

-
砗磲隶属于软体动物门,瓣鳃纲,帘蛤目,砗磲科,目前有2属10种,广泛分布于印度洋和西太平洋的珊瑚礁海域,我国岛礁海域分布有6种[1-2]。砗磲是珊瑚礁生态系统的重要组成部分,对维护珊瑚礁系统生态平衡具有重要的生态学意义。由于砗磲固定生活在热带浅层的透明海域,易被捕捞,因此,砗磲资源容易遭受破坏,某些海域部分的砗磲品种已经濒临绝迹[3-4]。20世纪 60年代,我国西沙群岛海域砗磲资源很丰富,虽然逐年减少,但仍保持一定数量。20世纪90年代以后,西沙群岛海域很难再看到国家一级保护动物库氏砗磲的存在[5]。进入21世纪以后,受经济利益驱使,渔民通过暴力采挖珊瑚礁的手段获取砗磲贝壳,致使砗磲赖以生存的珊瑚礁栖息环境受到极大破坏,导致西沙群岛海域砗磲濒临绝迹[6]。此外,全球温室效应、海洋生态环境恶化、生物资源过度开发等都在加速破坏珊瑚礁生态系统,影响砗磲的存活[7]。目前,南海砗磲资源现状如何,仅见海南岛近岸海域的砗磲资源调查,如崔丹等[8]和刘春胜等[9]分别对海南岛近岸的砗磲进行了调查,结果表明海南岛周边砗磲资源量不容乐观,三亚珊瑚礁国家自然保护区砗磲密度仅为0.009个·m−2,临高和文昌等部分海域砗磲已经绝迹,而针对西沙海域砗磲的资源调查未见报道。笔者通过采用截线样带法对西沙群岛海域砗磲资源分布状况和生存环境进行了调查,以期为我国砗磲资源的保护提供基础资料。
-
调查区域西沙群岛位于我国南海中北部,海南岛东南方,是南海诸岛中最大的群岛,居北纬15°46′~17°08′,东经111°11′~112°54′,西沙群岛海域面积达50万平方公里,由宣德和永乐2个群岛组成,陆地总面积约10 km2 [10]。西沙群岛海域拥有中国典型的湿地生态系统,尤其是珊瑚礁生态系统,为各种海洋生物资源提供栖息环境,是渔业资源产卵场、索饵场与洄游通道,是热带海洋生物聚集区,其中砗磲是珊瑚礁生态系统中重要的组成部分[11]。2017年6—7月开展野外调查。根据西沙主要岛礁的地理位置共布设27个调查站位,站位基本覆盖西沙群岛海域,其中宣德环礁海域5个,分别为西沙洲、赵述岛、北岛、南沙洲和永兴岛;永乐环礁海域7个,分别为晋卿岛、石屿、银屿、全富岛、珊瑚岛、甘泉岛和羚羊礁;离散岛礁5个,分别为华光礁、北礁、玉琢礁、盘石屿和浪花礁。
-
