

-
糖为生命体活动的提供能源物质和碳来源,参与调节植物生理过程,如种子萌发、光合作用、开花等诸多生理过程[1 − 2]。糖能影响渗透势,在植物响应生物和非生物胁迫中发挥作用[3 − 4]。在环境压力中,干旱胁迫是植物生长和生产力最不利的因素之一[5]。根系中可溶性糖的运输和积累促进了根冠比的增加,从而增强了抗旱性[6]。糖在细胞间的转运是由转运蛋白介导的。到目前为止,已经确定了三种糖转运蛋白,包括单糖转运体MST(monosaccharide transporter)、蔗糖转运体SUT(sucrose transporter)和糖外排转运体SWEET(sugars will eventually be exported transporter)[7]。
SWEET蛋白可以顺浓度梯度介导细胞蔗糖的摄取和流出,具有双向跨膜运输的功能[8]。真核生物SWEET蛋白包含2个保守的MtN3/saliva跨膜结构域,包含7个α-螺旋跨膜结构域(transmembrane,TMs),由3个结构域多肽按3+1+3构型融合而成[9 − 10]。水稻OsSWEET2b的低温电镜结构显示,N端以TM1-TM3-TM2的形式构成三螺旋束(triple-helix bundles),C端THB由TM5-TM7-TM6组成[11]。其中,3-TM结构域携带高度保守的脯氨酸(P)和谷氨酰胺基序(Q),SWEET也因此被称为PQ环家族转运蛋白[12]。SWEET成员可划为4个分枝(Clade Ⅰ、Ⅱ、Ⅲ、Ⅳ),属于同一分支的基因具有相似的结构和功能[13]。其中,Clade Ⅲ的SWEET成员参与蔗糖运输,Clade Ⅰ、Ⅱ和Ⅳ的SWEET成员主要运输己糖[14]。此外,研究发现Clade Ⅳ SWEET定位于液泡膜上,在拟南芥中特异性输出果糖[15]。SWEET基因于2010年首次在拟南芥中被发现[16]。此后在多种植物中均鉴定出SWEET基因,如草莓(Fragaria ×ananassa)[17]、大麦(Hordeum vulgare)[18]、辣椒(Capsicum annuum)[19]等。当植物暴露于低温、干旱、盐分等各种非生物胁迫时,其生长发育过程会受到严重影响[20]。茶树CsLHY具有转录激活因子的作用,结合CsSWEET17的启动子正向调控茶树的抗寒性[21]。CsSWEET16促使糖在液泡中的区室化来提高抗寒性[22]。在干旱胁迫下,拟南芥sweet17突变体表现出侧根生长减少,侧根发育相关转录因子的表达量降低,导致sweet17突变体的耐旱性受损[23]。
