


-
钾是植物生长发育所必需的一种大量元素,它在多种生理过程中都起着关键作用,包括水分吸收、蒸腾作用、酶激活和光合作用等[1−2]。植物主要通过根系从土壤中吸收钾离子(K+),但土壤中钾元素的分布不均及有效钾含量低下常常成为限制作物生长和产量的关键因素[3]。植物对钾离子的吸收与转运依赖于根系细胞膜上一系列钾离子转运蛋白和通道蛋白[4−5]。目前已发现的K+转运蛋白包括HKT(high-affinity K+ transporter)、KUP/HAK/KT(K+- uptake permease/high-affinity K+ transporter/K+ transporter)、CPAs(cation-protein antiporters)、CHX(cation/H+ exchanger)和KEA(K+-efflux antiporter)等家族[2, 6]。其中,KUP/HAK/KT家族成员最多,作为H+/K+同向转运载体,主要负责植物根部细胞对K+的高亲和吸收,在植物生长发育、逆境响应及信号转导中发挥重要作用[7−10]。目前该家族已在拟南芥[11]、辣椒[12]、水稻[13]、大豆[14]等多种作物中被陆续鉴定。拟南芥中鉴定到13个KUP/HAK/KT基因,其中KUP6亚家族的转运蛋白通过维持细胞生长过程中的钾平衡及应对干旱胁迫反应在调节渗透压方面发挥关键作用[15]。
烟草(Nicotiana tabacum L.)是一种喜钾作物,钾不仅能显著改善烟叶的颜色和燃烧性,还能增加其韧性和柔软度[16]。因此,烟叶中较高的钾含量能提高烟叶的产量和品质。烟草主要通过根系从土壤中吸收K+,随后将K+积累在叶片中。挖掘调控烟草K+吸收和运输的关键基因,对于烟草钾含量的定向改良具有重要意义[17]。目前对烟草钾营养分子机制的研究,特别是对KUP/HAK/KT家族关键成员的功能解析仍相对滞后。尽管已有研究报道了烟草KUP/HAK/KT家族的全基因组鉴定[18−19],但仅有个别基因如NtHAK1[20−22]、NtHAK5[23]和NtKT12[24]等被克隆并进行了表达分析,还有大量其他成员具体的生物学功能尚待通过实验进行验证。
本研究在深入分析EMS诱变获得的高钾氯比烟草株系转录组数据的基础上筛选出NtKUP6基因,以豫中烟区主栽品种中烟100为研究对象,克隆了KUP/HAK/KT家族成员NtKUP6基因,通过生物信息学分析了NtKUP6蛋白特性与启动子顺式作用元件;亚细胞定位分析结果表明,NtKUP6蛋白定位在细胞膜上,并利用qRT-PCR检测了NtKUP6基因的组织表达特性及其对非生物胁迫与ABA的响应模式;进一步通过病毒诱导的基因沉默技术(VIGS)及过表达分析初步探究了NtKUP6基因在烟草钾积累中的生物学功能。为阐明烟草钾吸收与胁迫应答的分子机制提供新的理论依据,并为烟草钾营养性状的遗传改良提供有价值的候选基因。
-
以豫中烟区主栽品种中烟100(Nicotiana tabacum L.)为研究对象。植株于2024年种植在国家烟草基因研究中心的人工气候室,采集旺长期烟株的根、茎、叶和盛花期烟株的花等组织器官,经液氮速冻后,−80℃冰箱保存备用。每个组织至少3个生物学重复。亚细胞定位、瞬时沉默及过表达实验的材料均为本氏烟草。培养条件:温度(26±1)℃,相对湿度60%,16 h光照培养,8 h黑暗培养。
-
中烟100种子消毒后点在1/2 MS固体培养基上,萌发生长2周后选取生长良好且长势相同的幼苗进行非生物胁迫处理。选取175 mmol·L−1 NaCl、200 mmol·L−1甘露醇和4℃分别进行盐、干旱和冷胁迫处理。取样时间为0 、1 、3 、12 、24 、48 和96 h,取样部位为全部整株幼苗,每个处理均设置3个生物学重复,液氮冷冻后置于-80℃超低温冰箱保存。
-
使用Polysaccharide/polyphenol Plant Total RNA Mini Kit(GeneBetter)试剂盒提取烟草中烟100根部总RNA。使用Transcriptor First Strand cDNA Synthesis Kit(Roche)试剂盒反转录获得cDNA用于克隆NtKUP6基因。使用Primer Premier 5.0软件设计NtKUP6基因的扩增引物(表1)。50 μL的基因扩增体系包含cDNA 1 μL、上下游引物各1 μL、dNTP Mixture 6 μL、5×PrimerSTAR GXL Buffer 10 μL、PrimeSTAR GXL DNA Polymerase 1 μL及ddH2O 30 μL。PCR反应程序为98℃变性10 s,55℃退火15 s,68℃延伸30 s,35个循环。PCR产物经1%的琼脂糖凝胶电泳检测后,将胶回收纯化的PCR产物连接至pMD18-T载体后转化大肠杆菌DH5α感受态。经PCR验证的阳性克隆菌送北京六合华大基因科技有限公司测序。
表 1 本研究所用引物信息
Table 1. Primer sequences used in this study
