

-
实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)因其重复性、特异性和灵敏度等方面的优点,常被研究人员用于评估目的基因的表达水平[1-2]。在利用qRT-PCR检测目的基因的表达水平时,需选择稳定表达的内参基因作为内源性的参照基因对结果进行校正,以降低样本之间的差异[3]。因此,内参基因的表达应在所需检测的全部样本中是稳定的,且不受试验条件的影响。近年的研究表明,不同试验条件或不同植物组织部位中稳定表达的内参基因有差异[4-5]。例如,柳杉(Cryptomeria japonica)在脱落酸(Abscisic acid,ABA)处理下,稳定表达的内参基因为ACT,但在赤霉素(Gibberellins,GA)、水杨酸(Salicylic acid,SA)和茉莉酸甲酯(Methyl jasmonate,MeJA)处理下ACT的稳定性较差[6];管花肉苁蓉(Cistanche tubulosa)不同组织部位中表达最稳定的内参基因是DNAJ,但在ABA和GA处理下,DNAJ的稳定性不如EF-1α[7]。上述研究结果表明,随着试验条件的变化稳定内参基因的表达也可能会出现波动,因此,需要根据不同的试验条件筛选稳定的内参基因。
植物的生长发育和环境适应性受多种植物激素的综合调控。例如,生长素响应因子(auxin response factor,ARF)通过生长素(auxin,IAA)和GA 信号串扰共同调控番茄果实成熟[8];ARF7通过IAA和SA信号串扰调控拟南芥的侧根发育以响应病原菌的入侵[9];基因BZR1(brassinozale-resistant 1)通过油菜素内酯(brassinosteroid,BR)和GA信号串扰调节水稻的生长发育[10]。上述研究结果表明,植物激素信号途径的关键基因可能受到多种植物激素信号的调控。因此,在分析和验证植物激素响应基因的表达水平时,应优先选择在多种植物激素处理条件下表达水平均较为稳定的内参基因[11-12]。
番茄(Solanum lycopersicum)是遗传学和分子生物学的研究模型。近年来,番茄内参基因的筛选已有较多的研究报道,但关于外源植物激素处理的种类较少。例如,白圣懿等[13]的研究仅针对ABA、 MeJA和SA等3种外源植物激素处理下的稳定内参基因进行了筛选;韩晓雪等[14]对番茄不同组织部位中内参基因的稳定性进行了筛选,但研究内容未涉及不同外源植物激素处理条件,因此仍需筛选番茄在不同外源植物激素处理和不同组织部位中稳定表达的内参基因。基于此,本研究以番茄不同组织部位及IAA、GA、ABA、细胞分裂素(cytokinin,CTK)、SA、BR和乙烯(ethylene,ETH)7种植物激素为处理条件,以DANJ、EF-1α、ACT、UBI、APT、CAC、TIP41和RPL8作为候选内参基因,利用geNorm、NormFinder、BestKeeper、Delta CT和RefFinde等算法综合评估候选内参基因的稳定性,研究结果可为后续番茄响应外源激素处理的基因表达网络分析和分子调控机制研究提供理论支撑。
-
本试验中的番茄品种为 “Alisa Craig”(AC)。番茄种子用5%的次氯酸钠溶液消毒5 min,无菌水清洗3次,再用75%的乙醇消毒30 s,无菌水清洗3次。番茄种子在无菌培养皿中萌发后,将发芽的种子转移到1/4强度的霍格兰营养液中生长。置于光周期为光:暗=12 h:12 h、温度为(24±1) ℃的培养箱中。
-
选择14 d龄长势一致且健康的水培番茄幼苗进行处理,外源植物激素的处理浓度参考Xu等[12]的方法,使用10 μmol·L−1 IAA、1 mmol·L−1 SA、0.1 μmol·L−1 BR及10 μmol·L−1 ETH,参考Omena-Garcia等[15]的方法,使用10 μmol·L−1 GA,参考Hu等[16]的方法,使用1 μmol·L−1 ABA,参考Lu等[17]的方法,使用10 μmol·L−1 CTK。参考Albuquerque等[18]的方法收集不同处理组0 、3和6 h番茄的根、茎和叶片组织提取RNA并反转录为cDNA,然后将同一处理条件下的根、茎和叶片组织的cDNA按照时间段等量混合用于qRT-PCR分析。不同组织部位的样品为收集的正常水培条件下番茄的根、茎和叶片组织样品。每个处理均重复3次。样品置于液氮中,−80 ℃冰箱保存备用。
-
使用植物组织总RNA提取试剂盒(天根,N2730,中国)提取番茄不同处理组的总RNA,提取步骤按照试剂盒的说明书进行。使用ToloScript 第一链 cDNA 合成试剂盒(ToLoBio,中国上海)合成 cDNA,操作步骤参考说明书,cDNA 置于−20 ℃冰箱保存备用。
-
选择DNAJ蛋白(DnaJ-like protein/Protein binding/folding, DNAJ)、延伸因子(Elongation factor 1-α,EF-1α)、肌动蛋白(Actin,ACT)、泛素(Ubiquitin 3,UBI)、腺嘌呤磷酸核糖基转移酶(Adenine phosphoribosyltransferase,APT)、网格蛋白连接复合物介质(Clathrin adaptor complexes medium subunit,CAC)、相互作用蛋白(TIP41-like protein,TIP41)和60S核糖体蛋白(60S ribosomal protein L8,RPL8)等8个常见的内参基因作为候选内参,基因及引物序列来源于文献,引物由北京六合华大基因科技有限公司合成,详细信息见表1。
表 1 候选内参基因引物序列
Table 1. Primer sequences of candidate reference genes
