


-
由稻黄单胞菌(Xanthomonas oryzae)的致病变种水稻细菌性条斑病菌(X. oryzae pv. oryzicola)引起的条斑病是水稻生产上重要的细菌病害,严重威胁中国水稻的育种和生产安全。水稻细菌性条斑病是中国禁止入境的检疫性病害[1-3]。因此,开发快速、准确、高效的水稻细菌性条斑病菌的检测技术,对水稻细菌性条斑病菌及时进行有效的防治,控制水稻细菌性条斑病菌病害的发生及发展,对减少经济损失具有十分重要的意义。围绕水稻细菌性条斑病菌,前人已经开发了多种分子检测技术。一些研究表明实时荧光PCR检测技术可用于检测水稻细菌性条斑病[4-11]。重组酶聚合酶扩增(RPA)技术是近年发展起来的一种代表性的等温扩增技术[12],该方法在37~42 ℃的恒定温度条件下,15~20 min即可完成无需变温热循环的核酸扩增[13-14]。实时荧光RPA(Exo-RPA)是一种通过添加荧光基团(carboxy fluorescein,FAM)和淬灭基团(black hole quencher,BHQ)探针、四氢呋喃(tetrahydrofuran,THF)位点和3′端阻断物的实时监控荧光值变化的检测技术。Exo-RPA原理是exo酶识别切割探针的THF位点,将荧光基团和淬灭基团裂解,释放荧光产生荧光信号来实时监测荧光值的变化,得到荧光扩增曲线[15]。LFD-RPA(lateral flow dipstick-recombinase polymerase amplification)技术是在RPA技术的基础上加入核酸外切酶Ⅳ(endonuclease Ⅳ,nfo)、nfo荧光探针以及附着于3′端生物素(Biotin)标记的下游引物对RPA产物进行检测。基于双抗体夹心原理,在nfo-RPA反应中,探针是关键的组成部分,通常由寡核苷酸组成,包括四氢呋喃(THF)残基、5′端荧光基团(FAM)、3′端修饰基团及下游引物5′端标记的抗原性生物素。在反应过程中,nfo酶识别并裂解探针,产生新的3′−羟基,作为聚合酶延伸的启动位点。随后,与生物素标记的下游引物反应,使扩增产物同时携带FAM和Biotin标记[16]。随后,扩增产物被捕获在加样垫上包被有抗FAM特异性抗体的胶体金颗粒,这些颗粒与扩增产物中的FAM标记结合,形成带有标记的扩增产物——胶体金颗粒复合物。接着,复合物在侧流层析试纸条上进行层析。测试线上包被有链霉亲和素(SA),与FAM标记的扩增产物结合进行显色,形成阳性测试条带;与此同时,控制线上包抗体,当胶体金颗粒经过时,与抗体结合产生阳性控制条带[13],实现目标生物分子的检测。LFD-RPA具有操作简单、快速、无需特殊仪器和试剂等优点,检测结果可在5~10 min直接用肉眼看到,适合在现场和资源受限的情况下应用[17]。目前该重组酶聚合酶技术已被广泛应用于病原体鉴定[18-21]、环境监测[20]、食品安全监测[22-23]和疾病诊断[24]等领域。本研究对RPA引物及探针组合进行筛选,同时对Exo-RPA和LFD-RPA的反应条件(检出限、时间和温度)进行优化,最后评估Exo-RPA和LFD-RPA方法的特异性,完成针对水稻细菌性条斑病菌的Exo-RPA及LFD-RPA快速检测方法的建立。
-
准备水稻细菌性条斑病菌(Xanthomonas oryzae pv. oryzicola)、水稻白叶枯病菌(X. oryzae pv. oryzae)和果胶杆菌(Pectobacterium aroidearum)L6、斯氏泛菌(Pantoea stewartii)SJM1和健康水稻叶片等实验材料,提取DNA用于检测。水稻细菌性条斑病菌特异性序列DNA标准品重组质粒pUC57-AvrRxo1由金斯瑞生物公司根据Genbank序列XOCgx_RS03090合成。
-
引物和探针由上海生工生物工程有限公司合成,并稀释至10 μmol·L−1备用;LB液体培养基(胰蛋白胨10 g,酵母粉提取物5 g,NaCl 10 g,蒸馏水1.0 L,pH7.4),LB固体培养基(LB液体和琼脂20 g);pMD18-T载体[宝日医生物技术(北京)有限公司,北京];大肠杆菌(Escherichia coli)DH5α感受态细胞(上海唯地生物技术有限公司,上海);PCR扩增产物回收纯化试剂盒(重鼎生物技术有限公司,浙江宁波),快速质粒小提试剂盒(天根生化科技有限公司,北京);基因组DNA提取试剂盒(天根生化科技有限公司,北京);多糖多酚植物基因组DNA提取试剂盒(天根生化科技有限公司,北京);Taq DNA 聚合酶(天根生化科技有限公司,北京);TwistAmpTM 试剂盒(TwistDx公司, 英国);RAA-nfo核酸扩增试剂(试纸条型)(杭州众测生物科技有限公司);DL500分子质量标记[宝日医生物技术(北京)有限公司,北京]。蓝光切胶仪、PCR仪、Micro Drop超微量分光光度计和QuantStudioTM 5实时荧光定量PCR仪。