引物名称 Primer 引物序列(5'-3') Sequence 用途 Usage NtKUP6-F ATGGATCTTGAAACAGGGGT For cloning vector NtKUP6-R TCATACATTGTAAACCATT NtKUP6-qF GACTGTTGTTGGAACGCCTTCT For qRT-PCR NtKUP6-qR TCTTTACGCTCTGATGGTCC Nt26S-qF GAAGAAGGTCCCAAGGGTTC Nt26S-qR TCTCCCTTTAACACCAACGG NtKUP6-VIGS-F AGATCGACGACAAGACCCTGCAGGTCATGCTGGTTACGACTTGC For VIGS vector construction NtKUP6-VIGS-R TTCTGAGGAGAAGAGCCCTGCAGGATTGCTGGGATTCCGGATACAAG -
利用ProtParam(
https://web.expasy.org/protparam/ )对NtKUP6蛋白进行理化性质分析。利用TMHMM-2.0(https://services.healthtech.dtu.dk/services/TMHMM-2.0/ )对NtKUP6蛋白进行跨膜结构预测。利用Plant-mPLoc在线工具 (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )对NtKUP6蛋白进行亚细胞定位预测。利用NetPhos-3.1(https://services.healthtech.dtu.dk/services/NetPhos-3.1/ )对NtKUP6蛋白进行磷酸化位点预测。利用SOPMA(https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html )对NtKUP6蛋白进行二级结构预测。利用SWISS-MODEL(https://swissmodel.expasy.org/ )对NtKUP6蛋白进行三级结构预测。利用PlantCARE(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )对NtKUP6基因的上游2000 bp启动子区进行顺式作用元件分析。 -
将NtKUP6基因克隆至pC1300s-35S-GFP载体上构建重组表达载体pC1300s-35S-NtKUP6-GFP,转化农杆菌(Agrobacterium tumefaciens)GV3101菌株,注射本氏烟草,暗光培养48 h后使用激光共聚焦显微镜观察GFP荧光信号。
-
选取旺长期的根、茎、叶和盛花期的花等4种不同组织的样品,液氮速冻后-80℃保存。提取不同组织样品的RNA,反转录为cDNA,以此为模板进行NtKUP6基因在各组织的表达分析。另提取保存的非生物胁迫处理样品的RNA,反转录cDNA后,以此为模板进行NtKUP6基因在非生物胁迫下的表达分析。以Nt26S为内参基因,用Primer Premier 5.0软件设计引物(表1),进行实时荧光定量PCR。反应体系按照罗氏的FastStart Universal SYBR Green Master(ROX)预混液说明书配制。反应程序:94℃预变性30 s;94℃变性5 s,60℃退火20 s,72℃延伸10 s,45个循环。每个样品设置3个重复,使用的荧光定量PCR仪型号是Roche LightCycler 96,运用2-ΔΔCt方法计算基因相对表达量。
-
根据NtKUP6基因序列设计特异性引物NtKUP6-VIGS-F/ NtKUP6-VIGS-R(表1),以根部cDNA为模板,通过PCR扩增得到目的片段。将目的片段连接到pTRV2载体上,转化大肠杆菌DH5α,测序获得阳性克隆TRV2-NtKUP6后,提取质粒转化GV3101农杆菌感受态。
瞬时沉默方法参考文献[25],选取叶片大小一致的本氏烟,将菌液注满展平的叶片,注射后的烟苗黑暗培养过夜后恢复至正常光照培养。注射一周左右pTRV2-PDS(阳性对照)的烟苗出现白化表型,注射两周左右选取长势较好的6株新生叶片,液氮速冻后,−80℃保存备用,后续进行NtKUP6基因的定量分析及钾离子含量测定。
-
利用引物NtKUP6-F/NtKUP6-R(表1)将NtKUP6基因克隆至表达载体pDT1上,测序获得正确的pDT1-NtKUP6重组质粒,转化GV3101农杆菌感受态。将注射菌液于室温避光静置3~5 h,注射本氏烟,空载体菌株为对照。注射后的烟苗黑暗培养过夜后恢复至正常光照培养,注射后72 h采集长势较好的注射叶片每种6株,进行后续基因表达检测和钾离子含量测定。
-
将保存的VIGS植株及过表达叶片样品冷冻干燥后,使用混合型振荡研磨仪研磨至粉末状,称取约
0.0500 g(精确至0.1 mg)粉末,置于10 mL 5%(体积分数)醋酸中,30 ℃震荡萃取30 min,然后用定性滤纸过滤,滤液经稀释后用美国安莱立思Alalis MP6500型台式pH计测定钾离子含量。 -
本研究所有试验均设置至少3个生物学重复,试验原始数据利用Excel进行整理,然后使用Graph Pad Prim 8.0软件进行图表绘制,所有实验结果均以平均值±标准差(Mean±SD)表示。对两个独立样本的比较采用独立样本t检验分析组间差异显著性,对多组样本比较采用单因素方差分析(One-way ANOVA)结合Duncan氏多重比较法分析组间差异。