基因
Gene基因ID
Gene ID引物序列5′—3′(F and R)
Primer sequence参考文献
ReferencesDNAJ Solyc04g081530 GAGCACACATTGAGCCTTGAC [19] CTTTGGTACATCGGCATTCC EF-1α Solyc05g693059 GATTGACAGACGTTCTGGTAAGGA [20] ACCGGCATCACCATTCTTCA ACT Solyc03g078400 GTCCTCTTCCAGCCATCCAT [20] ACCACTGAGCACAATGTTACCG UBI Solyc11g056940 TCGTAAGGAGTGCCCTAATGCTGA [21] CAATCGCCTCCAGCCTTGTTGTAA APT Solyc04g077970 GAACAGACAAGATTGAGATGCATGTA [20] CCACGAGGGCACGTTCA CAC Solyc08g006960 CCTCCGTTGTGATGTAACTGG [22] ATTGGTGGAAAGTAACATCATCG TIP41 Solyc10g049850 ATGGAGTTTTTGAGTCTTCTGC [19] GCTGCGTTTCTGGCTTAGG RPL8 Solyc10g006580 CCGAAGGAGCTGTTGTTTGTA [22] ACCTGACCAATCATAGCACGA -
以番茄不同组织的cDNA等量混合为模板,对8个候选内参基因进行PCR扩增,扩增反应体系10.0 µL:2×Taq Plus Master Mix(Dye Plus)5.0 µL,10 µmol·L−1上、下游引物各0.4 µL,100 ng·µL−1 cDNA模板0.2 µL,RNase-Free ddH2O补足至10.0 µL。扩增程序:95 ℃预变性3 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸15 s,以上3个步骤30个循环;72 ℃延伸10 min,待PCR反应完成之后,使用1%琼脂糖凝胶电泳检测扩增产物。以所有样本的cDNA为模板,根据qRT-PCR系统的熔解曲线进行引物特异性检测。每个样本3次重复。
-
以所有样本的cDNA为模板在qTower3实时荧光定量PCR系统(Analytikjena,GER)中使用ChamQ Universal SYBR qPCR Master Mix 试剂盒(Vazyme)检测候选内参基因的Ct值,反应体系15.0 µL:2×Taq Plus Master Mix 7.5 µL,10 µmol·L−1正、反向引物各0.3 µL,100 ng·µL−1 cDNA模板1.0 µL,RNase-free ddH2O补足至15.0 µL。扩增程序:95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火延伸45 s,以上2个步骤进行40个循环,60 ℃再延伸15 s,每个样本3次重复。
-
根据qRT-PCR得到的结果绘制候选内参基因Ct值的箱线图。候选内参基因的稳定性分析使用geNorm、NormFinder、BestKeeper、Delta CT等算法评估,通过RefFinder(
http://www.heartcure.com.au/reffinder/ )在线程序综合分析候选内参基因的稳定性。 -
选择长势一致的14 d苗龄番茄幼苗,将幼苗置于10 μmol·L−1 IAA的营养液中处理,收集处理后0 、3 和6 h的番茄根、茎和叶部组织,按照1.3的方法提取总RNA和合成cDNA,以cDNA为模板,根据IAA处理和不同组织中筛选的最稳定和最不稳定的基因为内参,使用qRT-PCR仪检测样本中SlGH3.4基因的表达情况,体系和程序同1.6,数据采用2−△△Ct法计算,每个样本重复3次,目的基因引物序列参考Chen等[23]。
-
使用Graphpad Prism软件绘图;使用SPSS 26.0软件进行显著性分析(P<0.05)。
-
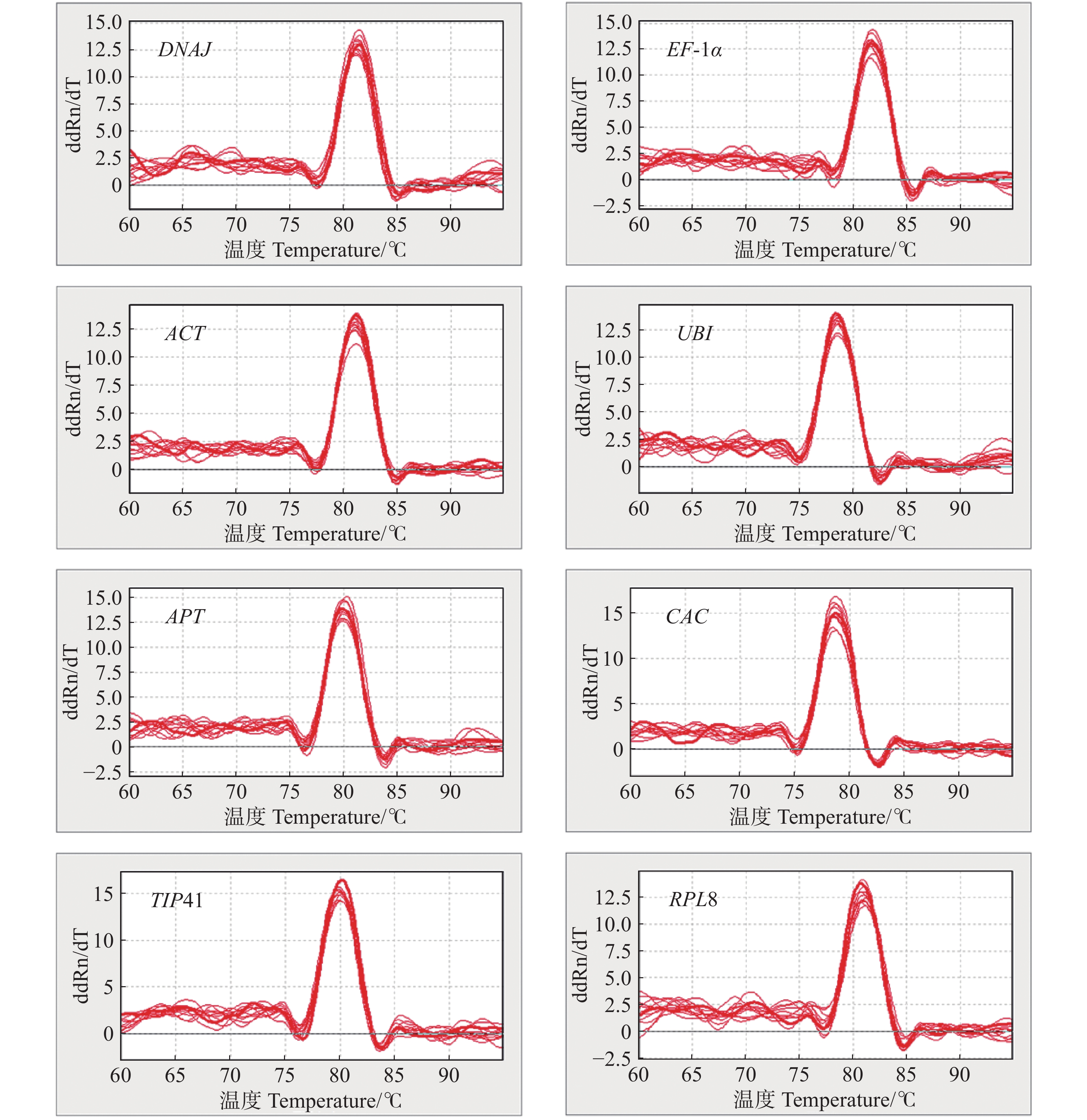
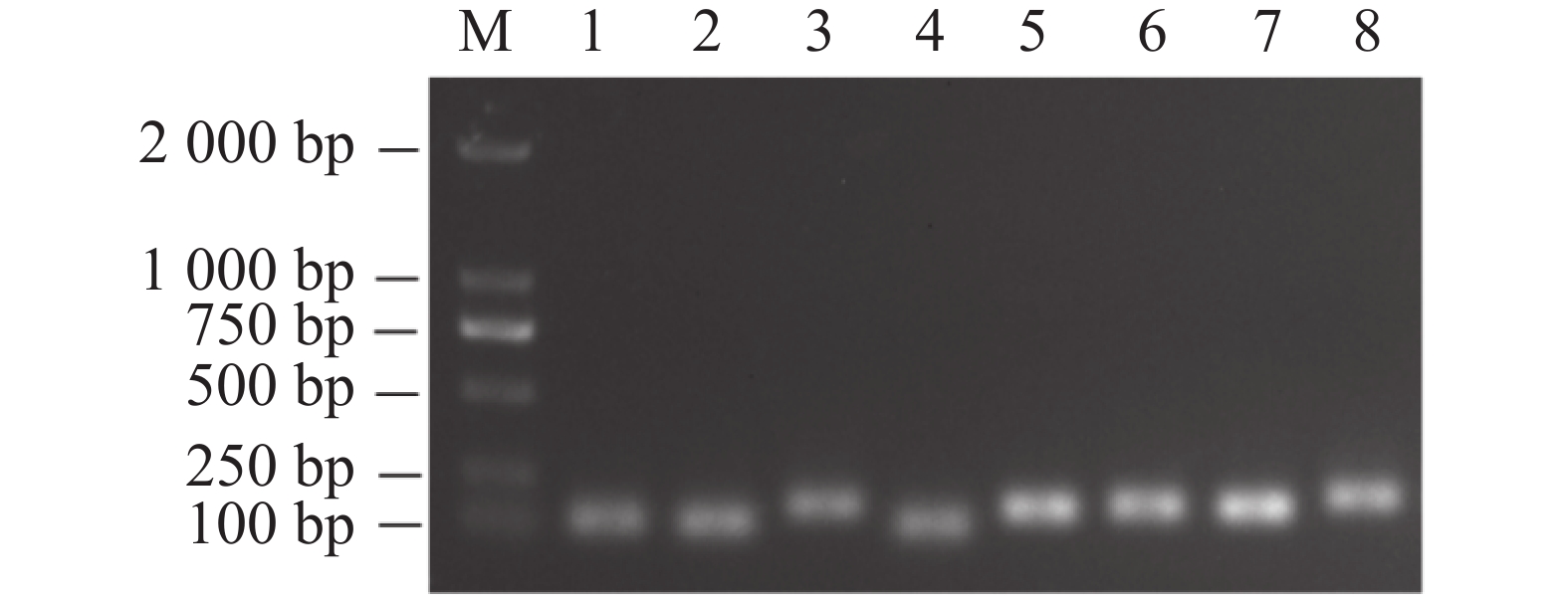
以不同组织等量混合的cDNA为模板,对8个候选内参基因进行PCR扩增,并使用1%的琼脂糖凝胶进行电泳检测扩增产物的特异性。如图1所示,挑选的8个候选内参基因PCR的扩增产物条带单一,无引物二聚体,且PCR扩增片段的大小与预测一致。由图2可知,8个内参基因的熔解曲线都呈现单峰,表明了候选内参基因引物具有较高的特异性和可靠性,可进行后续实验。

图 1 候选内参基因PCR扩增结果
Figure 1. Results of routine PCR amplification of candidate reference genes in tomato root, stem and leaf tissues
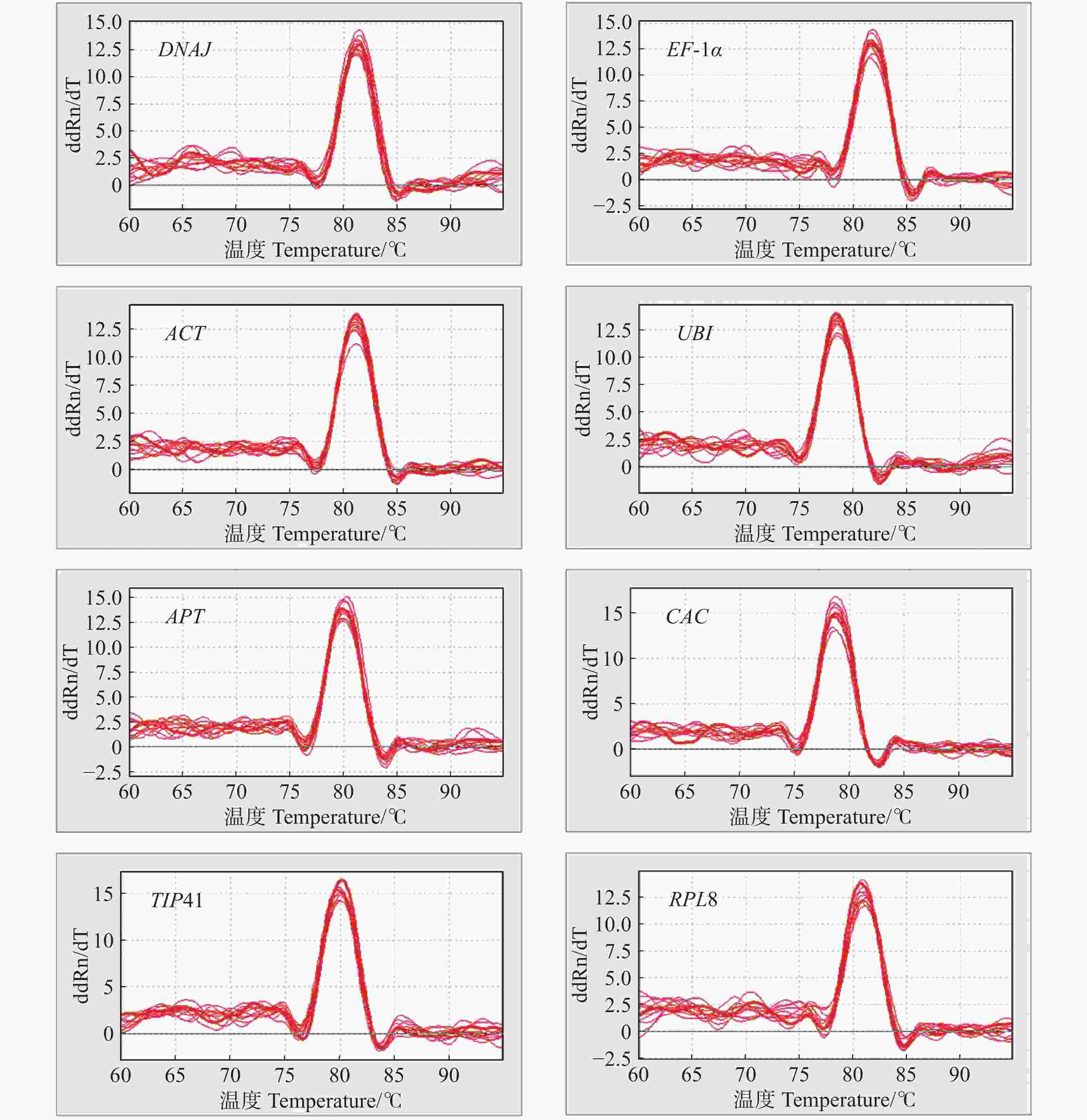
图 2 候选内参基因的熔解曲线
Figure 2. Melting curve of tomato candidate reference genes
-
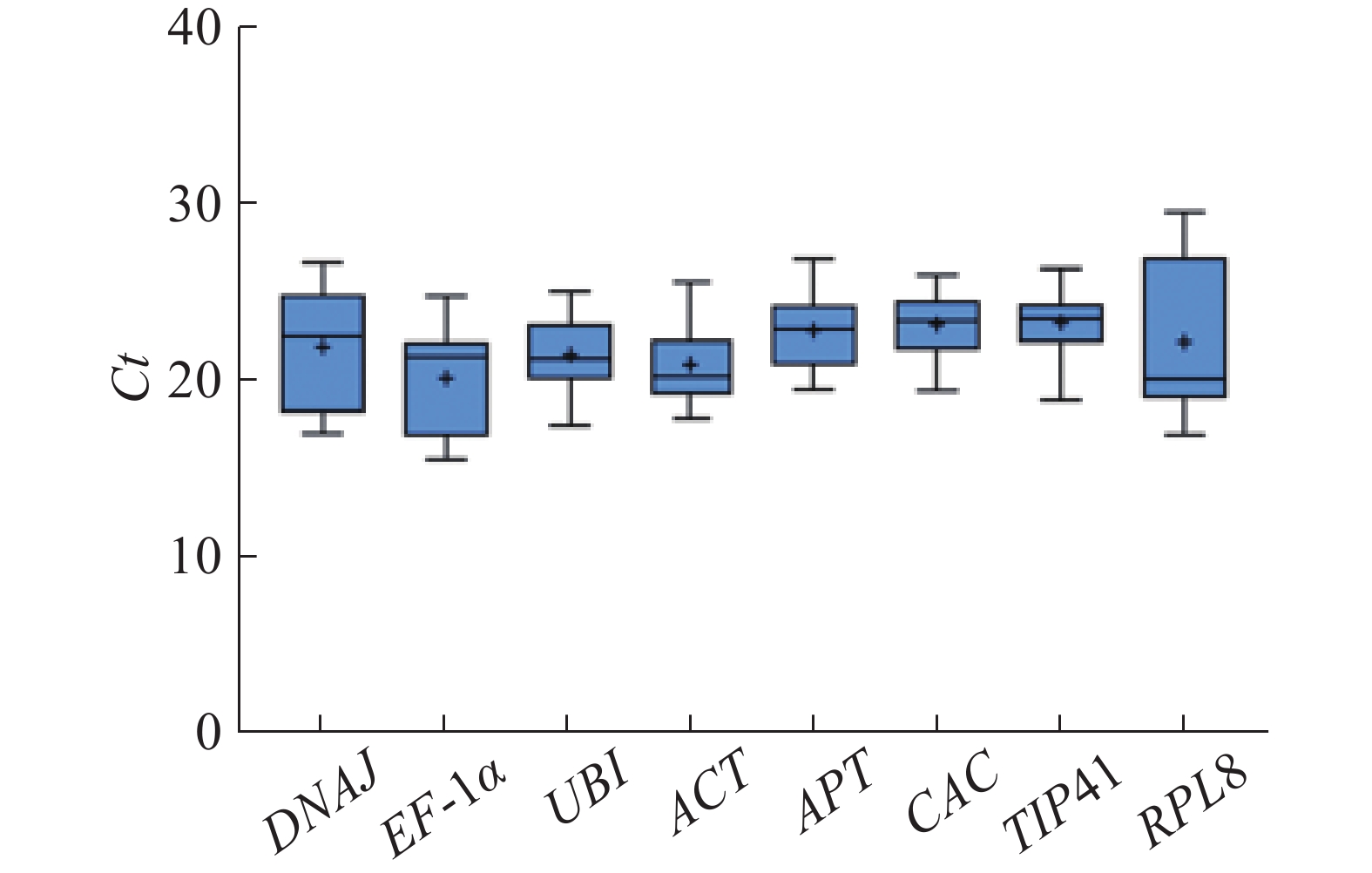
根据所有样品中的Ct值分布情况绘制箱线图(图3)。Ct值越低,说明基因的表达丰度越高。 8个候选内参基因的Ct值范围为15.42~29.55,其中EF-1α的平均Ct值最低(平均Ct值20.18),表达丰度最高,TIP41的表达丰度最低(平均Ct值23.32)。RPL8和DNAJ基因的波动范围较大,在所有样品中表达均不稳定。其他基因的Ct值离散程度大体相同,没有明显区别。
