

-
环二鸟苷酸(c-di-GMP)是细菌中普遍存在的第二信使分子,首次在葡糖酸醋酸杆菌中被发现,是纤维素合酶的异构激活因子[1-2]。其后的研究结果表明,c-di-GMP参与调节各种生物过程如致病性、运动性、生物被膜、胞外多糖及胞外酶等[3-4]。胞内c-di-GMP水平受含有GGDEF、EAL或HD-GYP结构域蛋白的调控:GGDEF蛋白负责c-di-GMP合成,而EAL或HD-GYP蛋白参与c-di-GMP的分解[5]。GGDEF结构域含有约180个氨基酸残基,具有保守的Gly-Gly-Asp-Glu- Phe氨基酸残基序列[6]。在新月形杆菌(Caulobacter crescentus)中,GGDEF结构域蛋白PleD具有双鸟苷酸环化酶活性,调控细菌的运动功能,促进细胞从运动转向静止[7-9]。EAL结构域长约250个氨基酸,Glu–Ala–Leu序列高度保守,可调控细菌生物被膜的形成和运动性[10]。有时GGDEF和EAL结构域共存于同一个蛋白中,可能只有其中1个结构域有功能。例如,铜绿假单胞菌BifA含有GGDEF和EAL结构域,但其没有c-di-GMP合成却有分解活性[5]。稻黄单胞菌水稻致病变种(Xanthomonas oryzae pv. oryzae, Xoo)是引起水稻白叶枯病的病原菌[11],可对水稻造成严重的产量损失。C-di-GMP代谢相关蛋白同样参与Xoo致病力的调控,如:① GGDEF结构域蛋白GdpX1具有合成c-di-GMP的活性,调节Xoo的运动能力、胞外多糖的含量以及致病力[12];② EAL结构域蛋白PXO_03877具有降解c-di-GMP的活性,增强细菌的致病性、运动性、胞外多糖合成以及生物被膜的形成能力[13];③ GGDEF/EAL结构域蛋白PdeR与组氨酸激酶PdeK组成双组份系统,磷酸化的PdeR具有分解c-di-GMP的能力,pdeR突变降低Xoo的致病性、胞外多糖及生物被膜的含量,但对运动性和胞外酶无明显影响[14]。本研究分析了Xoo中另一个GGDEF/EAL结构域蛋白CrxV对运动性、生物被膜、胞外多糖以及胞外酶等毒力因子的调控作用,为进一步分析c-di-GMP在Xoo与水稻互作中的作用奠定基础。
-
Xoo野生型菌株PXO99A、大肠杆菌DH5a、基因敲除载体pK18mobSacB、过表达载体菌株pHM1等均由笔者所在的实验室保存;一般化学药品购自广州化学试剂公司;PCR试剂购自诺唯赞生物科技有限公司;内切酶和连接酶购自NEB生物科技有限公司;快速质粒小提试剂盒、琼脂凝胶DNA回收试剂盒等购自天根生物科技有限公司;IR24水稻用于检测Xoo的致病力;用SMART软件(
http://smart.embl-heidelberg.de/ )分析蛋白保守结构域。 -
将crxV基因上下游各约500 bp的序列通过PCR扩增、酶切(HindIII/XbaI与XbaI/EcoRI)回收后连接到自杀性载体pK18mobSacB(HindIII/EcoRI酶切),获得敲除载体pK-crxV,电击转化PXO99A菌株,采用同源重组2次交换的方法构建突变体ΔcrxV[15-16]。利用PCR方法扩增crxV基因片段,酶切(HindIII/EcoRI)回收后连接互补载体pHM1(HindIII/EcoRI酶切),然后转化ΔcrxV获得互补菌株C-ΔcrxV(引物序列见表1)。为便于比较,野生型菌株和ΔcrxV也转入空载体pHM1。采用剪叶法检测致病力,将突变体菌株ΔcrxV-pHM1、野生型菌株PXO99A-pHM1和互补菌株C-ΔcrxV接种至生长期为60 d的水稻叶片上,14 d后统计水稻的发病情况。