-
使用细菌基因组DNA提取试剂盒操作说明书所描述的步骤提取水稻细菌性条斑病菌DNA。使用Micro Drop超微量分光光度计对DNA质量浓度进行检测。最终将 DNA 质量浓度稀释至94.974 ng·μL−1,用于后续试验。利用EcoRI/SacI技术将基因克隆到pUC57中,重组质粒名为pUC57-AvrRxo1;使用快速质粒小提试剂盒(TIANprep Rapid Mini Plasmid Kit)提取和纯化pUC57-AvrRxo1质粒。采用RPA方法检测重组质粒pUC57-AvrRxo1是否插入水稻细菌性条斑病菌297 bp的特异性基因序列XOCgx_RS03090。RPA引物序列为XocRPA-3F(5′-ACGCACTCGAATTTCATAAGAAGCTTAAGGA-3′),XocRPA-3R(5′-GTACATCCGTTCATATATGTCGTGTCGCTCT-3′)。根据TwistAmp® Basic kit试剂盒说明书进行Basic-RPA方法检测,反应体系:总反应体积50 μL,包括向装有冻干粉的反应管中加入29.5 μL再水化缓冲液(Rehydration buffer)、正反向引物(10 μmol·L−1)各2.4 μL和ddH2O 12.2 μL,反应体积达到46.5 μL,同时设置阴性对照(NC);将混合液旋涡振荡简短离心后,用移液枪将1 μL质粒模板加入混合液中;将2.5 μL醋酸镁离子(280 mmol·L−1)加在管盖中,小心关闭管盖,用微型离心机将醋酸镁离子(280 mmol·L−1) 离心到混合液中;立即放入PCR仪器中,在39 ℃下等温扩增反应;反应进行4 min后,取出反应管颠倒混匀并短暂离心,再放回PCR仪器继续反应26 min。RPA扩增后的产物,使用3%(1.2 g琼脂糖,40 mL 1×TBE缓冲液)琼脂糖凝胶电泳进行结果检测,并使用MEGA v11和Snap Gene软件对产物测序后的序列进行序列比对。重组质粒作为标准核酸,用超微量分光光度计测定质粒拷贝数。标准核酸质粒拷贝数计算结果为4.96×1010 copies·μL−1。
$$ 质粒拷贝数({\mathrm{copies}}·\text{μL}^{−1})=(\rho \times 6.02\times10^{23}\times10^{−9})/{\mathrm{M}} 。 $$ 式中,ρ表示重组质粒的质量浓度(ng·μL−1),M表示平均分子质量。
-
根据水稻细菌性条斑病菌的特异性基因序列XOCgx_RS03090并基于RPA引物和荧光探针的设计原则,利用软件Oligo v7.0设计筛选获得RPA引物XocRPA-3和Exo-RPA及LFD-RPA探针组合。引物和探针由上海生工生物工程有限公司合成。
-
使用水稻细菌性条斑病菌重组质粒DNA作为扩增反应模板,按照TwistAmp®Basic Kit试剂盒说明书进行Basic-RPA扩增。对水稻细菌性条斑病菌XocRPA-3引物的Basic-RPA特异性扩增产物进行回收纯化后,与pMD18-T Vector在16 ℃条件下连接低温连接仪,转化到DH5α感受态细胞中,37 ℃过夜培养。挑取白色单克隆菌落转至含氨苄青霉素的LB液体培养基中,37 ℃、180 r·min−1 恒温振荡培养60 min。肉眼可见浑浊后,经Sanger方法测序,使用MEGA v11软件将测序结果与水稻细菌性条斑病菌的特异性基因序列XOCgx_RS03090进行比对。
-
