




-
香蕉枯萎病又称巴拿马病或黄叶病,其病原菌为尖孢镰刀菌古巴专化型(Fusarium oxysporum f. sp. cubense),其中4号生理小种(Foc4)危害最严重。香蕉枯萎病是典型的真菌维管束病害,主要通过土壤、染病的繁殖材料、灌溉水和农具等传播,已造成了全球香蕉产业的严重经济损失[1-2]。目前香蕉枯萎病的主要防治措施有生物防治[3-5]、化学防治[6]、抗病品种选育[7-8]等。香蕉枯萎病的致病相关基因在致病过程中的作用也得到了研究,如过氧化氢酶、氯离子电导蛋白、多聚半乳糖醛酸酶和丝裂原活化蛋白激酶MAPK等方面取得的研究进展[9-13],为筛选靶向药物防控香蕉枯萎病提供了借鉴。海藻糖(trehalose) 是一种非还原性二糖,在细菌、真菌和植物中广泛存在,可作为储能物质,为细胞代谢提供能量,并涉及对各种外界环境胁迫的应答,以协助生物体顺利度过逆境 [14-17]。海藻糖酶是真菌中海藻糖唯一的水解酶,在代谢过程中起着关键作用,而人和动物体中均无此代谢途径。以海藻糖酶为靶标位点设计杀菌剂,可以提高农药的作用效率和对非靶标生物的安全性。根据其最适pH和调节特性,真菌海藻糖酶分为酸性(非调节性)和中性(调节性)海藻糖酶[18]。在酵母细胞中,海藻糖是由中性海藻糖酶分解而非酸性海藻糖酶,并且两者之前不具有同源性[19-20]。目前在不同真菌中对中性海藻糖酶基因的功能也得到了系列研究,如在构巢曲霉、根内球囊霉、灰葡萄孢和酿酒酵母等真菌中获得中性海藻糖酶基因敲除突变体,并对突变体的功能进行了研究分析[21-24]。氰烯菌酯是一类对镰刀菌具有特异抑菌活性的新型氰基丙烯酸脂类杀菌剂,具有保护、治疗、内吸、输导等作用,能有效抑制镰刀菌菌丝生长,使分生孢子生长畸形,有望替代其他多类杀菌剂[25-28]。氰烯菌酯对禾谷镰刀菌有较强的抑制活性,以对氰烯菌酯具有抗性的禾谷镰刀菌YP-1为材料,对其进行测序,发现YP-1中有132个基因发生突变,其中包括中性海藻糖酶基因 [29]。为了探索中性海藻糖酶NTH1基因在香蕉枯萎病菌致病过程中的作用及其是否参与尖孢镰刀菌对氰烯菌酯的抗药性,笔者采用基因敲除技术敲除了香蕉枯萎病菌的中性海藻糖酶基因NTH1,并通过敲除突变体的表型及对氰烯菌酯的敏感性进行测定分析,旨在丰富该基因的具体功能知识,并为开发农药防治的靶标基因提供科学依据。
-
香蕉枯萎病4号生理小种(F. oxysporum f. sp. cubense race 4,Foc4)野生型菌株37,大肠杆菌E.coli DH5a,pCT 74质粒均由本实验室保存。盆栽巴西蕉幼苗(株高20~30 cm,4~5枚叶片)。
-
94.9%氰烯菌酯原药(西北农林科技大学刘西莉教授赠)用DMSO(二甲基亚砜)溶解,配成10 g·L−1贮存液。Congo Red(刚果红,CR)购于BIO BASIC INC公司;Calcofluor White Stain(荧光增白剂,CFW)购于sigma-aldrich公司;Hygromycin B(潮霉素B,50 mg·L−1)溶液购于Wako公司。
Chitinase(几丁质酶)、Driselase (崩溃酶)、Lysing enzyme (溶壁酶)均购自sigma-aldrich公司;LA Taq、Premix TaqTM、克隆载体pMDTM19-T Vector购自TaKaRa公司;真菌DNA提取试剂盒和总RNA提取试剂盒购自OMEGA公司;琼脂糖凝胶DNA回收试剂盒、质粒提取试剂盒购自TIANGEN公司。
培养基:PDA 培养基、PDB培养基、LB 培养基、MM培养基、LR培养基、SR培养基、Top Agar培养基。
-
PTC200 PCR仪(MJ RESEARCH);Quantstudio 6荧光定量PCR仪;电泳仪DY CP-31E;微量紫外可见分光度计Thermo Scientific NanoDrop 2000c;涡旋振荡器IKA MS3 basic;全自动样品快速研磨仪Tissueiyser-64;Eppendorf台式高速离心机5417R / 5810R;双目生物显微镜XCP-2 C、尼康生物智能荧光显微镜Ni-E;生物大分子分析仪Fusion FX7。