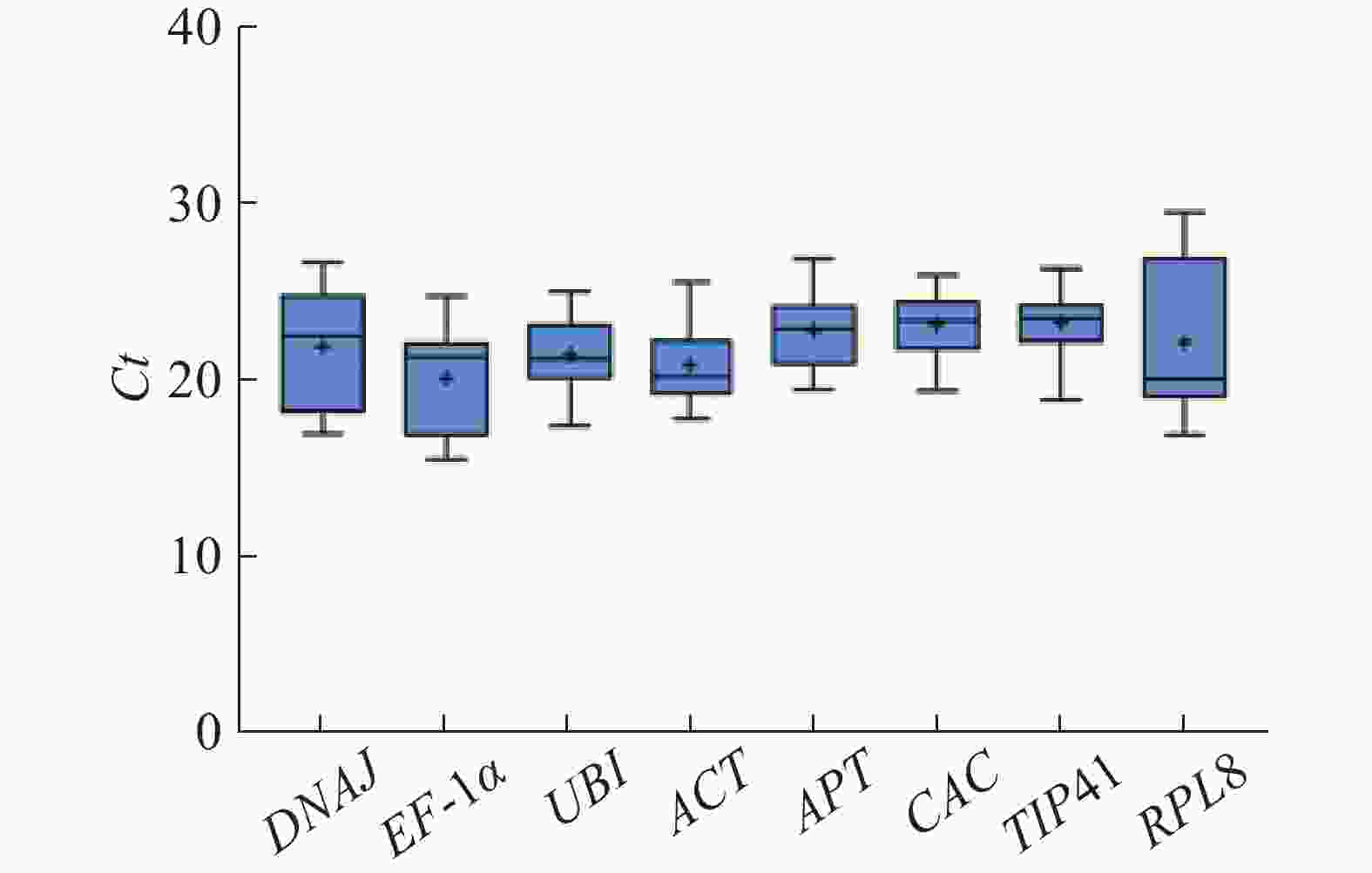
图 3 所有样品中8个候选内参基因的Ct值
Figure 3. Ct values of 8 candidate reference genes in all the samples
-
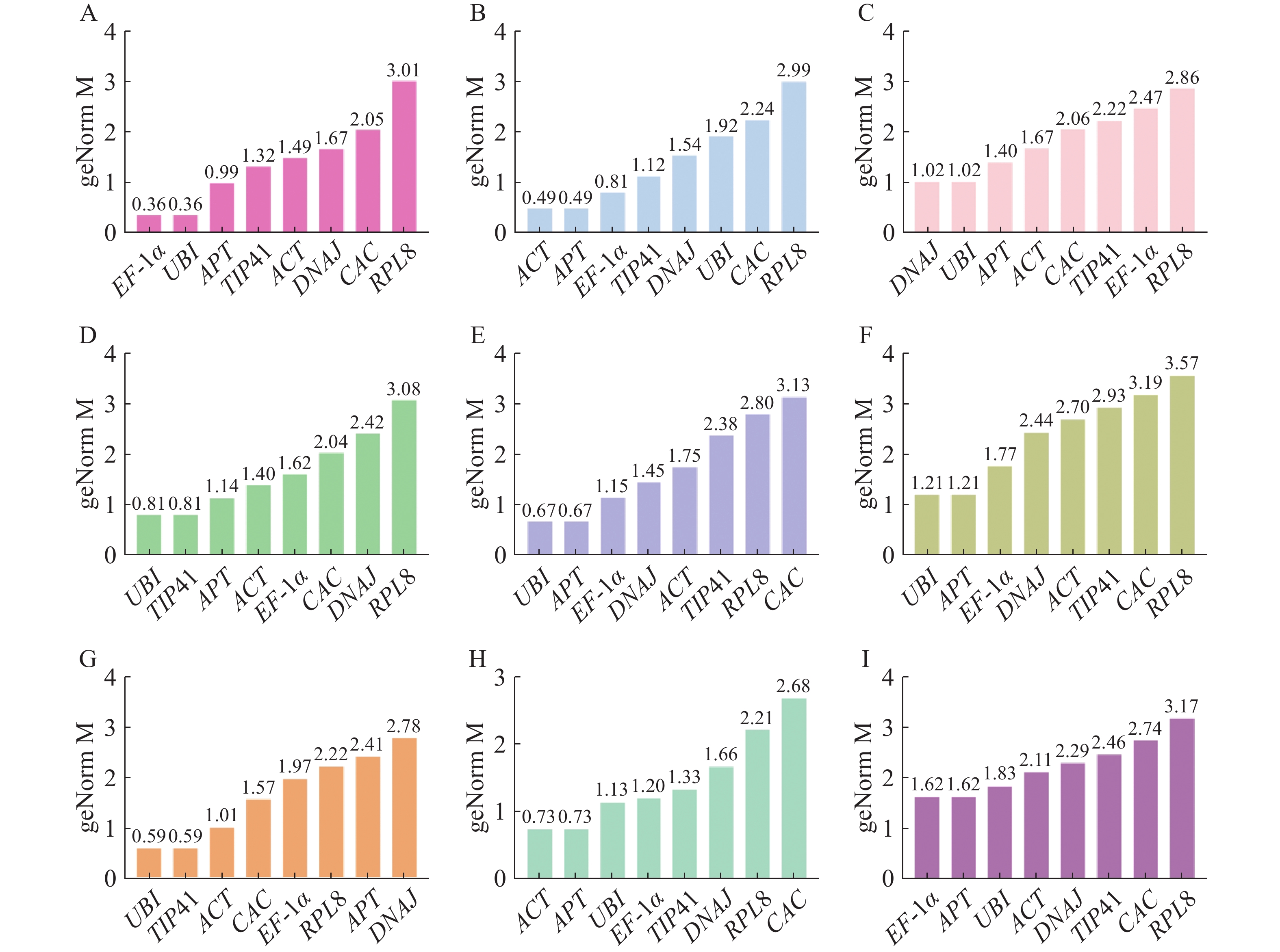
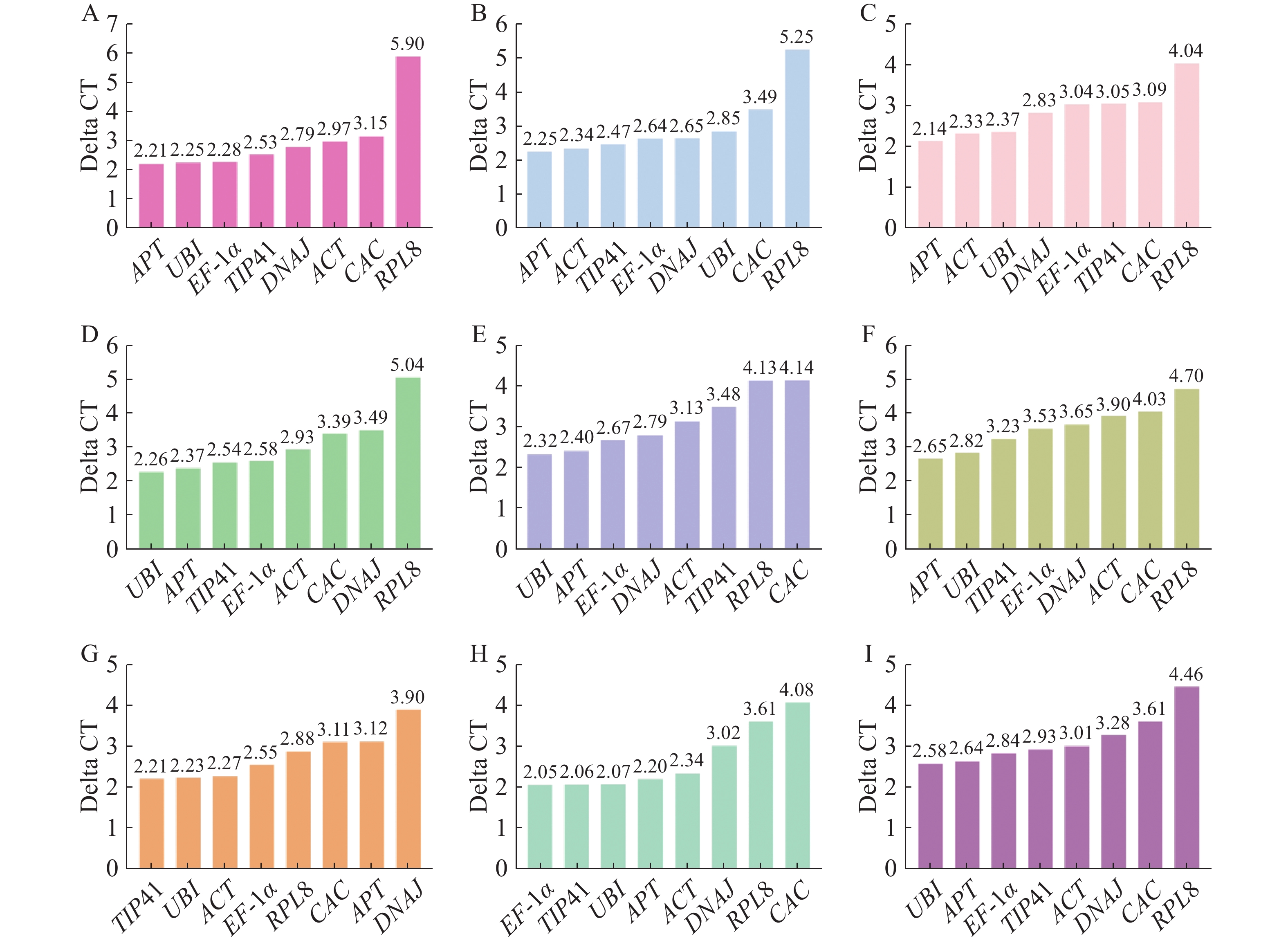
geNorm分析是用M值(稳定值)评估内参基因的稳定性,阈值为1.5,且M值越小稳定性越好[24]。根据geNorm分析结果(图4),在生长素处理下EF-1α和UBI表达最稳定;赤霉素处理和不同组织中ACT和APT表达最稳定;脱落酸处理下,DNAJ和UBI表达最稳定;细胞分裂素和乙烯处理下的最佳内参基因是UBI和TIP41;油菜素内酯和水杨酸处理下APT和UBI是最稳定的内参基因。在所有样品中,候选内参基因的表达稳定性强弱排序为EF-1α=APT>UBI>ACT>DNAJ>TIP41>CAC>RPL8,APT和EF-1α是最稳定的内参基因。
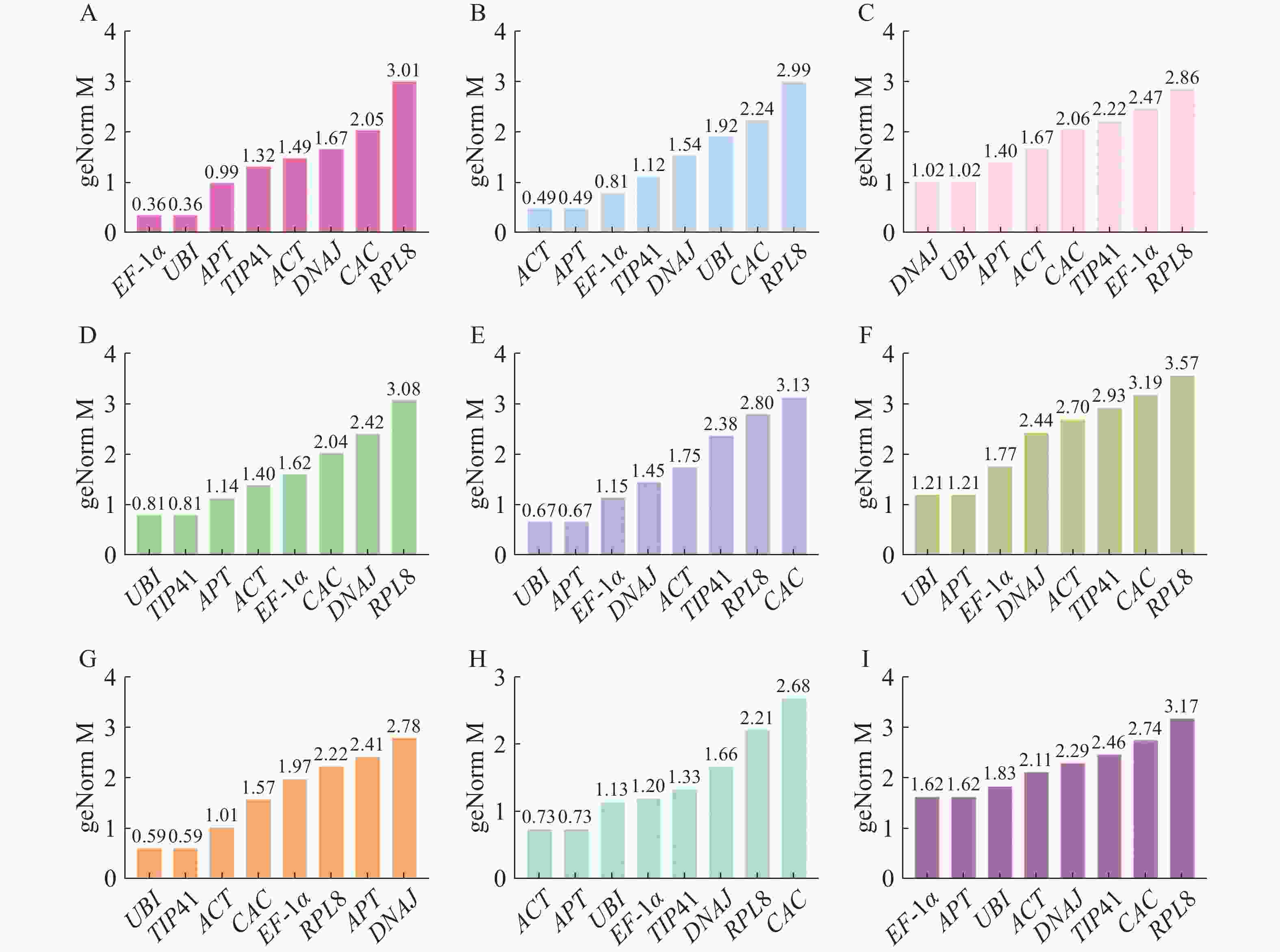
图 4 geNorm分析不同处理下番茄幼苗中候选内参基因的稳定性
Figure 4. geNorm analysis of stability of candidate internal reference genes in tomato seedlings under different treatments
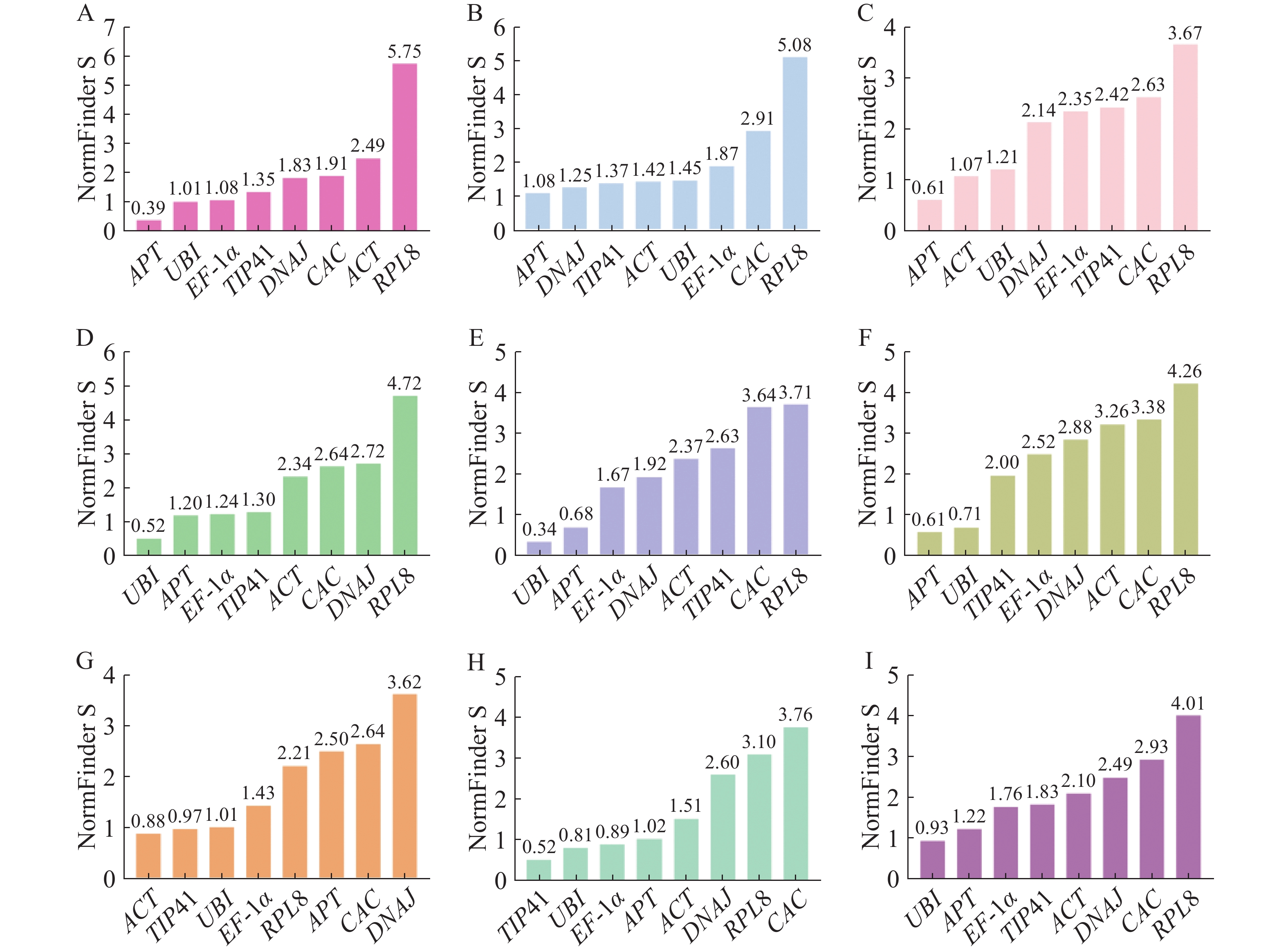
NormFinder分析是通过方差分析(S值)评估内参基因的稳定性,S值越小稳定性越好[25]。如图5所示,在生长素、赤霉素、脱落酸和油菜素内酯处理下的最佳内参基因为APT;乙烯处理下的最佳内参基因为ACT;不同组织中最稳定的内参基因为TIP41。在所有样品中,UBI是表达最稳定的内参基因,同时也是细胞分裂素和水杨酸处理下最稳定的内参基因。
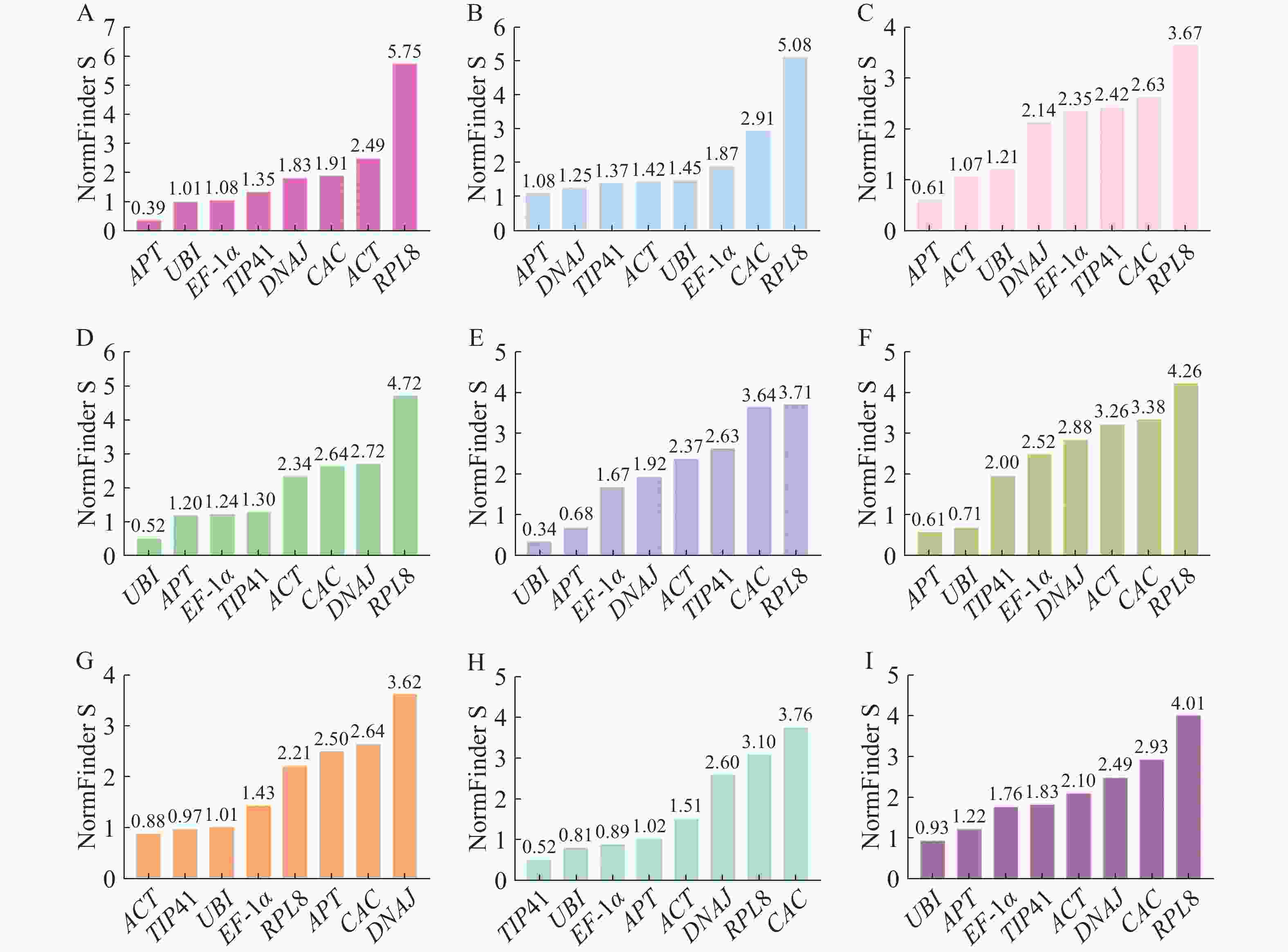
图 5 NormFinder分析不同处理下番茄幼苗中候选内参基因的稳定性
Figure 5. NormFinder analysis of stability of candidate internal reference genes in tomato seedlings under different treatments
-
用BestKeeper软件测算SD值(标准偏差)和CV值(变异系数)来评估内参基因表达的稳定性,SD值和CV值越小,内参基因的稳定性越高[26]。从表2可知,在赤霉素和水杨酸处理下SD值和CV值最小的内参基因为CAC;脱落酸处理和不同组织中SD值和CV值最小的内参基因分别是ACT和APT;其余处理下SD值和CV值最小的内参基因均为TIP41。此外,BestKeeper软件还可以对内参基因的Ct值测算相关性系数(r),SD值和CV值越小且r值较高的内参基因越稳定[27]。在脱落酸处理下ACT的r值为0.652;不同组织中APT的r值为0.991;其余处理下排名最高的内参基因r值均较低,但最终排名需要综合SD值和CV值分析。因此,在赤霉素和水杨酸处理下最稳定的内参为CAC;脱落酸处理下最稳定的内参为ACT;不同组织中最稳定内参为APT;其余处理下最稳定的内参均为TIP41。
表 2 BestKeeper分析结果
Table 2. BestKeeper analysis results
生长素 Auxin 赤霉素 Gibberellin 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.86 3.68 0.311 CAC 0.73 3.07 0.001 1 CAC 1.06 4.52 0.001 TIP41 1.10 4.66 0.668 2 APT 1.11 5.39 0.778 APT 1.93 8.73 0.840 3 UBI 1.94 9.36 0.924 ACT 1.96 9.29 0.788 4 EF-1α 2.06 10.65 0.942 UBI 2.33 10.85 0.845 5 ACT 2.24 10.66 0.433 EF-1α 2.69 13.32 0.857 6 DNAJ 2.75 12.47 0.985 DNAJ 2.82 13.07 0.994 7 RPL8 4.35 19.59 0.180 RPL8 4.73 21.43 0.593 8 细胞分裂素 Cytokinin 脱落酸 Abscisic acid 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.52 2.12 0.001 ACT 0.32 1.60 0.652 1 UBI 0.62 2.87 0.299 CAC 0.84 3.46 0.001 2 CAC 1.35 5.79 0.001 TIP41 1.08 4.68 0.001 3 APT 1.36 6.21 0.528 APT 1.49 6.68 0.999 4 EF-1α 1.81 8.95 0.948 UBI 1.90 8.72 0.852 5 ACT 2.11 9.61 0.422 DNAJ 2.09 9.81 0.691 6 DNAJ 2.95 12.61 0.989 EF-1α 3.06 14.77 0.996 7 RPL8 3.81 16.59 0.312 RPL8 3.82 16.68 0.801 8 表 2 续表2 Tab.2 Continued