铁皮石斛(Dendrobium catenatum)作为中国传统名贵中药材,具有保护心脏、保护胃肠道、抗肿瘤、免疫调节、抗菌、抗氧化和抗疲劳等作用[24]。野生铁皮石斛主要分布于岩石和树皮上,经常遇到恶劣的环境,已被列为稀有濒危植物。在生产中,常以模拟野生种植模式在石头、树桩和林下种植,时常受到周期性干旱胁迫影响[25]。但目前关于铁皮石斛在干旱环境下的生存机制的研究,尤其是铁皮石斛SWEET基因家族在干旱胁迫下的功能研究还未见报道。因此,本研究在基因组水平上对DcSWEET基因家族成员进行鉴定,利用生物信息学方法对DcSWEET基因的系统发育树、保守基序、基因结构、顺式作用元件等进行综合分析。利用公共数据库中的转录组数据分析了DcSWEET基因在干旱胁迫下的相对表达水平,并利用RT-qPCR进行验证。研究结果可为铁皮石斛抗旱分子育种和提高铁皮石斛对干旱胁迫的耐受性提供参考。
-
从NCBI GenBank数据库下载铁皮石斛基因组注释文件(PRJNA262478)[26]。从Pfam数据库(
http://pfam.xfam.org/ , v35.0)查询MtN3/saliva结构域(PF03083)的隐马尔可夫模型,利用Hmmersearch进行搜索,保留E≤1×10−10的序列。通过Bio-linux bioinformatics documentation system查询D. catenatum蛋白数据库,初步获得DcSWEET候选蛋白。最后,提交至SMART(http://smart.embl.de/smart/batch.pl )和NCBI数据库(https://www.ncbi.nlm.nih.gov/ )的 CDD 在线工具来确定是否具有MtN3/saliva和PQ-loop保守结构域[27]。使用ExPASy(https://www.expasy.org/ )分析DcSWEET蛋白的氨基酸数、分子量、理论pI。使用PSORT工具预测DcSWEET蛋白的亚细胞定位。使用Clustalw 2.0程序对拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)以及铁皮石斛SWEET蛋白序列进行多重序列比对,将比对结果导入MEGA11,使用泊松模型的最大似然法构建SWEET蛋白的系统发育树,用在线网站Evolview(
