

-
芒果(Mangifera indica),又被称作杧果,属于漆树科(Anacardiaceae)芒果属(Mangifera)常绿乔木,是著名的热带水果。其果实具有特殊香气,果肉细腻香甜,营养物质丰富,素有“热带果王”的美誉,且其适应能力极强,在全球热带和亚热带地区被广泛种植栽培。中国芒果的主栽区域主要分布在四川、广东、广西、云南、台湾以及海南等省份(自治区)[1]。近些年来,芒果产业发展迅速,截至2022年,我国芒果种植面积达39.52万hm2,总产量为431.7万t,产值达235.2亿元[2]。可以说,芒果产业已成为主栽地区的支柱性产业。
外观色泽是果实的重要品质指标,其往往起到吸引消费者购买的作用。芒果果皮颜色具有多种,绿色、黄色、红色(带红晕以及橙红色)。目前,以‘贵妃’、‘Zill’和‘Ruby’等为代表性的红色芒果品种深受消费者的青睐。芒果果皮的红色源自于花青苷的积累,其主要成分为矢车菊素−3−O−半乳糖苷[3]。此外,显著影响芒果果皮颜色的物质还有叶绿素和类胡萝卜素[4]。植物中花青苷的生物合成途径目前已被广泛研究,经由苯丙烷代谢和类黄酮生物合成途径催化合成[5]。该生物合成途径的产物转化主要依靠一些重要的酶发挥作用。其中,主要包括苯丙氨酸解氨酶(PAL)、肉桂酸−4−羟化酶(C4H)、4−香豆酰辅酶A连接酶(4CL)、查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、黄烷酮−3−羟化酶(F3H)、类黄酮−3’−羟化酶(F3’H)、二氢黄酮醇−4−还原酶(DFR)、花青素合成酶(ANS)以及UDP−葡萄糖:类黄酮3−O−葡萄糖基转移酶(UFGT)[6]。其中编码合成这些酶的基因被称为结构基因,而这些基因主要受到转录因子的调控。其中,由MYB(MYB类转录蛋白),bHLH(碱性螺旋−环−螺旋蛋白)以及WD40(WD40重复蛋白)转录因子所形成的MYB-bHLH-WD40 (MBW)复合体在花青苷的生物合成途径中发挥着重要的调控作用[7]。除了内在基因的调控,花青苷的生物合成同样受到外界环境因子的影响,其中光照是最重要的环境因子之一。已有研究表明,光照可以通过光强、光质以及光周期等不同因素影响花青苷的生物合成[8]。在对苹果(Malus domestica)的研究中发现,光照条件好的果实会在生长过程中积累更多的花青苷,并且光照上调了花青苷生物合成相关的结构基因[9]。在荔枝(Litchi chinensis)中,发现套袋能够抑制相关结构基因的表达,进而抑制花青苷的积累,而在去袋重新照射
阳光后,花青苷又会迅速积累[10]。除了采前自然光照射,采后补光同样能够显著诱导花青苷物质的积累。例如在采后对梨(Pyrus pyrifolia)进行UV-B补光试验中发现,补光均促进了果皮中花青苷物质的积累[11-12]。在对芒果的研究中发现,采后UV-A和UV-B并组合白光进行补光能够显著诱导芒果果皮中花青苷的生物合成[13-14]。除了紫外光,可见光补光同样具有诱导花青苷积累的作用。例如对梨果实的一项研究中发现,在蓝光的诱导下,Py4ERF24和Py12ERF96上调表达,且能够与PyMYB114发生互作,进而共同促进PyUFGT靶基因的表达从而促进花青苷的积累[15]。
目前,有关光调控花青苷生物合成的分子机制已有大量报道,植物主要通过光受体来感受外界环境所传递的光信号。所发现的植物光受体主要分为3类:分别是感知红光和远红光的光敏色素(Phytochromes,PHYs);感知UV-A和蓝光的隐花色素(Cryptochromes,CRYs)和向光素(Phototropins,PHOTs);以及感知UV-B信号的UVR8[16]。光受体在接受到光信号后会接着与调控光信号途径的COP1-HY5/HYH蛋白模块发挥作用。HY5和HYH已经被确定为光信号转导途径的正向调控因子,其可以与光信号诱导花青苷生物合成途径中的相关基因启动子的T/G-box(CACGTT)、C-box(GTCANN)、以及E-box(CAATTG)结合,并激活花青苷的合成与调控[17]。黑暗环境下COP1会在细胞核中积累,并通过泛素化作用降解调控花青苷生物合成的正向因子(HY5等),进而抑制花青苷的积累,而在光照环境下COP1会被转移至细胞质,正向调控因子无法被泛素化降解从而大量积累,进而促进花青苷的合成[18]。此外值得注意的是,高强度的光照也可能作为胁迫因子对植物造成威胁,例如其可以导致植物体内活性氧(ROS)的积累进而对植物造成胁迫[19]。而植物可通过自身的抗氧化酶系统降低这种胁迫,其主要包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等抗氧化酶系统[20]。
在红色芒果品种生产过程中,常因树冠茂密或果实套袋需求,导致芒果果实光照不足,从而面临着色不良的问题[21]。此外,芒果在转运过程中同样无法获得有效光照,因色泽问题导致其经济效益大打折扣。因此,开发一种能有效促进芒果着色的技术,对芒果产业具有重要应用价值。目前,已有关于采后利用UV-A和UV-B光照促进芒果着色的研究己有报道,但蓝光对芒果着色及其他品质的影响仍有待深入探究。本研究以红色芒果品种‘贵妃’为对象,探讨采后蓝光处理(以黑暗处理为对照)对其品质和抗氧化活性的影响,旨在通过精准可控的采后环境条件,系统分析蓝光处理对芒果外观色泽、色素含量、果肉内在品质、抗氧化酶活性以及花青苷生物合成相关结构基因和转录调控因子的影响。研究结果不仅有助于揭示相较于黑暗环境,蓝光调控芒果品质及相关生理分子响应的机制,也为采前补光技术的研发提供理论依据。
-
以青熟期(盛花后110 d)芒果品种‘贵妃’作为本采后蓝光处理试验的研究材料,芒果采摘自海南省三亚市崖州区的商业果园。选取6棵无病虫害、树形大小相似且树龄一致(12年)的芒果树,每棵果树分别采摘30个果实。采摘后迅速将果实转移至实验室,挑选大小均匀、果体健康的果实分为两组,每组90个果实。随后将两组果实分别置于培养箱(Boxun,BIC-400,上海)中进行不同处理,其中处理组采用蓝光灯管(LED,T5,5W,宁波江南仪器厂)持续照射(波长453.2 nm,光强110 μmol·m-2·s-1),对照组果实置于黑暗环境中,每天处理24 h。培养箱培养条件设置为温度17℃,湿度80%。每30个果实视为1个生物学重复,每组共设3个生物学重复。在试验处理后的0、6、24、72、144及216 h时,分别从每个生物学重复(30个果实)中随机选取5个芒果果实进行取样,将该5个果实的样品混合后视为1个生物学重复,即每个处理组在每个时间点获得3个混合样本,对应3个生物学重复。处理组取蓝光照射的上果面进行取样,对照组随机取果面。果皮取样时,使用水果削皮刀轻刮果皮,尽量减少附带果肉组织;果肉取样时,确保不附带果皮和果核组织。样品迅速置于液氮中冷冻,随后存放于−80℃超低温冰箱中,用于后续试验。
-
采用便携式色度计(Linshang,LS170,深圳)测定芒果果实的颜色指数,其中,L*值代表亮度,a*值的从负到正代表着颜色由绿转红,b*值的从负到正代表着颜色由蓝转黄。芒果果实的总可溶性固形物含量、总有机酸含量和固酸比由芒果糖酸度计(ATAGO,PAL-BX/ACID15,日本东京)测定。首先将被蓝光照射的果肉组织使用削皮刀刮下并置于研钵中,且确保不附带果皮和果核组织。随后将果肉研磨匀浆,并使用纱布过滤出果汁。测定方法为使用果汁测得总可溶性固形物含量,随后将果汁稀释50倍后测得有机酸含量,固−酸比为总可溶性固形物含量与有机酸含量的比值。
-
芒果果皮总花青素含量的测定参照已发表的文献[22]的方法。称取0.2 g样品,加入1.2 mL盐酸−甲醇提取液(V盐酸︰V甲醇 = 1︰99,4℃预冷),震荡混匀后于4℃黑暗环境下提取4 h。随后在4℃条件下以12 000 r·min−1离心10 min,取上清液,采用紫外分光光度计(新世纪,T6,中国北京)测定530 nm和600 nm处的吸光度OD530和OD600,以OD530与OD600的差值表示单位鲜质量下的花青苷含量。
叶绿素和类胡萝卜素含量的测定方法如下:称取0.1 g样品,加入1.5 mL 体积分数95%的乙醇,旋涡混合后提取40 min,随后以12 000 r·min−1离心10 min,取上清液,测定665、649和470 nm处的吸光度,计算方法参照文献[23],结果以mg·g −1为单位表示。
-
过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)活性以及总抗氧化能力(T-AOC)的测定均使用了相应的测定试剂盒(格锐思,苏州),测定方法参照了生产厂家所提供的使用说明书(
https://www.geruisi-bio.com.cn/pro/index/classid/1 ),搜索货号分别为:POD:G0107W;CAT:G0105W;SOD:G0101W;T-AOC:G0115W。 -