根据TwistAmpTM exo试剂盒说明书,进行Exo-RPA扩增反应。在50 μL的反应体积中,分别将29.5 μL再水化缓冲液(Rehydration buffer)、正反向引物(10 μmol·L−1)各2.1 μL、荧光探针(10 μmol·L−1)0.6 μL和 ddH2O 12.2 μL 加入到含有冻干粉的反应管中,反应体积为46.5 μL,同时设置阴性对照(NC),将混合液旋涡振荡简短离心后,用移液枪转移到0.2 mL PCR反应管中,将1 μL质粒DNA模板加入混合液中,随后将2.5 μL醋酸镁离子(280 mmol·L−1)加在管盖中,小心关闭管盖,用微型离心机将醋酸镁离子(280 mmol·L−1)离心到混合液中,并立即放入QuantStudioTM 5实时荧光定量PCR仪中,在39 ℃下恒温扩增20 min,每30 s采集1次荧光信号,观察读取荧光信号。为了避免光照影响荧光染料的强度,操作过程中探针要注意避光。
-
按1.6所述配置Exo-RPA反应体系,将水稻细菌性条斑病菌质粒标准品按10倍梯度依次稀释,经预实验后,分别设置4.96、4.96×101、4.96×102、4.96×103和4.96×104 copies·μL−1 5个梯度的1 μL质粒DNA模板,进行反应灵敏度检测。每个质粒DNA模板灵敏度检测设置3次重复,同时设置1个阴性对照(NC),总计进行16个RPA反应。使用QuantStudioTM5 实时荧光定量PCR仪,在39 ℃下恒温扩增20 min,每30 s采集1次荧光值,观察读取荧光信号,确认是否对水稻细菌性条斑病菌进行了有效扩增,进而确定实时荧光定量检测方法对Exo-RPA检测的灵敏度。
-
按1.6所述配置Exo-RPA反应体系。使用以1 μL 4.96×102 copies·μL−1水稻细菌性条斑病菌质粒为扩增模板的Exo-RPA反应体系,放入QuantStudioTM 5 实时荧光定量PCR仪器中,分别在35、37、39、41、43 ℃共5个温度梯度,恒温扩增20 min,同时检测荧光信号强度,确定最佳扩增温度。
-
使用细菌基因组DNA提取试剂盒操作说明书所描述的步骤提取水稻细菌性条斑病菌、水稻白叶枯病菌、果胶杆菌L6和斯氏泛菌SJM1的DNA。使用多糖多酚植物基因组DNA提取试剂盒提取健康的水稻嫩叶基因组DNA,然后在QuantStudio TM5荧光定量PCR仪器中进行反应,反应体系同1.6,RPA反应在最佳反应条件39 ℃恒温扩增20 min,每30 s采集1次荧光值,观察读取荧光信号,确认是否对水稻细菌性条斑病菌进行了有效扩增,进而确定 Exo-RPA 检测系统的特异性和实际检测效果。
-
使用水稻细菌性条斑病菌XOCgx_RS03090基因序列设计的引物,根据RAA-nfo核酸扩增试剂盒说明书进行LFD-RPA方法检测。在50 μL反应体积中,分别将25 μL A Buffer、正反向引物(2 μmol·L−1)各2.0 μL、荧光探针(2 μmol·L−1)0.6 μL和 ddH2O 12.9 μL加入到含有冻干粉的反应管中,再向反应管中加入5 μL DNA,最后向反应管管盖上加入2.5 μL B Buffer,盖上管盖,上下颠倒轻甩混匀5~6次,低速离心10 s,将反应管放入恒温为39 ℃的PCR仪中,孵育30 min。反应结束后,按照一次性核酸检测试纸条使用说明书进行操作。依据所需检测的样本量,提取对应数量的检测试纸条,并在吸收垫处加注明记号,每根检测试纸条限一次性使用;取8~10 μL核酸扩增产物,点在试纸条的样品垫区域;将试纸条的样品垫部分朝下,插入含有100 μL缓冲液(TE或水或PBS)的1.5 mL灭菌试管中,在3~5 min内观察并记录试纸条上的反应区域,得出检测结果。检测结果:如试纸条检测带和控制带均出现颜色变化,表示检测为阳性反应;若仅控制带显色,则结果为阴性。
-
将水稻细菌性条斑病菌的重组质粒按照10倍梯度依次稀释,经预实验后,水稻细菌性条斑病分别设置4.96×10−1、4.96、4.96×101、4.96×102和4.96×103 copies·μL−1 5个浓度梯度的质粒DNA模板,进行反应灵敏度检测。反应体系按1.10操作,每个质粒DNA模板浓度灵敏度检测设置3次重复,同时设置1个阴性对照(NC)。通过观察检测带的清晰度和颜色深度,确定LFD-RPA可检测到的最低反应浓度。
-
为确定最优反应温度,以4.96×103 copes·μL−1 DNA为模板,反应时间为30 min,温度优化试验的梯度设置:35、37、39、41、43 ℃,共5个温度梯度。另外,每个温度梯度均单独设置3个阴性对照,反应体系按1.10操作,每个温度梯度重复3次实验,反应进行扩增,将试纸条放入稀释液中进行测定观察结果。
-
