





-
主持人:郭志强
琼枝藻(Betaphycus gelatinae)俗称琼枝麒麟菜,隶属于红藻门、真红藻纲、杉藻目、红翎菜科、琼枝藻属,主要分布于中国、菲律宾、日本、印度尼西亚等地[1-2]。琼枝藻在中国自然分布于海南岛、东沙群岛和台湾岛等热带和亚热带海区[1],主要养殖栽培区域为海南岛周边海域[3],是当前中国麒麟菜养殖业的主要品种之一。琼枝藻多呈紫红或黄绿色,藻体表面光滑,具不规则对生、互生或叉状分枝,偶有羽状分枝,枝近于扁平,腹面常具有疣状或圆锥状突起[4]。琼枝藻是提取卡拉胶的优质原料,含胶量高达30%[5],被广泛应用到食品、日化、医药等领域[5-11]。近年来,受养殖空间的限制,我国麒麟菜的产量处于一个相对较低的水平,2019年仅420 t[12];而受人类活动和自然水域环境变化的影响,我国麒麟菜的自然资源也呈现出显著萎缩趋势。
琼枝藻的生活史属于同型世代类型,其有性繁殖需经过孢子体、配子体和果孢子体3个交替的生活史阶段[13];而无性繁殖则为营养繁殖,能够通过组织的培养产生与其亲本在基因型上几乎完全一致的新个体。在养殖生产过程中,琼枝藻与异枝江蓠(Gracilaria bailinae)、龙须菜(Gracilariopsis lemaneiformis)、长心卡帕藻(Kappaphycus alvarezii)等红藻类似,主要靠营养繁殖的方式进行扩繁和增殖,即通过多次“分苗”的方式获得生产所需的苗种,并直接在养殖海域中进行连续栽培。这种生产模式基本没有苗种培育的环节,存在着种质来源不稳定、质量退化等一系列问题。因此,琼枝藻的苗种技术是当前影响养殖产业发展的重要因素之一。利用组织培养的方法进行快速繁殖或种质保存是高等植物研究中常用的技术手段[14-20],组培技术在大型海藻中的应用已经在条斑紫菜(Porphyra yezoensis)[21]、羊栖菜(Sargassum fusiforme)[22]、长心卡帕藻[23]、海带(Laminaria japonica)[24]、异枝江蓠[25]等品种上有所报道,主要体现在藻体不同部位、组织大小、消毒方式和培养条件对苗种培育的效果上,并取得了较好的效果和积累了一定的经验。目前,有关琼枝藻的研究多集中在环境因子对其生长生理的影响[26-30]、卡拉胶及多糖的提取[31-34]、营养成分分析[10]等,而有关组织繁殖方面的研究鲜有报道。本研究通过开展琼枝藻不同切段部位和切段长度的组织在不同温度条件下的培养实验,比较出芽和生长情况,旨在探索琼枝藻组织切段培养的适宜条件,为琼枝藻的种质保存和快速扩繁提供依据。
-
琼枝藻取自海南省海洋与渔业科学院琼海科研基地(110°40′07.2″E,19°21′58.2″N),挑选颜色正常、生长旺盛的藻体洗净,自然海水中暂养7 d后开始实验。暂养温度24 ℃,盐度28,光照强度5 000 lx,光周期为光照(L):黑暗(D)=12 h∶12 h。
-
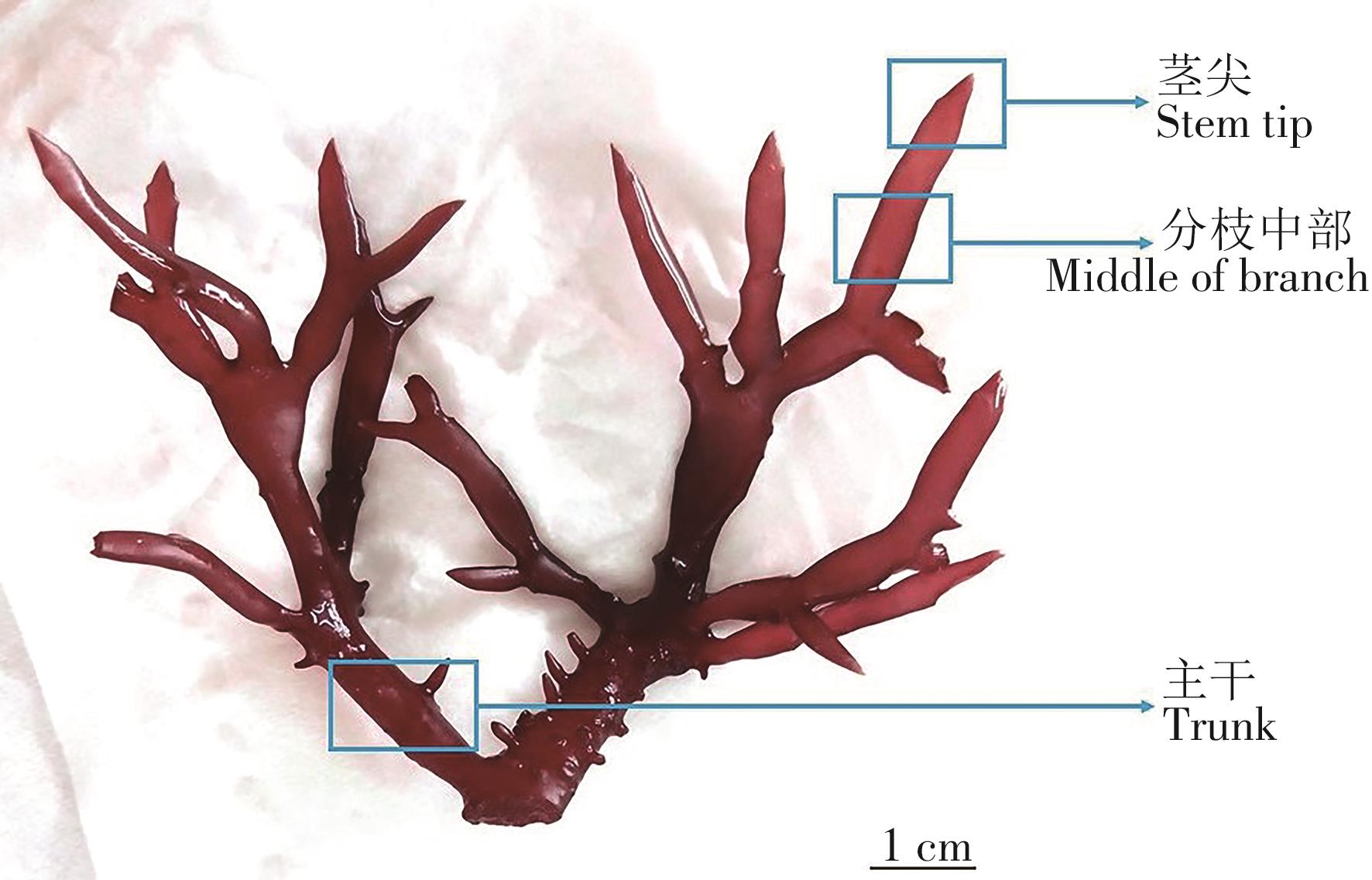