-
以烟草中烟100根部cDNA为模板,经PCR扩增,PCR产物的琼脂糖电泳检测结果如图1所示,获得了一条大小约2500bp的DNA片段。目的片段回收后测序,分析发现该段序列包含一个完整的开放阅读框(ORF),全长
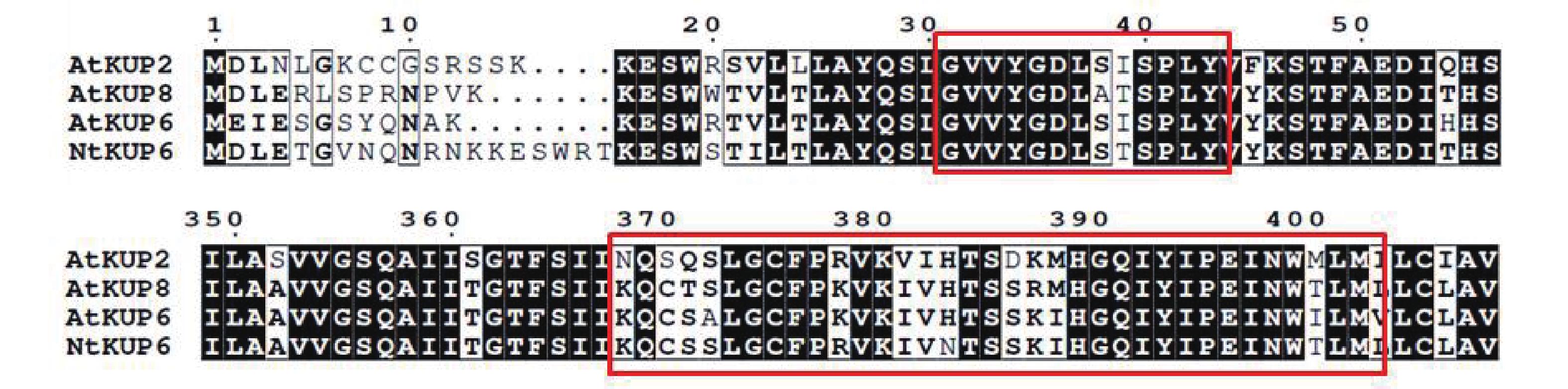
2355 bp,编码784个氨基酸。使用Clustal W将NtKUP6与NCBI中下载拟南芥KUP6亚家族蛋白进行比对分析,发现NtKUP6具有KUP/HAK/KT家族的保守区域GXXXGDXXXSPLY和KQXXALGCFPKXKIVHTSXKXXGQIYIPENWILM [26](图2)。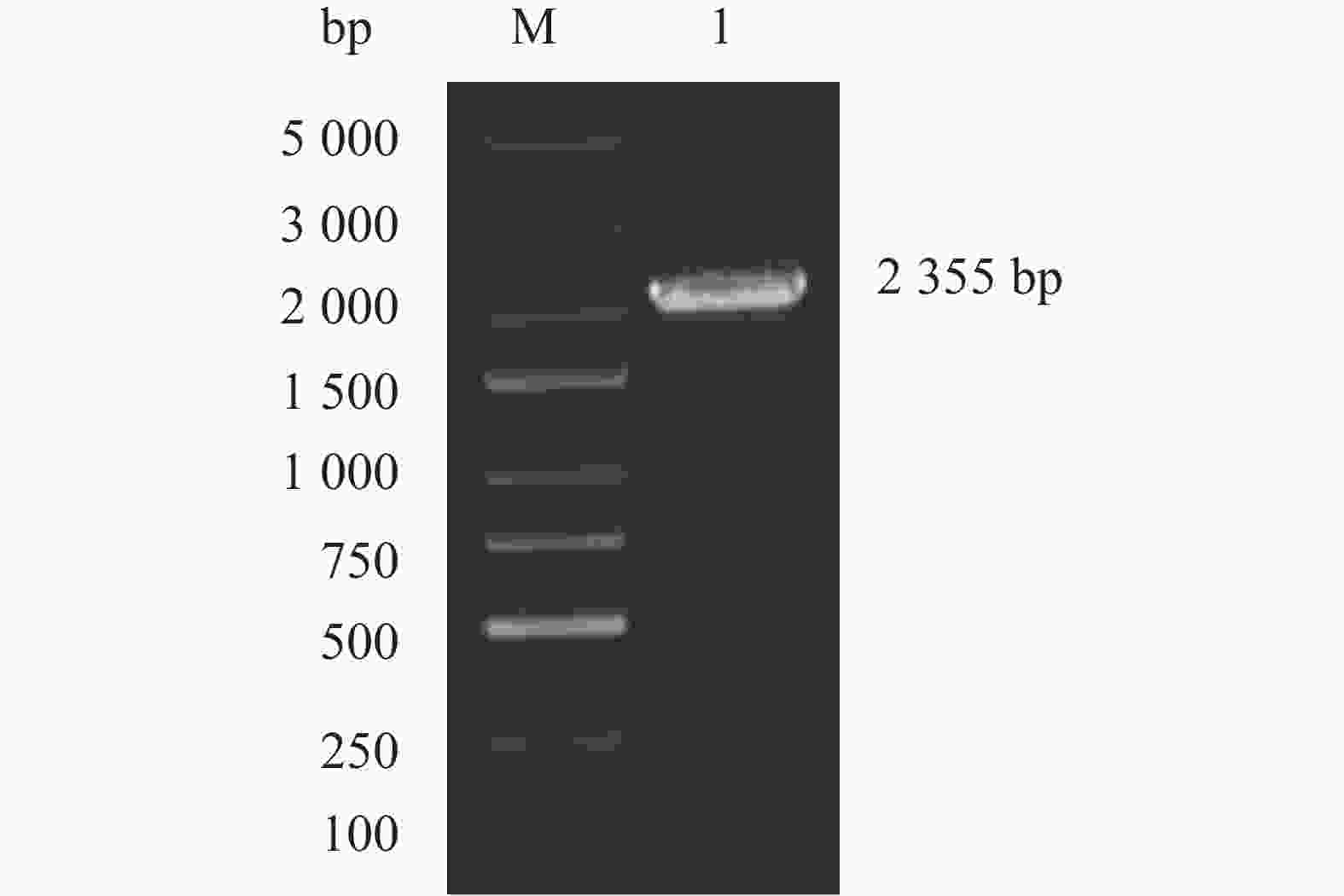
图 1 烟草NtKUP6基因的PCR扩增结果
Figure 1. Electropherogram of the full-length sequence of NtKUP6 gene in Nicotiana tabacum
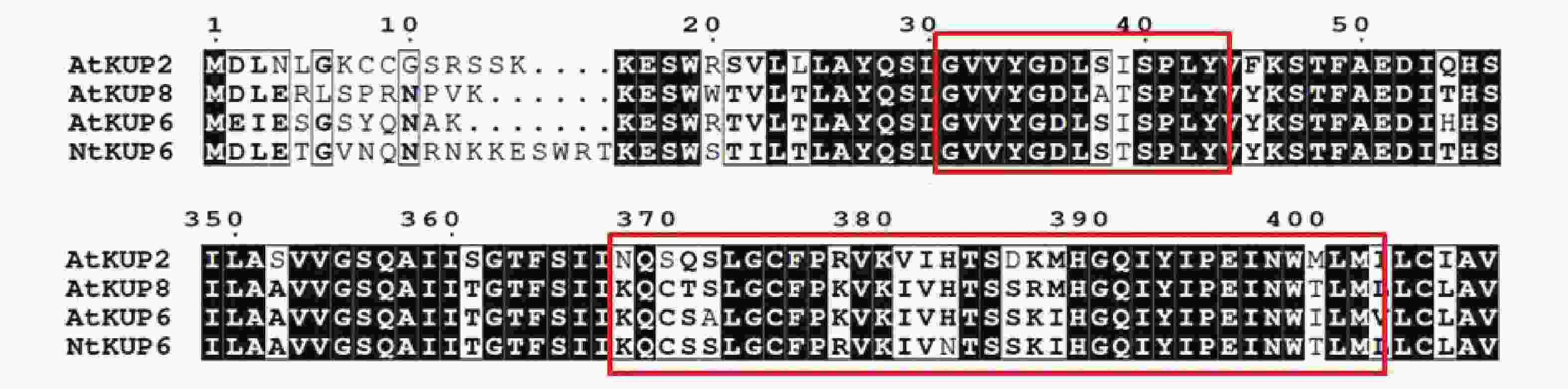
图 2 烟草NtKUP6与拟南芥KUP6亚家族氨基酸序列的多重比对
Figure 2. Alignment of amino acid sequence of NtKUP6 and KUP6 subfamily members in Arabidopsis thaliana
-
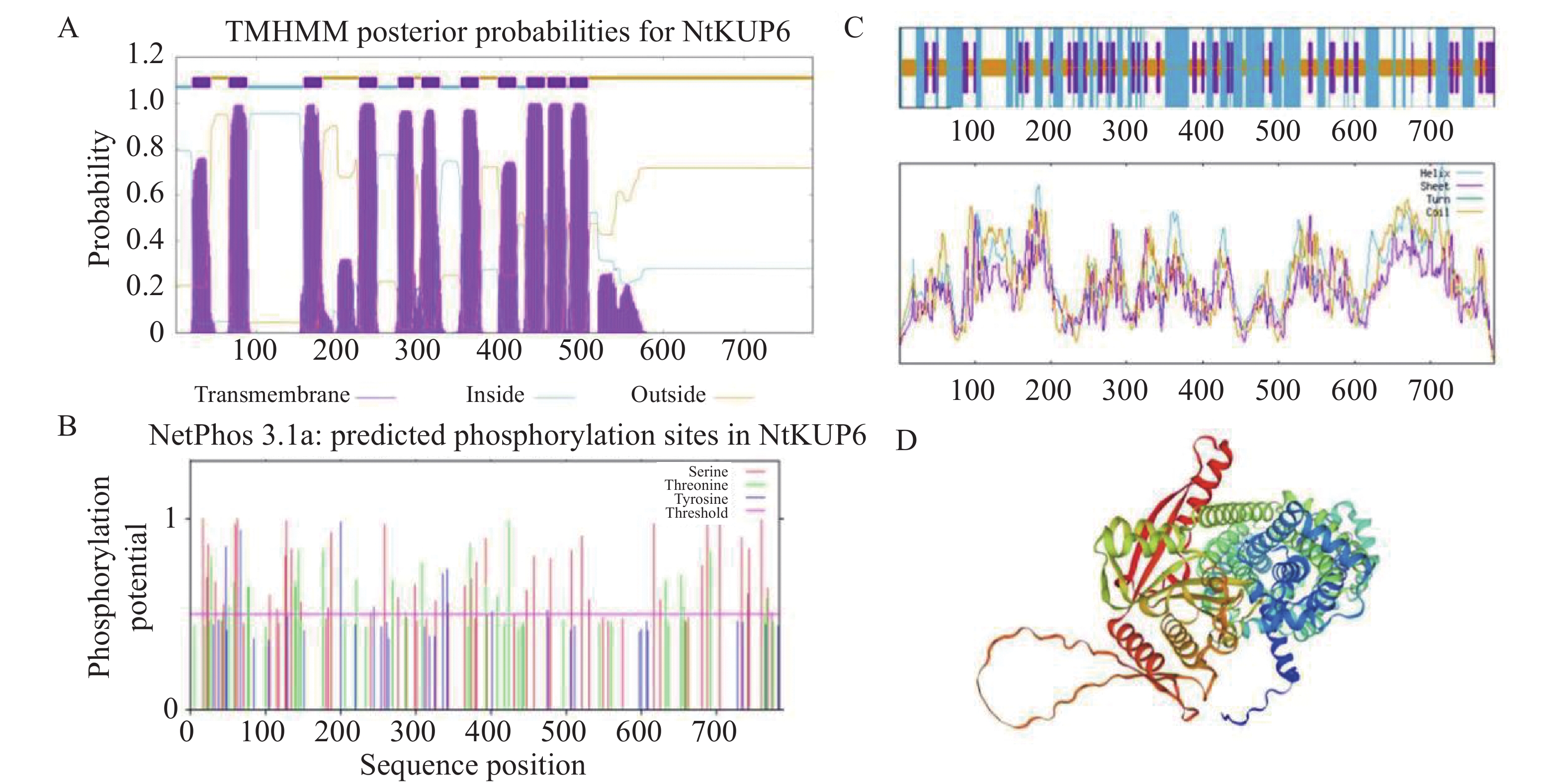
使用在线分析工具ProtParam对NtKUP6蛋白序列进行分析,该蛋白相对分子质量为87.9 kDa,理论等电点为8.76,不稳定系数为37.48,说明该蛋白属于稳定蛋白。NtKUP6蛋白包含了20种氨基酸,含量最高的是亮氨酸(11.2%),含量最低的是色氨酸(1.4%)。脂肪族氨基酸指数为107.86,具有高脂肪族性质。利用TMHMM-2.0对NtKUP6蛋白进行跨膜结构分析,结果显示,该蛋白共有11个跨膜结构域(图3-A),其中第2、3跨膜区之间存在1个胞质环(Loop)结构,C端具有较长的胞外区。利用Plant-mPLoc预测NtKUP6蛋白的亚细胞定位于细胞膜、液泡。利用NetPhos-3.1对NtKUP6蛋白进行磷酸化位点预测,共发现44个丝氨酸、23个苏氨酸和7个酪氨酸位点(图3-B)。二级结构分析显示,α-螺旋(40.82%,320个氨基酸)和无规则卷曲(41.71%,327个氨基酸)是构成NtKUP6蛋白的主要结构元件,延伸链(17.47%,137个氨基酸)则分散在整个蛋白中(图3-C)。进一步三级结构预测以V4VGN3.1.A柑橘(Citrus clementina)钾离子通道蛋白三级结构(AlphaFold DB model)为参考模型,相似度为82.63%,GMQE值为0.77,蛋白含有丰富的无规则卷曲和α-螺旋(图3-D),与二级结构预测结果一致。
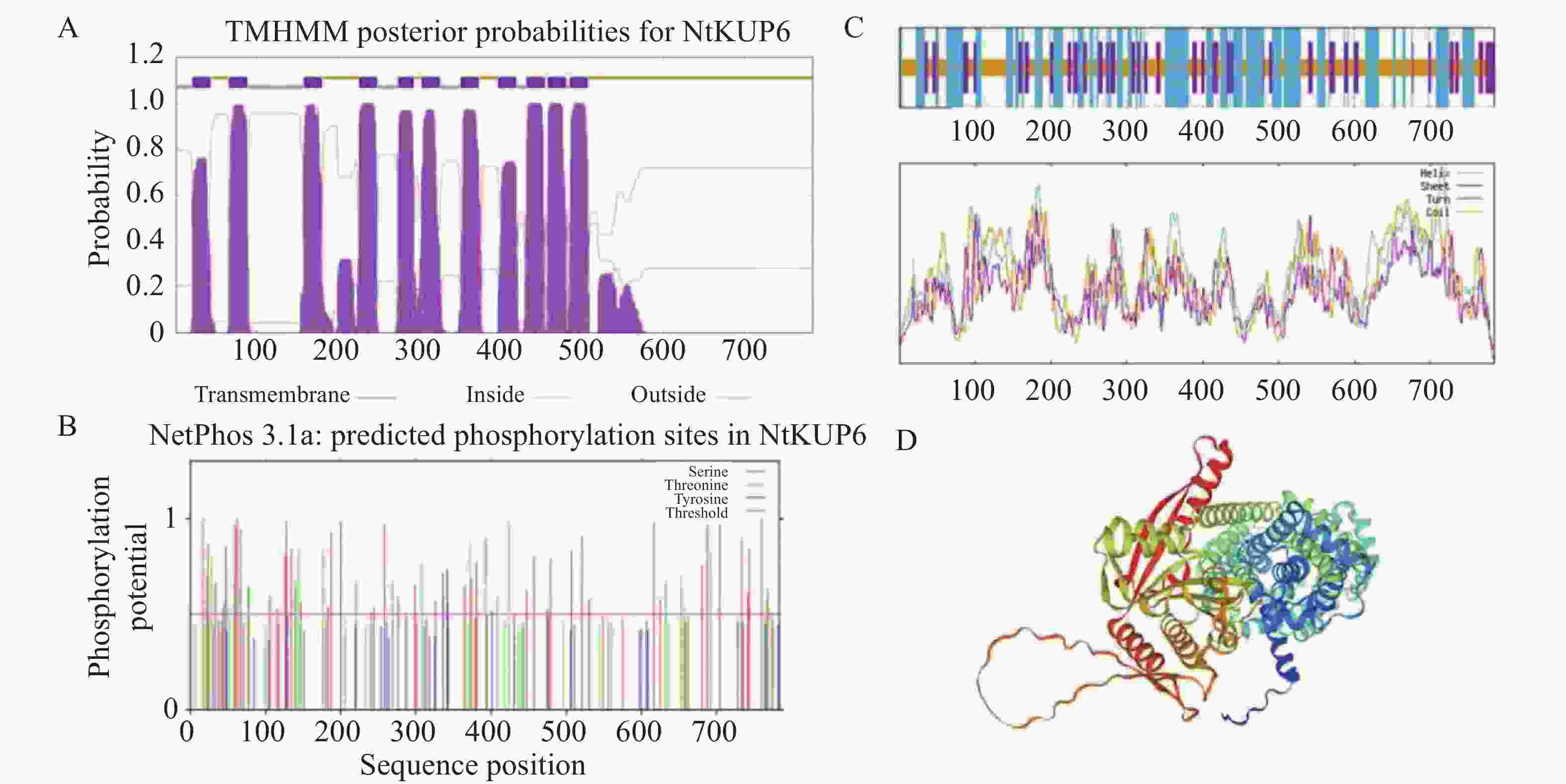
图 3 NtKUP6蛋白生物信息学分析
Figure 3. Bioinformatics analysis of the NtKUP6 protein in tobacco
顺式作用元件分析结果如表2所示,NtKUP6基因启动子区含有干旱响应的MBS元件和防御应激响应的TC-rich repeats元件, ABA响应的ABRE元件和GA响应的P-box元件,以及参与光响应的AT1基序等作用元件。
表 2 NtKUP6基因启动子区顺式作用元件
Table 2. The cis-acting elements in the promoter region of the NtKUP6 gene
种类
Function元件
Elements描述
Descriptions数量
Quantities胁迫相关
Stress-relatedMBS 参与干旱诱导性的MYB结合位点
(MYB binding site involved in drought-inducibility)1 TC-rich repeats 防御和应激反应元件
(cis-acting element involved in defense and stress responsiveness)3 调节相关
Regulate-relatedCAAT-box 启动子和增强子普遍存在的元件
(common cis-acting element in promoter and enhancer regions)38 TATA-box 据转录起始位点30个碱基的中心元件
(core promoter element around -30 of transcription start)36 光相关