使用植物总RNA提取试剂盒(Tiangen, DP441,北京)提取芒果果皮样品的总RNA,试验操作步骤按照试剂盒厂商提供的使用说明书进行。随后测得cDNA浓度,依据cDNA浓度将总量为1 μg的RNA逆转录为cDNA,逆转录试验使用HiScript Ⅲ All-in-one RT SuperMix(Vazyme,R333,中国南京)试剂盒并按照厂商提供的使用说明书中的试验步骤进行。随后将逆转录得到的cDNA原液稀释20倍得到qPCR试验反应所需要的模版,随后将由5.5 μL的反应模版、1 μL的正向引物和反向引物(10 µM)和7.5μL的SYBR Premix Ex TaqTM Ⅱ(Takara,RR820A,日本京都)组成总体积为15 μL的反应体系。qPCR试验反应程序请参照之前所发表论文中方法描述[24]。qPCR试验使用了实时荧光定量PCR仪器(qTOWER3 G,德国耶拿)。基因的相对表达量采用2−ΔΔCt的方法进行计算,芒果β-actin基因被作为内参基因进行归一化处理[25]。
-
引物的设计由在线网站工具primer3(
https://bioinfo.ut.ee/primer3-0.4.0/ )完成。设计完成的引物使用TBtools软件内置的“primer check”工具检查其是否具有特异性[26]。引物的合成工作由擎科生物科技股份有限公司(Tsingke Biotech Co.,Ltd.,北京)完成(表1)。并最终通过检测引物的溶解曲线以确定引物是否特异和适用。表 1 引物序列信息
Table 1. Primer sequence information
引物名称 Primer 引物序列 Forward primer sequence(5′-3′) PAL-LOC123193566-F GAGCACATTTTAGCTGGTAGCG PAL-LOC123193566-R CAATCATTTTGGTTGCAGCACG PAL-LOC123209119-F GGAGTGACAGTGGAGCTGAG PAL-LOC123209119-R CCGGCGTTCAAGAATCTAATGA 4CL-LOC123205016-F TATTAGACGTTGAGGGTTGGCTC 4CL-LOC123205016-R GCTACTTCATCTTTTTGCGGGAC 4CL-LOC123211966-F GGAGAATCTTACGCTCACTGGGTT 4CL-LOC123211966-R CCAAAAACATTACAACAAACTCGACCG C4H-LOC123211848-F ATTTTGTCGAGGAGAGGAAGAAACT C4H-LOC123211848-R CTCAATCGACCATAGAGTTGTCTCG CHS-LOC123221035-F TTCAAGCGCATGTGTGAAAA CHS-LOC123221035-R CTTCTTTACCCAGCTTAGGGAC CHS-LOC123210196-F GAAATTCAAACGCATGTGTGAC CHS-LOC123210196-R CTTTGGAACTTCAACCACCACAA CHS-LOC123196898-F ACATTTGCGCGAAGTTGGTC CHS-LOC123196898-R GTTTGAGCTCCACCTGGTCA CHI-LOC123192937-F TGAGAGTTGTAGTGATCAAGGAGC CHI-LOC123192937-R CGCAGGGAAATGGAATGTGATAAT F3H-LOC123199749-F CACATCGGTGATCAGATTGAGATTC F3H-LOC123199749-R TTTAGGTGGATCGTCTTCTCCAAC F3H-LOC123199757-F GGAATCGATGAAGTTGATGGCA F3H-LOC123199757-R ATCTTGAACCGTTTCTCCCTGG F3'H-LOC123204634-F CTTGTTCGACAGGGGGAGGT F3'H-LOC123204634-R GATTTGAACTCATCGGCCTTCG DFR-LOC123228680-F CCACAAAGATGACTGGATGGATGT DFR-LOC123228680-R TTGGTGAAAGTGCAGAGATCAAAC ANS-LOC123211400-F AAATATGCAAACGACCAGGTCTC ANS-LOC123211400-R TATTCGCTAGTCGCCACTGTGT UFGT1-LOC123200085-F GCACATTCTTCGGTTGGTGT UFGT1-LOC123200085-R ACAAGATTGCTCTCAGGGCT UFGT3-LOC123220308-F TTCCTGAACGTTGGCCCCTT UFGT3-LOC123220308-R TTCTTCTGCCCGTCAAGCCA MYB1-LOC123197673-F TGGTCACTAATTGCGGGTAGG MYB1-LOC123197673-R TTTGAGGTGTTCGTCGCAAG bHLH2-LOC123210251-F TGGTTTCCGGTTGTGAAGTG bHLH2-LOC123210251-R ACCTTCTGCTGGTTGGTTGG WD40-LOC123193015-F GTTTGACCCCGAAACCCTCT WD40-LOC123193015-R ATCAGCAGAAACCGAAGCGA CRY1-LOC123205540-F TGAAGACTCCACGGCTGAAT CRY1-LOC123205540-R GTGAGACTGTGGATGCCTCT CRY2-LOC123220743-F ATGTGGGAAATGGAAGCTGC CRY2-LOC123220743-R ATGTGCGGCCTTTCTTCTTC COP1-LOC123196711-F AGTCGATTGAGGGTGATTGC COP1-LOC123196711-R ACCTTGATGCGTCTGGAAAC HY5-LOC123207422-F CGTCCAGCAGTGAGAGATCT HY5-LOC123207422-R TCCTGATGGCTGAACTCCTG HYH-LOC123221145-F CACCATGGTTGAAGCACAAG HYH-LOC123221145-R CGTCGGGAACCGTAAGTAAA β-actin-LOC123199782-F ATCTGCTGGAAGGTGCTGAG β-actin-LOC123199782-R CCAAGCAGCATGAAGATCAA -
数据均以均值±标准差表示。数据采用SPSS 27.0软件(SPSS, Chicago, IL, USA)进行Student’s t-test分析。P<0.05(*)和P<0.01(**)分别被认为数据统计分析具有显著性差异和极显著性差异。
-
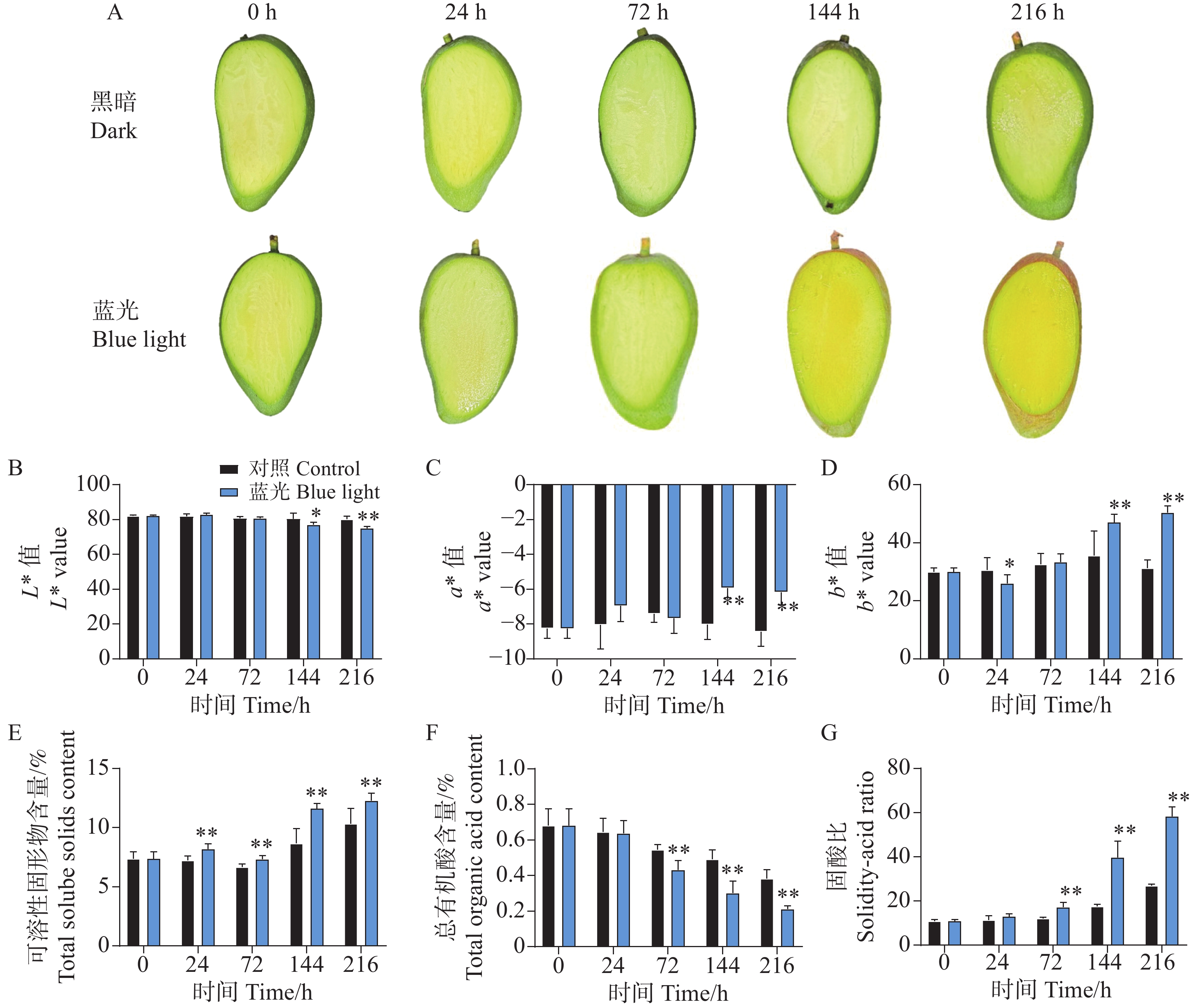
与处理组相比,随着处理时间的增加,处于黑暗环境的对照组芒果果皮没有发生转色现象,而处理组果实在光照144 h时表现出明显的红色色素积累,在处理进行到216 h时红色程度进一步增强(图1-A)。通过对果皮表面颜色指数测定结果分析表明,与对照组相比,处理组的着色现象显著降低了果皮的亮度L*值和黄色颜色指数b*值,但大大增加了红色颜色指数a*值(图1-B—D)。通过对果皮中不同色素物质含量的测定发现,处理组加快了果皮中叶绿素a和叶绿素b的降解(图1-E—F)。同时,在处理进行到144 h和216 h时,发现处理组促进了芒果果皮中类胡萝卜素的积累(图1-G)。对果皮中的花青苷物质含量测定结果分析发现,从处理进行到24 h开始,相比较于对照组,处理组芒果果皮中的花青苷开始缓慢积累,在144 h和216 h时,花青苷含量剧烈增加(图1-H)。
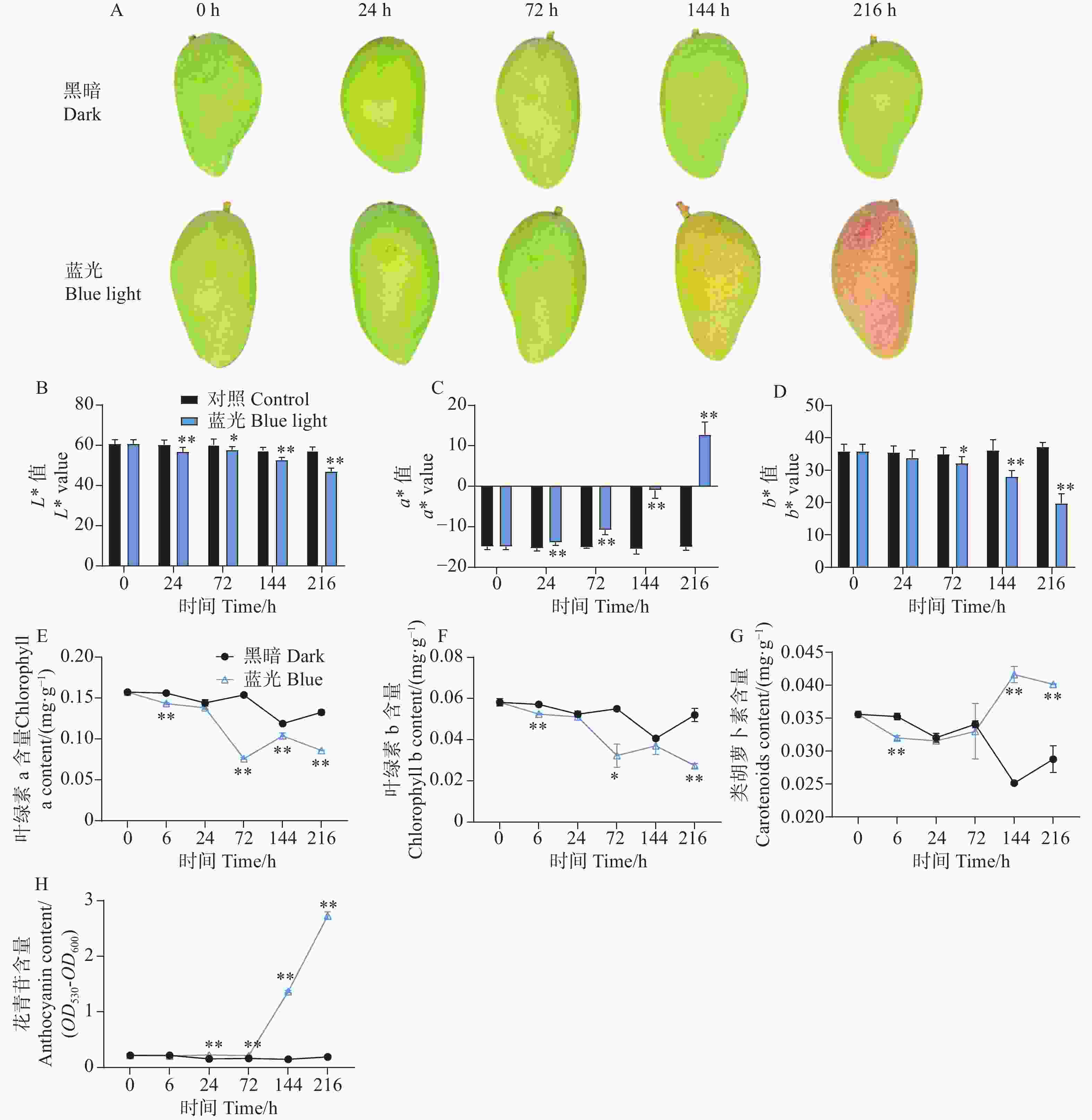
图 1 采后蓝光处理对‘贵妃’芒果果皮着色及色素含量的影响
Figure 1. Effects of postharvest blue light treatment on peel coloration and pigment content of ‘Guifei’ mango
-
相比较于对照组,处理组下的芒果果肉在处理进行到144 h时果肉出现明显转黄,而对照组则无明显变化(图2-A)。对果肉的颜色指数测定结果也表明,在第144 h和216 h时,处理组下的果肉表现出更高的b*值,说明果肉更为偏黄色。对L*和a*值的测定结果表明,在144 h和216 h时,处理组显著降低了L*值和增加了a*值(图2-B—D)。同时发现,处理组下的芒果果肉在24~216 h整个处理期间均表现出更高的可溶性固形物含量(图2-E);而在72~216 h期间表现出更低的总有机酸含量(图2-F);在整体上,处理组下的芒果果肉在72~216 h表现出更高且不断增加的固-酸比(图2-G)。说明蓝光处理能够促进芒果果实的成熟。