采用双因子实验设计,2个自变量为切段部位(茎尖、分枝中部和主干)(图1)和切段长度(5 mm、10 mm和15 mm),因变量为增重率、个体出芽数和出芽密度。3个切段部位,3个切段长度,共设置9个处理组,每个处理组设3个重复,每个重复10个切段。各处理组置于智能型光照培养箱(GXZ-300D,宁波江南仪器厂,宁波)中进行恒温培养,温度、盐度、光照强度和光周期与暂养条件相同。实验所用培养液为自然海水砂滤后,经高温高压灭菌制备。

图 1 琼枝藻形态结构及切段部位
Figure 1. The morphological structure and cutting section of B. gelatinae
-
选择藻体的分枝中部进行培养,切段长度为10 mm,培养温度设置为21、24、27、30、33 ℃,共5个处理组,每个处理组设3个重复,每个重复10个切段。各处理组置于智能型光照培养箱中进行恒温培养,盐度、光照强度和光周期与暂养条件相同。实验所用培养液为自然海水砂滤后,经高温高压灭菌制备。
-
实验第1天,对各实验组切段的发芽数和鲜质量进行统计和称量;切段培养第42天,统计各实验组的出芽总数、存活切段总数、出芽切段数和鲜质量。
按照下列公式进行计算:
个体出芽数(个)=出芽总数/存活切段总数,
成活率=存活切段总数/切段总数×100%,
出芽率=出芽切段数/存活切段总数×100% ,
出芽密度(个·mm-1)=个体出芽数/切段长度,
增重率=(最终鲜质量-初始鲜质量)/初始鲜质量×100% 。
-
利用Excel 2019和SPSS 18.0软件进行数据处理及统计分析。采用双因子方差分析(Two-way ANOVA)检验2个自变量(切段部位和长度)对生长和出芽的影响;运用Duncan多重比较检验不同处理间的差异性(P<0.05)。
-
琼枝藻不同部位及长度的切段下端均无出芽,表现出明显的极性;培养42 d后,茎尖伸长生长明显,芽体从两侧出芽为主;分枝中部和主干部位则无明显伸长生长,而是上端切面膨大,芽体从切面皮层出芽生长,数量多且集中;藻体切段也由实验初期的紫红色转变为绿色(图2)。

图 2 琼枝藻切段培养42 d后的形态结构图
Figure 2. The morphological structure of B. gelatinae cultured for 42 days.
-
双因子方差分析结果显示(表1),琼枝藻切段培养的增重率受切段部位影响极显著(F=146.425,P<0.01);同时,切段长度同样影响着琼枝藻体切段的增重率(F=6.170,P<0.01);但琼枝藻切段部位与长度之间没有显著的互作效应(F=2.677,P=0.065)。
表 1 琼枝藻切段部位和长度对增重率影响的双因子方差分析结果
Table 1. Results from the 2-way ANOVA examining the effect of B. gelatinae cutting section and its length on the weight gain rate.
