

-
森林生态系统的碳储量约占全球陆地生态系统的40%[1-2],森林的呼吸作用约消耗了自身光合作用固定有效碳的50%[3],而地上部分碳收支的10%~42%由树木茎干的呼吸所贡献[4],在成熟的森林中茎干呼吸超过了叶片呼吸的总量[5-6]。可见,树干呼吸在森林生态系统的碳循环和碳平衡研究中不能被忽视,因此,准确估算树干呼吸显得十分重要。树干呼吸释放的CO2通量有2个来源:其一为树干细胞代谢底物呼吸作用所产生;其二为土壤呼吸所产生并通过树木根部液流运输到树干中的CO2[7]。先前研究多通过测定胸径高度局部树干CO2释放速率(Es)及树木边材积来扩展估算整株树木及群落呼吸通量[8-9],群落树干呼吸的估算模型以温度和Q10为基础。然而,有研究提出,森林中的Es、温度及Q10在不同的高度、朝向存在明显差异[10-11],石新立等[12]和许飞等[13]认为,Es具有种内和种间差异以及明显的时空动态变化,因树干释放的CO2来源和去向是一个复杂的过程[14],Es垂直分布格局及其影响因素的报道较少,这使估算单株及群落水平树干呼吸的通量存在不确定性。研究表明,在时间尺度上Es主要表现为昼夜和季节变化,而温度是Es时间变化格局的主要影响因子[15-16],通过影响树木中酶、基质和腺苷酸的生理生化过程[14]使Es表现出规律性变化。在垂直高度上,木质部解剖特征是导致Es差异的主要因素[12-13]。树干呼吸的直接影响因子有环境因子和生物因子,而纬度、海拔和地形等因子则间接影响树干呼吸[5]。本研究选择对海南人工橡胶(Hevea brasiliensis)林样树不同垂直高度的树干CO2释放速率(Es)进行1年的原位监测,旨在研究橡胶树树干呼吸速率空间和时间变化特征,提高该地区人工林生态系统呼吸估算准确性。另外,同步监测了样树的树干液流密度(Fd),还通过显微切片法观察各高度树干木质部导管解剖结构,并进行统计分析,试图探讨橡胶树干CO2释放变化特征的机理。
-
本研究地点选于海南岛西北部农业农村部儋州热带作物科学观测实验站(位于中国热带农业科学院试验场三队内,19°32′47″N,109°28′30″E),海拔为114 m,地势平坦,年均温度23.5~24.1℃,属于热带海岛季风气候,旱雨季明显,其中5~10月份是雨季,11月~翌年4月为旱季,年均降水量约1607 mm,降水多集中于7~9月。样地栽种的橡胶林品系为PR107,种植于2001年,2009年开始割胶。试验样地土壤为砂质粘壤土,土层厚度约100 cm。橡胶林结构单一,行间距7 m×3 m,上层为橡胶林乔木层,林冠高度约16 m,下层多为1年生草本层,高度0.5 m左右,主要有弓果黍(Cyrtococcum patens)、山麻杆(Alchornea rugosa)、露籽草(Ottochloa nodosa)、竹节草(Chrysopogon aciculatus)、杯苋(Cyathula prostrata)、含羞草(Momosa pudica)等[17]。
-