油菜素内酯 Brassinolide 乙烯 Ethylene 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.78 3.57 0.001 TIP41 0.41 1.83 0.001 1 CAC 1.47 6.57 0.001 UBI 0.51 2.43 0.001 2 APT 1.77 7.82 0.986 ACT 0.94 4.70 0.416 3 UBI 2.04 9.57 0.906 CAC 1.50 6.75 0.001 4 DNAJ 2.49 11.83 0.512 RPL8 1.75 7.84 0.345 5 ACT 2.60 12.06 0.267 EF-1α 1.77 8.54 0.910 6 EF-1α 3.34 16.02 0.988 APT 2.44 10.62 0.785 7 RPL8 4.28 19.29 0.739 DNAJ 2.90 13.08 0.598 8 水杨酸 Salicylic acid 不同组织 Different tissues 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r CAC 1.43 6.07 0.001 APT 1.23 5.03 0.991 1 TIP41 1.65 7.04 0.001 CAC 1.32 5.87 0.001 2 UBI 1.67 7.69 0.984 ACT 1.83 9.13 0.712 3 APT 1.94 8.62 0.987 UBI 1.94 8.72 0.926 4 ACT 1.94 8.90 0.420 TIP41 2.21 9.87 0.770 5 DNAJ 2.31 10.96 0.765 EF-1α 2.28 11.92 0.964 6 EF-1α 2.56 12.64 0.931 RPL8 2.91 13.70 0.541 7 RPL8 3.79 17.38 0.754 DNAJ 3.69 16.03 0.998 8 所有样品 All Samples 排名 Rank 基因 Gene SD CV r TIP41 1.17 5.01 0.194 1 CAC 1.37 5.92 0.001 2 UBI 1.68 7.80 0.839 3 ACT 1.82 8.68 0.466 4 APT 1.88 8.46 0.803 5 EF-1α 2.50 12.41 0.894 6 DNAJ 2.86 13.03 0.782 7 RPL8 3.68 16.56 0.550 8 注:所有样品表示不同组织和不同外源植物激素处理下的样品。
Note: All samples represent those under different plant parts and different exogenous plant hormone treatments. -
Delta CT(ΔCt)法是根据基因表达差异的可重复性对候选基因稳定性进行排序,计算公式为Q=EΔCt,Delta CT值越小,内参基因稳定性越好[28]。分析结果(图6)表明,在生长素、赤霉素、脱落酸和油菜素内酯处理下,APT均能稳定表达;乙烯处理下的最佳内参基因为TIP41;不同组织中表达最稳定的内参基因为EF-1α。在所有样品中、细胞分裂素和水杨酸处理下UBI表达最稳定。
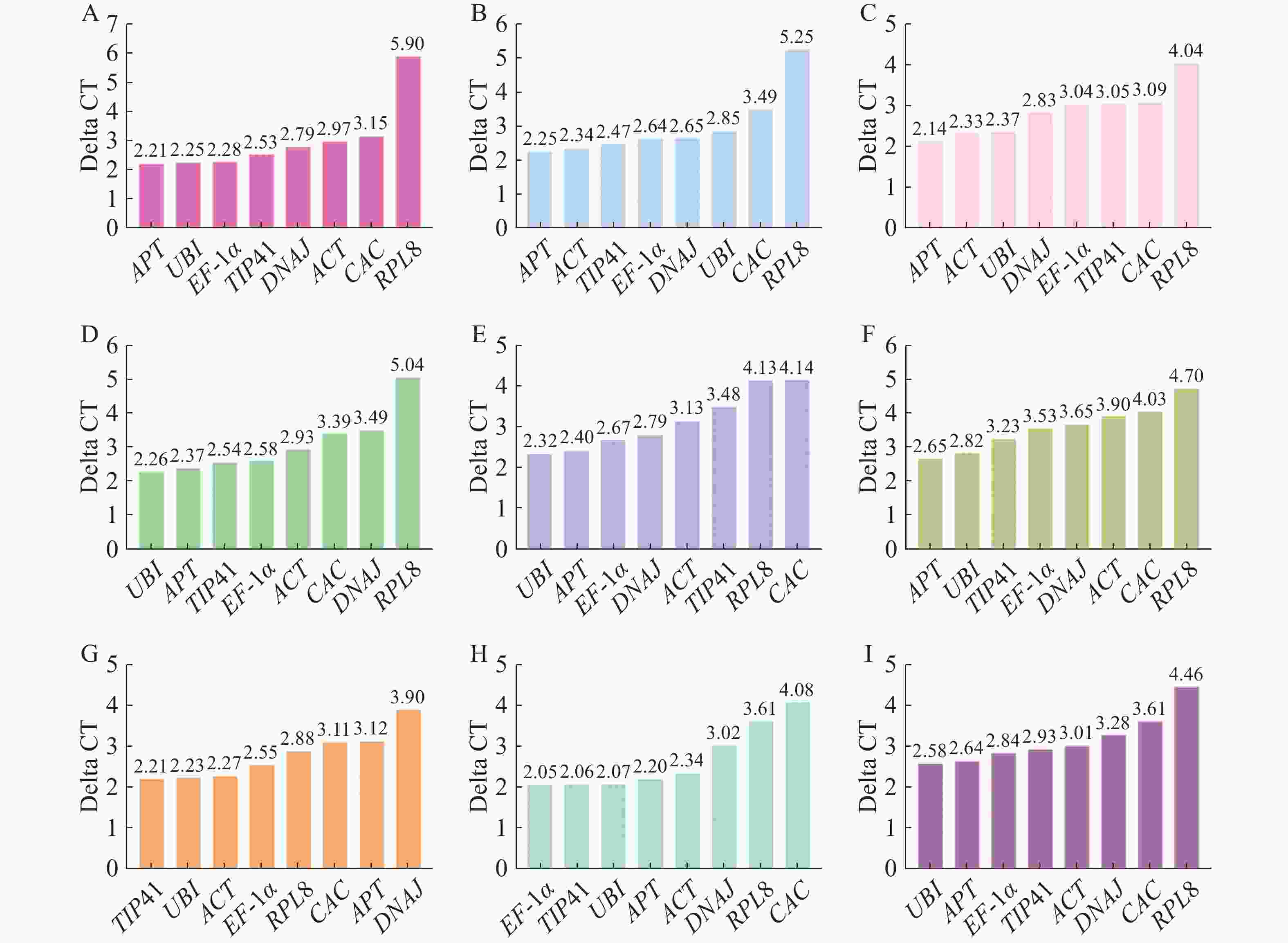
图 6 Delta CT 分析结果
Figure 6. Delta CT analysis results
-
geNorm、NormFinder、BestKeeper、Delta CT等4种算法的结果有一定的偏差,是4种算法的原理和程序不同导致。因此,本研究利用RefFinder在线程序对这4种算法进行综合评估。由表3可知,在生长素、赤霉素、脱落酸、油菜素内酯处理下和不同组织中APT表达最稳定;乙烯处理下表达最稳定的内参基因为TIP41;在所有样品中、细胞分裂素和水杨酸处理下最稳定的内参基因为UBI。
表 3 候选内参基因表达稳定性综合分析结果
Table 3. Comprehensive analysis results of the expression stability of candidate reference genes
处理
Treatment综合排名
Comprehensive ranking排序
Sort处理
Treatment综合排名
Comprehensive ranking排序
Sort生长素
AuxinAPT 1 油菜素内酯
BrassinolideAPT 1 UBI 2 UBI 2 EF-1α 3 TIP41 3 TIP41 4 EF-1α 4 CAC 5 DNAJ 5 DNAJ 6 CAC 6 ACT 7 ACT 7 RPL8 8 RPL8 8 赤霉素
GibberellinAPT 1 乙烯
EthyleneTIP41 1 ACT 2 UBI 2 TIP41 3 ACT 3 CAC 4 EF-1α 4 DNAJ 5 CAC 5 EF-1α 6 RPL8 6 UBI 7 APT 7 RPL8 8 DNAJ 8 细胞分裂素
CytokininUBI 1 水杨酸
Salicylic acidUBI 1 TIP41 2 APT 2 APT 3 EF-1α 3 EF-1α 4 DNAJ 4 ACT 5 TIP41 5 CAC 6 CAC 6 DNAJ 7 ACT 7 RPL8 8 RPL8 8 脱落酸
Abscisic acidAPT 1 不同组织
