大学实验动物福利与伦理要求,并经海南大学动物伦理委员会批准(批准文号:HNUAUCC-2024-00212)。
-
大肠杆菌(Escherichia coli)DH5α感受态细胞(B528413)、BL21(DE3)感受态细胞(B528414)、ELISA酶标板(F605031)、EL-TMB显色试剂盒(C520026),甲酰胺(A501904),Evans blue(E713BA0028)均购自生工生物工程(上海)股份有限公司;小鼠免疫球蛋白E(IgE)酶联免疫检测试剂盒(H107-1-1)购自南京建成生物;金牌超量无内毒素质粒大提试剂盒(CW2104M)购自康为世纪生物科技有限公司;即用型BCA蛋白浓度测定试剂盒(25FM0200)购自赛文创新(北京)生物科技有限公司;通用型组织固定液(中性)(GP24103082084)购自武汉赛维尔生物科技有限公司;氯化乙酰甲胆碱(F2406065)购自上海阿拉丁生化科技股份有限公司;异丙基-β-D-硫代半乳糖苷(1106H054)购自北京索莱宝科技有限公司。
pET-28a-Der p 1- Der f 1-Blo t 5、pET-28a-PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP、PVAX1-PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP、PVAX1均由擎科生物全基因合成。实验中涉及的其他试剂均购自国药集团化学试剂有限公司。
-
在GenBank(
https://www.ncbi.nlm.nih.gov/genbank/ )上查找尘螨主要变应原Der p 1(U11695)、Der f 1(AB034946)、Blo t 5(U59102)的基因序列,按照Der p 1- Der f 1-Blo t 5的顺序连接,进行密码子优化后,利用Nde I和Xho I两个酶切位点将其插入到原核表达载体pET-28a上,由擎科生物全基因合成。 -
使用在线工具SVMTriP(
http://sysbio.unl.edu/SVMTriP )、ABCPred(https://webs.iiitd.edu.in/raghava/abcpred/ )、Predicted Antigenic Peptides(http://imed.med.ucm.es/Tools/antigenic.pl )等对尘螨变应原Der p 1、Der f 1、Blo t 5成熟蛋白的氨基酸序列做抗原表位预测,综合筛选出Der p 1的T1(E45–67)、T2(E94–143),Der f 1的T3(E18–31)、T4(E75–94)、T5(E206–223),Blo t 5的T6(E1–20)、T7(E76–91)、T8(E106–115),将筛选出的8个表位用GS连接子按T1~T8的顺序连接,并在N端加上通用表位PADRE序列,进行密码子优化后,利用Nde I和Xho I两个酶切位点将其插入到原核表达载体pET-28a上,及利用Hind Ⅲ和Xba Ⅰ两个酶切位点将其插入到真核表达载体PVAX1上,由擎科生物全基因合成。 -
分别取1 μL原核表达质粒pET-28a-Der p 1- Der f 1-Blo t 5、pET-28a-PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP加入到大肠杆菌BL21(DE3)感受态细胞中,冰上静置30 min,42℃金属浴热激45 s,冰上静置2 min,随后加入700 μL无抗性的LB液体培养基,于37℃,200 r·min−1恒温摇床培养1 h后,5 000 r·min−1离心1 min收集菌体,弃大部分上清,留下约300 μL重悬沉淀,取上述菌液100 μL涂布于含卡那霉素的LB固体平板,37℃倒置培养12~16 h至长出单个分散均匀的乳白色圆形菌落。
-
分别取1 μL真核表达质粒PVAX1-PADRE-GS-( Der p 1- Der f 1-Blo t 5)EP、PVAX1加入到大肠杆菌DH5α感受态细胞中,冰上静置30 min,42℃金属浴热激45 s,冰上静置2 min,随后加入700 μL无抗性的LB液体培养基,于37℃,200 r·min−1恒温摇床培养1 h后,5 000 r·min−1离心1 min收集菌体,弃大部分上清,留下约300 μL重悬沉淀,取上述菌液100 μL涂布于含卡那霉素的LB固体平板,37℃倒置培养12~16 h至长出单个分散均匀的乳白色圆形菌落。
-