采用国际上通用的截线样带法,在各岛礁调查站位不同深度布设一条50 m×0.5 m的条状断面,断面平行于海岸线。由于虫黄藻的光合作用需要合适的光照,水太深光照则无法到达,因此,砗磲的垂直分布受与其共生的虫黄藻的限制,根据岛礁调查站位的海底地形及海流波动等因素,设置砗磲调查条状断面在20 m水深以内[12-13]。在每个站位布设5个水深,分别是0 m(潮间带)、5 m、10 m 、15 m、20 m。为尽可能较详细和准确地评估每一站位砗磲生态现状,潜水员在每个样带潜水10~25 min,同时利用水下高清摄影机和照相机对布设站点进行拍摄、照相,然后在实验室内根据影像资料和现场调查记录的真实信息进行分析[14]。砗磲种类鉴定参照《世界名贝鉴赏图谱》和《中国海洋贝类图鉴》等贝类图鉴进行鉴定[15-16]。珊瑚的调查方法与砗磲的相同,同时根据影像资料计算各调查断面珊瑚覆盖率[6]。
-
采用Margalef种类丰富度指数D(Richness index)[17]、Shannon-Wiener多样性指数H′(Diversity index)[18]、Pileou均匀度指数J'(Evenness index)[19]对不同站位的砗磲物种多样性进行分析。
$$ \begin{gathered} D=(S-1) / \ln N ;\\ H^{\prime}=-\sum\nolimits_{i=1}^S\left(n_i / N\right) \ln \left(n_i / N\right) ; \\ J^{\prime}=H^{\prime} / \ln S \;, \end{gathered} $$ 式中,S为物种种类数;N为物种总个体数;ni为第i种个体数占总个体数的比例。
$$\rho=N i / S \;,$$ 式中,ρ为砗磲密度,S为调查样带面积,Ni为i物种在样带面积内的个体数。
所有数据进行统计分析,采用统计软件SPSS19.0单因素方差分析(One-way ANOVA,Tukey氏检验),比较不同站位间的各项指标数据(物种多样性指数、砗磲密度和珊瑚覆盖率)的差异[20],差异显著性水平设置P<0.05。砗磲密度与珊瑚覆盖率进行相关性分析:相关系数R的绝对值在0.8以上,表示有很强的相关性;在0.3到0.8之间,表示有弱的相关性;在0.3以下,表示没有相关性。
-
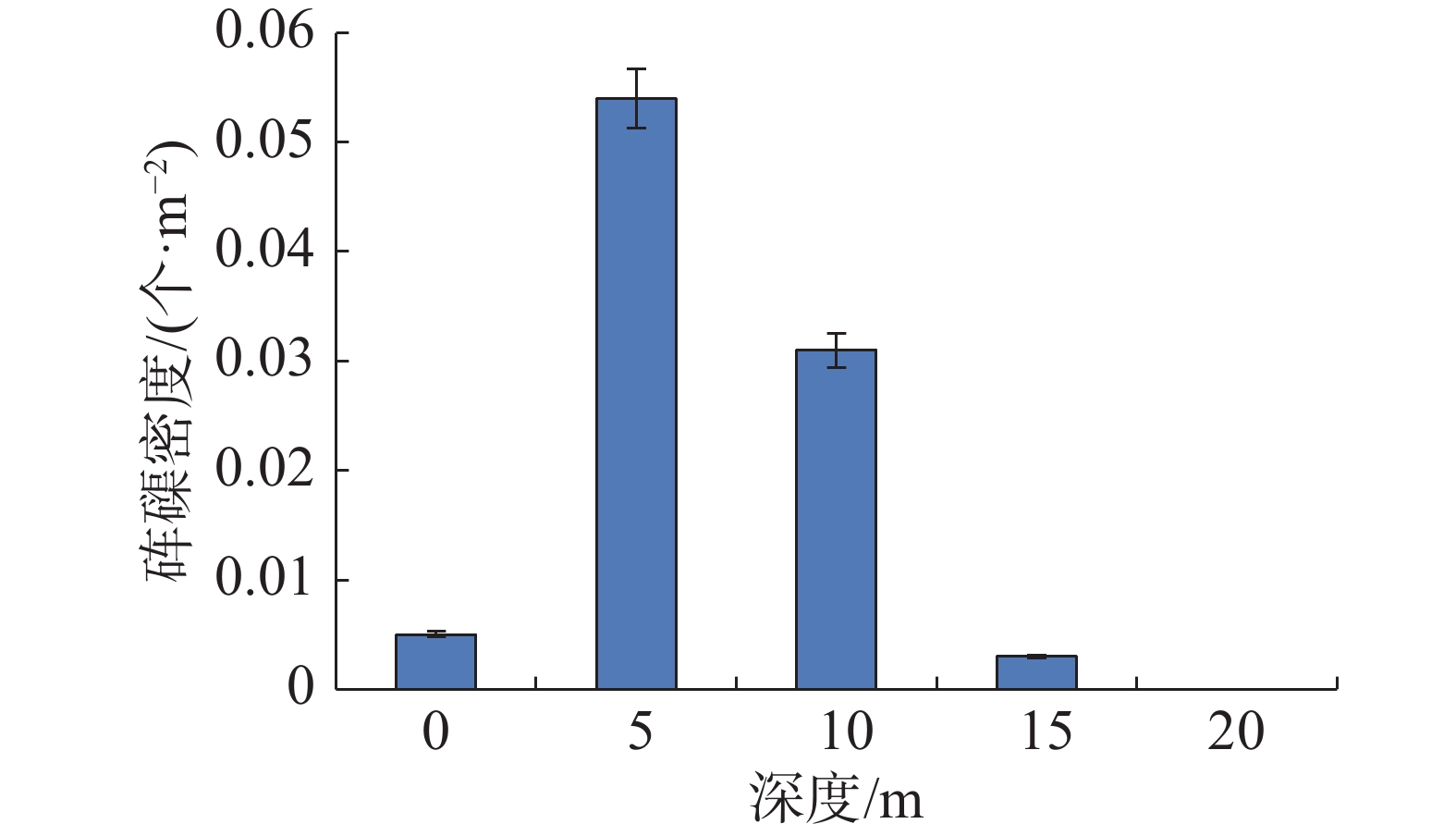
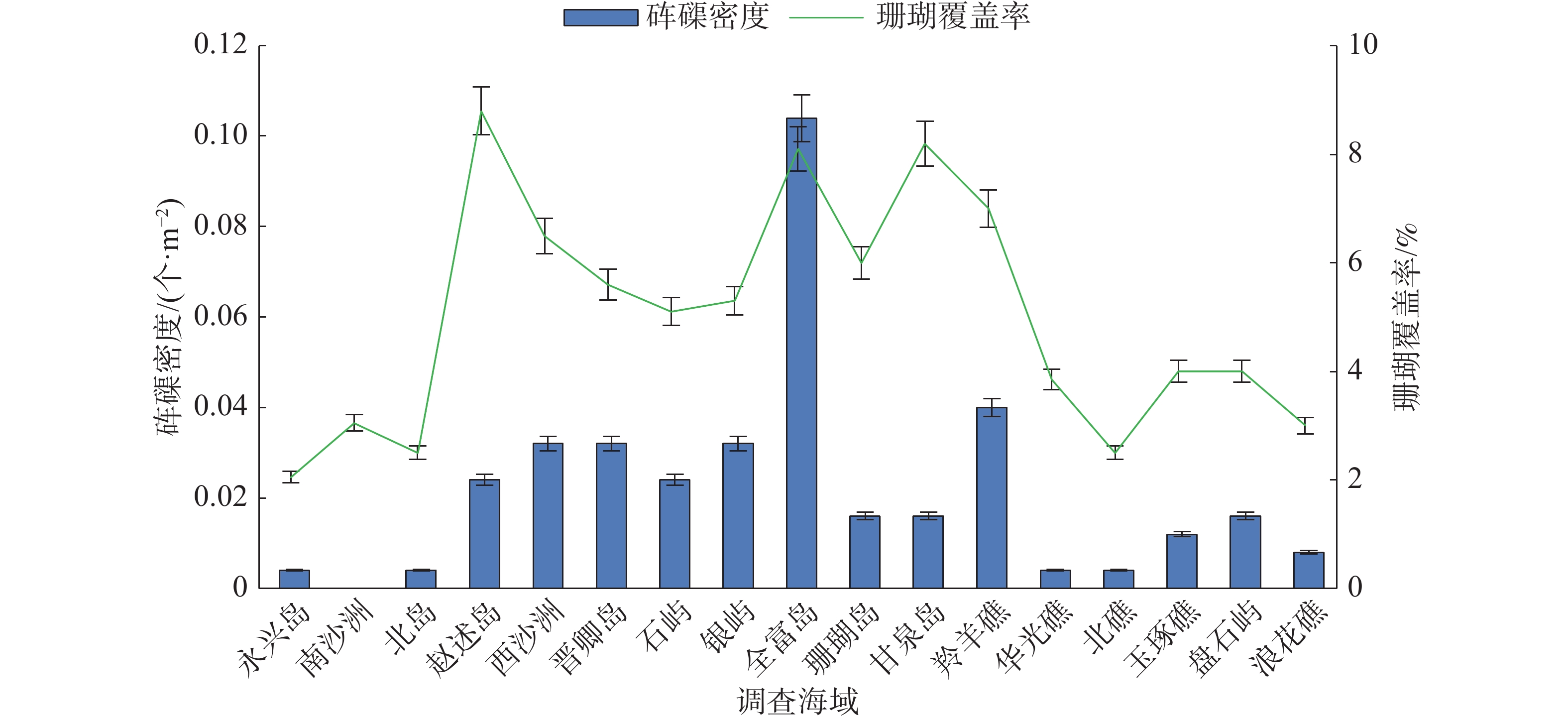