Different tissuesAPT 1 ACT 2 TIP41 2 UBI 3 EF-1α 3 DNAJ 4 UBI 4 CAC 5 ACT 5 TIP41 6 CAC 6 EF-1α 7 DNAJ 7 RPL8 8 RPL8 8 所有样品
All SamplesUBI 1 APT 2 EF-1α 3 TIP41 4 ACT 5 CAC 6 DNAJ 7 RPL8 8 注:所有样品表示不同组织和不同外源植物激素处理下的样品。 Note: All samples represent those under different plant parts and different exogenous plant hormone treatments. -
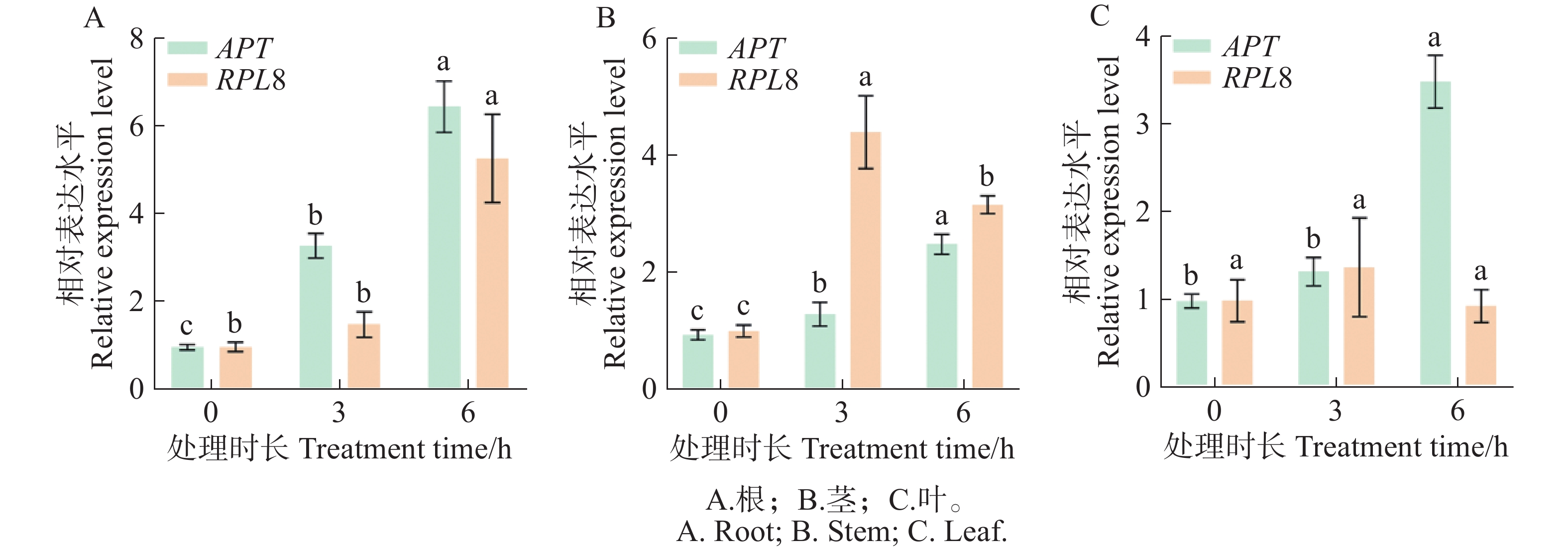
IAA作为促进和影响植物发育及生理变化的激素,几乎影响着植株形态发育的全过程[29]。GH3基因是早期响应IAA的关键基因之一[30]。以IAA处理和不同组织中,综合排名最稳定的内参基因APT和最不稳定的内参基因RPL8对生长素响应基因GH3.4进行不同组织部位的表达分析。结果(图7)表明,在IAA的诱导下,以稳定的APT为内参基因时,SlGH3.4基因在番茄根、茎和叶片中的表达模式都呈上升趋势,表达差异显著。以不稳定的RPL8为内参基因时,根部表达模式呈上升趋势,而茎部和叶片的表达模式呈先上升后下降的趋势,表达量明显变化。以上结果表明筛选出的稳定内参基因较准确和可靠。
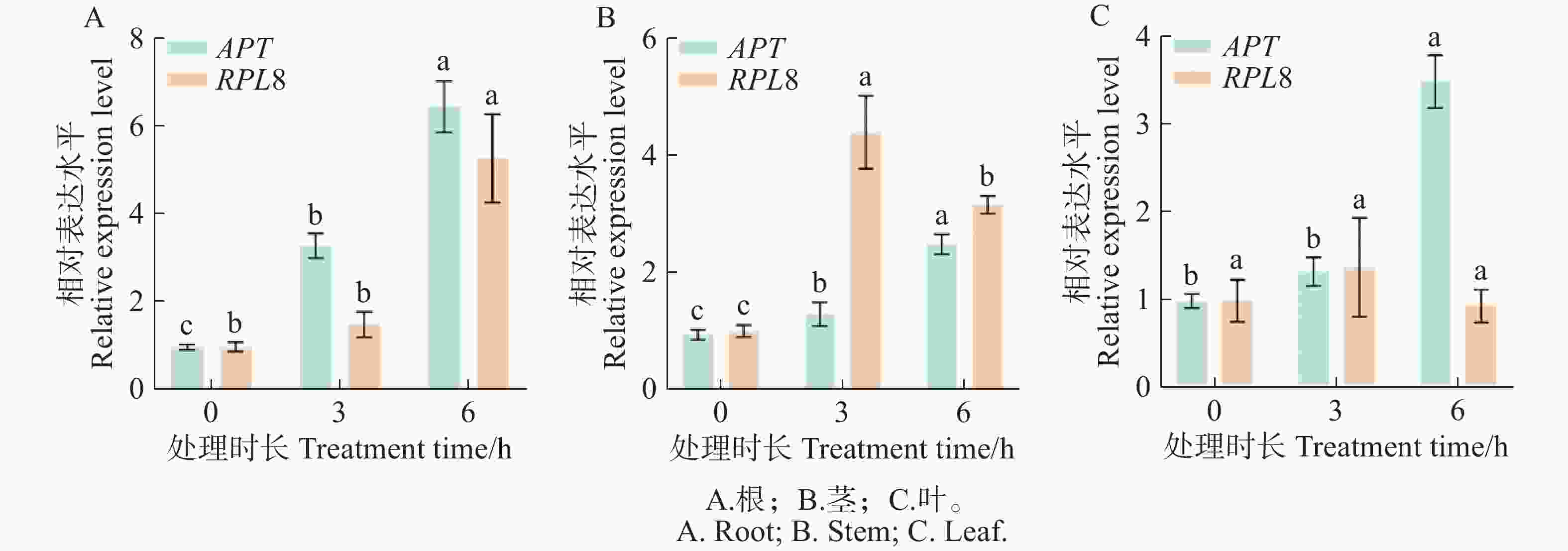
图 7 外源IAA诱导下SlGH3.4基因的表达模式
Figure 7. Expression patterns of SlGH3.4 gene induced by exogenous IAA
-
qRT-PCR是评价目的基因表达水平的重要技术手段,而内参基因的稳定性是保证试验结果精确度的关键影响因素[31]。同一物种在不同试验条件下或者不同组织中内参基因的稳定性也存在差异,因此选择合适的内参基因来减少试验中的误差是极其重要的[32]。例如,宋雄[33]发现欧芹(Petroselinum crispum )在SA处理下ACT和EF-1α表达的稳定性最佳,但本研究结果显示,番茄在SA处理条件下ACT的稳定性不如EF-1α,这与白圣懿等[13]的报道一致。此外,黄丽萍等[34]的研究结果显示,APT和UBI在香瓜茄不同组织中表达稳定性最好。在本研究中,APT和UBI不仅在番茄不同组织中表达较稳定,还在不同植物激素条件下表达最稳定,而在紫花苜蓿中[35],不同植物激素条件下ACT的表达最稳定,这进一步说明不同试验条件下稳定表达的理想内参基因并不存在。综上所述,本研究根据多种植物激素处理和不同组织中各内参基因稳定性的综合排名,发现APT和UBI的表达相对较稳定。
-
内参基因稳定性的筛选方法因其各自的原理和程序不同,故得出的结果也不同。番茄自交系“Hawaii 7996”在接种青枯菌菌株 ‘RS 489’ 后进行内参基因的稳定性筛选,在geNorm和NormFinder算法中最稳定的内参基因均为TIP41,而BestKeeper分析结果显示最稳定的内参基因为UBI[18]。本研究中,在geNorm、NormFinder和Delta Ct算法中SA和BR处理下最稳定的内参基因均为APT和UBI,而BestKeeper分析结果显示最稳定的内参基因分别为CAC和TIP41。geNorm、NormFinder和Delta Ct的分析结果类似,但是与BestKeeper分析略有差异,这种现象也存在于石蒜属和无患子植物中[4,36]。因此,为了减小软件算法原理不同所造成的差异,利用RefFinder在线程序进行综合分析,从而综合评估出最稳定的内参基因[37]。
-
为了确定分析结果的准确性,选择生长素响应基因GH3.4对筛选出的内参基因进行验证。GH3基因编码的酰胺合成酶是生长素动态平衡的调节因子,SlGH3.4基因在IAA处理和丛枝菌根(AM)真菌定植的根中被强烈诱导[23,38]。通过qRT-PCR检测最稳定的内参基因APT和最不稳定的内参基因RPL8对SlGH3.4基因的表达水平。结果显示,在外源IAA处理下,选择稳定的基因为内参时,SlGH3.4基因在番茄不同组织中的表达趋势一致,证明了所筛选的内参基因适用于分析IAA信号转导相关基因的表达水平。而选择不稳定的基因作为参考时,SlGH3.4基因在茎部和叶片的表达量发生明显变化,说明使用不稳定的内参导致基因表达的分析结果不可靠,这与岳炜楠等[35]、Liu等[39]的研究结果一致。因此,在选择不同处理及不同组织样品内参基因时,需要结合多种算法的综合分析结果并对结果进行验证,以提高结果的准确性。
-
本研究利用geNorm、NormFinder、BestKeeper、Delta CT以及RefFinder等算法对番茄不同组织及不同激素处理下内参基因的稳定性进行综合评估,结果表明,在IAA、GA、ABA、CTK、SA、BR、ETH等7个激素处理条件下、不同组织部位及所有样品中,均表达较稳定的内参基因为APT和UBI。使用稳定的内参基因APT和不稳定的内参基因RPL8对筛选的结果进行验证,发现选择稳定的内参基因可更准确展示目的基因的表达水平。综上所述,本研究为后续番茄在不同激素条件下和不同组织中筛选合适的内参基因提供了一定的理论参考。
番茄不同组织及不同激素处理下内参基因的筛选
DOI: 10.15886/j.cnki.rdswxb.20240140
 CSTR: 32425.14.j.cnki.rdswxb.20240140
CSTR: 32425.14.j.cnki.rdswxb.20240140