表 1 引物序列
Table 1. Primers used in this study
引物 Primer 序列 Sequences 用途 Usages CrxVFF GCACAAGCTTTCGCAGTTCGTGCTGGCAGT crxV deletion CrxVFR GCACTCTAGAATGCAGGCGCATCGACCTCA CrxVRF GCACTCTAGATGCATGCGCCAACTCGGCAT CrxVRR GCACAATTCCAAGGCCTTCAAGAGCTACG CrxVhmF TATGACCATGATTACGCCAAGCTTGATGCGCCTGCATACCCGCATC ΔcrxV complement CrxVhmR TGTAAAACGACGGCCAGTGAATTCATCAACTGTGCCAGGAACTG 注:“—”表示酶切位点
Note: restriction enzyme sites are underlined -
参照文献[17]的方法检测胞外酶的活性,将突变体菌株ΔcrxV-pHM1、野生型菌株PXO99A-pHM1和互补菌株C-ΔcrxV置28 ℃条件下液体培养48 h后,取1 mL培养物用ddH2O洗涤2遍,用1 mL ddH2O重悬菌体。取3 µL重悬液分别接种至纤维素检测培养基、淀粉酶检测培养基和蛋白酶检测培养基上,在28 ℃条件下静置培养5 d。PGA培养基:1 g·mL−1 Tryptone、1 g·mL−1葡萄糖、0.1 g·mL−1 L-谷氨酸钠、1.5% agar;纤维素检测培养基为PGA培养基+0.5%羧甲基纤维素;淀粉酶检测培养基为PGA培养基+0.1%可溶性淀粉;蛋白酶检测培养基为PGA培养基+2%脱脂奶粉。纤维素酶活性测定方法: 0.1%刚果红染色30 min,用1mol·L−1的NaCl溶液洗2次,观察酶水解直径(cm);淀粉酶活性测定方法:1:100 I2/KI (0.08% mol·L−1 I2,3.2 mol·L−1 KI)溶液染色10 min,观察酶水解直径(cm);蛋白酶活性测定方法:直接在蛋白培养基上观察水解光圈(cm)。每个试验重复3次。
-
参照文献[18]的方法检测游动性。用牙签蘸取1.3节中的菌重悬液,垂直接种于含0.25%琼脂的游动性培养基(0.03%Tryptone, 0.03%Yeast-extract, 0.25%agar)底部,在28 ℃条件下静置培养4 d,测量菌株在培养基表面游动的直径(cm)并统计。重复3次。
-
参照文献[19-20]的方法检测生物被膜的形成及胞外多糖的产生。取单克隆细菌在PSA中培养48 h后,取2 mL菌在5 500 r·min−1条件下离心3 min,收集菌体,用M210培养基(5 g·L−1蔗糖; 8 g·L−1酶水解酪素; 4 g·L−1 Yeast-extract; 3 g·L−1 K2HPO4;0.3 g·L−1 MgSO4 · 7H2O,pH7.0)洗涤2次,再加入M210重悬,把重悬液的OD600调至0.5。生物被膜实验:取2 mL重悬液(OD600=0.5)接种至玻璃试管,在28 ℃下静置培养4 d,缓慢倒出玻璃试管底的培养基,用H2O洗涤3遍,0.1%结晶紫染色液染色30 min,再用H2O洗涤3遍,加入3 mL90%乙醇溶解,在OD590下测吸光值。胞外多糖实验:取2 µL重悬液(OD600=0.5)接种至PGA培养基表面,在28 ℃下静置培养4 d,观察菌落表面形态。试验重复3次。