挑取1.4所述的2种原核表达质粒的单克隆菌落至10 mL卡那霉素抗性LB液体培养基中,37℃,200 r·min−1过夜培养。次日早上取5 mL过夜培养的菌液加至250 mL卡那抗性LB培养基中,37℃,200 r·min−1培养至OD600为0.6~0.8时,加入终浓度为1 mmol·L−1的异丙基-β-D-硫代半乳糖苷(Isopropyl-β-D-1-Thiogalac-topyranoside,IPTG),37℃,180 r·min−1诱导表达6 h。诱导结束后收集菌体,加入25 mL包涵体洗涤缓冲液 Ⅰ 重悬菌体沉淀,采用细胞超声破碎仪在冰水浴上进行超声破碎,破碎结束后,4℃,12 000 r·min−1离心15 min收集沉淀。
包涵体蛋白的洗涤步骤为,第1次洗涤:加入25 mL洗涤缓冲液Ⅰ重悬沉淀,反复吹打至沉淀全部散开,8 000 r·min−1,4℃,离心15 min收集沉淀,重复洗涤1次;第2次洗涤:加入25 mL洗涤缓冲液Ⅱ重悬沉淀,同样条件离心收集,重复洗涤1次;第3次洗涤:加入25 mL洗涤缓冲液Ⅲ重悬沉淀,离心收集沉淀。包涵体蛋白的溶解步骤为,加入5 mL溶解缓冲液Ⅰ重悬沉淀,再加入1 mL溶解缓冲液Ⅱ,吹打混匀后,4℃,8 000 r·min−1离心15 min收集上清液。
包涵体蛋白的复性步骤为,将上清液装入透析袋中,4℃下分别在复性缓冲液Ⅰ/Ⅱ/Ⅲ中进行降梯度复性,每个梯度透析12 h,将复性后的蛋白置于PBS中透析除盐。通过SDS-PAGE检测包涵体蛋白的表达及纯化情况,并使用BCA蛋白定量法测定蛋白浓度及含量。
-
菌体活化:在10 mL含有卡那霉素的LB液体培养基中加入60 μL保存的目的甘油菌,于37℃,180 r·min−1条件下,培养4~6 h。质粒扩增:取上述已活化的菌液5 mL加入到250 mL含有卡那霉素的LB液体培养基中,于37℃,180 r·min−1的条件下,培养12~16 h。后续操作使用金牌超量无内毒素质粒大提试剂盒进行提取,提取后的质粒在超微量分光光度计测定浓度后,置于−20℃保存备用。
-
将适应性饲养1周后的BALB/c小鼠分为6组,每组6只,正常组不做任何处理,其余5组均使用Der p 1- Der f 1-Blo t 5尘螨融合变应原蛋白诱导过敏小鼠模型。造模方法:(1)腹腔致敏:在第0、7天对小鼠腹腔注射100 μg的尘螨融合变应原Der p 1- Der f 1-Blo t 5(含5 mg的氢氧化铝佐剂),变应原蛋白浓度为1 g·L-1,用量均为100 μL·只−1;(2)气管内灌入:在第14、16、18天通过给小鼠气管灌入100 μL的Der p 1- Der f 1-Blo t 5蛋白(1 g·L−1溶于PBS),对其肺部组织进行激发,共3次。上述的致敏操作中,100 μg·只−1为国内外过敏小鼠的常规、有效且可重复的注射剂量,可在BALB/c小鼠中稳定诱导血清特异性IgE水平升高及气道高反应性[21]。
气管内激发参照Mateus Casaro等[22]的方法,具体操作为:(1)麻醉小鼠后用橡皮筋挂住其门牙,使身体自然下垂并与实验台面呈90°角,确保气管不被挤压。(2)用镊子移开小鼠舌头以暴露气管区域,将20 µL毛细管平行于气管缓缓插入,使用注射器将变应原溶液通过毛细管注入,若小鼠出现呼吸急促的现象,则表明气管内滴注成功。
在气管灌入结束后的第3天,通过检测正常组和致敏组小鼠的气道高反应性变化、血清总IgE水平及分析肺部组织切片的炎症情况,评估是否成功建立尘螨诱导的过敏小鼠模型。造模流程如图1所示。
图 1 尘螨诱导过敏小鼠的造模流程图
Figure 1. Flowchart for modeling dust mite-induced allergic mice
-
对上述处理后的小鼠进行随机分组,正常组不作变动(n=6),致敏后的小鼠随机分为5组(n=6),分别是Allergy、GS-DNA、GS-Protein、PVAX1+GS-Protein、GS-DNA+GS-Protein。在第25、39天时,注射疫苗进行过敏小鼠模型的治疗,正常组(Naïve)不做任何处理,致敏组(Allergy)注射PBS,200 μL·只-1,其余组的疫苗用量为,DNA疫苗组(100 μg GS-DNA),蛋白疫苗组(100 μg GS-Protein),空载质粒/蛋白疫苗组(100 μg PVAX1+100 μg GS-Protein),联合免疫组(100 μg GS-DNA+100 μg GS-Protein),颈部皮下100 μL、腿部肌肉各50 μL进行多点注射。所有的小鼠分组及疫苗治疗的注射剂量均参照文献[21]进行设计。
在第54天时,通过检测小鼠血清总IgE水平、气道高反应性变化、耳朵染料渗漏面积及肺部组织切片结果,综合分析DNA疫苗和蛋白疫苗联合免疫对尘螨过敏小鼠模型的治疗效果。疫苗治疗流程如图2所示。

图 2 尘螨过敏小鼠模型的治疗流程图
Figure 2. Treatment flowchart for a mouse model of dust mite allergy
-