Screening of internal reference genes of tomato under different plant parts and hormone treatments
-
摘要: 番茄(Solanum lycopersicum)是遗传学和分子生物学的研究模型。近年来,番茄内参基因的筛选已有较多的研究报道,但关于外源植物激素处理的种类较少,因此,本研究尝试筛选番茄在不同外源植物激素处理和不同组织部位中稳定表达的内参基因。为筛选番茄不同组织部位及植物激素处理下稳定表达的内参基因,以DANJ、EF-1α、ACT、UBI、APT、CAC、TIP41和 RPL8作为候选内参基因,在生长素、赤霉素、脱落酸、细胞分裂素、水杨酸、油菜素内酯、乙烯和不同组织部位8种试验条件下,利用geNorm、NormFinder、BestKeeper、Delta CT和RefFinder等算法对候选内参基因的稳定性进行综合评估,并通过生长素响应基因SlGH3.4对内参基因的稳定性进行验证。结果显示,APT是生长素、赤霉素、脱落酸、油菜素内酯处理和不同组织部位中表达最稳定的内参基因,UBI是全部样品中、细胞分裂素和水杨酸处理下最稳定的内参基因;TIP41是乙烯处理下表达最稳定的内参基因。最后,以综合排名较稳定的APT作为内参基因时发现,SlGH3.4基因的表达水平在生长素(IAA)处理条件下显示出相似的趋势,而稳定性较差的RPL8基因未能对目的基因的表达量进行准确校正。该研究结果可为后续番茄响应外源激素处理的基因表达网络分析和分子调控机制研究提供理论支撑。Abstract: Solanum lycopersicum is a model plant for research in genetics and molecular biology. Recently selection of tomato internal reference genes has been reported but with less types of exogenous hormones used for treatment. An attempt was hence made to select internal reference genes with stable expression in different tissues of tomato and under treatment with plant hormones. DANJ, EF-1α, ACT, UBI, APT, CAC, TIP41 and RPL8 were used as candidate internal reference genes, and selected under the 8 experimental conditions: exogenous hormone treatment with auxin, gibberellin, abscisic acid, cytokinin, salicylic acid, brassinolide and ethylene and various plant parts. The stability of the candidate reference genes was comprehensively evaluated using algorithms such as geNorm, NormFinder, BestKeeper, Delta CT and RefFinder. The stability of the reference genes was validated using the auxin-responsive gene SlGH3.4. The results showed that APT is the most stable reference gene expressed in auxin, gibberellin, abscisic acid, brassinosteroid treatment and various plant parts, that UBI is the most stable reference gene expressed in all samples under cytokinin and salicylic acid treatment, and that TIP41 is the most stable reference gene expressed under ethylene treatment. Finally, when APT, which has a relatively stable comprehensive ranking, was used as an internal reference gene, it was found that the expression level of SlGH3.4 gene showed a similar trend under IAA treatment conditions, while the less stable RPL8 gene failed to accurately correct the expression level of the target gene. All these results may provide theoretical support for the analysis of gene expression networks and molecular regulatory mechanisms in the response of tomato to exogenous hormone treatment.