-
根据SMART预测结果,CrxV含有GGDEF、EAL、HAMP结构域(图1)。GGDEF和EAL可能参与c-di-GMP的合成与分解,而HAMP可能通过调控CrxV的二聚体化影响GGDEF和EAL的活性,因此,CrxV可能参与c-di-GMP的代谢,调控细菌对外界环境的响应能力。

图 1 CrxV的保守结构域
Figure 1. The conserved domains of CrxV
-
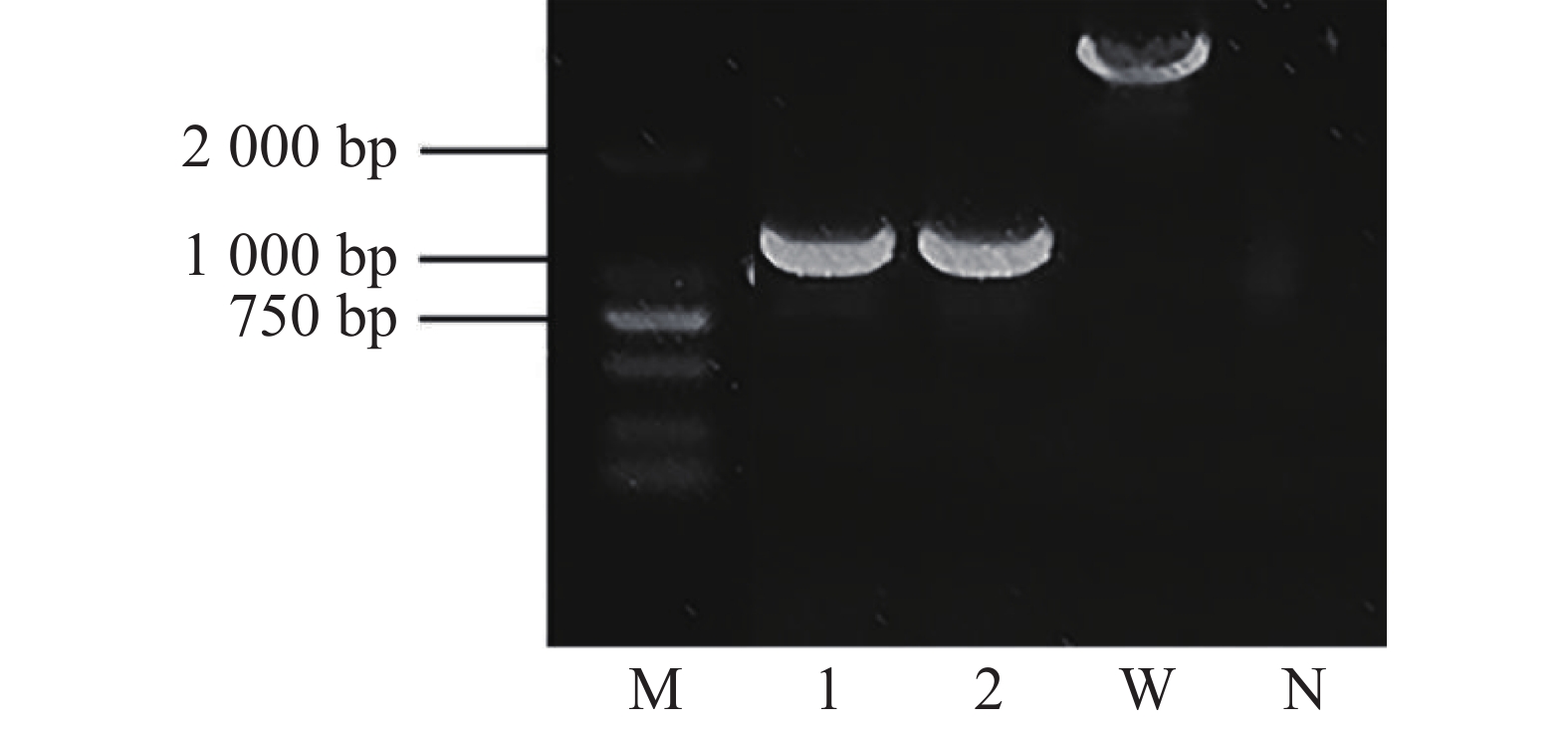
为了分析crxV的功能,本实验采用同源重组2次交换法构建了crxV的缺失突变体(图2)。结果显示:crxV突变体基因组扩增产物约为1 000 bp,而野生型PXO99A基因组扩增片段比突变体长约1 500 bp(与crxV基因大小一致),说明crxV突变体构建成功。
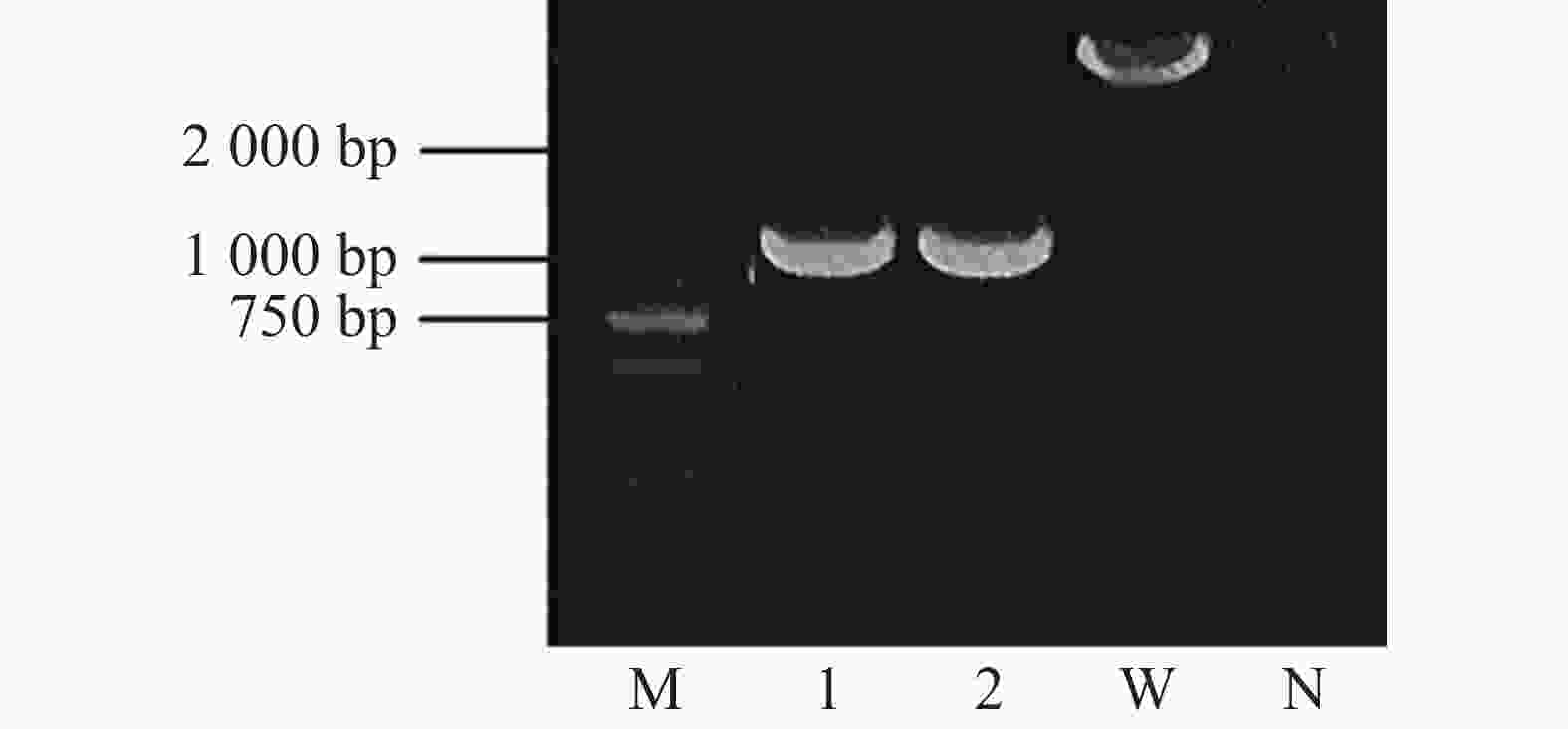
图 2 PCR 验证 crxV 突变体(ΔcrxV)
Figure 2. PCR confirmation of the crxV mutant
-
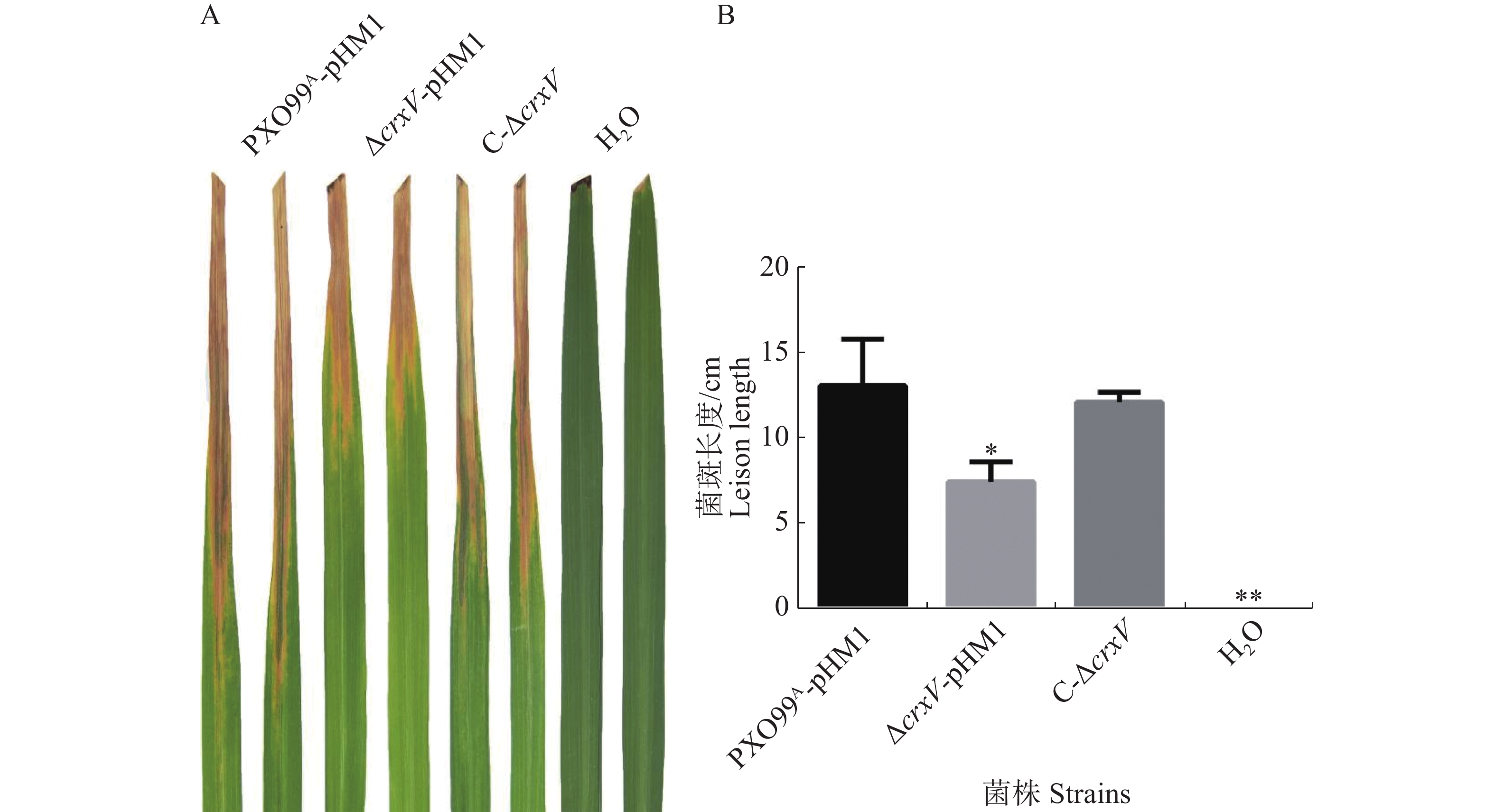
为了分析基因对Xoo的致病性,本实验构建了ΔcrxV的互补菌株C-ΔcrxV。致病性分析结果发现,ΔcrxV的病斑长度比野生型PXO99A的病斑长度减短了3 cm左右,互补菌株的致病力与野生型基本一致(图3)。说明CrxV是Xoo侵染所必需的。
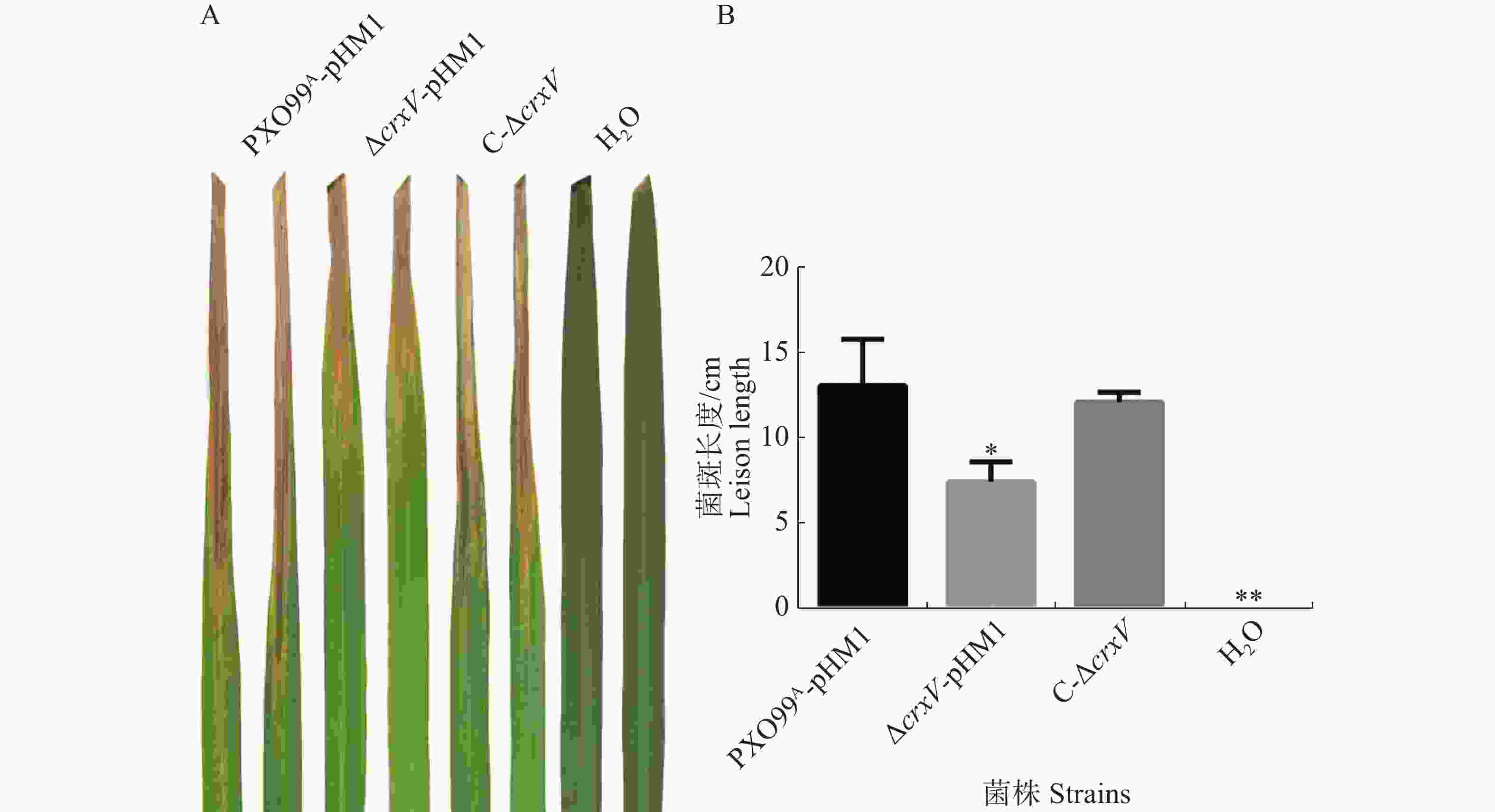
图 3 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV的致病性分析
Figure 3. Virulence analysis of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
-
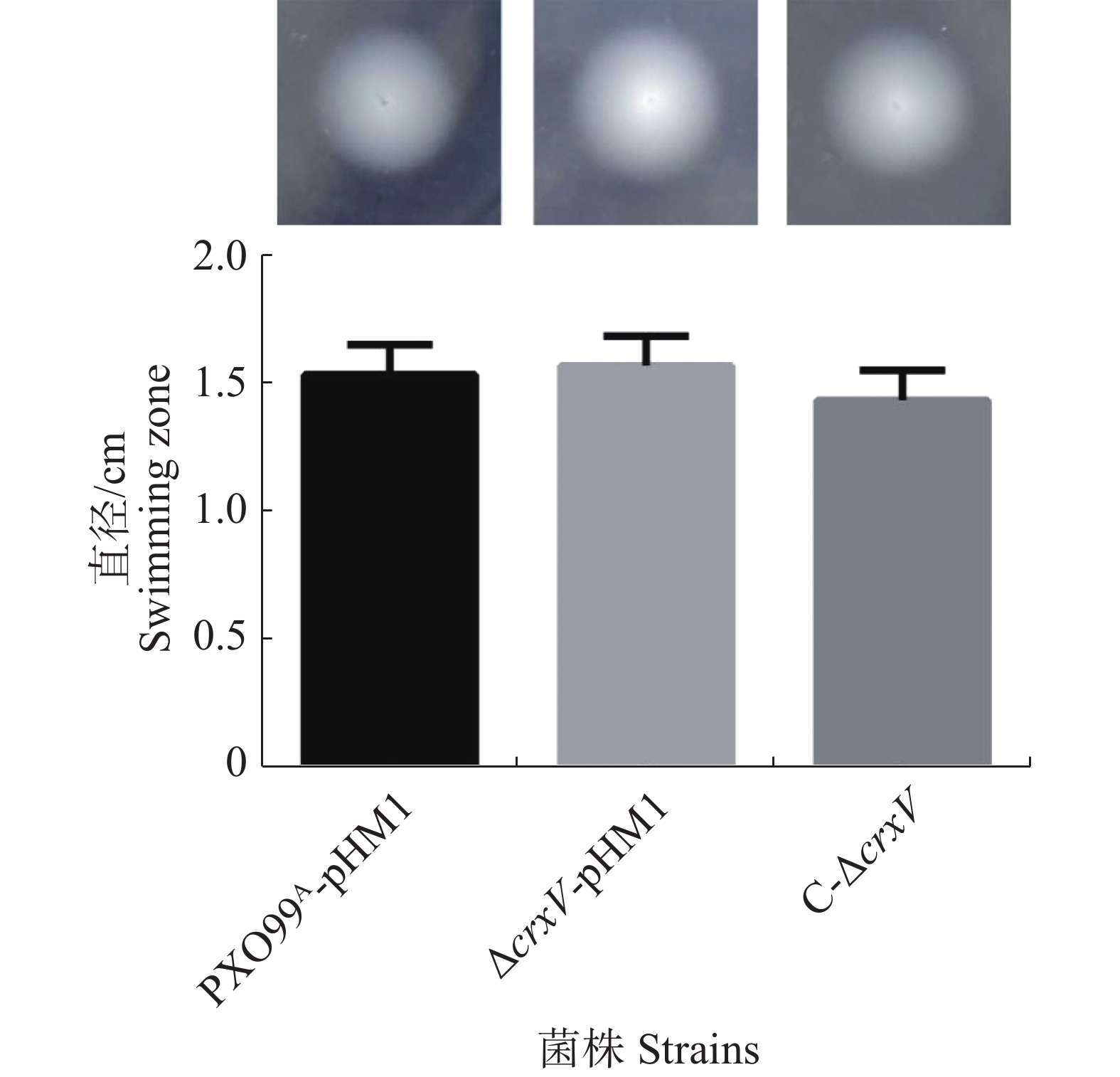
运动性是Xoo的重要毒力因子之一,且受c-di-GMP调控[21],为此有必要分析crxV突变是否影响Xoo的运动能力。PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV的游动能力分析结果(图4)表明,这3个菌株之间泳动能力没有显著差异,说明CrxV不调控Xoo的运动性。
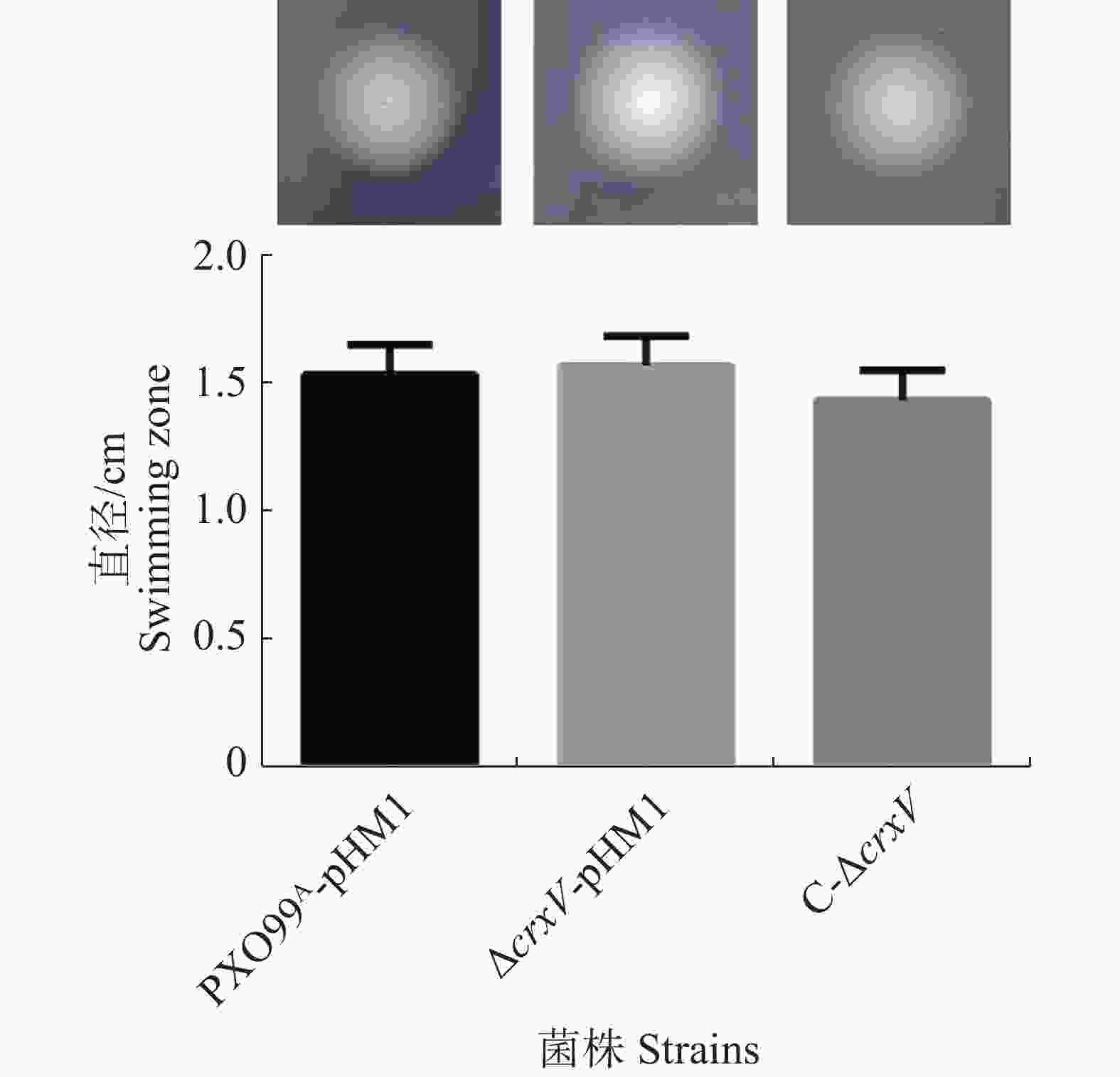
图 4 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV的游动能力
Figure 4. The swimming abilities of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
-