采用Li-6400 IRGA(Li-cor Inc.,NE,USA)与土壤呼吸气室相连接直接测定树干的Es ,通过原位测定的方法,挑选出生长正常PR107品系样树3株,2020年5月测量的样树基本信息见表1。围绕样树搭建1个14 m高钢管脚手架,带手扶楼梯和多层测量平台。因该样地内PR107品系橡胶主干分叉多在6 m以下,故将PVC呼吸环安装在6 m以下并选择了3个垂直高度,即在未开割的样树茎干表面离地1.5 、3.0 、4.5 m处分别安装高5 cm、内径为10.4 cm的PVC环,安装方向保持一致。在安装前先清理松脱的表皮和附着物,随后根据树干表面形状将PVC环打磨成与之相吻合的弧形,再用100%中性透明防水硅酮胶均匀地涂抹在接合处,确保将PVC环固定在树干表面上密闭。自2019年9月至2020年8月,每月上旬和下旬各测量1次,测量时间选在晴朗的上午9:00~11:00间,用Li-6400土壤呼吸气室嵌入上述PVC环内测量Es,每次测量3个循环。为确保测量时的密闭性,在土壤呼吸气室探头上加1个胶垫,测量时用橡胶带将呼吸气室在树干上捆扎紧实。树干Es监测的数据由机器主机收集、存储。
表 1 橡胶样树树高和不同高度直径
Table 1. The height and diameter of sample trees at different heights
编号 树高/m 1.5 m处
直径/cm3.0 m处
直径/cm4.5 m处
直径/cmTJ301 15.2 19.7 21.1 19.8 TJ302 14.8 19.5 17.7 16.1 TJ303 15.5 21.7 19.5 19.5 -
树干液流密度(Fd)测定采用1987年法国学者Granier发明的热消散探针法(TDP)[18-19]。其原理为:上部探针电流加热,下部探针不加热,树干液流带走探针的热量,利用两针之间的温差来推算液流密度。TDP茎液流计安装在距地面1.5~2.0 m处树干边材进行连续测定,采用12 V直流电压供电,采集频率为10 Hz测定间隔为15 min,数据记录采用数据采集器(CR1000,Campbell Scientific,USA),TDP探针外面包裹防强光辐射和防潮的材料,避免因太阳辐射和降水引起液流数据的误差。
-
为保证样树Es与Fd定期原位监测不受影响,在试验样地内选出3株生理特征相近的样树(TJ88、TJ268、TJ270)通过伐木分别获取1.5、3.0、4.5 m处3个高度的圆盘(厚度3~5 cm),圆盘木质部由外至内(距离木材韧皮部3~5 cm)选取3个约0.5 cm×1 cm×1 cm(横×弦×径向)的样品木材小块用于制作切片观察。树干木质部导管解剖结构采用橡胶树木质部石蜡切片制作方法[20],具体步骤为:取材→固定→脱水→透明→浸蜡→包埋→切片→展片→脱蜡→染色→显微观察。通常在低倍镜下测量木质部的密度,在高倍镜下测量木质部导管孔径的尺寸,测量并统计样品切片的所有导管数量及导管直径。
-
本研究中仪器直接测量的树干释放CO2通量值,因树干表面积和PVC呼吸环体积不同,需要校正[15]。校正公式[21]如下:
$$ E_{s}'=\left(V+V'-\pi(d / 2)^{2} l\right) / V \times S / S' \times E_{s}, $$ (1) 式中,Es和Es′分别为校正前后的呼吸速率值(μmol·m−2·s−1),V和V′分别为呼吸气室及PVC呼吸环的体积(cm3),S和S′分为呼吸气室及PVC呼吸环覆盖树干的表面积(cm2),d和l分别为嵌入PVC呼吸环中呼吸气室的外径和深度(cm)。