-
Key words:
- tomato /
- hormone /
- internal reference genes /
- qRT-PCR /
- gene expression analysis
-
图 3 所有样品中8个候选内参基因的Ct值
Fig. 3 Ct values of 8 candidate reference genes in all the samples
图 4 geNorm分析不同处理下番茄幼苗中候选内参基因的稳定性
A.生长素处理;B.赤霉素处理;C.脱落酸处理;D.细胞分裂素处理;E.水杨酸处理;F.油菜素内酯处理;G.乙烯处理;H.不同组织;I.所有样品。
Fig. 4 geNorm analysis of stability of candidate internal reference genes in tomato seedlings under different treatments
A. Auxin treatment; B. Gibberellin treatment; C. Abscisic acid treatment; D. Cytokinin treatment; E. Salicylic acid treatment; F. Brassinosteroid treatment; G. Ethylene treatment; H. Different plant parts; I. All the samples.
图 5 NormFinder分析不同处理下番茄幼苗中候选内参基因的稳定性
A.生长素处理;B.赤霉素处理;C.脱落酸处理;D.细胞分裂素处理;E.水杨酸处理;F.油菜素内酯处理;G.乙烯处理;H.不同组织;I.所有样品。
Fig. 5 NormFinder analysis of stability of candidate internal reference genes in tomato seedlings under different treatments
A. Auxin treatment; B. Gibberellin treatment; C. Abscisic acid treatment; D. Cytokinin treatment; E. Salicylic acid treatment; F. Brassinosteroid treatment; G. Ethylene treatment; H. Different plant parts; I. All the samples.
图 6 Delta CT 分析结果
A.生长素处理;B.赤霉素处理;C.脱落酸处理;D.细胞分裂素处理;E.水杨酸处理;F.油菜素内酯处理;G.乙烯处理;H.不同组织;I.所有样品。
Fig. 6 Delta CT analysis results
A. Auxin treatment; B. Gibberellin treatment; C. Abscisic acid treatment; D. Cytokinin treatment; E. Salicylic acid treatment; F. Brassinosteroid treatment; G. Ethylene treatment; H. Different plant parts; I. All the samples.
图 7 外源IAA诱导下SlGH3.4基因的表达模式
注:不同字母表示同一内参基因校正下SlGH3.4基因表达的差异显著性(P<0.05)。
Fig. 7 Expression patterns of SlGH3.4 gene induced by exogenous IAA
Note: Different letters indicate significant differences in SlGH3.4 gene expression under the same reference gene correction (P<0.05).
表 1 候选内参基因引物序列
Table 1 Primer sequences of candidate reference genes
基因
Gene基因ID
Gene ID引物序列5′—3′(F and R)
Primer sequence参考文献
ReferencesDNAJ Solyc04g081530 GAGCACACATTGAGCCTTGAC [19] CTTTGGTACATCGGCATTCC EF-1α Solyc05g693059 GATTGACAGACGTTCTGGTAAGGA [20] ACCGGCATCACCATTCTTCA ACT Solyc03g078400 GTCCTCTTCCAGCCATCCAT [20] ACCACTGAGCACAATGTTACCG UBI Solyc11g056940 TCGTAAGGAGTGCCCTAATGCTGA [21] CAATCGCCTCCAGCCTTGTTGTAA APT Solyc04g077970 GAACAGACAAGATTGAGATGCATGTA [20] CCACGAGGGCACGTTCA CAC Solyc08g006960 CCTCCGTTGTGATGTAACTGG [22] ATTGGTGGAAAGTAACATCATCG TIP41 Solyc10g049850 ATGGAGTTTTTGAGTCTTCTGC [19] GCTGCGTTTCTGGCTTAGG RPL8 Solyc10g006580 CCGAAGGAGCTGTTGTTTGTA [22] ACCTGACCAATCATAGCACGA  下载: 导出CSV
下载: 导出CSV
表 2 BestKeeper分析结果
Table 2 BestKeeper analysis results
生长素 Auxin 赤霉素 Gibberellin 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.86 3.68 0.311 CAC 0.73 3.07 0.001 1 CAC 1.06 4.52 0.001 TIP41 1.10 4.66 0.668 2 APT 1.11 5.39 0.778 APT 1.93 8.73 0.840 3 UBI 1.94 9.36 0.924 ACT 1.96 9.29 0.788 4 EF-1α 2.06 10.65 0.942 UBI 2.33 10.85 0.845 5 ACT 2.24 10.66 0.433 EF-1α 2.69 13.32 0.857 6 DNAJ 2.75 12.47 0.985 DNAJ 2.82 13.07 0.994 7 RPL8 4.35 19.59 0.180 RPL8 4.73 21.43 0.593 8 细胞分裂素 Cytokinin 脱落酸 Abscisic acid 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.52 2.12 0.001 ACT 0.32 1.60 0.652 1 UBI 0.62 2.87 0.299 CAC 0.84 3.46 0.001 2 CAC 1.35 5.79 0.001 TIP41 1.08 4.68 0.001 3 APT 1.36 6.21 0.528 APT 1.49 6.68 0.999 4 EF-1α 1.81 8.95 0.948 UBI 1.90 8.72 0.852 5 ACT 2.11 9.61 0.422 DNAJ 2.09 9.81 0.691 6 DNAJ 2.95 12.61 0.989 EF-1α 3.06 14.77 0.996 7 RPL8 3.81 16.59 0.312 RPL8 3.82 16.68 0.801 8
下载: 导出CSV
2 续表2 Tab.2 Continued
油菜素内酯 Brassinolide 乙烯 Ethylene 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r TIP41 0.78 3.57 0.001 TIP41 0.41 1.83 0.001 1 CAC 1.47 6.57 0.001 UBI 0.51 2.43 0.001 2 APT 1.77 7.82 0.986 ACT 0.94 4.70 0.416 3 UBI 2.04 9.57 0.906 CAC 1.50 6.75 0.001 4 DNAJ 2.49 11.83 0.512 RPL8 1.75 7.84 0.345 5 ACT 2.60 12.06 0.267 EF-1α 1.77 8.54 0.910 6 EF-1α 3.34 16.02 0.988 APT 2.44 10.62 0.785 7 RPL8 4.28 19.29 0.739 DNAJ 2.90 13.08 0.598 8 水杨酸 Salicylic acid 不同组织 Different tissues 排名 Rank 基因 Gene SD CV r 基因 Gene SD CV r CAC 1.43 6.07 0.001 APT 1.23 5.03 0.991 1 TIP41 1.65 7.04 0.001 CAC 1.32 5.87 0.001 2 UBI 1.67 7.69 0.984 ACT 1.83 9.13 0.712 3 APT 1.94 8.62 0.987 UBI 1.94 8.72 0.926 4 ACT 1.94 8.90 0.420 TIP41 2.21 9.87 0.770 5 DNAJ 2.31 10.96 0.765 EF-1α 2.28 11.92 0.964 6 EF-1α 2.56 12.64 0.931 RPL8 2.91 13.70 0.541 7 RPL8 3.79 17.38 0.754 DNAJ 3.69 16.03 0.998 8 所有样品 All Samples 排名 Rank 基因 Gene SD CV r TIP41 1.17 5.01 0.194 1 CAC 1.37 5.92 0.001 2 UBI 1.68 7.80 0.839 3 ACT 1.82 8.68 0.466 4 APT 1.88 8.46 0.803 5 EF-1α 2.50 12.41 0.894 6 DNAJ 2.86 13.03 0.782 7 RPL8 3.68 16.56 0.550 8 注:所有样品表示不同组织和不同外源植物激素处理下的样品。
Note: All samples represent those under different plant parts and different exogenous plant hormone treatments.
下载: 导出CSV
表 3 候选内参基因表达稳定性综合分析结果
Table 3 Comprehensive analysis results of the expression stability of candidate reference genes
处理
Treatment综合排名
Comprehensive ranking排序
Sort处理
Treatment综合排名
Comprehensive ranking排序
Sort生长素
AuxinAPT 1 油菜素内酯
BrassinolideAPT 1 UBI 2 UBI 2 EF-1α 3 TIP41 3 TIP41 4 EF-1α 4 CAC 5 DNAJ 5 DNAJ 6 CAC 6 ACT 7 ACT 7 RPL8 8 RPL8 8 赤霉素
GibberellinAPT 1 乙烯
EthyleneTIP41 1 ACT 2 UBI 2 TIP41 3 ACT 3 CAC 4 EF-1α 4 DNAJ 5 CAC 5 EF-1α 6 RPL8 6 UBI 7 APT 7 RPL8 8 DNAJ 8 细胞分裂素
CytokininUBI 1 水杨酸
Salicylic acidUBI 1 TIP41 2 APT 2 APT 3 EF-1α 3 EF-1α 4 DNAJ 4 ACT 5 TIP41 5 CAC 6 CAC 6 DNAJ 7 ACT 7 RPL8 8 RPL8 8 脱落酸
Abscisic acidAPT 1 不同组织
Different tissuesAPT 1 ACT 2 TIP41 2 UBI 3 EF-1α 3 DNAJ 4 UBI 4 CAC 5 ACT 5 TIP41 6 CAC 6 EF-1α 7 DNAJ 7 RPL8 8 RPL8 8 所有样品
All SamplesUBI 1 APT 2 EF-1α 3 TIP41 4 ACT 5 CAC 6 DNAJ 7 RPL8 8 注:所有样品表示不同组织和不同外源植物激素处理下的样品。 Note: All samples represent those under different plant parts and different exogenous plant hormone treatments.