Light-relatedAT1-motif 光响应元件(light responsive element) 1 ATCT-motif 光响应元件(light responsive element) 1 Box 4 光响应元件(light responsive element) 3 G-box 光响应元件(light responsive element) 1 GT1-motif 光响应元件(light responsive element) 1 MRE 光响应元件(light responsive element) 1 TCCC-motif 光响应元件(light responsive element) 1 TCT-motif 光响应元件(light responsive element) 1 生长相关
Growth-relatedARE 厌氧响应元件(light responsive element) 3 GC-motif 缺氧特异性诱导元件
(enhancer-like element involved in anoxic specific inducibility)1 激素相关
Hormone-relatedABRE 脱落酸应答元件
(cis-acting element involved in the abscisic acid responsiveness)2 P-box 赤霉素响应元件
(gibberellin-responsive element)1 -
为明确NtKUP6蛋白的亚细胞定位,本研究构建了NtKUP6-GFP融合表达载体,转化农杆菌后注射本氏烟叶片,以细胞膜mCherry为marker,通过激光共聚焦显微镜观察绿色荧光信号,结果如图4所示,NtKUP6-GFP融合蛋白在细胞边缘呈现清晰的环状绿色荧光信号,与膜蛋白marker蛋白mCherry的红色荧光信号叠加后呈现黄色荧光,表明NtKUP6蛋白定位于细胞膜。

图 4 NtKUP6蛋白亚细胞定位
Figure 4. Subcellular localization of NtKUP6 protein
-
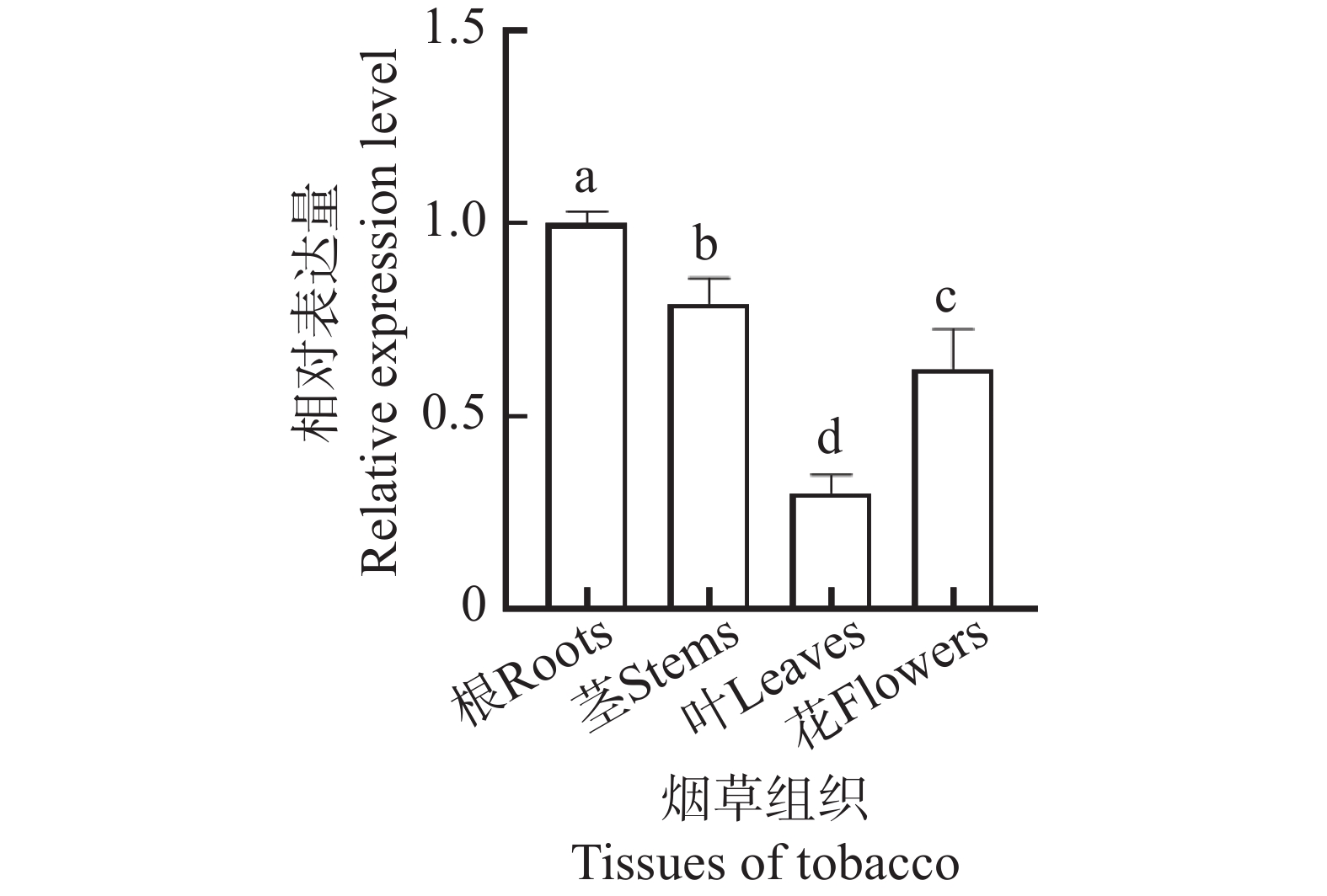
对NtKUP6基因在烟草根、茎、叶、花等组织器官的表达进行了qRT-PCR分析,结果如图5所示,NtKUP6基因在不同组织器官均有表达,且根部相对表达量最高。
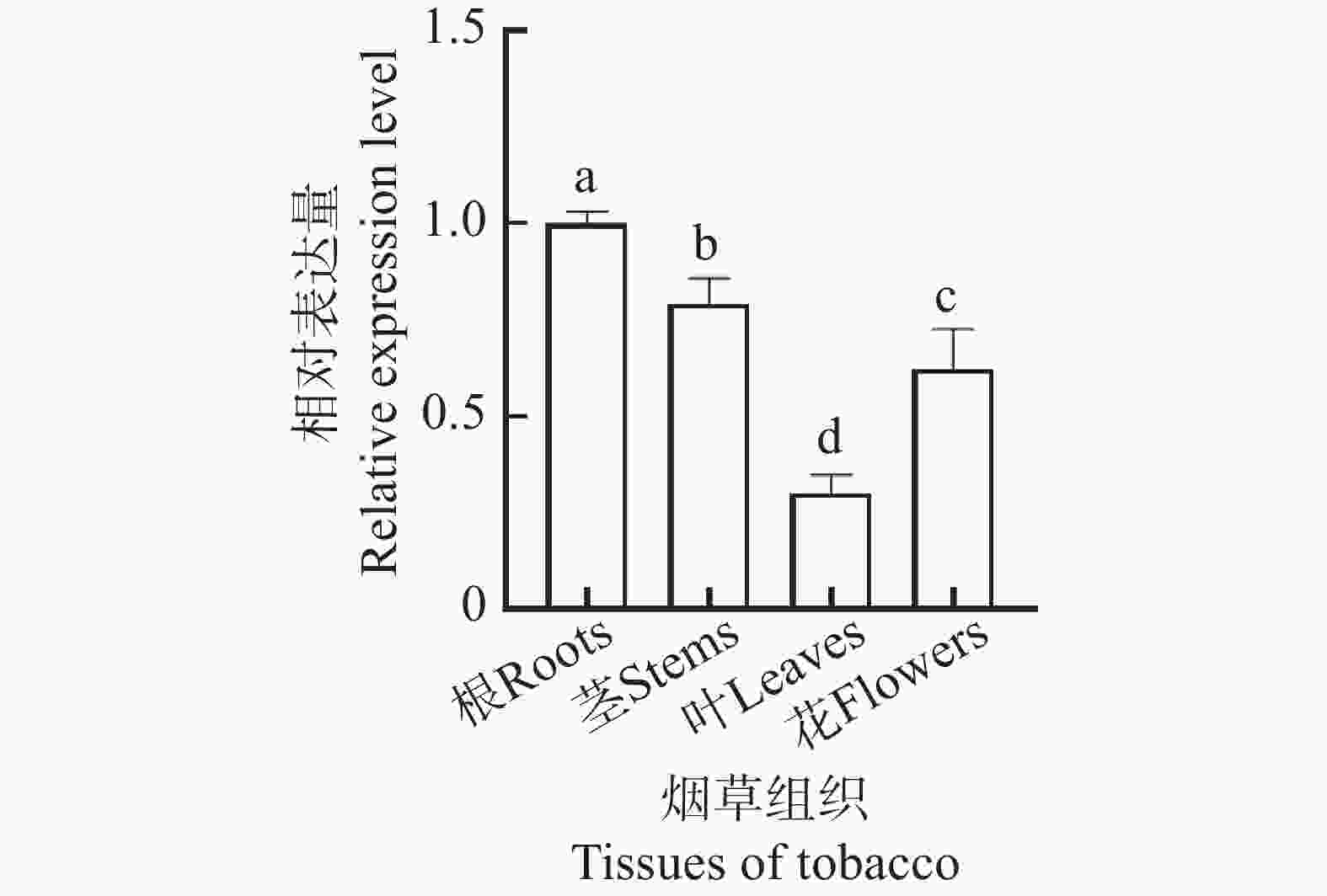
图 5 NtKUP6基因组织表达分析
Figure 5. Tissue expression analysis of NtKUP6 gene
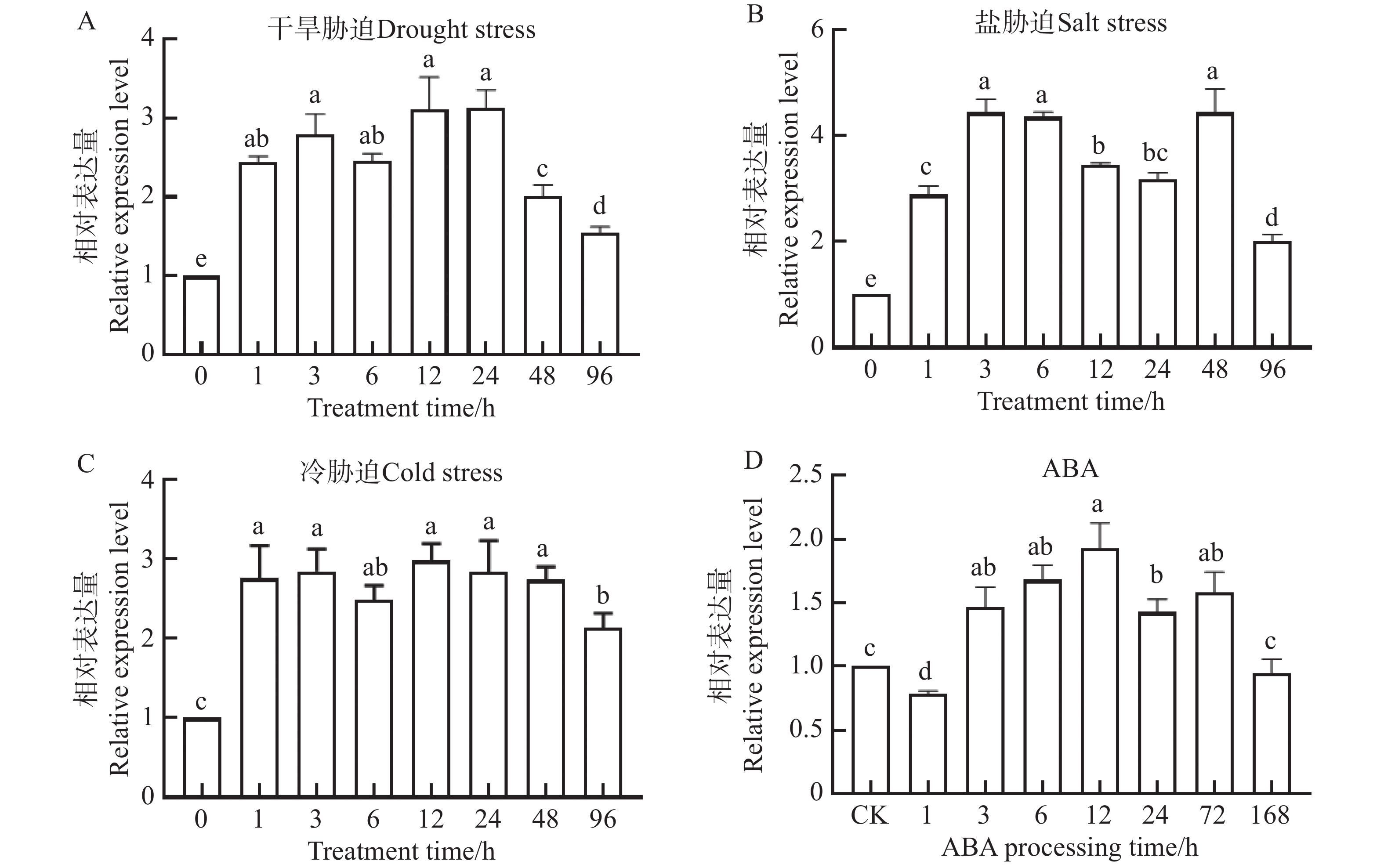
启动子分析表明,NtKUP6基因启动子含有干旱等胁迫响应元件及脱落酸(Abscisic Acid, ABA)等激素响应元件,提示该基因可能参与干旱等非生物胁迫应答及ABA信号调控。为验证这一可能性,利用qRT-PCR检测了NtKUP6基因在干旱、盐、低温及ABA处理下的表达情况,结果如图6所示。在干旱胁迫处理下,NtKUP6基因表达显著升高,在处理12 ~24 h维持较高水平后,随着处理时间的增加NtKUP6基因的表达量又迅速下降;在盐胁迫处理下,NtKUP6基因表达量显著升高,处理3 h表达量达到最高;冷胁迫处理下,随着处理时间的增加,NtKUP6基因表达量一直维持在较高水平;在ABA处理下,NtKUP6基因表达量与干旱处理类似,经历一个先升高再降低的过程。总之,NtKUP6基因对各种非生物胁迫及ABA处理均有响应。
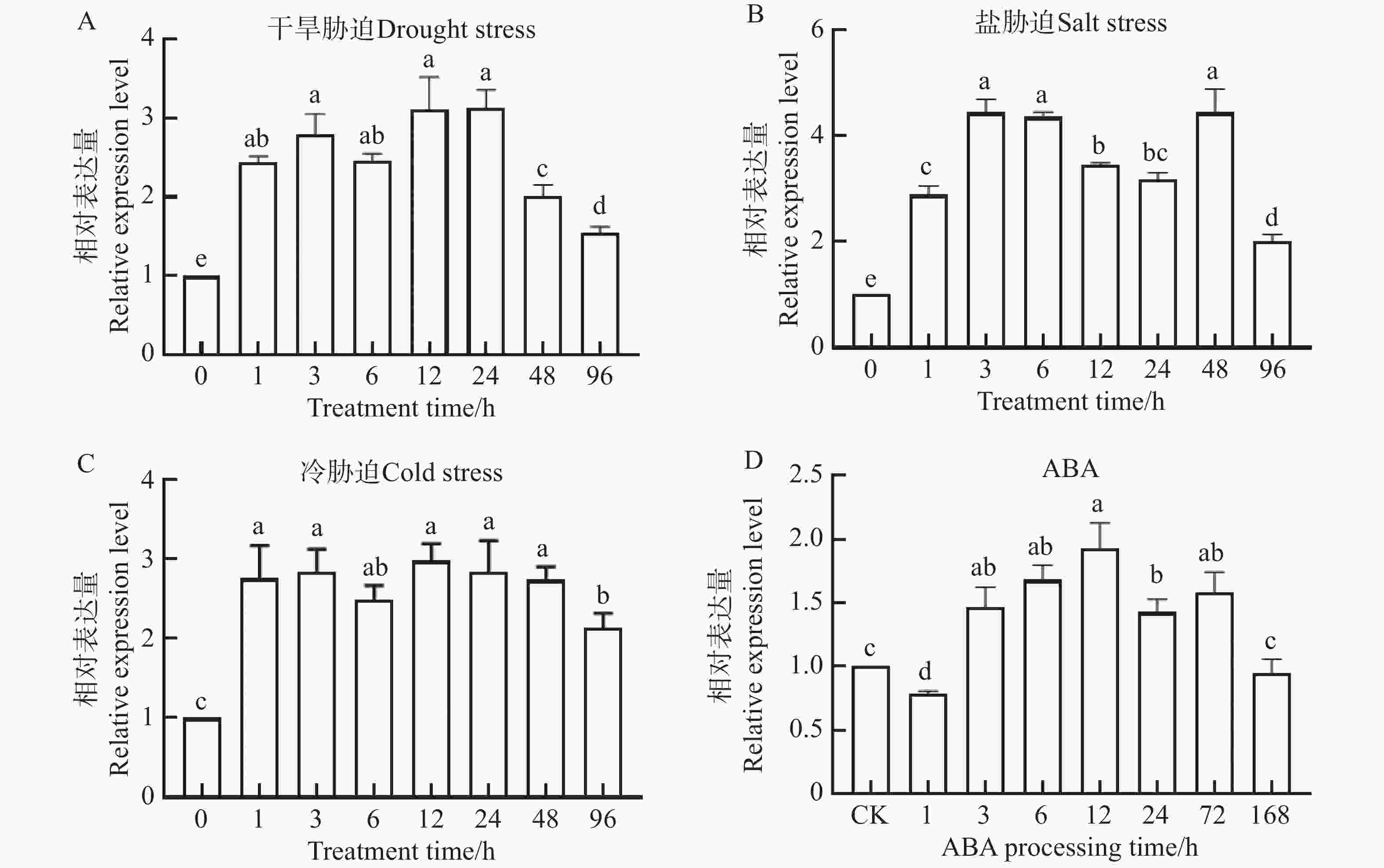
图 6 NtKUP6基因在非生物胁迫及ABA处理下表达分析
Figure 6. Expression analysis of the NtKUP6 gene under abiotic stress and ABA treatment
-
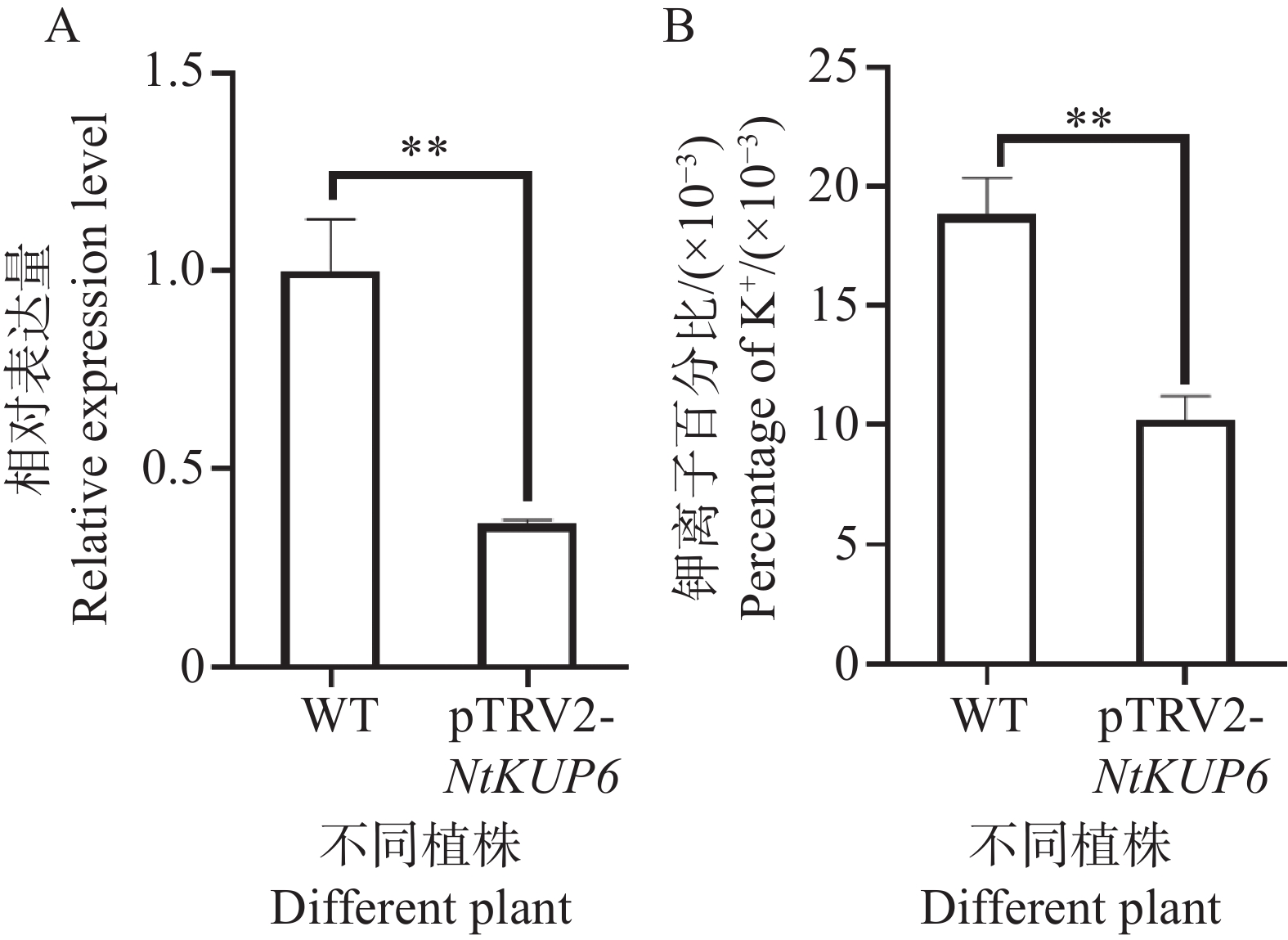
为了进一步明确NtKUP6基因的功能,构建了NtKUP6基因的VIGS瞬时沉默载体,转化农杆菌后注射本氏烟,生长10 d左右取新长出的叶片检测沉默效率并测定叶片钾离子含量,结果表明,相对于pTRV2,pTRV2:NtKUP6沉默植株中NtKUP6基因的表达量降低了69%(图7-A),且沉默植株叶片钾离子含量降低了39.3%(图7-B)。
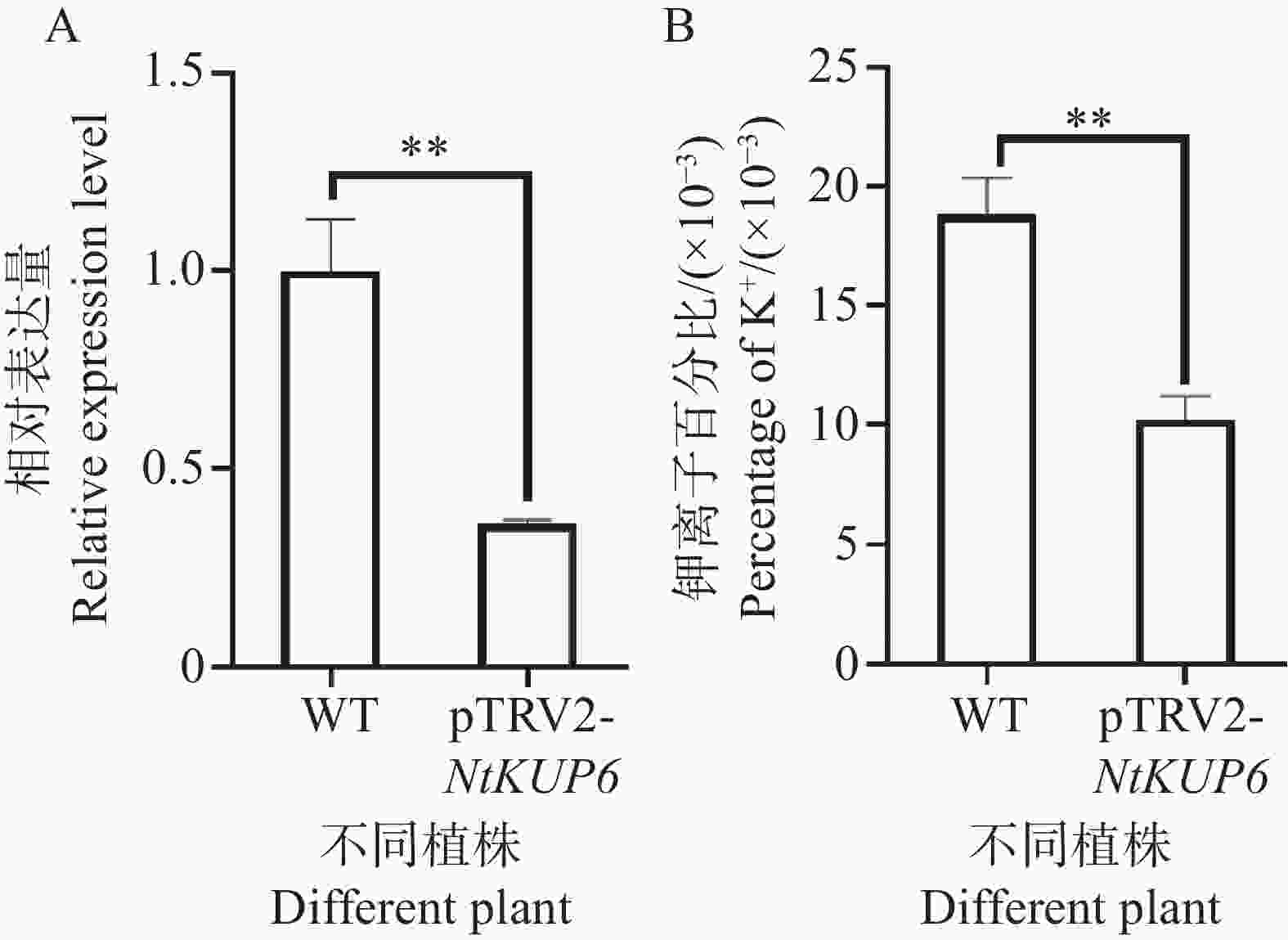
图 7 NtKUP6基因VIGS沉默植株钾离子含量测定
Figure 7. Determination of K+ contents in NtKUP6 gene VIGS silenced plants
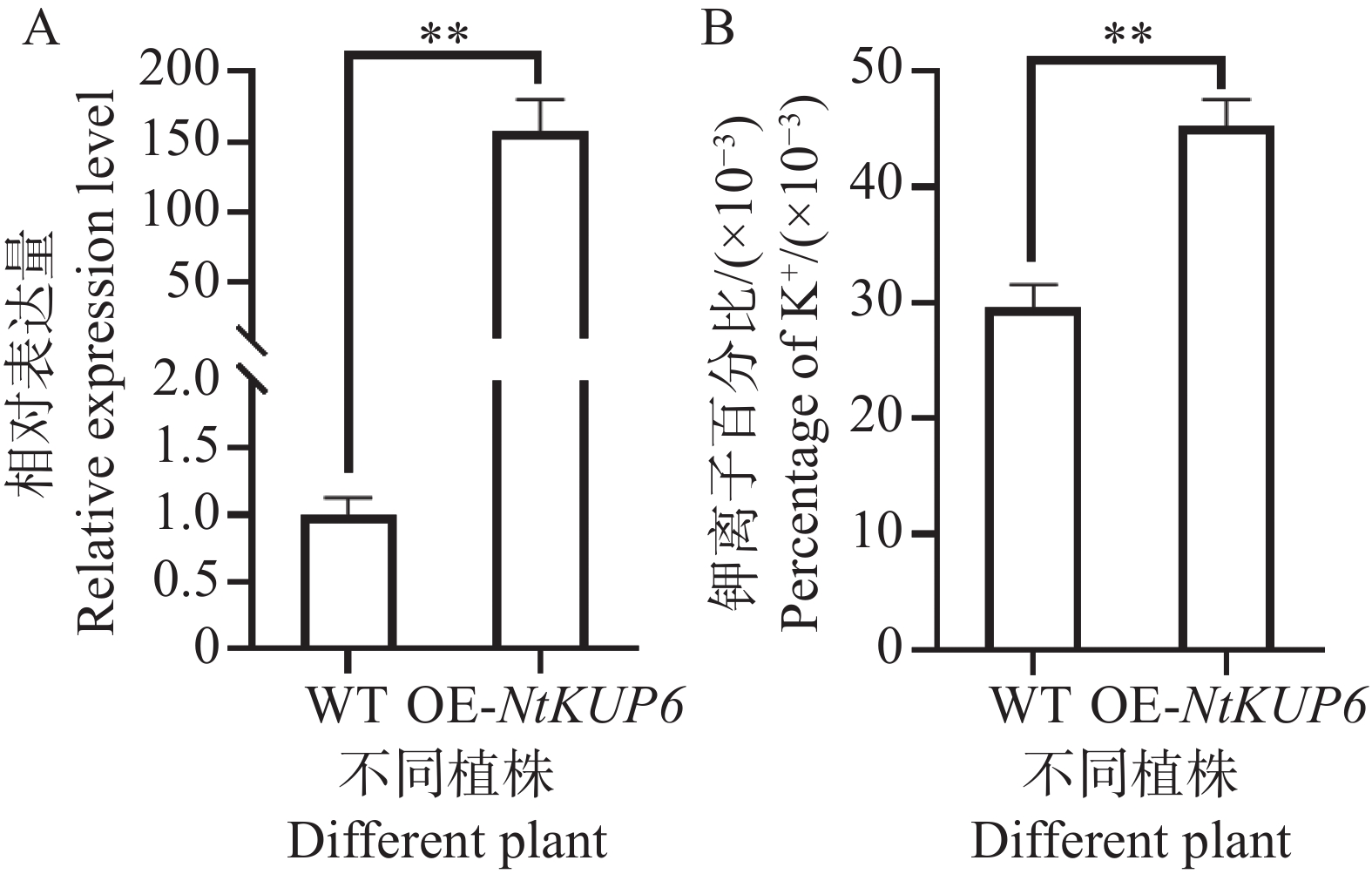
对NtKUP6基因在本氏烟中过表达,分别在本氏烟注射后72 h采样注射叶片,对采集的叶片进行基因表达检测和钾离子含量测定,结果如图7所示。与对照相比,过表达植株叶片中NtKUP6基因的表达量升高了近160倍(图8-A),且过表达植株叶片钾离子含量升高了约53%(图8-B)。
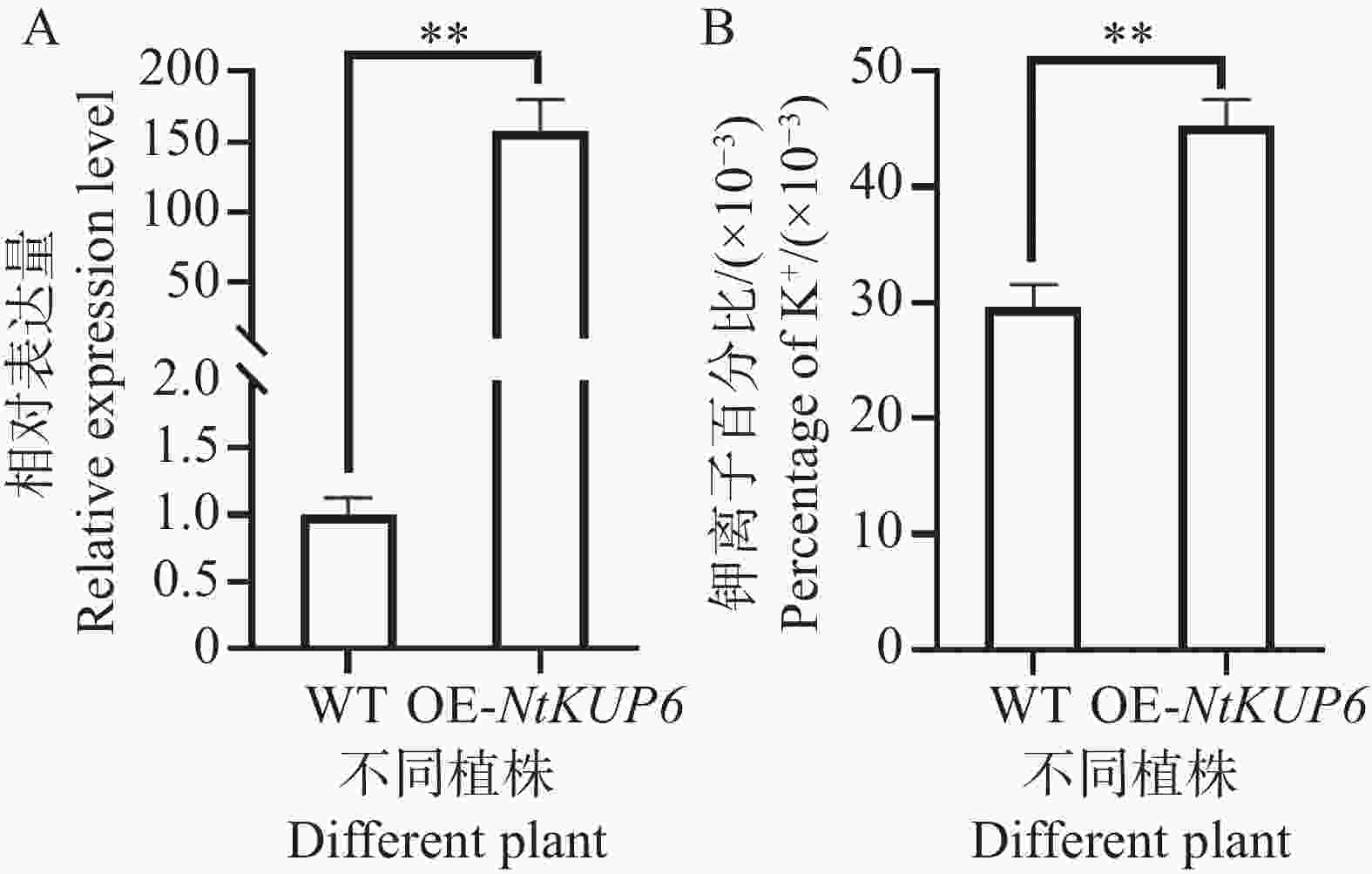
图 8 NtKUP6基因在本氏烟的过表达植株钾离子含量测定
Figure 8. Determination of K+ contents in NtKUP6 gene overexpression plants