变异源Source of variations df 均方差MS F P 切段部位Cutting section 2 3 497.028 146.425 0.000 切段长度Cutting length 2 147.348 6.170 0.009 部位×长度Section × length 4 63.922 2.677 0.065 总计Total 27 切段部位和切段长度对琼枝藻切段培养的成活率(均为100%)和出芽率(均为100%)均无影响。在体质量变化方面,茎尖切段的增重率显著高于分枝中部和主干切段(P<0.05),说明茎尖的伸长生长量大于分枝中部和主干切段芽体生长的质量;同一切段部位,不同切段长度的增重率在总体上无显著差异(P>0.05)(图3)。

图 3 不同切段长度和切段部位的琼枝藻增重率
Figure 3. The weight gain rate at different cutting length and cutting section of B. gelatinae.
-
双因子方差分析结果显示(表2),切段的部位和长度均显著影响着切段的个体出芽数(部位:F=156.253,P<0.01;长度:F=59.589,P<0.01);同时,切段的个体出芽数也显著受到切段部位和长度的互作影响(F=11.131,P<0.01)。
表 2 琼枝藻切段部位和长度对个体出芽数影响的双因子方差分析结果
Table 2. Results from the 2-way ANOVA examining the effect of B. gelatinae cutting section and its length on the number of buddings.
变异源Source of variations df 均方差MS F P 切段部位Cutting section 2 111.634 156.253 0.000 切段长度Cutting length 2 42.573 59.589 0.000 部位×长度Section × length 4 7.953 11.131 0.000 总计Total 27 切段部位和长度对个体出芽数的影响如图4所示,同一切段长度,不同切段部位的出芽数由高到低排列,依次为主干、分枝中部和茎尖。Duncan多重比较结果显示,分枝中部和主干的个体出芽数均显著高于茎尖(P<0.05);5 mm和15 mm组切段中的分枝中部和主干的个体出芽数无显著差异(P>0.05)。同一切段部位,个体随着切段长度的增加,个体出芽数均呈现一定的增长趋势,15 mm组的个体出芽数分别为茎尖(5.90 ± 0.67)个、分枝中部(10.16 ± 1.14)个和主干(11.90 ± 0.88)个。Duncan多重比较结果显示,茎尖切段不同长度间的个体出芽数差异显著(P<0.05);分枝中部切段5和10 mm组显著低于15 mm组(P<0.05);主干切段10和15 mm组则显著高于5 mm组(P<0.05)。

图 4 不同切段长度和切段部位的琼枝藻个体出芽数
Figure 4. The number of buddings at different cutting length and cutting section of B. gelatinae.