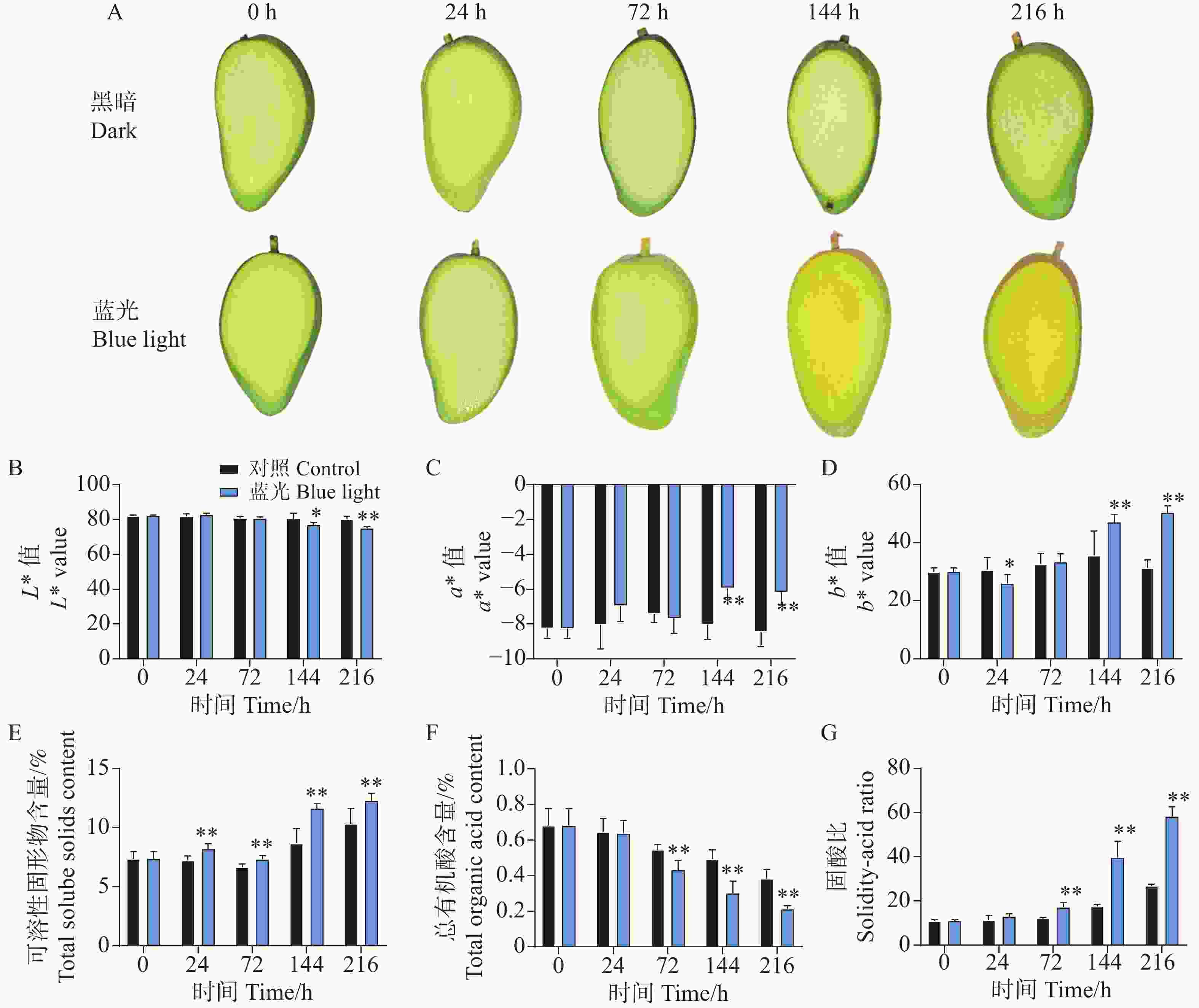
图 2 采后蓝光处理对‘贵妃’芒果果肉着色及品质的影响
Figure 2. Effects of postharvest blue light treatment on pulp coloration and quality of ‘Guifei’ mango
-
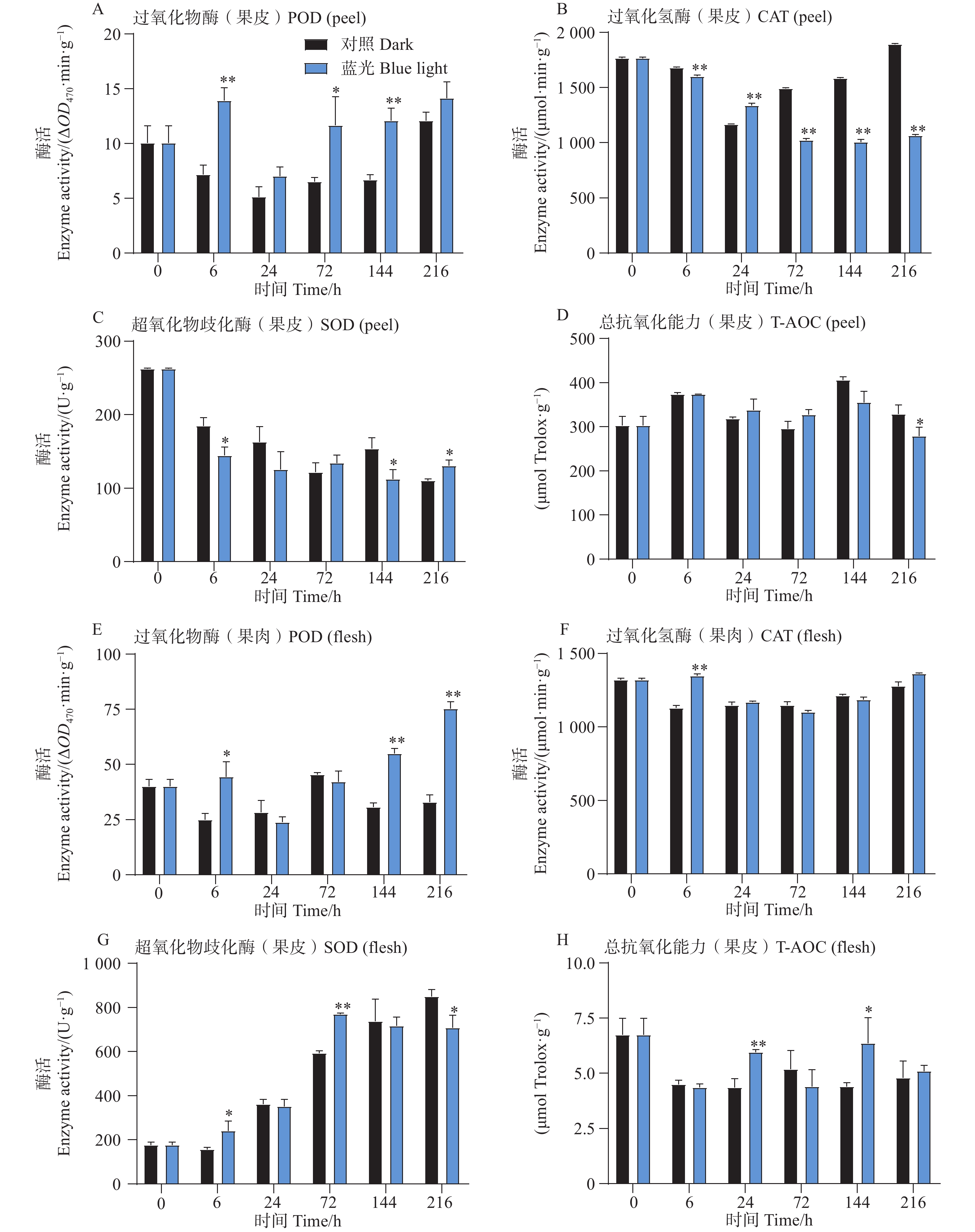
相比较于对照组,试验发现在6、72和144h时处理组芒果果皮中含有的POD活性更高,而在果肉中,POD活性整体呈上升趋势,且在6、144和216h时POD活性要比对照组高(图3-A,E)。此外发现,芒果果皮CAT活性在蓝光处理过程中呈现下降趋势,而对照组中的CAT活性则呈现先下降后上升的趋势,除了24 h,其余时间点处理组果皮中的CAT活性均低于对照组果皮(图3-B)。而T-AOC在果皮中无显著差异趋势, 而在24和144 h处理组果肉中,T-AOC较高(图3-D,H)。此外对于SOD活性,发现在处理组和对照组果皮中其整体呈现下降趋势,而在果肉中则相反,且处理组果肉中的SOD活性在前期高于对照组,在后期则开始出现低于对照组的现象(图3-C,G)。
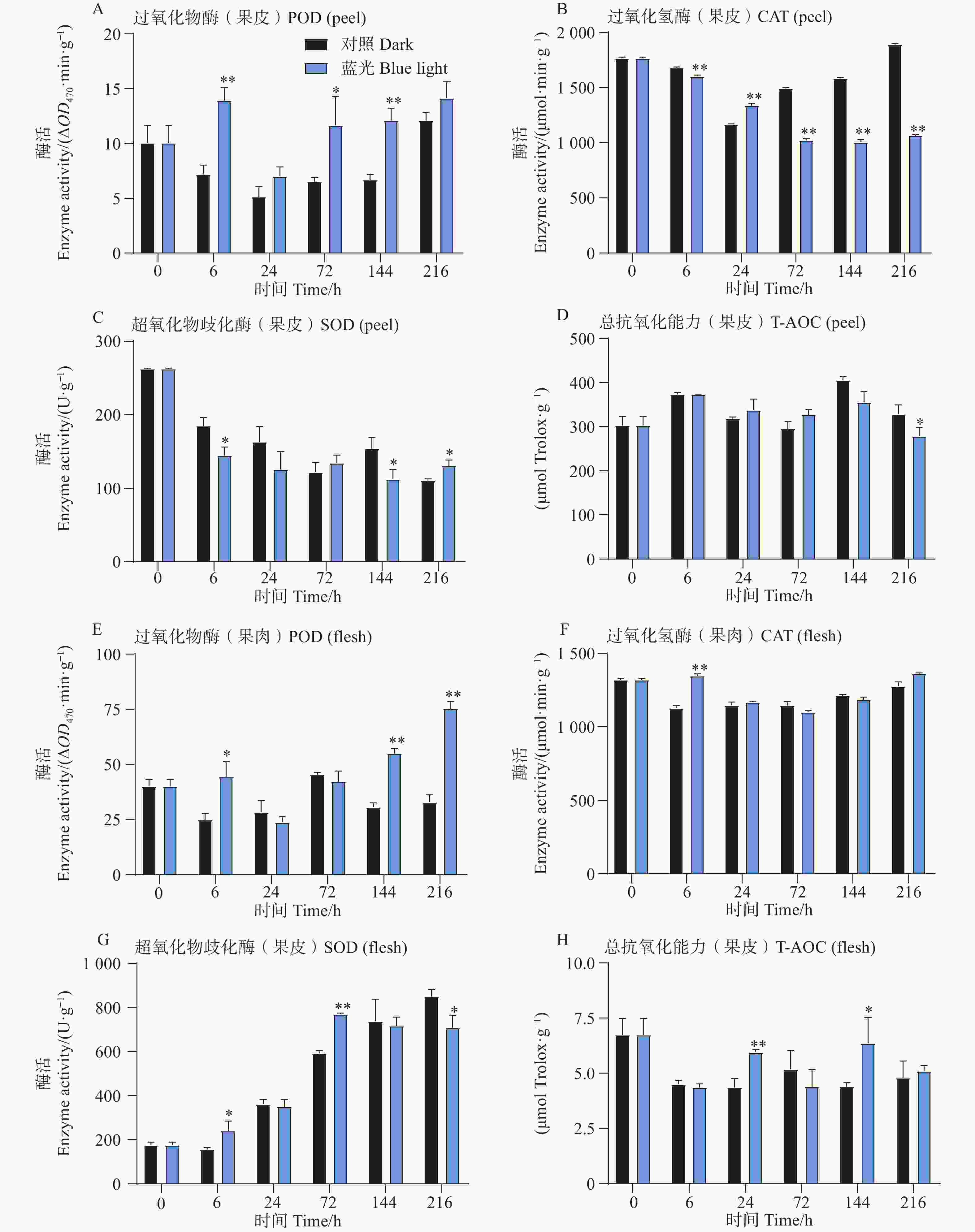
图 3 采后蓝光对芒果果实中抗氧化酶活性及总抗氧化能力的影响
Figure 3. Effects of postharvest blue light treatment on antioxidant enzyme activities and total antioxidant capacity in mango fruit
-
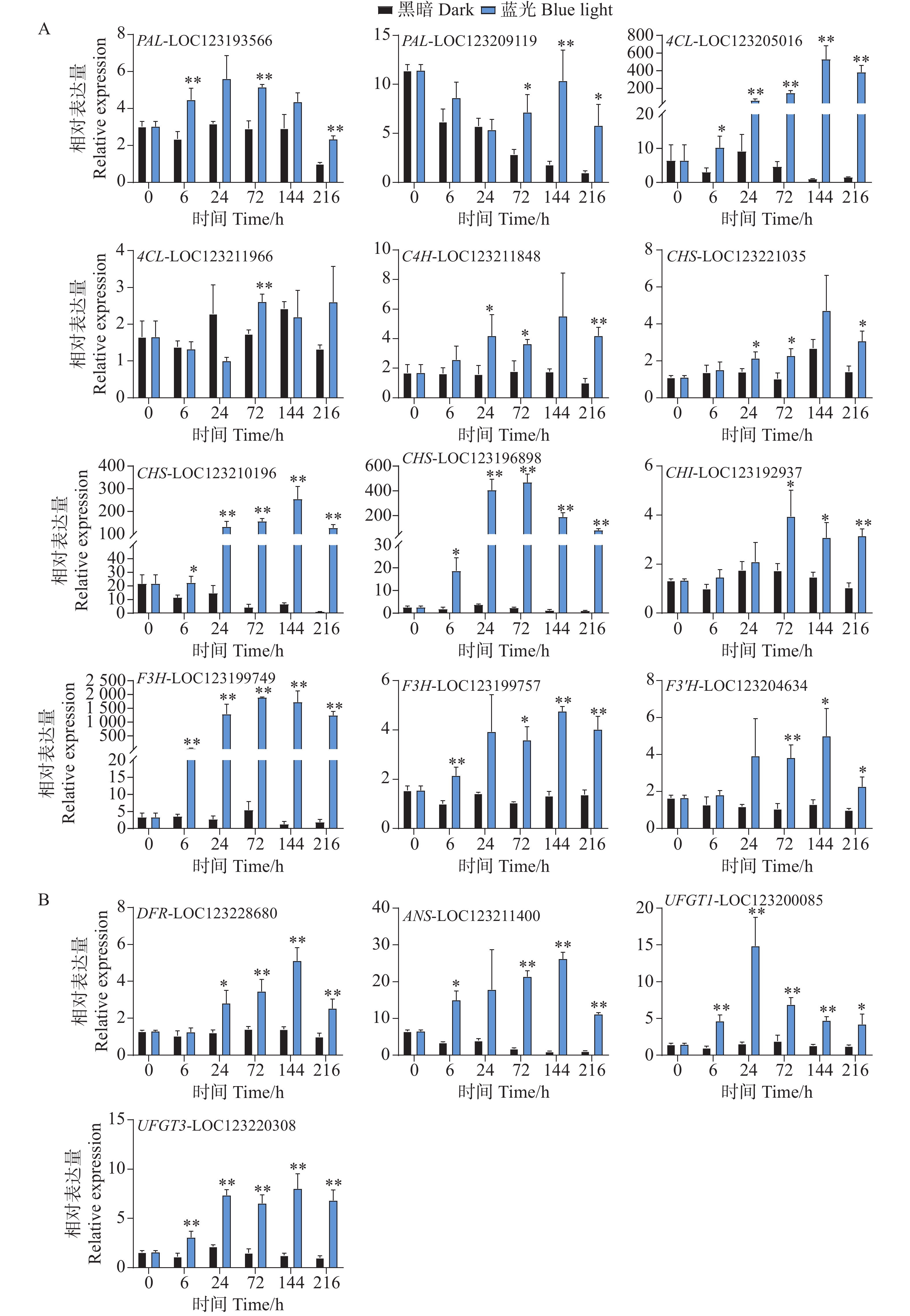
基因表达水平分析反映了花青苷生物合成通路结构基因对蓝光和黑暗处理的差异响应。对于早期合成基因,在蓝光处理后,所有基因均变现出不同程度的光响应表达模式。其中,PAL-LOC123193566基因在第6、72以及216 h处理组中表现出更高的表达水平,而PAL-LOC123209119基因在处理后期的72到216 h表达水平更高(图4-A)。4CL-LOC123205016基因在处理组的整个时期均表现出光响应模式,其基因表达水平在144 h时达到顶峰,是对照组的528倍(图4-A)。C4H-LOC123211848和CHS-LOC123221035基因的表达水平均在24、72和216 h显著高于对照组(图4-A)。而CHS-LOC123210196、CHS-LOC123196898和F3H-LOC123199749基因在整个处理时期均表现出强烈的受光诱导现象(图4-A)。其中CHI-LOC123192937和F3'H-LOC123204634基因均在后期表现出较高的表达水平模式(图4-A)。F3H-LOC123199757基因在6h开始被蓝光诱导表达,并在72、144和216h显著高于对照组的表达水平(图4-A)。
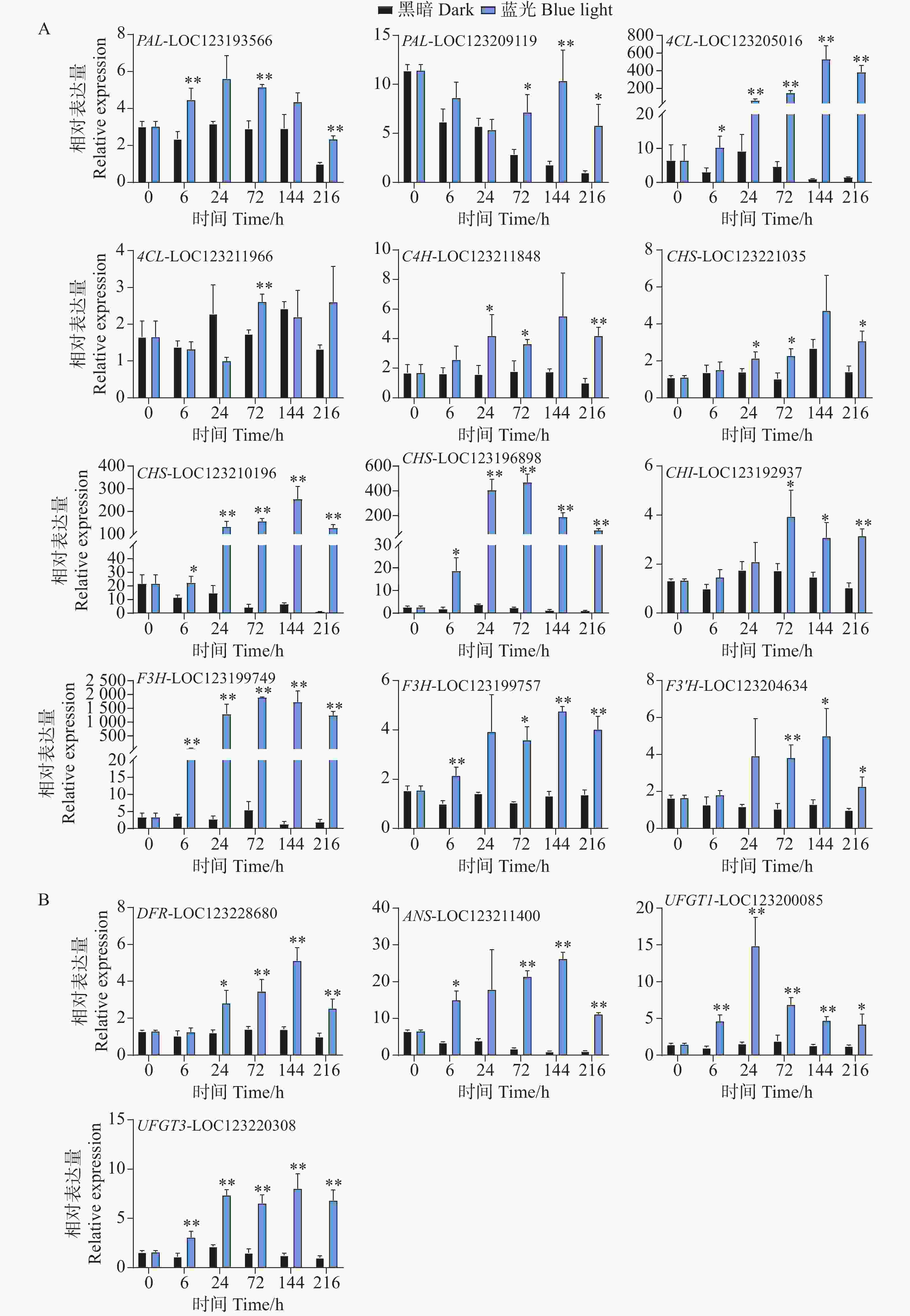
图 4 采后蓝光处理对芒果花青苷生物合成途径相关结构基因表达水平的影响分析
Figure 4. Analysis of the effect of postharvest blue light treatment on the expression levels of structural genes related to the anthocyanin biosynthesis pathway in mango
对于花青苷后期合成基因,DFR-LOC123228680基因在24 h时,处理组中的表达水平开始显著高于对照组,并在144 h达到顶峰,直至216 h(图4-B)。ANS-LOC123211400基因在蓝光处理6 h时就出现上调现象,直至216 h(图4-B)。而花青苷生物合成关键结构基因UFGT1-LOC123200085和UFGT3-LOC123220308均在处理整个时期都表现出蓝光诱导表达的现象(图4-B)。
-
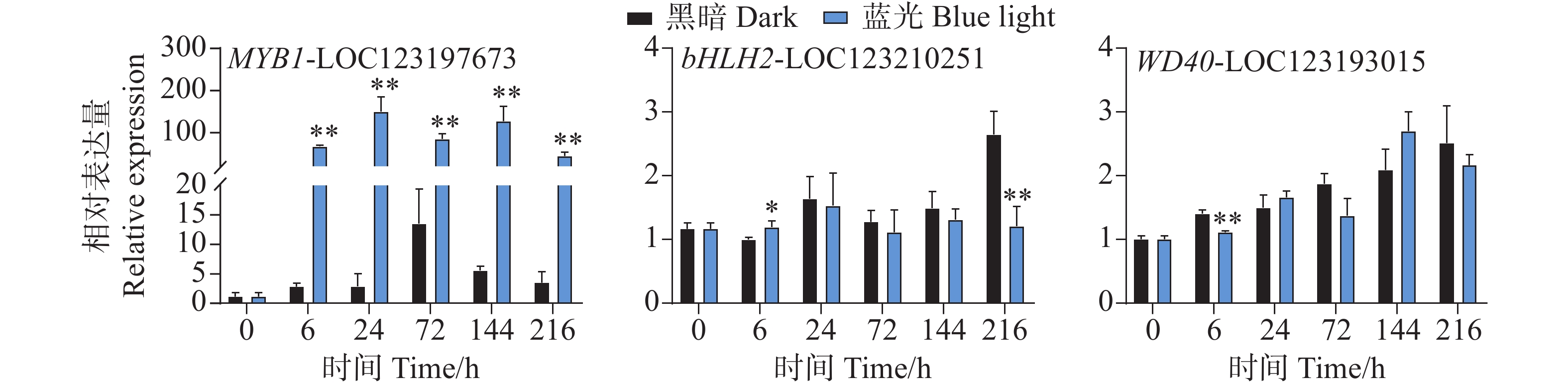
在花青苷生物合成过程中结构基因会受到转录因子的调控,其中MYB、bHLH和WD40转录因子在这个过程中发挥着主要作用。qPCR试验结果分析发现,在采后蓝光诱导下,芒果果皮中MYB和bHLH以及WD40转录因子的基因表达模式存在着很大不同。相比于对照组,其中MYB1-LOC123197673基因在处理组中的所有时期均表现出强烈的光响应模式现象,基因表达水平被显著诱导(图5)。其基因表达水平在24 h时达到顶峰,是对照组的52倍(图5)。而在对bHLH2-LOC123210251和WD40-LOC123193015的基因表达模式分析中发现,它们并未表现出由蓝光引起的差异表达趋势(图5)。相比于对照组,其中bHLH2-LOC123210251基因在6 h时轻微受到蓝光诱导表达,在216h时其表达水平又显著低于对照组,而在其它时期,并未表现出显著性差异(图5)。WD40-LOC123193015基因在6 h时处理组中的表达水平低于对照组,而在其它时期其基因表达并未表现出显著差异(图5)。
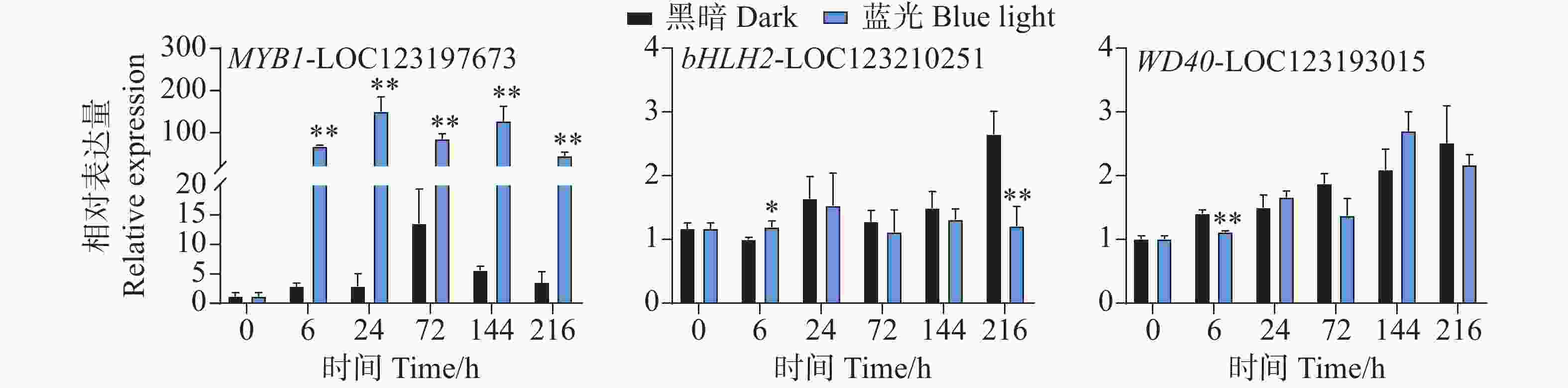
图 5 采后蓝光处理对转录调控因子MYB、bHLH和WD40基因表达水平的影响
Figure 5. Effects of postharvest blue light treatment on the expression levels of the transcriptional regulatory factors MYB, bHLH, and WD40 genes
-
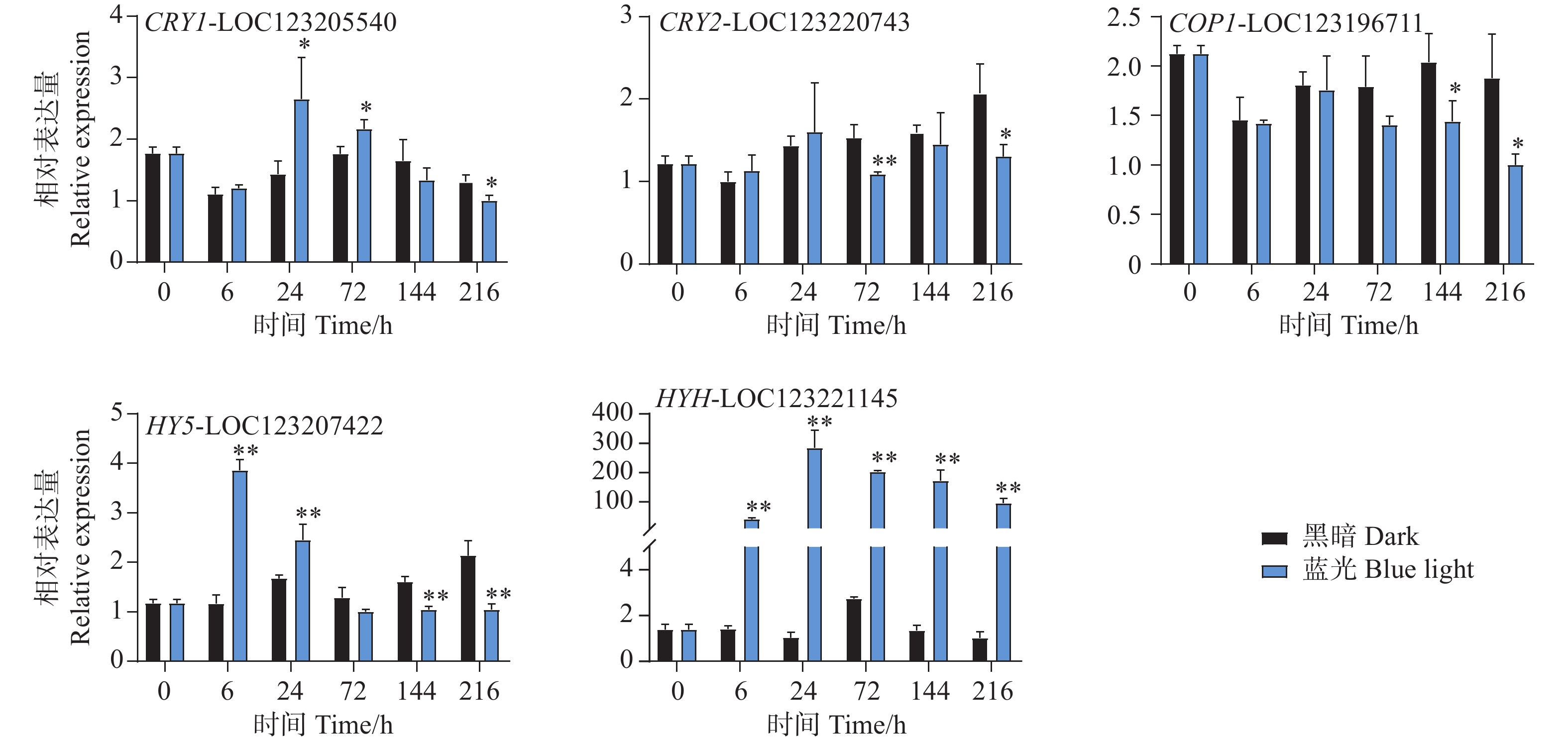
本研究通过蓝光对芒果进行采后处理,且花青苷的合成积累主要受到光信号的诱导。因此,对光信号通路一些重要基因进行了基因表达分析。qPCR试验结果显示,光信号受体隐花色素的编码基因CRY1-LOC123205540和CRY2-LOC123220743的表达模式存在差异。其中CRY1-LOC123205540在试验进行后的6 h和72 h在处理组中被显著诱导表达,且在6 h时基因表达水平达到顶峰,随后逐渐下降,并在216 h时显著低于对照组(图6)。而CRY2-LOC123220743在所有时期并未出现受到蓝光诱导表达的现象,且在72 h和216 h时其基因表达水平处理组显著低于对照组。COP1-LOC123196711基因在处理前期的处理组和对照组中的表达水平并未存在差异,而在处理后期的144 h和216 h时,其基因表达开始出现受蓝光抑制的现象,显著低于对照组(图6)。光形态关键调控因子HY5-LOC123207422和HYH-LOC123221145的基因表达模式在本试验中存在差异。其中HY5基因在6 h时就显著受到蓝光的诱导表达,并持续至24 h,随试验进行其表达水平开始出现下调,并于144 h和216 h时显著低于对照组(图6)。而HYH基因在整个试验时期,其基因表达均受到蓝光的剧烈诱导,显著高于黑暗条件下的基因表达水平,在24 h时其基因表达达到顶峰,是对照组的284倍(图6)。
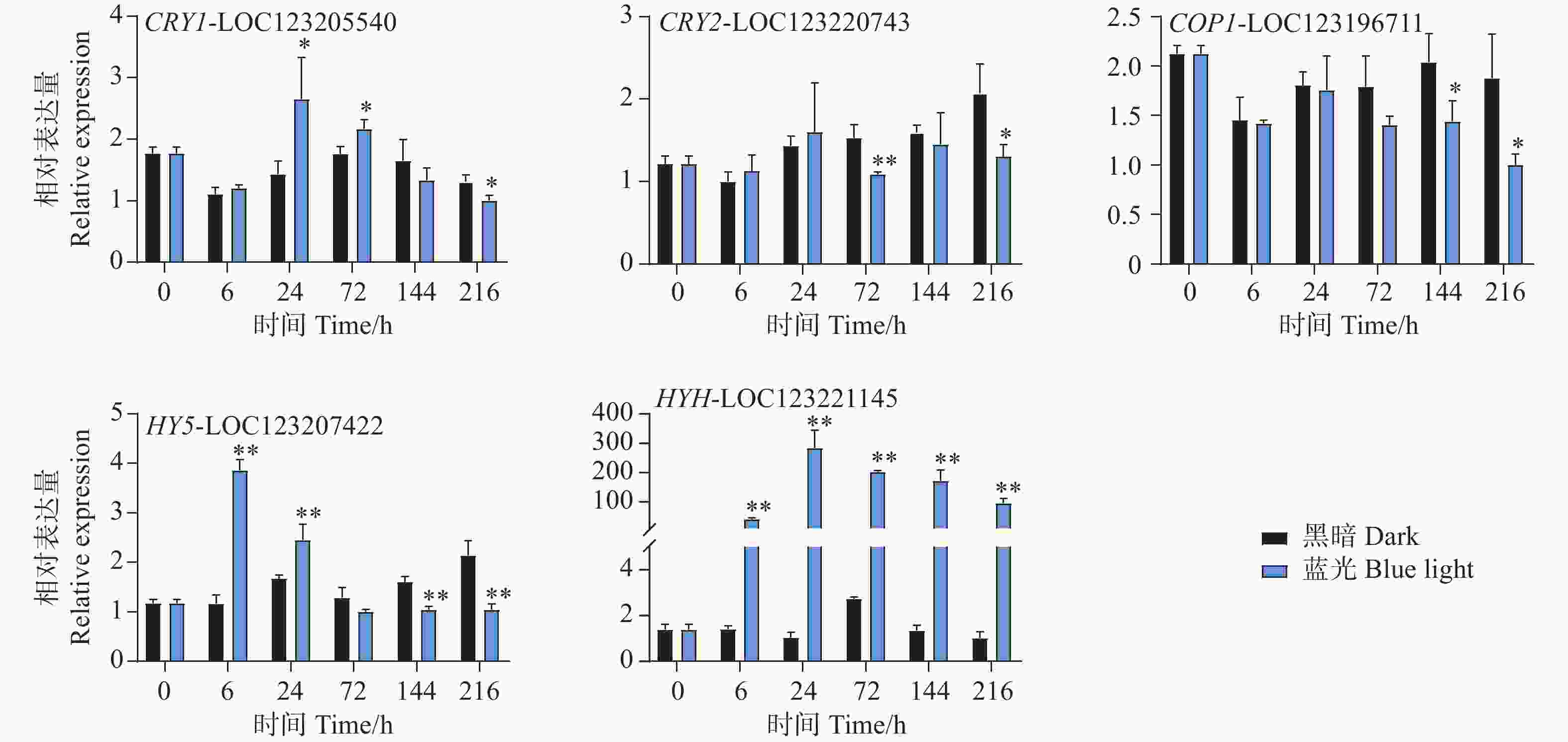
图 6 采后蓝光处理对光信号通路相关基因表达水平的影响
Figure 6. Effects of postharvest blue light treatment on the expression levels of genes related to the light signaling pathway
-
光照是调控植物生长发育最重要的环境因素之一,在植物初级与次生代谢途径中发挥关键作用。它不仅是光合作用的能量来源,也作为外界信号为植物传递信息。大量研究表明,植物花青苷的积累依赖光的诱导。采前生产中,苹果套袋果实中花青苷含量较低,去袋后则迅速积累[27]。采后补光试验也得到一致结果:UV-A和UV-B照射可显著诱导蓝莓(Vaccinium corymbosum)花青苷积累[28],蓝光照射则促进黑枸杞(Lycium ruthenicum)中花青苷积累[29]。先前研究发现,采后UV-A/白光和UV-B/白光补光均能促进芒果果皮着色。本研究发现,蓝光同样能诱导芒果果皮着色:处理144 h后果皮开始明显转色,颜色指数a*值显著升高,总花青苷含量测定也证实着色伴随花青苷大量积累。类似结果在甜椒(Capsicum annuum)[30]、生菜(Lactuca sativa)[31]和大豆(Glycine max)[32]的研究中亦有报道。进一步对芒果其他色素测定显示,果实转色伴随叶绿素a和叶绿素b降解及类胡萝卜素积累,这些变化共同支撑了蓝光处理下的转色过程。