-
根据文献[18]建立的以下经验公式,将记录的温差电势转换为液流密度值:
$$ F_{d} = 4.284 ×[(\Delta T_{{\rm{max}}} - \Delta T) /\Delta T] ^{1.231} \text{,} $$ (2) 式中,Fd 为瞬时液流密度(kg ·dm−2·s−1), ΔTmax 为上下探针之间的最大昼夜温差, ΔT是瞬时温度差,此公式适用于任何树种[19]。
-
Li-6400仪器运算并记录树干呼吸速率值,用MS Excel2010进行数据整理,采用Origin 2017C软件进行绘图,用IBM SPSS Statistics 25软件进行统计分析,比较不同数据组间的差异用单因素方差分析(One-way ANOVA)。
-
从图1-a中可见,橡胶树3个不同垂直高度Es在1年的观测期间均呈现出季节性单峰变化的趋势,自2019年10月起呈逐渐下降趋势,2020年4月达到月均Es的最小值,此时在空间上呈现Es3.0>Es4.5> Es1.5格局;随后Es呈逐渐上升的趋势且在2020年8月达到月均峰值,同时呈现Es3.0>Es1.5>Es4.5格局。从整个趋势看,Es1.5的最大值为2.200 μmol·m−2·s−1以及Es3.0的最大值为2.372 μmol·m−2·s−1均出现在2020年8月,而Es4.5最大值为2.054 μmol·m−2·s−1出现在2019年10月。最大呼吸速率值相比,Es3.0比Es1.5高7.81%,Es3.0比Es4.5高15.48%。图1-b中显示,Fd在不同月份的变动幅度整体呈现出与Es一致的单峰季节性变化趋势,Fd月平均值随着旱季呈现递减趋势而随着雨季呈现递增趋势,最低值出现在2020年4月而最高值出现在2019年10月。
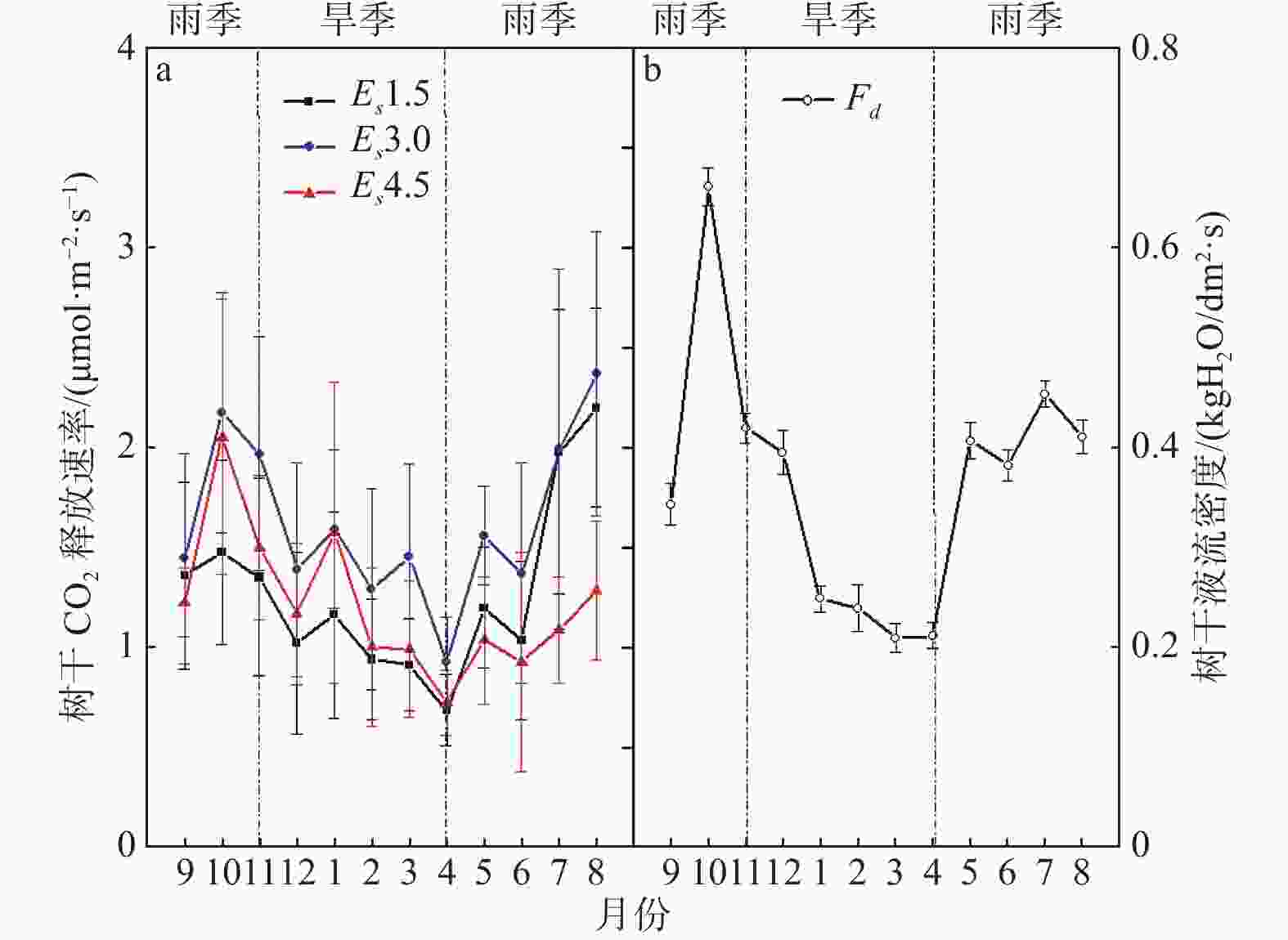
图 1 橡胶树不同高度树干CO2释放速率与树干液流季节动态变化
Figure 1. Seasonal variations of stem CO2 flux rate at different heights and stem sap flow of Hevea brasiliensis
-
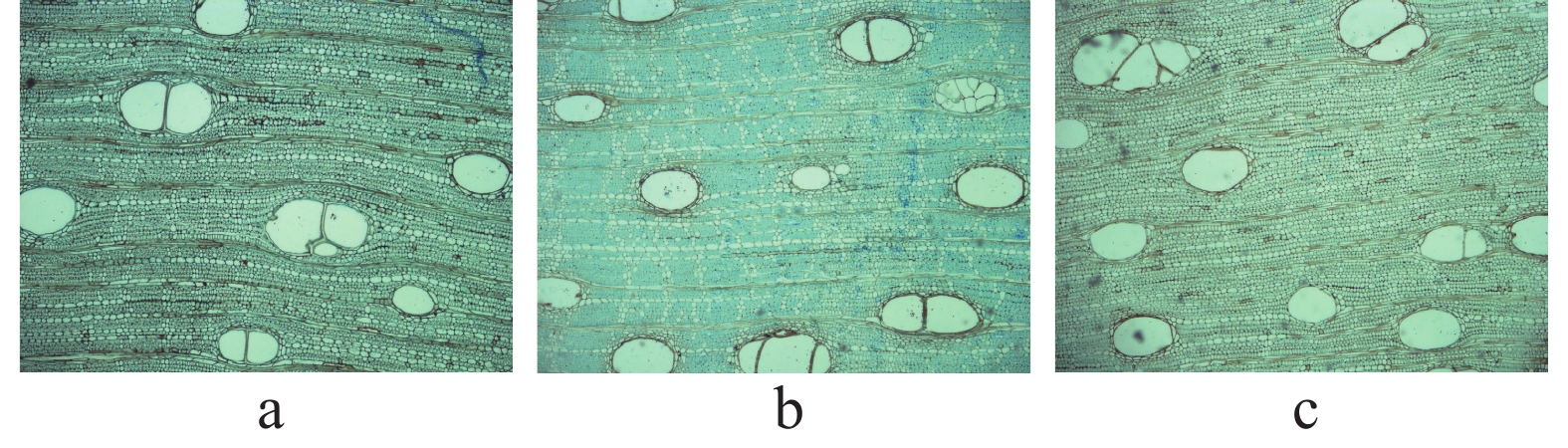
在电子显微镜下观察了3 棵样树不同高度木质部切片样品,从切片样品(图2)分析可见,3个垂直高度取样的木质部横切面上均可见色深的管细胞群,半环孔材,年轮不显明。TJ-88和TJ-270两株样树导管密度呈现3.0 m>4.5 m>1.5 m趋势,而TJ-268样树导管密度呈现4.5 m>3.0 m>1.5 m趋势。3株样树树干导管直径均呈现1.5 m>4.5 m>3.0 m趋势。经过对比,3株样树不同垂直高度树干的导管密度和直径明显不同(表2,表3)。

图 2 橡胶树不同高度树干木质部解剖照片
Figure 2. Anatomy photographs of stem xylem at different heights of Hevea brasiliensis
表 2 橡胶树干不同垂直高度木质部结构的差异