-
将野生型菌株在PDA培养基上培养7 d后,收集菌丝。按照Fungal DNA Kit(OMEGA)说明书提取基因组DNA。
-
用DNAMAN7.0软件进行引物设计,并由北京六合华大基因科技股份有限公司合成引物(表1)。
表 1 本实验的引物序列
Table 1. Sequences of the primers used in the study
引物名称 Primer 序列 Sequence(5′→3′) HYG-F CTTGGCTGGAGCTAGTGGAGGT HYG-R CCCGGTCGGCATCTACTCTATTC HYG-R1 GGATGCCTCCGCTCGAAGTA HYG-F1 CGTTGCAAGACCTGCCTGAA HPT-LBCK GACAGACGTCGCGGTGAGTT HPT-RBCK TCTGGACCGATGGCTGTGTAG FOC4NTH1-LBCK TAAACCCAGCCACGACAC FOC4NTH1-HPH-LB-R ACCTCCACTAGCTCCAGCCAAGCCGTTGACTTACACCAGCAG FOC4NTH1-HPH-RB-F GAATAGAGTAGATGCCGACCGGGCTGACCTACCAGCACTCTCG FOC4NTH1-RBCK GGCAAGCCACTCATCTTCA FOC4NTH1-LB-F ACATTGAGCAACATCCGAGT FOC4NTH1- RB-R TGATGTCCTCCAAACTCACG FOC4NTH1-CKFP ACCTCTTCGTGGTCTGTCCT FOC4NTH1-CKRP ATCTGTTTCCGCCCATACTC FOC4NTH1-F CTCCTTCTCACCGCTCTATTA FOC4NTH1-R CGTATGTGTTCGGCTCAAA 在NCBI(
http://www.ncbi.nlm.nih.gov )数据库中通过F.graminearum PH-1 neutral trehalase NTH1基因(GenBank登录号: XM_011320499.1)在香蕉枯萎病菌热带4号生理小种菌株的全基因组数据库 (GenBank登录号: JH658279.1)中进行Blast分析,并设计cDNA全长引物对:FOC4NTH1-F/FOC4NTH1-R,扩增出香蕉枯萎病4号生理小种的NTH1基因。将连接转化后的阳性克隆产物送至深圳华大基因科技有限公司测序。测序结果在NCBI(
http://www.ncbi.nlm.nih.gov )数据库中进行同源性序列分析,使用MEGA7.0软件以Neighbor-Joining(NJ)构树法构建系统进化树。 -
采用Split-Marker重组技术构建基因敲除片段。以野生型菌株基因组DNA为模板,扩增NTH1基因上下游片段,引物为FOC4NTH1-LBCK和FOC4NTH1-HPH-LB-R、FOC4NTH1-HPH-RB-F和FOC4NTH1-RBCK(表1)。以pCT74质粒的DNA为模板,用引物HYG-F和HYG-R扩增潮霉素(HYG)抗性基因片段。将扩增得到的NTH1基因上游和下游片段分别与扩增的潮霉素(HYG)抗性基因片段混合为模板,引物FOC4 NTH1-LB-F和HYG-R1、HYG-F1和FOC4NTH1-RB-R(表1)扩增用于同源重组的上游和下游片段。
-
参照王飞燕等[30]的方法,稍作改进,制备香蕉枯萎病菌的原生质体。将在PDB培养基中振荡培养3 d的野生型菌株,用3层灭菌擦镜纸过滤菌液,37 ℃,150 r·min−1振荡酶解菌丝3 h,过滤收集原生质体。原生质体转化参照徐齐军等[31]的方法,稍作改进。各取100 μL的上下游扩增重组片段,加入到300 μL的原生质体中(1.0×108个·mL−1),混匀后,缓慢滴加2 mL 40% PTC溶液。最后加入到再生培养基(含有100 mg·L−1 Ampicillin和100 mg·L−1 Hygromycin B)中,随后将长出的单菌落挑取到新的含潮霉素抗性的PDA平板上,待转化子在含有潮霉素抗性平板上培养3代后可进行后续实验。
-