西沙群岛海域各岛礁调查站位砗磲密度和珊瑚覆盖率见图1,在各岛礁海域,除了南沙洲均调查到砗磲,砗磲主要分布于宣德环礁海域的赵述岛和西沙洲,永乐环礁海域的晋卿岛、石屿、银屿、全富岛和羚羊礁。在全富岛砗磲密度最高(0.104个·m−2),其次是西沙洲、晋卿岛和羚羊礁,砗磲密度均为0.040 个·m−2。在不同深度砗磲密度见图2,砗磲主要分布于5~10 m水深,其中5 m深度砗磲密度最高(0.054 个·m−2);其次是10 m深度(0.031 个·m−2)。在宣德环礁海域,以永兴岛为起点,砗磲密度沿着各岛礁顺序呈递增的趋势,永乐环礁海域砗磲密度无明显规律变化,永乐环礁海域砗磲平均密度显著大于宣德环礁海域。在27个调查站位中,全富岛砗磲密度显著大于其他岛礁海域(P<0.05),西沙群岛海域砗磲平均密度为0.026 个·m−2。
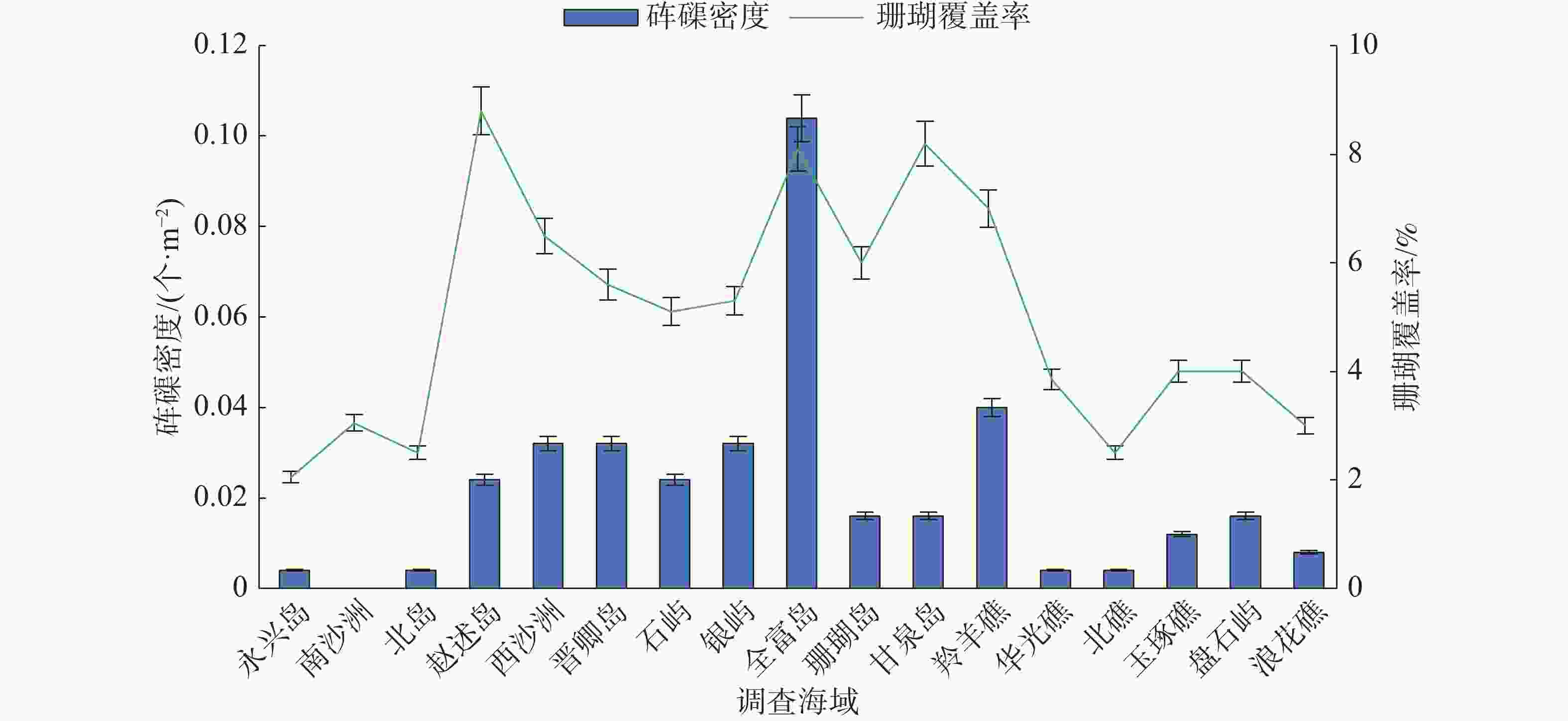
图 1 各岛礁海域砗磲密度与珊瑚覆盖率
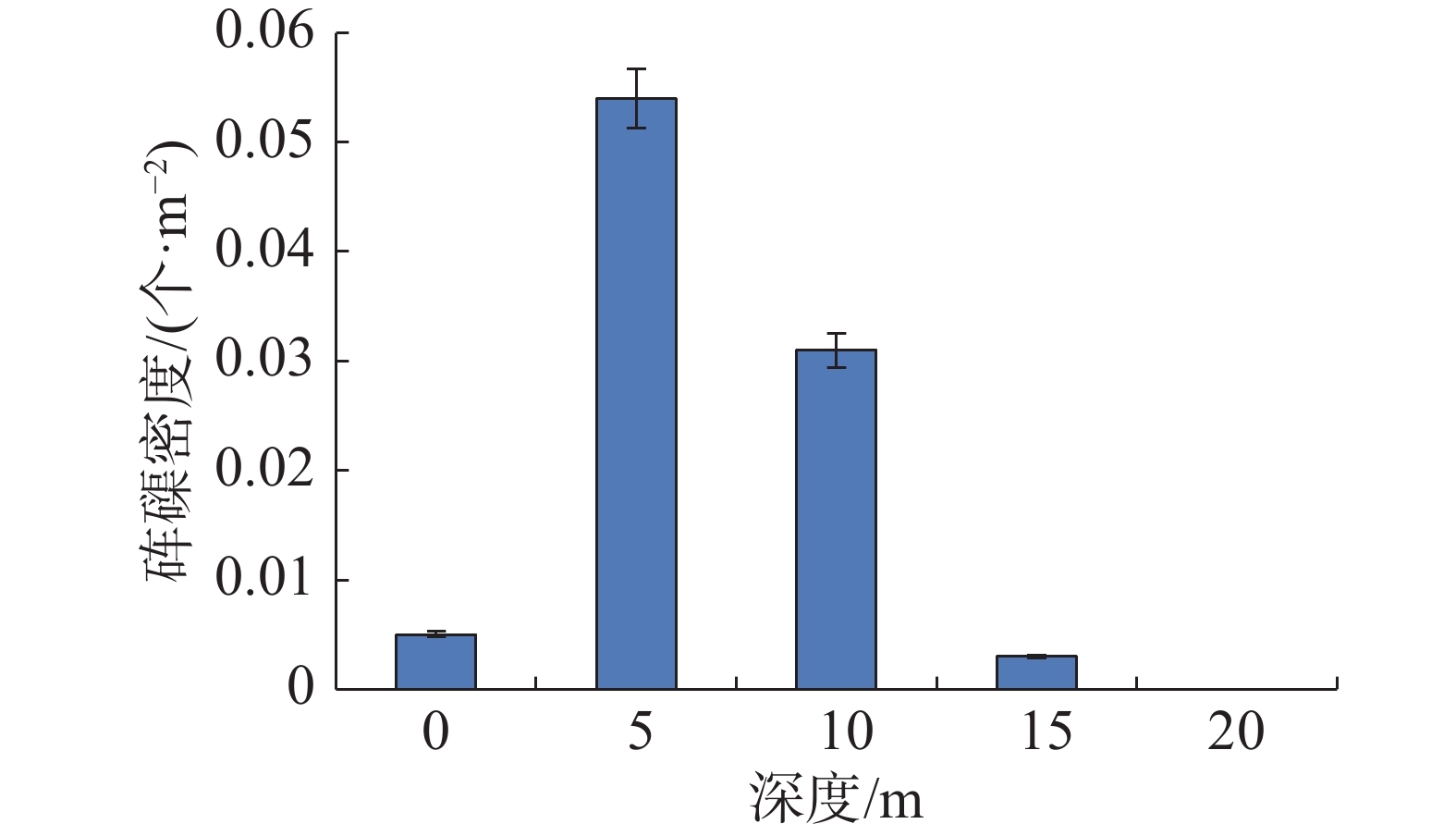
图 2 不同深度砗磲资源密度分布
-
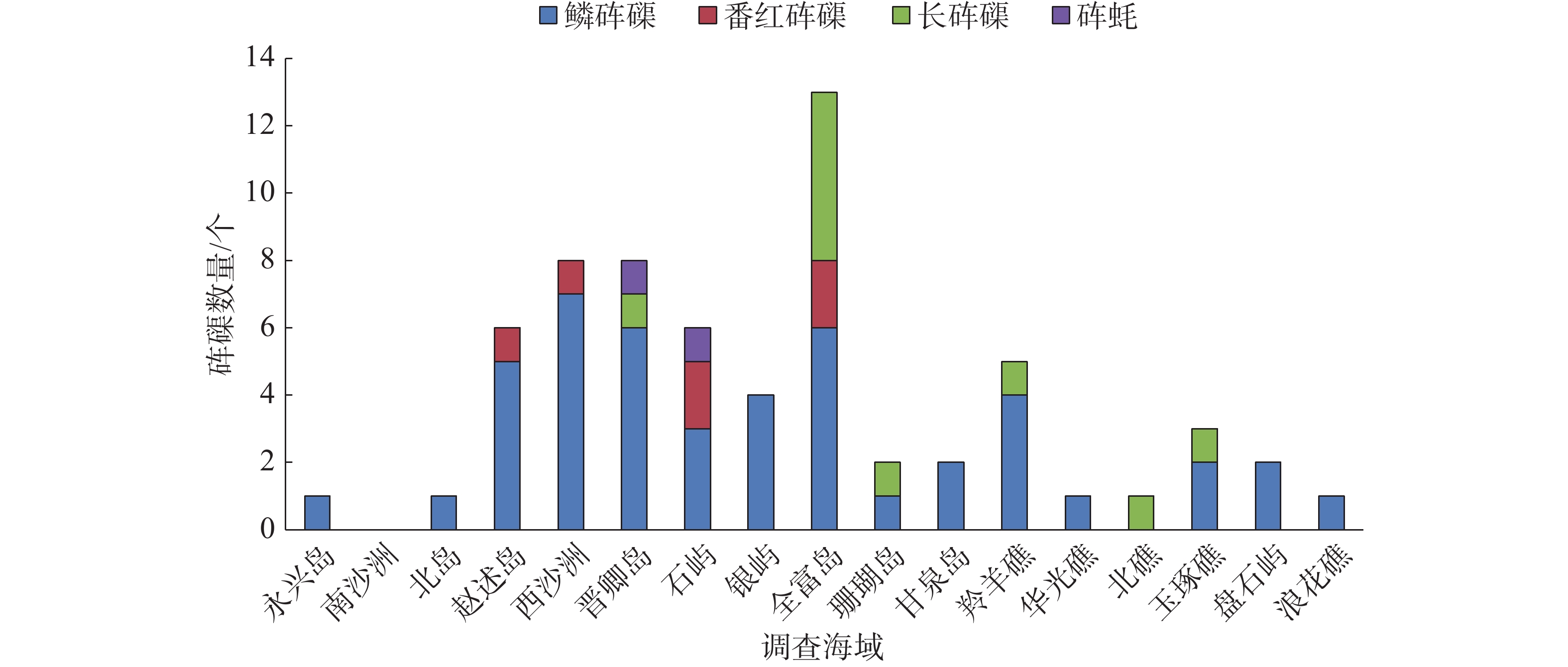
通过影像资料和形态学分析,共鉴定出4种砗磲,分别是鳞砗磲(Tridacna squamosa)、番红砗磲(Tridacna crocea)、长砗磲(Tridacna maxima)和砗蚝(Hippopus hippopus)。64个砗磲样品中,鳞砗磲46个,占样品总数的71.9%;长砗磲10个,占样品总数的15.6%;番红砗磲6个,占样品总数的9.4%;砗蚝2个,占样品总数的3.1%。对调查海域砗磲种类进行统计分析,在全富岛、晋卿岛和石屿各出现3个种类砗磲,在赵述岛、西沙洲、珊瑚岛、羚羊礁和玉琢礁各出现2个种类砗磲,在南沙洲未调查到砗磲,其余各岛礁均只出现1个种类砗磲,各岛礁海域具体出现砗磲种类见图3。

图 3 调查海域砗磲种类及数量分布
-
在27个调查站位中,按照地理位置,可代表性划分为8块群落区域,不同群落区域砗磲物种多样性指数见表1。分析结果表明,全富岛的砗磲物种多样性指数显著高于其他岛礁海域(P<0.05),丰富度指数和多样性指数分别为0.62和1.14;其次是石屿,砗磲物种多样性指数仅略低于全富岛,丰富度指数和多样性指数分别为0.61和1.01;西沙洲砗磲物种多样性指数最低,其丰富度指数和多样性指数分别为0.22和0.38。全富岛、石屿、珊瑚岛和玉琢礁均匀度指数均较高,说明在这些岛礁海域不同种类砗磲之间的数量差异较小,不同种类砗磲分布较为均匀。
表 1 调查海域砗磲物种多样性指数