Table 2. Variation of stem xylem of Hevea brasiliensis at different vertical heights
垂直高度/ m 导管密度/(个·cm−2) 导管直径/μm TJ-88 TJ-268 TJ-270 TJ-88 TJ-268 TJ-270 1.5 (25±7)c (21±9)c (15±3)c (352.41±28.02)a (404.13±18.14)a (376.73±7.70)a 3.0 (32±4)a (29±7)b (42±9)a (299.06±25.12)c (339.44±27.55)c (309.48±35.2)c 4.5 (27±7)b (30±9)a (38±9)b (342.01±50.76)b (361.02±33.43)b (328.69±33.16)b 注:同一列不同小写字母表示不同垂直高度间存在显著差异(P<0.05)。 -
因伐木取样并制作木质部切片选在2020年5月份,故选择本试验期间5月份的Fd和Es数据进行相关性分析,结果显示,5月份Fd与各垂直高度(1.5、3.0、4.5 m)Es及导管直径呈现显著正相关关系、而与导管密度相关性并不显著,不同垂直高度木质部的导管直径均与Fd呈现显著正相关关系,1.5、3.0 m处木质部的导管直径与各垂直高度Es呈现显著正相关关系、而4.5 m处木质部的导管直径与Es未呈现显著相关关系(表3)。结果表明,Fd是Es的主要影响因子,而木质部的导管直径又是影响Fd的主要因子,木质部导管直径是影响液流和树干CO2释放速率的主要因子。
表 3 树干液流密度分别与树干CO2释放通量、导管密度及导管直径的相关性分析
Table 3. Analysis of correlation of stem sap flow density with stem CO2 efflux and the density and diameter of xylem vessels
Fd Es1.5 Es3.0 Es4.5 1.5 m
导管密度3.0 m
导管密度4.5 m
导管密度1.5 m
导管直径3.0 m
导管直径4.5 m
导管直径Fd 1 Es1.5 0.979** 1 Es3.0 0.983** 0.999** 1 Es4.5 0.973** 0.999** 0.999** 1 1.5 m导管密度 0.079 −0.573 −0.591 −0.550 1 3.0 m导管密度 0.183 −0.21 0.002 −0.49 −0.808 1 4.5 m导管密度 0.173 0.452 0.471 0.426 −0.990 0.883 1 1.5 m导管直径 0.904** 0.972** 0.967** 0.978** −0.366 −0.254 0.230 1 3.0 m导管直径 0.992** 0.997** 0.998** 0.994** −0.639 0.62 0.524 0.950** 1 4.5 m导管直径 0.921** 0.415 0.395 0.441 0.507 −0.918 −0.624 0.616 0.338 1 注:*代表显著水平(*=P<0.05, **=P<0.01)。 -