切段长度相同,不同切段部位的出芽密度变化与个体出芽数一致。切段部位相同,不同切段长度的出芽密度变化与个体出芽数呈相反的趋势。切段长度越长,单位长度内的出芽个数越少,5 mm组出芽密度总体上显著高于其他各组(P<0.05),具有一定的出芽优势(图5)。

图 5 不同切段长度和切段部位的琼枝藻切段出芽密度
Figure 5. The budding density at different cutting length and cutting section of B. gelatinae.
-
实验范围内,琼枝藻藻体切段生长适温为21 ~ 27 ℃,在该温度范围内,藻体切段的成活率和出芽率均达到100%,且随温度的增加,个体出芽总数逐渐增加,个体出芽总数为(4.50 ± 0.16)~(9.16 ± 0.28)个;而30 ℃组,其存活率虽为100%,但出芽率仅为66.66%,个体出芽总数较低,在实验第15 d后,切段出现轻微腐烂;33 ℃组,在实验第15 d,藻体切段均腐烂死亡,成活率为0。因此,琼枝藻藻体切段最适生长温度为27 ℃,个体出芽总数为(9.16 ± 0.28)个,且与其他处理组差异显著(P<0.05)(表3)。
表 3 温度对琼枝藻藻体切段出芽和生长的影响
Table 3. Effects of temperature on budding and growth of B. gelatinae.
温度/℃Temperature 成活率/%Survival rate 出芽率/%Germination rate 个体出芽数/个 Number of buddings 上端 侧面 下端 总数 21 100 100 4.50 ± 0.16a 0 0 4.50 ± 0.16a 24 100 100 7.40 ± 0.37b 0 0 7.40 ± 0.37b 27 100 100 9.16 ± 0.28c 0 0 9.16 ± 0.28c 30 100 66.66 3.13 ± 0.33d 0 0 3.13 ± 0.33d 33 0 - - - - - 注: 33 ℃时切段成活率为0,相关出芽指标无法测得,所以用“-”表示。Note: The survival rate was 0 when the temperature at 33 ℃, so used “-” indicated no detected to the relevant budding index. -
在大型海藻组织培养中,外植体产生愈伤组织的能力或再生能力与材料所选取的部位有明确的关系。本研究所选取的茎尖、分枝中部和主干3个部位的实验结果证明,选择主干部位作为切段培养的材料是最佳选择,主干部位具有更高的增殖潜力,这与早年尤丽莉[35]开展的龙须菜切段培养实验结果一致,而陈海红等[25]近年来通过对异枝江蓠的切段培养对这一结论进行了重新印证。茎尖切段主要以切段的伸长生长为主,重量变化明显,增重率显著大于分枝中部和主干,而发芽较少,这可能是相关激素调节的结果,并未能形成愈伤组织,而是促使其向上伸长生长。
在实验中发现,分枝中部和主干部位藻体的上端切面均明显膨大,芽体从切面皮层出芽生长。而在长心卡帕藻切段培养中,其新生芽的发生主要集中在切面髓部,而不是切面皮层[23]。愈伤组织形成位置的差异与不同部位细胞的分生能力有关,琼枝藻藻体切面皮层内侧富含薄壁细胞,分生能力较强,利于形成再生芽;长心卡帕藻藻体切面的髓部细胞分生能力则较强;而是否存在海藻种间生长调节机制差异的影响,尚需进一步研究。本实验中,所有藻体切段的出芽均在上端面和侧面,下端切面无芽生成,表现出明显的极性,而陈昌生等[36]在对5种江蓠切段的研究中也发现了此现象,均存在上端生长优势,上端切面利于分化增殖,产生新芽,进而形成新枝。但在对粗枝软骨藻(Chondria crassicaulis)的研究中却发现上下端切面均可形成芽[37],因而海藻切段的增值分化,存在着一定的种间差异。
组织培养切段不仅受切段部位的影响,切段长度同样影响培养的效果。有研究发现羊栖菜1 mm以内的组织碎片出芽生苗较好[22];唐贤明等[23]在对长心卡帕藻的研究中也发现短的切段有助于提高出芽的数量;梅俊学等[21]认为藻体出苗与切块大小有关,切块小,则表面积大,更有利于新陈代谢,进而使更多的细胞转化成苗。在本研究中,各个长度切段的成活率和出芽率均为100%,切段长度并不影响其成活率和出芽率,但随着切段长度的增加,个体出芽数也随之增加。笔者认为,在不添加外源激素的条件下,琼枝藻的发芽和生长只能通过内源激素调节,内源激素的含量可能影响着琼枝藻出芽生长的数量,因而激素调节可能对琼枝藻组织培养有着重要的作用。
不同植物体在长期的自然进化和环境适应中对所生存的环境有着不同的要求,研究显示琼枝藻27 ℃时生长最快,温度升高或降低均会影响其生长速率,长期低于18 ℃或高于36 ℃则会造成死亡[26]。组织切段培养诱导生芽需根据植物体所适宜生长的温度来进行适当调整培养温度,以诱导愈伤组织的形成和发生。本研究中,培养温度为27 ℃时切段的出芽情况最好,温度低于或高于此温度,均影响出芽数量;30 ℃时出芽率受到影响,33 ℃时切段无法成活,这与梁磊等[26]关于琼枝藻生长温度的研究基本一致,因此在对海藻组织切段培养温度的选择上可参考其生长温度。在对长心卡帕藻的切段培养研究中发现,最适生长的温度为20 ℃,高于或低于此温度均影响其生长[23];20℃也同样是鼠尾藻(Sargassum thunbergia)假根再生不定芽的最适温度[38];这些研究结果与琼枝藻的培养结果相似,而温度上的差异可能是海藻种间差异造成的。