采用竞争法检测样本中IgE的含量:向预先包被了抗体的酶标孔中加入小鼠血清样本,再加入生物素标记的识别抗原,在37℃下孵育30 min,两者与固相抗体竞争结合形成免疫复合物,经PBST洗涤除去未结合的生物素抗原,然后加入亲和素-HRP,在37℃下孵育30 min,亲和素-HRP与生物素抗原结合,洗涤后结合的HRP催化TMB(四甲基联苯胺)成蓝色,随后在酸的作用下转化成黄色,在450 nm波长下有吸收峰,吸光值与样本中抗原的浓度成负相关。
-
通过FinePointe WBP全身体积描记仪,测量小鼠暴露于不同浓度的氯化乙酰甲胆碱(Methacholine Chloride,Mch)时的肺部呼吸阻力,评估致敏和疫苗治疗后各组小鼠的气道高反应性(Airway Hyperresponsiveness,AHR)变化。实验开始前,创建程序:适应时间30 min,雾化时间2 min,反应时间3 min,恢复时间2 min,雾化体积均为20 μL。待仪器装置连接好后,将小鼠置于腔体内进行适应,随后依次雾化PBS及不同浓度的Mch,Mch的浓度由低到高依次为3.125、6.25、12.5、25和50 g·L−1。
-
对小鼠尾静脉注射200 μL,0.5%的Evans blue染料,反应30 min后将小鼠麻醉,使用23G穿刺针分别在小鼠的两侧耳朵中心处进行穿刺,并滴加20 μL的尘螨融合变应原蛋白Der p 1- Der f 1-Blo t 5(500 μg·mL−1溶于PBS),1 h后麻醉处死小鼠,拍照观察耳朵染料的渗漏情况,使用Image J对染料渗漏面积进行统计。
拍照结束后,剪下小鼠的耳部组织并浸入400 μL的甲酰胺溶液中,置于63℃水浴锅温育32 h,以提取耳部组织的Evans blue染料。温育结束后取出耳部组织,每个样品吸取100 μL加至96孔板中,设置2个复孔,于620 nm波长处测量其吸光度值。
-
麻醉处死小鼠后,取出左肺于通用型组织固定液中进行固定、包埋、切片及染色。通过苏木精-伊红染色(H&E)观察肺组织中炎症细胞的浸润程度。过碘酸雪夫氏染色(PAS)用于统计支气管内的平均杯状细胞数。马松染色(Masson)用于评估肺组织的胶原纤维沉积程度。评分细则(表1)参照魏春洁等[23- 24]的评分方法制定。
表 1 肺部组织切片评分细则
Table 1. Scoring criteria for lung tissue sections
得分
ScoreH&E染色
H&E stainingPAS染色
PAS stainingMasson染色
Masson staining0 支气管周围无炎症细胞浸润
No inflammatory cell infiltration around the bronchi管腔内无杯状细胞
No goblet cells in the lumen胶原纤维沉积含量正常
Normal collagen fiber deposition content.1 支气管周围偶尔出现炎症细胞
Occasionally inflammatory cells appear around the bronchi管腔内有1~5个杯状细胞
The lumen contains 1~5 goblet cells.轻度胶原纤维沉积
Mild collagen fiber deposition.2 支气管周围环绕着1~5层炎症细胞
The bronchi are surrounded by 1~5 layers of inflammatory cells管腔内有6~20个杯状细胞
The lumen contains 6~20 goblet cells.中度胶原纤维沉积
Moderate collagen fiber deposition.3 支气管周围有超过5层炎症细胞浸润
There are more than 5 layers of inflammatory cell infiltration around the bronchi管腔内有超过20个杯状细胞
The lumen contains more than 20 goblet cells.重度胶原纤维沉积
Severe collagen fiber deposition. -
使用Graphpad Prism 9.5.0软件进行数据分析处理。多组间比较采用单因素方差(one-way ANOVA)进行分析。
-
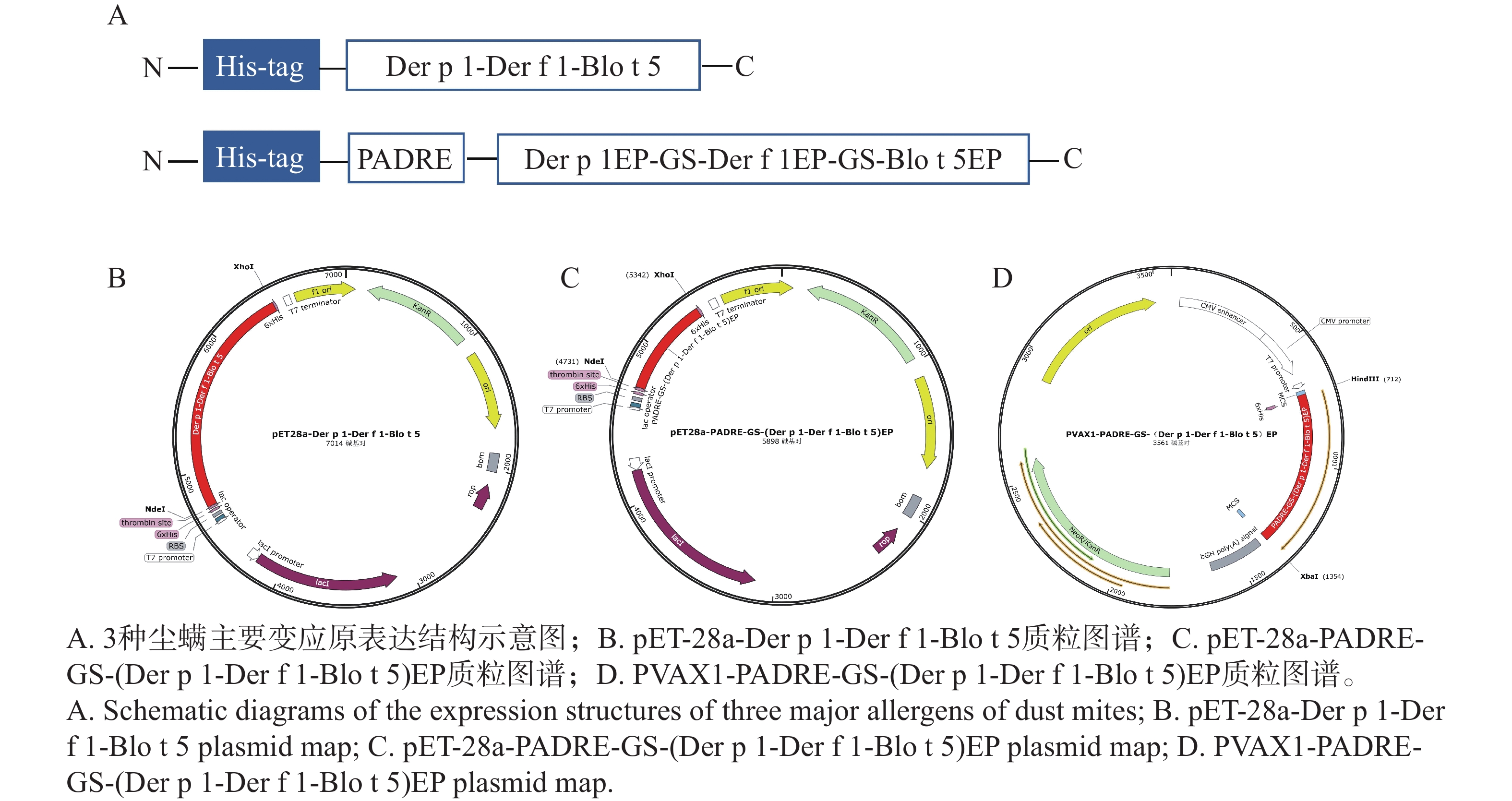
原核表达载体pET-28a-Der p 1- Der f 1-Blo t 5、pET-28a-PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP,真核表达载体PVAX1-PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP的完整质粒图谱如图3-B、C和D所示。
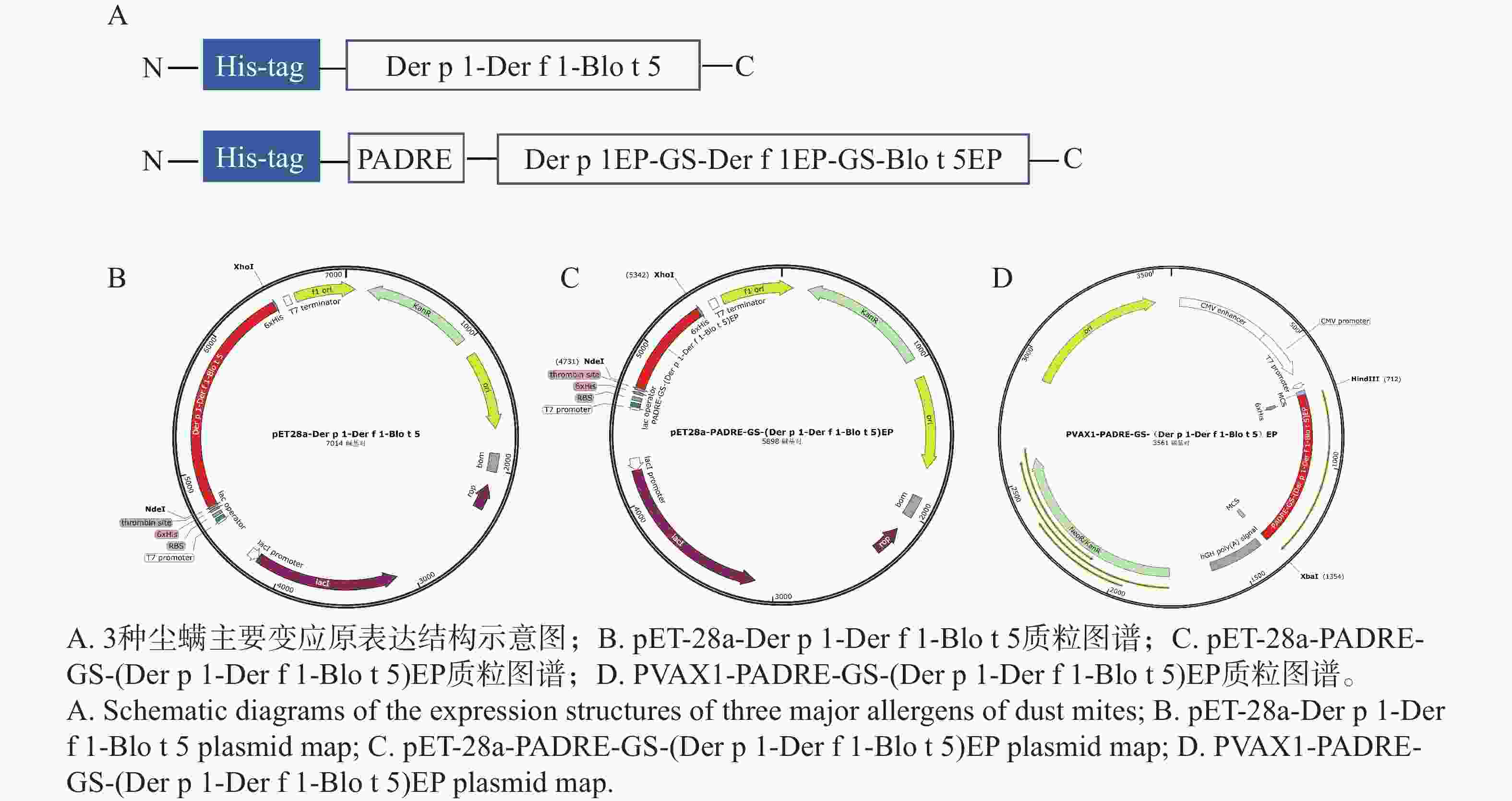
图 3 尘螨主要变应原表达载体的构建