同时,蓝光处理也影响芒果果肉品质。处理144 h后,果肉肉眼可见更黄,颜色指数b*值升高;可溶性固形物含量提高,有机酸含量降低,固-酸比升高。芒果属呼吸跃变型水果,可溶性固形物含量呈先升后降趋势,因此在实际应用中需进一步明确光照处理的时间窗口,避免因成熟过度影响果实品质与商业价值。对桃(Prunus persica)的研究发现,采后UV-B补光可影响果肉次生代谢产物合成,但UV-B无法穿透果皮辐射至果肉,推测果皮与果肉间存在信号传导通路介导果肉生理变化[33]。据此推测,芒果果皮与果肉间也可能存在类似跨组织信号传导机制。
综上,与黑暗处理相比,采后蓝光处理可显著提升芒果果皮与果肉品质,同时加速果实成熟。未来可基于此开发改良芒果着色及相关品质的应用技术,助力产业提质增效。例如,在海南秋冬红皮芒果生产中,果实常因光照不足导致着色及品质不佳,蓝光处理不仅能改善着色,还可促进成熟,实现提前上市,契合产业经济需求。此外需注意,光源具有方向性,果实背面无法直接受光,易出现“半边红”现象(与大田芒果背光面呈绿色一致),因此实际应用中应优化光源布局,确保受光均匀。未来研究需以此为基础,进一步优化光照处理工艺。
-
在本研究中,蓝光处理下果皮和果肉中POD活性的升高表明蓝光激活了芒果果皮抗氧化系统,从而对活性氧(ROS)产生响应与抵御机制。这与蓝光能够激活植物抗氧化系统的相关报道相符。例如在对草莓(Fragaria × ananassa)的研究中,蓝光处理激活了果实的抗氧化系统并提高了抗氧化酶的活性[34]。在本研究中,CAT活性在果皮蓝光处理组中出现了持续下降的现象,而对照组CAT活性出现了先降低后上升的现象且在中后期(72、144和216 h)高于蓝光处理组。推测这种现象可能是因为蓝光抑制了果皮过氧化氢的酶清除途径,导致了过氧化氢的积累,进而激活了其他抗氧化途径(例如POD途径)从而维持抗氧化能力平衡。果肉在24和144 h出现了阶段性提升总抗氧化能力的现象。推测蓝光可能通过非酶抗氧化物质(例如类黄酮和酚类)的合成增强果实的抗氧化能力。例如在植物面对胁迫时,会产生大量ROS,ROS能够作为信号激活抗氧化系统,诱导花青苷等物质的积累从而清除ROS[35]。此外,SOD活性在果皮和果肉中整体呈现相反的变化趋势,凸显了果实不同组织对光信号响应的差异性。总的来说,与黑暗处理相比,蓝光处理激活了抗氧化组织特异性反应系统,后续研究可结合自由基动态检测与多组学分析,进一步解析蓝光信号与抗氧化系统的互作网络机制。
-
在本研究中,与黑暗处理相比,花青苷生物合成通路的所有结构基因在蓝光处理下均表现出了明显的光诱导表达模式。对于花青苷早期合成结构基因,4CL、CHS以及F3H基因所表现出的光响应模式强度最为强烈。结构基因的作用较为保守,对此已有大量相关报道。在对美洲山杨(Populus tremuloides)4CL基因的研究当中,Pt4CL2被鉴定其在表皮细胞调控类黄酮物质的积累[36]。在对砂梨的采后UV-B/白光补光中,PpCHS1以及PpCHS2基因受到光信号的诱导表达[11]。在拟南芥(Arabidopsis thaliana)中,F3H基因的缺失会导致植株花色转白,且花青苷含量减少[37]。此外,在花青苷后期合成结构基因当中,UFGT作为花青苷合成途径最后一个结构基因尤为重要。在本研究中,UFGT1和UFGT3在所有时期均被蓝光诱导表达。在葡萄中,UFGT基因已被证实与花青苷的合成具有直接促进关系[38]。
花青苷的生物合成不仅受结构基因的影响,同时受到转录因子的调控。其中MYB、bHLH和WD40转录因子以及组成的MBW复合蛋白是花青苷合成通路主要的调控因子。在本研究中,MYB1基因的表达受到蓝光的强烈诱导,而bHLH和WD40的表达模式则并未出现被蓝光诱导的表达模式。这是因为bHLH和WD40往往并不单独参与转录调控,而是以蛋白的形式与MYB共同发挥作用。而WD40蛋白本身不具有催化能力,但其是MBW蛋白复合物形成不可或缺的媒介[39]。在拟南芥中,TT2(AtMYB123),TT8(AtbHLH42)以及TTG1(WD40)所组成的MBW复合体能够结合UFGT启动子进而促进花青苷的生物合成[40]。而MYB转录因子则可以单独发挥作用,例如在苹果中,光信号通过诱导MdMYB1的表达,进而促进花青苷的积累[41]。
-
为应对复杂的光信号环境,高等植物进化出一套精细的光受体系统。其中,蓝光受体CRY在光信号响应中发挥重要作用。本研究中,芒果果实在蓝光处理24 h和72 h时CRY1基因表达水平显著高于对照,表明蓝光信号被成功接收。已有研究证实,光受体是植物在光诱导下合成花青苷的关键。拟南芥中缺失光受体的cry1cry2突变体无法参与蓝光诱导的光形态建成,因而无法合成花青苷[42]。此外,本研究发现,蓝光显著诱导芒果果皮中HY5(处理早期)和HYH(处理全时期)的表达,并抑制COP1的表达(处理后期),这与前人研究结果一致。COP1一方面通过介导HY5的降解抑制花青苷积累,另一方面在光照条件下被转移至细胞质,从而丧失泛素化降解功能[43]。这一机制与蓝光诱导下芒果果实积累花青苷的现象相符。在桃中,PpHY5通过与自身启动子中的E-box元件互作,正调控花青苷合成结构基因PpCHS和PpMYB10.1的表达,从而促进花青苷积累[44]。HYH作为HY5的同源基因,在桃中也已被证实参与花青苷的调控[45]。上述结果表明,光信号转导途径在花青苷生物合成中发挥着重要作用。
-
本研究证实,与黑暗处理相比,采后蓝光处理可有效提升‘贵妃’芒果的果实品质。在外观品质方面,蓝光显著促进果皮着色,其生理机制主要涉及叶绿素降解、类胡萝卜素积累以及花青苷的大量合成。在果肉内在品质方面,蓝光处理提高了可溶性固形物含量,同时降低了有机酸含量,从而加速果实成熟进程。在抗氧化响应方面,蓝光激活了果实组织特异性防御系统,在一定程度上增强了芒果果实的抗氧化能力。分子机制分析表明,相较于黑暗处理,蓝光处理上调了花青苷生物合成通路中相关结构基因及转录因子的表达,同时诱导了光信号通路相关转录因子的转录水平。本研究系统揭示了蓝光相较于黑暗处理调控芒果果实着色与品质形成的生理及分子网络,为开发芒果采后物理增色技术及深入开展基因调控网络机制研究提供了理论依据。
Effect of postharvest blue light treatment on the quality and antioxidant activity of mango 'Guifei'
-
摘要: 芒果(Mangifera indica)果皮色泽是衡量外观品质和市场竞争力的重要指标。采后补光作为一种物理调控手段,对改善红色芒果品种着色具有应用价值。本研究以青熟期‘贵妃’芒果为材料,系统探讨了采后蓝光处理(以黑暗为对照)对果实品质、抗氧化活性及花青素生物合成的影响机制。结果表明,蓝光处理能有效促进果皮中花青素和类胡萝卜素含量增加以及叶绿素的降解,进而促进了果皮的着色。同时,果肉色泽及内在品质也得到同步改善。此外,抗氧化系统相关指标测定显示,蓝光激活了果实组织特异的抗氧化响应,果皮与果肉中过氧化物酶活性出现增强,果肉总抗氧化能力阶段性提升。花青苷合成相关分子机制表明,蓝光处理全面上调了花青素合成通路关键结构基因及转录因子MYB1的表达,此外蓝光受体基因CRY1、转录因子HY5与HYH被显著诱导,而负调控因子COP1表达在后期受到抑制。Abstract: The peel color of mango (Mangifera indica) is an important indicator of appearance quality and market competitiveness. Postharvest supplemental light treatment has an application value in improving the coloration of red mango varieties. In this context green-mature mango 'Guifei' fruits were treated with blue light to systematically analyze the effects and underlying mechanisms of postharvest blue light treatment (with darkness as the control) on their quality, antioxidant activity, and anthocyanin biosynthesis. The results showed that the blue light treatment effectively promoted the accumulation of anthocyanins and carotenoids in the peel, as well as the degradation