本研究结果显示,海南人工橡胶树Es在时间尺度上随原位测定的Fd呈现一致性的干湿季节性变化,与大多数树种一致[21],主要受温度和树木生长的影响[22-23],因试验样地处于热带地区,雨热同期使样树增加了蒸腾作业和液流速率,树干液流显著影响了树干中CO2的运输和释放。维管植物木质部的主要功能是传输水分与支撑植物体[24],导管管腔直径直接影响植物导水能力。泊肃叶定律(Poiseuille’S law)指出,管腔半径增加,相应导水能力呈4次方增加,故随着管腔直径的下降或增加将导致导水效率呈现大幅减少或增加的趋势[25]。本研究通过对不同垂直高度木质部切片进行显微观察对比,3个垂直高度树干的导管密度和直径明显不同,木质部传导能力呈现差异,进而影响树干液流和CO2的传导。然而,本研究结果中木质部导管的密度和直径并未呈现随着垂直梯度改变而变化的趋势,有研究表明,阔叶树的木质部导管分布无规律,其大小过渡急或不呈梯度[26],本研究结果与其一致。
在空间上,橡胶树树干Es在旱季呈现Es3.0>Es4.5> Es1.5的趋势,而湿季呈现Es3.0>Es1.5>Es4.5格局,Es最大值出现在2020年8月3.0 m处,在1年的观测期内Es并未呈现一致的梯度变化。而3株样树的导管直径均呈现1.5 m>4.5 m>3.0 m趋势,表明树干木质部解剖结构变化趋势并不能较好解释Es在垂直梯度上的变化,而其他环境因素和生理因素如树干温度、边材生长状况、非结构性碳水化合物 (non-structural carbohydrates,NSC) 分布情况、CO2在树干液流传导过程中的阻力等可能会影响Es垂直方向上的变化,这些生理生化的活动往往需要通过呼吸作用为其提供碳骨架和能量物质[27],从而导致局部树干表面较高的CO2释放速率。此外,树干3.0 m处Es大于1.5 m处和4.5 m处,这可能与树干内部活细胞的分布有关[22],局部生理活性高往往也会使该处树干表面会产生较高的CO2释放速率[10]。另外,通常每年4~11月是海南橡胶割胶季节,树干表面受损后会加速使储存在树体不同器官或组织内的非结构性碳水化合物(NSC)包括可溶性糖和不溶性的淀粉等物质代谢,为植物的生长和代谢过程提供能量,进而影响着树木的生长状况以及树干表面CO2释放通量等关键过程[28]。
虽然,本研究结果反映了Es和树干液流之间的关系以及影响,但是这两个过程的环境因素应进行更多和更深入的研究,小气候和生物效应对所研究的相关性的影响似乎更为复杂。本研究结果反映了树干木质结构与树干液流之间的关系以及对树干CO2释放通量的影响,除了木质部导管特征影响树干向上CO2传输,王秀伟和毛子军[29]提出树干中CO2横向扩散的阻力同样值得关注,尽管已知树干CO2释放径向扩散的阻力依赖于皮层、周皮层和树皮层的保护作用,但影响横向运动阻力的生理结构还需深入研究。树干表面CO2释放通量季节性变化以及空间变化的格局是环境因素和树木生理因素共同作用的结果,是理解森林生态系统碳收支过程和平衡的重要因素,其估算的准确程度有助于对热带人工林生态系统的碳平衡提供有力的理论支撑,需要得到重视。
致谢:海南省环境科学研究院董璐女士为本研究绘图和切片整理方面提供的无私帮助、中国热带农业科学院橡胶研究所周世俊先生在石蜡切片和显微观察等方面提供的技术支持,一并致谢!