https://evolgenius.info/evolview-v2 )美化。 -
采用TBtools以及MEME在线工具(
http://meme-suite.org/tools/meme )分析27个DcSWEET蛋白序列的保守基序。利用TBtools软件[28]从铁皮石斛基因组gff注释文件中提取DcSWEET基因结构文件,提取DcSWEET各基因起始密码子(ATG)上游2000 bp序列作为启动子序列,并利用PlantCARE 分析顺式作用元件。利用TBtools对27个DcSWEET保守基序、基因结构和顺式元件进行可视化分析。 -
基于SRA数据库中的RNA-seq数据(SRP132541)分析干旱胁迫下DcSWEET基因的表达模式。该数据包含基质不同含水量(0、10%~15%和30%~35%)下的基因表达量,根据基因ID筛选DcSWEET基因的表达数据,并使用TPM值构建TBtools热图。根据RNA-seq数据的TPM值,使用Origin软件(version 2022b)分析DcSWEET基因共表达关系[29] ,根据相关系数,使用Cytoscape(版本3.8.1)进行共表达网络分析,以鉴定核心基因。用于相关性分析的数据是4次重复的平均值。
-
利用实时荧光定量PCR(RT-qPCR)技术分析响应干旱胁迫的核心DcSWEET基因的表达模式。以实验室保存的20% PEG8000干旱胁迫下的cDNA为模板[30],分析DcSWEET基因在干旱处理3、6、9、12、24、48 h后的基因表达情况。引物序列如表1所示,引物由生工生物工程(上海)股份有限公司合成。使用ChamQ Universal SYBR qPCR Master Mix试剂(诺唯赞生物科技有限公司, 南京)进行PCR扩增。RT-qPCR扩增条件为:95 ℃初始变性5 min;95 ℃变性30 sec,60 ℃退火延伸45 s,40个循环。结束后读取荧光信号:95℃ 15 s,60℃ 15 s,95℃ 15 s,进行熔解曲线分析。试验设置3次生物学重复。以铁皮石斛Actin基因作为内参基因,采用2-ΔΔCt方法计算基因的相对表达量[25]。