of chlorophyll, thereby facilitating peel coloration. Concurrently, the flesh color and internal quality were also improved. Furthermore, assays of antioxidant system-related indicators revealed that the blue light activated tissue-specific antioxidant responses in the fruit. Peroxidase activity was enhanced in both the peel and flesh, and the total antioxidant capacity of the flesh was increased in a stage-specific manner. Mechanistic studies on anthocyanin biosynthesis indicated that the blue light treatment comprehensively upregulated the expression of key structural genes involved in the anthocyanin biosynthesis pathway, as well as the transcription factor MYB1. Additionally, the expression of the blue light receptor gene CRY1 and the transcription factors HY5 and HYH was significantly induced, while the expression of the negative regulator COP1 was suppressed at the late stage.
-
Key words:
- Mango(Mangifera indica) /
- blue light /
- anthocyanin /
- antioxidation /
- light signaling
-
图 2 采后蓝光处理对‘贵妃’芒果果肉着色及品质的影响
Fig. 2 Effects of postharvest blue light treatment on pulp coloration and quality of ‘Guifei’ mango
图 3 采后蓝光对芒果果实中抗氧化酶活性及总抗氧化能力的影响
Fig. 3 Effects of postharvest blue light treatment on antioxidant enzyme activities and total antioxidant capacity in mango fruit
图 4 采后蓝光处理对芒果花青苷生物合成途径相关结构基因表达水平的影响分析
Fig. 4 Analysis of the effect of postharvest blue light treatment on the expression levels of structural genes related to the anthocyanin biosynthesis pathway in mango
图 5 采后蓝光处理对转录调控因子MYB、bHLH和WD40基因表达水平的影响
Fig. 5 Effects of postharvest blue light treatment on the expression levels of the transcriptional regulatory factors MYB, bHLH, and WD40 genes
图 6 采后蓝光处理对光信号通路相关基因表达水平的影响
Fig. 6 Effects of postharvest blue light treatment on the expression levels of genes related to the light signaling pathway
表 1 引物序列信息
Table 1 Primer sequence information
引物名称 Primer 引物序列 Forward primer sequence(5′-3′) PAL-LOC123193566-F GAGCACATTTTAGCTGGTAGCG PAL-LOC123193566-R CAATCATTTTGGTTGCAGCACG PAL-LOC123209119-F GGAGTGACAGTGGAGCTGAG PAL-LOC123209119-R CCGGCGTTCAAGAATCTAATGA 4CL-LOC123205016-F TATTAGACGTTGAGGGTTGGCTC 4CL-LOC123205016-R GCTACTTCATCTTTTTGCGGGAC 4CL-LOC123211966-F GGAGAATCTTACGCTCACTGGGTT 4CL-LOC123211966-R CCAAAAACATTACAACAAACTCGACCG C4H-LOC123211848-F ATTTTGTCGAGGAGAGGAAGAAACT C4H-LOC123211848-R CTCAATCGACCATAGAGTTGTCTCG CHS-LOC123221035-F TTCAAGCGCATGTGTGAAAA CHS-LOC123221035-R CTTCTTTACCCAGCTTAGGGAC CHS-LOC123210196-F GAAATTCAAACGCATGTGTGAC CHS-LOC123210196-R CTTTGGAACTTCAACCACCACAA CHS-LOC123196898-F ACATTTGCGCGAAGTTGGTC CHS-LOC123196898-R GTTTGAGCTCCACCTGGTCA CHI-LOC123192937-F TGAGAGTTGTAGTGATCAAGGAGC CHI-LOC123192937-R CGCAGGGAAATGGAATGTGATAAT F3H-LOC123199749-F CACATCGGTGATCAGATTGAGATTC F3H-LOC123199749-R TTTAGGTGGATCGTCTTCTCCAAC F3H-LOC123199757-F GGAATCGATGAAGTTGATGGCA F3H-LOC123199757-R ATCTTGAACCGTTTCTCCCTGG F3'H-LOC123204634-F CTTGTTCGACAGGGGGAGGT F3'H-LOC123204634-R GATTTGAACTCATCGGCCTTCG DFR-LOC123228680-F CCACAAAGATGACTGGATGGATGT DFR-LOC123228680-R TTGGTGAAAGTGCAGAGATCAAAC ANS-LOC123211400-F AAATATGCAAACGACCAGGTCTC ANS-LOC123211400-R TATTCGCTAGTCGCCACTGTGT UFGT1-LOC123200085-F GCACATTCTTCGGTTGGTGT UFGT1-LOC123200085-R ACAAGATTGCTCTCAGGGCT UFGT3-LOC123220308-F TTCCTGAACGTTGGCCCCTT UFGT3-LOC123220308-R TTCTTCTGCCCGTCAAGCCA MYB1-LOC123197673-F TGGTCACTAATTGCGGGTAGG MYB1-LOC123197673-R TTTGAGGTGTTCGTCGCAAG bHLH2-LOC123210251-F TGGTTTCCGGTTGTGAAGTG bHLH2-LOC123210251-R ACCTTCTGCTGGTTGGTTGG WD40-LOC123193015-F GTTTGACCCCGAAACCCTCT WD40-LOC123193015-R ATCAGCAGAAACCGAAGCGA CRY1-LOC123205540-F TGAAGACTCCACGGCTGAAT CRY1-LOC123205540-R GTGAGACTGTGGATGCCTCT CRY2-LOC123220743-F ATGTGGGAAATGGAAGCTGC CRY2-LOC123220743-R ATGTGCGGCCTTTCTTCTTC COP1-LOC123196711-F AGTCGATTGAGGGTGATTGC COP1-LOC123196711-R ACCTTGATGCGTCTGGAAAC HY5-LOC123207422-F CGTCCAGCAGTGAGAGATCT HY5-LOC123207422-R TCCTGATGGCTGAACTCCTG HYH-LOC123221145-F CACCATGGTTGAAGCACAAG HYH-LOC123221145-R CGTCGGGAACCGTAAGTAAA β-actin-LOC123199782-F ATCTGCTGGAAGGTGCTGAG β-actin-LOC123199782-R CCAAGCAGCATGAAGATCAA  下载: 导出CSV