为确定最优反应时间,以4.96×103 copes·μL−1 DNA为模板,反应温度为39 ℃,时间优化试验的梯度设置0、5、10、15、20 min共5个时间梯度。另外,每个时间梯度均独立设置1个阴性对照,反应体系按1.10操作,每个时间梯度重复3次实验。对反应进行扩增,将试纸条放入稀释液中进行测定观察结果。
-
为验证水稻细菌性条斑病菌LFD-RPA检测方法的特异性,以水稻细菌性条斑病菌、水稻白叶枯病菌、斯氏泛菌、果胶杆菌L6和水稻叶片基因组DNA为模板进行LFD-RPA扩增实验,ddH2O为阴性对照,反应体系按1.10操作,每个样品重复3次实验,39 ℃反应30 min进行LFD-RPA扩增,验证LFD-RPA方法的特异性。
-
根据RPA引物和Exo-RPA、LFD-RPA探针组合的设计原则和预试验验证结果,开发出一组水稻细菌性条斑病菌RPA引物和Exo-RPA、LFD-RPA探针组合。引物及探针组合具体序列如表1所示。
表 1 水稻细菌性条斑病菌RPA引物和探针信息
Table 1. Basic information of RPA primers and probe for X. oryzae pv. oryzicola
引物及探针
Primers & Probe类型 Category 序列 Sequences (3′-5′) XocRPA-F3 正向引物 The forward primer F: 5′-ACGCACTCGAATTTCATAAGAAGCTTAAGGA-3′ XocRPA-R3 反向引物 The reverse primer R: 5′-GTACATCCGTTCATATATGTCGTGTCGCTCT-3′ XocRPA-3-Exo-P 探针 Probe 5′-CGCCGATGCTGTCGGCAAAGAACGGTTCGA-[FAM-dT]--
THF--[BHQ-dT]-CGAGGAGCTGGAGAA-3′-spacer-3′XocRPA-3-LFD-p 探针 Probe 5′-FAM-CGCCGATGCTGTCGGCAAAGAACGGTTCGAT-
THF-TCGAGGAGCTGGAGAA-C-3′-spacer-3′ -
以水稻细菌性条斑病菌的XOCgx_RS03090基因重组质粒pUC57-AvrRxo1转入大肠杆菌DH5α宿主的菌液为模板,进行RPA产物扩增。 使用MEGA v11和Snap Gene软件将经Sanger法测得的RPA产物序列与GenBank下载的水稻细菌性条斑病菌的特异性基因序列XOCgx_RS03090进行比对,结果表明,包含水稻细菌性条斑病菌的特异性基因序列XOCgx_RS03090已成功插入重组质粒中。
-
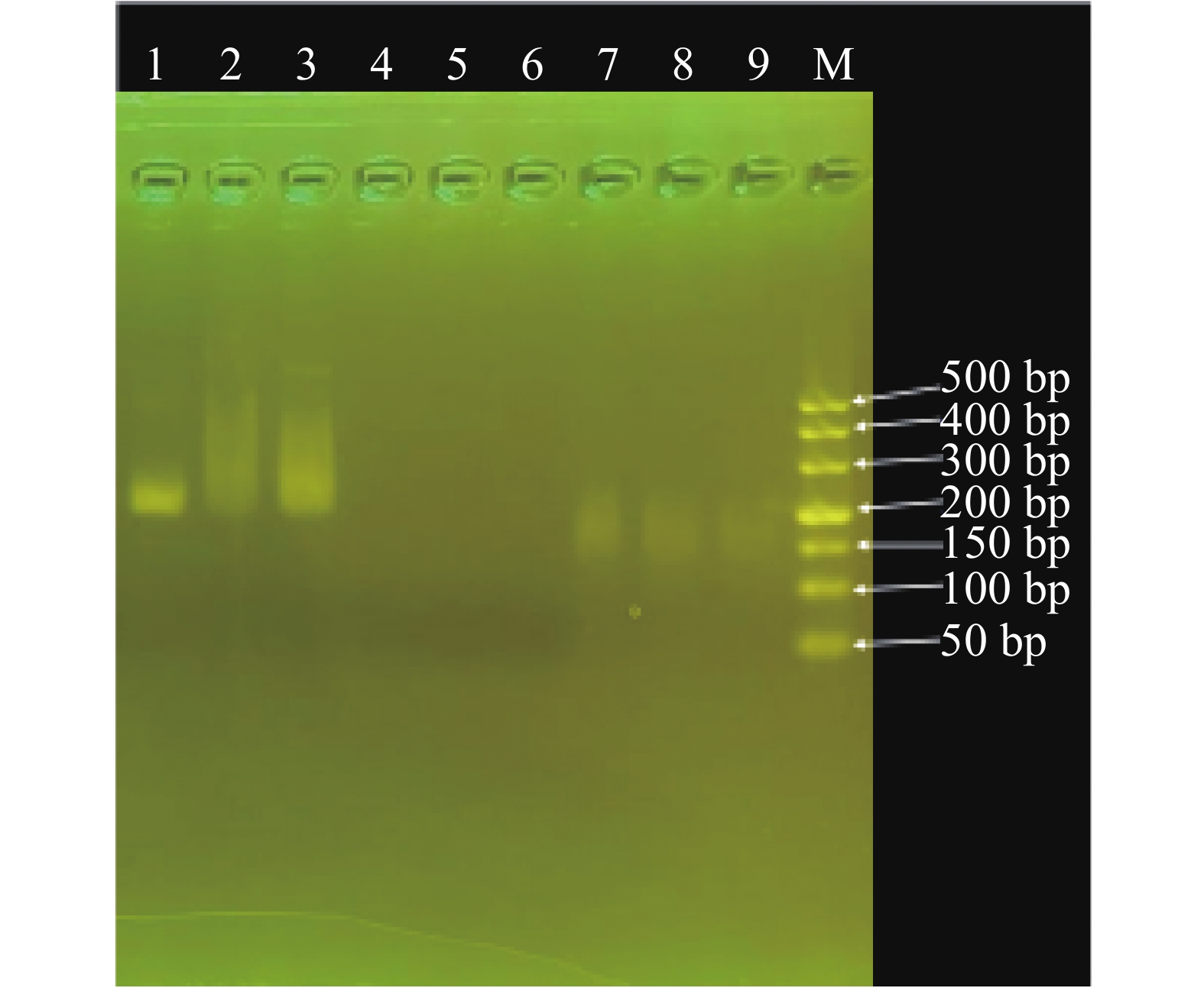
以重组质粒DNA(pUC57-AvrRxo1)为扩增模板,使用XocRPA-3引物进行Basic-RPA扩增,经3%琼脂糖凝胶电泳扩增后,扩增产物条带明亮、较清晰,扩增产物分子质量大小约为200 bp;构建重组质粒, 经过Sanger法测序,使用MEGA v11软件进行序列比对。结果表明,重组质粒DNA (pUC57-AvrRxo1)包含已插入目的片段,标准品质粒能用于RPA扩增反应体系的检测及优化(图1)。
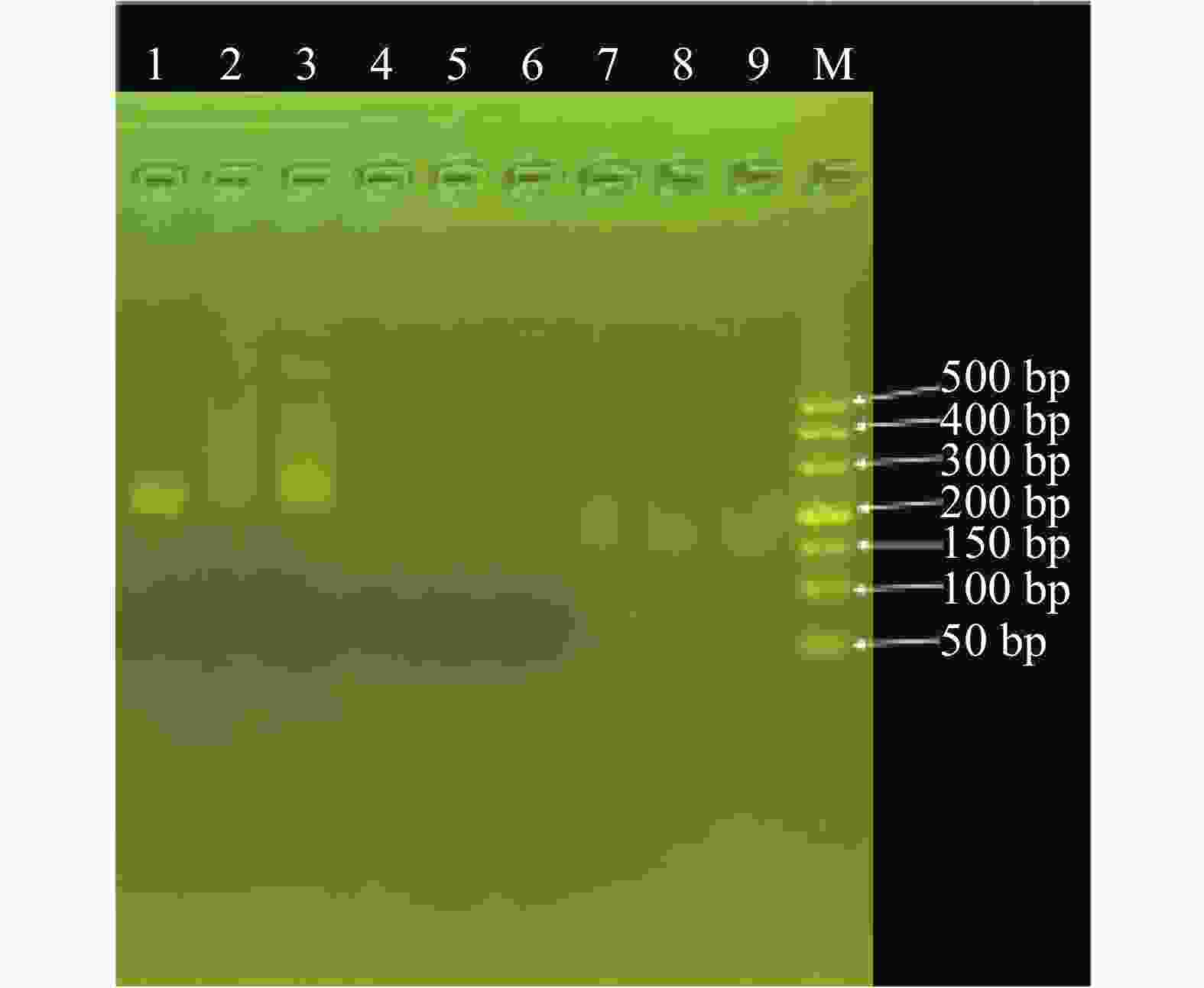
图 1 水稻细菌性条斑病菌重组质粒pUC57-AvrRxo1扩增产物的3%琼脂糖凝胶电泳图
Figure 1. 3% agarose gel electrophoresis of the pUC57-AvrRxo1 amplification product of the recombinant plasmid of X. oryzae pv. oryzicola
-
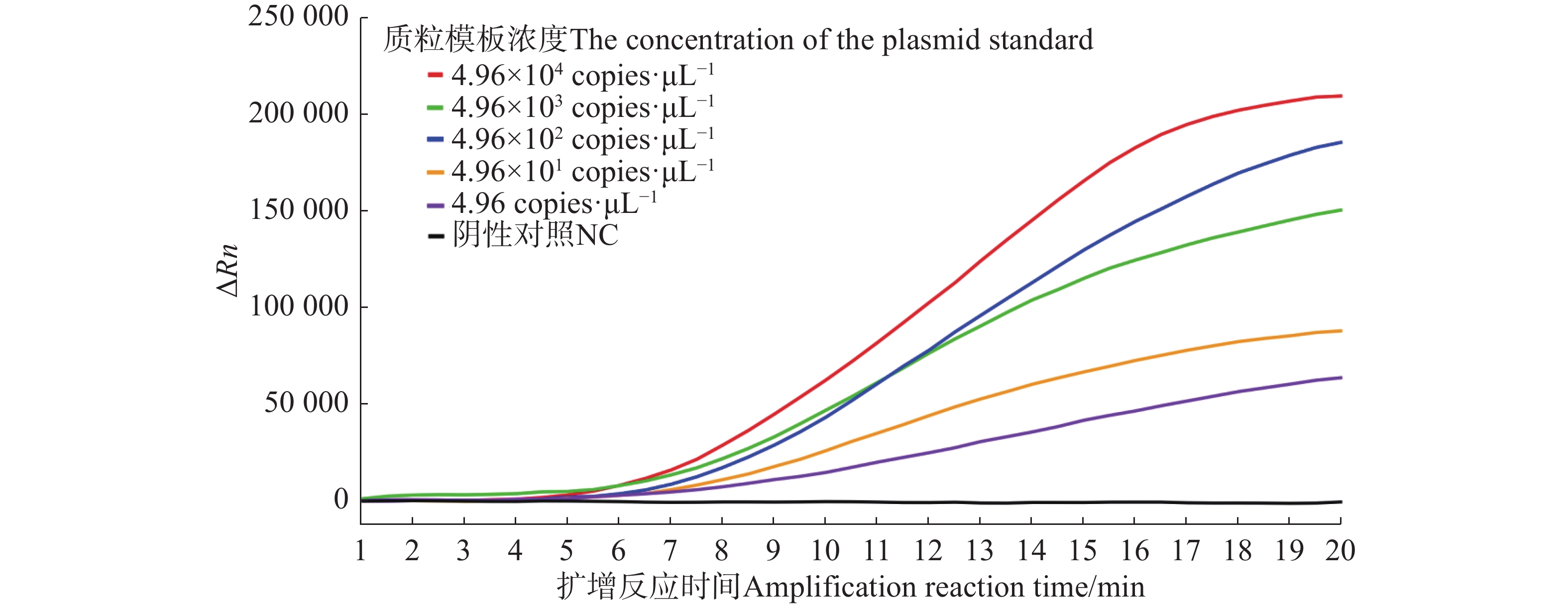
由荧光信号曲线(图2)可知,从采集荧光信号(即0 min)开始计时可在12 min之内检测到4.96~4.96×104 copies·μL−1 5个浓度的样品,最低检测限为4.96 copies·μL−1。
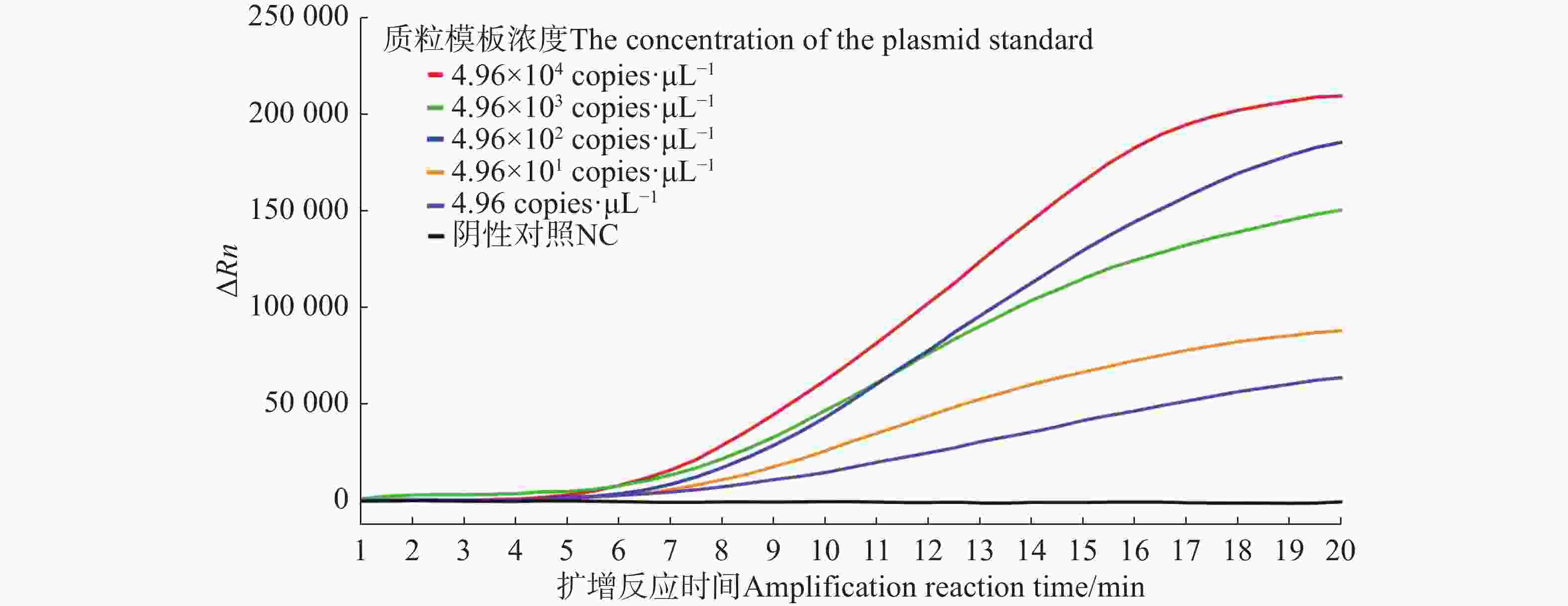
图 2 水稻细菌性条斑病菌不同浓度质粒标准品的Exo-RPA扩增曲线图