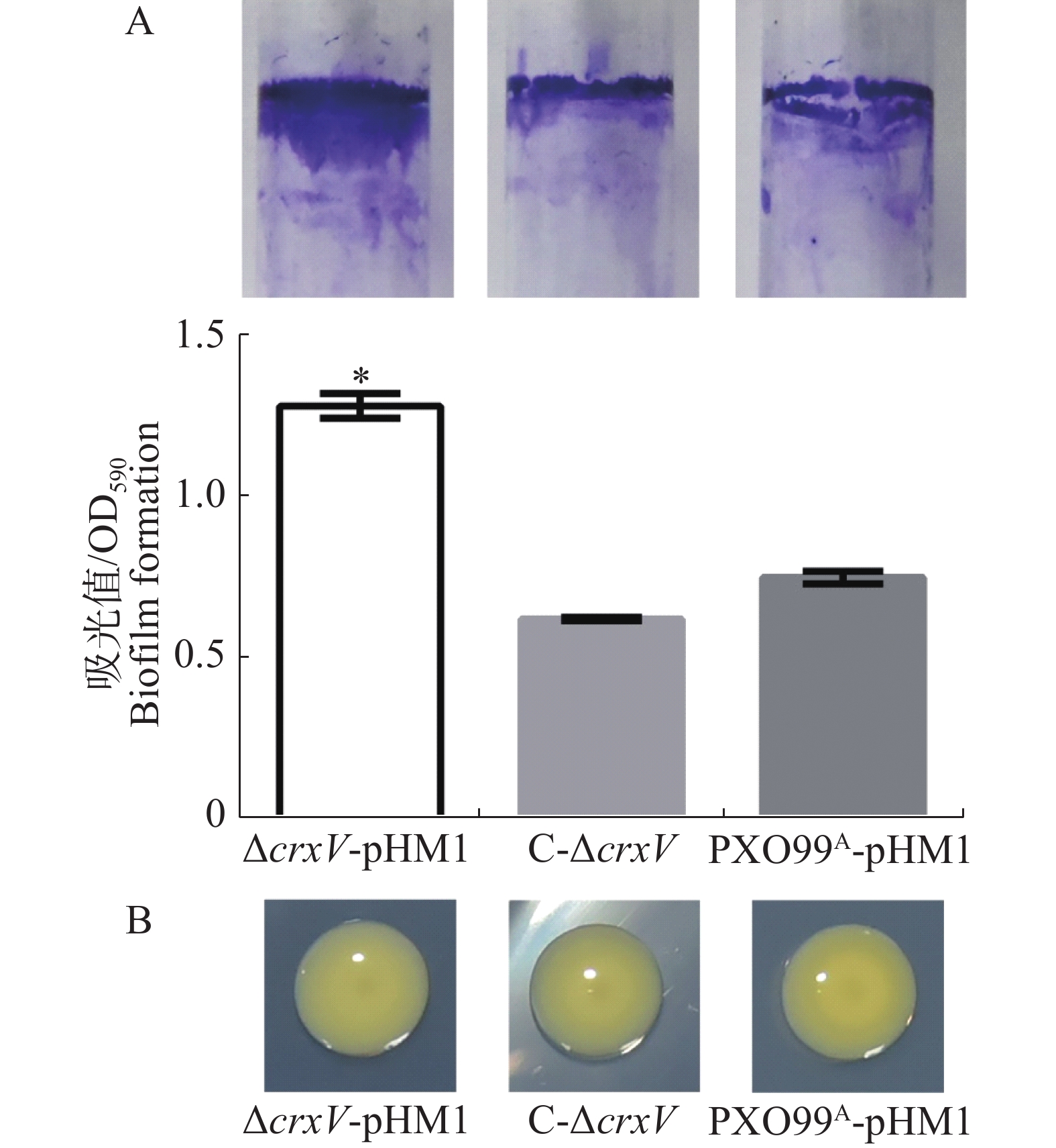
胞外多糖和生物被膜是Xoo的毒力因子,与细菌的致病性密切相关[14]。为分析胞外多糖的产生和生物被膜的形成,本实验将菌株接种至PGA固体培养基表面和M210液体培养基中,静置培养4 d后发现,PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV菌落表面无明显差别,而ΔcrxV-pHM1的生物被膜形成的量比PXO99A-pHM1高出了2倍,回补菌株与野生型菌株的生物被膜形成量接近一样(图5),说明CrxV负调控Xoo生物被膜的形成,但可能不调控细菌胞外多糖的产生。
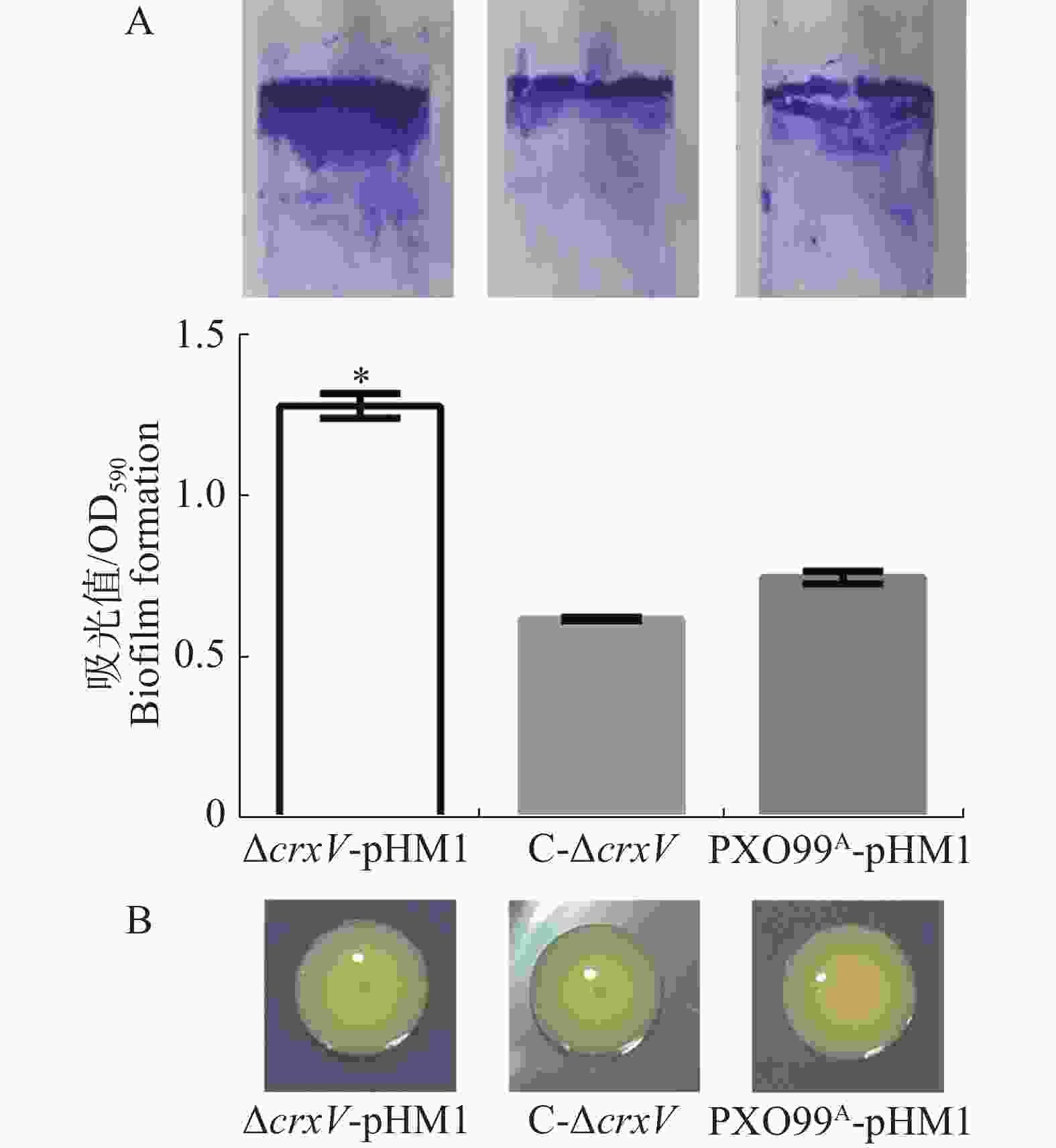
图 5 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV产生生物被膜(A)及胞外多糖(B)的能力
Figure 5. The biofilm (A) and extracellular polysaccharide (B) contents of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
-
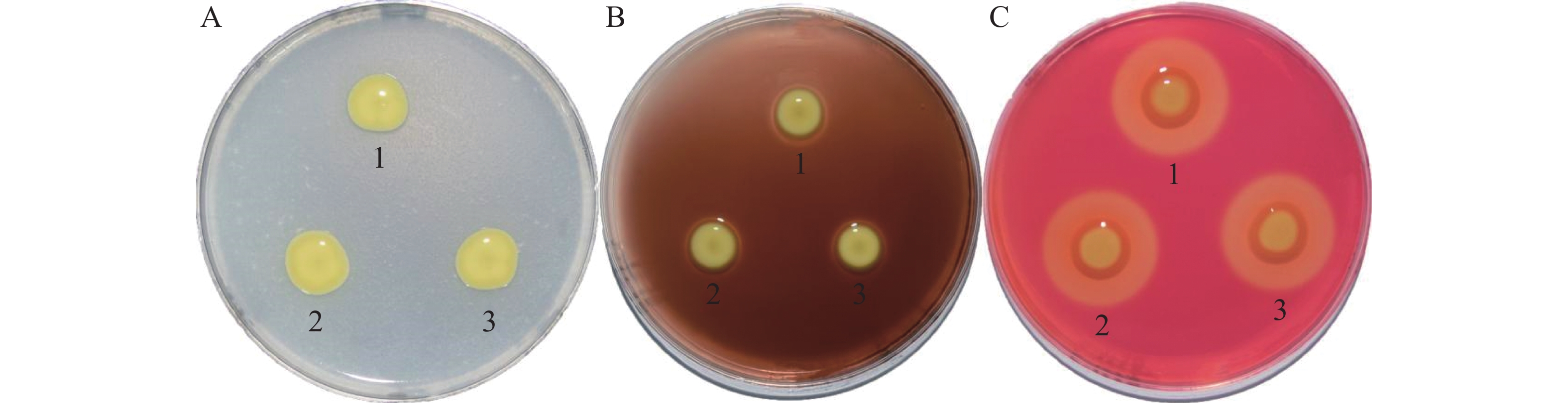
胞外酶通过Ⅱ型分泌系统将毒力因子分泌至胞外,促进细菌对寄主的致病力,胞外酶合成或分泌可能受c-di-GMP调控[22-23]。如图6所示,PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV菌株间的胞外纤维素酶、淀粉酶和蛋白酶活性均没有显著差别,说明CrxV不调控Xoo胞外酶的合成或分泌。
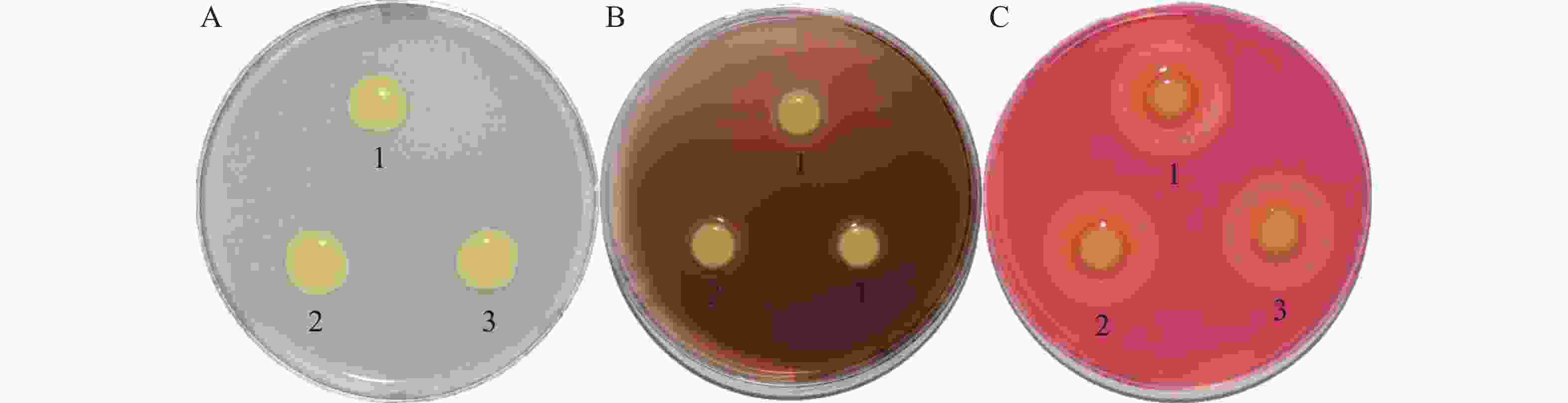
图 6 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV胞外酶活性分析
Figure 6. The extracellular enzyme activities of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
-
C-di-GMP分别由GGDEF结构域蛋白负责合成和EAL/HD-GYP结构域蛋白负责降解,胞内受体通过感知c-di-GMP水平调控细胞多个代谢途径,影响细菌的生长及环境适应能力。通常低水平的c-di-GMP提高细菌的运动能力,高水平的c-di-GMP则有利于生物被膜的形成,调节细菌的致病能力[4]。在Xoo中, GGEDF结构域蛋白GdpX1,通过负调控胞外多糖的产生和运动性从而抑制Xoo的致病性[12]。EAL结构域蛋白PXO_03877具有c-di-GMP水解酶活性,正调控致病性、生物膜的形成和胞外多糖的产生[13]。PdeR含有保守的GGDEF和EAL结构域,作为响应调节因子与组氨酸激酶PdeK组成双组份系统,正调控Xoo致病性和胞外多糖的产生[24]。同PdeR类似,CrxV也含有保守的GGDEF和EAL结构域,但其具体工作还不清楚。由于致病性与c-di-GMP的含量可能成负相关[13,24], crxV突变导致Xoo致病力降低,因此CrxV可能作为c-di-GMP分解酶,其GGDEF结构域可能没有酶活性。