下载: 导出CSV
-
[1] 陈业渊, 党志国, 林电, 等. 中国杧果科学研究70年[J]. 热带作物学报, 2020, 41(10): 2034−2044. https://doi.org/10.3969/j.issn.1000-2561.2020.10.010 doi: 10.3969/j.issn.1000-2561.2020.10.010 [2] 冯焕德, 朱敏, 汪秀华, 等. 海南杧果主产区果园土壤pH值和有机质分布特征及其与土壤养分的关系[J]. 中国热带农业, 2025(1): 44−53. https://doi.org/10.3969/j.issn.1673-0658.2025.01.008 doi: 10.3969/j.issn.1673-0658.2025.01.008 [3] Zhu W C, Wu H X, Yang C K, et al. Postharvest light-induced flavonoids accumulation in mango (Mangifera indica L. ) peel is associated with the up-regulation of flavonoids-related and light signal pathway genes [J]. Frontiers in Plant Science, 2023, 14: 1136281. https://doi.org/10.3389/fpls.2023.1136281 doi: 10.3389/fpls.2023.1136281 [4] Chen M M, Gu H, Wang L R, et al. Exogenous ethylene promotes peel color transformation by regulating the degradation of chlorophyll and synthesis of anthocyanin in postharvest Mango fruit [J]. Frontiers in Nutrition, 2022, 9: 911542. https://doi.org/10.3389/fnut.2022.911542 doi: 10.3389/fnut.2022.911542 [5] Petroni K, Tonelli C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs [J]. Plant Science, 2011, 181(3): 219−229. https://doi.org/10.1016/j.plantsci.2011.05.009 doi: 10.1016/j.plantsci.2011.05.009 [6] Jaakola L. New insights into the regulation of anthocyanin biosynthesis in fruits [J]. Trends in Plant Science, 2013, 18(9): 477−483. https://doi.org/10.1016/j.tplants.2013.06.003 doi: 10.1016/j.tplants.2013.06.003 [7] Dubos C, Stracke R, Grotewold E, et al. MYB transcription factors in Arabidopsis [J]. Trends in Plant Science, 2010, 15(10): 573−581. https://doi.org/10.1016/j.tplants.2010.06.005 doi: 10.1016/j.tplants.2010.06.005 [8] Chen M, Chory J, Fankhauser C. Light signal transduction in higher plants [J]. Annual Review of Genetics, 2004, 38: 87−117. https://doi.org/10.1146/annurev.genet.38.072902.092259 doi: 10.1146/annurev.genet.38.072902.092259 [9] Feng F J, Li M J, Ma F W, et al. Phenylpropanoid metabolites and expression of key genes involved in anthocyanin biosynthesis in the shaded peel of apple fruit in response to sun exposure [J]. Plant Physiology and Biochemistry, 2013, 69: 54−61. https://doi.org/10.1016/j.plaphy.2013.04.020 doi: 10.1016/j.plaphy.2013.04.020 [10] Wei Y Z, Hu F C, Hu G B, et al. Differential expression of anthocyanin biosynthetic genes in relation to anthocyanin accumulation in the pericarp of Litchi chinensis Sonn [J]. PLoS One, 2011, 6(4): e19455. https://doi.org/10.1371/journal.pone.0019455 doi: 10.1371/journal.pone.0019455 [11] Qian M J, Zhang D, Yue X Y, et al. Analysis of different pigmentation patterns in ‘Mantianhong’ (Pyrus pyrifolia Nakai) and ‘Cascade’ (Pyrus communis L. ) under bagging treatment and postharvest UV-B/visible irradiation conditions [J]. Scientia Horticulturae, 2013, 151: 75−82. https://doi.org/10.1016/j.scienta.2012.12.020 doi: 10.1016/j.scienta.2012.12.020 [12] Sun Y W, Qian M J, Wu R Y, et al. Postharvest pigmentation in red Chinese sand pears (Pyrus pyrifolia Nakai) in response to optimum light and temperature [J]. Postharvest Biology and Technology, 2014, 91: 64−71. https://doi.org/10.1016/j.postharvbio.2013.12.015 doi: 10.1016/j.postharvbio.2013.12.015 [13] Li F L, Tahir M M, Yang C K, et al. Postharvest UV-A treatment promotes mango fruit pigmentation and ripening in a dose-dependent manner [J]. Postharvest Biology and Technology, 2025, 230: 113836. https://doi.org/10.1016/j.postharvbio.2025.113836 doi: 10.1016/j.postharvbio.2025.113836 [14] Yang C K, Wang X W, Zhu W C, et al. Postharvest white light combined with different UV-B doses differently promotes anthocyanin accumulation and