Figure 2. Exo-RPA amplification curves based on different DNA concentrations of X. oryzae pv. oryzicola plasmid standards
-
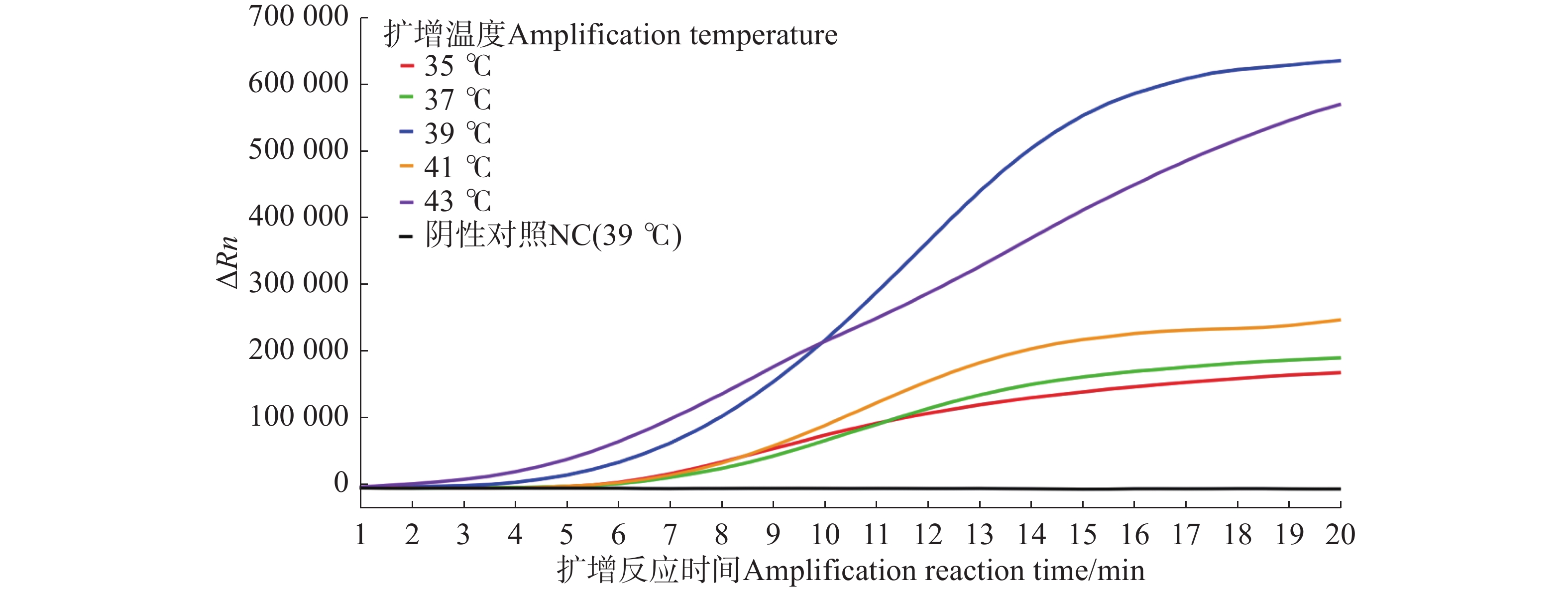
由荧光信号曲线(图3)可知,扩增5 min后,35、37、39、41、43 ℃均检测到荧光信号,且随着反应时间的增加荧光强度逐渐增强。综合考虑起峰时间和荧光信号强度,最终确定反应的最佳扩增温度为39 ℃。
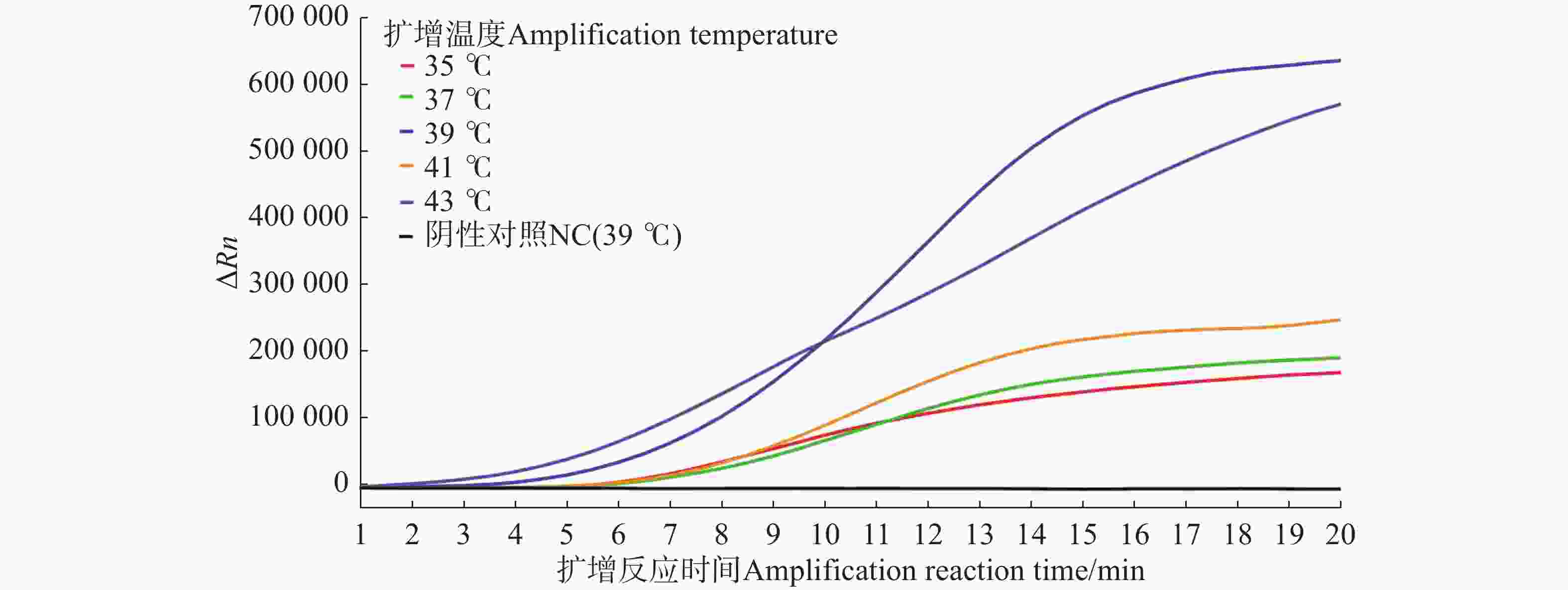
图 3 水稻细菌性条斑病菌不同温度条件下质粒标准品的Exo-RPA扩增曲线图
Figure 3. Exo-RPA application curves plot of X. oryzae pv. oryzicola plasmid standards under different temperature conditions
-
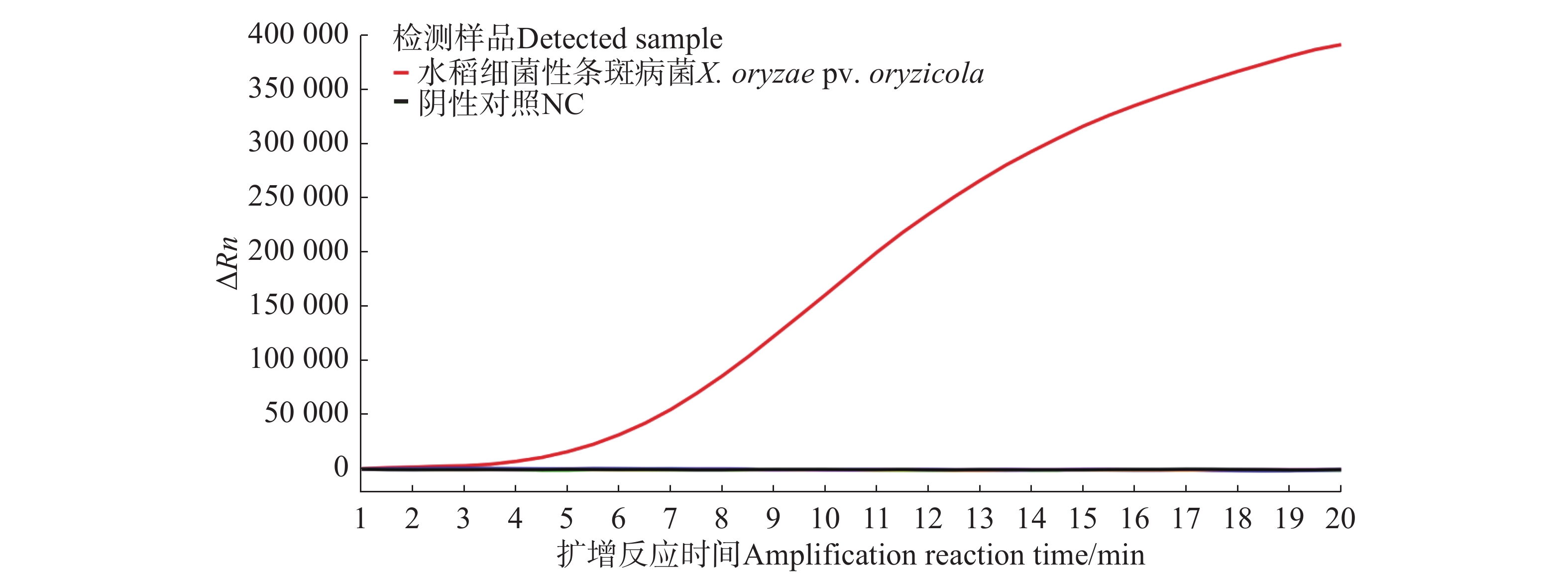
由荧光信号曲线(图4)可知,随着反应时间的增加,水稻细菌性条斑病菌样品在4 min后荧光强度逐渐增强,检测结果均为阳性,其他非目标样品在4 min后荧光强度无明显扩增现象,均为阴性,表明建立的Exo-RPA方法可以很好检测水稻细菌性条斑病菌。
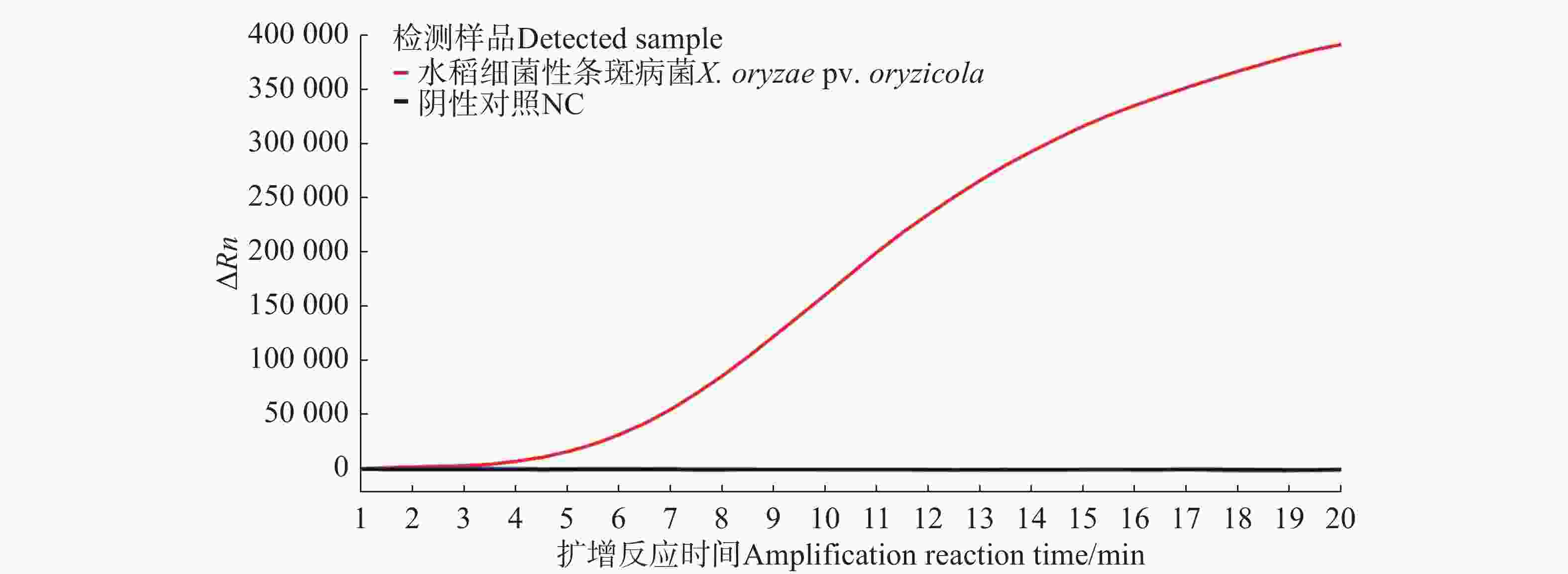
图 4 水稻细菌性条斑病菌Exo-RPA特异性验证和实际样品检测扩增曲线图
Figure 4. Specificity validation and amplification curve of actual sample detection of Exo-RPA of X. oryzae pv. oryzicola
-
将水稻细菌性条斑病重组质粒pUC57-AvrRxo1模板浓度对LFD-RPA检测方法的灵敏性进行评估,浓度稀释到4.96 copies·μL−1 均能看到清晰的检测带,LFD-RPA的最低检测限为4.96 copies·μL−1(图5)。
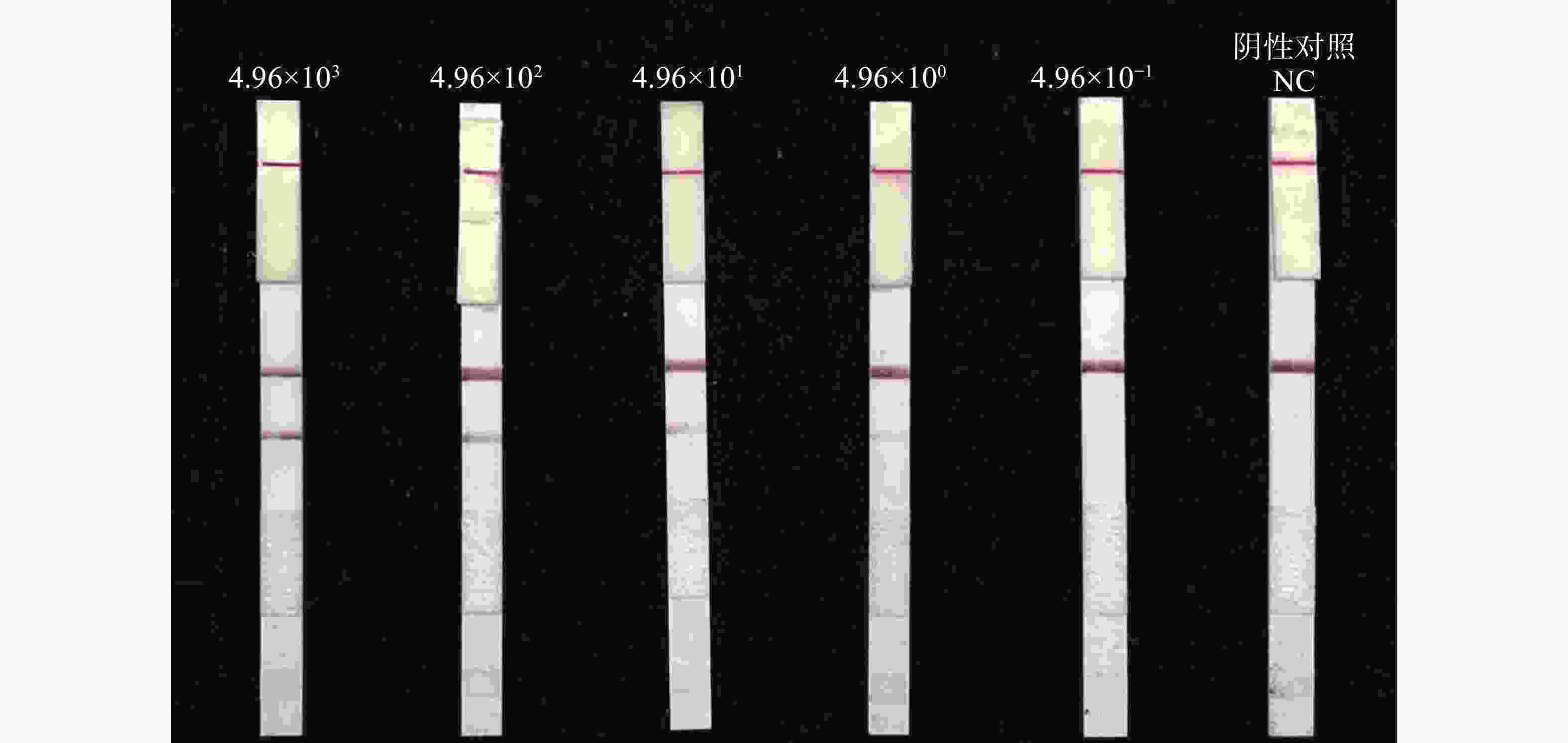
图 5 水稻细菌性条斑病菌质粒标准品pUC57-AvrRxo1 LFD-RPA灵敏度检测
Figure 5. LFD-RPA sensitivity determination of standard recombinant plasmid pUC57-AvrRxo1
-
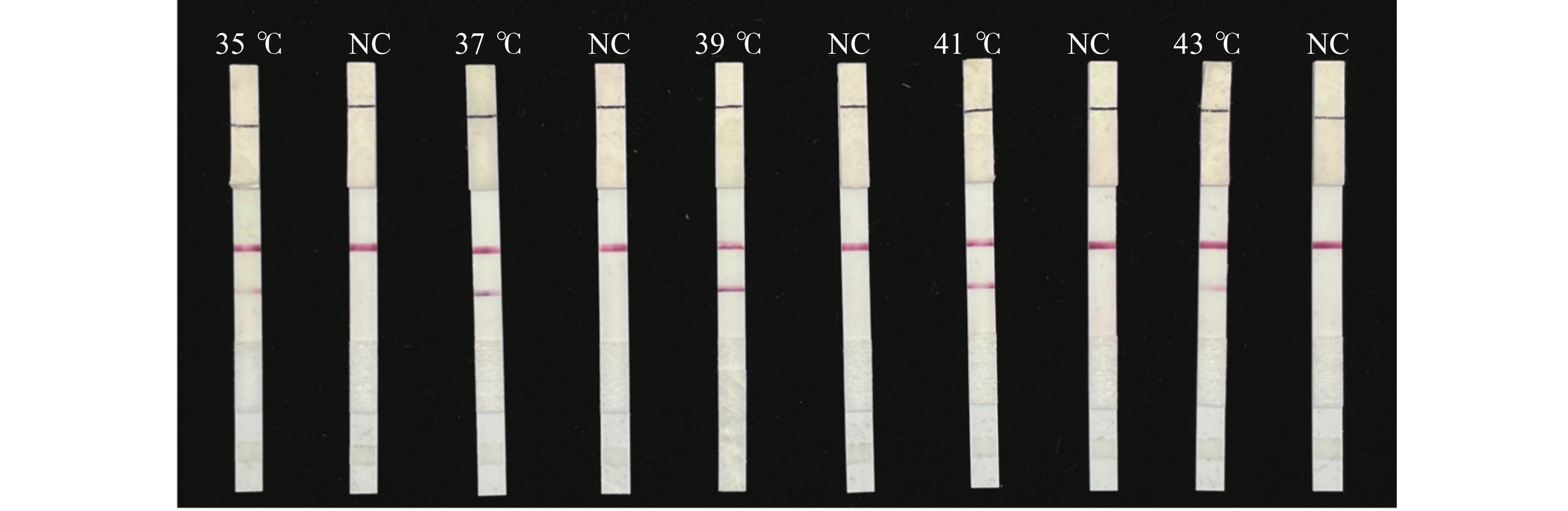
由图6可知,反应在35~43 ℃条件下均出现检测带,39 ℃检测带颜色最明显。这表明LFD-RPA能够在常温条件下进行检测,确定最佳反应温度为39 ℃。
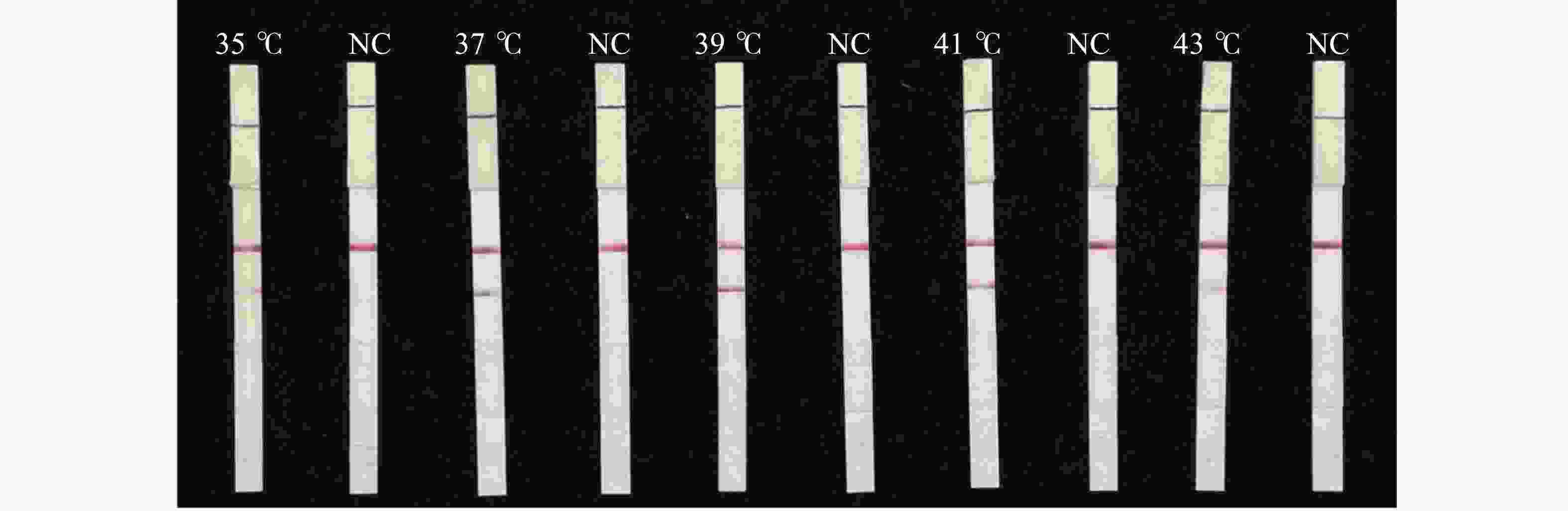
图 6 水稻细菌性条斑病菌LFD-RPA检测反应温度优化
Figure 6. Optimization of reaction temperature for LFD-RPA assay of X. oryzae pv. oryzicola
-
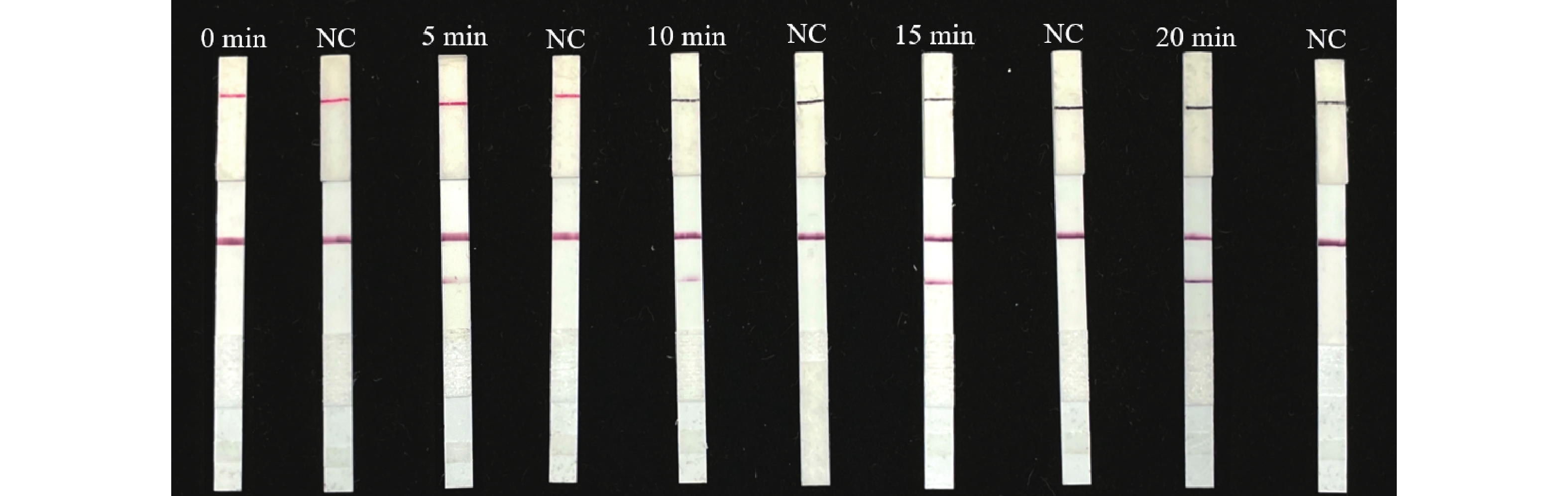
由图7可知,在反应5 min时即可在检测靶标样品的侧流层析试纸条上观察到阳性检测带,说明在5 min时所产生的扩增产物即可被试纸条法检出。此外,在5~20 min反应时间内均可观察到检测带,考虑到反应时间、扩增产物的稳定性及肉眼观察的清晰可辨别程度,确定最佳反应时间为20 min。

图 7 水稻细菌性条斑病菌LFD-RPA检测反应时间优化
Figure 7. Optimization of reaction time for LFD-RPA assay of X. oryzae pv.oryzicola
-
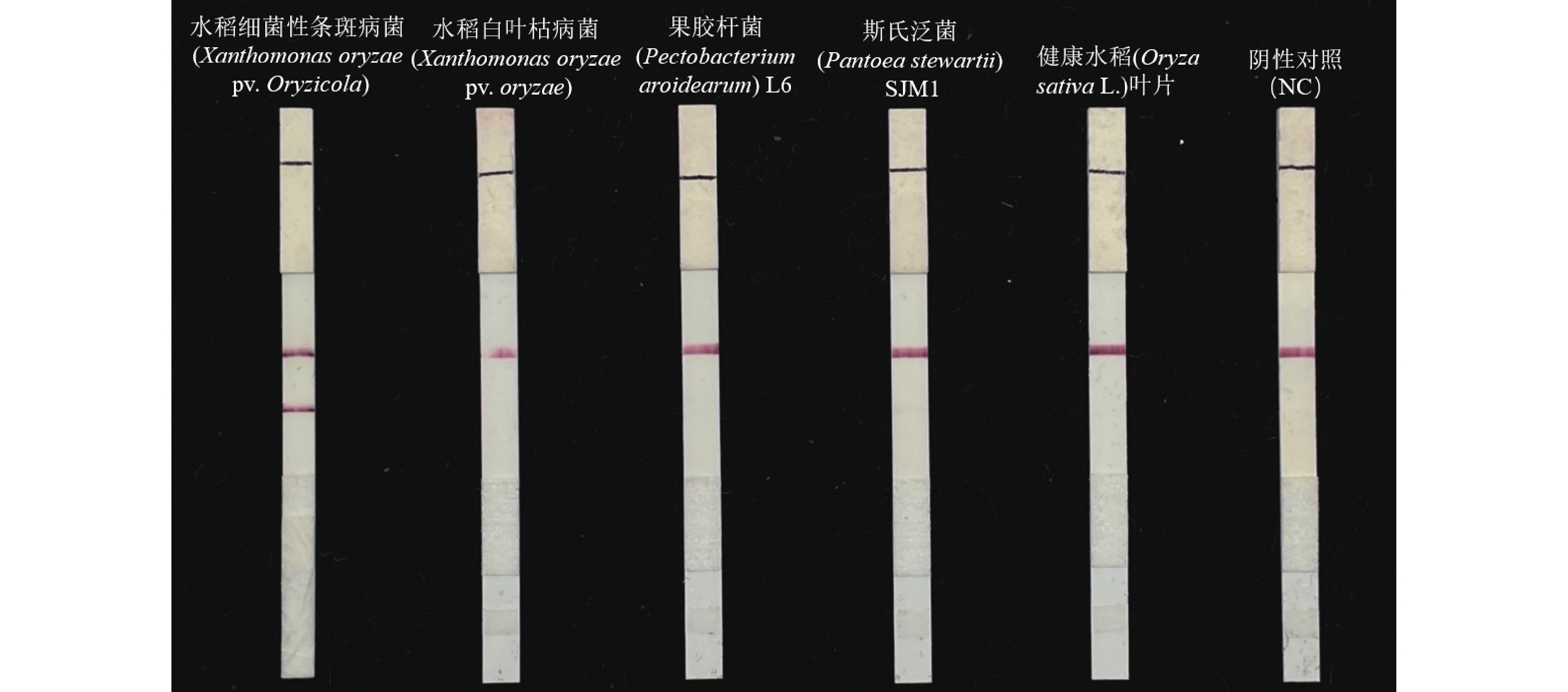
在39 ℃ 恒温及20 min扩增时间的最佳反应条件下,进行LFD-RPA方法针对水稻细菌性条斑病菌的检测特异性验证,只有检测水稻细菌性条斑病菌的LFD-RPA显示红色检测带,其他均未出现条带,结果为阴性(图8)。这说明本体系未与所选的其他病原菌出现交叉反应,表明本套nfo-RPA特异性引物及探针组合能够准确区分水稻细菌性条斑病菌及其他病原菌菌株,具有良好的特异性。