Figure 3. Construction of the expression vector of the main allergens of dust mites
-
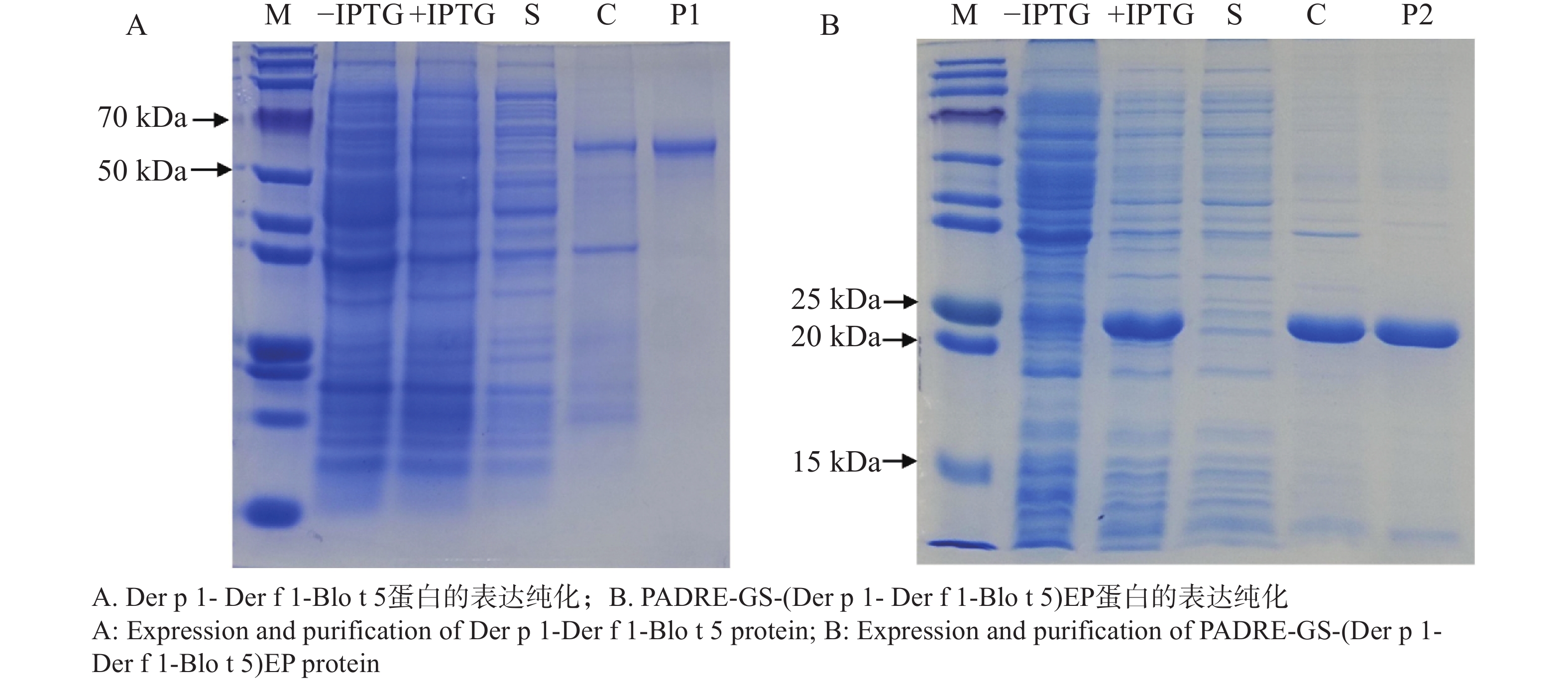
原核表达载体转化后,挑取阳性单克隆培养,经IPTG诱导后表达出重组蛋白Der p 1- Der f 1-Blo t 5、PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP。SDS-PAGE电泳显示,纯化后得到单一的目的蛋白,条带分别出现在65 kDa、22 kDa,表明成功表达并纯化得到了Der p 1- Der f 1-Blo t 5、PADRE-GS-(Der p 1- Der f 1-Blo t 5)EP重组蛋白。
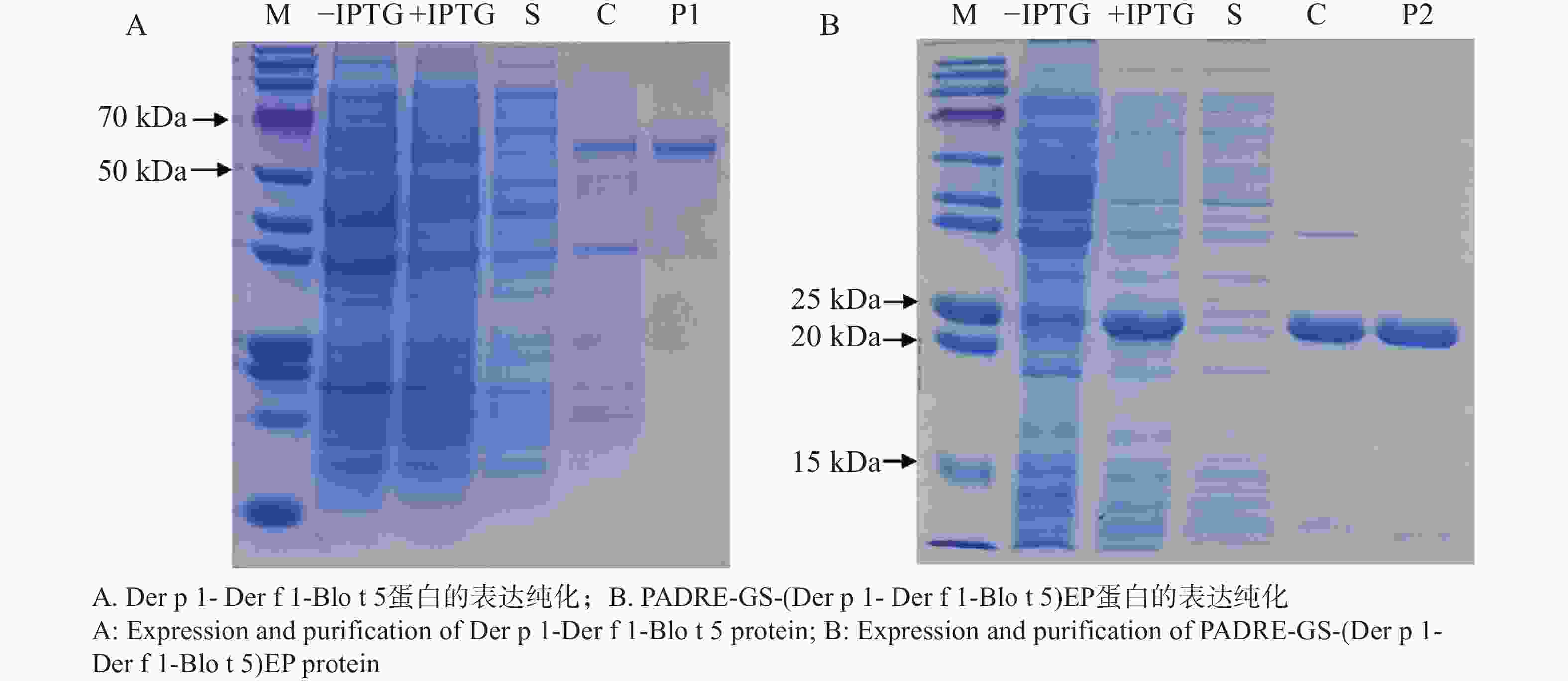
图 4 SDS-PAGE电泳检测重组蛋白的表达与纯化
Figure 4. SDS-PAGE electrophoresis for the detection of expression and purification of the recombinant protein
-
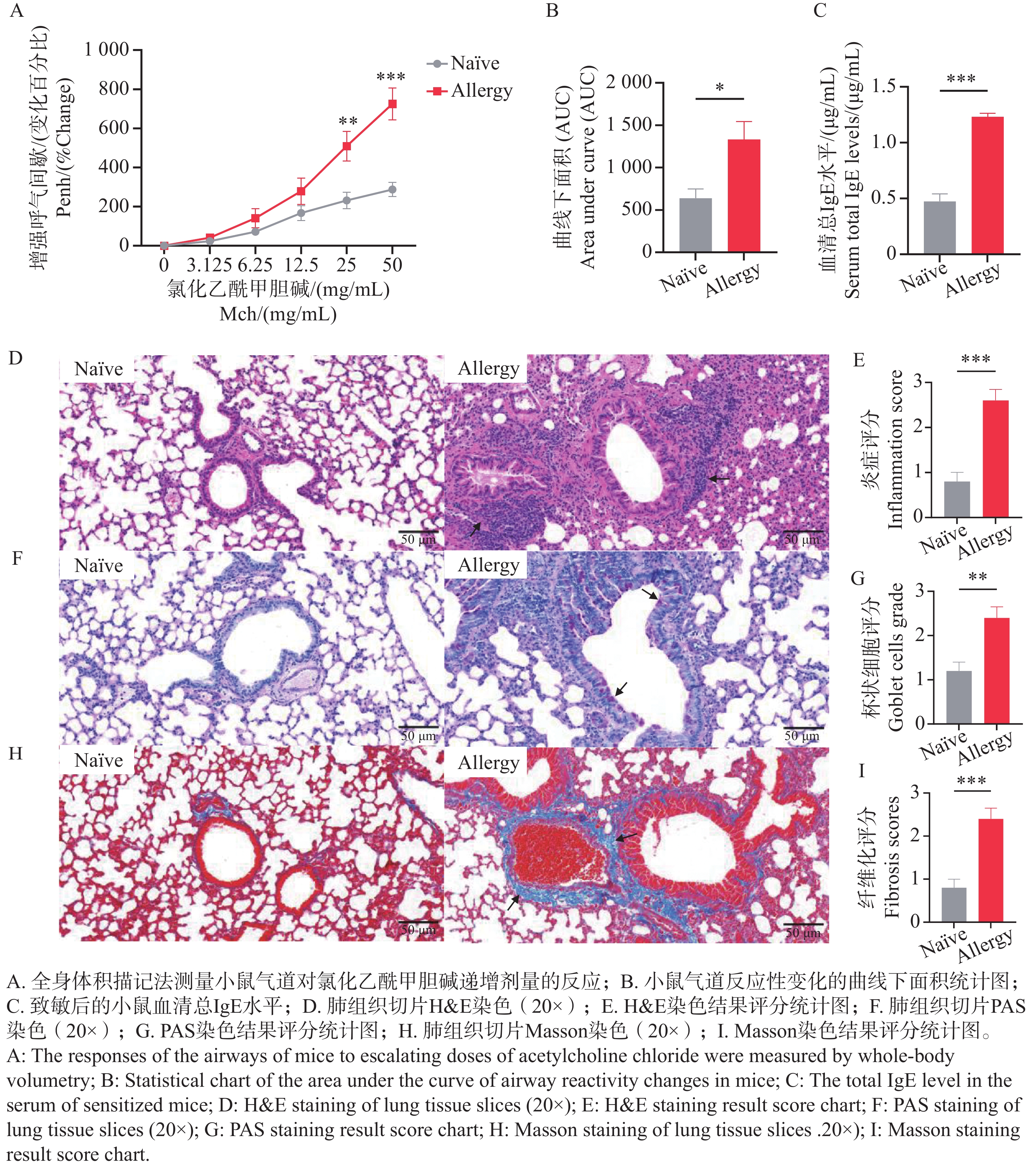
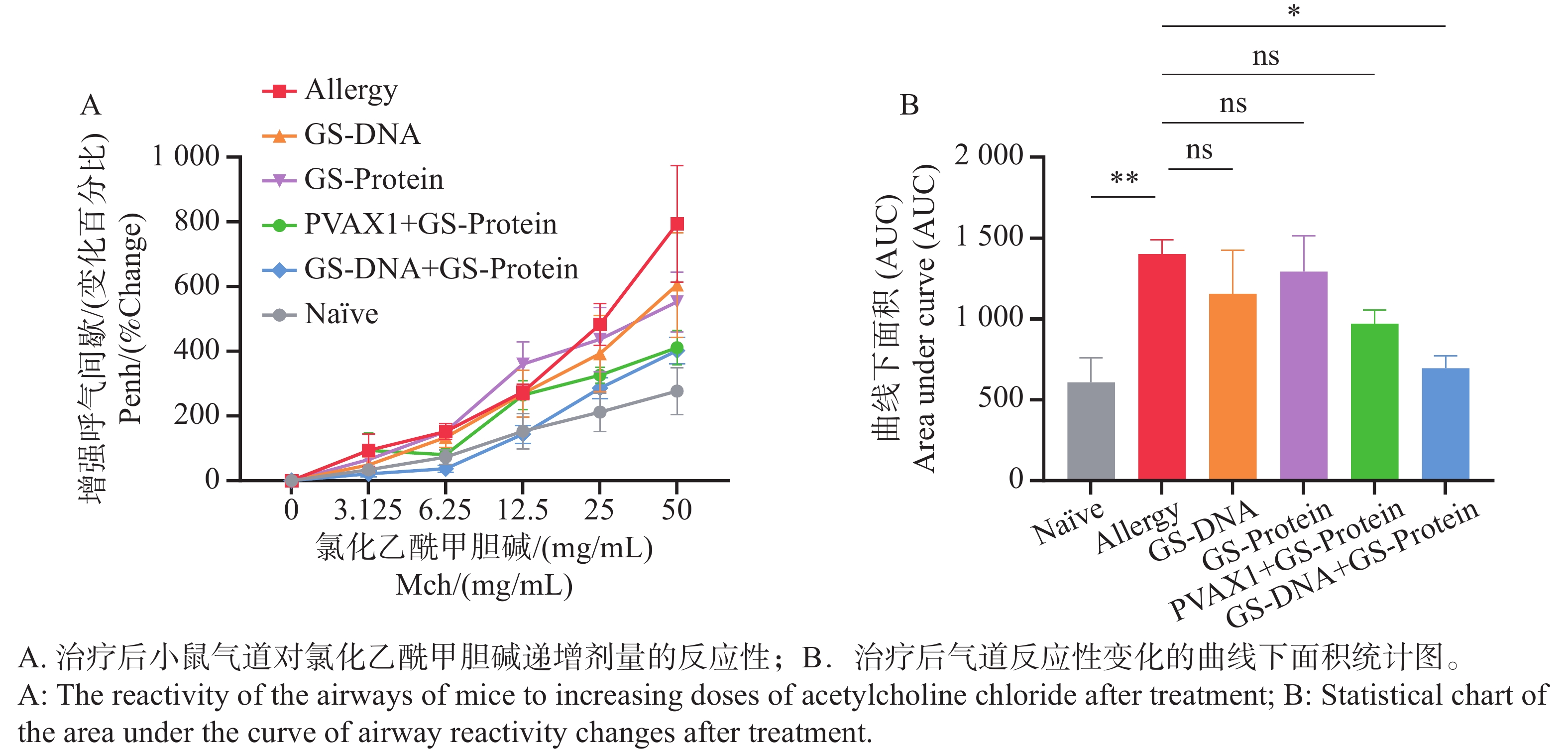
气道高反应性(Airway Hyperresponsiveness,AHR)是过敏性哮喘等呼吸道疾病的核心病理特征之一,表现为支气管平滑肌收缩、气道阻力升高及呼吸功能异常,也是动物模型中判断造模及干预效果的重要指标[25]。通过全身体积描记法,测量致敏后小鼠暴露于不同浓度氯化乙酰甲胆碱时的Enhanced Pause(Penh)值与雾化PBS时的Penh值的百分比,及Penh值百分比的曲线下面积衡量AHR的变化。结果表明(图5-A、B),与Naïve组相比,致敏组小鼠的气道呼吸阻力显著增大,Penh值百分比的曲线下面积统计显著增加,提示小鼠经过腹腔注射和气管灌入变应原后,诱导出了AHR状态。
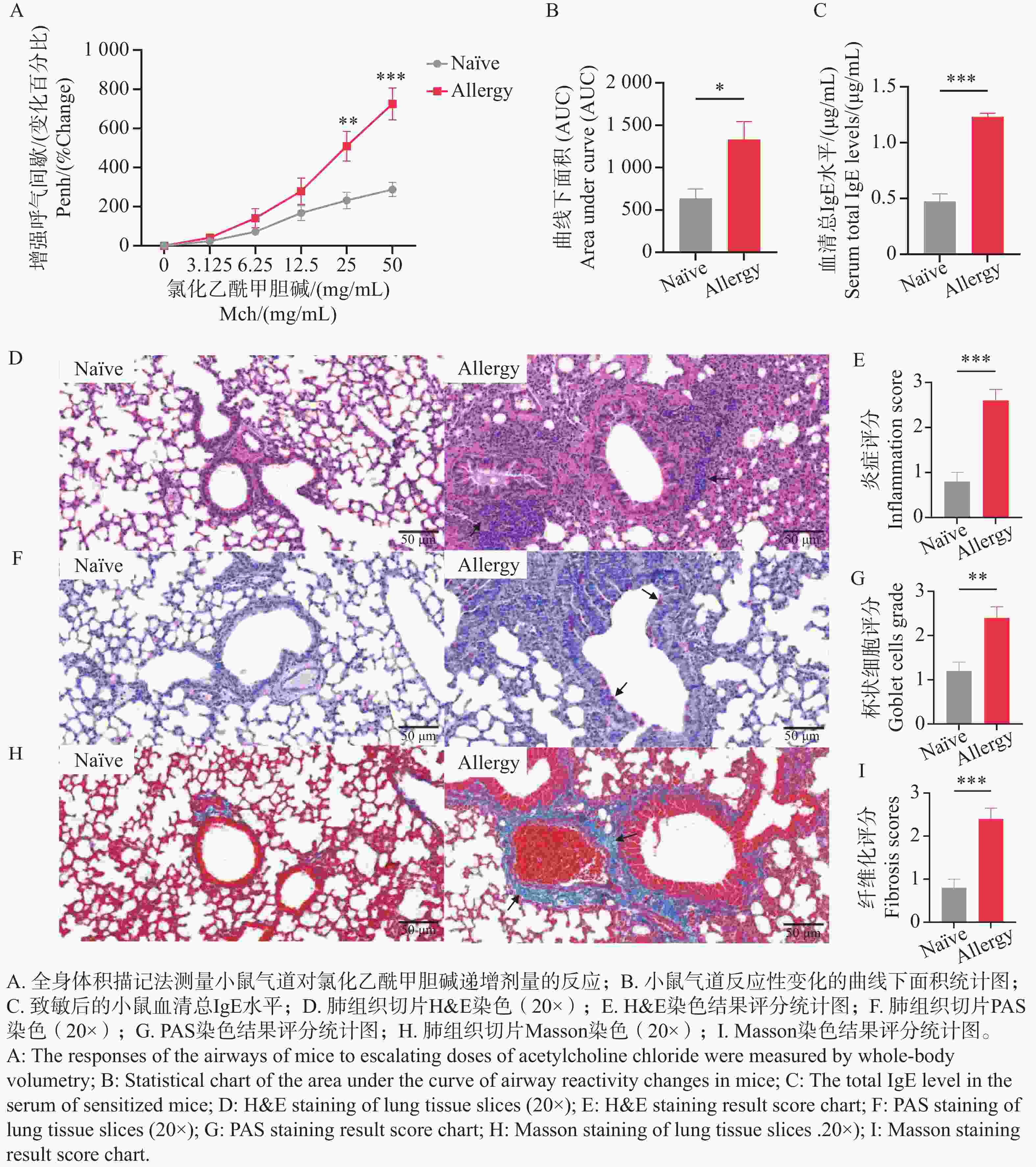
图 5 过敏小鼠模型的建立
Figure 5. Establishment of an allergic mouse model
-
IgE是评估过敏反应的一个重要病理指标,在最后一次气管灌入3 d后,采眼眶血检测血清总IgE水平,结果表明(图5-C),与Naïve组相比,致敏组小鼠血清总IgE水平极显著升高,提示小鼠体内可能发生了过敏反应。
-
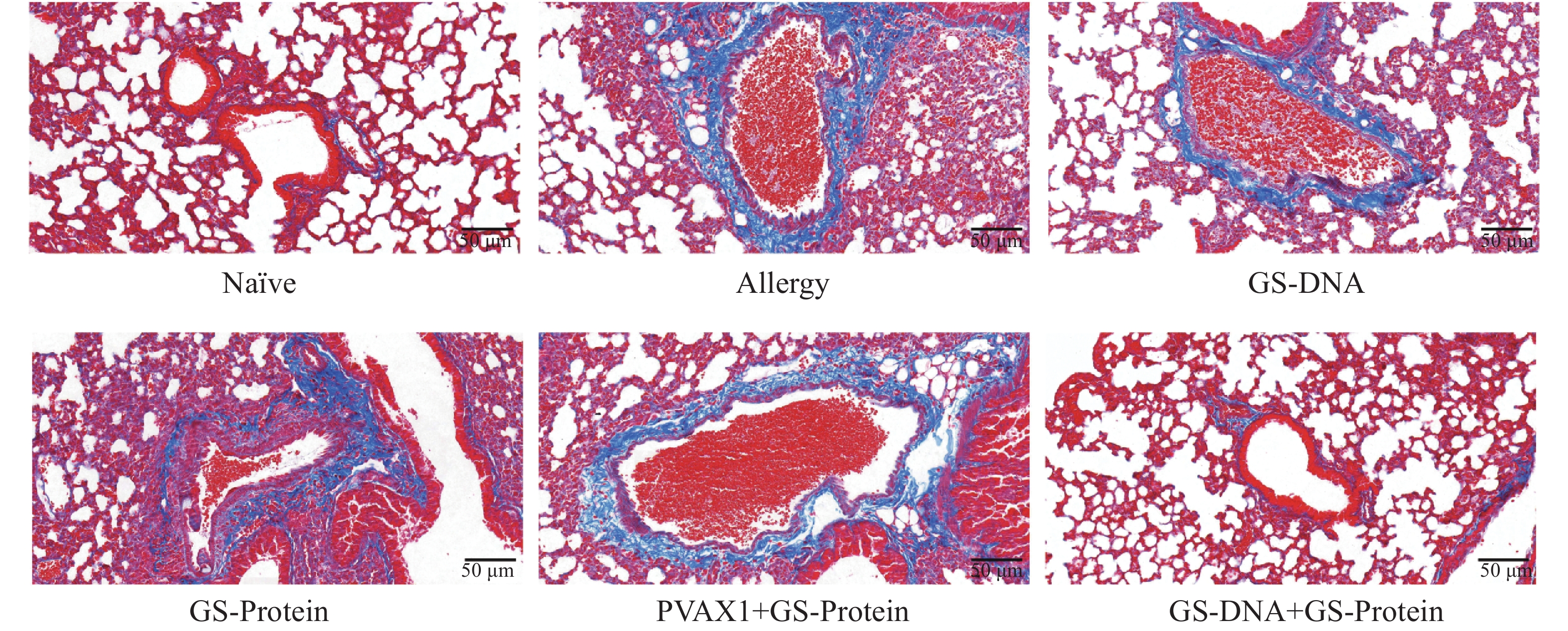
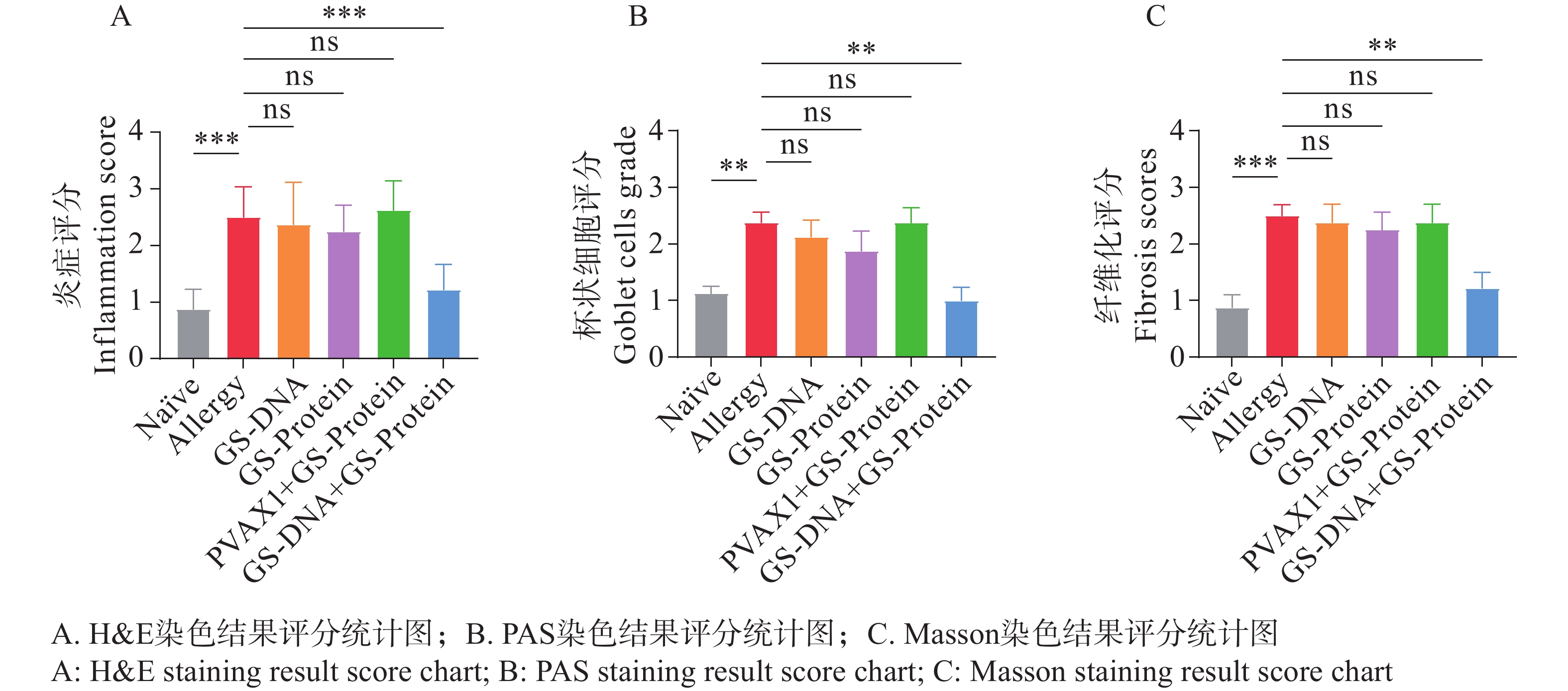
病理学切片是研究过敏性哮喘的重要工具,通过观察肺组织结构及细胞的变化,评估造模后小鼠的过敏情况。使用颈椎脱臼法处死小鼠后,取左肺于通用型组织固定液中固定,通过H&E、PAS及Masson染色观察肺组织中的炎症细胞浸润程度、支气管腔内杯状细胞数及胶原纤维化情况。H&E染色结果显示,致敏组小鼠肺组织支气管周围可见 5~8 层浸润的炎性细胞,炎症评分显著高于Naïve组(图5-D、E);PAS染色结果显示,致敏组支气管腔内杯状细胞数相较于Naïve组显著增加,提示黏液高分泌(图5-F、G);Masson染色结果显示,致敏组蓝色胶原纤维环绕气道壁,纤维化评分较Naïve组显著升高(图5-H、I)。上述3种切片结果说明,尘螨融合变应原可诱导出具有典型气道炎症、黏液过度分泌及早期重塑的过敏小鼠模型。
-
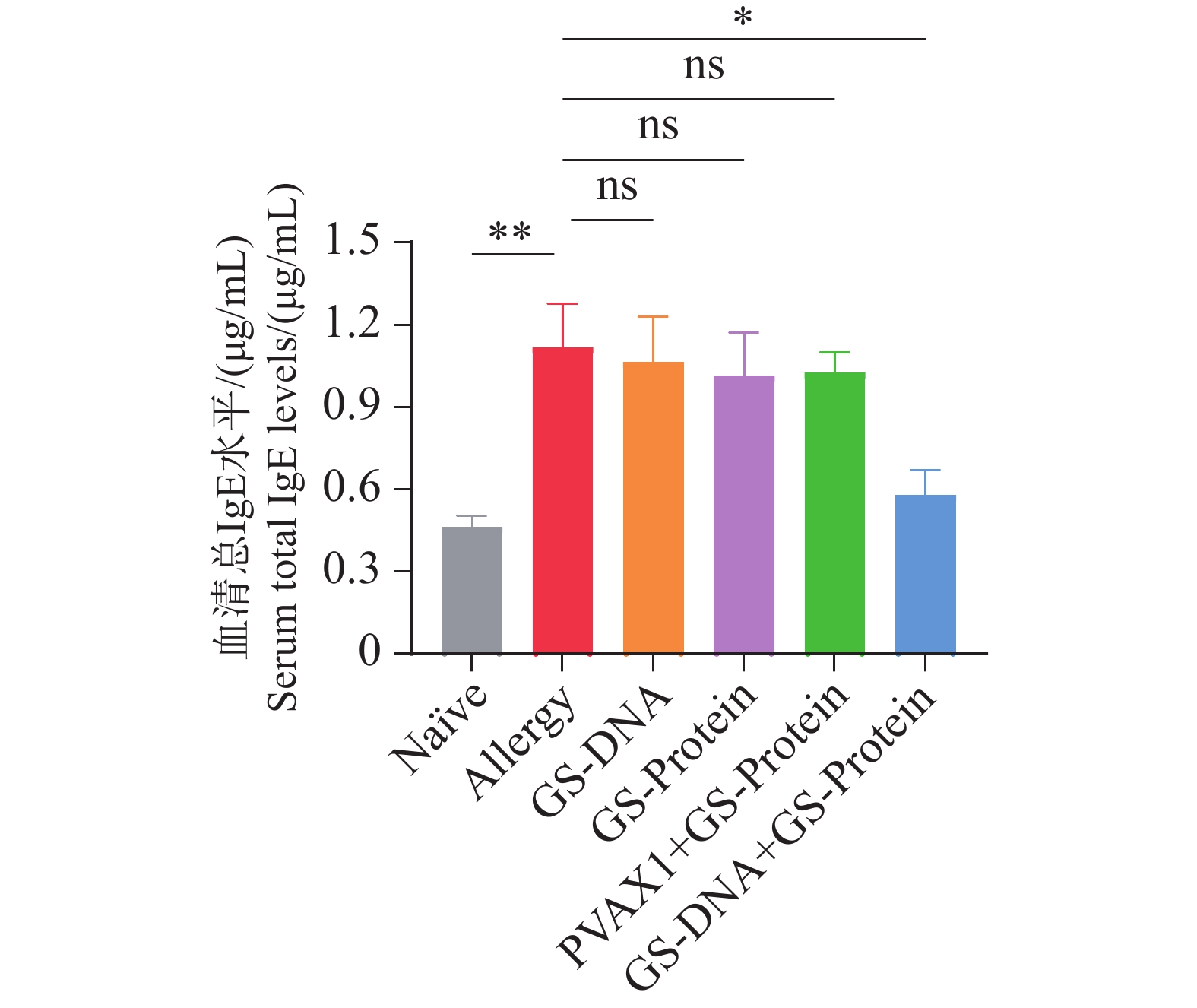
在最后一次过敏原气道激发1周后,对小鼠眼眶采血检测血清的总IgE水平。结果表明(图6),经疫苗治疗后,Naïve组和联合免疫组(GS-DNA+GS-Protein)小鼠血清中的IgE水平显著低于致敏组,而其他疫苗对照组的IgE水平与致敏组相比无统计学差异。IgE水平降低与AHR改善(图7)、耳部染料渗漏减少(图8)及肺组织炎症/黏液/纤维化评分下降(图12)的结果相一致,提示联合免疫对小鼠速发型过敏反应和气道炎症均有抑制作用。
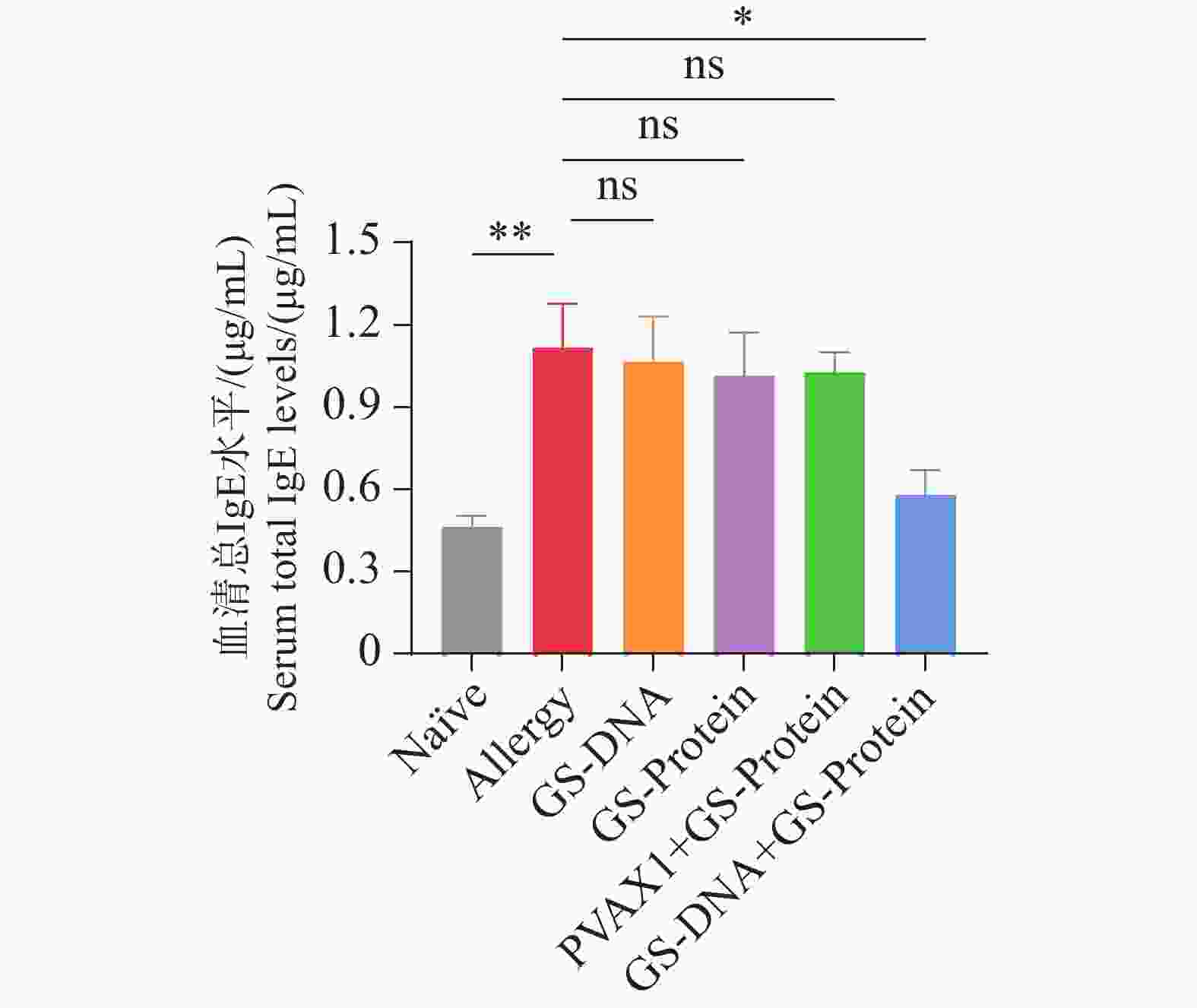
图 6 治疗后的小鼠血清总IgE水平
Figure 6. The total serum IgE level of mice after treatment
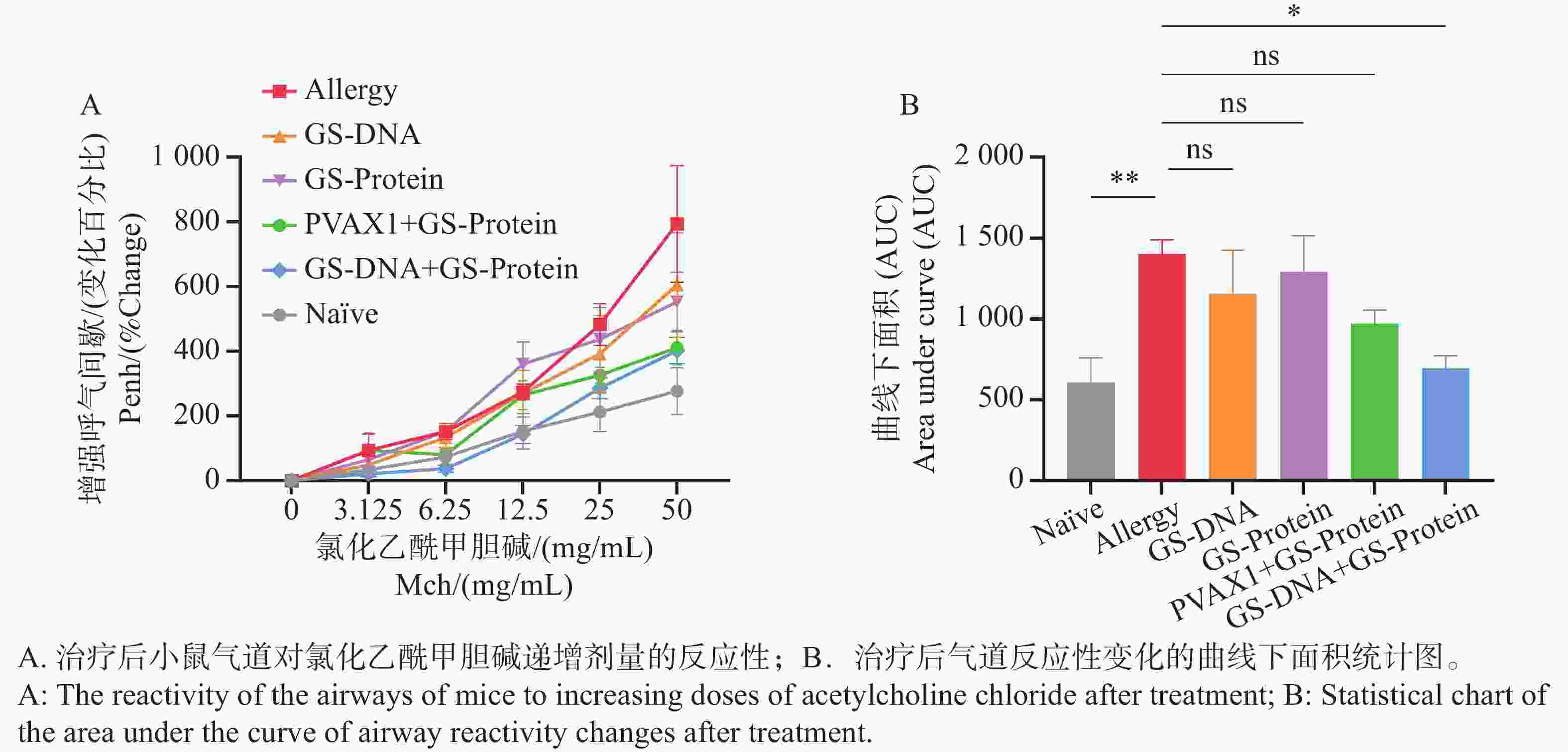
图 7 疫苗治疗后小鼠的气道高反应性变化
Figure 7. Airway hyperresponsiveness changes in mice after vaccine treatment