-
钾离子转运蛋白在植物维持K+稳态和应对非生物胁迫过程中发挥着核心作用[15]。其中,KUP/HAK/KT家族是负责高亲和性吸收的关键成员。本研究从烟草中烟100中成功克隆了NtKUP6基因,并对其功能进行了初步探索。序列分析结果表明,NtKUP6蛋白具有KUP/HAK/KT家族的典型保守结构域,并与拟南芥KUP6亚家族成员高度同源[27],说明NtKUP6在进化上的保守性和功能上的相似性。生物信息学分析结果表明,NtKUP6是一个稳定的疏水性蛋白,具有11个预测的跨膜结构域,这与NtKUP6作为膜定位转运蛋白的功能定位相符。进一步的亚细胞定位结果预测了NtKUP6可能定位于细胞膜、液泡膜,说明该蛋白不仅负责从外界吸收K+,也可能参与细胞内K+的区隔化储存。这个暗示在离子稳态调节中具有重要意义。
组织表达模式分析结果表明,NtKUP6基因在烟草根部的相对表达量最高,且在茎、叶、花等组织中均有一定表达(图4),这与其他物种中多数KUP/HAT/KT家族成员的表达模式相似[7,10],体现了该家族基因功能的进化保守性。烟草根系是钾离子吸收的主要器官,NtKUP6在根中高表达的特性,与NtKUP6作为HAK/KUP/KT家族成员参与根部高亲和钾吸收的功能定位一致,这说明NtKUP6基因是烟草从土壤中获取K+的核心转运蛋白。而NtKUP6基因在地上部组织的广泛表达,推测NtKUP6可能同时参与K+在植物体内的长距离运输及不同组织中的分配,为叶片等经济器官的钾积累提供保障。
多项研究表明HAK/KUP/KT蛋白与非生物胁迫有关[5]。如拟南芥kup268突变体对干旱胁迫敏感,且KUP6受到ABA信号通路的直接调控,通过平衡K+稳态实现渗透调节进而抵御干旱胁迫[28]。在水稻中过表达OsHAK5能够增加烟草BY2细胞中的K+积累并增强耐盐性[29]。对NtKUP6基因启动子区进行分析发现,含有干旱响应的MBS元件、ABA响应的ABRE元件以及防御与应激相关的TC-rich repeats等多种顺式作用元件(表2),这为NtKUP6基因该基因响应非生物胁迫及激素信号提供了分子基础。随后的qRT-PCR实验结果证实了这一推测,NtKUP6基因对干旱、盐、冷及外源ABA处理均呈现显著的响应特征,但不同胁迫下的表达模式存在明显差异。在干旱与ABA处理下,NtKUP6基因表达呈“先升后降“趋势,推测NtKUP6基因在胁迫早期通过快速上调表达增强钾吸收与稳态维持,以提升细胞渗透压、减少水分流失,而后期表达下调可能是植物通过反馈调节避免钾过量积累造成的代谢负担,且ABA响应元件ABRE的存在说明NtKUP6基因可能通过ABA信号通路介导干旱胁迫的适应性反应。在盐胁迫下,NtKUP6基因在处理3 h表达量达到峰值,后续逐渐下降,这一快速响应特征可能与短期盐胁迫下植物快速积累K+以平衡Na+毒性、维持细胞膜稳定性相关,体现了NtKUP6基因在盐胁迫早期离子平衡调节中的关键作用。在冷胁迫下,NtKUP6基因表达持续维持高水平,推测K+作为渗透调节物质和酶活性保护剂,NtKUP6的持续高表达可通过稳定细胞内钾浓度,减轻低温对细胞膜结构和代谢酶活性的损伤,这与冷胁迫下植物对钾稳态的长期需求相符,也为烟草低温逆境适应性提供了新的调控线索。
为了验证NtKUP6在烟草K+积累中的功能,通过VIGS技术获得瞬时沉默植株,结果显示沉默植株NtKUP6的表达量降低69%,叶片中K+含量也随之下降39.3%;而本氏烟中过表达NtKUP6后,基因表达量升高近160倍,叶片K+含量显著提升约53%。这一结果与水稻OsHAK16的功能类似[30],证明NtKUP6基因对于烟草叶片中的K+积累具有关键调控的作用。结合NtKUP6基因的组织表达与胁迫响应特征,基因瞬时沉默后叶片K+含量的显著降低,可能与根系钾吸收能力下降或体内K+转运受阻有关。根中高表达保障基础钾吸收,胁迫诱导表达强化逆境下的钾稳态调节,最终通过调控钾积累实现对烟草生长发育及抗逆性的支撑。
当然,本研究仍存在一定局限性,NtKUP6在木质部装载或韧皮部再转运中的具体作用尚未明确,NtKUP6与其他钾转运蛋白的协同调控机制也有待探究。未来通过CRISPR或过表达等方法获得稳定遗传突变体,可更全面地评估该基因在烟草整个生育期中对钾吸收、分配及抗逆性的综合影响;同时结合转录组测序、酵母双杂交、非损伤微测(NMT)等技术,进一步解析NtKUP6参与的调控通路及互作蛋白,为深入阐明烟草钾营养与抗逆协同调控的分子机制提供更充分的依据。
Cloning and functional analysis of the tobacco potassium transporter NtKUP6 Gene
-
摘要: 钾是影响烟草产量与品质的关键元素。为探究烟草钾转运蛋白基因NtKUP6的功能,本研究以烟草(中烟100)为材料,克隆获得编码区全长为