采用CTAB[32]法提取NTH1敲除转化子的基因组DNA和野生型菌株基因组DNA为模板,以HYG-F/HYG-R、FOC4NTH1-LBCK/HPT -LBCK、HPT-RBCK/FOC4NT H1-RBCK(表1)为正筛引物,以FOC4NTH1 - CKFP/FOC4NTH1-CKRP(表1)为负筛引物,进行PCR扩增,并将得到的PCR产物进行琼脂糖凝胶电泳,根据扩增的条带鉴定出Δnth1敲除突变体。
参照杨军等[33]的方法,稍作改进。将Δnth1接种到PDB培养基中,28 ℃,180 r·min−1振荡培养3 d。过滤收集孢子,制备分生孢子悬浮液,并将孢子浓度调至106,107,108 个·mL−1,并分别涂布到TOP培养基(含有100 mg·L−1 Ampicillin和100 mg·L−1 Hygromycin B)上,28 ℃培养10 h,在显微镜下,用挑针将单个分生孢子挑取到新的PDA培养基(含有100 mg·L−1 Ampicillin和100 mg·L−1 Hygromycin B)上。
-
分别在PDB培养基中振荡培养3 d的野生型菌株和Δnth1菌株,用3层灭菌擦镜纸过滤收集分生孢子,使用血球计数板计数,加入适量的无菌水重悬分生孢子并调整孢子浓度至2×106个·mL−1。采用伤根侵菌的盆栽致病性测定法。每个菌株接种15株蕉苗,并以水作为对照,试验重复3次。接种45 d后参照MOHAMED等[34]的病情分级标准,进行病情指数统计分析。
-
分别将野生型菌株和Δnth1菌株接种于含有不同浓度氰烯菌酯的PDA培养中,28 ℃培养7 d,每个处理重复3个培养皿,试验重复3次,最终计算野生型菌株和Δnth1的抑制中浓度EC50。
-
(1)生长速率:将野生型菌株和Δnth1菌株接种到MM培养基平板上,28 ℃培养6 d,并每隔1天进行拍照观察和测量菌落直径,每次3个平板,重复3次。(2)产孢量:将在PDA培养基上培养7 d的野生型菌株和Δnth1菌株,用10 mL的无菌水从平板上收集新鲜的分生孢子,然后使用血球计数板计数,每次3个平板,重复3次。(3)孢子萌发:参照Feng等[35]的方法,稍作改进。用10 mL的无菌水分别收集野生型菌株和Δnth1菌株在PDA平板上的分生孢子(培养7 d),并将孢子浓度调至106个·mL−1,置于28 ℃潮湿培养箱中培养。分别于8,12,24 h用显微镜进行观察并记录,试验重复3次,最后测定分生孢子萌发率。
-
在PDA培养基上活化7 d的野生型菌株和Δnth1菌株,用打孔器制取菌落边缘处6 mm的菌块,分别接种于含有Congo Red(终浓度100 mg·L−1),D-Glucose(终浓度1 mol·L−1),SDS(十二烷基硫酸钠,终浓度0.1 mmol·L−1),Calcofluor White Stain(终浓度100 mg·L−1),Sorbitol(山梨醇,终浓度1.2 mol·L−1),NaCl(终浓度0.7 mol·L−1),MgCl2(终浓度0.1 mol·L−1),KCl(终浓度1 mol·L−1)和H2O2(终浓度4 mmol·L−1)的基本培养基(minimal medium,MM)平板上,以未做任何处理的MM平板作对照,28 ℃培养箱中培养6 d,每隔1天观察测量菌落直径大小并拍照,计算抑制率,每个处理重复3个培养皿,试验重复3次。
将在PDA培养基上活化7 d的野生型菌株和Δnth1菌株,接种于MM培养基上,并分别置于20 ℃,32 ℃培养箱中培养6 d,以28 ℃为对照,每个处理重复3个培养皿,试验重复3次,观察在低温和高温条件下对菌株生长的影响。
-
香蕉枯萎病菌的NTH1基因全长4 383 bp,cDNA全长2 217 bp,编码738个氨基酸,序列登录号:XM_031206259。在NCBI(
http://www.ncbi.nlm.nih.gov )数据库通过Blast搜索,利用GenBank中已知的氨基酸序列进行同源性分析,发现香蕉枯萎病菌中性海藻糖酶基因的氨基酸序列,与禾谷镰刀菌(F. graminearum)和假禾谷镰刀菌(F. pseudograminearum)等多种真菌的中性海藻糖NTH1基因具有高度相似性(图1)。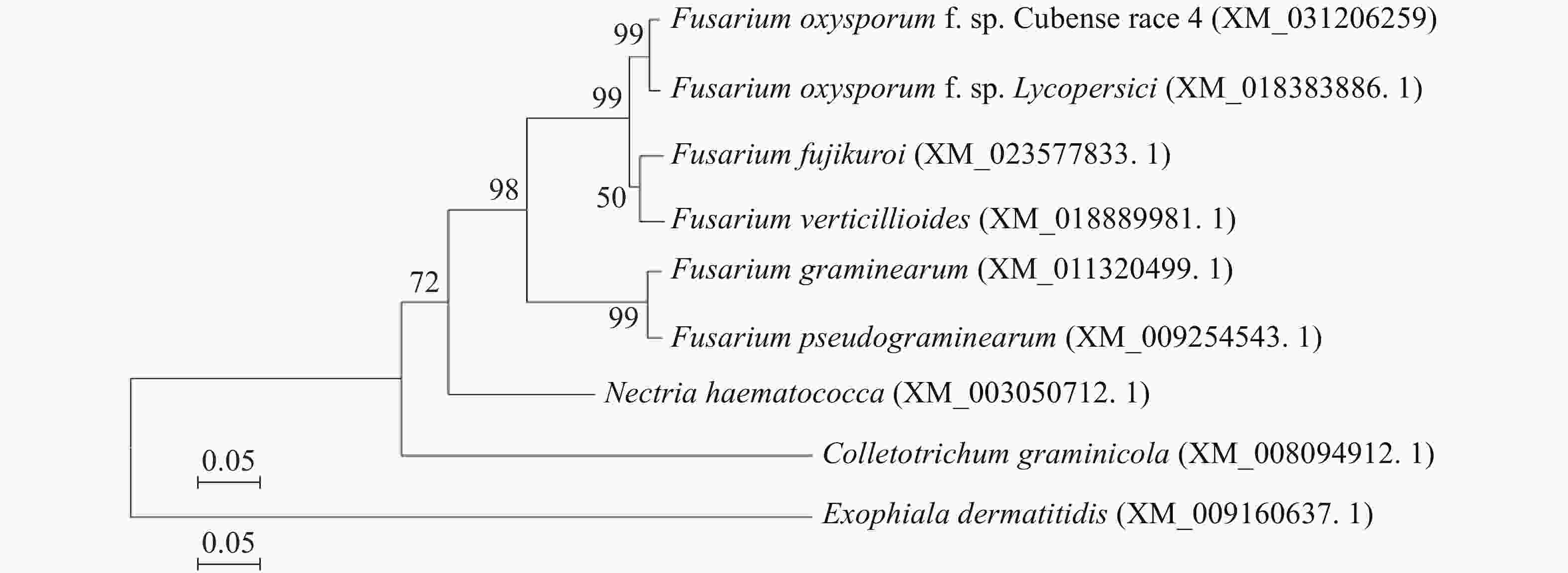