图 8 水稻细菌性条斑病菌特异性检测
Figure 8. RPA-LFD specificity validation and detection of actual samples of X. oryzae pv. oryzicola
-
水稻含有丰富的碳水化合物、蛋白质、维生素和矿物质等营养成分,是中国主粮作物。水稻细菌性条斑病是具有爆发性和破坏性的传染性病害。本研究通过水稻细菌性条斑病菌特异性基因序列XOCgx_RS03090设计RPA引物及Exo-RPA和LFD-RPA探针,建立了Exo-RPA荧光检测体系和LFD-RPA检测体系。将本实验设计的水稻细菌性条斑病菌RPA正反向引物XocRPA-3F与XocRPA-3R经过NCBI Basic Local Alignment Search Tool (
https://blast.ncbi.nlm.nih.gov/Blast.cgi )比对后,仅比对中的水稻细菌性条斑病菌基因组同源相似度达到100% (E-value=1×10−5)。这提示本研究团队设计的Exo-RPA引物具有仅针对水稻细菌性条斑病菌的特异性。目前,在检测水稻细菌性条斑病菌中,应用实时荧光PCR技术检测该病害的最低质粒DNA浓度为30. 6 fg·μL−1及菌悬浮液为1.0×103 cfu·mL−1,该方法虽然有高特异性,但检测成本相对较高[4]。应用Taqman荧光定量PCR检测水稻细菌性条斑病菌最低灵敏度为2.0×101 copies·μL−1,检测时长约为2 h,但该方法容易出现非特异性扩增,比较费时费力,成本高[6]。应用padlock探针和Macroarray技术相结合检测水稻细菌性条斑病菌,操作过程繁琐且检测时间长,检测成本高[8]。应用多重PCR检测水稻细菌性条斑病菌最低灵敏度为103 cfu·mL−1,但由于在同一反应中扩增多个目标序列,增加了交叉污染的可能性,引物设计难度大,非特异性产物可能增加,因为不同目标序列之间的扩增效率可能存在差异,多重PCR技术在定量分析方面可能存在一定难度[9]。应用PMA-PCR诊断技术检测水稻细菌性条斑病菌最低灵敏度为2×104 cfu·mL−1,但PMA-PCR涉及多个步骤,包括PMA处理、洗涤、PCR扩增和数据分析等[11]。这些步骤需要仔细的操作和适当的实验条件,因此相对于传统的PCR方法来说更为复杂。虽然PMA能够与死细胞中的DNA结合并阻止其PCR扩增,但某些情况下,死细胞可能无法被完全去除或区分。这可能导致假阳性结果的出现,特别是在样本中存在大量死细胞时。应用LAMP诊断技术检测水稻细菌性条斑病菌最低灵敏度为2.0×102 copies·μL−1,检测时长为40~60 min,具有高灵敏度,稳定性好,但检测时间长,引物设计复杂,需要针对靶基因的特定区域设计4条特异引物,增加了引物设计的难度,扩增过程涉及多个循环,容易发生非特异性扩增和污染问题,导致假阳性结果[6]。应用RPA/Cas12a检测水稻细菌性条斑病,通过荧光定量PCR仪检测水稻细菌性条斑病菌检测限为10 fg·μL−1,通过可视化检测,DNA检测限为1 fg·μL−1[25]。
-
RPA 检测技术反应迅速、操作简单,无需复杂仪器,能在便携式微型荧光定量 PCR 仪上恒温扩增现场检测,20 min内即可检测到极低质量浓度病原体特异性DNA,具有更高的灵敏度和准确性及更短的反应时间。本研究建立的水稻细菌性条斑病菌 Exo-RPA 检测体系与之前的快速核酸检测方法相比,具有可在接近人体温度或室温的39 ℃进行扩增反应,检测灵敏度更高、操作步骤更简便的优点,具备针对水稻细菌性条斑病菌的快速检测能力。
水稻细菌性条斑病菌重组酶聚合酶扩增快速检测方法的建立
DOI: 10.15886/j.cnki.rdswxb.20250004
 CSTR: 32425.14.j.cnki.rdswxb.20250004
CSTR: 32425.14.j.cnki.rdswxb.20250004
Recombinase polymerase amplification assay for rapid detection of Xanthomonas oryzae pv. oryzicola in rice
-