2355 bp的NtKUP6基因。生物信息学分析结果表明,NtKUP6基因编码的蛋白具有11个跨膜结构域,且启动子区含有MBS、ABRE等与非生物胁迫及激素响应相关的顺式作用元件。亚细胞定位表明NtKUP6蛋白定位在细胞膜上。表达模式分析结果显示,NtKUP6基因在根中表达量最高,并能被干旱、高盐、低温和ABA处理显著诱导。功能验证方面,通过病毒诱导的基因沉默技术(VIGS)沉默NtKUP6基因表达后,烟草叶片中的钾离子含量显著降低(约39.3%);在本氏烟中过表达NtKUP6基因,烟草叶片中的钾离子含量显著升高(53%)。综上,NtKUP6是一个在烟草钾积累及非生物胁迫响应中发挥重要功能的钾转运蛋白基因,为解析烟草钾营养分子机制提供了理论依据。Abstract: Potassium is a key element affecting the yield and quality of tobacco. To investigate the function of the tobacco potassium transporter gene NtKUP6, the NtKUP6 gene with a full coding sequence length of 2,355 bp was cloned from tobacco Zhongyan 100 (Nicotiana tabacum L.). Bioinformatics analysis indicated that its encoded protein possesses 11 transmembrane domains, and that its promoter region contains cis-acting elements such as MBS and ABRE, which are associated with abiotic stress and hormone responses. Subcellular localization indicated that NtKUP6 was located on the cell membrane. Expression pattern analysis revealed that this gene is most highly expressed in the roots and can be significantly induced by drought, high salt, low temperature, and ABA treatments. For functional verification, transient interference with NtKUP6 expression via virus-induced gene silencing (VIGS) resulted in a significant reduction of potassium ion content in tobacco leaves by 39.3%, whereas overexpression of NtKUP6 gene in N. benthamiana resulted in a 53% increase of potassium ion content in tobacco leaves. In conclusion, NtKUP6 is a potassium transporter gene that plays an important role in potassium accumulation and abiotic stress response in tobacco, providing a theoretical basis for elucidating the molecular mechanisms of potassium nutrition in tobacco.-
Key words:
- Tobacco /
- potassium transporter protein /
- NtKUP6 /
- abiotic stress /
- potassium accumulation
-
图 2 烟草NtKUP6与拟南芥KUP6亚家族氨基酸序列的多重比对
Fig. 2 Alignment of amino acid sequence of NtKUP6 and KUP6 subfamily members in Arabidopsis thaliana
图 5 NtKUP6基因组织表达分析
注:NtKUP6基因在根中的表达量设置为1.0。所有数据至少包含3个生物重复,每个生物重复中大约含有3个不同植株组织样品。采用单因素方差分析(One-way ANOVA)结合Duncan氏多重比较法分析差异显著性,不同字母表示不同组织的差异显著性(P< 0.05)。
Fig. 5 Tissue expression analysis of NtKUP6 gene
Note: The expression level of the NtKUP6 gene in the root was set as 1.0. All data included at least three biological replicates, with approximately three different plant tissue samples in each biological replicate. One-way ANOVA combined with Duncan's multiple range test was used to analyze the significance of differences. Different letters indicated significant differences among different tissues (P< 0.05).