图 1 NTH1基因系统进化分析
Figure 1. Phylogenetic analysis of NTH1
-
第1轮PCR扩增的NTH1基因上下游片段和潮霉素(HYG)抗性基因片段:以野生型菌株基因组DNA为模板,以引物FOC4NTH1-LBCK和FOC4NTH1- HPH-LB-R、FOC4NTH1-HPH-RB-F和FOC4NTH1-RBCK,扩增NTH1基因的上下游片段,大小为1 903 bp和2 055 bp,同时以pCT 74质粒的DNA为模板,用引物HYG-F和HYG-R扩增出潮霉素(HYG)抗性基因片段,大小为1 376 bp(图2-B)。
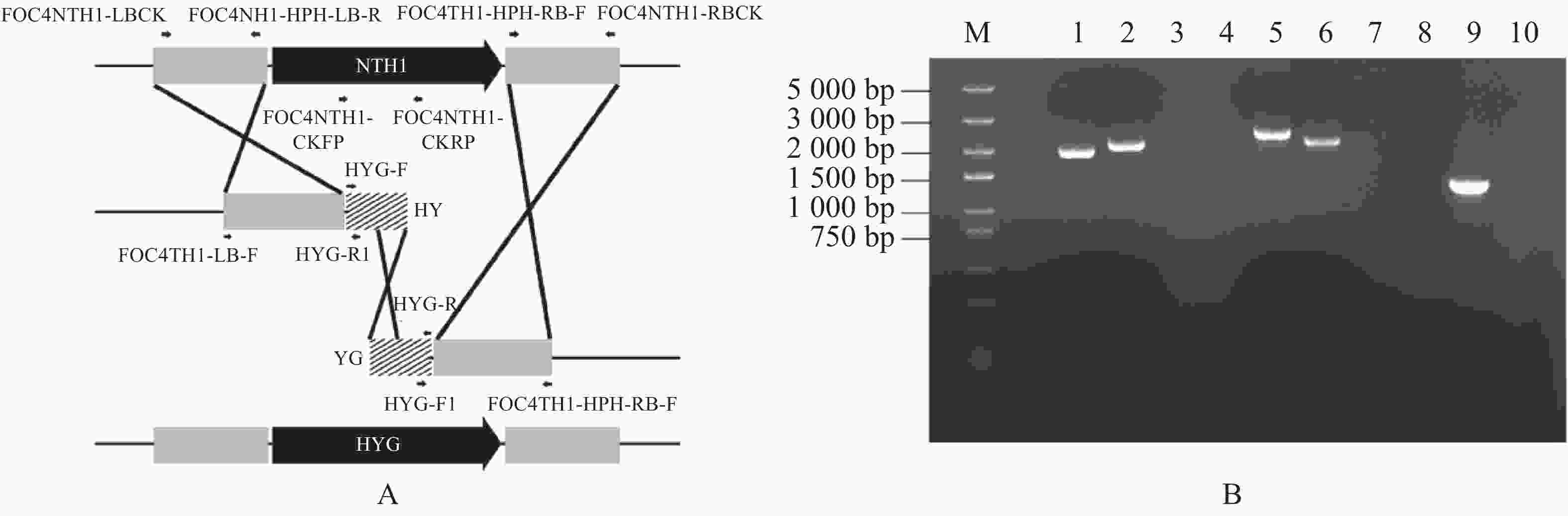
图 2 Split-Marker重组技术的片段扩增
Figure 2. Fragment amplification in Split-Marker recombination
第2轮PCR重叠片段扩增:NTH1基因上游扩增片段和潮霉素(HYG)抗性基因扩增片段混合物为模板,以引物FOC4NTH1-LB-F和HYG-R1,扩增重叠的上游片段,大小为2 379 bp;NTH1基因下游扩增片段和潮霉素(HYG)抗性基因扩增片段混合物为模板,再用引物HYG-R1和FOC4NTH1-RB-R,扩增重叠的下游片段,大小为2 138 bp(图2-B)。
-
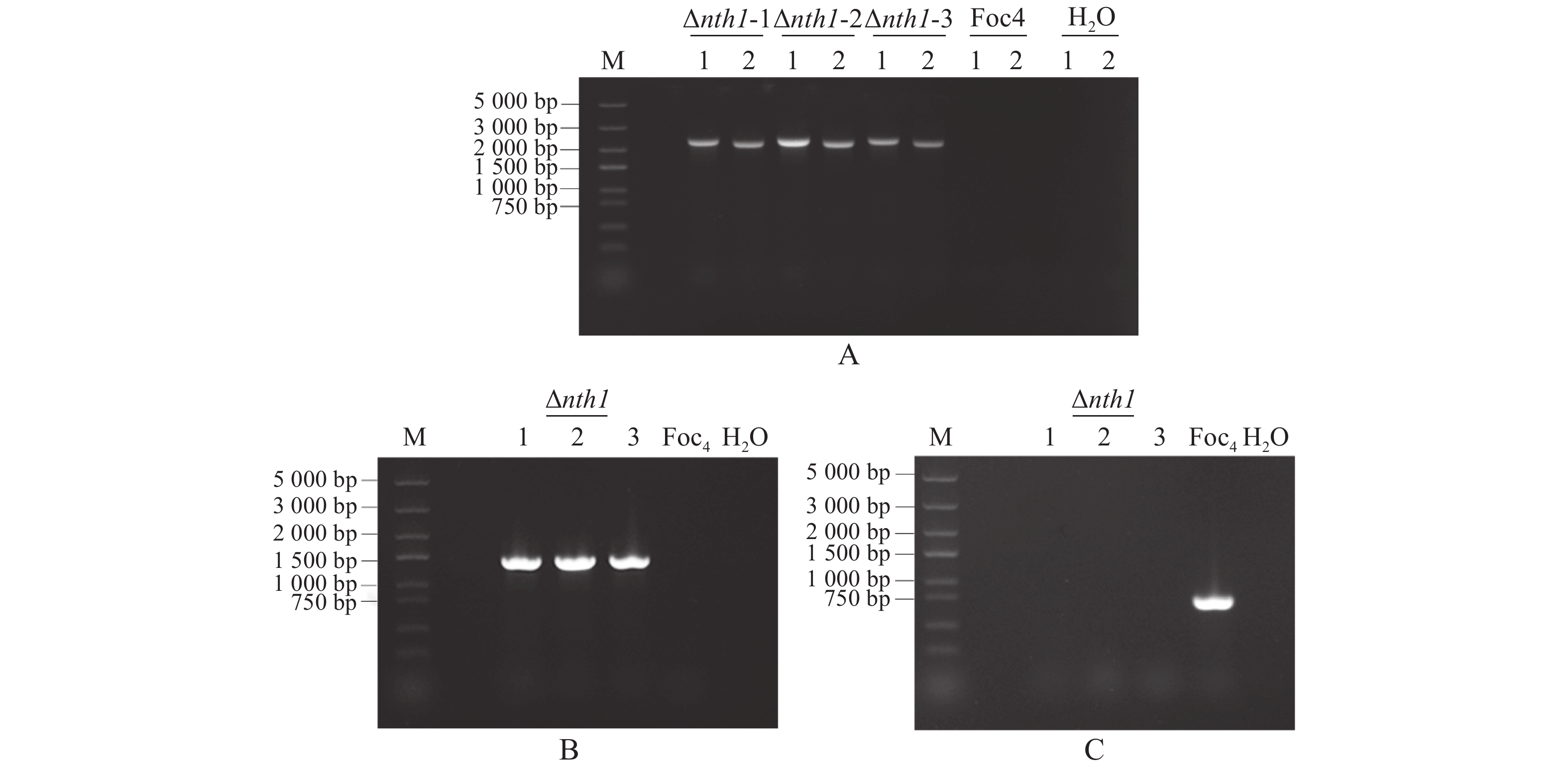
NTH1敲除转化子在潮霉素抗性培养基上培养3代后,提取基因组DNA并以此为模板进行PCR扩增,以野生型基因组DNA为对照,用正筛引物HYG-F /HYG-R,FOC4NTH1-LBCK/HPT-LBCK,HPT-RBCK/FOC4NTH1-RBCK进行PCR扩增,阳性NTH1基因敲除转化子应有条带,大小分别为1 376,2 297,2 160 bp,对照组野生型DNA模板无条带(图3-A,B)。负筛引物FOC4NTH1-CKFP/ FOC4NTH1-CKRP进行PCR扩增,阳性NTH1敲除转化子应没有条带,而对照组野生型DNA模板有条带,大小为737 bp(图3-C)。从图3可知,3个转化子已被鉴定为阳性,分别命名为Δnth1-1,Δnth1-2,Δnth1-3,选取其中的Δnth1-1菌株进行后续实验。
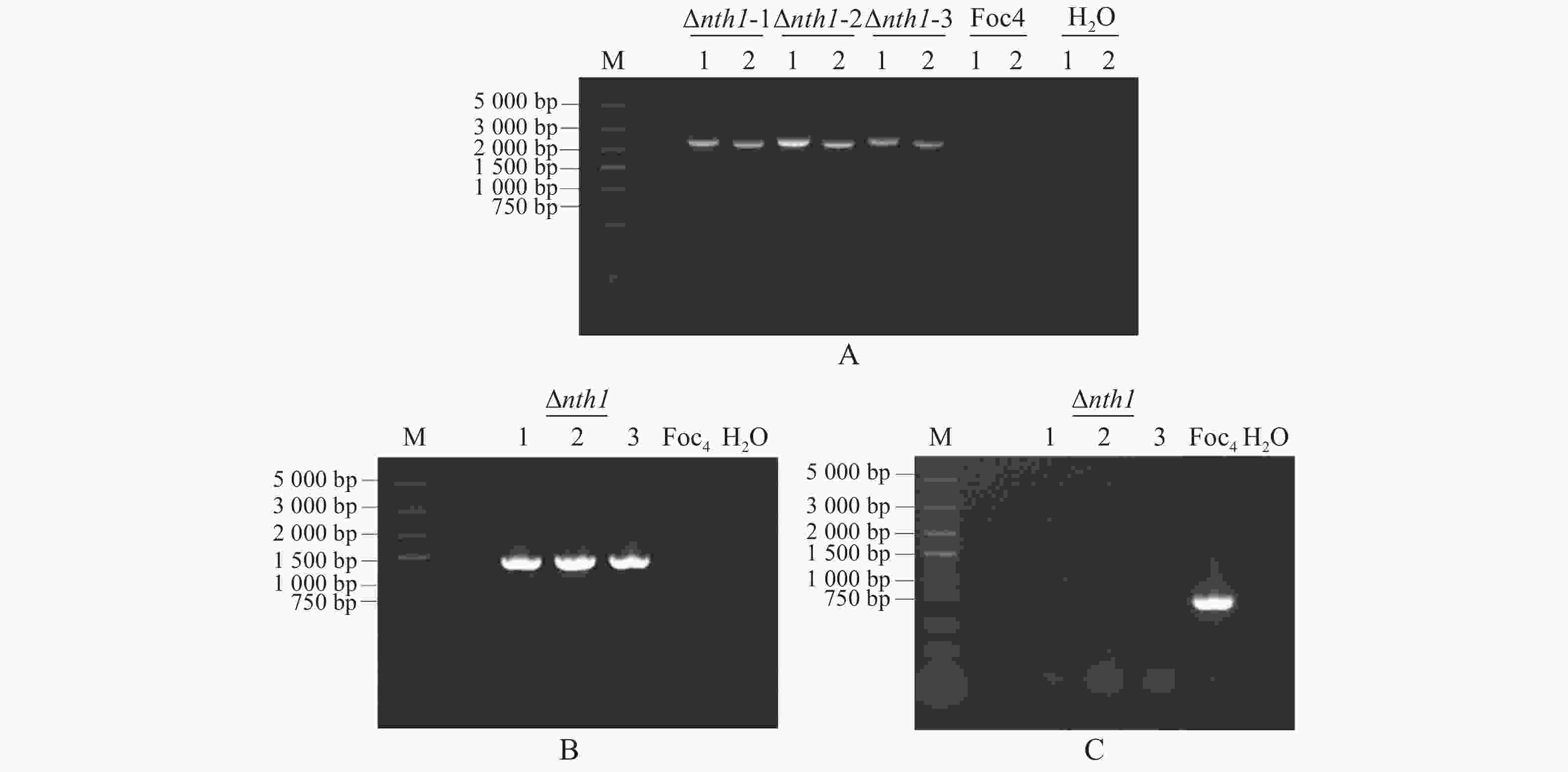
图 3 NTH1敲除突变体的筛选验证
Figure 3. Screening and validation of NTH1 knockout mutant
-
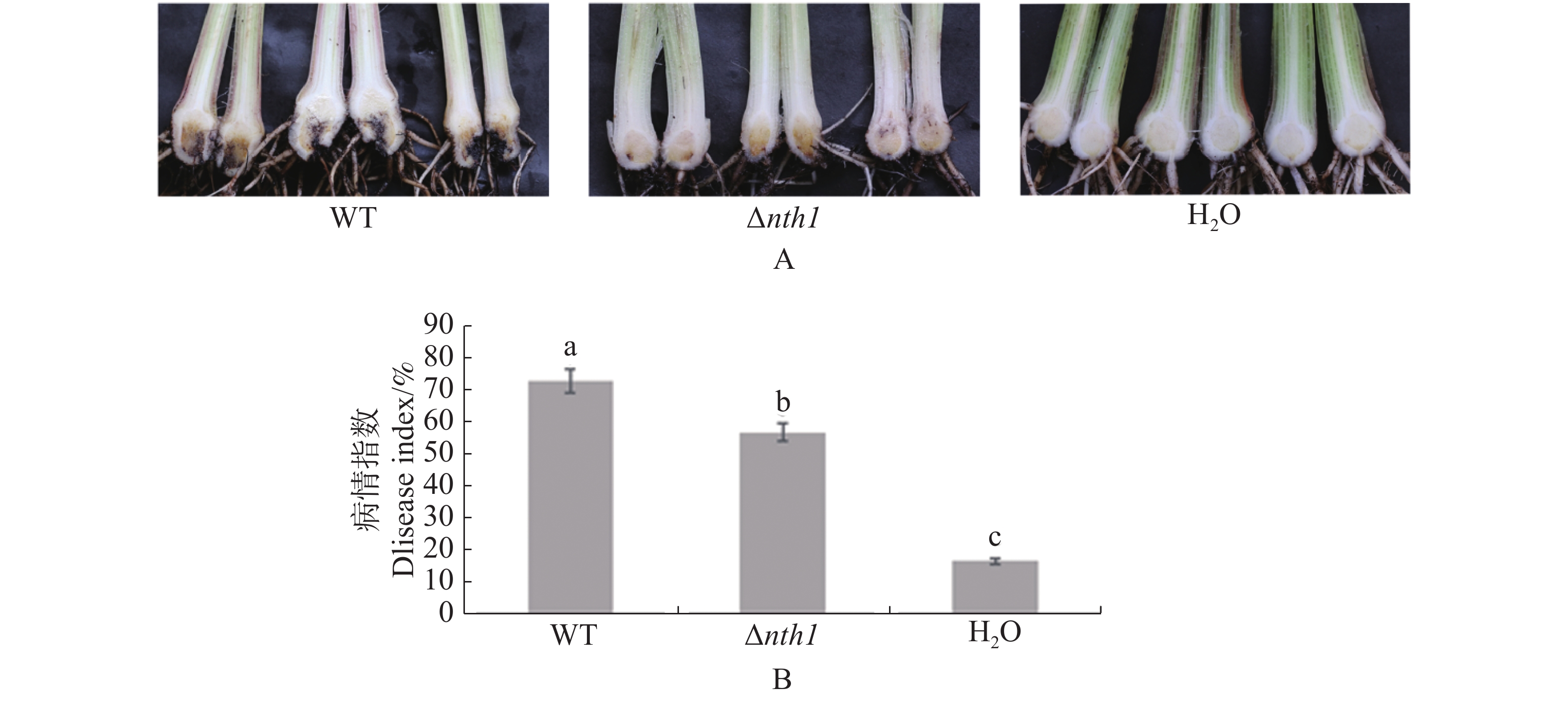
分别将野生型菌株和Δnth1菌株接种巴西蕉苗,45 d后观察发病情况。发现对照组巴西蕉苗无病害症状出现,而接种野生型菌株的巴西蕉苗外围叶片黄化,出现枯萎现象,且植株假茎下部出现褐色斑点状或条状斑块,球茎切开后观察到严重的褐化现象,有的植株枯萎死亡,接种突变体的部分巴西苗少数叶片出现小面积黄色斑块,球茎有部分褐化现象(图4-A)。结果(图4-B)显示,在病情指数方面,Δnth1菌株(56.6)低于野生型菌株(72.6)。
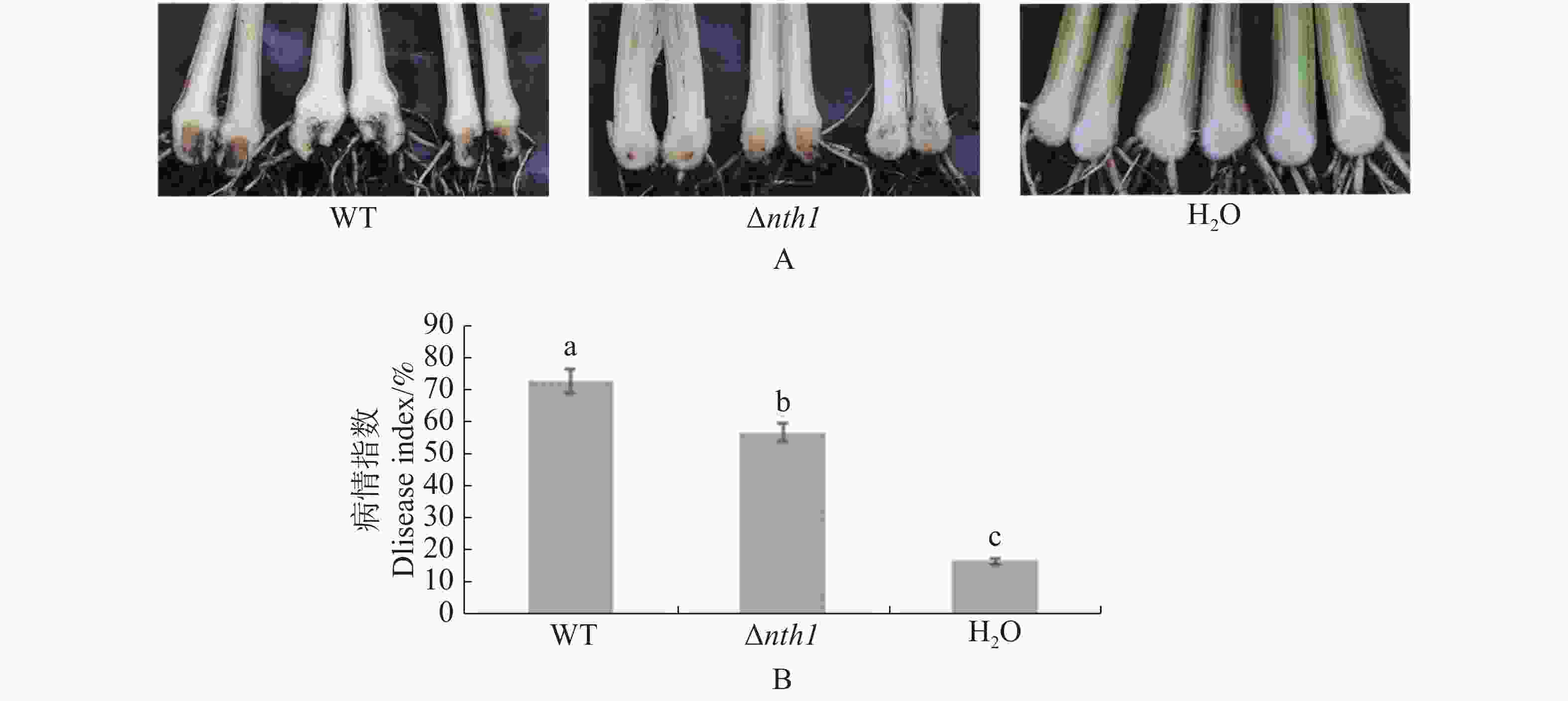
图 4 Δnth1的致病力测定
Figure 4. Pathogenicity test of Δnth1
-
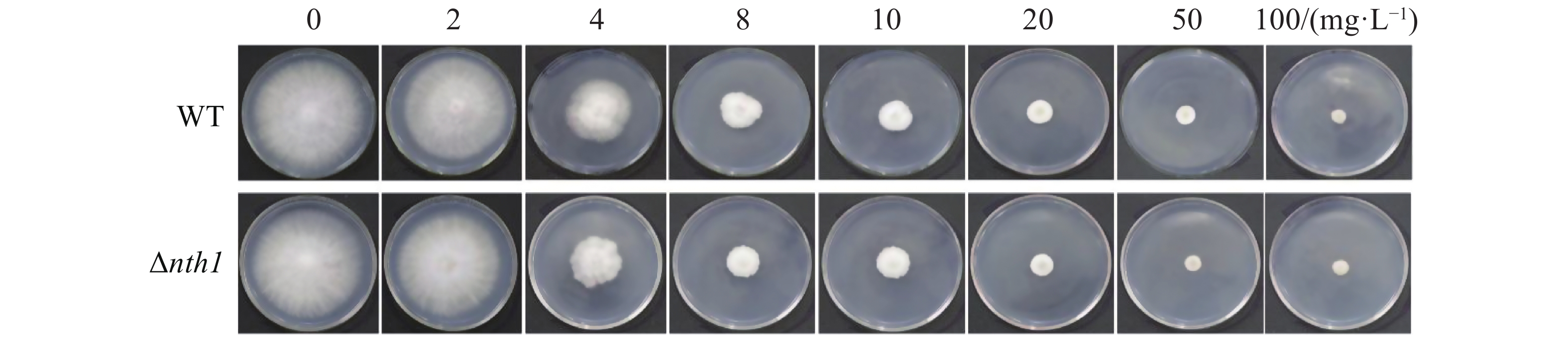
将野生型菌株和Δnth1菌株分别接种于含有2,4,8,10,20,50,100 mg·L−1氰烯菌酯的PDA平板中,培养7 d,分别测定野生型菌株和Δnth1菌株对氰烯菌酯的敏感性,并计算在不同氰烯菌酯浓度的生长抑制率(图5,6)。结果表明,Δnth1对氰烯菌酯的敏感性(EC50=6.07 mg·L−1)与野生型对氰烯菌酯的敏感性(EC50=6.22 mg·L−1)相比没有明显差异。
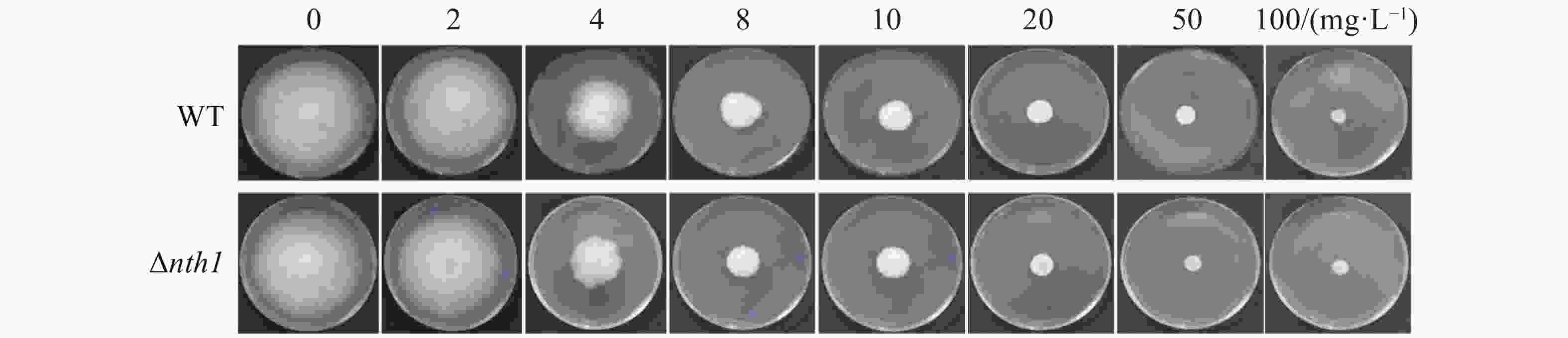
