



-
三叶草斑潜蝇[Liriomyza trifolii(Burgess)]隶属双翅目(Diptera)、潜蝇科(Agromyzidae)、斑潜蝇属(Liriomyza),是危害全世界农作物和园艺作物的检疫性害虫,同时也是中国一种重要的入侵害虫[1-3]。该虫起源于北美洲,20世纪70年代后,随着国际贸易的发展,迅速在全球扩散[2]。2005年12月,三叶草斑潜蝇在中国广东省中山市蔬菜基地首次报道[4],次年4月在海南省海口市等4个地区陆续发现[5]。三叶草斑潜蝇寄主范围广泛,有20科300多种作物[6]。幼虫和成虫均能对寄主植物进行危害,雌成虫用产卵器刺穿植物叶片上的植物组织,并在植物叶片表面形成白色斑点;幼虫在寄主植物叶片内潜食,叶片表面形成一条不规则的白色“蛇状”虫道,损坏叶肉组织和影响叶绿素的正常合成,降低植物的光合作用,严重时可导致寄主植物叶片大量脱落直至整株枯萎[4, 6]。三叶草斑潜蝇为多食性害虫,具有寄主范围广、适应性强、世代重叠严重等特点,能在田间迅速扩散与蔓延[7]。豇豆[Vigna unguiculata (L.) Walp.]起源于非洲,一种喜温耐热蔬菜,是海南省冬季瓜菜特色支柱产业之一[8-10]。据调查发现,三叶草斑潜蝇在海南省田间主要危害豇豆、丝瓜等瓜菜,造成产量损失[5]。大量化学农药及其不合理的使用导致海南三叶草斑潜蝇产生抗药性[11],同时也造成食品安全和环境污染等问题。因此,发展三叶草斑潜蝇的绿色防控技术对保障人民的“菜篮子”具有重要意义。
过氧化氢酶(Catalase,CAT)是昆虫抗氧化酶系统中一种重要抗氧化酶,其主要作用是保护生物免受代谢过程中产生的活性氧(Reactive oxygen species,ROS)如过氧化氢(H2O2)、超氧阴离子自由基(O2−)等引起的氧化损伤[12-13]。各种环境胁迫条件,如高低温、紫外线和农药暴露等都会诱导ROS的产生,导致昆虫受到不可逆的氧化损伤[14-15]。昆虫为应对这些逆境胁迫会做出一些响应,如演化出复杂的抗氧化酶系统,包括过氧化氢酶、超氧化物歧化酶(Superoxide dismutase,SOD)和过氧化物酶(Peroxidase,POD)等,共同协同清除ROS[16]。一般情况下,昆虫抗氧化酶体系与ROS处于动态平衡状态,以此维持正常生理活动,使其体内不受自由基毒害[17]。目前研究证明,CAT在昆虫适应温度胁迫中发挥重要作用。短时温度胁迫CAT基因均有不同的表达情况,在针对黄野螟(Heortia vitessoides Moore)CAT基因对高温胁迫的响应研究中,发现热应激(35、37 ℃)胁迫2 h导致H2O2增加,能提高HvCAT基因的表达,通过RNAi干扰HvCAT基因证实了其参与抵抗热应激引起的氧化损伤[18]。粘虫(Mythimna separata)幼虫在高温(33、35、37 ℃)胁迫2 h时CAT基因表达上调,而39 ℃胁迫2 h表达量会出现下调现象,该结果表明粘虫的氧化胁迫反应可以应对一定的高温,但是不能处理过高的温度[19]。CAT基因在长时间温度胁迫的表达情况,桃蚜(Myzus persicae)在27、30、33、36 ℃分别暴露1、3、6、10 h,热胁迫导致CAT基因的转录水平上调[20]。同样,巴氏新小绥螨(Neoseiulus barkeri)在36、38、40 ℃进行2、4、6 h的转录表达分析,表明CAT基因在热应激下均表现出显著的上调[21]。通过对烟粉虱(Bemisia tabaci)的研究发现,CAT基因可以通过消除过量的ROS来帮助烟粉虱适应长期的平均高温(30、35 ℃)和短期的极高温(39、41 ℃)[22]。在35 ℃及以上的温度,CAT活性被诱导并赋予寄生蜂(Aphelinus asychis)耐热性,以抵抗致命的热应激[23]。
海南由于其独特的地理位置及气候特点,使得该地区气温常年高于中国其他地区。冬季瓜菜的害虫防治措施目前仍以化学农药为主,农药、高温会诱导害虫体内产生大量的ROS[17, 24]。相较于近缘物种美洲斑潜蝇、南美斑潜蝇等,三叶草斑潜蝇具有较强的温度适应性[25 − 26]和抗药性[11],它的形成可能与其抗氧化酶的表达调控有关。目前缺乏三叶草斑潜蝇抗氧化酶的研究报道,其CAT基因的序列结构和功能也尚不明确,因此,分析三叶草斑潜蝇CAT的理化性质、序列结构和表达模式有助于了解其分子功能特征。基于此,本研究克隆了三叶草斑潜蝇CAT基因,并进行了生物信息学等相关分析,利用实时荧光定量PCR技术测定了LtCAT基因在三叶草斑潜蝇不同发育阶段及不同温度胁迫处理下的表达模式,旨在为进一步研究三叶草斑潜蝇过氧化氢酶基因的生理功能和作用机制提供参考。
-
本实验所用三叶草斑潜蝇于2023年2月在海南省三亚市崖城镇(109°14′E,18°37′N)豇豆地中采集获得。通过采集带有三叶草斑潜蝇幼虫的豇豆叶片后,待化蛹后收集至果蝇管,成虫羽化后用质量分数1%的蜂蜜水进行饲喂,接入放置干净豇豆苗的昆虫200目纱帐饲养笼(长50 cm×宽50 cm×高50 cm)中进行室内继代饲养。室内饲养条件为温度(26 ± 1) ℃、相对湿度(65 ± 5)%、光照16L∶8D。