antioxidant capacity in mango peel [J]. LWT, 2024, 203: 116385. https://doi.org/10.1016/j.lwt.2024.116385 doi: 10.1016/j.lwt.2024.116385 [15] Ni J B, Bai S L, Zhao Y, et al. Ethylene response factors Pp4ERF24 and Pp12ERF96 regulate blue light-induced anthocyanin biosynthesis in ‘Red Zaosu’ pear fruits by interacting with MYB114 [J]. Plant Molecular Biology, 2019, 99(1): 67−78. https://doi.org/10.1007/s11103-018-0802-1 doi: 10.1007/s11103-018-0802-1 [16] Casal J J. Photoreceptor signaling networks in plant responses to shade [J]. Annual Review of Plant Biology, 2013, 64: 403−427. https://doi.org/10.1146/annurev-arplant-050312-120221 doi: 10.1146/annurev-arplant-050312-120221 [17] An J P, Qu F J, Yao J F, et al. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple [J]. Horticulture Research, 2017, 4: 17023. https://doi.org/10.1038/hortres.2017.23 doi: 10.1038/hortres.2017.23 [18] Lau O S, Deng X W. The photomorphogenic repressors COP1 and DET1: 20 years later [J]. Trends in Plant Science, 2012, 17(10): 584−593. https://doi.org/10.1016/j.tplants.2012.05.004 doi: 10.1016/j.tplants.2012.05.004 [19] Hideg É, Jansen M A K, Strid Å. UV-B exposure, ROS, and stress: inseparable companions or loosely linked associates? [J]. Trends in Plant Science, 2013, 18(2): 107−115. https://doi.org/10.1016/j.tplants.2012.09.003 doi: 10.1016/j.tplants.2012.09.003 [20] Abdulfatah H. Non-Enzymatic Antioxidants in Stressed Plants: A Review [J]. Journal of University of Anbar for Pure Science, 2022, 16(2): 25−37. https://doi.org/10.37652/juaps.2022.176435 doi: 10.37652/juaps.2022.176435 [21] Karanjalker G R, Ravishankar K V, Shivashankara K S, et al. Influence of bagging on color, anthocyanin and anthocyanin biosynthetic genes in peel of red colored Mango Cv. ‘Lily’ [J]. Erwerbs-Obstbau, 2018, 60(4): 281−287. https://doi.org/10.1007/s10341-018-0371-0 doi: 10.1007/s10341-018-0371-0 [22] Zhang D L, Quantick P C. Effects of chitosan coating on enzymatic browning and decay during postharvest storage of litchi (Litchi chinensis Sonn. ) fruit [J]. Postharvest Biology and Technology, 1997, 12(2): 195−202. https://doi.org/10.1016/S0925-5214(97)00057-4 doi: 10.1016/S0925-5214(97)00057-4 [23] Lichtenthaler H K, Wellburn A R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents [J]. Biochemical Society Transactions, 1983, 11(5): 591−592. https://doi.org/10.1042/bst0110591 doi: 10.1042/bst0110591 [24] Shi B, Wu H X, Zheng B, et al. Analysis of light-independent anthocyanin accumulation in Mango (Mangifera indica L. ) [J]. Horticulturae, 2021, 7(11): 423. https://doi.org/10.3390/horticulturae7110423 doi: 10.3390/horticulturae7110423 [25] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method [J]. Methods, 2001, 25(4): 402−408. https://doi.org/10.1006/meth.2001.1262 doi: 10.1006/meth.2001.1262 [26] Chen C J, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194−1202. https://doi.org/10.1016/j.molp.2020.06.009 doi: 10.1016/j.molp.2020.06.009 [27] Ma C Q, Liang B W, Chang B, et al. Transcriptome profiling of anthocyanin biosynthesis in the peel of “Granny Smith” apples (Malus domestica) after bag removal [J]. BMC Genomics, 2019, 20(1): 353. https://doi.org/10.1186/s12864-019-5730-1 doi: 10.1186/s12864-019-5730-1 [28] Yang J F, Li B B, Shi W J, et al. Transcriptional activation of anthocyanin biosynthesis in developing fruit of blueberries (Vaccinium corymbosum L. ) by preharvest and postharvest UV irradiation [J]. Journal of Agricultural and Food Chemistry, 2018, 66(42): 10931−10942. https://doi.org/10.1021/acs.jafc.8b03081 doi: 10.1021/acs.jafc.8b03081 [29] Zeng H T, Zheng T, Peng X, et al. Transcriptomic and targeted metabolomics analysis of detached Lycium ruthenicum leaves reveals mechanisms of anthocyanin biosynthesis induction through light quality and sucrose treatments [J]. Metabolites, 2023, 13(9): 1004. https://doi.org/10.3390/metabo13091004 doi: 10.3390/metabo13091004 [30] 高金汇. 