图 5 野生型和Δnth1在含有不同浓度氰烯菌酯的PDA平板上的菌落生长情况
Figure 5. Colony growth of WT and Δnth1 on PDA supplemented with phenamacril at different concentrations

图 6 野生型和Δnth1在不同氰烯菌酯浓度下的菌落生长抑制率
Figure 6. Inhibition rates of WT and Δnth1 under different concentrations of phenamacril
-
将野生型菌株和Δnth1菌株接种到MM培养基平板上,每隔一天测量记录菌落生长直径(图7-A)。结果表明,Δnth1菌株与野生型菌株相比,菌落生长速度下降,差异显著(图7-B)。产孢量计算结果表明Δnth1的孢子数(0.9×107 个·mm−2)与野生型的孢子数(1.073×107个·mm−2)差异不显著。孢子萌发率测定显示Δnth1菌株的孢子萌发率显著降低(图7-C)。

图 7 Δnth1的生物学表型分析
Figure 7. Biological phenotypic analysis of Δnth1
-
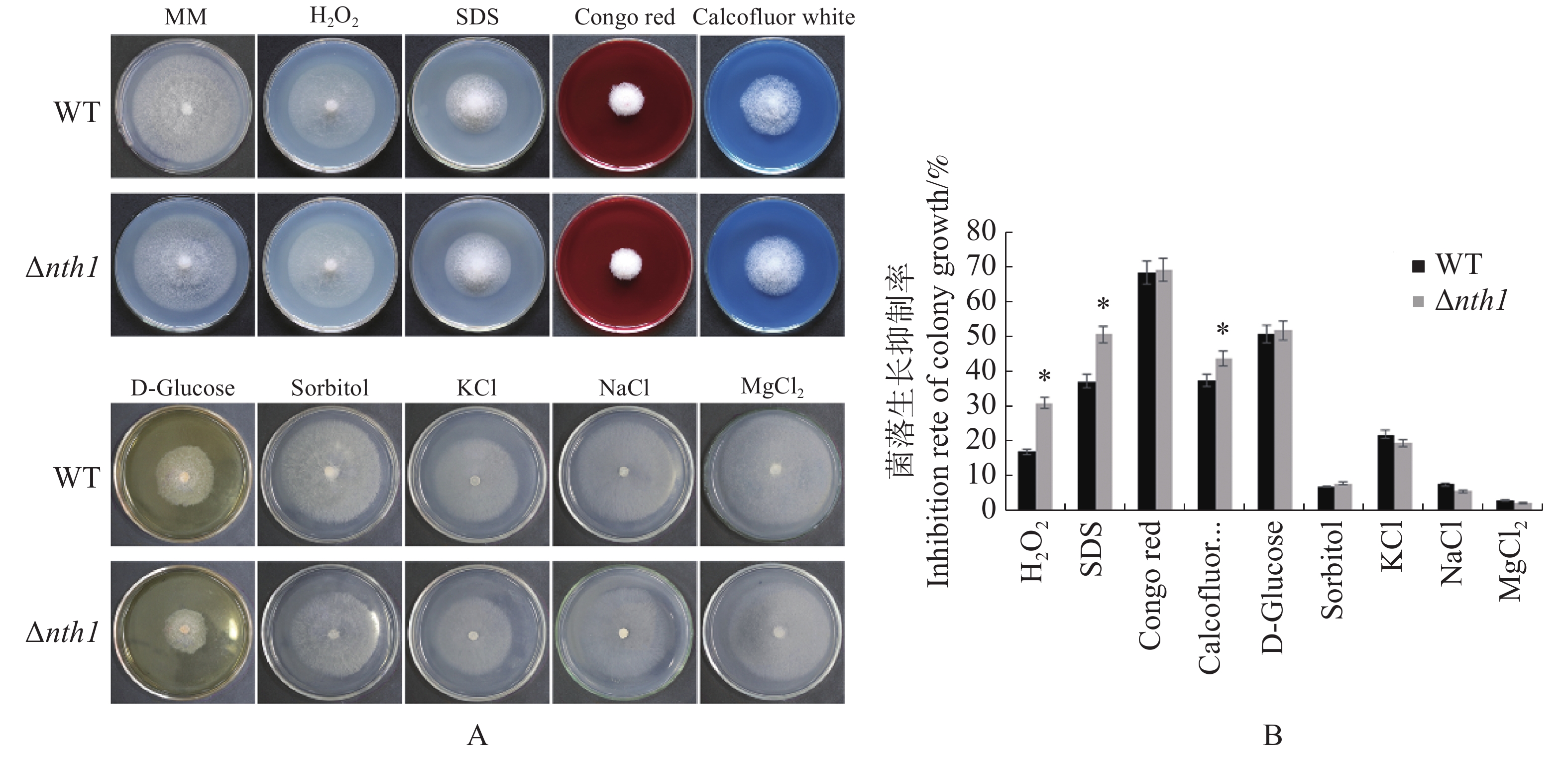
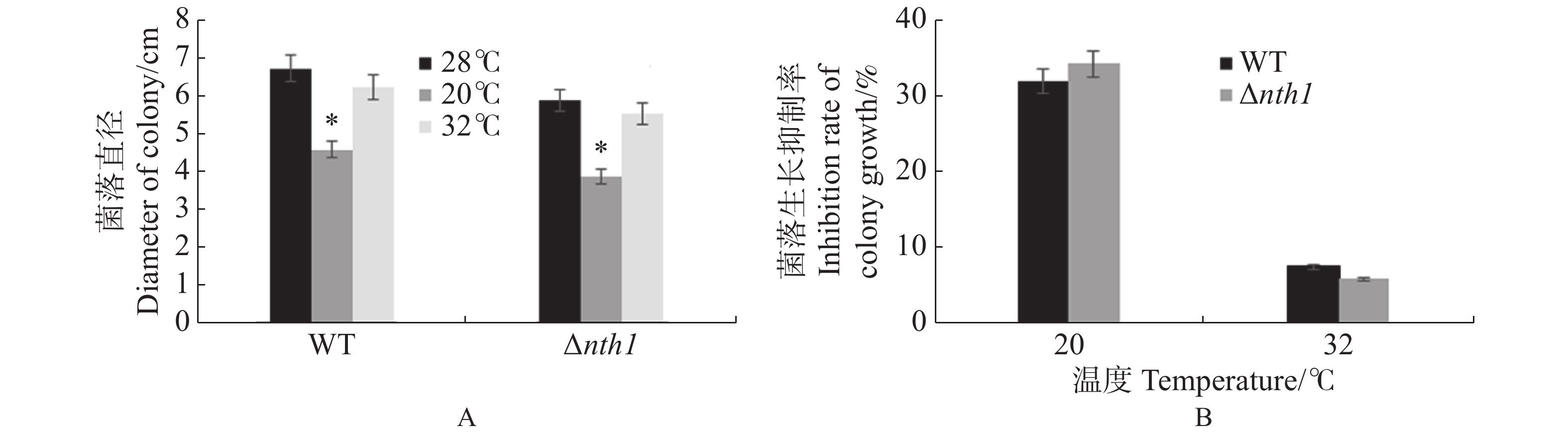
在含有100 mg·L−1 CR(Congo Red),0.7 mol·L−1 NaCl,0.1 mol·L−1 MgCl2和1 mol·L−1 KCl的胁迫因子存在时,野生型与Δnth1的生长均受到相同程度的抑制,差异不明显;在含有1 mol·L−1 D-Glucose 和1.2 mol·L−1/L Sorbitol的培养基中,野生型与Δnth1的生长均受到相同程度的抑制;在含有100 mg·L−1 CFW(Calcofluor White),4 mmol·L−1 H2O2,0.1 mmol·L−1 SDS的培养基上,野生型与Δnth1的生长均受到不同程度的抑制,与野生型相比,突变体受抑制程度更明显(图8);在20 ℃低温和32 ℃高温条件下,Δnth1的菌落生长形态和大小,与野生型相比无明显差异(图9)。结果表明,Δnth1对细胞壁选择性压力和氧化压力敏感,但对渗透压、高糖和高、低温胁迫的敏感度与野生型没有差异。
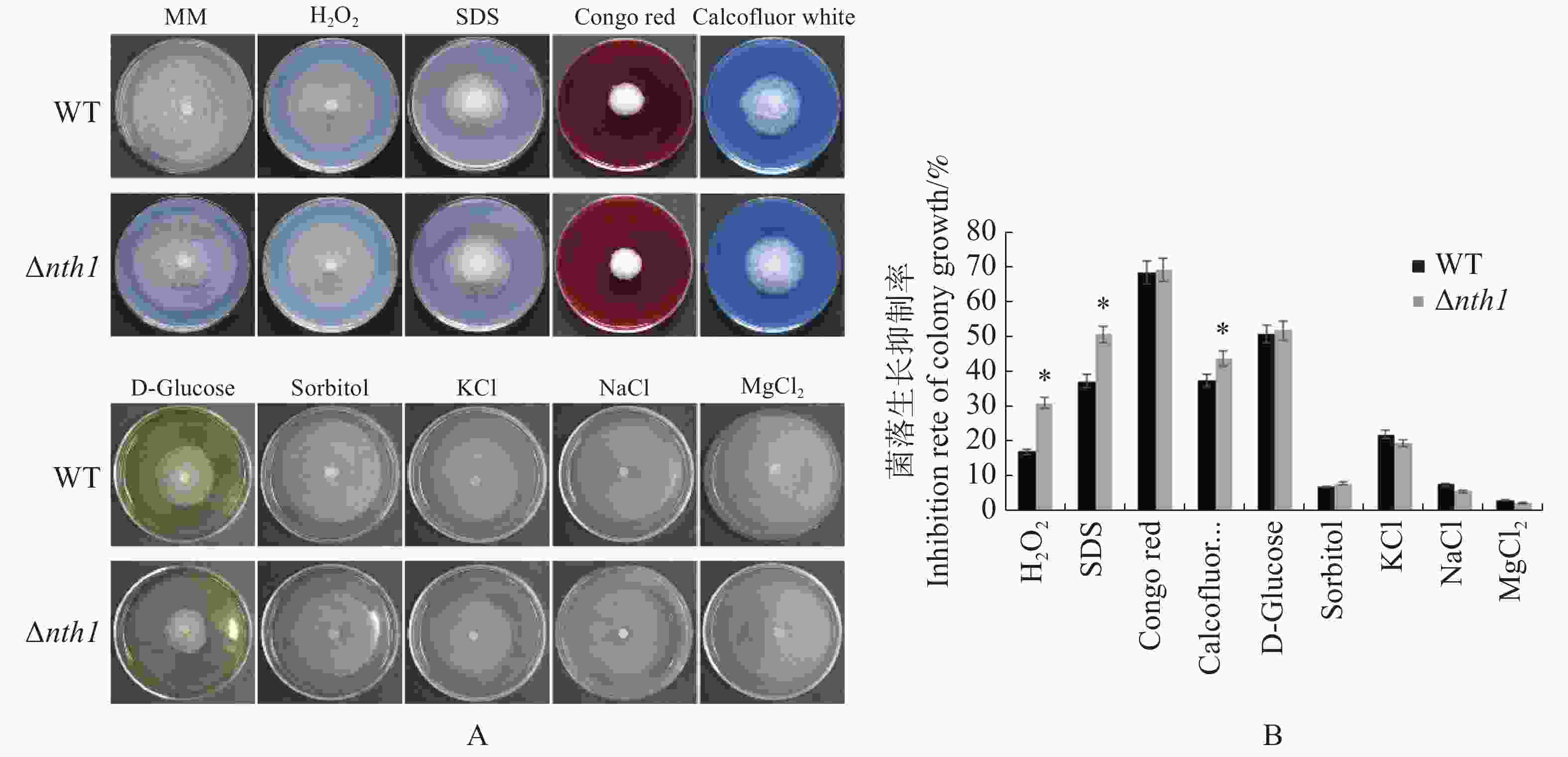
图 8 Δnth1对胁迫因子的敏感性测定
Figure 8. Determination of Δnth1 sensitivity to stress factors

图 9 Δnth1的高、低温胁迫敏感性分析
Figure 9. Sensitivity analysis of Δnth1 to high or low temperature stress
-
本研究通过Split-Marker同源重组技术获得香蕉枯萎病菌的中性海藻糖NTH1基因敲除突变体Δnth1,表型鉴定结果表明,与野生型菌株相比,Δnth1菌株对氰烯菌酯抗性没有明显差异,但NTH1基因的缺失导致香蕉枯萎病菌的致病力减弱,与FOSTER[36]“稻瘟病菌的该基因缺失会引起致病力下降”的研究结果一致。与野生型菌株相比,Δnth1菌株的生长速度和孢子萌发率均下降,这与DOEHLEMANN[23]“在灰葡萄孢中NTH1基因缺失导致孢子萌发率下降”的研究结果一致。与野生型菌株相比,Δnth1菌株对H2O2的氧化应激作用更明显,这与PEDREÑ[37]“酿酒酵母中NTH1基因缺失对H2O2引起的氧化应激响应较强,突变体的菌落生长受到更明显抑制”的研究结果一致。同时,Δnth1菌株在高糖、低温和高温条件下的生长,与野生型菌株相比没有明显差异,这与吕烨等[38]测试的酵母NTH1基因缺失突变株耐冷冻、耐高糖和耐高温能力优于野生型的结果不一致。编码海藻糖酶基因的数量会由于在不同菌中而存在差异,而发挥不同的功能,如在稻瘟病菌中,中性海藻糖基因的缺失使其定值于植物组织的能力降低,而致病力下降[33]。与之相反的是,在白色丝状酵母和隐球菌中,中性海藻糖酶突变体并没有明显的致病力差异,该基因并不是抗真菌药的潜在靶标[39-40]。尽管海藻糖酶是海藻糖唯一的水解酶,但由于影响真菌适应性的因素和其他酶的存在,使得不同真菌的表型存在差异。