-
总RNA提取所用Trizol Reagent试剂购自美国赛默飞世尔科技公司;纯化PCR产物试剂盒(E.Z.N.A® Cycle-Pure Kit)购自美国Omega Bio-Tek公司;感受态细胞DH5α、普通cDNA合成试剂盒(PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit)和荧光cDNA反转录试剂盒(PrimeScriptTM RT reagent Kit with gDNA Eraser)均购自宝日医生物技术(北京)有限公司;DL2000 Plus DNA Marker、2×Rapid Taq Master Mix、5×min TA/Blunt-Zero Cloning Kit、ChamQ Universal SYBR qPCR Master Mix均购自南京诺唯赞生物科技股份有限公司;由北京擎科生物科技有限公司合成引物。
-
利用Trizol试剂提取三叶草斑潜蝇50头成虫总RNA。质量分数1%琼脂糖凝胶电泳检测RNA质量,Micro Drop分光光度计检测RNA浓度和纯度,保存于−80 ℃备用。按照普通反转录说明书完成cDNA合成,保存于−20 ℃备用。
通过NCBI的GenBank数据库下载7种昆虫的CAT基因序列作为查询序列(query),利用TBtools软件通过BLAST在三叶草斑潜蝇转录组数据(未发表)中进行比对,并且通过NCBI搜索保守结构域(Batch CD-Search)鉴定为三叶草斑潜蝇CAT基因序列,根据序列设计三叶草斑潜蝇全长引物(表1)。以三叶草斑潜蝇反转录cDNA为模板,采用2×Rapid Taq Master Mix进行PCR扩增。反应体系(50 μL):dd H2O 19 μL,上下游引物(10 μmol·L−1)各2 μL,cDNA 2 μL,2×Rapid Taq Master Mix 25 μL。PCR程序:94 ℃预变性3 min;94 ℃变性15 s,58 ℃退火15 s,72 ℃延伸2 min,共35个循环;72 ℃延伸5 min。对PCR产物进行1%琼脂糖凝胶电泳检测,并利用Cycle-Pure Kit纯化试剂盒对目标条带的PCR产物进行纯化,纯化PCR产物连接至5 min TA/Blunt-Zero载体,转化到DH5α感受态细胞中,接着在含有氨苄卡那霉素(AMP)的LA固体培养基上进行培养,并通过挑斑、摇菌,经菌液PCR确认为阳性后,菌液送上海生工生物工程有限公司测序。
表 1 本研究所用引物信息
Table 1. Primer information used in this study
引物名称
Primer引物序列(5′−3′)
Primer sequence(5′−3′)产物长度/bp
Product length/bp用途
UseLtCAT-F F:ATTCAGCCAGACGGATCAGC 1731 全长克隆
(RT-PCR)LtCAT-R R:ACTCAATAGACTTGCTTTGCTGG qLtCAT-F F:CATTCCAGAACGTGTCGTGC 148 荧光定量
(RT-qPCR)qLtCAT-R R:GCCCACAGTGGAGAAACGTA β-actin-F[27] F:TTGTATTGGACTCTGGTGACGG 73 β-actin-R[27] R:GATAGCGTGAGGCAAAGCATAA 注:使用Primer Premier 5.0软件设计引物。 Note: Primers were designed using Primer Premier 5.0 software. -
利用在线网站ProtParam(
https://web.expasy.org/protparam/ )进行相对分子质量、等电点、氨基酸残基等理化特性分析;利用在线网站ScanProsite(https://prosite.expasy.org/scanprosite/ )进行基因家族位点、核心结构域、近端活性位点等分析;使用DNAman 9.0软件进行序列可视化及多序列比对;利用在线网站SignalP-6.0(https://services.healthtech.dtu.dk/services/SignalP-6.0/ )进行信号肽、N−糖基化位点的预测分析;通过在NCBI数据库(https://www.ncbi.nlm.nih.gov/ )查找下载序列,采用邻接法(Neighbor-joining,NJ)在MEGA 11.0软件进行聚类分析(Bootstrap 1 000次) 绘制系统发育树;利用SWISS-MODEL在线网站(https://swissmodel.expasy.org/ )进行拉马钱德兰图绘制和蛋白质三维结构预测。 -