图 6 NtKUP6基因在非生物胁迫及ABA处理下表达分析
注:A. 干旱胁迫;B. 盐胁迫;C. 冷胁迫;D. ABA处理。NtKUP6基因在0 h的表达量设置为1.0。所有数据至少包含三个生物重复,每个生物重复中约有10株小幼苗。采用单因素方差分析(One-way ANOVA) 结合 Duncan 氏多重比较法分析差异显著性,不同字母表示P< 0.05的差异显著性
Fig. 6 Expression analysis of the NtKUP6 gene under abiotic stress and ABA treatment
Note: A; Drought stress; B: Salt stress; C: Cold stress; D: ABA treatment. The expression level of the NtKUP6 gene at 0 h was set to 1.0. All data included at least three biological replicates, and each biological replicate contained approximately 10 young seedlings. One-way ANOVA was used in combination with Duncan's multiple comparison method to analyze the significance of differences. Different letters indicated significant differences at P< 0.05.
图 7 NtKUP6基因VIGS沉默植株钾离子含量测定
注:A. NtKUP6基因沉默效率(NtKUP6基因在对照植株中的表达量设置为1.0);B. VIGS植株叶片钾离子含量。所有数据至少包含三个生物重复,每个生物重复中至少含有3个植株的叶片。采用独立样本 t 检验分析差异显著性,**代表P< 0.01。
Fig. 7 Determination of K+ contents in NtKUP6 gene VIGS silenced plants
Note: A: Silencing efficiency of the NtKUP6 gene (the expression level of the NtKUP6 gene in the control plants was set at 1.0); B: Potassium ion content in the leaves of VIGS plants. All data included at least three biological replicates, and each biological replicate contained at least 3 plant leaves. The differences in significance were analyzed using the independent sample t-test, and ** indicates P< 0.01.
图 8 NtKUP6基因在本氏烟的过表达植株钾离子含量测定
注:A. NtKUP6基因在过表达植株叶片的表达量(NtKUP6基因在对照植株中的表达量设置为1.0);B. 过表达植株叶片钾离子含量。所有数据至少包含3个生物重复,每个生物重复中至少含有3个植株的叶片。采用独立样本 t 检验分析差异显著性,**代表P< 0.01。
Fig. 8 Determination of K+ contents in NtKUP6 gene overexpression plants
Note: A: The expression level of the NtKUP6 gene in the leaves of overexpressing plants (the expression level of the NtKUP6 gene in the control plants was set to 1.0); B: The potassium ion content in the leaves of overexpressing plants. All data include at least three biological replicates, and each biological replicate contains at least 3 plant leaves. The significance of differences was analyzed using the independent sample t-test, and ** represents P< 0.01.