表 1 本试验中实时荧光定量PCR引物序列
Table 1. Primer sequences of quantitative real time PCR in this experiment
基因ID
Gene ID上游引物(5′→3′)
Forward primers(5′→3′)下游引物(5′→3′)
Reversed primers(5′→3′)DcSWEET1 TGGTGAAGACAAAAAGTGTGGAG AGAGCACTCCCACAACCATTT DcSWEET2 GTCTTCCCTACTTGTATTCCCTTCT CATTCCACACATTTTGAGTTTCTTA DcSWEET4 ATTTCTCTCTCTTGCTTCCTTCTTT CGATCTCTGTGGTCTTTTTACGT DcSWEET5 TGGAGAAAGAAGGCAGTAGAACA GATAAGCACCCTCAAGCACAATA DcSWEET7 CGGTGGGTCATAGTCACGAGA GCATACGCCATTCAGAAAGCT DcSWEET9 CCGAATGTATTGGGTTTCCTGT ATCTTATCTTTCACCTGGTTTGTCC DcSWEET11 GGTGATGGGATCTTTGTGTGC CTCTCTATGTCTGTTTCCTTGGCT DcSWEET12 GCCCCAAAGAAGGTCAAGATAC GCGAAGACGCAAACAGAGAAG DcSWEET13 TAGGATTCACATTTGGGGCG TCATACAGGGCTCATCTCATTTTTA DcSWEET14 ATCATGTATCTCCTCTATGCCCCT AGAAAGCAACGCAAATCCAAC DcSWEET15 GTGTTACCTTCTCTGTGAGCGTCT GAGTCCGCAAGTATTATTTCAGTCA DcSWEET16 ACTGCCGAATGCTAAACCAAC AACTCATCTCCAGCCGAAAAA DcSWEET17 GGATTTGGAGCATCTACTCTGTG CTCTCTATGTCTGTTTCCTTGGC DcSWEET18 GTGTCGCCAATACCAACTTTTAGA CCAAAAATACACCATAGGAGACACA DcSWEET21 GTGTCGCCAATACCAACTTTTAGA