用于实时荧光定量PCR(RT-qPCR)的样品采集(每个处理各20头,设3个生物学重复)。不同温度处理样品:三叶草斑潜蝇2日龄蛹在13、17、25、33、37 ℃处理1 h、在17、25、33 ℃分别处理1、6、12 h。不同发育阶段样品:三叶草斑潜蝇3龄幼虫、2日龄蛹、1日龄成虫。样品处理后用液氮速冻,保存于−80 ℃备用。总RNA提取操作同1.3,按照荧光反转录(PrimeScript RT reagent Kit with gDNA Eraser)进行cDNA合成,将cDNA稀释100倍作为模板。反应体系(20.0 μL):cDNA模板2.0 μL,DEPC水7.2 μL,上下游引物(10 μmol·L−1)各0.4 μL,2×ChamQ Universal SYBR qPCR Master Mix 10.0 μL。PCR程序:95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火与延伸30 s,共40个循环。根据克隆获得的三叶草斑潜蝇CAT基因序列来设计荧光定量引物(表1),选择三叶草斑潜蝇β-actin[27]作为内参基因。以25 ℃室温处理作为对照,利用2−△△CT法[28]计算LtCAT基因在三叶草斑潜蝇不同发育阶段和不同温度胁迫处理的相对表达量。每个样品设3个生物学重复和3个技术重复。实时荧光定量数据利用Excel 2023软件进行计算与分析,并利用GraphPad Prism 9.0软件作图,利用SPSS 26.0软件中单因素ANOVA检验的Duncan氏新复极差法(P < 0.05)进行各个处理之间的差异显著性分析。
-
根据三叶草斑潜蝇全长引物进行PCR扩增,经质量分数1%琼脂糖凝胶电泳分析,获得单一条带,目的基因片段大小符合预期结果(图1)。将克隆获得的序列在NCBI数据库中进行BLAST分析,与地中海实蝇CAT基因同源性最高,达84.57%。三叶草斑潜蝇CAT基因序列包含1个
1542 bp的开放阅读框(Open Reading Frame,ORF),共编码513个氨基酸。对基因序列的保守结构域分析表明,LtCAT基因具有catalase家族保守结构域(22~504氨基酸位点),其中包含1个核心结构域(26~411氨基酸位点),该区域有6个CAT家族特征位点(39~62、102~120、123~140、142~160、307~334、339~365氨基酸位点);另外还具有1个近端活性位点(62~78氨基酸位点)以及1个近端血红素配体特征(352~360氨基酸位点)(图2)。
图 1 三叶草斑潜蝇CAT基因的RT-PCR产物电泳图
Figure 1. Electrophoresis map of RT-PCR product of CAT gene in Liriomyza trifolii

图 2 三叶草斑潜蝇CAT基因序列分析
Figure 2. Sequence analysis of CAT gene of Liriomyza trifolii
-
经ProtParam软件预测三叶草斑潜蝇CAT蛋白相对分子质量为57 762.19 Da,理论等电点为8.40,正、负电荷氨基酸残基(Arg+Lys)、(Asp+Glu)分别为59、56个,不稳定系数为26.72,脂肪系数为69.94,体外半衰期为30 h,分子式为C2589H3959N721O756S15。SignalP−6.0预测结果显示,三叶草斑潜蝇CAT基因无信号肽,推测其并非分泌蛋白,N−糖基化位点位于319氨基酸位点。
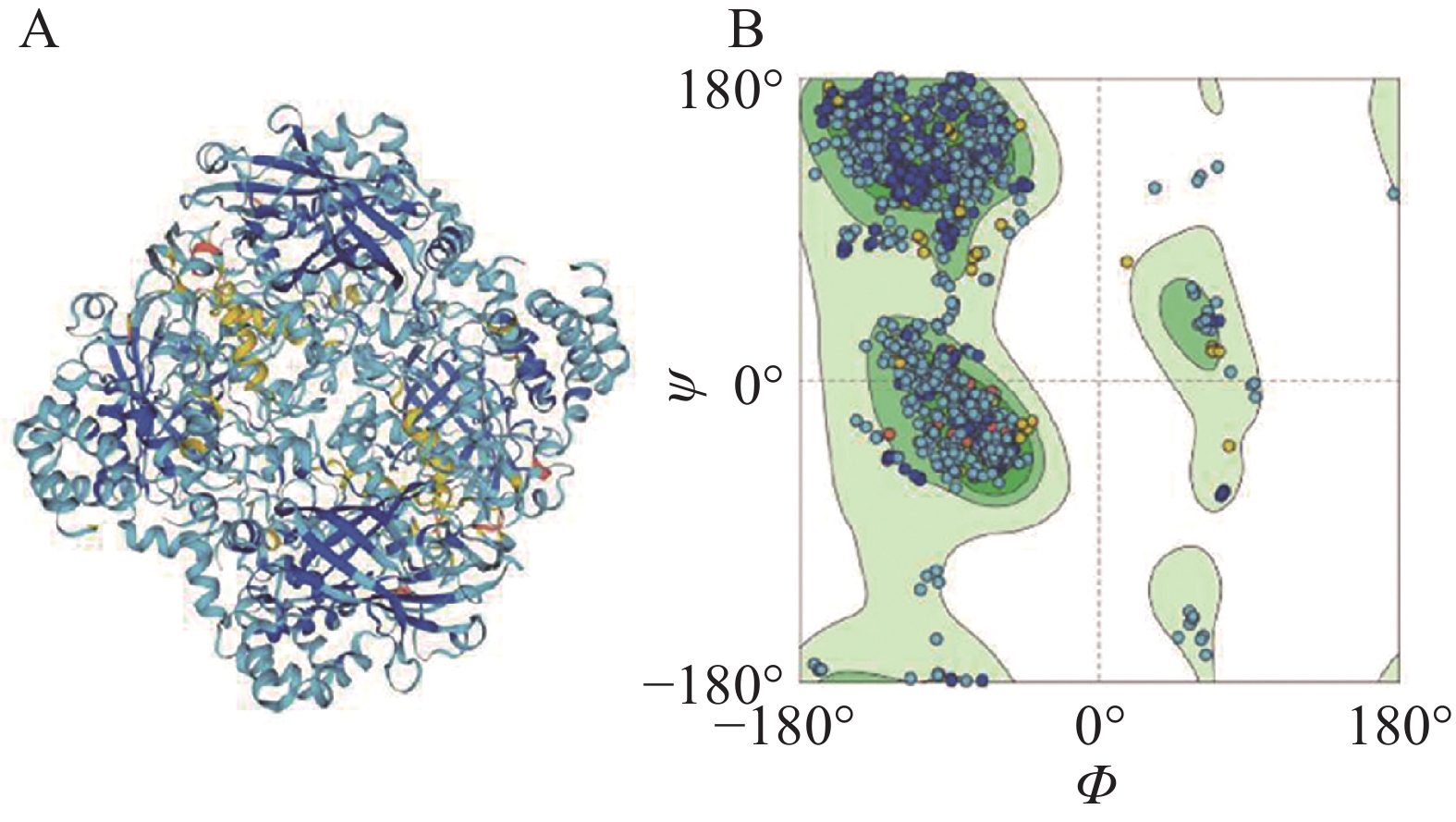
在SWISS-MODEL在线网站以3re8.1.A Catalase为模板,对LtCAT的蛋白结构3D模型预测,序列相似性为68.95%,模型质量评估值为0.85 ± 0.05,使用拉马钱德兰图对蛋白结构3D模型进行质量评估,结果表明该蛋白的预测3D结构模型评估质量与拉马钱德兰图的评价一致(图3)。
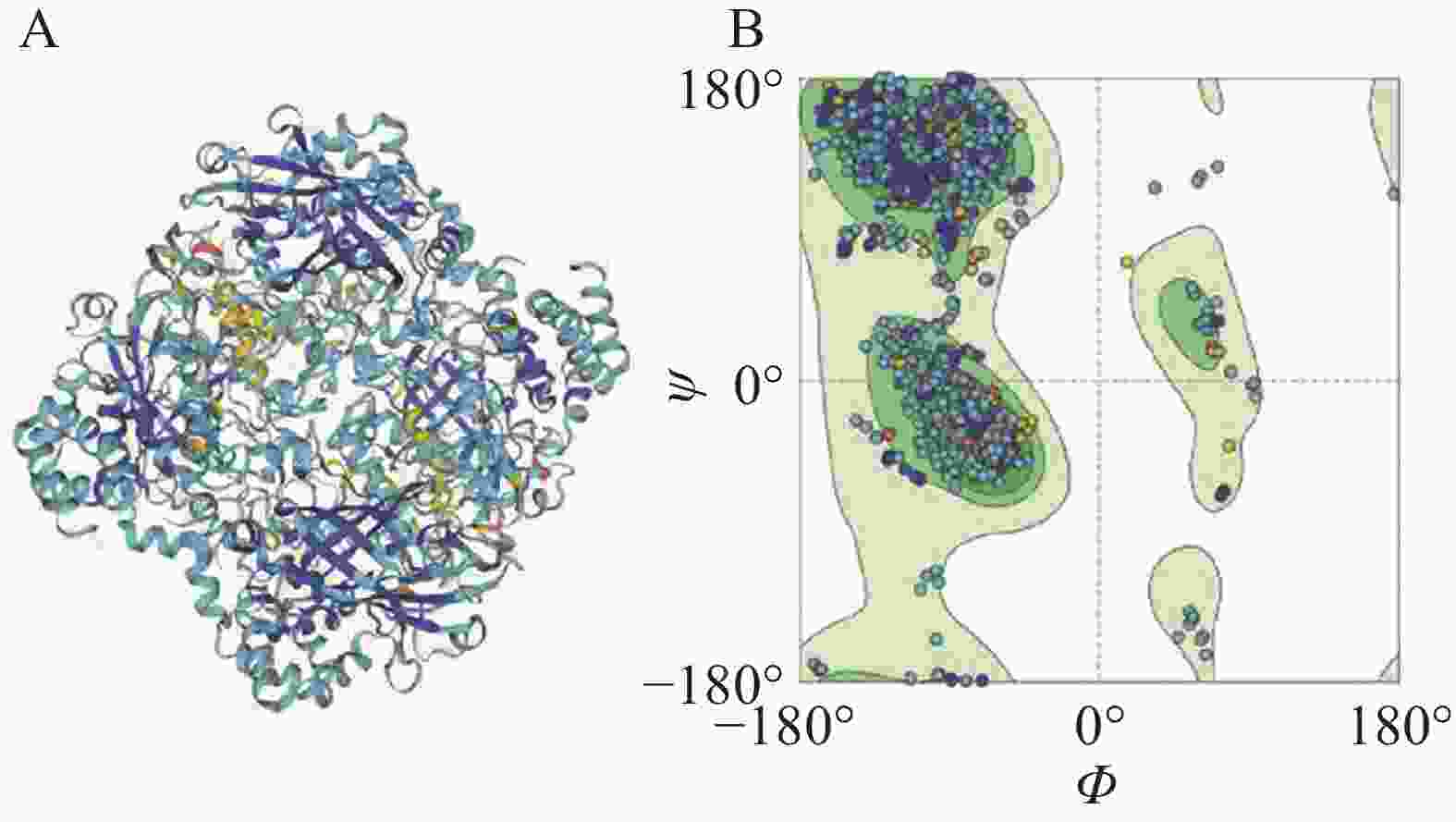
图 3 使用SWISS-MODEL预测LtCAT基因的蛋白质结构
Figure 3. The predicted protein structure of LtCAT gene by SWISS-MODEL
-
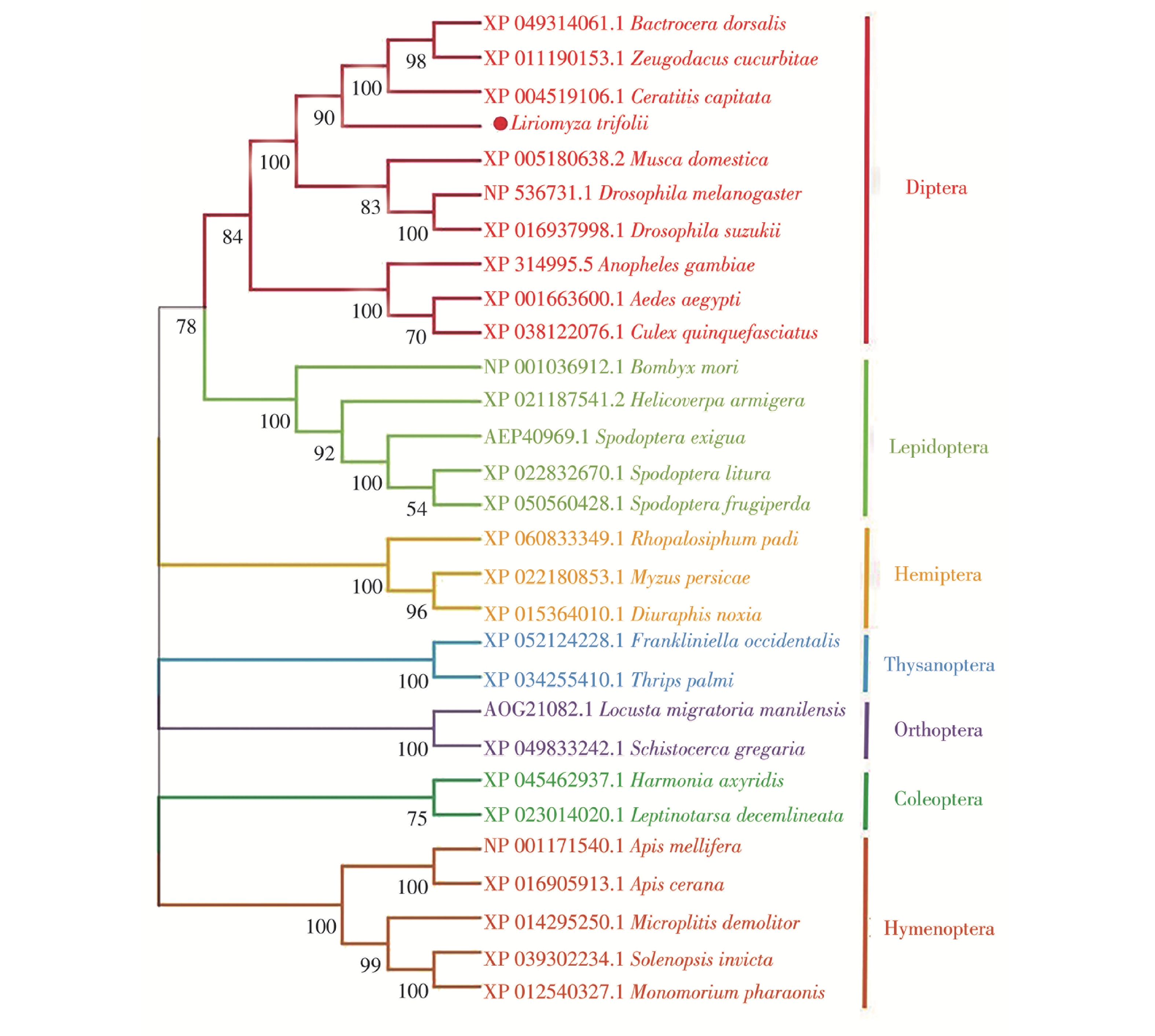
通过在NCBI查找并下载双翅目、鳞翅目、半翅目等7个目28种不同昆虫的CAT基因编码的氨基酸序列,与LtCAT基因编码的氨基酸序列共同进行系统发育树的构建。结果表明,相同目的昆虫CAT基因均以较高置信度聚为一支,说明昆虫CAT基因在昆虫属的分类阶元上具有高度保守性(图4)。其中,三叶草斑潜蝇CAT基因与橘小实蝇(Bactrocera dorsalis)、瓜实蝇(Zeugodacus cucurbitae)、地中海实蝇(Ceratitis capitata)聚为一支,置信度90%,而后与黑腹果蝇(Drosophila melanogaster)、斑翅果蝇(Drosophila suzukii)和家蝇(Musca domestica)共聚1个分支,置信度100%,表明其与其他蝇类的CAT基因进化关系更近。