群落区域 丰富度指数 多样性指数 均匀度指数 赵述岛 0.28±0.05 0.46±0.03 0.66±0.04 西沙洲 0.22±0.03 0.38±0.04 0.54±0.06 晋卿岛 0.42±0.04 0.74±0.09 0.67±0.08 石屿 0.61±0.10 1.01±0.12 0.90±0.09 全富岛 0.62±0.08 1.14±0.09 0.92±0.14 珊瑚岛 0.50±0.03 0.69±0.11 0.94±0.10 羚羊礁 0.32±0.02 0.50±0.06 0.72±0.07 玉琢礁 0.44±0.04 0.63±0.07 0.91±0.05 -
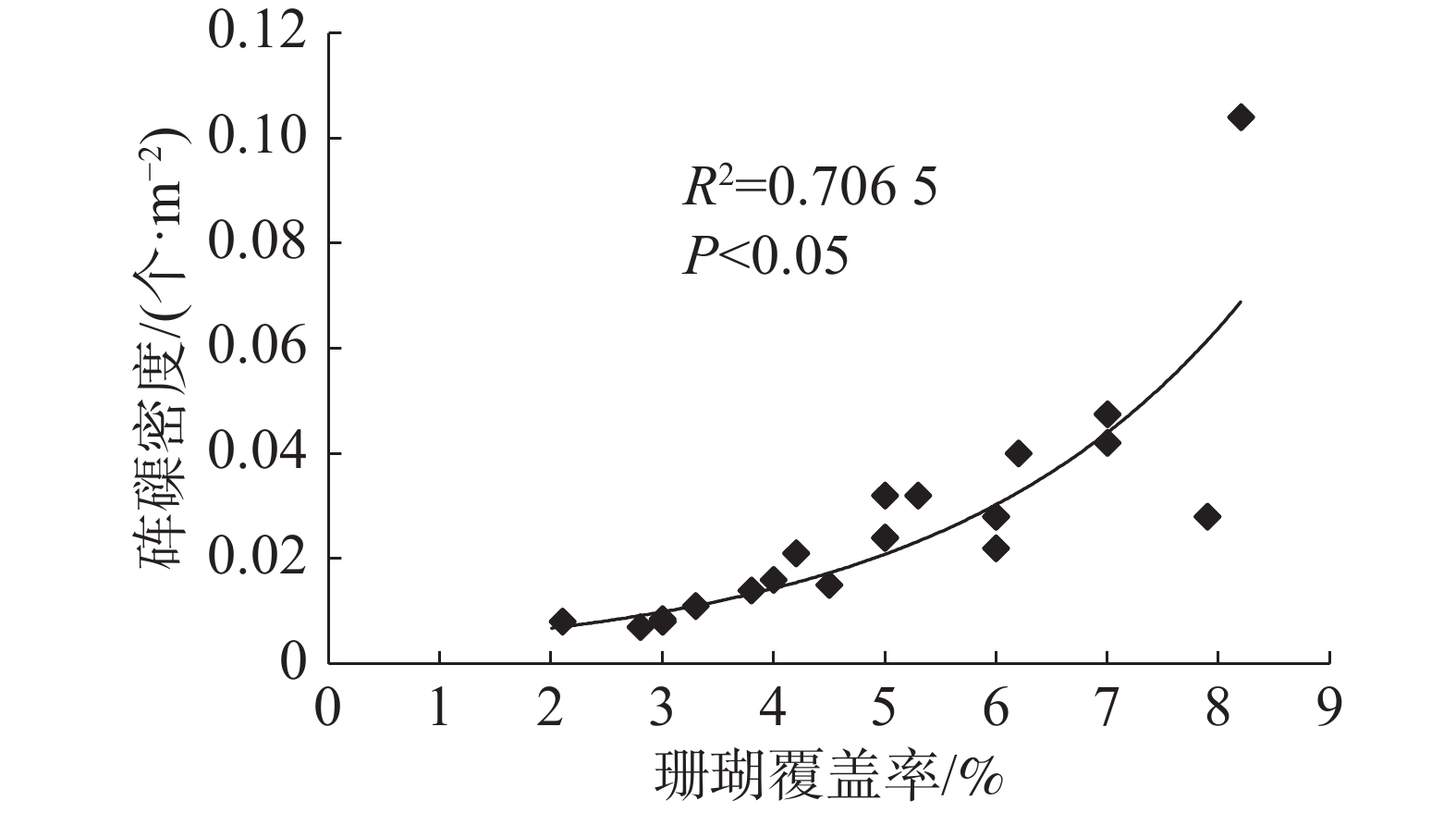
各岛礁海域砗磲密度和珊瑚覆盖率见图1。珊瑚覆盖率变化范围为2.0%~8.2%,平均覆盖率为4.5%。全富岛珊瑚覆盖率较高,达8.1%;羚羊礁、西沙洲和晋卿岛的珊瑚覆盖率也相对较高,达5.5%以上。珊瑚覆盖率较高的岛礁海域对应砗磲密度也较高,砗磲密度分布趋势与珊瑚覆盖率变化相似。将各岛礁海域砗磲密度与珊瑚覆盖率进行相关性分析(图4),结果显示R2为0.706 5,相关系数R的绝对值大于0.8(为0.840 5),砗磲密度与珊瑚覆盖率具有显著正相关性(P<0.05),说明砗磲密度随着珊瑚覆盖率增加而增大,珊瑚覆盖率是影响砗磲资源分布的重要因素。
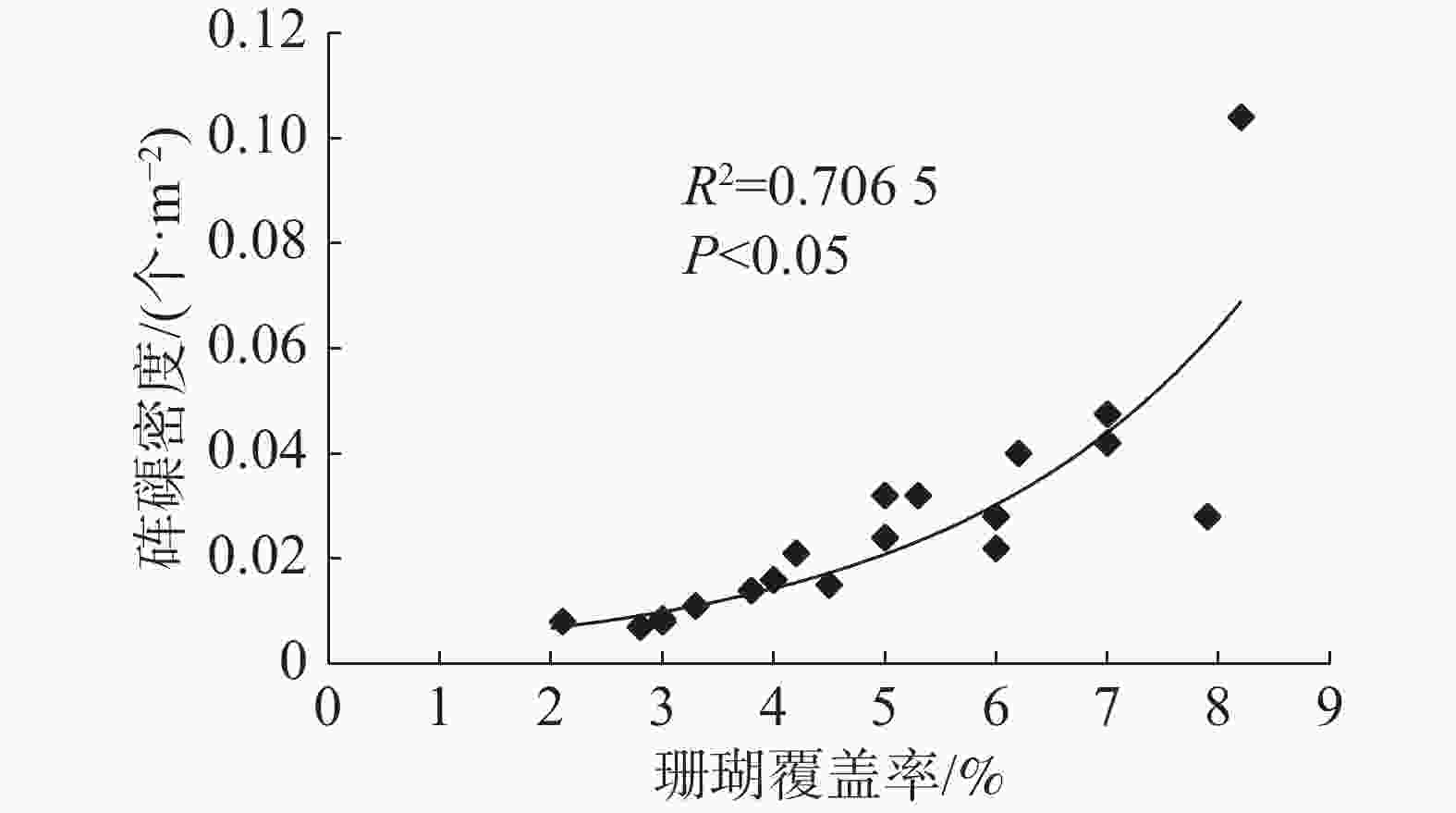
图 4 砗磲密度与珊瑚覆盖率相关性
-
砗磲是珊瑚礁生态系统中的优势生物,具有造礁、固礁和护礁功能,砗磲贝壳在珊瑚礁中占相当比例,为珊瑚的生长提供物理支撑[21-22]。本研究结果显示,砗磲主要分布于赵述岛、西沙洲、晋卿岛、石屿、银屿、全富岛和羚羊礁,其中在全富岛砗磲密度和物种多样性较高,对应全富岛珊瑚覆盖率也较高,砗磲密度与珊瑚覆盖率具有很强的正相关性,砗磲分布主要是受珊瑚覆盖率影响。刘春胜等[9]对三亚近海砗磲分布特征进行研究,结果表明砗磲主要分布于活珊瑚区,砗磲分布与珊瑚覆盖率密切相关,珊瑚覆盖率高的地方,砗磲分布多;李萌等[23]指出蜈支洲海域的珊瑚、礁石和砂石底质中均有砗磲,珊瑚礁区砗磲分布最多,砗磲密度与珊瑚覆盖率显著相关。本研究结论与以上观点相似。本研究结果显示,砗磲主要分布于5~10 m水深,在5 m水深砗磲密度最大,在20 m水深未调查到砗磲,砗磲密度随着水深增加呈下降趋势。吴钟解等[24]对海南西瑁洲岛周边海域造礁石珊瑚进行调查,发现造礁石珊瑚覆盖率随着水深增加而下降,这与本研究砗磲水深分布规律一致,表明砗磲密度主要受不同深度珊瑚覆盖率影响。