-
琼枝藻在进行离体组织培养时,可优先考虑利用藻体主干部位进行培养,培养藻体长度为15 mm,培养温度为27 ℃,可达到较好的繁殖育苗效果。本研究结果可为琼枝藻的种质保存和快速扩繁提供数据支撑。
切段部位、长度和培养温度对琼枝藻繁殖的影响
DOI: 10.15886/j.cnki.rdswxb.20230102
 CSTR: 32425.14.j.cnki.rdswxb.20230102
CSTR: 32425.14.j.cnki.rdswxb.20230102
Effects of explant position, section length and culture temperature on the propagation of Betaphycus gelatinae
-
摘要: 为了探究琼枝藻(Betaphycus gelatinae)组织切段培养的适宜条件,以琼枝藻为材料,研究不同切段部位(茎尖、分枝中部和主干)和切段长度(5、10、15 mm)的外植体切段在不同培养温度(21、24、27、30、33 ℃)下的出芽和生长。不同切段部位和切段长度的成活率和出芽率均达100%。不同切段部位出芽数由大到小依次为主干、分枝中部和茎尖;茎尖以伸长生长为主,主干和分枝中部则以上端切面膨大发芽为主;芽体形成于上端切面皮层,下端切面无芽体生成,表现出明显的极性。不同切段长度出芽数由大到小依次为15 mm组、10 mm组和5 mm组,15 mm组的个体出芽数分别为茎尖(5.90 ± 0.67)个、分枝中部(10.16 ± 1.14)个、主干(11.90 ± 0.88)个;5 mm组出芽密度最高。琼枝藻切段培养适宜的生长温度为(21 ~ 27)℃,在此范围内,成活率和出芽率均为100%,且出芽数随温度的升高而增加;超过27 ℃,藻体切段的成活率和出芽率均下降。研究结果表明,琼枝藻最适组织切段培养部位为主干,长度为15 mm,培养温度为27 ℃。Abstract: Betaphycus gelatinae was cut into sections of different lengths from different explants and cultured at different temperatures to explore the suitable conditions for its tissue culture to determine the effects of different explants (stem tip, main branch and trunk), section lengths (5 mm, 10 mm and 15 mm) and culture temperatures (21 ℃, 24 ℃, 27 ℃, 30 ℃ and 33 ℃) on bud emergence and growth of the explants of B. gelatinae. The results showed that the survival rate and bud emergence rate of different explants and section lengths were all up to 100%. The number of buds in different explants was in the order of the trunk > the main branch > the stem tip. The stem tip mainly grew elongate, while the main branch and trunk were mainly expanded and formed buds. The buds were formed on the upper cut, not on the lower cut, showing obvious polarity. The number of the buds in the sections of different lengths was in the order of the sections 15 mm long > the sections 10 mm long > the sections 5 mm long. The number of individual buds in the sections 15 mm long was 5.90 ± 0.67 in the stem tip, 10.16 ± 1.14 in the main branch and 11.90 ± 0.88 in the trunk. The highest budding density was in the sections 5 mm long. The suitable