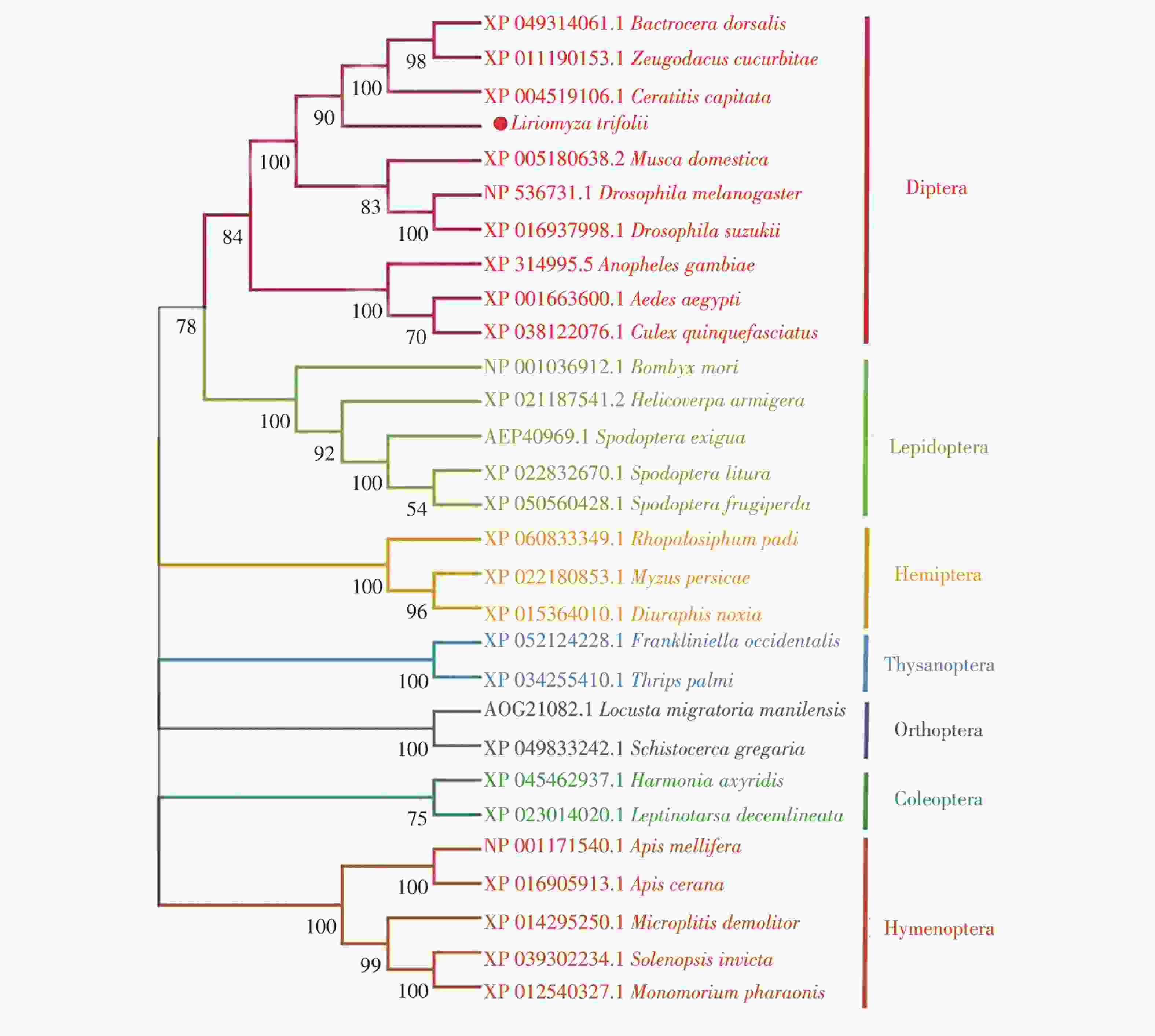
图 4 三叶草斑潜蝇与不同昆虫CAT蛋白序列的系统发育分析
Figure 4. Phylogenetic analysis of CAT protein sequences of Liriomyza trifolii and other different insects
-
根据系统发育树的同源进化分析,选择同源性高的昆虫进行多序列比对(图4)。三叶草斑潜蝇与黑腹果蝇等9种双翅目昆虫的CAT蛋白序列进行多序列比对,结果表明,三叶草斑潜蝇和9种双翅目昆虫的蛋白质序列一致性达84.38%,其中三叶草斑潜蝇与地中海实蝇序列相似度最高,序列一致性达82.85%,其次是橘小实蝇序列一致性达82.07%;三叶草斑潜蝇和冈比亚按蚊相似度最低,序列一致性仅70.04%,而与模式物种黑腹果蝇进行蛋白质序列比较,相似度为76.61%(图5)。

图 5 10种双翅目昆虫CAT蛋白序列的多序列比对
Figure 5. Multiple sequence alignment of CAT protein sequences of 10 Diptera insects
-
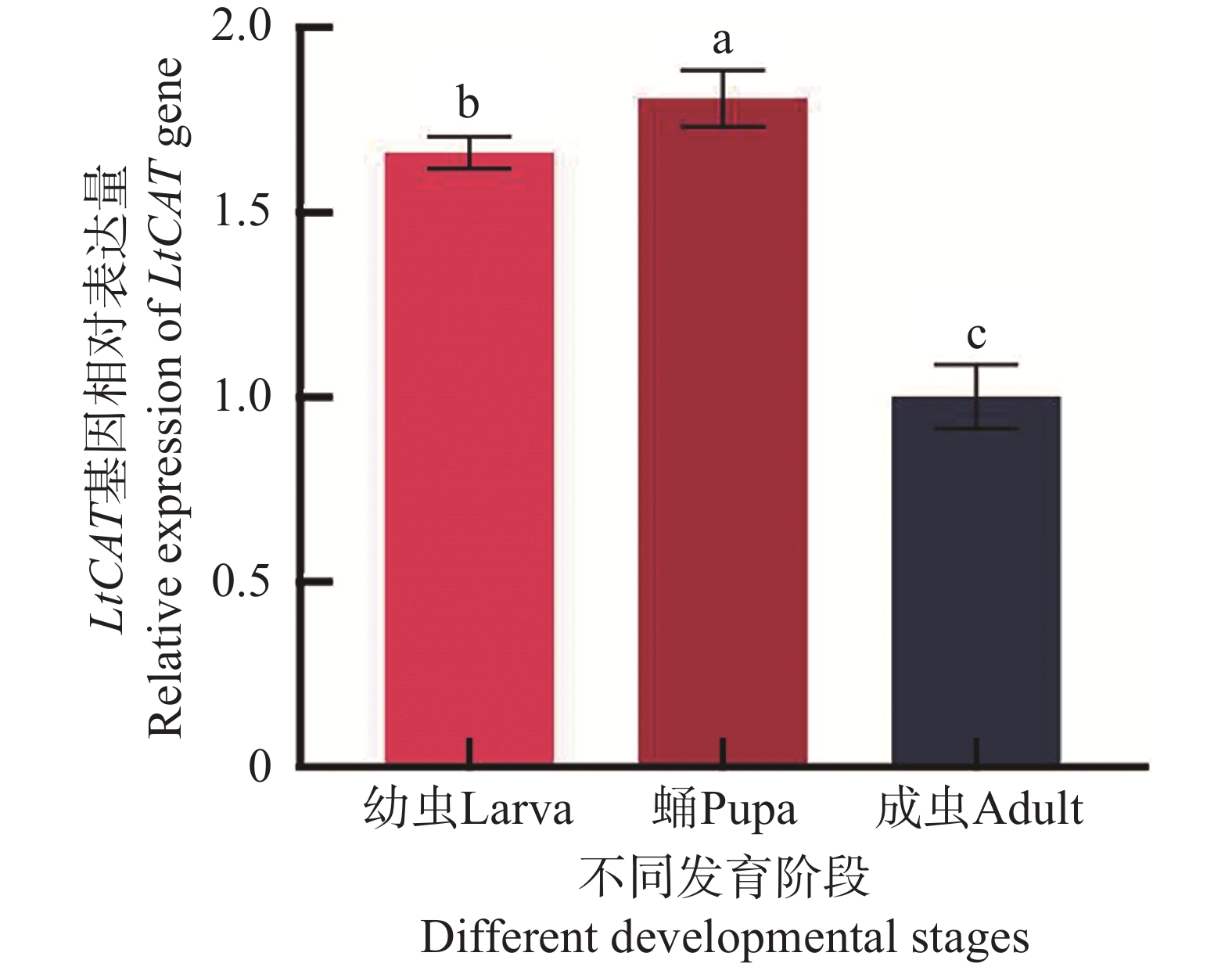
LtCAT基因在三叶草斑潜蝇幼虫、蛹、成虫发育阶段均有表达,表达水平呈先上升后下降的趋势,其蛹期表达量最高,显著高于幼虫期和成虫期(P < 0.05)(图6)。
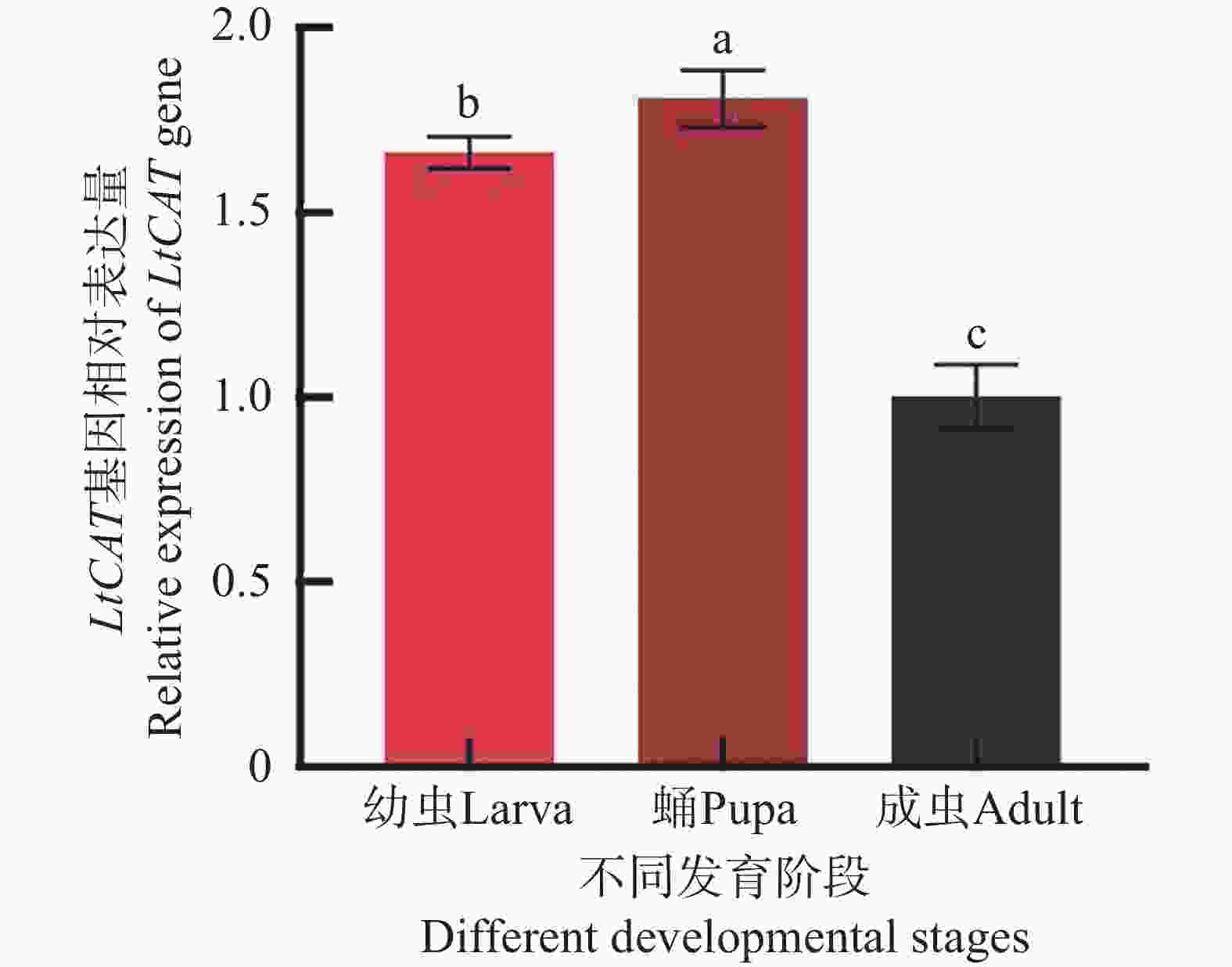
图 6 LtCAT基因在三叶草斑潜蝇不同发育阶段的表达
Figure 6. Gene expression of LtCAT of Liriomyza trifolii at different developmental stages
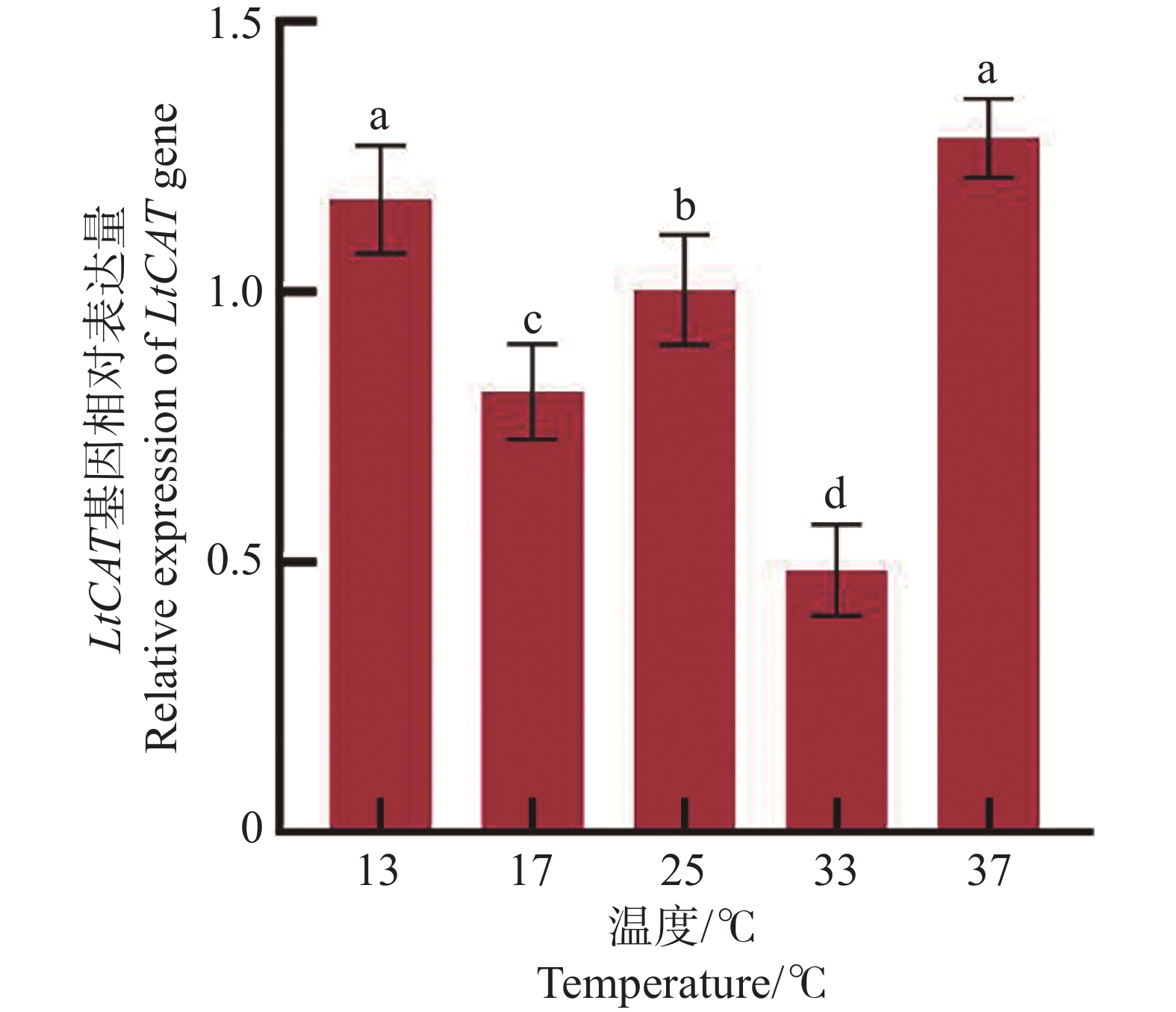
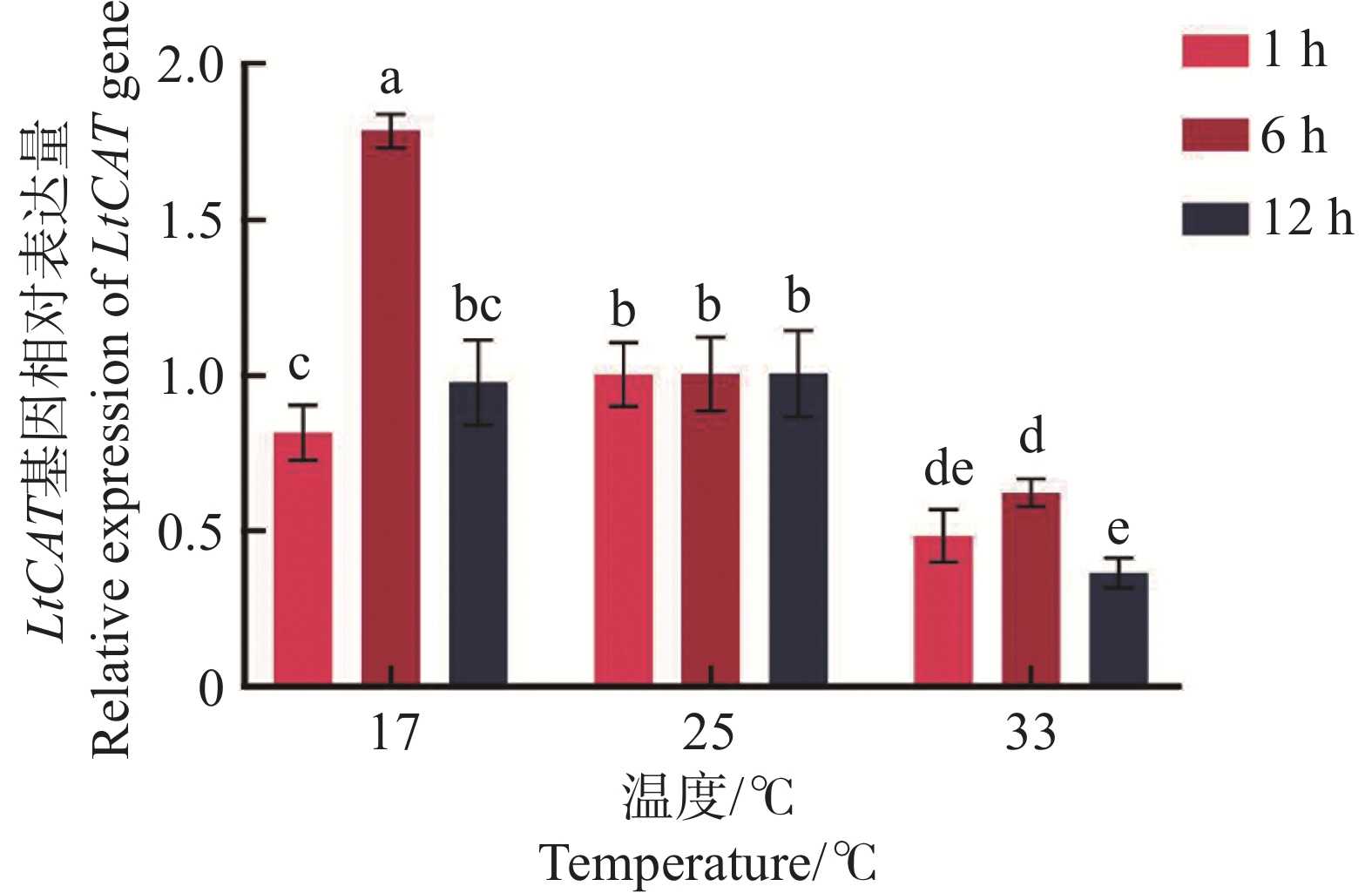
三叶草斑潜蝇2日龄蛹分别在13、17、25、33、37 ℃ 5个温度梯度短时间胁迫1 h后,LtCAT基因表达水平呈现波动变化(图7)。在低温13 ℃和高温37 ℃时,LtCAT的表达量最高,且两者之间无显著差异;两者的表达水平显著高于17、25、33 ℃的表达水平(P < 0.05)。低温17 ℃和高温33 ℃相比于25 ℃对照,LtCAT基因表达量分别下降了31%和52%。表明三叶草斑潜蝇2日龄蛹在低温和高温短时间胁迫时均积极做出了响应。
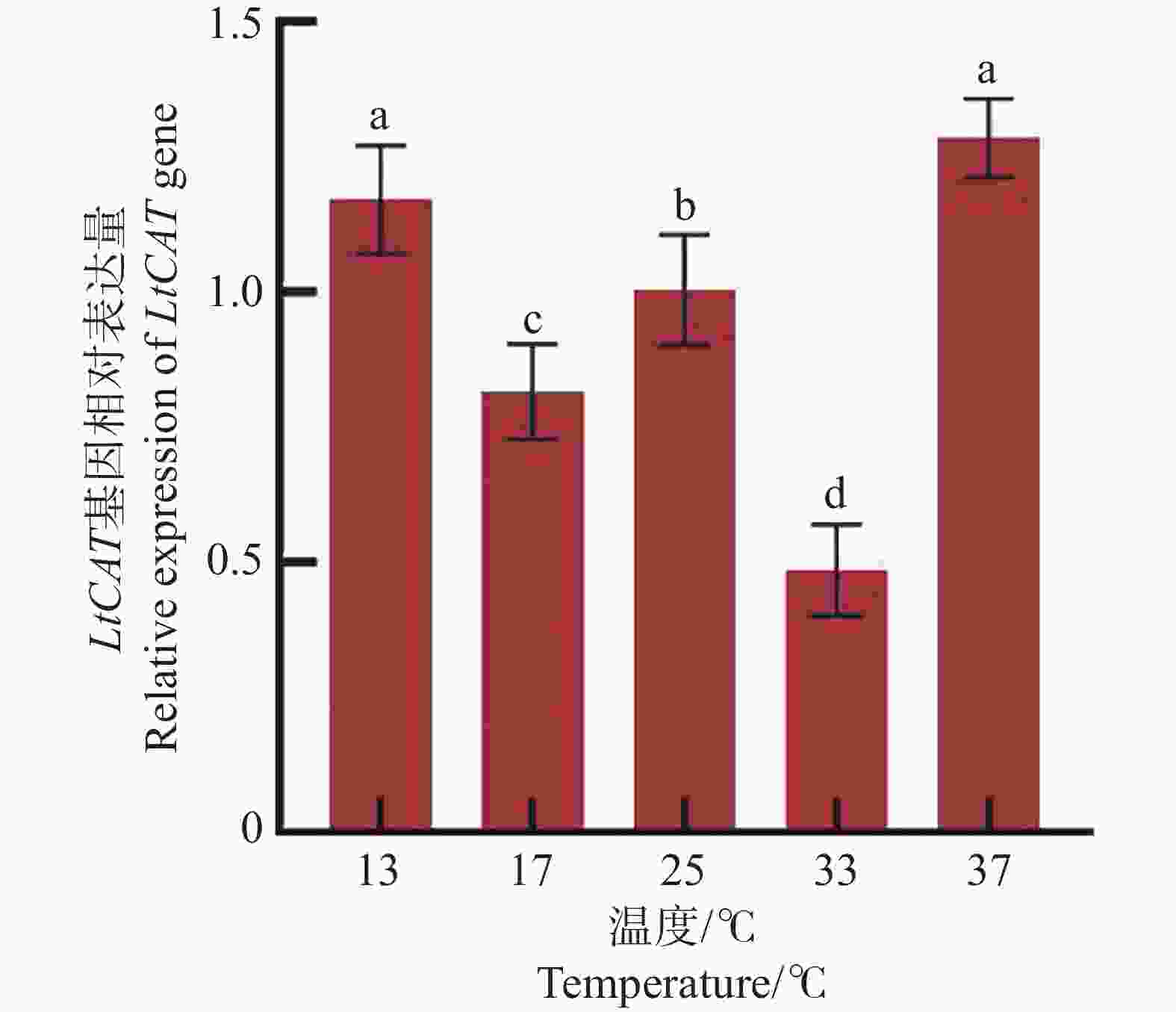
图 7 不同温度短时处理1 h的LtCAT基因表达
Figure 7. Gene expression of LtCAT treated at different temperatures for 1 h
相较于室温25 ℃,三叶草斑潜蝇在17 ℃和33 ℃处理不同时间的表达模式不同(图8)。在17 ℃处理时,LtCAT基因随胁迫时间的延长,其表达水平呈先上升后下降的趋势。17 ℃处理6 h时,LtCAT基因表达水平达到最高,显著的高于其他处理(P < 0.05);12 h时趋近与对照(室温25 ℃)。
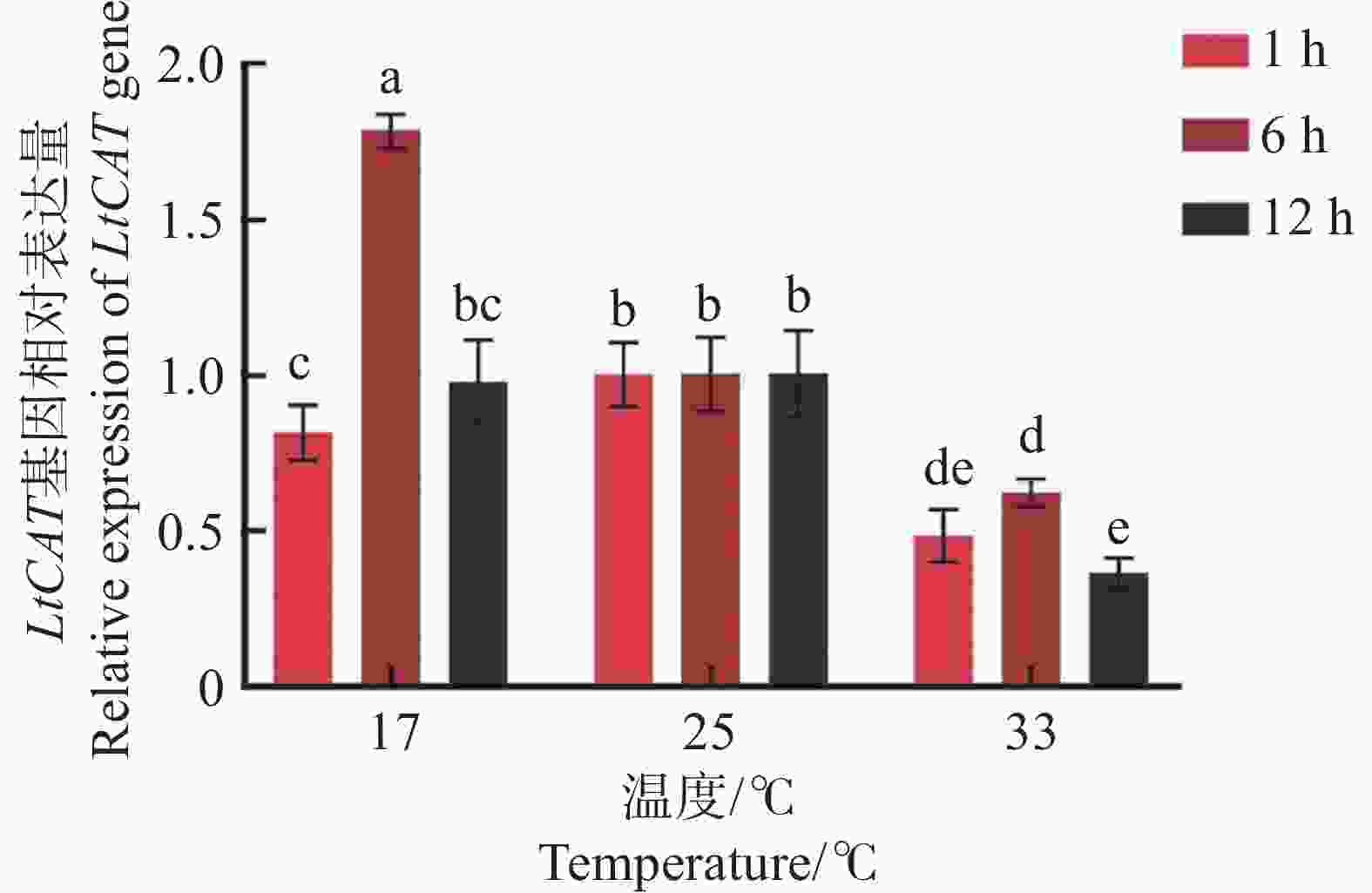
图 8 17、25、33 ℃处理不同时间后的LtCAT基因表达
Figure 8. The gene expression of LtCAT after treatment at 17,25, 33 ℃ for different hours
而在高温33 ℃胁迫后,三叶草斑潜蝇响应迅速,且持续表现出下降的趋势。结果表明,LtCAT基因在受到低温胁迫时具有一定的响应过程,6 h后被诱导高表达;而在受到高温胁迫时响应迅速,且高温对LtCAT基因的表达具有一定的抑制作用。
-
随着地球气候变暖,温度胁迫成为入侵昆虫成功定殖的关键屏障,同时入侵昆虫对高温的适应性促进了它们的全球扩散[29 − 30]。以往对三叶草斑潜蝇在温度适应的分子机制研究主要集中在热休克转录因子(HSF)[31]、小热休克蛋白(sHSPs)[32]、热激蛋白(hsp20、hsp40、hsp60、hsp70、hsp90)[33]等基因。热应激会破坏生物体内活性氧(ROS)水平的平衡,导致体内发生氧化应激反应,抗氧化酶在有效清除活性氧中起着至关重要的作用[21]。过氧化氢酶(CAT)是关键的抗氧化酶,在抗氧化途径中起着抵抗ROS的第一道防线[22, 34]。