近十几年来,由于砗磲工艺品热销,渔民受经济利益驱使,在珊瑚礁盘大肆采挖砗磲贝壳[2,5]。李元超等[25]于2013年和2014年对西沙北礁进行珊瑚调查,两次调查期间共统计到40多艘小船每天进出礁坪采挖砗磲贝,破坏珊瑚礁生态系统,2012年和2014年珊瑚覆盖率分别为64.75%和52.83%。本次调查得到2017年西沙北礁珊瑚覆盖率约为4.50%,可知从2012—2017年,西沙北礁珊瑚覆盖率急剧下降。本研究西沙北礁砗磲分布较少,密度显著低于其他调查站位,笔者认为珊瑚覆盖率急剧下降是影响西沙北礁砗磲资源分布较低的主要原因。张振冬等[26]通过比较分析多年西沙造礁珊瑚监测结果,发现2007—2015年造礁珊瑚覆盖率呈现急剧下降趋势,覆盖率从2007年的53.8%下降到2015年的2.7%,近10年间下降了96%。本研究结果显示,近十几年来西沙海域珊瑚覆盖率急剧下降是造成砗磲资源处于濒危状态的重要原因。此外,由于全球气候变暖、海水酸化、长棘海星爆发等原因,造成海洋生态环境不断恶化,导致珊瑚覆盖率急剧下降[27-28],这些都是造成西沙海域砗磲资源严重下降的重要原因。
-
目前砗磲科所有物种已列入世界自然保护联盟濒危物种红色名录(IUCN红色名录)和濒危野生动植物种国际贸易公约附录II(CITES Appendix II)。我国发布的《国家重点保护野生动物名录》也已把库氏砗磲列为一级保护野生动物,把鳞砗磲、无鳞砗磲、长砗磲、番红砗磲和砗蚝列入二级保护野生动物,在本研究调查到的4种砗磲均属于二级保护野生动物。海南省发布的《海南省珊瑚礁和砗磲保护规定》已将上述名录中的所有砗磲物种纳入保护范围,明确规定对破坏珊瑚礁、砗磲的违法行为进行具体行政处罚,并规定构成犯罪的,依法追究刑事责任[29]。国内外发布的这些名录和法规为砗磲保护提供了强有力的政策支持,而本文基于西沙砗磲资源的调查则从科学角度为砗磲资源保护提供基础数据。建议在砗磲保护政策制定中,应强调重点保护珊瑚礁,恢复砗磲栖息地,坚决打击非法采挖珊瑚和捕捞砗磲的行为,开展砗磲增殖放流活动,补充周边海域砗磲种群数量。
本研究结果显示,西沙永乐环礁海域调查到的砗磲资源量占总量的62.5%,其中在全富岛砗磲的数量、密度和物种多样性指数均显著高于其他岛礁海域,其次是羚羊礁海域。根据本次西沙珊瑚礁调查数据,在全富岛和羚羊礁的珊瑚覆盖率也相对较高,说明该岛礁海域珊瑚礁生态系统良好,更有利于砗磲的栖息生长,可为砗磲的保育提供保障。因此,笔者建议在永乐环礁的全富岛和羚羊礁周边海域建立西沙砗磲种质资源保护区,加强重点保护。此外,建议西沙当地政府尽快向国家申报砗磲自然保护区,禁止采挖珊瑚和砗磲贝化石,禁止捕捞活体砗磲,同时开展砗磲苗种的孵化和培育工作,为今后实施砗磲增殖放流提供苗种,以期能尽快恢复西沙野生砗磲种群数量、保护濒危砗磲资源和维护珊瑚礁生态系统平衡。