虽然研究发现Xoo中影响致病性的c-di-GMP代谢酶都可能调控诸如运动性、胞外多糖和胞外酶等毒力因子的合成或分泌[13,24],但crxV突变并不影响这些因子的形成或分泌。有趣的是,crxV突变导致Xoo生物被膜含量显著增加。生物被膜形成和解聚是细菌致病重要的调节过程,缺一不可。因此,CrxV可能负调控生物被膜的解聚,进而正调控病原菌的致病力。以上结果虽然显示CrxV可能通过介导c-di-GMP代谢,影响病原菌生物被膜的含量,调控病原菌的致病力,但CrxV的酶活性及其在生物被膜形成与解聚中的作用还不清楚,尚需做进一步研究。
CrxV Regulates the Virulence of Xanthomonas oryzae pv. oryzae
-
摘要: 为分析GGDEF和EAL结构域的蛋白CrxV对水稻黄单胞菌(Xanthomonas oryzaepv.oryzae,Xoo)致病性及相关致病因子的影响,通过同源重组法构建了crxV的突变体及其互补菌株;采用剪叶法比较了突变体、野生型和互补菌株对水稻叶片的侵染能力;分析了这3个菌株产生相关致病因子(运动性、胞外多糖、生物被膜和胞外酶)的能力。结果显示,crxV突变体的致病力比野生型和互补菌株的低。crxV突变导致病原菌生物被膜含量显著升高,但对运动性、胞外多糖和胞外酶均无明显影响。结果表明,CrxV可能通过负调控生物被膜影响Xoo的致病性。Abstract: Proteins containing GGDEF and EAL domains are involved in the synthesis and degradation of the cell´s second messenger c-di-GMP. They are important regulators for bacteria to respond to the external environment, and may be involved in the interaction between pathogen and host. We analyzed the effect of CrxV containing a GGDEF and an EAL domain on the virulence and some related virulent factors of Xanthomonas oryzae pv. oryzae (Xoo), a causal agent of bacterial leaf blight of rice. We used the homology recombination method to construct crxV mutant (ΔcrxV), and then the leaf clipping method to analyze the infection ability of the wild-type strain, ΔcrxV, and its complementary strain (C-ΔcrxV) on rice leaves. We also compared the motility, the contents of biofilm, extracellular polysaccharides and extracellular enzymes of these