本研究克隆得到的三叶草斑潜蝇CAT基因序列,ORF长度为1 542 bp,共编码513个氨基酸,预测相对分子质量为57.76 kDa,与二化螟CAT(56.76 kDa)、家蚕CAT(56.89 kDa)和飞蝗CAT(57.49 kDa)序列相似度较高且分子量大小相近[35 − 37]。LtCAT基因具有1个典型的CAT家族保守结构域(22-504氨基酸位点)。系统发育树结果表明:三叶草斑潜蝇和橘小实蝇、瓜实蝇、地中海实蝇亲缘关系更近,CAT基因同源性高达90%。同时在多序列比对发现三叶草斑潜蝇与黑腹果蝇、冈比亚按蚊等9种双翅目昆虫的CAT基因序列一致性达84.38%。表明CAT基因在不同物种之间较为保守。
三叶草斑潜蝇在幼虫、蛹、成虫3个不同发育阶段均有表达,且表达水平有差异,在蛹期表达量最高,其次为幼虫期,最低为成虫期(图6)。胡振等[38]的研究表明,甜菜夜蛾(Spodoptera exigua)CAT基因在低龄幼虫的表达显著低于蛹期;同时,粘虫(Mythimna separata)和二化螟(Chilo suppressalis)的CAT基因同样在不同龄期的幼虫表达量均大于成虫[19, 39],这与本研究结果一致,推测蛹期暴露在环境中,遇到的外界胁迫因素较多,容易引起CAT的高表达,而幼虫阶段处于活跃发育阶段,体内H2O2的积累量较成虫高,因此也需要较多的CAT用以清除体内ROS[40]。
不同温度短时胁迫1 h的表达分析发现,LtCAT基因在13、17、25 ℃表达谱为下降后再上升的趋势,与黏虫(M. separata)的CAT基因在10、15、20、30 ℃温度胁迫1 h的表达谱类似,呈现下降后上升的趋势[41]。而LtCAT基因在温度17、25、33 ℃处理1 h呈先上升后下降的趋势,与粘虫(M. separata)3龄幼虫在不同高温31、33、35、37、39 ℃处理2 h,随着温度的增加,呈现先上升后下降的趋势[19]结果类似。LtCAT基因在25、33、37 ℃处理1 h的表达谱为下降后再上升的趋势,与寄生蜂(A. asychis)雌雄成虫的CAT基因在25、32.5、35 ℃表达趋势一致,寄生蜂在25 ℃到32.5 ℃表达量下降,35 ℃时表达量上升[23]。但是与二化螟(C. suppressalis)的CAT基因在27、33、36 ℃表达趋势不同,该二化螟表达量是呈先上升再下降的趋势[35, 39]。CAT基因受到外界不同温度胁迫后,首先在昆虫体内积极响应导致表达量升高,随着温度的升高,昆虫机体活力下降,基因表达量反而会出现下降现象。昆虫体内的热感受器(神经细胞)分布因物种和个体生命阶段的不同而有很大差异[42]。
不同温度长时胁迫1、6、12 h的表达分析发现,LtCAT基因在17、33 ℃处理1、6、12 h的表达谱,随着温度胁迫时间从1 h延长到6 h,三叶草斑潜蝇2日龄蛹体内氧化应激水平提高,但紧随着时间延长到12 h时,表达量出现下降的趋势,这与朱彤等[21]的研究结果类似,巴氏新小绥螨(N. barkeri)在40 ℃分别进行2、4和6 h的温度胁迫,表达量在4 h时上升诱导了体内的CAT氧化应激,6 h时下降,呈现随着时间从2 h到4 h的延长,表达量上调,到6 h表达量下调。研究结果表明,三叶草斑潜蝇CAT基因在应对高温和低温以及短时和长时处理时的响应是不同的,抗氧化酶CAT基因转录上调、下调表达可作为生物体内氧化应激反应的指示。
本研究克隆三叶草斑潜蝇Liriomyza trifolii的CAT基因,并对其进行了生物信息学等相关分析,同时,使用荧光定量PCR技术测定了CAT基因对三叶草斑潜蝇的温度胁迫和发育表达分析。本研究将为三叶草斑潜蝇的CAT基因功能研究提供重要的基础理论依据,能够给未来深入了解CAT基因的分子机制以及进一步挖掘基因功能提供一定的理论基础和数据支持。
-
本研究通过克隆得到三叶草斑潜蝇的CAT基因序列,命名为LtCAT,ORF长度为1 542 bp,共编码513个氨基酸;LtCAT基因在三叶草斑潜蝇的幼虫、蛹、成虫3个发育阶段体内均有表达,蛹期表达量较高;LtCAT基因在应对高低温以及短时和长时处理时的响应是不同的,抗氧化酶CAT基因转录上调、下调表达可作为生物体内氧化应激反应的指示。
三叶草斑潜蝇过氧化氢酶基因的克隆、表达及其对温度的响应
DOI: 10.15886/j.cnki.rdswxb.20240033
 CSTR: 32425.14.j.cnki.rdswxb.20240033
CSTR: 32425.14.j.cnki.rdswxb.20240033
Molecular cloning, expression pattern of catalase gene in Liriomyza trifolii and its response to temperature stress