A survey of giant clams in the waters at Xisha Islands
-
摘要: 砗磲是珊瑚岛礁的构成物种,对维护珊瑚礁生态系统平衡起着重要作用。近年来,由于非法捕捞和海洋生态环境影响,对我国西沙砗磲资源造成严重威胁。为掌握西沙海域砗磲资源现状,笔者于2017年6—7月采用截线样带法,对西沙主要岛礁海域砗磲的种类、分布及其与环境相关性进行了分析。结果显示,调查海域共发现4 种砗磲:鳞砗磲(Tridacna squamosa)、长砗磲(T. maxima)、番红砗磲(T. crocea)和砗蚝(Hippopus hippopus),分别占总数的71.9%、15.6%、9.4%和3.1%。西沙不同岛礁海域砗磲密度、物种多样性指数存在显著差异,砗磲平均密度为0.026个·m−2。砗磲密度与珊瑚覆盖率呈显著正相关(P<0.05,R2=0.7065),表明珊瑚覆盖率是影响砗磲分布的重要因素。Abstract: Giant clams are the largest living marine bivalves inhabiting coral reefs, and perform a significant ecological role in maintaining the balance of coral reef ecosystem. In recent years, giant clams in the waters at Xisha Islands, China are seriously threatened due to overfishing and the deterioration of the marine environment in the coastal areas. In order to master the present status of giant clam resources at Xisha Islands, a survey of giant clams in the waters at the main islands of Xisha Islands were made by using the method of line intercept transect to determine the species and distribution of the giant clams at Xisha Islands in the South China Sea from June to July 2017, and then analyze the correlation of the giant clams with the environment in the waters at the main islands of Xisha Islands. The results showed that there were four species of the giant clams in the survey area, including Tridacna squamosa, Tridacna crocea, Tridacna maxima and Hippopus hippopus, accounting for 71.9%, 15.6%, 9.4% and 3.1% of the total giant clams, respectively. The giant clams in the waters of different main islands of Xisha Islands are significantly different in habitat density and species diversity index. The average density of the giant clams is 0.026 ind/m2. Correlation analysis showed that there was a significant positive correlation between habitat density and coral reef coverage (P<0.05, R2 = 0.7065). This shows that coral reef coverage is an important factor affecting the distribution of giant clams. This result can provide scientific basis for the conservation of giant clam resources in Xisha Islands in China.