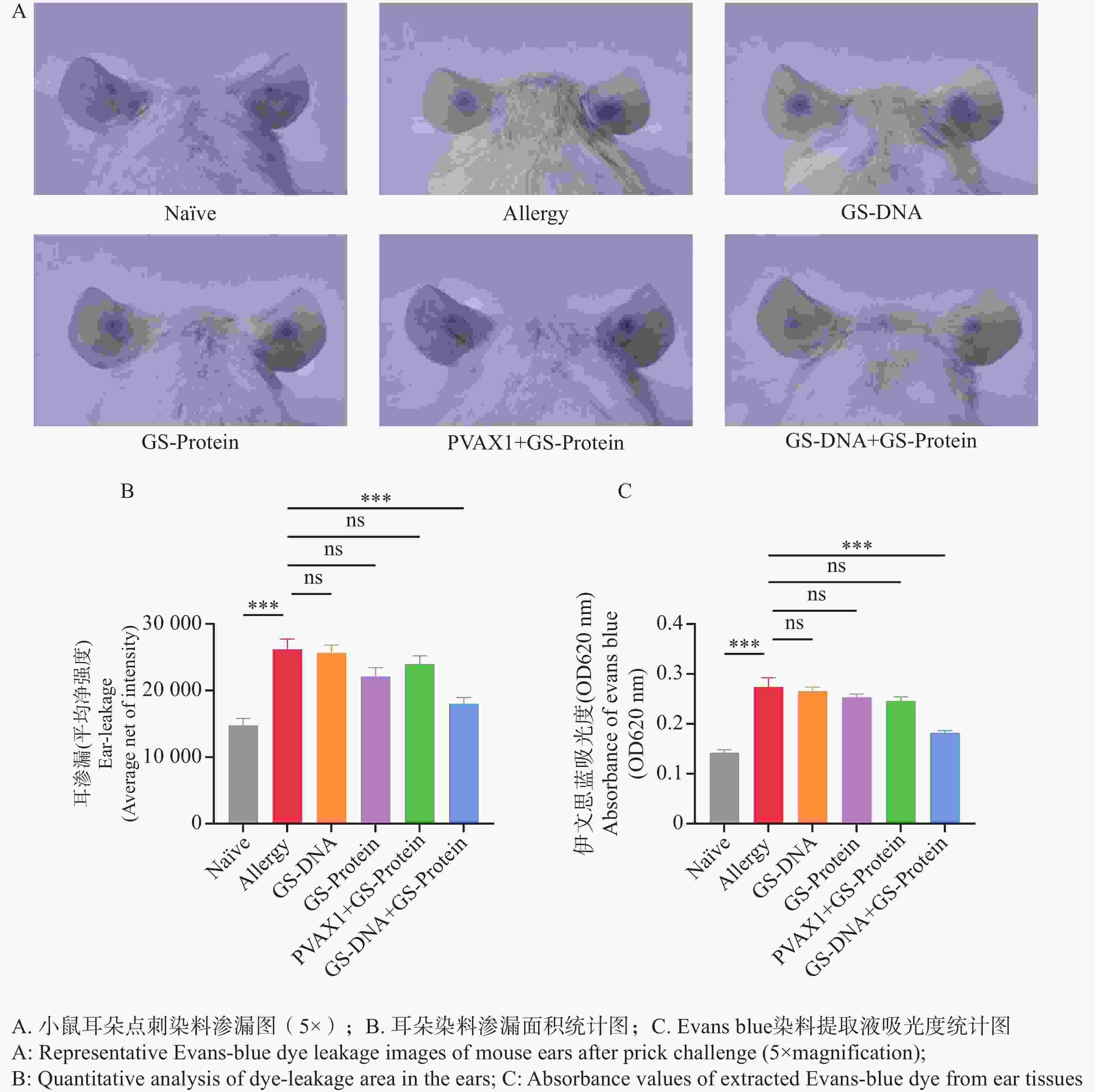
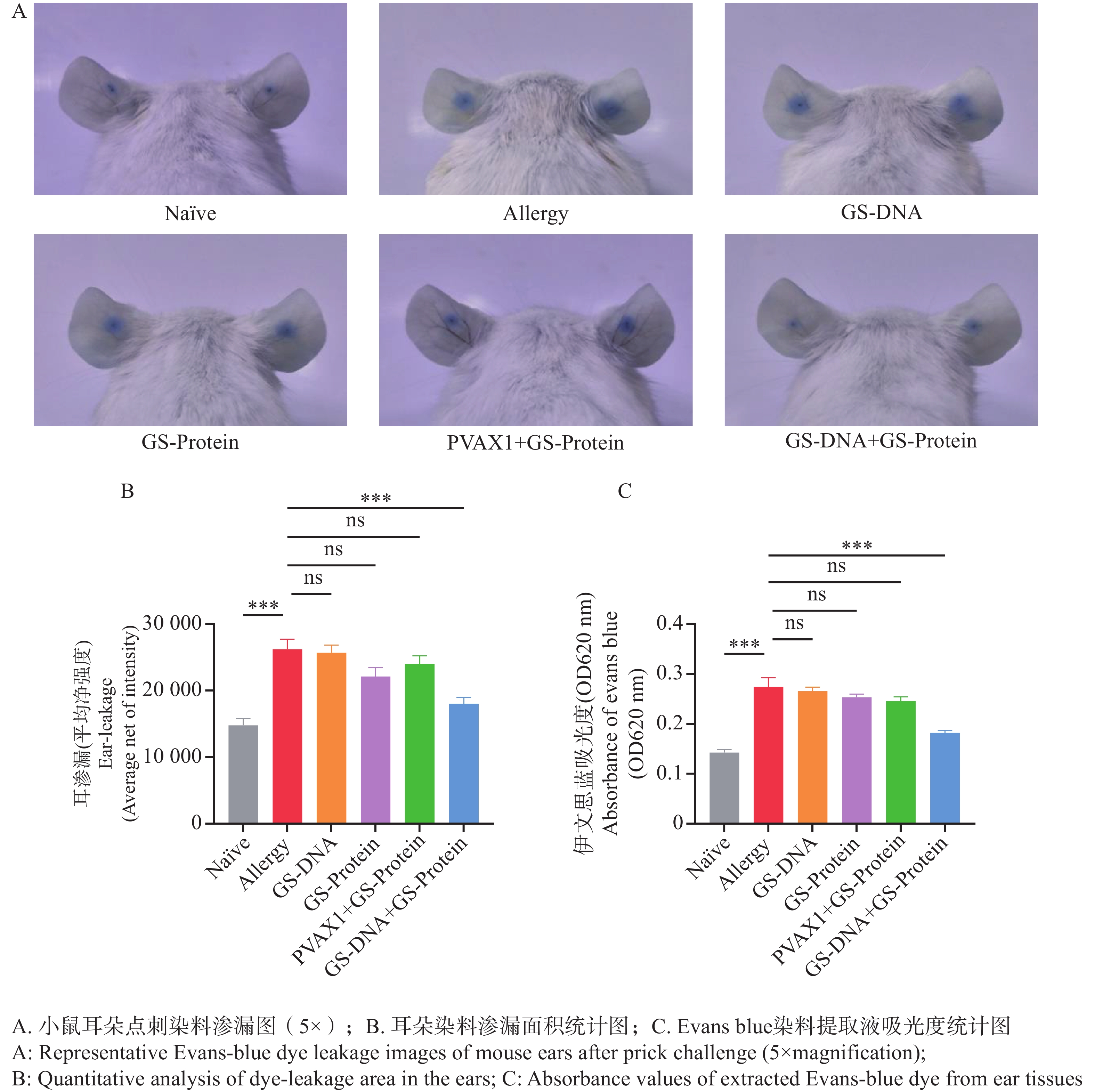
图 8 耳朵点刺试验
Figure 8. Ear-prick test
-
结果表明(图7-A),经疫苗治疗后,与致敏组相比,DNA组(GS-DNA)和蛋白组(GS-Protein)以及空载质粒/蛋白组(PVAX1+GS-Protein)的对照组小鼠的气道呼吸阻力有所降低,但无显著性差异;联合免疫组(GS-DNA+GS-Protein)的气道呼吸阻力与致敏组相比显著下降,且恢复到与Naïve组无明显差异。曲线下面积的统计结果与其一致(图7-B)。上述结果表明,DNA疫苗和蛋白疫苗联合免疫能够有效缓解尘螨融合变应原诱发的气道高反应性。
-
在过敏反应中,IgE与肥大细胞和嗜碱性粒细胞交联后可释放组胺,组胺可引起毛细血管扩张和通透性增加。在对小鼠进行耳朵点刺试验后,通过计算抗原蛋白刺激后点刺部位的染料渗漏面积及荧光强度,来评估各组疫苗对尘螨过敏小鼠模型的治疗效果。结果表明(图8-A、B),与致敏组相比,Naïve组和联合免疫组(GS-DNA+GS-Protein)的染料渗漏面积极显著减少,而其他疫苗对照组的染料渗漏面积与致敏组相比无显著性差异。
剪下小鼠耳部组织浸入甲酰胺溶液中以充分提取染料,经水浴锅温育后,在OD620 波长处测量提取液的吸光度值。结果表明(图8-C),Naïve组和联合免疫组(GS-DNA+GS-Protein)小鼠耳朵染料提取液的吸光度值显著低于致敏组小鼠,而其他疫苗对照组与致敏组相比无显著性差异。
-
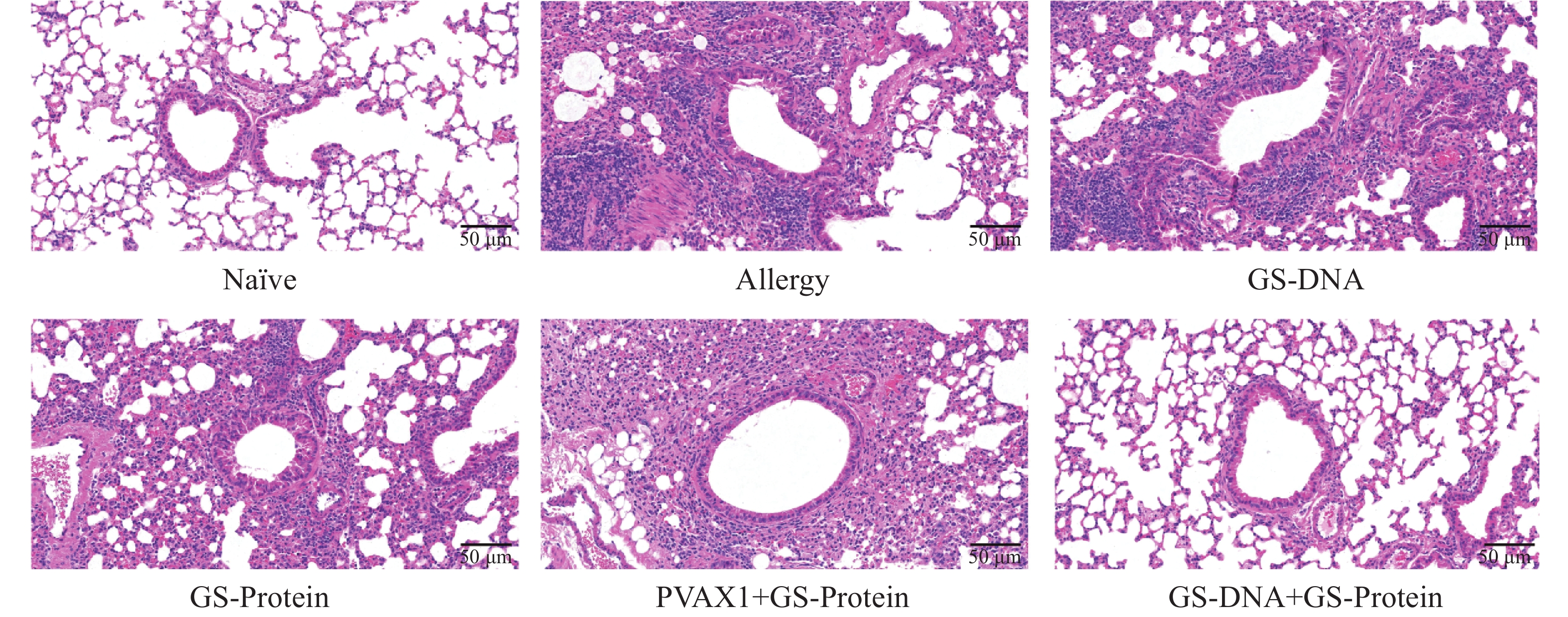
通过颈椎脱臼法处死小鼠后,取左肺于通用型组织固定液中固定并切片。通过H&E、PAS以及Masson染色,综合分析DNA疫苗和蛋白疫苗联合免疫对尘螨过敏小鼠模型的肺组织炎症的影响。H&E染色结果显示(图9、图12-A),与Naïve组相比,致敏组小鼠的肺组织中仍有较多的炎症细胞浸润;与致敏组相比,联合免疫组(GS-DNA+GS-Protein)小鼠经治疗后肺组织的炎症细胞浸润程度显著降低,其他3个疫苗对照组与致敏组相比无显著性差异。
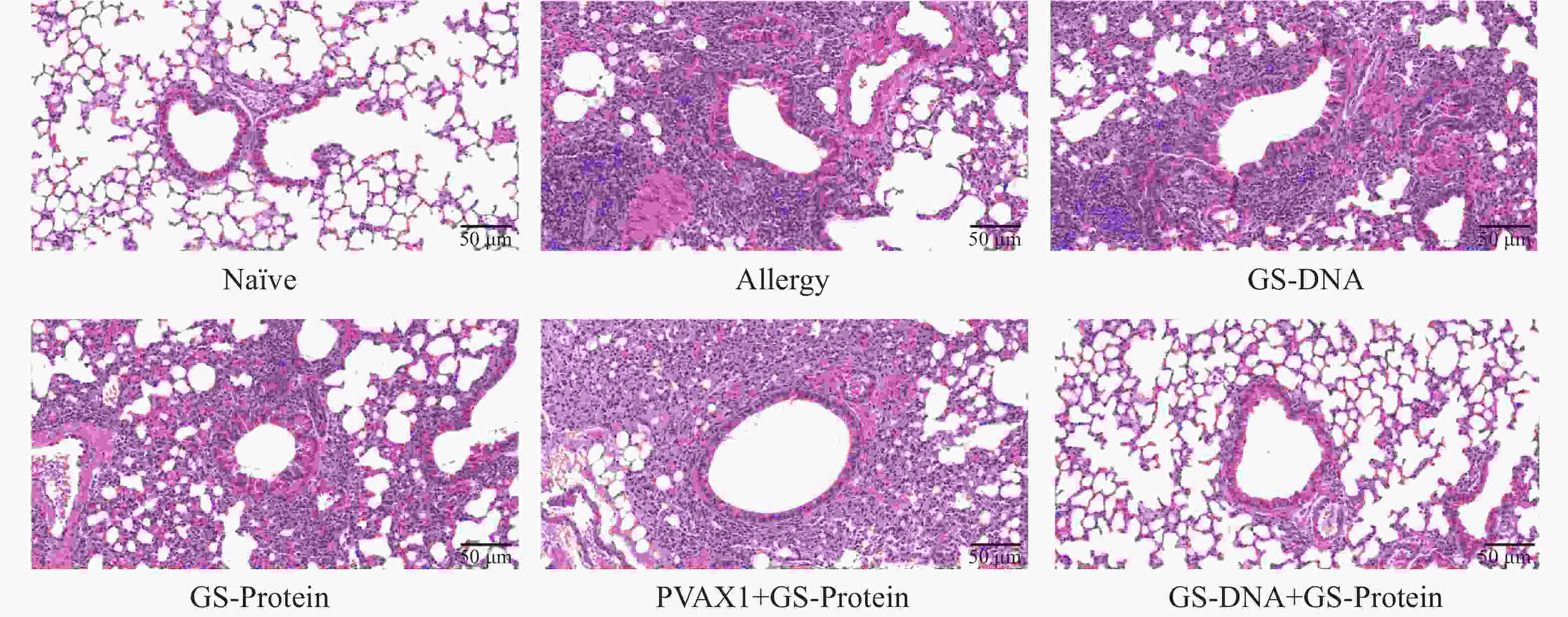
图 9 肺组织切片H&E染色(20×)
Figure 9. H&E staining of lung tissue slices (20×)
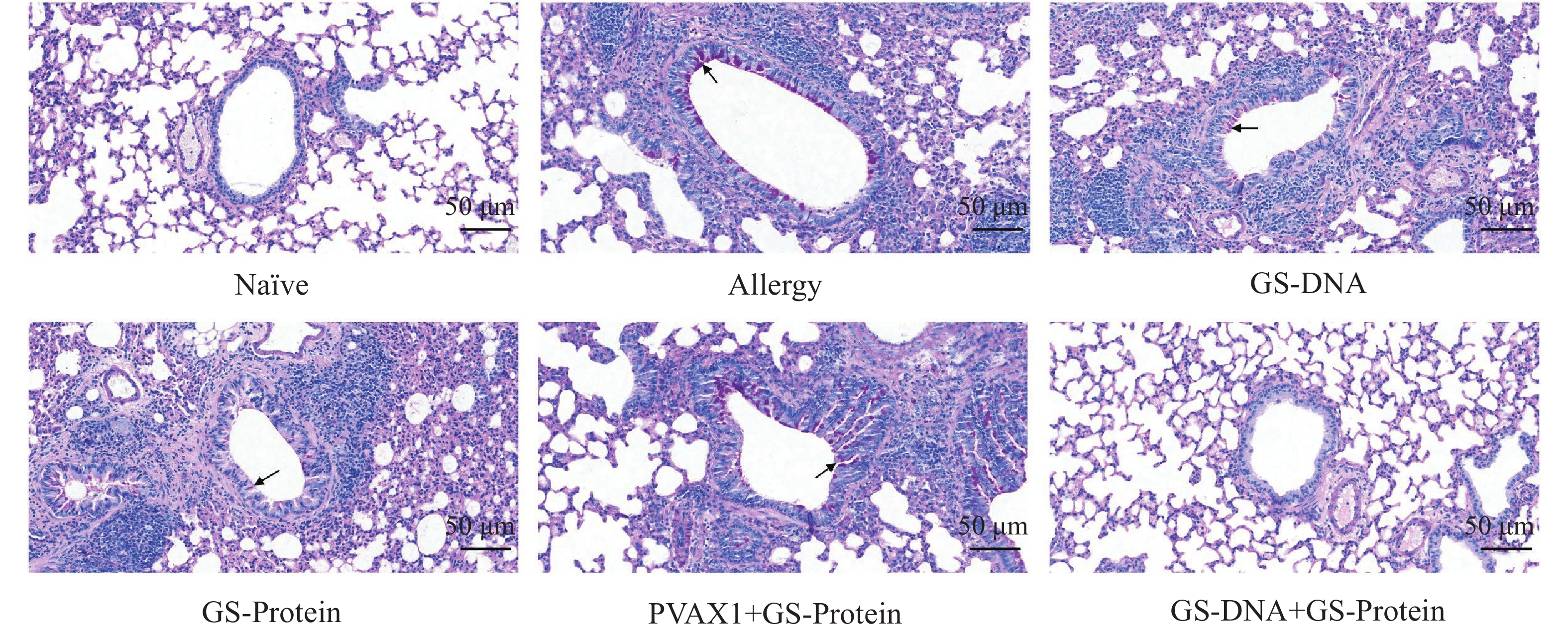
杯状细胞在呼吸系统和免疫反应中具有重要作用,受到过敏原刺激或有炎症反应时会迅速增加,同时分泌黏蛋白,黏蛋白会被PAS染成紫红色。PAS染色结果显示(图10、图12-B),联合免疫组(GS-DNA+GS-Protein)小鼠支气管内的平均杯状细胞数与致敏组相比显著减少,而其他3个疫苗对照组与致敏组相比无显著性差异。Masson染色结果显示(图11、图12-C),与致敏组相比,联合免疫组(GS-DNA+GS-Protein)小鼠肺部胶原纤维沉积程度显著下降。
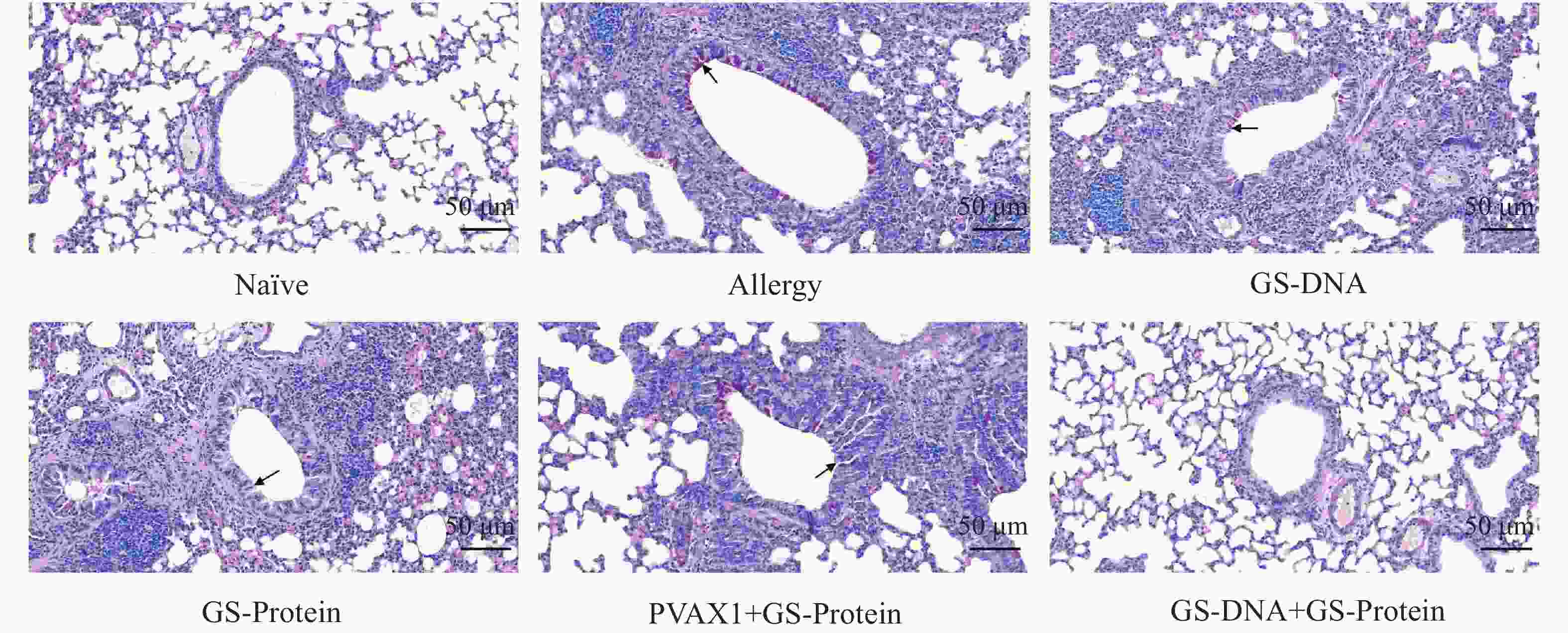
图 10 肺组织切片PAS染色(20×)
Figure 10. PAS staining of lung tissue slices (20×)
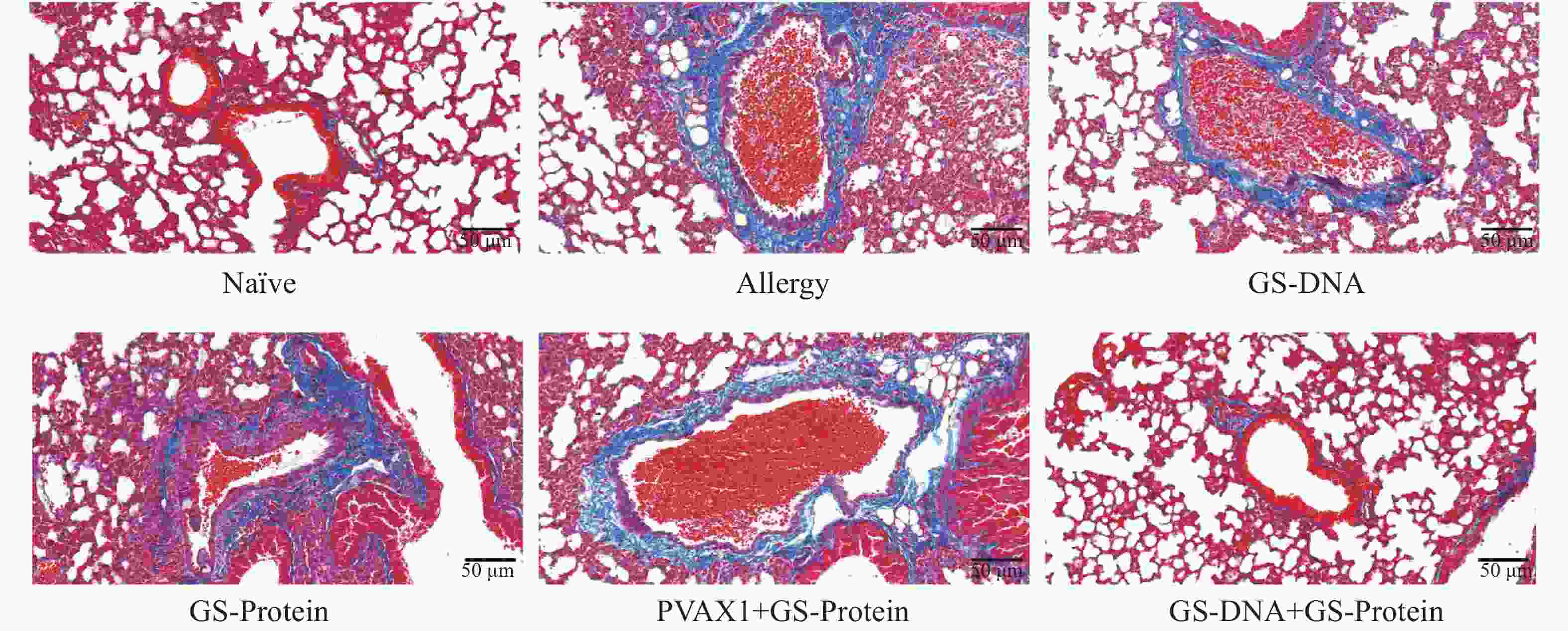
图 11 肺组织切片Masson染色(20×)
Figure 11. Masson staining of lung tissue slices (20×)
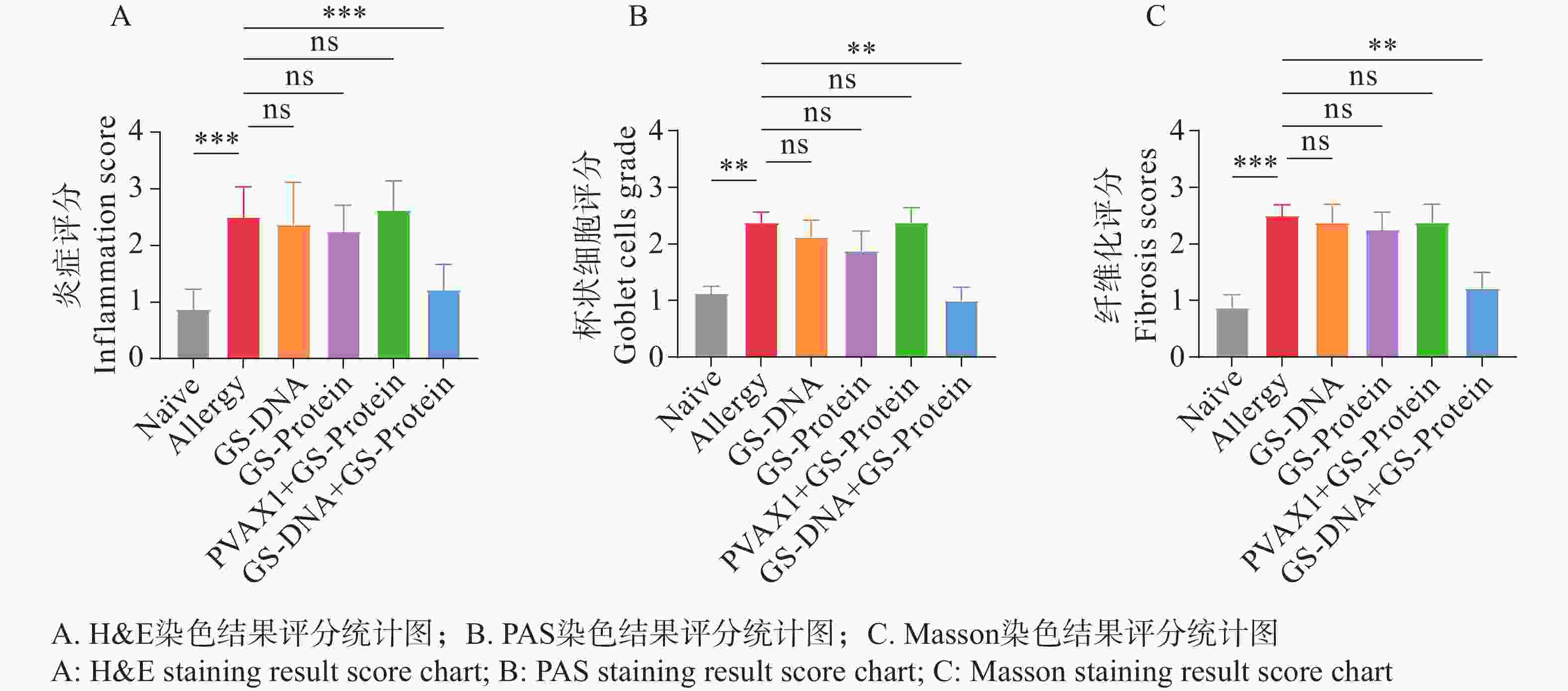
图 12 肺组织切片染色结果评分统计图
Figure 12. Statistical chart of lung tissue slice staining results
综合H&E、PAS、Masson染色的切片结果,可见联合免疫组(GS-DNA+GS-Protein)小鼠的炎症细胞浸润减少、杯状细胞数减少且黏液分泌降低、胶原沉积减少且纤维化程度减轻,相对应的染色结果评分均显著下降,说明联合免疫可有效缓解过敏小鼠的肺组织炎症损伤程度。
-
近年来,过敏性疾病如支气管哮喘、过敏性鼻炎、特应性皮炎的发病率逐渐上升,全球约20%的人口患有过敏性疾病[26]。从免疫应答极性调节的角度来看,尘螨过敏属于典型的Th2型免疫优势应答疾病,而IgE作为I型变态反应的核心效应分子,是由Th2型细胞诱导产生的,并在速发型变态反应中起核心作用。DNA疫苗通过载体携带过敏原表位基因,能在宿主细胞内持续表达抗原,从而通过内源性抗原提呈途径激活细胞免疫,诱导Th1型细胞因子(如IFN-γ)的分泌来拮抗Th2型细胞的分化,从源头上阻断IgE的过度产生,纠正免疫失衡状态。蛋白疫苗则直接提供成熟的外源性抗原,快速激活体液免疫系统,诱导抗体产生,在联合免疫的策略中,蛋白疫苗可能弥补了单独DNA疫苗免疫原性较弱的不足。