表 1 本研究所用引物信息
Table 1 Primer sequences used in this study
引物名称 Primer 引物序列(5'-3') Sequence 用途 Usage NtKUP6-F ATGGATCTTGAAACAGGGGT For cloning vector NtKUP6-R TCATACATTGTAAACCATT NtKUP6-qF GACTGTTGTTGGAACGCCTTCT For qRT-PCR NtKUP6-qR TCTTTACGCTCTGATGGTCC Nt26S-qF GAAGAAGGTCCCAAGGGTTC Nt26S-qR TCTCCCTTTAACACCAACGG NtKUP6-VIGS-F AGATCGACGACAAGACCCTGCAGGTCATGCTGGTTACGACTTGC For VIGS vector construction NtKUP6-VIGS-R TTCTGAGGAGAAGAGCCCTGCAGGATTGCTGGGATTCCGGATACAAG  下载: 导出CSV
下载: 导出CSV
表 2 NtKUP6基因启动子区顺式作用元件
Table 2 The cis-acting elements in the promoter region of the NtKUP6 gene
种类
Function元件
Elements描述
Descriptions数量
Quantities胁迫相关
Stress-relatedMBS 参与干旱诱导性的MYB结合位点
(MYB binding site involved in drought-inducibility)1 TC-rich repeats 防御和应激反应元件
(cis-acting element involved in defense and stress responsiveness)3 调节相关
Regulate-relatedCAAT-box 启动子和增强子普遍存在的元件
(common cis-acting element in promoter and enhancer regions)38 TATA-box 据转录起始位点30个碱基的中心元件
(core promoter element around -30 of transcription start)36 光相关
Light-relatedAT1-motif 光响应元件(light responsive element) 1 ATCT-motif 光响应元件(light responsive element) 1 Box 4 光响应元件(light responsive element) 3 G-box 光响应元件(light responsive element) 1 GT1-motif 光响应元件(light responsive element) 1 MRE 光响应元件(light responsive element) 1 TCCC-motif 光响应元件(light responsive element) 1 TCT-motif 光响应元件(light responsive element) 1 生长相关
Growth-relatedARE 厌氧响应元件(light responsive element) 3 GC-motif 缺氧特异性诱导元件
(enhancer-like element involved in anoxic specific inducibility)1 激素相关
Hormone-relatedABRE 脱落酸应答元件
(cis-acting element involved in the abscisic acid responsiveness)2 P-box 赤霉素响应元件
(gibberellin-responsive element)1
下载: 导出CSV
-
[1] Mishra S, Bisht D, Amtmann A, et al. Potassium deficiency and hormone signalling in plants [J]. Plant, Cell & Environment, 2025, 1-17. doi:10.1111/pce.70165 (查阅网上资料,未找到本条文献卷期页码信息,请确认) [2] Mostofa M G, Rahman M M, Ghosh T K, et al. Potassium in plant physiological adaptation to abiotic stresses [J]. Plant Physiology and Biochemistry, 2022, 186: 279−289. https://doi.org/10.1016/j.plaphy.2022.07.011 doi: 10.1016/j.plaphy.2022.07.011 [3] Ashley M K, Grant M, Grabov A. Plant responses to potassium deficiencies: a role for potassium transport proteins [J]. Journal of Experimental Botany, 2006, 57(2): 425−436. https://doi.org/10.1093/jxb/erj034 doi: 10.1093/jxb/erj034 [4] Shabala S, Cuin T A. Potassium transport and plant salt tolerance [J]. Physiologia Plantarum, 2008, 133(4): 651−669. https://doi.org/10.1111/j.1399-3054.2007.01008.x doi: 10.1111/j.1399-3054.2007.01008.x [5] 宋秀娟, 岳文浩, 刘磊, 等. 植物高亲和性钾离子转运蛋白研究进展[J]. 浙江大学学报(农业与生命科学版), 2025, 51(4): 561−571. https://doi.org/10.3785/j.issn.1008-9209.2024.03.191 doi: 10.3785/j.issn.1008-9209.2024.03.191 [6] Lhamo D, Chao W, Gao Q F, et al. Recent advances in genome-wide analyses of plant potassium transporter families [J]. Current Genomics, 2021, 22(3): 164−180. https://doi.org/10.2174/1389202922666210225083634 doi: 10.2174/1389202922666210225083634 [7] 杨中敏, 王艳. 钾离子转运载体KUP/HAK/KT家族功能的研究进展[J]. 生物技术, 2014, 24(3): 100−103. https://doi.org/10.3969/j.issn.1004-311X.2014.03.0074 doi: 10.3969/j.issn.1004-311X.2014.03.0074 [8] Wang Y, Wu W H. Regulation of potassium transport and signaling in plants [J]. Current Opinion in Plant Biology, 2017, 39: 123−128. https://doi.org/10.1016/j.pbi.2017.06.006 doi: 10.1016/j.pbi.2017.06.006 [9] 柴薇薇, 王文颖, 崔彦农, 等. 植物钾转运蛋白KUP/HAK/KT家族研究进展[J]. 植物生理学报, 2019, 55(12): 1747−1761. https://doi.org/10.13592/j.cnki.ppj.2019.0133 doi: 10.13592/j.cnki.ppj.2019.0133 [10] Cui Y X, Zhang X Q, Yang M X, et al. Current understanding of HAK potassium transporters in plant development and stress tolerance [J]. Journal of Plant Physiology, 2025, 313: 154617. https://doi.org/10.1016/j.jplph.2025.154617 doi: 10.1016/j.jplph.2025.154617 [11] Quintero F J, Blatt M R. A new family of K+ transporters from Arabidopsis that are conserved across phyla [J]. FEBS Letters, 1997, 415(2): 206−211. https://doi.org/10.1016/S0014-5793(97)01125-3 doi: 10.1016/S0014-5793(97)01125-3 [12] Martinez-Cordero M A, Martínez V, Rubio F. Cloning and functional characterization of the high-affinity K+ transporter HAK1 of pepper [J]. Plant Molecular Biology, 2004, 56(3): 413−421. https://doi.org/10.1007/s11103-004-3845-4 doi: 10.1007/s11103-004-3845-4 [13] Gupta M, Qiu X H, Wang L, et al. KT/HAK/KUP potassium transporters gene family and their whole-life cycle expression profile in rice (Oryza sativa) [J]. Molecular Genetics and Genomics, 2008, 280(5): 437−452. https://doi.org/10.1007/s00438-008-0377-7 doi: 10.1007/s00438-008-0377-7 [14] 晁毛妮, 温青玉, 张晋玉, 等. 大豆KUP/HAK/KT钾转运体基因家族的鉴定与表达分析[J]. 西北植物学报, 2017, 37(2): 239−249. https://doi.org/10.7606/j.issn.1000-4025.2017.02.0239 doi: 10.7606/j.issn.1000-4025.2017.02.0239 [15] Osakabe Y, Arinaga N, Umezawa T, et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis [J]. The Plant Cell, 2013, 25(2): 609−624. https://doi.org/10.1105/tpc.112.105700 doi: 10.1105/tpc.112.105700 [16] Mo Z J, Luo W, Pi K, et al. Comparative transcriptome analysis between inbred lines and hybrids provides molecular insights into K+ content heterosis of tobacco (Nicotiana tabacum L. ) [J]. Frontiers in Plant Science, 2022, 13: 940787. https://doi.org/10.3389/fpls.2022.940787 doi: 10.3389/fpls.2022.940787 [17] 鲁逸飞, 李立芹, 任学良, 等. 烟草(Nicotiana tabacum)钾吸收的分子机制研究进展[J]. 分子植物育种, 2019, 17(22): 7572−7578. https://doi.org/10.13271/j.mpb.017.007572 doi: 10.13271/j.mpb.017.007572 [18] Song Z B, Wu X F, Gao Y L, et al. Genome-wide analysis of the HAK potassium transporter gene family reveals asymmetrical evolution in tobacco (Nicotiana tabacum) [J]. Genome, 2019, 62(4): 267−278. https://doi.org/10.1139/gen-2018-0187 doi: 10.1139/gen-2018-0187 [19] 石远帅, 陆俊, 丁丹, 等. 烟草钾转运蛋白KUP基因家族鉴定与表达分析[J]. 江苏农业科学, 2025, 53(4): 100−110. https://doi.org/10.15889/j.issn.1002-1302.2025.04.012 doi: 10.15889/j.issn.1002-1302.2025.04.012 [20] 鲁黎明, 杨铁钊. 烟草钾转运体基因NtHAK1的克隆及表达模式分析[J]. 核农学报, 2011, 25(3): 469−476. [21] 谭颖, 秦利军, 赵丹, 等. 共转化法获得HAK1基因高表达烟草提高植株钾吸收能力[J]. 植物生理学报, 2013, 49(7): 689−699. https://doi.org/10.13592/j.cnki.ppj.2013.07.012 doi: 10.13592/j.cnki.ppj.2013.07.012 [22] 秦利军, 宋拉拉, 赵丹, 等. 超量表达烟草高亲和钾离子转运体蛋白基因(NtHAK1)提高烟草盐胁迫能力[J]. 农业生物技术学报, 2015, 23(12): 1576−1587. https://doi.org/10.3969/j.issn.1674-7968.2015.12.006 doi: 10.3969/j.issn.1674-7968.2015.12.006 [23] 李晨依, 喻奇伟, 罗贞宝, 等. 烟草高亲和钾转运蛋白基因NtHAK5克隆及表达模式分析[J]. 南方农业学报, 2022, 53(6): 1493−1501. https://doi.org/10.3969/j.issn.2095-1191.2022.06.002 doi: 10.3969/j.issn.2095-1191.2022.06.002 [24] 李姣, 许力, 鲁黎明, 等. 烟草钾转运体NtKT12的克隆及表达分析[J]. 华北农学报, 2016, 31(2): 65−70. https://doi.org/10.7668/hbnxb.2016.02.012 doi: 10.7668/hbnxb.2016.02.012 [25] Zhang H, Jin J J, Jin L F, et al. Identification and analysis of the chloride channel gene family members in tobacco (Nicotiana tabacum) [J]. Gene, 2018, 676: 56−64. https://doi.org/10.1016/j.gene.2018.06.073 doi: 10.1016/j.gene.2018.06.073 [26] 杨中敏, 王艳. 盐穗木钾转运蛋白基因HcKUP12的克隆及在盐胁迫下的表达分析[J]. 植物科学学报, 2015, 33(4): 499−506. https://doi.org/10.11913/PSJ.2095-0837.2015.40499 doi: 10.11913/PSJ.2095-0837.2015.40499 [27] Ahn S J, Shin R, Schachtman D P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake [J]. Plant Physiology, 2004, 134(3): 1135−1145. https://doi.org/10.1104/pp.103.034660 doi: 10.1104/pp.103.034660 [28] Osakabe Y, Arinaga N, Umezawa T, et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis [J]. The Plant Cell, 2013, 25(2): 609−624. (查阅网上资料,本条文献与第15条文献重复,请确认) https://doi.org/10.1105/tpc.112.105700 [29] Horie T, Sugawara M, Okada T, et al. Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells [J]. Journal of Bioscience and Bioengineering, 2011, 111(3): 346−356. https://doi.org/10.1016/j.jbiosc.2010.10.014 doi: 10.1016/j.jbiosc.2010.10.014 [30] Feng H M, Tang Q, Cai J, et al. Rice OsHAK16 functions in potassium uptake and translocation in shoot, maintaining potassium homeostasis and salt tolerance [J]. Planta, 2019, 250(2): 549−561. https://doi.org/10.1007/s00425-019-03194-3 doi: 10.1007/s00425-019-03194-3 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 215
- HTML全文浏览量: 101
- 被引次数: 0