strains. The results showed that the ΔcrxV mutant had lower virulence than the wild-type and complementary strains. Compared with the wild-type and complementary strains, the ΔcrxV mutant was significantly higher in biofilm content, but no significant difference was observed in its motility, extracellular polysaccharides and extracellular enzymes. These results demonstrate that the CrxV may positively regulate the virulence of Xoo through negative regulation of biofilm formation.
-
Key words:
- CrxV /
- C-di-GMP /
- biofilm /
- virulence /
- Xanthomonas oryzae pv. oryzae
-
图 2 PCR 验证 crxV 突变体(ΔcrxV)
M:DL2000 Marker;1、2:ΔcrxV;W:野生型 PXO99A;N:阴性对照
Fig. 2 PCR confirmation of the crxV mutant
M: DL2000 Marker; lanes 1 and 2: ΔcrxV;W: wild-type PXO99A; N: negative control.
图 3 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV的致病性分析
A:接种水稻14 d后,野生型菌株PXO99A-pHM1、突变体菌株ΔcrxV-pHM1和互补菌株C-ΔcrxV的病斑特征,H2O为空白对照。B:接种14 d后的水稻病斑长度/cm (*: P<0.05, **: P<0.01)。
Fig. 3 Virulence analysis of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
A: The lesions caused by PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV (H2O: mock control). B: The lesion lengths (cm) of the infected IR24 leaves after 14 days’ inoculation of the indicated Xoo strains (*: P<0.05, **: P<0.01, compared withPXO99A-pHM1).
图 4 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV的游动能力
上图:细菌在平板上的游动表型;下图:游动直径的统计分析。