-
摘要: 三叶草斑潜蝇[Liriomyza trifolii (Burgess)]是中国瓜菜上的重要入侵害虫之一。为进一步探讨三叶草斑潜蝇对温度胁迫的适应性机制,对三叶草斑潜蝇的过氧化氢酶(Catalase,CAT)序列特征、结构和系统发育等进行了生物信息学分析,并利用实时荧光定量PCR检测了LtCAT基因在不同发育阶段、温度胁迫下的表达情况。本研究克隆获得了三叶草斑潜蝇CAT基因序列,其开放阅读框长度为1 542 bp,共编码513个氨基酸。系统发育树表明,三叶草斑潜蝇与橘小实蝇(Bactrocera dorsalis)、瓜实蝇(Zeugodacus cucurbitae)、地中海实蝇(Ceratitis capitata)CAT基因亲缘关系更近,聚为一支,置信度达90%。LtCAT基因在幼虫、蛹、成虫均有差异表达,在蛹期表达量最高;温度胁迫处理后,LtCAT基因表达随着温度变化积极响应。本研究为后续深入研究三叶草斑潜蝇CAT基因的功能提供依据。Abstract: Liriomyza trifolii (Burgess) is one of the important invasive pests infesting melons and vegetables in China. An attempt was made to further explore the adaptive mechanism of L. trifolii to temperature stress. The sequence characteristics, structure and phylogenetic analysis of catalase (CAT) in L. trifolii were analyzed by using bioinformatics, and the expression of LtCAT gene at different developmental stages and under temperature stress was detected by real-time fluorescence quantitative PCR. The CAT gene sequence of L. trifolii was cloned, and its open reading frame length was
1542 bp, encoding 513 amino acids. The construction of phylogenetic tree showed that the CAT gene of L. trifolii was closely related to those of Bactrocera dorsalis, Zeugodacus cucurbitae and Ceratitis capitata, and hence clustered into one branch with a confidence level of 90%. The LtCAT gene was differentially expressed in larvae, pupae and adults, and the expression level was the highest in pupae. After temperature stress treatment, the expression of LtCAT gene responded positively to temperature changes. This study provides a basis for further study on the function of CAT gene in L. trifolii.-
Key words:
- Liriomyza trifolii /
- antioxidase /
- temperature stress /
- sequence analysis /
- expression pattern
-
图 2 三叶草斑潜蝇CAT基因序列分析
注:灰色区域为CAT家族区域位点(22~504氨基酸位点),其中红色方框为CAT家族特征位点(39~62、102~120、123~140、142~160、307~334、339~365氨基酸位点);黑色括号区域为CAT核心结构域(26~411氨基酸位点);蓝色横线为CAT近端活性位点(62~78氨基酸位点);蓝色虚线为CAT近端血红素配体特征位点(352~360氨基酸位点)。
Fig. 2 Sequence analysis of CAT gene of Liriomyza trifolii