CCAAAAATACACCATAGGAGACACA DcSWEET26 ACTTCTCATCACAGACATCACCTTC CACACTCTTTGTTCTTATCACCCTT DcSWEET27 TTGTGTTGGATTCTGGTGATGGTGT TTTCCCGTTCTGCTGTTGTTGTGAA DcActin TTGTGTTGGATTCTGGTGATGGTGT TTTCCCGTTCTGCTGTTGTTGTGAA -
本研究中从铁皮石斛中共鉴定到27个DcSWEET基因,根据DcSWEET家族成员与水稻和拟南芥SWEET蛋白的关系,将其命名为DcSWEET1 ~ 27。DcSWEET基因编码的蛋白氨基酸数为201(DcSWEET5)~296(DcSWEET11),分子量为22.44(DcSWEET5)~33.11(DcSWEET11)kDa,理论等电点pI为8.47(DcSWEET25)~9.91(DcSWEET16)。DcSWEET5是编码氨基酸最少、分子量最小的蛋白,DcSWEET11编码氨基酸最多,DcSWEET3分子量最大。27个DcSWEET蛋白的等电点都在7以上。亚细胞定位预测到24个(89%)DcSWEET蛋白定位于细胞质膜上,3(11%)个DcSWEET蛋白定位在叶绿体上(表2)。
表 2 铁皮石斛DcSWEET蛋白的理化性质
Table 2. Physiochemical properties of D. catenatum SWEET proteins
基因名
Gene Name位置
Locus长度
Length分子量/kDa
MW/kDa等电点
pI亚细胞定位
Subcellular localization组
GroupDcSWEET1 NW_021319548.1:92365...96183 211 23.2 8.95 plasma membrane Ⅰ DcSWEET2 NW_021320155.1:178296..180115 237 26.62 8.87 plasma membrane Ⅰ DcSWEET3 NW_021319455.1:1525477..1538903 277 30.99 9.25 plasma membrane Ⅰ DcSWEET4 NW_021319579.1:2296589..2300708 248 27.45 9.63 