本研究中,致敏组小鼠表现出典型的Th2型反应特征,即血清总IgE水平显著升高,而经过联合疫苗治疗后,IgE水平显著下降,这一关键指标的变化不仅与IgE在促进过敏反应中的重要角色一致[27],同时也提示联合免疫策略可能抑制了Th2型免疫反应的过度活跃,促使免疫反应向Th1型方向偏移,说明联合疫苗可以通过不同路径共同调节Th1/Th2平衡。值得注意的是,本实验中单独的DNA组(GS-DNA)在降低IgE和缓解症状方面效果有限,这提示单独的Th1偏移诱导可能不足以完全逆转已建立的过敏反应,而DNA疫苗与蛋白疫苗的联合应用发挥了关键的细胞免疫与体液免疫协同效应,从而实现对免疫极性的有效重塑。
免疫应答极性的重塑可改善过敏小鼠的各项病理生理指标。在局部速发型变态反应方面,当发生局部过敏反应时,IgE与肥大细胞表面FcεR I结合可引发肥大细胞脱颗粒,释放组胺、白三烯等炎症介质,从而引起毛细血管通透性增加[28]。在本实验中,尘螨致敏小鼠耳廓点刺变应原后,观察到伊文思蓝渗漏面积显著增加;而联合疫苗免疫干预后,血清IgE水平显著降低,导致了效应阶段肥大细胞和嗜碱性粒细胞表面结合的IgE抗体减少,从而在再次接触过敏原时,阻断肥大细胞脱颗粒及后续的炎症级联反应,直接表现为小鼠耳朵的伊文思蓝渗漏面积明显减小。表明联合方案可通过降低IgE水平,有效抑制局部速发型变态反应及炎症介质释放。
在气道功能方面,本研究通过WBP全身体积描记法测量了小鼠的气道呼吸阻力,发现致敏组气道阻力显著高于Naïve组,暗示了致敏组小鼠可能是由于肺部Th2型免疫反应的增强导致大量炎症细胞浸润,从而堵塞肺泡,因而易受刺激物刺激而引起强烈的气道收缩,呼吸阻力增大,同时也证明造模后成功诱导了小鼠AHR状态。进一步对疫苗干预后的气道反应性进行评估,结果表明联合免疫方案能有效降低AHR,提高肺功能,这一改善可以归因于联合免疫诱导的Th1型偏移抑制了气道内的Th2型炎症环境,减少了炎症细胞对气道的浸润与阻塞,从而达到防治过敏性气道炎症的目的。
理想的哮喘动物模型应能精确模拟人类哮喘的核心病理生理学特征,主要包括:由肥大细胞、嗜酸性粒细胞及T淋巴细胞等多种免疫细胞介导的慢性气道炎症;对多种刺激物产生的气道高反应性;以及以气道壁结构改变为特征的慢性气道重塑[29]。在本研究中,与Naïve组相比,致敏组小鼠肺组织可见明显的炎症细胞浸润及气道病理损害,其他三个疫苗对照组也呈现相同的趋势;而联合免疫组(GS-DNA+GS-Protein)的肺部炎症程度明显减轻,支气管内平均杯状细胞数减少,胶原纤维沉积程度降低,炎症评分也较致敏组显著下降,表明联合免疫治疗方案可通过调节免疫应答极性,改善肺部的免疫微环境,不仅缓解了急性炎症,还抑制了由Th2型细胞因子驱动的气道重塑过程。值得一提的是,除了DNA组(GS-DNA)和蛋白组(GS-Protein),本研究还增加了空载质粒/蛋白(PVAX1+GS-Protein)对照组,由各项实验结果可看出,空载质粒/蛋白对照组共同免疫并不能抑制尘螨融合变应原诱发的过敏反应,这说明联合免疫缓解小鼠过敏反应的关键在于免疫后DNA疫苗表达与蛋白疫苗相同的抗原,而与载体骨架上的元件无关[21]。
-
本研究利用尘螨融合变应原Der p 1- Der f 1- Blo t 5成功诱导了过敏小鼠模型,并采用DNA疫苗与蛋白疫苗联合免疫的治疗策略来评估对过敏小鼠模型的免疫治疗效果。结果表明,联合免疫方案主要是通过细胞免疫与体液免疫的协同作用,重塑免疫应答极性,在降低IgE、抑制速发型变态反应(耳朵渗漏)、改善肺功能以及减轻慢性气道炎症等方面发挥了综合治疗效应,为研究同时抑制多种尘螨诱发过敏反应的疫苗提供了动物模型参考及理论依据。
尽管本研究证实了联合免疫策略在尘螨过敏小鼠模型中具有良好的治疗效果,但主要侧重于表型指标的观察(如IgE水平、气道阻力、病理切片等),尚未对深层的免疫学机制进行系统验证,仍存在一定的局限性。未来可通过检测关键的Th1/Th2型细胞因子(如IFN-γ、IL-4、IL-5、IL-13等)的变化,流式细胞术深入分析脾脏淋巴细胞亚群(如Th1、Th2、Treg细胞)的比例,以及检测阻断性抗体(各IgG亚类)的水平,深入探究联合免疫重塑免疫应答极性,诱导免疫耐受的具体细胞机制与分子基础。
Efficacy of a combined DNA and protein vaccine immunotherapy for house dust mite-induced allergy in a mouse model
-