growth temperature for tissue culture of B. gelatinae ranged from 21℃ to 27 ℃, under which the survival rate and bud emergence rate were both 100%, and the number of buds increased with the temperature. The survival rate and bud emergenc rate of the sections were decreased when the temperature was above 27 ℃. For optimal tissue culture B. gelatinae was cultured at 27 ℃ with the trunk sections 15 mm long as explants.
-
Key words:
- Betaphycus gelatinae /
- explant /
- tissue culture /
- bud proliferation /
- temperature
注释:1) 钟云芳 -
图 2 琼枝藻切段培养42 d后的形态结构图
a、b、c分别为茎尖的5 mm组、10 mm组和15 mm组;d、e、f分别为分枝中部的5 mm组、10 mm组和15 mm组;g、h、i分别为主干部位的5 mm组、10 mm组和15 mm组。
Fig. 2 The morphological structure of B. gelatinae cultured for 42 days.
a, b and c are 5 mm group, 10 mm group and 15 mm group in the stem tip, respectively. d, e and f are the 5 mm group, 10 mm group and 15 mm group in the middle of branch, respectively. g, h and i are 5 mm group, 10 mm group and 15 mm group in the trunk, respectively.
图 3 不同切段长度和切段部位的琼枝藻增重率
同一切段长度不同小写字母表示切段部位之间差异显著(P<0.05),同一切段部位不同大写字母表示切段长度之间差异显著(P<0.05)。图4和图5同此。
Fig. 3 The weight gain rate at different cutting length and cutting section of B. gelatinae.
The same cutting length of B. gelatinae, different lowercase letters indicate significant differences on the cutting section (P < 0.05). The same cutting section of B. gelatinae, different capital letters indicate significant differences (P < 0.05). The same as following figures.
图 4 不同切段长度和切段部位的琼枝藻个体出芽数
Fig. 4 The number of buddings at different cutting length and cutting section of B. gelatinae.
图 5 不同切段长度和切段部位的琼枝藻切段出芽密度
Fig. 5 The budding density at different cutting length and cutting section of B. gelatinae.
表 1 琼枝藻切段部位和长度对增重率影响的双因子方差分析结果
Table 1 Results from the 2-way ANOVA examining the effect of B. gelatinae cutting section and its length on the weight gain rate.
变异源Source of variations df 均方差MS F P 切段部位Cutting section 2 3 497.028 146.425 0.000 切段长度Cutting length 2 147.348 6.170 0.009 部位×长度Section × length 4 63.922 2.677 0.065 总计Total 27  下载: 导出CSV
下载: 导出CSV
表 2 琼枝藻切段部位和长度对个体出芽数影响的双因子方差分析结果
Table 2 Results from the 2-way ANOVA examining the effect of B. gelatinae cutting section and its length on the number of buddings.