plasma membrane Ⅱ DcSWEET5 NW_021320057.1:1532934..1534265 201 22.44 9.34 plasma membrane Ⅱ DcSWEET6 NW_021318787.1:354097..355486 235 26.27 9.03 plasma membrane Ⅱ DcSWEET7 NW_021318787.1:348952..350461 236 26.37 9.14 plasma membrane Ⅱ DcSWEET8 NW_021320141.1:81668..104479 283 31.65 9.39 plasma membrane Ⅱ DcSWEET9 NW_021319221.1:10426..13058 262 29.22 9.49 plasma membrane Ⅲ DcSWEET10 NW_021319083.1:1983587..1985446 271 30.5 9.25 plasma membrane Ⅲ DcSWEET11 NW_021415225.1:997761..1000664 296 33.11 9.52 chloroplast Ⅳ DcSWEET12 NW_021319195.1:493114..496184 266 29.94 8.87 plasma membrane Ⅲ DcSWEET13 NW_021318711.1:2482437..2484846 265 29.55 9.27 plasma membrane Ⅲ DcSWEET14 NW_021318711.1:2495228..2496861 265 29.63 9.00 plasma membrane Ⅲ DcSWEET15 NW_021319091.1:54815..57648 249 27.59 9.13 plasma membrane Ⅲ DcSWEET16 NW_021415225.1:997761..1000664 254 28.71 9.91 chloroplast Ⅳ DcSWEET17 NW_021415225.1:997761..1000664 283 31.54 9.50 chloroplast Ⅳ DcSWEET18 NW_021320019.1 :9075754..9077812 237 26.58 8.99 