摘要: 旨在分析DNA疫苗与蛋白疫苗联合免疫对尘螨融合变应原诱导的过敏小鼠模型的治疗效果。本研究通过构建尘螨主要变应原融合表位真核表达质粒及原核表达质粒,制备DNA和蛋白联合疫苗。利用尘螨融合变应原Der p 1- Der f 1-Blo t 5诱导过敏小鼠模型,将其随机分为致敏组(Allergy)和疫苗组,每组6只,另外设置正常组(Naïve)为对照。其中,疫苗组注射相应的疫苗,致敏组注射等体积PBS(磷酸盐缓冲液),正常组则不做任何处理。通过检测小鼠血清总IgE(免疫球蛋白E)水平、气道高反应性变化、耳朵染料渗漏面积、肺部切片病理特征分析等,综合评估疫苗的治疗效果。结果表明,与致敏组相比,DNA疫苗与蛋白疫苗联合免疫组小鼠的血清总IgE水平显著下降、气道高反应性显著降低、耳部染料渗漏面积显著减少,肺组织炎症细胞浸润有所缓解、支气管内平均杯状细胞数减少、胶原纤维沉积程度减轻。综上所述,DNA疫苗与蛋白疫苗联合免疫对尘螨过敏小鼠模型具有较好的免疫治疗效应,可为尘螨过敏症防治型疫苗的研发提供重要的参考。Abstract: An attempt was made to analyze the therapeutic effect of combined DNA and protein vaccines on a mouse model of dust mite allergy induced by a fusion allergen. In this attempt eukaryotic and prokaryotic expression plasmids of the major dust mite allergen fusion epitope were constructed to prepare a combined DNA and protein vaccine. A mouse model of dust mite allergy was induced by the fusion allergen Der p 1-Der f 1-Blo t 5. The mice were randomly divided into an allergy group (Allergy) and a vaccine group, with 6 mice in each group. A normal group (Naïve) was set as a control. The vaccine group was injected with the corresponding vaccine, the allergy group was injected with an equal volume of PBS (phosphate buffered saline), and the normal group received no treatment. The therapeutic effect of the vaccine was comprehensively evaluated by detecting the total IgE (immunoglobulin E) level in the mouse serum, changes in airway hyperresponsiveness, the area of dye leakage in the ear, and pathological features of lung sections. The results showed that compared with the allergy group, the combined DNA and protein vaccine group had significantly lower total IgE levels of serum, significantly reduced airway hyper-responsiveness, had significantly smaller area of dye leakage in the mouse ear, alleviated lung tissue inflammatory cell infiltration, reduced average goblet cell numbers in the bronchus, and decreased collagen fiber deposition. It is concluded that the combined DNA and protein vaccine has a good immunotherapeutic effect on the dust mite allergy mouse model, which can provide an important reference for the development of preventive and therapeutic vaccines for dust mite allergy.