摘要: 为便于水稻细菌性条斑病菌(Xanthomonas oryzae pv. oryzicola,Xoc)的早期快速检测,本研究根据GenBank公开报道的水稻细菌性条斑病菌特异性基因序列(序列号:XOCgx_RS03090)设计了重组酶聚合酶扩增(RPA)引物、Exo荧光探针组合及nfo探针组合,并构建含有Xoc基因序列的重组质粒作为阳性对照;建立了水稻细菌性条斑病菌的重组酶聚合酶扩增法(Exo-RPA)和侧流层析试纸条检测法(LFD-RPA)的技术体系,并评估了上述两种不同终端RPA检测方法的灵敏度和特异性,通过优化反应温度、反应时间明确了其对RPA方法扩增效果的影响。结果表明,Exo-RPA和LFD-RPA检测法对水稻细菌性条斑病菌检测的灵敏度均为4.96 copies·μL−1,反应的最佳温度均为39 ℃;优化后的Exo-RPA方法在12 min内检测出水稻细菌性条斑病菌,优化后的LFD-RPA可以在5 min内检测出水稻细菌性条斑病菌;与其他植物病原菌菌株一起进行检测,RPA检测方法可特异地检测出目标病原菌。该方法灵敏度高,特异性强,检测速度快,有望在水稻细菌性条斑病病原监测与病害的早期诊断上得到应用。Abstract: For early and rapid detection of Xanthomonas oryzae pv. oryzicola (Xoc), the cause of bacterial leaf streak in rice, specific recombinase polymerase amplification (RPA) primers, and Exo fluorescent and nfo probes were designed and screened, respectively, according to the publicly available gene sequence XOCgx_RS03090 of Xoc genome sequence in GenBank. The recombinant plasmid containing Xoc gene was constructed as a positive control. One fluorescent Exo-RPA assay and the other lateral flow strip detection (LFD-RPA) assay were established for rapid detection of Xoc. The sensitivity and specificity of the RPA methods were assessed, and the amplification effects of the RPA methods were explored by optimizing the reaction temperature and reaction time. The results showed that the sensitivity of both the Exo-RPA and LFD-RPA assays for the detection of Xoc were both 4.96 copies·μL−1, and that the optimal temperatures for the reaction were both 39 ℃. Xoc was detected within 12 min by the optimized Exo-RPA method, and within 5 min by the optimized LFD-RPA. The Exo-RPA and LFD-RPA assays developed were highly specific and could specifically and uniquely identify Xoc among plant pathogens in test. Both the RPA methods are highly sensitive, specific and fast, which pose a high potential to the monitoring of the target pathogen and early diagnosis of the disease.