变异源Source of variations df 均方差MS F P 切段部位Cutting section 2 111.634 156.253 0.000 切段长度Cutting length 2 42.573 59.589 0.000 部位×长度Section × length 4 7.953 11.131 0.000 总计Total 27
下载: 导出CSV
表 3 温度对琼枝藻藻体切段出芽和生长的影响
Table 3 Effects of temperature on budding and growth of B. gelatinae.
温度/℃Temperature 成活率/%Survival rate 出芽率/%Germination rate 个体出芽数/个 Number of buddings 上端 侧面 下端 总数 21 100 100 4.50 ± 0.16a 0 0 4.50 ± 0.16a 24 100 100 7.40 ± 0.37b 0 0 7.40 ± 0.37b 27 100 100 9.16 ± 0.28c 0 0 9.16 ± 0.28c 30 100 66.66 3.13 ± 0.33d 0 0 3.13 ± 0.33d 33 0 - - - - - 注: 33 ℃时切段成活率为0,相关出芽指标无法测得,所以用“-”表示。Note: The survival rate was 0 when the temperature at 33 ℃, so used “-” indicated no detected to the relevant budding index.
下载: 导出CSV
-
[1] 匡梅, 曾呈奎, 夏邦美. 中国麒麟菜族的分类研究[J]. 海洋科学集刊, 1999,(00): 168-189,235-236,243-248. [2] 曾呈奎, 毕列爵. 藻类名词及名称[M]. 2版.北京:科学出版社, 2005. [3] 邢诒炫, 曾俊, 吴翔宇, 等. 三种热带经济海藻养殖现状与应用前景[J]. 海洋湖沼通报, 2019,(6): 112-120. [4] 钱树本. 海藻学[M]. 青岛: 中国海洋大学出版社, 2014: 244-245. [5] 邓春梅, 吴祖件, 何兰珍, 等. 碱法预处理琼枝麒麟菜提取卡拉胶的工艺优化[J]. 食品工业科技, 2017, 38(22): 178-183. [6] ALISTE A J, VIEIRA F F, DEL MASTRO N L. Radiation effects on agar, alginates and carrageenan to be used as food additives [J]. Radiation Physics and Chemistry, 2000, 57(3-6): 305-308. [7] BONNAUD M, WEISS J, MCCLEMENTS D J. Interaction of a food-grade cationic surfactant (lauric arginate) with food-grade biopolymers (pectin, carrageenan, xanthan, alginate, dextran, and chitosan) [J]. Journal of Agricultural and Food Chemistry, 2010, 58(17): 9770-9777. [8] PING H, MING L, DONGMEI C, et al. Morphological comparison and composition analysis of wild and farmed Eucheuma gelatinae in Hainan Province[J]. Animal Husbandry and Feed Science, 2015, (6): 3. [9] WEINER M L. Food additive carrageenan: Part II: A critical review of carrageenan in vivo safety studies [J]. Critical Reviews in Toxicology, 2014, 44(3): 244-269. [10] 李继伟, 杨贤庆, 潘创, 等. 琼枝麒麟菜的营养成分分析与评价[J]. 食品与发酵工业, 2020, 46(15): 265-269. [11] 余华军, 伍俊, 吴尚, 等. 麒麟菜多肽对血小板聚集及角叉菜胶诱导的小鼠尾部血栓形成的影响[J]. 解放军医学杂志, 2018, 43(2): 96-100. [12] 农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2020. [13] 梁磊. 