综上所述,NTH1基因具有高度保守性,但由于在不同菌中,使其发挥了不同的功能。目前,国内关于植物病原真菌海藻糖酶基因的功能研究尚少。在香蕉枯萎病菌中,NTH1基因缺失突变体对氰烯菌酯的敏感性和野生型相比不存在明显差异,而对巴西蕉苗致病力显著下降。Δnth1菌株对细胞壁选择性压力和氧化压力敏感,但在高糖、高盐和高低温胁迫条件下与野生型菌株相比没有明显差异。因此,笔者推测该基因可能调控香蕉枯萎病菌的分生孢子萌发和生长,参与细胞壁合成和氧化应激反应,并且在致病力方面发挥重要作用,因为海藻糖的储存不是细胞防御高盐、高糖和高低温胁迫的唯一保护剂,NTH1在香蕉枯萎病菌中的具体调控机制还需进一步研究。
Knockout and Functional Analysis of Neutral Trehalase NTH1 Gene in Fusarium oxysporum f. sp. cubense Race 4
-
摘要: 为了研究中性海藻糖酶在香蕉枯萎病菌4号生理小种(Foc4-37)致病和耐受不良环境中的作用,笔者构建了中性海藻糖酶编码基因敲除突变体Δnth1,并对敲除突变体的致病力、生物学特性和对氰烯菌酯的抗药性开展测定分析。结果表明:与野生型相比,Δnth1菌落生长缓慢、孢子萌发率下降,对巴西蕉苗致病力明显减弱,但产孢量没有显著差异,对氰烯菌酯的敏感性(EC50=6.07 mg·L−1)也无明显变化。由此推断NTH1基因参与调控香蕉枯萎病菌的分生孢子萌发和生长,参与细胞壁合成和氧化应激反应的应答,不参与FOC对渗透压力、高糖和高低温胁迫的应答。Abstract: In order to study the role of neutral trehalase in the pathogenicity and intolerance environment of Fusarium oxysporum f. sp.cubense race 4 (Foc4-37), a neutral trehalase-encoding gene knockout mutant Δnth1 was constructed, and its pathogenicity, biological characteristics and resistance to phenamacril were determined. The results showed the neutral trehalase-encoding gene knockout mutant Δnth1 was lower in colony growth rate and conidia germination rate, and significantly lower in pathogenicity to banana (Cavendish, AAA) than the wild-type strain. However, there was no significant difference in conidia yield and sensitivity to phenamacril (EC50 = 6.07 mg·L−1) between the mutant and the wild type strain. It was inferred that NTH1 gene was involved in regulating the growth and conidia germination of Fusarium oxysporum f. sp., and responding to the cell wall synthesis and oxidative stress, but not to the osmotic pressure, high sugar stress and temperature stress.