Fig. 4 The swimming abilities of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
Up panel: swimming phenotypes of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV; Down panel: the diameters of swimming zones of the indicated strains. All the experiments were repeated triple times. Values are given as means ± SD.
图 5 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV产生生物被膜(A)及胞外多糖(B)的能力
A:液体静置培养4 d后生物被膜含量分析。上图:气液表面生物被膜的结晶紫染色;下图:生物膜含量的定量分析(*: P<0.05)。B: PGA固体培养基上菌落的生长情况及表型特征。
Fig. 5 The biofilm (A) and extracellular polysaccharide (B) contents of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
A: The biofilm contents of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV 4 days after incubation (*: p<0.05). Up panel: the crystal violet staining biofilm synthesized in the liquid-gas interfaces; Down panel: the biofilm contents of the indicated strains (OD590). All the experiments were repeated triple times. Values are given as means ± SD. *: P<0.05, compared withPXO99A-pHM1. B: The colony phenotypes (4 days after incubation) of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV.
图 6 PXO99A-pHM1、ΔcrxV-pHM1和C-ΔcrxV胞外酶活性分析
A:蛋白酶活性;B:淀粉酶活性;C:纤维素酶活性。1:PXO99A-pHM1;2:ΔcrxV-pHM1;3:C-ΔcrxV。
Fig. 6 The extracellular enzyme activities of PXO99A-pHM1, ΔcrxV-pHM1 and C-ΔcrxV
A: protease activities; B: amylase activities; C: cellulase activities.1:PXO99A-pHM1;2:ΔcrxV-pHM1;3:C-ΔcrxV. All the experiments were repeated triple times with same results.