琼枝麒麟菜生物学特性及其养殖技术研究[D]. 海口: 海南大学, 2010. [14] 赵霞, 陈怡超, 曾晓鹏, 等. 海南杜鹃的组织快繁体系优化[J]. 分子植物育种, 2022, 20(13): 4447-4454. [15] 张佳奇, 高玉福, 徐博, 等. 长白山特有彩叶树种紫花槭的组培快繁[J]. 植物生理学报, 2021, 57(8): 1701-1707. [16] 曾文丹, 严华兵, 曹升, 等. 何首乌离体快繁技术体系的建立[J]. 南方农业学报, 2018, 49(12): 2494-2499. [17] 赵春晓, 王春艳, 苏秀红, 等. 一种卷丹高效快繁体系的建立[J]. 植物生理学报, 2020, 56(1): 101-108. [18] 周小雪, 邓洪平, 汤绍虎. 我国特有珍稀濒危植物崖柏的离体快繁[J]. 植物生理学报, 2020, 56(5): 990-996. [19] 曾文丹, 陆柳英, 谢向誉, 等. 优良木薯品种新选048离体快繁技术[J]. 南方农业学报, 2014, 45(6): 950-954. [20] 闻永慧, 陈双秀, 汪琼, 等. 红纹凤仙花离体快繁技术体系的建立[J]. 北方园艺, 2021(15): 57-64. [21] 梅俊学, 费修绠. 条斑紫菜离体组织再生苗的研究[J]. 海洋科学, 2000(6): 39-43. [22] 朱仲嘉, 谭立佐, 翟世宽. 羊栖菜马尾藻组织培养芽生苗[J]. 水产学报, 1992 (3): 275-277. [23] 唐贤明, 王瑛睿, 刘翠, 等. 长心卡帕藻切段培养的初步研究[J]. 南方水产科学, 2014, 10(2): 36-41. [24] 王希华, 秦松, 曾呈奎. 海带愈伤组织的高效率诱导[J]. 海洋与湖沼, 1999,(6): 652-657. [25] 陈海红, 陈伟洲. 异枝江蓠切段培养的初步研究[J]. 海洋湖沼通报, 2017 (6): 146-152. [26] 梁磊, 方哲, 黄惠琴, 等. 温度对琼枝麒麟菜生长及色素含量的影响[J]. 海洋科学, 2014, 38(3): 87-90. [27] 童立豪, 吴翔宇, 曾俊, 等. 光质对琼枝生长和生理特性的影响[J]. 中国渔业质量与标准, 2020, 10(6): 17-23. [28] 曾俊, 童立豪, 吴翔宇, 等. 不同营养盐条件对琼枝生长的影响[J]. 渔业科学进展, 2021, 42(4): 168-176. [29] 童立豪, 吴翔宇, 黄良夫, 等. 光照强度与琼枝藻生长、光合色素和颜色值的相关性分析[J]. 南方水产科学, 2021, 17(5): 79-85. [30] 张帅.环境因子对琼枝生长与光合生理的影响[D].三亚: 海南热带海洋学院, 2023. [31] HOANG T H, DANG X C , LE H T, et al. Carrageenan of red algae Eucheuma gelatinae: extraction, antioxidant activity, rheology characteristics, and physicochemistry characterization[J]. Molecules, 2022, 27(4): 1268. [32] 王哲, 赵彩博, 肖娟. 超声波辅助盐酸水解提取琼枝麒麟菜卡拉胶工艺优化[J]. 海南大学学报(自然科学版), 2023, 41(4): 376-386. [33] 石硕, 孙康婷, 李继伟, 等. 琼枝麒麟菜多糖的分离纯化、结构表征及降血糖活性[J]. 大连海洋大学学报, 2023, 38(5): 866-873. [34] 李继伟, 杨贤庆, 许加超, 等. 超声波辅助酶法提取琼枝麒麟菜多糖及其理化性质研究[J]. 南方农业学报, 2020, 51(12): 3030-3039. [35] 尤丽莉. 龙须菜切段再生、原生质体分离培养及果孢子与四分孢子的收集和培养的研究[D]. 海口: 华南热带农业大学, 2005. [36] 陈昌生, 章景荣, 林爱福. 五种江蓠切段离体培养的研究[J]. 福建水产, 1987(2): 19-24. [37] 吴勋建, 贺丽虹, 宋毅, 等. 粗枝软骨藻Chondria crassicaulis组织培养初探[J]. 生态科学, 2007(4): 337-342. [38] 张俊涛. 鼠尾藻的生理生态学特性及组织培养技术的初步研究[D]. 青岛: 中国海洋大学, 2012. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 675
- HTML全文浏览量: 581
- PDF下载量: 18
- 被引次数: 0