-
Key words:
- giant clam /
- resource distribution /
- coral reef coverage /
- Xisha Islands
-
表 1 调查海域砗磲物种多样性指数
群落区域 丰富度指数 多样性指数 均匀度指数 赵述岛 0.28±0.05 0.46±0.03 0.66±0.04 西沙洲 0.22±0.03 0.38±0.04 0.54±0.06 晋卿岛 0.42±0.04 0.74±0.09 0.67±0.08 石屿 0.61±0.10 1.01±0.12 0.90±0.09 全富岛 0.62±0.08 1.14±0.09 0.92±0.14 珊瑚岛 0.50±0.03 0.69±0.11 0.94±0.10 羚羊礁 0.32±0.02 0.50±0.06 0.72±0.07 玉琢礁 0.44±0.04 0.63±0.07 0.91±0.05  下载: 导出CSV
下载: 导出CSV
-
[1] LUCAS J S, LINDSAY S R. Density of clams and depth reduce growth in grow-out culture of Tridacna gigas. [J]. ACIAR Proceedings-Australian Centre for International Agricultural Research, 1993, 95(9): 67 − 73. [2] 董杨, 李向民. 砗磲资源保护、开发及其产业化发展前景[J]. 水产科学, 2015, 34(3): 195 − 200. [3] CABAITAN P C, GOMEZ E D, ALINO P M. Effects of coral transplantation and giant clam restocking on the structure of fish communities on degraded patch reefs [J]. Journal of Experimental Marine Biology and Ecology, 2008, 357(1): 85 − 98. doi: 10.1016/j.jembe.2008.01.001 [4] MUNRO J L, HESLINGA G A. Prospects for the commercial cultivation of giant clams (Bivalvia: Tridacnidae) [J]. Proceedings of the Gulf and Caribbean Fisheries Institute, 1982: 122 − 134. [5] 周勤, 王玉芬, 姜秀凤. 砗磲研究现状及在我国南海开展养殖的前景[J]. 热带海洋学报, 2014, 33(2): 87 − 93. doi: 10.3969/j.issn.1009-5470.2014.02.012 [6] 黄晖, 尤丰, 练健生, 等. 西沙群岛海域造礁石珊瑚物种多样性与分布特点[J]. 生物多样性, 2011, 19(6): 710 − 715. [7] 王红勇, 孔伟. 保护南海砗磲贝类资源刻不容缓[J]. 河北渔业, 2015(4): 62 − 62. doi: 10.3969/j.issn.1004-6755.2015.04.022 [8] 崔丹, 王辉, 刘君, 等. 海南岛沿岸海域砗磲资源调查及保护前景展望[J]. 海洋通报, 2019, 38(5): 527 − 532. doi: 10.11840/j.issn.1001-6392.2019.05.006 [9] 刘春胜, 吴川良, 李秀保, 等. 三亚近海诺亚砗磲分布特征及底质对其外套膜颜色的影响[J]. 水生生物学报, 2021, 45(3): 636 − 642. doi: 10.7541/2021.2020.017 [10] 黄晖, 董志军, 练健生. 论西沙群岛珊瑚礁生态系统自然保护区的建立[J]. 热带地理, 2008, 28(6): 540 − 544. doi: 10.3969/j.issn.1001-5221.2008.06.010 [11] 林鸿民, 陈惠. 西沙群岛海洋环境资源保护与利用[J]. 中国发展, 2017, 17(1): 46 − 49. [12] 邓远球, 邓岳文, 梁飞龙. 砗磲人工繁育研究进展[J]. 水产养殖, 2019(11): 18 − 21. doi: 10.3969/j.issn.1004-2091.2019.11.007 [13] NEO M L, ECKMAN W, VICENTUAN K, et al. The ecological significance of giant clams in coral reef ecosystems [J]. Biological Conservation, 2015, 181: 111 − 123. doi: 10.1016/j.biocon.2014.11.004 [14] 梁文, 黎广钊, 谭趣孜, 等. 一种适合珊瑚礁生态区用海项目的礁系生物群落调查方法[J]. 广西科学院学报, 2012, 28(3): 212 − 215. doi: 10.3969/j.issn.1002-7378.2012.03.013 [15] 顾茂彬. 世界名贝鉴赏图谱[M]. 郑州: 河南科学技术出版社, 2005. [16] 张素萍. 中国海洋贝类图鉴[M]. 北京: 海洋出版社, 2008. [17] MARGALEF D R. Information theory in ecology [J]. United Kingdom:International Journal of General Systems, 1958, 3: 36 − 71. [18] SHANNON E C, WEAVER W. The mathematical theory of communication[M]. Urbana: University of Illinois Press, 1948: 125. [19] PIELOU E C. Species-diversity and pattern-diversity in the study of ecological succession [J]. Journal of Theoretical Biology, 1966, 10(2): 370 − 383. doi: 10.1016/0022-5193(66)90133-0 [20] 吴程宏, 张羽翔, 赵海龙, 等. 海南文昌冯家湾人工鱼礁区渔业资源养护效果初步评估[J]. 海洋湖沼通报, 2020(6): 158 − 167. doi: 10.13984/j.cnki.cn37-1141.2020.06.020 [21] 喻子牛. 砗磲人工繁育、资源恢复与南海岛礁生态牧场建设[J]. 科技促进发展, 2020, 16(2): 231 − 236. [22] ANDRFOUT S, WYNSBERGE S V, MAZOUNI N G, et al. Climate variability and massive mortalities challenge giantclam conservation and management efforts in French Polynesiaatolls [J]. Biological Conservation, 2013(160): 190 − 199. [23] 李萌, 李秀保, 顾志峰, 等. 海南岛近海砗磲(Tridacninae)资源时空变化 ——以蜈支洲岛为例[J]. 海洋与湖沼, 2021, 52(6): 1521 − 1529. [24] 吴钟解, 张光星, 陈石泉, 等. 海南西瑁洲岛周边海域造礁石珊瑚空间分布及其生态系统健康评价[J]. 应用海洋学学报, 34, 1: 133 − 140. [25] 李元超, 韩有定, 陈石泉, 等. 砗磲采挖对珊瑚礁生态系统的破坏——以西沙北礁为例[J]. 应用海洋学学报, 2015, 34(4): 518 − 524. doi: 10.3969/J.ISSN.2095-4972.2015.04.009 [26] 张振冬, 邵魁双, 杨正先, 等. 西沙珊瑚礁生态承载状况评价研究[J]. 海洋环境科学, 2018, 37(4): 487 − 492. doi: 10.12111/j.cnki.mes20180403 [27] 吴钟解, 王道儒, 李元超, 等. 西沙监控区浮游植物生态现状[J]. 热带作物学报, 2010, 31(6): 1020 − 1025. doi: 10.3969/j.issn.1000-2561.2010.06.027 [28] 李元超, 吴钟解, 梁计林, 等. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 2019, 64(33): 3478 − 3484. [29] 海南省人大常委会. 海南省珊瑚礁和砗磲保护规定[N]. 海南日报, 2016−12−16 (A10). DOI: 10.28356/n.cnki.nhlrb.2016.000151. -

 点击查看大图
点击查看大图
图(4) / 表 (1)
计量
- 文章访问数: 376
- HTML全文浏览量: 137
- PDF下载量: 36
- 被引次数: 0