表 1 引物序列
Table 1 Primers used in this study
引物 Primer 序列 Sequences 用途 Usages CrxVFF GCACAAGCTTTCGCAGTTCGTGCTGGCAGT crxV deletion CrxVFR GCACTCTAGAATGCAGGCGCATCGACCTCA CrxVRF GCACTCTAGATGCATGCGCCAACTCGGCAT CrxVRR GCACAATTCCAAGGCCTTCAAGAGCTACG CrxVhmF TATGACCATGATTACGCCAAGCTTGATGCGCCTGCATACCCGCATC ΔcrxV complement CrxVhmR TGTAAAACGACGGCCAGTGAATTCATCAACTGTGCCAGGAACTG 注:“—”表示酶切位点
Note: restriction enzyme sites are underlined 下载: 导出CSV
下载: 导出CSV
-
[1] HEENGGE R. Principles of c-di-GMP signalling in bacteria [J]. Nature Reviews Microbiology, 2009, 7(4): 263 − 273. doi: 10.1038/nrmicro2109 [2] ROSS P, WEINHOUSE H, ALONI Y, et al. Regulation of cellulose synthesis in Acetobacter xylinumby cyclic diguanylic acid [J]. Nature, 1987, 325(6101): 279 − 281. doi: 10.1038/325279a0 [3] HENGGE R. Novel tricks played by the second messenger c-di-GMP in bacterial biofilm formation [J]. The EMBO Journal, 2013, 32(3): 322 − 323. doi: 10.1038/emboj.2012.351 [4] JENAL U, REINDERS A, LORI C. Cyclic di-GMP: second messenger extraordinaire [J]. Nature Reviews Microbiology, 2017, 15(5): 271 − 284. doi: 10.1038/nrmicro.2016.190 [5] OCHSNER U A, WILDERMAN P J, VASIL A I, et al. GeneChip® expression analysis of the iron starvation response in Pseudomonas aeruginosa: identification of novel pyoverdine biosynthesis genes [J]. Molecular Microbiology, 2002, 45(5): 1277 − 1287. doi: 10.1046/j.1365-2958.2002.03084.x [6] KARAOLIS D. K, RASHID M H, CHYTHANYA R., et al. c-di-GMP (3'-5'-cyclic diguanylic acid) inhibits Staphylococcus aureus cell-cell interactions and biofilm formation [J]. Antimicrob Agents Chemother, 2005, 49(3): 1029 − 1038. doi: 10.1128/AAC.49.3.1029-1038.2005 [7] ALDRIDGE P, JENAL U. Cell cycle-dependent degradation of a flagellar motor component requires a novel-type response regulator [J]. Molecular Microbiology, 1999, 32(2): 379 − 391. doi: 10.1046/j.1365-2958.1999.01358.x [8] HECHT G B, NEWTON A. Identification of a novel response regulator required for the swarmer-to-stalked-cell transition in Caulobacter crescentus [J]. Journal of Bacteriology, 1995, 177(21): 6223 − 6229. doi: 10.1128/jb.177.21.6223-6229.1995 [9] SOMMER J M, NEWTON A. Turning off flagellum rotation requires the pleiotropic gene pleD: pleA, pleC, and pleD define two morphogenic pathways in Caulobacter crescentus [J]. Journal of Bacteriology, 1989, 171(1): 392 − 401. doi: 10.1128/jb.171.1.392-401.1989 [10] DE SOUZA A A, IONESCU M, BACCARI C, et al. Phenotype overlap in Xylellafastidiosa Is controlled by the cyclic di-GMP Phosphodiesterase Eal in response to antibiotic exposure and diffusible signal factor-mediated cell-cell signaling [J]. Applied and Environmental Microbiology, 2013, 79(11): 3444 − 3454. doi: 10.1128/AEM.03834-12 [11] NINO-LIU D O, RONALD P C, BOGDANOVE A J. Xanthomonas oryzaepathovars: model pathogens of a model crop [J]. Molecular Plant Pathology, 2006, 7(5): 303 − 324. doi: 10.1111/j.1364-3703.2006.00344.x [12] YANG F, QIAN S, TIAN F, et al. The GGDEF-domain protein GdpX1 attenuates motility, exopolysaccharide production and virulence in Xanthomonas oryzaepv. oryzae [J]. Journal of Applied Microbiology, 2016, 120(6): 1646 − 1657. doi: 10.1111/jam.13115 [13] XUE D, TIAN F, YANG F, et al. Phosphodiesterase EdpX1 Promotes Xanthomonas oryzaepv. oryzaevirulence, exopolysaccharide production, and biofilm formation [J]. Applied and Environmental Microbiology, 2018, 84(22): e01717 − 18. [14] YANG F, TIAN F, SUN L, et al. A novel two-component system PdeK/PdeR regulates c-di-GMP turnover and virulence of Xanthomonas oryzaepv. oryzae [J]. Molecular Plant-Microbe Interactions, 2012, 25(10): 1361 − 1369. doi: 10.1094/MPMI-01-12-0014-R [15] LL Y R, ZOU H S, CHE Y Z, et al. A novel regulatory role of hrpD6 in regulating hrp-hrc-hpagenes in Xanthomonas oryzaepv. oryzicola [J]. Molecular Plant-Microbe Interactions, 2011, 24(9): 1086 − 1101. doi: 10.1094/MPMI-09-10-0205 [16] SCHAFER A, TAUCH A, JAGER W, et al. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum [J]. Gene, 1994, 145(1): 69 − 73. doi: 10.1016/0378-1119(94)90324-7 [17] ZHU PL. , ZHAO S, TANG J L, et al. ThersmA-like gene rsmAXooof Xanthomonasoryzaepv. oryzaeregulates bacterial virulence and production of diffusible signal factor [J]. Molecular Plant Pathology, 2011, 12(3): 227 − 237. doi: 10.1111/j.1364-3703.2010.00661.x [18] DILUZIO W R, TURNER L, MAYER M, et al. Escherichia coli swim on the right-hand side [J]. Nature, 2005, 435(7046): 1271 − 1274. doi: 10.1038/nature03660 [19] YU C, CHEN H, TIAN F, et al. A ten gene-containing genomic island determines flagellin glycosylation: implication for its regulatory role in motility and virulence of Xanthomonas oryzae pv. oryzae [J]. Molecular Plant Pathology, 2018, 19(3): 579 − 592. doi: 10.1111/mpp.12543 [20] TANG J L, LIU Y N, BARBER C E, et al. Genetic and molecular analysis of a cluster of rpfgenes involved in positive regulation of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar campestris [J]. MGG Molecular & General Genetics, 1991, 226(3): 409 − 417. [21] TIAN F, YU C, LI H, et al. Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas oryzaepv. oryzae [J]. Microbiological Research, 2015, 170: 177 − 183. doi: 10.1016/j.micres.2014.07.002 [22] BÜTNER D, BONAS U. Regulation and secretion of Xanthomonas virulence factors [J]. FEMS Microbiology Reviews, 2010, 34(2): 107 − 133. doi: 10.1111/j.1574-6976.2009.00192.x [23] JHA G , RAJESHWARI R , SONTI R V. Bacterial type two secretion system secreted proteins: Double-edged swords for plant pathogens[J]. Molecular Plant-Microbe Interactions, 2005, 18 (9): 891-898. [24] LI H, XUE D, TIAN F, et al. Xanthomonas oryzaepv. oryzae response regulator triP regulates virulence and exopolysaccharide production via interacting with c-di-GMP phosphodiesterase pdeR [J]. Molecular Plant-Microbe Interactions, 2019, 32(6): 729 − 739. doi: 10.1094/MPMI-09-18-0260-R -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1075
- HTML全文浏览量: 351
- PDF下载量: 116
- 被引次数: 0