-
Key words:
- Fusarium oxysporum f. sp. cubense /
- neutral trehalase /
- gene knockout /
- pathogenicity
-
图 2 Split-Marker重组技术的片段扩增
A. NTH1基因敲除原理示意图;B.两轮PCR的片段扩增:1~2.第1轮PCR扩增的NTH1基因上下游片段,条带大小分别为1 903,2 055 bp;5~6. 第2轮PCR扩增重叠片段,条带大小分别为2 379,2 138 bp;9.第1轮PCR扩增的潮霉素(HYG)抗性基因片段,条带大小为1 376 bp;3~4,7~8,10.水对照;M.DNA Marker DL 5000。
Fig. 2 Fragment amplification in Split-Marker recombination
A. Schematic diagram of NTH1 gene knockout; B. Fragment amplification of two rounds of PCR: 1−2. The first round of PCR amplification of the upstream and downstream fragments of NTH1 gene with their band sizes being 1 903 bp and 2 055 bp, respectively; 5-6. The second round of PCR amplification of overlapping fragments with their band sizes being 2 379 bp and 2 138 bp, respectively; 9. The first round of PCR amplification of hygromycin (HYG) resistance gene fragments, with its band size being 1 376 bp; 3-4, 7-8, 10. H2O was used as control; M. DNA Marker DL 5000.
图 3 NTH1敲除突变体的筛选验证
A. NTH1敲除突变体上下游重组片段的正筛选:1~2. NTH1基因敲除突变体重组片段的PCR扩增,条带分别为2 297,2 160 bp(3个NTH1敲除转化子,Δnth1-1,-2,-3),H2O和野生型为对照,M. DNA Marker DL 5000;B. NTH1敲除突变体HYG片段的正筛选:NTH1敲除突变体潮霉素片段的PCR扩增,条带大小为1 376 bp(Δnth1-1,-2,-3),H2O和野生型为对照,M. DNA Marker DL 5000;C. NTH1敲除突变体内源基因的负筛选:NTH1敲除突变体内源基因的PCR扩增(Δnth1-1,-2,-3),H2O和野生型为对照,条带大小为737 bp,M. DNA Marker DL 5000。
Fig. 3 Screening and validation of NTH1 knockout mutant
A. Positive screening of upstream and downstream recombinant fragments of the NTH1 knockout mutant: 1−2. PCR amplification of the NTH1 knockout mutant recombinant fragments, their band sizes 2 297 bp and 2 160 bp, (Three NTH1 knockout transformants, Δnth1-1, -2, -3), respectively, H2O and WT used as controls, M. DNA Marker DL 5 000; B. Positive screening of the NTH1 knockout mutant HYG fragment: PCR amplification of hygromycin fragment of the NTH1 knockout mutant, its band size 1 376 bp (Δnth1-1, -2, -3), H2O and WT used as controls, M. DNA Marker DL 5 000; C. Negative screening of endogenous genes from the NTH1 knockout mutant: PCR amplification of endogenous genes from the NTH1 knockout mutant (Δnth1-1, -2, -3), H2O and WT used as controls, its band size 737 bp, M. DNA Marker DL 5 000.
图 4 Δnth1的致病力测定
A:野生型和Δnth1侵染巴西蕉苗后的球茎发病症状,H2O为对照(45 d);B:巴西蕉的病情指数,H2O为对照(45 d);图中误差线代表3次生物学重复的标准误差,不同小写字母代表在P<0.05显著差异;WT:香蕉枯萎病菌4号生理小种野生型菌株37。下同。
Fig. 4 Pathogenicity test of Δnth1
A: Infection of banana corms inoculated by wild type strain WT and Δnth1, with H2O as control (45 days);B: The disease indexes of the banana with H2O as control (45 days); The error bars in the figure represent the standard errors of 3 biological replicates. Different lowercase letters represent significant differences at P <0.05; WT: F. oxysporum f. sp. cubense race 4 wild type strain 37. Similarly hereinafter.
图 5 野生型和Δnth1在含有不同浓度氰烯菌酯的PDA平板上的菌落生长情况
Fig. 5 Colony growth of WT and Δnth1 on PDA supplemented with phenamacril at different concentrations
图 6 野生型和Δnth1在不同氰烯菌酯浓度下的菌落生长抑制率
*代表在P<0.05水平下差异显著,下同。
Fig. 6 Inhibition rates of WT and Δnth1 under different concentrations of phenamacril
* represents significant differences at P <0.05, similarly hereinafter.
图 7 Δnth1的生物学表型分析
A:野生型和Δnth1在PDA上的菌落生长情况(2,4,6 d);B:野生型和Δnth1菌落生长直径对比(2,4,6 d);C:野生型和Δnth1的分生孢子萌发率对比(8,12,24 h)。
Fig. 7 Biological phenotypic analysis of Δnth1
A: The colony growth of WT and Δnth1 on PDA (2, 4, 6 d); B: Comparison of colony growth diameter between WT and Δnth1(2, 4, 6 d); C: Comparison of conidial germination rates between WT and Δnth1 (8, 12, 24 h).
图 8 Δnth1对胁迫因子的敏感性测定
A:野生型和Δnth1在不同胁迫因子下的菌落生长情况(生长6 d);B:野生型和Δnth1在不同胁迫因子下的受抑制率。
Fig. 8 Determination of Δnth1 sensitivity to stress factors
A: Colony growth of WT and Δnth1 under different stress factors (6 d); B: Inhibition rates of WT and Δnth1 under different stress factors.
图 9 Δnth1的高、低温胁迫敏感性分析
A:野生型和Δnth1在高、低温条件下的菌落生长情况(6 d);B:野生型和Δnth1在高、低温胁迫因子下的受抑制率。
Fig. 9 Sensitivity analysis of Δnth1 to high or low temperature stress
A: Colony growth of WT and Δnth1 under high or low temperature (6 d); B: Inhibition rates of WT and Δnth1 under the stress factor of high or low temperature.