-
图 2 水稻细菌性条斑病菌不同浓度质粒标准品的Exo-RPA扩增曲线图
Fig. 2 Exo-RPA amplification curves based on different DNA concentrations of X. oryzae pv. oryzicola plasmid standards
图 3 水稻细菌性条斑病菌不同温度条件下质粒标准品的Exo-RPA扩增曲线图
Fig. 3 Exo-RPA application curves plot of X. oryzae pv. oryzicola plasmid standards under different temperature conditions
图 4 水稻细菌性条斑病菌Exo-RPA特异性验证和实际样品检测扩增曲线图
Fig. 4 Specificity validation and amplification curve of actual sample detection of Exo-RPA of X. oryzae pv. oryzicola
图 5 水稻细菌性条斑病菌质粒标准品pUC57-AvrRxo1 LFD-RPA灵敏度检测
注:浓度单位为质粒标准品浓度,copies·μL−1。
Fig. 5 LFD-RPA sensitivity determination of standard recombinant plasmid pUC57-AvrRxo1
Note: Unit of concentration, Concentration units of the plasmid copies·μL−1.
图 6 水稻细菌性条斑病菌LFD-RPA检测反应温度优化
注:35、37、39、41、43 ℃为检测反应温度;NC为阴性对照。
Fig. 6 Optimization of reaction temperature for LFD-RPA assay of X. oryzae pv. oryzicola
Note: The detection reaction temperatures are 35, 37, 39, 41 and 43 ℃; NC refers to the negative control.
图 7 水稻细菌性条斑病菌LFD-RPA检测反应时间优化
注:0、5、10、15、20 min为检测反应时间;NC为阴性对照。
Fig. 7 Optimization of reaction time for LFD-RPA assay of X. oryzae pv.oryzicola
Note: The detection reaction time is 0,5,10,15 and 20 min; NC refers to the negative control.
图 8 水稻细菌性条斑病菌特异性检测
Fig. 8 RPA-LFD specificity validation and detection of actual samples of X. oryzae pv. oryzicola
表 1 水稻细菌性条斑病菌RPA引物和探针信息
Table 1 Basic information of RPA primers and probe for X. oryzae pv. oryzicola
引物及探针
Primers & Probe类型 Category 序列 Sequences (3′-5′) XocRPA-F3 正向引物 The forward primer F: 5′-ACGCACTCGAATTTCATAAGAAGCTTAAGGA-3′ XocRPA-R3 反向引物 The reverse primer R: 5′-GTACATCCGTTCATATATGTCGTGTCGCTCT-3′ XocRPA-3-Exo-P 探针 Probe 5′-CGCCGATGCTGTCGGCAAAGAACGGTTCGA-[FAM-dT]--
THF--[BHQ-dT]-CGAGGAGCTGGAGAA-3′-spacer-3′XocRPA-3-LFD-p 探针 Probe 5′-FAM-CGCCGATGCTGTCGGCAAAGAACGGTTCGAT-
THF-TCGAGGAGCTGGAGAA-C-3′-spacer-3′ 下载: 导出CSV
下载: 导出CSV
-
[1] Wang W, Mauleon R, Hu Z, et al. Genomic variation in 3, 010 diverse accessions of Asian cultivated rice [J]. Nature., 2018, 557(7703): 43−49. https://doi.org/10.1038/s41586-018-0063-9 doi: 10.1038/s41586-018-0063-9 [2] Zhang L B, Zhu Q, Wu Z Q, et al. Selection on grain shattering genes and rates of rice domestication [J]. The New Phytologist, 2009, 184(3): 708−720. https://doi.org/10.1111/j.1469-8137.2009.02984.x doi: 10.1111/j.1469-8137.2009.02984.x [3] Ji Z Y, Zakria M, Zou L F, et al. Genetic diversity of transcriptional activator-like effector genes in Chinese isolates of Xanthomonas oryzae pv. Oryzicola [J]. Phytopathology., 2014, 104(7): 672−682. https://doi.org/10.1094/PHYTO-08-13-0232-R doi: 10.1094/PHYTO-08-13-0232-R [4] 廖晓兰, 朱水芳, 赵文军, 等. 水稻白叶枯病菌和水稻细菌性条斑病菌的实时荧光 PCR 快速检测鉴定[J]. 微生物学报, 2003, 43(5): 626. https://doi.org/10.13343/i.cnki.wsxb.2003.05.013 doi: 10.13343/i.cnki.wsxb.2003.05.013 [5] 韩阳, 张丽辉, 王永吉, 等. 水稻细菌性条斑病菌的实时荧光PCR检测技术研究[J]. 云南农业大学学报(自然科学), 2012, 27(3): 315−320. [6] 白飒. 检测西瓜噬酸菌和稻黄单胞菌的环介导等温扩增技术及荧光定量PCR方法的建立[D]. 南京: 南京农业大学, 2013. [7] 张华, 姜英华, 胡白石, 等. 利用PCR技术专化性检测水稻细菌性条斑病菌[J]. 植物病理学报, 2008, 38(1): 1−5. https://doi.org/10.3321/j.issn:0412-0914.2008.01.001 doi: 10.3321/j.issn:0412-0914.2008.01.001 [8] 田艳丽, 孟军, 范家勤, 等. 利用padlock 探针结合 Macroarray 技术对几种重要植物病原细菌的高通量检测[C]// 第四届中国植物细菌病害学术研讨会 论文集. 北京: 中国植物病理学会, 2008. [9] 莫瑾, 王哲, 周慧平, 等. 利用多重PCR技术快速检测4种水稻病原细菌[J]. 植物保护, 2021, 47(3): 160−164. [10] 刘雅婷, 岳凯, 李小林, 等. 水稻上两种种传病害的多重PCR检测试剂盒及其专用引物和多重PCR检测方法: CN201510292152.8[P]. 2015-06-02. [11] 于贵戌, 赵玉强, 姜培, 等. PMA-PCR方法进行水稻细菌性条斑病菌细胞活性检测的建立与初步应用[J]. 植物病理学报, 2023, 53(5): 944−949. [12] Yang B, Kong J, Fang X. Bandage like wearable flexible microfluidic recombinase polymerase amplification sensor for the rapid visual detection of nucleic acids [J]. Talanta, 2019, 204: 685−692 https://doi.org/10.1016/j.talanta.2019.06.031 doi: 10.1016/j.talanta.2019.06.031 [13] Piepenburg O, Williams C H, Stempie D L, et al. DNA detection using recombination proteins [J]. PLoS biology, 2006, 4(7): e204. https://doi.org/10.1371/journal.pbio.0040204 doi: 10.1371/journal.pbio.0040204 [14] 王季秋, 赵欣, 王照, 等. 实时荧光定量PCR检测技术及其在病原微生物检测中的应用[J]. 中国地方病防治杂志, 2016, 31(5): 517−519. [15] Euler M, Wang Y, Heidenreich D, et al. Development of a panel of recombinase polymerase amplification assays for detection of biothreat agents [J]. Journal of Clinical Microbiology, 2013, 51(4): 1110−1117. https://doi.org/10.1128/JCM.02704-12 doi: 10.1128/JCM.02704-12 [16] Daher R K, Stewart G, Boissinot M, et al. Recombinase polymerase amplification for diagnostic applications [J]. Clinical Chemistry, 2016, 62(7): 947−958. https://doi.org/10.1373/clinchem.2015.245829 doi: 10.1373/clinchem.2015.245829 [17] Yang X, Dong Y, Ma C, et al. Establishment of a visualized isothermal nucleic acid amplification method for on-site diagnosis of acute hepatopancreatic necrosis disease in shrimp farm [J]. Journal of fish Diseases., 2021, 44(9): 1293−1303. https://doi.org/10.1111/jfd.13388 doi: 10.1111/jfd.13388 [18] Behrmann O, Bavhmann I, Spiegel M, et al. Rapid detection of SARS-CoV-2 by low volume real-time single tube reverse transcription recombinase polymerase amplification using an exo probe with an internally linked quencher (Exo-IQ) [J]. Clinical Chemistry, 2020, 66(8): 1047−1054. https://doi.org/10.1093/clinchem/hvaa116 doi: 10.1093/clinchem/hvaa116 [19] Chi Y K, Zhao W, Ye M, et al. Evaluation of recombinase polymerase amplification assay for detecting Meloidogyne javanica [J]. Plant Disease, 2020, 104(3): 801−807. https://doi.org/10.1094/PDIS-07-19-1473-RE doi: 10.1094/PDIS-07-19-1473-RE [20] Xu H, Xia A, Wang D, et al. An ultraportable and versatile point-of-care DNA testing platform [J]. Science Advances, 2020, 6(17): eaaz7445. https://doi.org/10.1126/sciadv.aaz7445 doi: 10.1126/sciadv.aaz7445 [21] Strayer-Scherer A, Jones JB, Paret ML. Recombinase polymerase amplification assay for field detection of tomato bacterial spot pathogens [J]. Phytopathology, 2019, 109(4): 690−700. https://doi.org/10.1094/PHYTO-03-18-0101-R doi: 10.1094/PHYTO-03-18-0101-R [22] Kissenkotter J, Bohlken-Fascher S, Forrest M S, et al. Recombinase polymerase amplification assays for the identification of pork and horsemeat [J]. Food Chemistry, 2020, 322: 126759. https://doi.org/10.1016/j.foodchem.2020.126759 doi: 10.1016/j.foodchem.2020.126759 [23] Geng Y, Tan K, Liu L, et al. Development and evaluation of a rapid and sensitive RPA assay for specific detection of Vibrio parahaemolyticus in seafood [J]. BMC Microbiology, 2019, 19(1): 186. https://doi.org/10.1186/s12866-019-1562-z doi: 10.1186/s12866-019-1562-z [24] Zhang L, Peng J, Chen J, et al. Highly sensitive detection of low-abundance BRAF v600E mutation in fine-needle aspiration samples by Zip recombinase polymerase amplification [J]. Analytical Chemistry, 2021, 93(13): 5621−5628. https://doi.org/10.1021/acs.analchem.1c00405 doi: 10.1021/acs.analchem.1c00405 [25] 张海敏. 基于CRISPR/Cas12a的水稻白叶枯病菌和细菌性条斑病菌快速检测方法研究[D]. 武汉: 华中农业大学, 2023. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1350
- HTML全文浏览量: 656
- PDF下载量: 79
- 被引次数: 0