plasma membrane Ⅰ DcSWEET19 NW_021320141.1:137711..138871 259 28.92 8.99 plasma membrane Ⅱ DcSWEET20 NW_021320141.1:73730..79633 285 32 9.24 plasma membrane Ⅱ DcSWEET21 NW_021320019.1:9075754..9077812 223 25.5 9.30 plasma membrane Ⅰ DcSWEET22 NW_021319959.1:1593464..1596798 237 26.78 9.18 plasma membrane Ⅱ DcSWEET23 NW_021594465.1:9716..14348 237 26.51 8.92 plasma membrane Ⅱ DcSWEET24 NW_021320141.1:124587..125761 259 28.72 9.11 plasma membrane Ⅱ DcSWEET25 NW_021320141.1:170130..171297 259 28.93 8.47 plasma membrane Ⅱ DcSWEET26 NW_021319959.1:1778270..1781665 237 26.59 9.32 plasma membrane Ⅱ DcSWEET27 NW_021319959.1:1514664..1518903 295 33.08 8.53 plasma membrane Ⅱ -
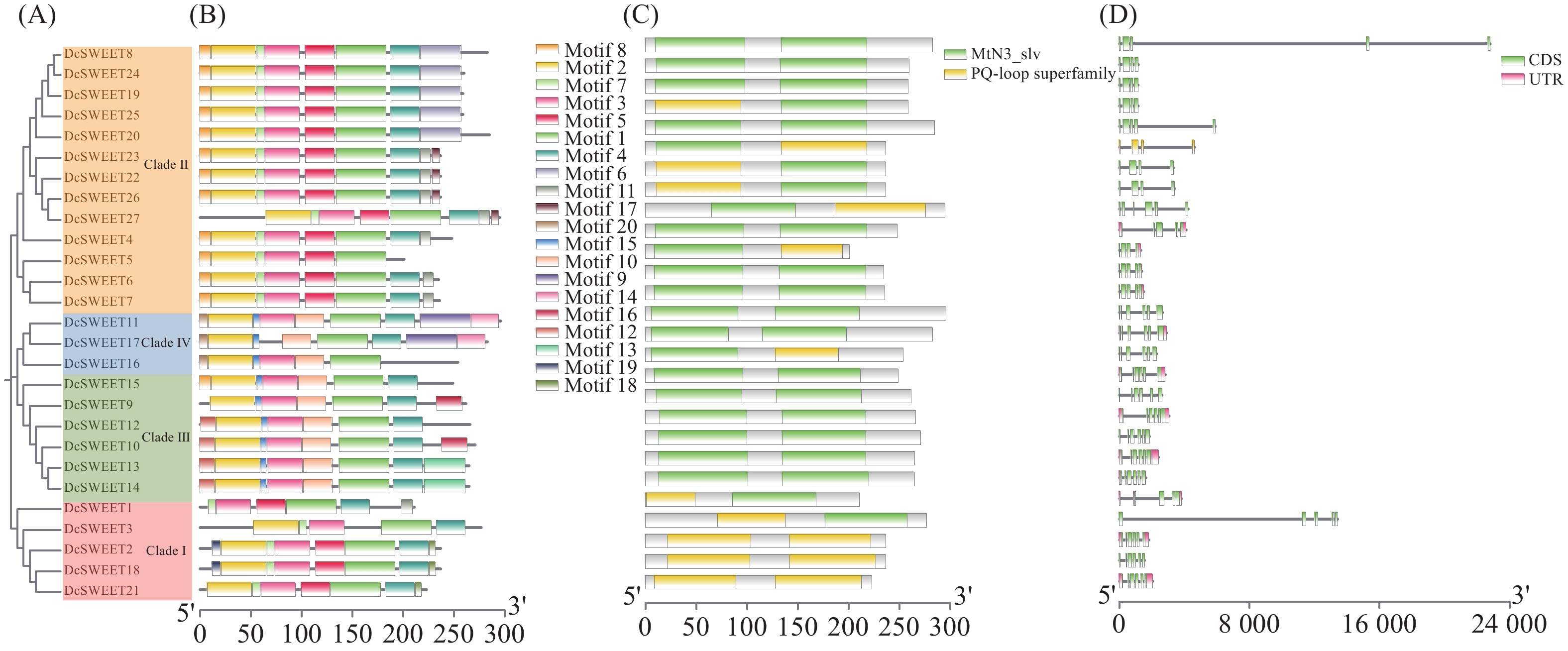
利用铁皮石斛(Dc)、拟南芥(At)和水稻(Os)的SWEET家族蛋白序列构建系统发育树(图1),铁皮石斛、拟南芥和水稻的SWEET蛋白均可分为4个亚组。铁皮石斛家族成员在四个亚组中的成员数分别为5、13、6和3,所占比例分别为19%(Clade Ⅰ)、48%(Clade Ⅱ)、22%(Clade Ⅲ)和11%(Clade Ⅳ)。而拟南芥AtSWEET家族成员在4个亚组中的数量分别为3个(18%)、5个(29%)、7个(41%)和2个(12%),水稻OsSWEET家族成员分别为6个(29%)、9个(43%)、5个(24%)和1个(4%)。可以看出,不同植物中SWETT基因在不同亚组中的分布具有相似性。