表 1 本实验的引物序列
Table 1 Sequences of the primers used in the study
引物名称 Primer 序列 Sequence(5′→3′) HYG-F CTTGGCTGGAGCTAGTGGAGGT HYG-R CCCGGTCGGCATCTACTCTATTC HYG-R1 GGATGCCTCCGCTCGAAGTA HYG-F1 CGTTGCAAGACCTGCCTGAA HPT-LBCK GACAGACGTCGCGGTGAGTT HPT-RBCK TCTGGACCGATGGCTGTGTAG FOC4NTH1-LBCK TAAACCCAGCCACGACAC FOC4NTH1-HPH-LB-R ACCTCCACTAGCTCCAGCCAAGCCGTTGACTTACACCAGCAG FOC4NTH1-HPH-RB-F GAATAGAGTAGATGCCGACCGGGCTGACCTACCAGCACTCTCG FOC4NTH1-RBCK GGCAAGCCACTCATCTTCA FOC4NTH1-LB-F ACATTGAGCAACATCCGAGT FOC4NTH1- RB-R TGATGTCCTCCAAACTCACG FOC4NTH1-CKFP ACCTCTTCGTGGTCTGTCCT FOC4NTH1-CKRP ATCTGTTTCCGCCCATACTC FOC4NTH1-F CTCCTTCTCACCGCTCTATTA FOC4NTH1-R CGTATGTGTTCGGCTCAAA  下载: 导出CSV
下载: 导出CSV
-
[1] 钟群有, 郑卓辉, 彭增明, 等. 香蕉枯萎病生物防治研究概述[J]. 广东农业科学, 2007(7): 64 − 65. doi: 10.3969/j.issn.1004-874X.2007.07.022 [2] PLOETZ R C. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. Cubense [J]. Phytopathology, 2006, 96(6): 648. doi: 10.1094/PHYTO-96-0648 [3] NEL B, STEINBERG C, LABUSCHAGNE N, et al. The potential of nonpathogenic Fusarium oxysporum and other biological control organisms for suppressing fusarium wilt of banana [J]. Plant Pathology, 2006, 55(2): 217 − 223. doi: 10.1111/j.1365-3059.2006.01344.x [4] THANGAVELU R, PALANISWAMI A, VELAZHAHAN R. Mass production of Trichoderma harzianum for managing fusarium wilt of banana. Agriculture [J]. Ecosystems and Environment, 2004, 103(1): 259 − 263. doi: 10.1016/j.agee.2003.09.026 [5] 汪军, 王国芬, 杨腊英, 等. 施用淡紫拟青霉与套作对香蕉枯萎病控病作用的影响[J]. 果树学报, 2013, 30(5): 857 − 864. [6] 刘磊, 梁昌聪, 覃和业, 等. 香蕉枯萎病田间发病株的高效灭菌方法筛选[J]. 植物保护学报, 2015, 42(3): 362 − 369. [7] 孙雪丽, 郝向阳, 王天池, 等. 香蕉枯萎病防控和抗病育种研究进展[J]. 果树学报, 2018, 35(7): 870 − 879. [8] 蒲金基, 刘晓妹, 曾会才. 香蕉抗枯萎病育种研究进展[J]. 中国南方果树, 2003(1): 31 − 34. doi: 10.3969/j.issn.1007-1431.2003.01.020 [9] 齐兴柱, 杨腊英, 黄俊生. Foc4的2个过氧化氢酶基因的克隆与表达分析及其引起的香蕉苗活性氧迸发研究[J]. 中国农学通报, 2012, 28(15): 163 − 169. doi: 10.3969/j.issn.1000-6850.2012.15.032 [10] 王飞燕. 香蕉尖孢镰刀菌fpd1基因敲除与功能研究[D].海口: 海南大学, 2015. [11] DONG Z, WANG Z. Isolation and characterization of an exopolygalacturonase from Fusarium oxysporum f.sp. cubense race 1 and race 4 [J]. BMC Biochemistry, 2011, 12(1): 51. doi: 10.1186/1471-2091-12-51 [12] PIETRO A D, GARCIA-MACEIRA F I, MEGLECZ E, et al. A MAP kinase of the vascular wilt fungus Fusarium oxysporum is essential for root penetration and pathogenesis [J]. Molecular Microbiology, 2001, 39(5): 1140 − 1152. doi: 10.1111/j.1365-2958.2001.02307.x [13] DING Z J, LI M, SUN F, et al. Mitogen-activated protein kinases are associated with the regulation of physiological traits and virulence in Fusarium oxysporum f. sp. cubense [J]. Plos One, 2015, 10(4): e0122634. doi: 10.1371/journal.pone.0122634 [14] 杨平, 李敏惠, 潘克俭, 等. 海藻糖的生物合成与分解途经及其生物学功能[J]. 生命的化学, 2006(3): 233 − 236. doi: 10.3969/j.issn.1000-1336.2006.03.014 [15] GANCEDO C, FLORES C L. The importance of a functional trehalose biosynthetic pathway for the life of yeasts and fungi [J]. FEMS Yeast Research, 2004, 4(4-5): 351 − 359. doi: 10.1016/S1567-1356(03)00222-8 [16] 谢夏青, 李志勇, 马继芳, 等. 谷子弯孢菌海藻糖酶基因(ClTRE)及其启动子的克隆和序列分析[J]. 生物技术通报, 2011(7): 154 − 159. [17] WOLSKA-MITASZKO B, MOLESTAK E. Properties of trehalase from different organs of alfalfa, Medicago sativa [J]. Acta Physiologiae Plantarum, 2005, 27(1): 53 − 60. doi: 10.1007/s11738-005-0036-0 [18] 任媛媛, 刘景芳, 戴秀玉, 等. 海藻糖代谢途径相关基因及生物工程[J]. 微生物学报, 2003(6): 821 − 825. doi: 10.3321/j.issn:0001-6209.2003.06.024 [19] MITTENBÜHLER K, HOLZER H. Purification and characterization of acid trehalase from the yeast suc2 mutant [J]. The Journal of biological chemistry, 1988, 263(17): 8537 − 8543. [20] KOPP M, MÜLLER H, HOLZER H. Molecular analysis of the neutral trehalase gene from Saccharomyces cerevisiae [J]. The Journal of biological chemistry, 1993, 268(7): 4766 − 4774. [21] RUIJTER G, FILLINGER S, THEVELEIN J, et al. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans [J]. Microbiology, 2001, 147(7): 1851 − 1862. doi: 10.1099/00221287-147-7-1851 [22] AURORA OCÓN, RÜDIGER HAMPP, REQUENA N. Trehalose turnover during abiotic stress in arbuscular mycorrhizal fungi [J]. New Phytologist, 2007, 174(4): 879 − 891. doi: 10.1111/j.1469-8137.2007.02048.x [23] DOEHLEMANN G. Trehalose metabolism is important for heat stress tolerance and spore germination of Botrytis cinerea [J]. Microbiology, 2006, 152(9): 2625 − 2634. doi: 10.1099/mic.0.29044-0 [24] GARRE E, MATALLANA E. The three trehalases Nth1p, Nth2p and Ath1p participate in the mobilization of intracellular trehalose required for recovery from saline stress in Saccharomyces cerevisiae [J]. Microbiology, 2009, 155(Pt 9): 3092 − 3099. [25] 郎玉成, 倪珏萍. 新型杀菌剂——氰烯菌酯(JS399-19)[J]. 世界农药, 2007(5): 52 − 53. doi: 10.3969/j.issn.1009-6485.2007.05.011 [26] 李恒奎, 周明国, 王建新, 等. 氰烯菌酯防治小麦赤霉病及治理多菌灵抗药性研究[J]. 农药, 2006(2): 92 − 94. doi: 10.3969/j.issn.1006-0413.2006.02.005 [27] 王龙根, 倪珏萍, 王凤云, 等. 新杀菌剂JS399-19的生物活性研究[J]. 农药, 2004(8): 380 − 383. doi: 10.3969/j.issn.1006-0413.2004.08.016 [28] 迟梦宇, 王健力, 黄金光. 禾谷镰刀菌肌球蛋白(MyosinⅠ)生物信息学分析及原核表达与纯化[J]. 青岛农业大学学报:自然科学版, 2018, 35(2): 111 − 118. [29] ZHENG Z, HOU Y, CAI Y, et al. Whole-genome sequencing reveals that mutations in myosin-5 confer resistance to the fungicide phenamacril in Fusarium graminearum [J]. Scientific Reports, 2015, 5: 8248. doi: 10.1038/srep08248 [30] 王飞燕, 郭立佳, 杨腊英, 等. 尖孢镰刀菌古巴专化型4号生理小种fpd1基因敲除与表型分析[J]. 热带作物学报, 2015, 36(8): 1462 − 1468. doi: 10.3969/j.issn.1000-2561.2015.08.017 [31] 徐齐君, 胡小平, 陈婷, 等. PEG介导的棉花枯萎病菌原生质体转化体系的建立[J]. 棉花学报, 2012, 24(3): 222 − 228. doi: 10.3969/j.issn.1002-7807.2012.03.005 [32] 漆艳香, 谢艺贤, 张欣, 等. 香蕉枯萎菌基因组DNA提取方法的研究[J]. 生物技术, 2004(6): 32 − 34. doi: 10.3969/j.issn.1004-311X.2004.06.016 [33] 杨军, 薛芳, 王海凤, 等. 水稻稻瘟病菌单孢分离技术及常见问题分析[J]. 山东农业科学, 2017, 49(2): 132 − 135. [34] MOHAMED A A, MAK C, LIEW K W, et al. Early evaluation of banana plants at nursery stage for fusarium wilts tolerance[C]// Molina A B, Masdek N H, Liew K W. Banana fusarium wilt management: towards sustainable cultivation. Rome: INIBAP Press, 2001: 174 − 185. [35] FENG H Q, LI G H, SHUN-WEN D, et al. The septin protein Sep4 facilitates host infection by plant fungal pathogens via mediating initiation of infection structure formation [J]. Environmental Microbiology, 2016, 19(5): 1730 − 1749. [36] FOSTER A J. Trehalose synthesis and metabolism are required at different stages of plant infection by Magnaporthe grisea [J]. EMBO (European Molecular Biology Organization) Journal, 2003, 22(2): 225 − 235. doi: 10.1093/emboj/cdg018 [37] PEDREÑ O Y, GIMENO-ALCAÑIZ J V, MATALLANA E, et al. Response to oxidative stress caused by H2O2 in Saccharomyces cerevisiae mutants deficient in trehalase genes [J]. Archives of Microbiology, 2002, 177(6): 494 − 499. doi: 10.1007/s00203-002-0418-2 [38] 吕烨, 肖冬光, 和东芹, 等. 酵母海藻糖酶缺失突株的构建及其耐性[J]. 微生物学报, 2008(10): 1301 − 1307. doi: 10.3321/j.issn:0001-6209.2008.10.005 [39] PENDREÑO Y, GONZÁLEZ‐PÁRRAGA P, CONESA S,, et al. The cellular resistance against oxidative stress (H2O2) is independent of neutral trehalase (Ntc1p) activity in Candda albicans [J]. FEMS Yeast Research, 2006, 6(1): 57 − 62. doi: 10.1111/j.1567-1364.2005.00025.x [40] BOTTS M R, HUANG M, BORCHARDT R K, et al. Developmental cell fate and virulence are linked to trehalose homeostasis in Cryptococcus neoformans [J]. Eukaryotic Cell, 2014, 13(9): 1158 − 1168. doi: 10.1128/EC.00152-14 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 525
- HTML全文浏览量: 225
- PDF下载量: 237
- 被引次数: 0