多组学联合分析蓝光诱导紫色甜椒果实花青苷的合成机制[D]. 泰安: 山东农业大学, 2024. [31] 熊伟任, 雷雨田, 杨永森, 等. 不同波长蓝光对紫叶生菜品质形成的影响[J]. 江苏农业科学, 2023, 51(1): 143−148. https://doi.org/10.15889/j.issn.1002-1302.2023.01.021 doi: 10.15889/j.issn.1002-1302.2023.01.021 [32] 胡钢. GmMYB75和GmMYB90响应蓝光调控大豆芽苗下胚轴中花青苷合成的机理研究[D]. 南京: 南京农业大学, 2023. https://doi.org/10.27244/d.cnki.gnjnu.2023.000101 [33] Santin M, Ranieri A, Hauser M T, et al. The outer influences the inner: postharvest UV-B irradiation modulates peach flesh metabolome although shielded by the skin [J]. Food Chemistry, 2021, 338: 127782. https://doi.org/10.1016/j.foodchem.2020.127782 doi: 10.1016/j.foodchem.2020.127782 [34] Xu F, Shi L Y, Chen W, et al. Effect of blue light treatment on fruit quality, antioxidant enzymes and radical-scavenging activity in strawberry fruit [J]. Scientia Horticulturae, 2014, 175: 181−186. https://doi.org/10.1016/j.scienta.2014.06.012 doi: 10.1016/j.scienta.2014.06.012 [35] Tohge T, Watanabe M, Hoefgen R, et al. The evolution of phenylpropanoid metabolism in the green lineage [J]. Critical Reviews in Biochemistry and Molecular Biology, 2013, 48(2): 123−152. https://doi.org/10.3109/10409238.2012.758083 doi: 10.3109/10409238.2012.758083 [36] Hu W J, Kawaoka A, Tsai C J, et al. Compartmentalized expression of two structurally and functionally distinct 4-coumarate: CoA ligase genes in aspen (Populus tremuloides) [J]. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(9): 5407−5412. https://doi.org/10.1073/pnas.95.9.5407 doi: 10.1073/pnas.95.9.5407 [37] Peer W A, Brown D E, Tague B W, et al. Flavonoid accumulation patterns of transparent testa mutants of arabidopsis [J]. Plant Physiology, 2001, 126(2): 536−548. https://doi.org/10.1104/pp.126.2.536 doi: 10.1104/pp.126.2.536 [38] Hall D, Yuan X X, Murata J, et al. Molecular cloning and biochemical characterization of the UDP-glucose: flavonoid 3-O-glucosyltransferase from concord grape (Vitis labrusca) [J]. Phytochemistry, 2012, 74: 90−99. https://doi.org/10.1016/j.phytochem.2011.10.007 doi: 10.1016/j.phytochem.2011.10.007 [39] Xu C, Min J R. Structure and function of WD40 domain proteins [J]. Protein & Cell, 2011, 2(3): 202−214. https://doi.org/10.1007/s13238-011-1018-1 doi: 10.1007/s13238-011-1018-1 [40] Baudry A, Heim M A, Dubreucq B, et al. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana [J]. The Plant Journal, 2004, 39(3): 366−380. https://doi.org/10.1111/j.1365-313X.2004.02138.x doi: 10.1111/j.1365-313X.2004.02138.x [41] Takos A M, Jaffé F W, Jacob S R, et al. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples [J]. Plant Physiology, 2006, 142(3): 1216−1232. https://doi.org/10.1104/pp.106.088104 doi: 10.1104/pp.106.088104 [42] Wang Q, Zuo Z C, Wang X, et al. Beyond the photocycle-how cryptochromes regulate photoresponses in plants? [J]. Current Opinion in Plant Biology, 2018, 45: 120−126. https://doi.org/10.1016/j.pbi.2018.05.014 doi: 10.1016/j.pbi.2018.05.014 [43] Li Y Y, Mao K, Zhao C, et al. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple [J]. Plant Physiology, 2012, 160(2): 1011−1022. https://doi.org/10.1104/pp.112.199703 doi: 10.1104/pp.112.199703 [44] Zhao Y, Min T, Chen M J, et al. The photomorphogenic transcription factor PpHY5 regulates anthocyanin accumulation in response to UVA and UVB irradiation [J]. Frontiers in Plant Science, 2021, 11: 603178. https://doi.org/10.3389/fpls.2020.603178 doi: 10.3389/fpls.2020.603178 [45] Zhao L, Sun J L, Cai Y M, et al. PpHYH is responsible for light-induced anthocyanin accumulation in fruit peel of Prunus persica [J]. Tree Physiology, 2022, 42(8): 1662−1677. https://doi.org/10.1093/treephys/tpac025 doi: 10.1093/treephys/tpac025 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 269
- HTML全文浏览量: 140
- 被引次数: 0