Vertical variation of stem CO2 efflux and its relationship with sap flow and xylem structure of rubber trees (Hevea brasiliensis)
-
摘要: 为了研究橡胶树(Hevea brasiliensis)干CO2释放速率(Es)时空动态变化特征及其影响因素,提高人工橡胶林生态系统呼吸计算的准确性。利用便携式红外气体分析系统(IRGA)对海南人工橡胶林样树不同垂直高度的Es进行为期1年的原位监测,包括1.5 m处Es1.5、3.0 m处Es3.0、4.5 m处Es4.5,并同步监测了样树的树干液流密度(Fd),还通过显微切片法观察各高度树干木质部导管解剖结构,并进行对比分析。结果表明,Es在旱季与湿季呈现明显的季节变化;10月~翌年4月Es3.0>Es4.5> Es1.5 ,5~9月Es3.0>Es1.5>Es4.5 ;橡胶树Es的差异主要受树干液流和木质部结构的影响。通过分析橡胶树树干各高度木质部导管密度和管腔直径的差异,可知管腔直径是Fd和Es的主要影响因子,随着管腔内径的增加,液流速度和Es呈增加趋势。Abstract: An attempt was made to study the temporal and spatial dynamic characteristics of stem CO2 efflux rate (Es) and its influencing factors for development of tree stem respiration flux models, which improve the accuracy of respiration calculations in the rubber plantation ecosystem. A portable infrared gas analyzer (IRGA) was used to monitor in situ Es of a rubber (Hevea brasiliensis) plantation for one year in Hainan Island. The Es measured include those at 1.5 m height (Es1.5), 3.0m height (Es3.0) and 4.5m height (Es4.5) of the sample rubber trees. In addition, stem sap flow density (Fd) was also monitored at the same time, and the anatomical structures of the xylem vessels at different heights of the sample rubber trees were observed and analyzed by using microsection method. The results showed that Es showed seasonal changes in dry and wet season, which were in the order of Es3.0>Es4.5>Es1.5 from October to April, and Es3.0>Es1.5>Es4.5 from May to September. The Es was significantly influenced by stem sap flow and xylem structure. Analysis of the differences between density and diameter of the xylem vessels of the sample rubber trees at different heights showed that the xylem vessel diameter was the main factor influencing Fd and Es. With the increase of xylem vessel diameter, the stem sap flow velocity tended to increase and then the Es tended to increase.
-
Key words:
- Hevea brasiliensis /
- stem CO2 efflux /
- sap flow /
- xylem
-
图 2 橡胶树不同高度树干木质部解剖照片
(a)1.5 m处木质部解剖图片,(b)3.0 m处木质部解剖图片,(c)4.5 m处木质部解剖图。
Fig. 2 Anatomy photographs of stem xylem at different heights of Hevea brasiliensis
(a) Anatomy photograph of the xylem at 1.5 m; (b) Anatomy photograph of the xylem at 3.0 m; (c) Anatomy photograph of the xylem at 4.5 m.