Note: The gray area is the CAT family region site (amino acid site 22-504), and the red box is the CAT family characteristic site (amino acid sites 39-62, 102-120, 123-140, 142-160, 307-334, 339-365); the black bracketed region is the CAT core domain (amino acid sites 26-411); the blue horizontal line is the proximal active site of CAT (amino acid site 62-78); the blue dotted line is the characteristic site of CAT proximal heme ligand (amino acid site 352-360).
图 3 使用SWISS-MODEL预测LtCAT基因的蛋白质结构
注:A.3D结构预测模型图;B. 拉马钱德兰图。
Fig. 3 The predicted protein structure of LtCAT gene by SWISS-MODEL
Note: A.3D structure prediction model diagram; B. Ramachandran plot.
图 4 三叶草斑潜蝇与不同昆虫CAT蛋白序列的系统发育分析
Fig. 4 Phylogenetic analysis of CAT protein sequences of Liriomyza trifolii and other different insects
图 5 10种双翅目昆虫CAT蛋白序列的多序列比对
注:图中黑色标注为高度保守区域(100%),红色标注为相似区域(≥75%),蓝色标注为相似区域(≥50%)。
Fig. 5 Multiple sequence alignment of CAT protein sequences of 10 Diptera insects
Note: In the figure, black is marked as a highly conserved region (100%), red is marked as a similar region (≥75%), and blue is marked as a similar region (≥50%).
图 6 LtCAT基因在三叶草斑潜蝇不同发育阶段的表达
注:不同字母表示各处理间具有显著性差异(P < 0.05),实验数据表示为平均数Mean ± 标准误差SEM。
Fig. 6 Gene expression of LtCAT of Liriomyza trifolii at different developmental stages
Note: Different letters indicate significant differences between treatments (P < 0.05), and the experimental data are expressed as Mean ± standard error (SEM).
图 7 不同温度短时处理1 h的LtCAT基因表达
注:不同字母表示各处理间具有显著性差异(P < 0.05),实验数据表示为平均数Mean ± 标准误差SEM,25 ℃作为对照。
Fig. 7 Gene expression of LtCAT treated at different temperatures for 1 h
Note: Different letters indicate significant differences between treatments (P < 0.05). The experimental data are expressed as Mean ± standard error (SEM), and 25 ℃ is used as a control.
图 8 17、25、33 ℃处理不同时间后的LtCAT基因表达
注:不同字母表示各处理间具有显著性差异(P < 0.05),实验数据表示为平均数Mean ± 标准误差SEM,25 ℃作为对照。
Fig. 8 The gene expression of LtCAT after treatment at 17,25, 33 ℃ for different hours
Note: Different letters indicate significant differences between treatments (P < 0.05). The experimental data are expressed as Mean ± standard error (SEM), and 25 ℃ is used as a control.