-
Key words:
- dust mite /
- allergy /
- combined immunization /
- vaccine /
- immunotherapy
-
图 3 尘螨主要变应原表达载体的构建
Fig. 3 Construction of the expression vector of the main allergens of dust mites
图 4 SDS-PAGE电泳检测重组蛋白的表达与纯化
Fig. 4 SDS-PAGE electrophoresis for the detection of expression and purification of the recombinant protein
图 6 治疗后的小鼠血清总IgE水平
注:正常组(Naïve);致敏组(Allergy);DNA组(GS-DNA);蛋白组(GS-Protein);空载质粒/蛋白组(PVAX1+GS-Protein);联合免疫组(GS-DNA+GS-Protein)(ns,P>0.05;*,P<0.05;**,P<0.01;***,P<0.001)。
Fig. 6 The total serum IgE level of mice after treatment
Note: Normal Group (Naïve); Allergy Group (Allergy); DNA Group (GS-DNA); Protein Group (GS-Protein); Empty Plasmid/Protein Group (PVAX1+GS-Protein); Combined Immunization Group (GS-DNA+GS-Protein) (ns: P>0.05; *: P<0.05; **: P<0.01; ***: P<0.001).
图 7 疫苗治疗后小鼠的气道高反应性变化
Fig. 7 Airway hyperresponsiveness changes in mice after vaccine treatment
图 9 肺组织切片H&E染色(20×)
注:正常组(Naïve);致敏组(Allergy);DNA组(GS-DNA);蛋白组(GS-Protein);空载质粒/蛋白组(PVAX1+GS-Protein);联合免疫组(GS-DNA+GS-Protein)。
Fig. 9 H&E staining of lung tissue slices (20×)
Note: Normal Group (Naïve); Allergy Group (Allergy); DNA Group (GS-DNA); Protein Group (GS-Protein); Empty Plasmid/Protein Group (PVAX1+GS-Protein); Combined Immunization Group (GS-DNA+GS-Protein).
图 10 肺组织切片PAS染色(20×)
注:正常组(Naïve);致敏组(Allergy);DNA组(GS-DNA);蛋白组(GS-Protein);空载质粒/蛋白组(PVAX1+GS-Protein);联合免疫组(GS-DNA+GS-Protein)。
Fig. 10 PAS staining of lung tissue slices (20×)
Note: Normal Group (Naïve); Allergy Group (Allergy); DNA Group (GS-DNA); Protein Group (GS-Protein); Empty Plasmid/Protein Group (PVAX1+GS-Protein); Combined Immunization Group (GS-DNA+GS-Protein).
图 11 肺组织切片Masson染色(20×)
注:正常组(Naïve);致敏组(Allergy);DNA组(GS-DNA);蛋白组(GS-Protein);空载质粒/蛋白组(PVAX1+GS-Protein);联合免疫组(GS-DNA+GS-Protein)。
Fig. 11 Masson staining of lung tissue slices (20×)
Note: Normal Group (Naïve); Allergy Group (Allergy); DNA Group (GS-DNA); Protein Group (GS-Protein); Empty Plasmid/Protein Group (PVAX1+GS-Protein); Combined Immunization Group (GS-DNA+GS-Protein).
表 1 肺部组织切片评分细则
Table 1 Scoring criteria for lung tissue sections
得分
ScoreH&E染色
H&E stainingPAS染色
PAS stainingMasson染色
Masson staining0 支气管周围无炎症细胞浸润
No inflammatory cell infiltration around the bronchi管腔内无杯状细胞
No goblet cells in the lumen胶原纤维沉积含量正常
Normal collagen fiber deposition content.1 支气管周围偶尔出现炎症细胞
Occasionally inflammatory cells appear around the bronchi管腔内有1~5个杯状细胞
The lumen contains 1~5 goblet cells.轻度胶原纤维沉积
Mild collagen fiber deposition.2 支气管周围环绕着1~5层炎症细胞
The bronchi are surrounded by 1~5 layers of inflammatory cells管腔内有6~20个杯状细胞
The lumen contains 6~20 goblet cells.中度胶原纤维沉积
Moderate collagen fiber deposition.3 支气管周围有超过5层炎症细胞浸润
There are more than 5 layers of inflammatory cell infiltration around the bronchi管腔内有超过20个杯状细胞
The lumen contains more than 20 goblet cells.重度胶原纤维沉积
Severe collagen fiber deposition. 下载: 导出CSV
下载: 导出CSV
-
[1] 李荣, 陈仪婷, 吕佳骏, 等. 上海市学龄儿童过敏性疾病流行现状及相关因素分析[J]. 中国学校卫生, 2021, 42(8): 1251−1256. https://doi.org/10.16835/j.cnki.1000-9817.2021.08.030 doi: 10.16835/j.cnki.1000-9817.2021.08.030 [2] Li F, Zhou Y, Li S, et al. Prevalence and risk factors of childhood allergic diseases in eight metropolitan cities in China: a multicenter study [J]. BMC Public Health, 2011, 11(1): 437. https://doi.org/10.1186/1471-2458-11-437 doi: 10.1186/1471-2458-11-437 [3] 中华医学会变态反应分会呼吸过敏学组, 中华医学会呼吸病学分会哮喘学组. 中国过敏性哮喘诊治指南(第一版, 2019年)[J]. 中华内科杂志, 2019, 58(9): 636−655. https://doi.org/10.3760/cma.j.issn.0578-1426.2019.09.004 doi: 10.3760/cma.j.issn.0578-1426.2019.09.004 [4] Gehring U, Heinrich J, Jacob B, et al. Respiratory symptoms in relation to indoor exposure to mite and cat allergens and endotoxins [J]. European Respiratory Journal, 2001, 18(3): 555−563. https://doi.org/10.1183/09031936.01.00096801 doi: 10.1183/09031936.01.00096801 [5] Bousquet. Global Initiative for Asthma (GINA) and its objectives [J]. Clinical & Experimental Allergy, 2010, 30(S1): 2−5. https://doi.org/10.1046/j.1365-2222.2000.00088.x doi: 10.1046/j.1365-2222.2000.00088.x [6] Sharma S, Lackie P M, Holgate S T. Uneasy breather: the implications of dust mite allergens [J]. Clinical & Experimental Allergy, 2003, 33(2): 163−165. https://doi.org/10.1046/j.1365-2222.2003.01605.x doi: 10.1046/j.1365-2222.2003.01605.x [7] Liao E C, Hsieh C W, Chang C Y, et al. Enhanced allergic inflammation of Der p 2 affected by polymorphisms of MD-2 promoter [J]. Allergy, Asthma & Immunology Research, 2015, 7(5): 497−506. https://doi.org/10.4168/aair.2015.7.5.497 doi: 10.4168/aair.2015.7.5.497 [8] Chua K Y, Stewart G A, Thomas W R, et al. Sequence analysis of cDNA coding for a major house dust mite allergen, Der p 1. Homology with cysteine proteases [J]. Journal of Experimental Medicine, 1988, 167(1): 175−182. https://doi.org/10.1084/jem.167.1.175 doi: 10.1084/jem.167.1.175 [9] Waldron R, Mcgowan J, Gordon N, et al. Proteome and allergenome of the European house dust mite Dermatophagoides pteronyssinus [J]. PLoS One, 2019, 14(5): e0216171. https://doi.org/10.1371/journal.pone.0216171 doi: 10.1371/journal.pone.0216171 [10] Thomas W R, Smith W A, Hales B J, et al. Characterization and immunobiology of house dust mite allergens [J]. International Archives of Allergy and Immunology, 2002, 129(1): 1−18. https://doi.org/10.1159/000065179 doi: 10.1159/000065179 [11] Yi F C, Shek L P, Cheong N, et al. Molecular cloning of Blomia tropicalis allergens - a major source of dust mite allergens in the tropics and subtropics [J]. Inflammation & Allergy - Drug Targets, 2006, 5(4): 261−266. https://doi.org/10.2174/187152806779010954 doi: 10.2174/187152806779010954 [12] Kuo I C, Cheong N, Trakultivakorn M, et al. An extensive study of human IgE cross-reactivity of Blo t 5 and Der p 5 [J]. Journal of Allergy and Clinical Immunology, 2003, 111(3): 603−609. https://doi.org/10.1067/mai.2003.167 doi: 10.1067/mai.2003.167 [13] 毕玉田, 吴奎, 王长征. 屋尘螨过敏原Der p1研究进展[J]. 重庆医学, 2007, 36(2): 171−173. https://doi.org/10.3969/j.issn.1671-8348.2007.02.040 doi: 10.3969/j.issn.1671-8348.2007.02.040 [14] 郑佩燕, 孙宝清. 屋尘螨致敏蛋白组分Derp1、Derp2和Derp10检测的临床意义[J]. 中华临床免疫和变态反应杂志, 2014, 8(2): 150−154. https://doi.org/10.3969/j.issn.1673-8705.2014.02.012 doi: 10.3969/j.issn.1673-8705.2014.02.012 [15] Penagos M, Durham S R. Duration of allergen immunotherapy for inhalant allergy [J]. Current Opinion in Allergy and Clinical Immunology, 2019, 19(6): 594−605. https://doi.org/10.1097/ACI.0000000000000585 doi: 10.1097/ACI.0000000000000585 [16] Passalacqua G, Durham S R. Allergic rhinitis and its impact on asthma update: allergen immunotherapy [J]. Rhinitis, Sinusitis, and Ocular Diseases, 2007, 119(4): 881−891. https://doi.org/10.1016/j.jaci.2007.01.045 doi: 10.1016/j.jaci.2007.01.045 [17] Stewart-Tull D E S. Immunopotentiating conjugates [J]. Vaccine, 1985, 3(1): 40−44. https://doi.org/10.1016/0264-410X(85)90010-6 doi: 10.1016/0264-410X(85)90010-6 [18] Alexander J, Fikes J, Hoffman S, et al. The optimization of helper T lymphocyte (HTL) function in vaccine development [J]. Immunologic Research, 1998, 18(2): 79−92. https://doi.org/10.1007/BF02788751 doi: 10.1007/BF02788751 [19] Gurunathan S, Wu C Y, Freidag B L, et al. DNA vaccines: a key for inducing long-term cellular immunity [J]. Current Opinion in Immunology, 2000, 12(4): 442−447. https://doi.org/10.1016/S0952-7915(00)00118-7 doi: 10.1016/S0952-7915(00)00118-7 [20] 方芳. 重组MUC1-MBP融合蛋白疫苗抗肿瘤作用机制研究[D]. 长春: 吉林大学, 2010. [21] Jin H L, Xiao C, Geng S, et al. Protein/DNA vaccine-induced antigen-specific Treg confer protection against asthma [J]. European Journal of Immunology, 2008, 38(9): 2451−2463. https://doi.org/10.1002/eji.200737899 doi: 10.1002/eji.200737899 [22] Casaro M, Souza V R, Oliveira F A, et al. OVA-induced allergic airway inflammation mouse model [J]. Methods in Molecular Biology, 2019, 1916: 297−301. https://doi.org/10.1007/978-1-4939-8994-2_28 doi: 10.1007/978-1-4939-8994-2_28 [23] 魏春洁, 徐琳皓, 高江瑞, 等. 猫、犬融合变应原Fel d 1-Can f 1过敏性哮喘小鼠模型的建立[J]. 热带生物学报, 2024, 15(5): 558−566. https://doi.org/10.15886/j.cnki.rdswxb.20240036 doi: 10.15886/j.cnki.rdswxb.20240036 [24] 肖正泮, 刘金莹, 韦双双, 等. 重组热带螨Blo t 21变应原诱导小鼠变态反应气道炎症模型的建立[J]. 热带生物学报, 2017, 8(3): 371−376. https://doi.org/10.15886/j.cnki.rdswxb.2017.03.020 doi: 10.15886/j.cnki.rdswxb.2017.03.020 [25] Nair P, Hargreave F E. Measuring bronchitis in airway diseases: clinical implementation and application: airway hyperresponsiveness in asthma: its measurement and clinical significance [J]. Airway Hyperresponsiveness in Asthma: Its Measurement and Clinical Significance, 2010, 138(S2): 38S−43S. https://doi.org/10.1378/chest.10-0094 doi: 10.1378/chest.10-0094 [26] Schoos A M M. Atopic diseases—Diagnostics, mechanisms, and exposures [J]. Pediatric Allergy and Immunology, 2024, 35(7): e14198. https://doi.org/10.1111/pai.14198 doi: 10.1111/pai.14198 [27] Hamelmann E, Tadeda K, Oshiba A, et al. Role of IgE in the development of allergic airway inflammation and airway hyperresponsiveness–a murine model [J]. Allergy, 1999, 54(4): 297−305. https://doi.org/10.1034/j.1398-9995.1999.00085.x doi: 10.1034/j.1398-9995.1999.00085.x [28] Sawaguchi M, Tanaka S, Nakatani Y, et al. Role of mast cells and basophils in IgE responses and in allergic airway hyperresponsiveness [J]. The Journal of Immunology, 2012, 188(4): 1809−1818. https://doi.org/10.4049/jimmunol.1101746 doi: 10.4049/jimmunol.1101746 [29] 周书林, 常维佳, 姜玉新, 等. 重组粉尘螨Ⅱ类变应原致敏小鼠肺部变应性炎症模型的建立[J]. 中国媒介生物学及控制杂志, 2012, 23(3): 221−224. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 379
- HTML全文浏览量: 182
- 被引次数: 0