表 1 橡胶样树树高和不同高度直径
Table 1 The height and diameter of sample trees at different heights
编号 树高/m 1.5 m处
直径/cm3.0 m处
直径/cm4.5 m处
直径/cmTJ301 15.2 19.7 21.1 19.8 TJ302 14.8 19.5 17.7 16.1 TJ303 15.5 21.7 19.5 19.5  下载: 导出CSV
下载: 导出CSV
表 2 橡胶树干不同垂直高度木质部结构的差异
Table 2 Variation of stem xylem of Hevea brasiliensis at different vertical heights
垂直高度/ m 导管密度/(个·cm−2) 导管直径/μm TJ-88 TJ-268 TJ-270 TJ-88 TJ-268 TJ-270 1.5 (25±7)c (21±9)c (15±3)c (352.41±28.02)a (404.13±18.14)a (376.73±7.70)a 3.0 (32±4)a (29±7)b (42±9)a (299.06±25.12)c (339.44±27.55)c (309.48±35.2)c 4.5 (27±7)b (30±9)a (38±9)b (342.01±50.76)b (361.02±33.43)b (328.69±33.16)b 注:同一列不同小写字母表示不同垂直高度间存在显著差异(P<0.05)。
下载: 导出CSV
表 3 树干液流密度分别与树干CO2释放通量、导管密度及导管直径的相关性分析
Table 3 Analysis of correlation of stem sap flow density with stem CO2 efflux and the density and diameter of xylem vessels
Fd Es1.5 Es3.0 Es4.5 1.5 m
导管密度3.0 m
导管密度4.5 m
导管密度1.5 m
导管直径3.0 m
导管直径4.5 m
导管直径Fd 1 Es1.5 0.979** 1 Es3.0 0.983** 0.999** 1 Es4.5 0.973** 0.999** 0.999** 1 1.5 m导管密度 0.079 −0.573 −0.591 −0.550 1 3.0 m导管密度 0.183 −0.21 0.002 −0.49 −0.808 1 4.5 m导管密度 0.173 0.452 0.471 0.426 −0.990 0.883 1 1.5 m导管直径 0.904** 0.972** 0.967** 0.978** −0.366 −0.254 0.230 1 3.0 m导管直径 0.992** 0.997** 0.998** 0.994** −0.639 0.62 0.524 0.950** 1 4.5 m导管直径 0.921** 0.415 0.395 0.441 0.507 −0.918 −0.624 0.616 0.338 1 注:*代表显著水平(*=P<0.05, **=P<0.01)。
下载: 导出CSV
-
[1] WARING R H, RUNNING S W. Forest Ecosystems: Analysis at Multiple Scales [M]. San Diego: Academic Press, 1998: 1 − 10. [2] MING X, TERRY A D, YE Q. A simple technique to measure stem respiration using a horizontally oriented soil chamber[J]. Canadian Journal of Forest Research, 2000, 30(10): 1555 −1560. [3] RYAN M G. Effects of climate change on plant respiration [J]. Ecological Applications, 1991, 1(2): 157 − 167. doi: 10.2307/1941808 [4] VOSE J, RYAN M. Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis [J]. Global Change Biology, 2002, 18(2): 182 − 193. [5] 马玉娥, 项文化, 雷丕锋. 林木树干呼吸变化及其影响因素研究进展[J]. 植物生态学报, 2007, 31(3): 403 − 412. doi: 10.3321/j.issn:1005-264X.2007.03.009 [6] WIESER G, GRUBER A, BAHN M, et al. Respiratory fluxes in a Canary Islands pine forest [J]. Tree Physiology, 2009, 29(3): 457 − 466. doi: 10.1093/treephys/tpp008 [7] TESKEY R O, MCGUIRE M A. Measurement of stem respiration of sycamore (Platanus occidental is L.) trees involves internal and external fluxes of CO2 and possible transport of CO2 from roots [J]. Plant, Cell and Environment, 2007, 30(15): 570 − 579. [8] DAMESIN C, CESCHIA E, LE G N, et al. Stem and branch respiration of beech: from tree measurements to estimations at the stand level [J]. New Phytologist, 2002, 153(1): 159 − 172. doi: 