表 1 本研究所用引物信息
Table 1 Primer information used in this study
引物名称
Primer引物序列(5′−3′)
Primer sequence(5′−3′)产物长度/bp
Product length/bp用途
UseLtCAT-F F:ATTCAGCCAGACGGATCAGC 1731 全长克隆
(RT-PCR)LtCAT-R R:ACTCAATAGACTTGCTTTGCTGG qLtCAT-F F:CATTCCAGAACGTGTCGTGC 148 荧光定量
(RT-qPCR)qLtCAT-R R:GCCCACAGTGGAGAAACGTA β-actin-F[27] F:TTGTATTGGACTCTGGTGACGG 73 β-actin-R[27] R:GATAGCGTGAGGCAAAGCATAA 注:使用Primer Premier 5.0软件设计引物。 Note: Primers were designed using Primer Premier 5.0 software.  下载: 导出CSV
下载: 导出CSV
-
[1] 王音, 问锦曾. 三叶草斑潜蝇发生动态及检疫[J]. 植物检疫, 1995, 9(1): 10 − 11. [2] 余道坚, 郑文华, 林朝森, 等. 警惕三叶斑潜蝇的侵入[J]. 中国进出境动植检, 1998(3): 40 − 42. [3] 雷仲仁, 朱灿健, 张长青. 重大外来入侵害虫三叶斑潜蝇在中国的风险性分析[J]. 植物保护, 2007, 33(1): 37 − 41. doi: 10.3969/j.issn.0529-1542.2007.01.010 [4] 汪兴鉴, 黄顶成, 李红梅, 等. 三叶草斑潜蝇的入侵、鉴定及在中国适生区分析[J]. 昆虫知识, 2006, 43(4): 540 − 545. doi: 10.3969/j.issn.0452-8255.2006.04.026 [5] 王凯歌, 益浩, 雷仲仁, 等. 两种外来入侵斑潜蝇在海南地区的竞争取代调查分析[J]. 中国农业科学, 2013, 46(22): 4842 − 4848. doi: 10.3864/j.issn.0578-1752.2013.22.023 [6] 何德良. 有害生物—三叶斑潜蝇[J]. 植物检疫, 2007, 21(2): 120 − 122. doi: 10.3969/j.issn.1005-2755.2007.02.024 [7] 陈洪俊, 李镇宇, 骆有庆. 检疫性有害生物三叶斑潜蝇[J]. 植物检疫, 2005, 19(2): 99 − 102. doi: 10.3969/j.issn.1005-2755.2005.02.010 [8] 吕延超, 廖道龙, 陈贻诵, 等. 海南蔬菜产业发展现状及其对策[J]. 特种经济动植物, 2020, 23(10): 44 − 47. doi: 10.3969/j.issn.1001-4713.2020.10.023 [9] 成善汉, 汪志伟, 朱国鹏. 海南省蔬菜产业发展现状及对策[J]. 中国蔬菜, 2019(6): 16 − 20. doi: 10.19928/j.cnki.1000-6346.2019.06.004 [10] 王硕, 吕宝乾, 王树昌, 等. 基于防虫网+的热区豇豆病虫害生态调控策略[J]. 热带农业科学, 2022, 44(7): 27 − 35. doi: 10.12008/j.issn.1009-2196.2024.07.005 [11] LI F, GONG X Y, YUAN L L, et al. Indoxacarb resistance-associated mutation of Liriomyza trifolii in Hainan, China[J]. Pesticide Biochemistry and Physiology, 2022, 183: 105054. doi: 10.1016/j.pestbp.2022.105054 [12] LIVINGSTONE D R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms[J]. Marine Pollution Bulletin, 2001, 42(8): 656 − 666. doi: 10.1016/S0025-326X(01)00060-1 [13] JIA F X, DOU W, HU F, et al. Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae)[J]. Florida Entomologist, 2011, 94(4): 956 − 963. doi: 10.1653/024.094.0432 [14] JOANISSE D, STOREY K. Oxidative stress and antioxidants in overwintering larvae of cold-hardy goldenrod gall insects[J]. Journal of Experimental Biology, 1996, 199(Pt 7): 1483-1491. doi: 10.1242/jeb.199.7.1483. [15] LANDIS G N, TOWER J. Superoxide dismutase evolution and life span regulation[J]. Mechanisms of Ageing and Development, 2005, 126(3): 365 − 379. doi: 10.1016/j.mad.2004.08.012 [16] AN M I, CHOI C Y. Activity of antioxidant enzymes and physiological responses in ark shell, Scapharca broughtonii, exposed to thermal and osmotic stress: effects on hemolymph and biochemical parameters[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2010, 155(1): 34 − 42. doi: 10.1016/j.cbpb.2009.09.008 [17] 冯宏祖, 刘映红, 何林, 等. 阿维菌素和温度胁迫对朱砂叶螨自由基及保护酶活性的影响[J]. 植物保护学报, 2008, 35(6): 530 − 536. doi: 10.3321/j.issn:0577-7518.2008.06.010 [18] CHENG J, WANG C Y, LYU Z H, et al. Identification and characterization of the catalase gene involved in resistance to thermal stress in Heortia vitessoides using RNA interference[J]. Journal of Thermal Biology, 2018, 78: 114 − 121. doi: 10.1016/j.jtherbio.2018.09.008 [19] 李鸿波, 戴长庚, 张昌容, 等. 粘虫过氧化氢酶基因的克隆与表达分析[J]. 昆虫学报, 2018, 61(2): 178 − 187. doi: 10.16380/j.kcxb.2018.02.004 [20] KHURSHID A, INAYAT R, TAMKEEN A, et al. Antioxidant enzymes and heat-shock protein genes of green peach aphid (Myzus persicae) under short-time heat stress[J]. Frontiers in Physiology, 2021, 12: 805509. doi: 10.3389/fphys.2021.805509 [21] ZHU T, LI W Z, XUE H, et al. Selection, identification, and transcript expression analysis of antioxidant enzyme genes in Neoseiulus barkeri after short-term heat stress[J]. Antioxidants, 2023, 12(11): 1998. doi: 10.3390/antiox12111998 [22] LIANG P, NING J, WANG W L, et al. Catalase promotes whitefly adaptation to high temperature by eliminating reactive oxygen species[J]. Insect Science, 2023, 30(5): 1293 − 1308. doi: 10.1111/1744-7917.13157 [23] LIU X, FU Z X, KANG Z W, et al. Identification and characterization of antioxidant enzyme genes in parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and expression profiling analysis under temperature stress[J]. Insects, 2022, 13(5): 447. doi: 10.3390/insects13050447 [24] 龙佳芝, 郭文秀, 季敏, 等. 高温胁迫对丽蚜小蜂死亡率和保护酶系的影响[J]. 草原与草坪, 2023, 43(4): 130 − 136. doi: 10.13817/j.cnki.cyycp.2023.04.017 [25] 常亚文. 三叶斑潜蝇的种间竞争优势和温度耐受性的分子机制[D]. 扬州: 扬州大学, 2021. doi: 10.27441/d.cnki.gyzdu.2021.001993. [26] CHANG Y W, ZHANG X X, LU M X, et al. Transcriptome analysis of Liriomyza trifolii (Diptera: Agromyzidae) in response to temperature stress[J]. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 2020, 34: 100677. doi: 10.1016/j.cbd.2020.100677 [27] CHANG Y W, CHEN J Y, LU M X, et al. Selection and validation of reference genes for quantitative real-time PCR analysis under different experimental conditions in the leafminer Liriomyza trifolii (Diptera: Agromyzidae)[J]. PLoS One, 2017, 12(7): e0181862. doi: 10.1371/journal.pone.0181862 [28] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the $ 2^{ - \Delta \Delta {C_{\rm{T}}}} $ method[J]. Methods, 2001, 25(4): 402 − 408. [29] ZHANG B C, ZHOU X H, ZHOU L Y, et al. A global synthesis of below-ground carbon responses to biotic disturbance: a meta-analysis[J]. Global Ecology and Biogeography, 2015, 24(2): 126 − 138. doi: 10.1111/geb.12235 [30] 彭露, 万方浩, 侯有明. 中国入侵昆虫预防与控制研究进展[J]. 应用昆虫学报, 2020, 57(2): 244 − 258. doi: 10.7679/j.issn.2095-1353.2020.028 [31] CHANG Y W, WANG Y C, ZHANG X X, et al. Transcriptional regulation of small heat shock protein genes by heat shock factor 1 (HSF1) in Liriomyza trifolii under heat stress[J]. Cell Stress and Chaperones, 2021, 26(5): 835 − 843. doi: 10.1007/s12192-021-01224-2 [32] CHANG Y W, ZHANG X X, LU M X, et al. Molecular cloning and characterization of small heat shock protein genes in the invasive leaf miner fly, Liriomyza trifolii[J]. Genes, 2019, 10(10): 775. doi: 10.3390/genes10100775 [33] CHANG Y W, CHEN J Y, LU M X, et al. Cloning and expression of genes encoding heat shock proteins in Liriomyza trifolii and comparison with two congener leafminer species[J]. PLoS One, 2017, 12(7): e0181355. doi: 10.1371/journal.pone.0181355 [34] KIM B Y, KIM H J, LEE K S, et al. Catalase from the white-spotted flower chafer, Protaetia brevitarsis: cDNA sequence, expression, and functional characterization[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2008, 149(1): 183 − 190. doi: 10.1016/j.cbpb.2007.09.007 [35] XU J, LU M X, HUANG D L, et al. Molecular cloning, characterization, genomic structure and functional analysis of catalase in Chilo suppressalis[J]. Journal of Asia-Pacific Entomology, 2017, 20(2): 331 − 336. doi: 10.1016/j.aspen.2016.08.006 [36] YAMAMOTO K, BANNO Y, FUJII H, et al. Catalase from the silkworm, Bombyx mori: gene sequence, distribution, and overexpression[J]. Insect Biochemistry and Molecular Biology, 2005, 35(4): 277 − 283. doi: 10.1016/j.ibmb.2005.01.001 [37] ZHANG X Y, LI Y H, WANG J X, et al. Identification and characteristic analysis of the catalase gene from Locusta migratoria[J]. Pesticide Biochemistry and Physiology, 2016, 132: 125 − 131. doi: 10.1016/j.pestbp.2016.03.010 [38] 胡振, 左洪亮, 李亚楠, 等. 甜菜夜蛾过氧化氢酶cDNA序列克隆、序列分析和表达特征[J]. 昆虫学报, 2011, 54(11): 1249 − 1257. doi: 10.16380/j.kcxb.2011.11.003 [39] LU Y H, BAI Q, ZHENG X S, et al. Expression and enzyme activity of catalase in Chilo suppressalis (Lepidoptera: Crambidae) is responsive to environmental stresses[J]. Journal of Economic Entomology, 2017, 110(4): 1803 − 1812. doi: 10.1093/jee/tox117 [40] 郑玉涛. 高温胁迫对西花蓟马抗氧化酶活性的影响及CAT基因的克隆与表达[D]. 扬州: 扬州大学, 2015. doi: 10.7666/d.Y2908420. [41] ALI A, RASHID M A, HUANG Q Y, et al. Response of antioxidant enzymes inMythimna separata(Lepidoptera: Noctuidae) exposed to thermal stress[J]. Bulletin of Entomological Research, 2017, 107(3): 382 − 390. doi: 10.1017/S0007485316001000 [42] BODLAH M A, IQBAL J, ASHIQ A, et al. Insect behavioral restraint and adaptation strategies under heat stress: an inclusive review[J]. Journal of the Saudi Society of Agricultural Sciences, 2023, 22(6): 327 − 350. doi: 10.1016/j.jssas.2023.02.004 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1644
- HTML全文浏览量: 590
- PDF下载量: 39
- 被引次数: 0