下载: 导出CSV
-
[1] BUSTIN S A, BENES V, NOLAN T, et al. Quantitative real-time RT-PCR-a perspective [J]. Journal of Molecular Endocrinology, 2005, 34(3): 597−601. https://doi.org/10.1677/jme.1.01755 doi: 10.1677/jme.1.01755 [2] GACHON C, MINGAM A, CHARRIER B. Real-time PCR: what relevance to plant studies? [J]. Journal of Experimental Botany, 2004, 55(402): 1445−1454. https://doi.org/10.1093/jxb/erh181 doi: 10.1093/jxb/erh181 [3] 张玉芳, 赵丽娟, 曾幼玲. 基因表达研究中内参基因的选择与应用[J]. 植物生理学报, 2014, 50(8): 1119−1125. https://doi.org/10.13592/j.cnki.ppj.2014.0201 doi: 10.13592/j.cnki.ppj.2014.0201 [4] 蒋婷婷, 高燕会, 童再康. 石蒜属植物实时荧光定量PCR内参基因的选择[J]. 园艺学报, 2015, 42(6): 1129−1138. https://doi.org/10.16420/j.issn.0513-353x.2014-0999 doi: 10.16420/j.issn.0513-353x.2014-0999 [5] MÜLLER O A, GRAU J, THIEME S, et al. Genome-wide identification and validation of reference genes in infected tomato leaves for quantitative RT-PCR analyses [J]. PLoS One, 2015, 10(8): e0136499. https://doi.org/10.1371/journal.pone.0136499 doi: 10.1371/journal.pone.0136499 [6] ZHANG Y T, ZHU L J, XUE J Y, et al. Selection and verification of appropriate reference genes for expression normalization in Cryptomeria fortunei under abiotic stress and hormone treatments [J]. Genes, 2021, 12(6): 791. https://doi.org/10.3390/genes12060791 doi: 10.3390/genes12060791 [7] 李铁铮, 王金铃, 刘晓, 等. 管花肉苁蓉实时荧光定量PCR分析中内参基因的选择和验证[J]. 植物生理学报, 2021, 57(4): 969−981. https://doi.org/10.13592/j.cnki.ppj.2019.0440 doi: 10.13592/j.cnki.ppj.2019.0440 [8] HU J H, ISRAELI A, ORI N, et al. The interaction between DELLA and ARF/IAA mediates crosstalk between gibberellin and auxin signaling to control fruit initiation in tomato [J]. The Plant Cell, 2018, 30(8): 1710−1728. https://doi.org/10.1105/tpc.18.00363 doi: 10.1105/tpc.18.00363 [9] KONG X P, ZHANG C L, ZHENG H H, et al. Antagonistic interaction between auxin and SA signaling pathways regulates bacterial infection through lateral root in Arabidopsis [J]. Cell Reports, 2020, 32(8): 108060. https://doi.org/10.1016/j.celrep.2020.108060 doi: 10.1016/j.celrep.2020.108060 [10] GAO J, CHEN H, YANG H F, et al. A brassinosteroid responsive miRNA-target module regulates gibberellin biosynthesis and plant development [J]. New Phytologist, 2018, 220(2): 488−501. https://doi.org/10.1111/nph.15331 doi: 10.1111/nph.15331 [11] 董汉松. 植物抗病防卫基因表达调控与诱导抗性遗传的机制[J]. 植物病理学报, 1996(4): 2−6. https://doi.org/10.13926/j.cnki.apps.1996.04.001 doi: 10.13926/j.cnki.apps.1996.04.001 [12] XU P P, FANG S, CHEN H Y, et al. The brassinosteroid-responsive xyloglucan endotransglucosylase/hydrolase 19 (XTH19) and XTH23 genes are involved in lateral root development under salt stress in Arabidopsis [J]. The Plant Journal, 2020, 104(1): 59−75. https://doi.org/10.1111/tpj.14905 doi: 10.1111/tpj.14905 [13] 白圣懿, 王晓敏, 刘文娟, 等. 不同激素处理下番茄实时荧光定量聚合酶链反应内参基因的筛选[J]. 浙江大学学报(农业与生命科学版), 2023, 49(1): 31−44. https://doi.org/10.3785/j.issn.1008-9209.2022.03.081 doi: 10.3785/j.issn.1008-9209.2022.03.081 [14] 韩晓雪, 韩佳轩, 姜晶. 番茄在非生物胁迫下实时定量RT-PCR中内参基因的筛选[J]. 分子植物育种, 2015, 13(4): 822−831. https://doi.org/10.13271/j.mpb.013.000822 doi: 10.13271/j.mpb.013.000822 [15] OMENA-GARCIA R P, OLIVEIRA MARTINS A, MEDEIROS D B, et al. Growth and metabolic adjustments in response to gibberellin deficiency in drought stressed tomato plants [J]. Environmental and Experimental Botany, 2019, 159: 95−107. https://doi.org/10.1016/j.envexpbot.2018.12.011 doi: 10.1016/j.envexpbot.2018.12.011 [16] HU E M, LIU M, ZHOU R, et al. Relationship between melatonin and abscisic acid in response to salt stress of tomato [J]. Scientia Horticulturae, 2021, 285: 110176. https://doi.org/10.1016/J.SCIENTA.2021.110176 doi: 10.1016/J.SCIENTA.2021.110176 [17] LU J Z, GUAN P X, GU J M, et al. Exogenous DA-6 improves the low night temperature tolerance of tomato through regulating cytokinin [J]. Frontiers in Plant Science, 2021, 11: 599111. https://doi.org/10.3389/fpls.2020.599111 doi: 10.3389/fpls.2020.599111 [18] ALBUQUERQUE G M R, FONSECA F C A, BOITEUX L S, et al. Stability analysis of reference genes for RT-qPCR assays involving compatible and incompatible Ralstonia solanacearum-tomato 'Hawaii 7996' interactions [J]. Scientific Reports, 2021, 11(1): 18719. https://doi.org/10.1038/s41598-021-97854-8 doi: 10.1038/s41598-021-97854-8 [19] EXPÓSITO-RODRÍGUEZ M, BORGES A A, BORGES-PÉREZ A, et al. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process [J]. BMC Plant Biology, 2008, 8(1): 131. https://doi.org/10.1186/1471-2229-8-131 doi: 10.1186/1471-2229-8-131 [20] ALFENAS-ZERBINI P, MAIA I G, FÁVARO R D, et al. Genome-wide analysis of differentially expressed genes during the early stages of tomato infection by a potyvirus [J]. Molecular Plant-Microbe Interactions, 2009, 22(3): 352−361. https://doi.org/10.1094/MPMI-22-3-0352 doi: 10.1094/MPMI-22-3-0352 [21] MASCIA T, SANTOVITO E, GALLITELLI D, et al. Evaluation of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in infected tomato plants [J]. Molecular Plant Pathology, 2010, 11(6): 805−816. https://doi.org/10.1111/j.1364-3703.2010.00646.x doi: 10.1111/j.1364-3703.2010.00646.x [22] CHOI S W, HOSHIKAWA K, FUJITA S, et al. Evaluation of internal control genes for quantitative Realtime PCR analyses for studying fruit development of dwarf tomato cultivar 'Micro-Tom' [J]. Plant Biotechnology, 2018, 35(3): 225−235. https://doi.org/10.5511/plantbiotechnology.18.0525a doi: 10.5511/plantbiotechnology.18.0525a [23] CHEN X, CHEN J D, LIAO D H, et al. Auxin-mediated regulation of arbuscular mycorrhizal symbiosis: a role of SlGH3.4 in tomato [J]. Plant Cell & Environment, 2022, 45(3): 955−968. https://doi.org/10.1111/pce.14210 doi: 10.1111/pce.14210 [24] VANDESOMPELE J, DE PRETER K, PATTYN F, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes [J]. Genome Biology, 2002, 3(7): research0034.1. https://doi.org/10.1186/gb-2002-3-7-research0034 doi: 10.1186/gb-2002-3-7-research0034 [25] ANDERSEN C L, JENSEN J L, ØRNTOFT T F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets [J]. Cancer Research, 2004, 64(15): 5245−5250. https://doi.org/10.1158/0008-5472.CAN-04-0496 doi: 10.1158/0008-5472.CAN-04-0496 [26] PFAFFL M W, TICHOPAD A, PRGOMET C, et al. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations [J]. Biotechnology Letters, 2004, 26(6): 509−515. https://doi.org/10.1023/B:BILE.0000019559.84305.47 doi: 10.1023/B:BILE.0000019559.84305.47 [27] 吴悠, 冀宏宇, 邓淑雯, 等. 三褶虾脊兰花发育相关基因RT-qPCR内参基因的筛选[J]. 园艺学报, 2024, 51(9): 2063−2074. https://doi.org/10.16420/j.issn.0513-353x.2023-0954 doi: 10.16420/j.issn.0513-353x.2023-0954 [28] 李佳妮, 张姝, 张永杰. 布莱克韦尔虫草实时荧光定量逆转录PCR内参基因的筛选[J]. 微生物学通报, 2025, 52(1): 219−229. https://doi.org/10.13344/j.microbiol.china.240343 doi: 10.13344/j.microbiol.china.240343 [29] 颜爽爽, 邱正坤, 余炳伟, 等. 植物生长素响应高温胁迫研究进展[J]. 园艺学报, 2020, 47(11): 2238−2246. https://doi.org/10.16420/j.issn.0513-353x.2020-0264 doi: 10.16420/j.issn.0513-353x.2020-0264 [30] 园园, 恩和巴雅尔, 齐艳华. 植物GH3基因家族生物学功能研究进展[J]. 植物学报, 2023, 58(5): 770−782. https://doi.org/10.11983/CBB22263 doi: 10.11983/CBB22263 [31] CARISEY A F, MACE E M, SAEED M B, et al. Nanoscale dynamism of actin enables secretory function in cytolytic cells [J]. Current Biology, 2018, 28(4): 489-502. e9. [32] CITRI A, PANG Z P, SÜDHOF T C, et al. Comprehensive qPCR profiling of gene expression in single neuronal cells [J]. Nature Protocols, 2011, 7(1): 118−127. https://doi.org/10.1038/nprot.2011.430 doi: 10.1038/nprot.2011.430 [33] 宋雄. 欧芹不同逆境条件下适宜内参基因的筛选[D]. 南京: 南京农业大学, 2016. [34] 黄丽萍, 李思鸿, 钟启文, 等. 香瓜茄多组织部位和病害胁迫条件下qRT-PCR内参基因的选择[J]. 青海大学学报, 2022, 40(3): 33−40. https://doi.org/10.13901/j.cnki.qhwxxbzk.2022.03.006 doi: 10.13901/j.cnki.qhwxxbzk.2022.03.006 [35] 岳炜楠, 胥通玉, 苗佳敏, 等. 紫花苜蓿不同组织及不同激素处理下内参基因的筛选[J/OL]. 草原与草坪, 1-16[2024-06-20]. http://kns.cnki.net/kcms/detail/62.1156.S.20240521.1243.002.html. [36] 徐圆圆, 赵国春, 郝颖颖, 等. 无患子RT-qPCR内参基因的筛选与验证[J]. 生物技术通报, 2022, 38(10): 80−89. https://doi.org/10.13560/j.cnki.biotech.bull.1985.2021-1616 doi: 10.13560/j.cnki.biotech.bull.1985.2021-1616 [37] 巨秀婷, 何金娣, 张梦洁, 等. 郁金香不同组织内参基因筛选及稳定表达分析[J]. 南方农业学报, 2023, 54(11): 3174−3185. https://doi.org/10.3969/j.issn.2095-1191.2023.11.005 doi: 10.3969/j.issn.2095-1191.2023.11.005 [38] LIAO D H, CHEN X, CHEN A Q, et al. The characterization of six auxin-induced tomato GH3 genes uncovers a member, SlGH3.4, strongly responsive to arbuscular mycorrhizal symbiosis [J]. Plant and Cell Physiology, 2015, 56(4): 674−687. https://doi.org/10.1093/pcp/pcu212 doi: 10.1093/pcp/pcu212 [39] LIU H L, LIU J, CHEN P, et al. Selection and validation of optimal RT-qPCR reference genes for the normalization of gene expression under different experimental conditions in Lindera megaphylla [J]. Plants, 2023, 12(11): 2185. https://doi.org/10.3390/plants12112185 doi: 10.3390/plants12112185 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1362
- HTML全文浏览量: 835
- PDF下载量: 90
- 被引次数: 0