10.1046/j.0028-646X.2001.00296.x [9] MASATAKE G A, HAJIME UTSUGI, TAKUYA K, et al. Estimation of whole-stem respiration, incorporating vertical and seasonal variations in stem CO2 efflux rate, of Chamaecyparis obtusa trees [J]. Journal of Forest Research., 2010, 15(2): 115 − 122. doi: 10.1007/s10310-009-0163-3 [10] TARVAINEN L, RANTFORS M, WALLIN G. Vertical gradients and seasonal variation in stem CO2 efflux within a Norway spruce stand [J]. Tree Physiology, 2014, 34(5): 488 − 502. doi: 10.1093/treephys/tpu036 [11] 赵广, 刘刚才, 朱万泽. 贡嘎山峨眉冷杉树干呼吸空间特征及其对温度的响应[J]. 生态学报, 2018, 38(8): 2732 − 2742. [12] 石新立, 王传宽, 许飞, 等. 四个温带树种树干呼吸的时间动态及其影响因子[J]. 生态学报, 2010, 30(15): 3994 − 4003. [13] 许飞, 王传宽, 王兴昌. 东北东部14个温带树种树干呼吸的种内种间变异[J]. 生态学报, 2011, 31(13): 3581 − 3589. [14] 黄玮, 朱锦懋, 阮宏华, 等. 树干CO2释放速率影响因素研究进展[J]. 生态学杂志, 2015, 29(4): 790 − 797. [15] AMTHOR J S. The McCree-de Wit-Penning de Vries-Thornley respiration paradigms: 30 years later [J]. Annals of Botany, 2000, 86(1): 1 − 20. doi: 10.1006/anbo.2000.1175 [16] ATKIN O K, TJOELKER M G. Thermal acclimation and the dynamic response of plant respiration to temperature [J]. Trends in Plant Science, 2003, 8(7): 343 − 351. doi: 10.1016/S1360-1385(03)00136-5 [17] 陈莉, 黄先寒, 兰国玉, 等. 中国橡胶林下植物物种组成与多样性分析[J]. 西北林学院学报, 2019, 34(2): 76 − 83. [18] GRANIRR A. Mesure du flux de sève brute dans le tronc du Douglas par une nouvelle méthode thermique [J]. Ann. Sci. For., 1987, 44(1): 1 − 14. [19] GRANIRR A. Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres [J]. Ann. Sci. for., 1985, 42(2): 81 − 88. [20] 蔡海滨, 涂敏, 胡彦师, 等. 一种优化的橡胶树木质部石蜡切片制作方法[J]. 热带农业科学, 2015, 35(6): 25 − 28. doi: 10.3969/j.issn.1009-2196.2015.06.005 [21] 易建学, 谢贵水, 王纪坤, 等. 不同品系与不同径围橡胶树树干呼吸的研究[J]. 安徽农业科学, 2012, 40(27): 13433 − 13436. doi: 10.3969/j.issn.0517-6611.2012.27.079 [22] 韩风森, 胡聃, 王晓琳, 等. 北京2种阔叶树不同高度枝干的呼吸速率及其对温度的敏感性[J]. 植物生态学报, 2015, 39(2): 197 − 205. doi: 10.17521/cjpe.2015.0019 [23] 朱丽薇, 赵平, 倪广艳, 等. 荷木树干CO2释放通量的个体间差异及树干液流的效应[J]. 应用与环境生物学报, 2011, 17(4): 447 − 452. [24] 张海燕, 王传宽, 王兴昌, 等. 白桦和紫椴树干非结构性碳水化合物的空间变异[J]. 应用生态学报, 2013, 24(11): 3050 − 3056. [25] 郑云普, 王贺新, 娄鑫, 等. 木本植物非结构性碳水化合物变化及其影响因子研究进展[J]. 应用生态学报, 2013, 25(4): 1188 − 1196. [26] ZIMMRNNANN M. Xylem Structure and the Ascent of Sap[M]. Berlin: Springer, 1983. [27] GEA-IZQU I G, FONTI P, CHERUBINI P, et al. Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability [J]. Tree Physiology, 2012, 32(4): 401 − 413. doi: 10.1093/treephys/tps026 [28] 赵玮, 张一平, 宋清海, 等. 橡胶树蒸腾特征及其与环境因子的关系[J]. 生态学杂志, 2014, 33(7): 1803 − 1810. [29] 王秀伟, 毛子军. 输导组织结构对液流速度和树干CO2释放通量的影响[J]. 北京林业大学学报, 2013, 35(4): 9 − 15. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 574
- HTML全文浏览量: 293
- PDF下载量: 61
- 被引次数: 0