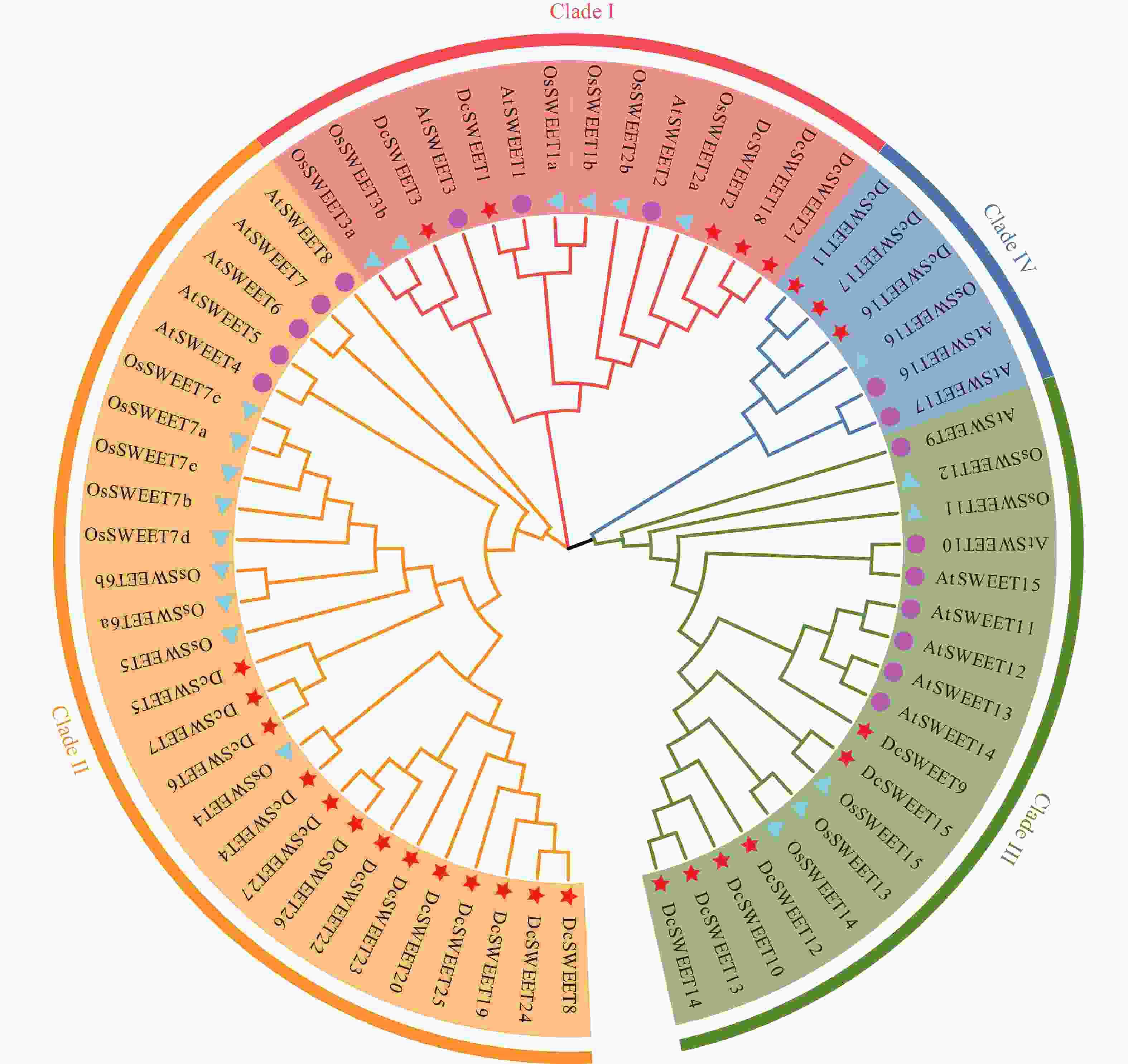
图 1 拟南芥、水稻和铁皮石斛SWEET蛋白的系统发育分析
Figure 1. Phylogenetic analyses of SWEET proteins from D. catenatum, A. thaliana and O. sativa
-
共鉴定得到20个基序(图2),DcSWEET蛋白几乎所有成员都包含motif 1、2、3、4,表明这些基序在DcSWEET转运体中是高度保守的。除了DcSWEET5和DcSWEET16外,DcSWEET成员都含有motif 4,这表明DcSWEET5和DcSWEET16可能在与其他成员共同进化的过程中丢失了motif 4(图2−B)。通过NCBI-CDD分析DcSWEET蛋白的保守结构域,发现所有的DcSWEETs都含有两个MtN3/saliva或PQ-loop superfamily(图2−C)。内含子–外显子结构分析表明,DcSWEET基因的内含子–外显子数量高度保守。27个成员中的内含子数量为3到5个之间。有14个(52%)DcSWEETs基因含有5个内含子,6个(22%)成员包含4个内含子,7个(26%)成员包含3个内含子(图2−D)。
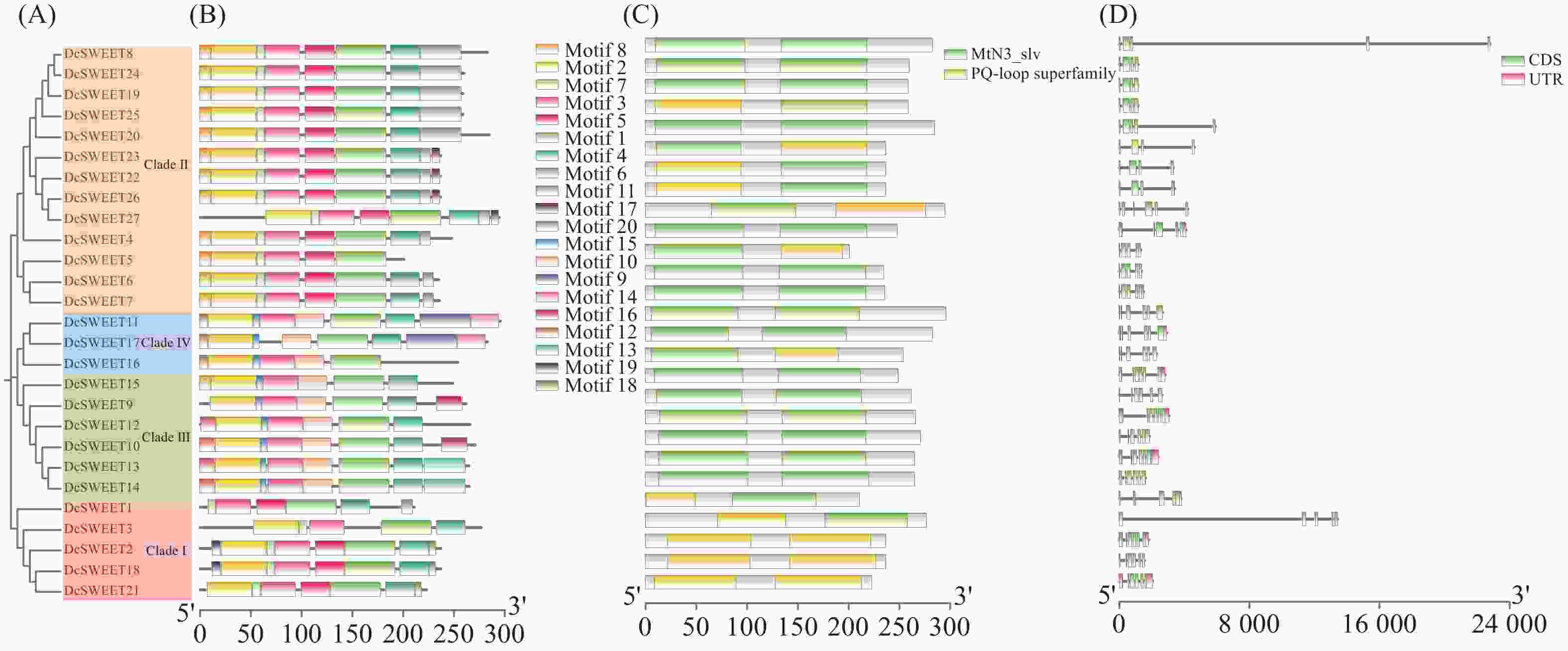
图 2 DcSWEET家族成员的基序、结构及系统发育关系
Figure 2. Motifs, structures and phylogenetic relationships of DcSWEET family members
-
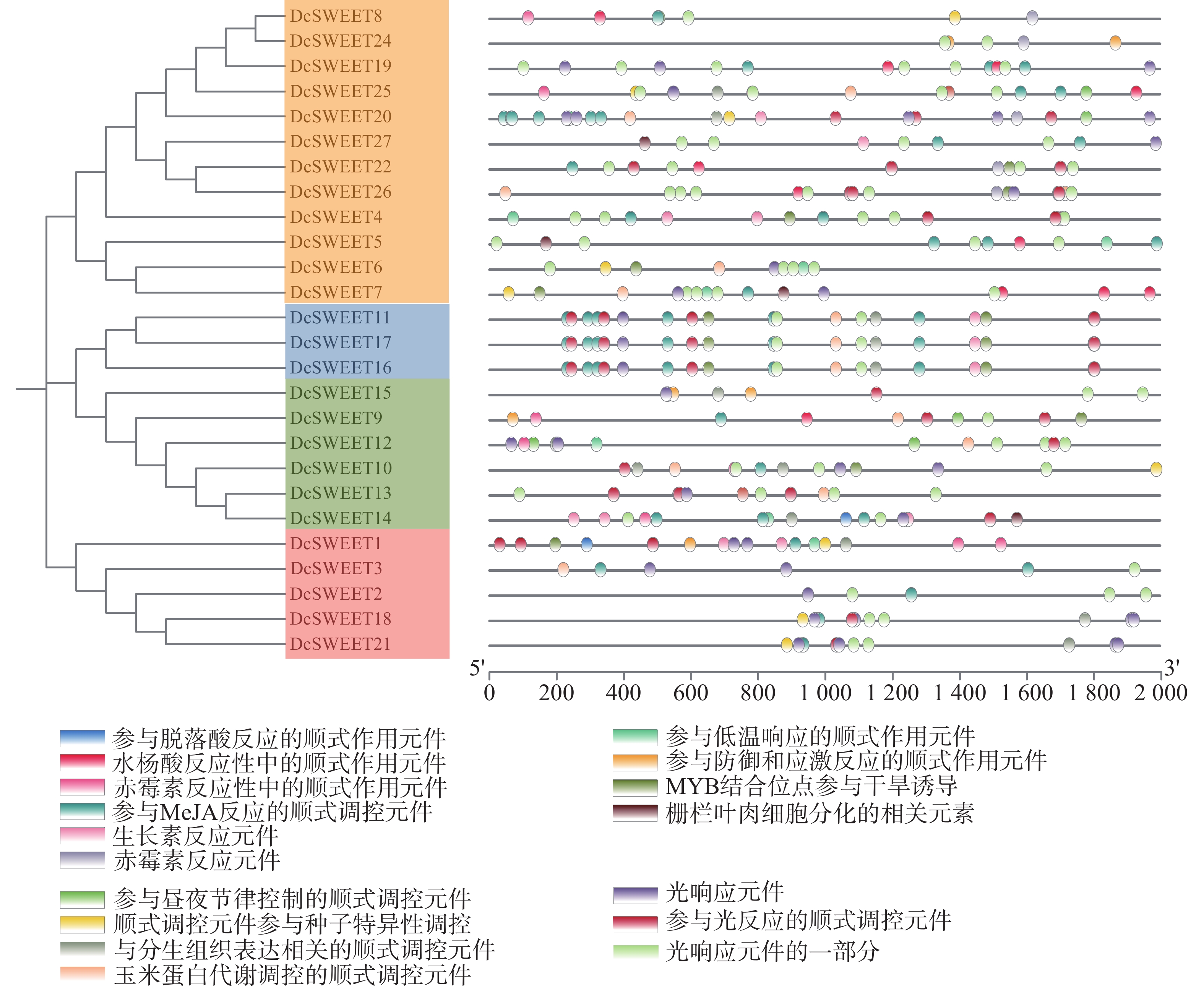
在DcSWEET启动子存在植物激素响应元件(脱落酸、水杨酸、赤霉素、生长素),逆境胁迫相关元件(低温、干旱、胁迫和防御响应元件)、植物生长发育相关元件(昼夜调控、分生组织、种子特异性调控、玉米蛋白代谢调控)、光响应元件(图3)。其中,在10个(37%)DcSWEET基因启动子中检测到参与低温响应的顺式作用元件,在11个(41%)DcSWEET基因启动子中检测到参与干旱诱导的MYB结合位点,在5个(19%)DcSWEET基因启动子中存在防御响应元件。说明大多数的DcSWEET基因参与对非生物胁迫的响应。
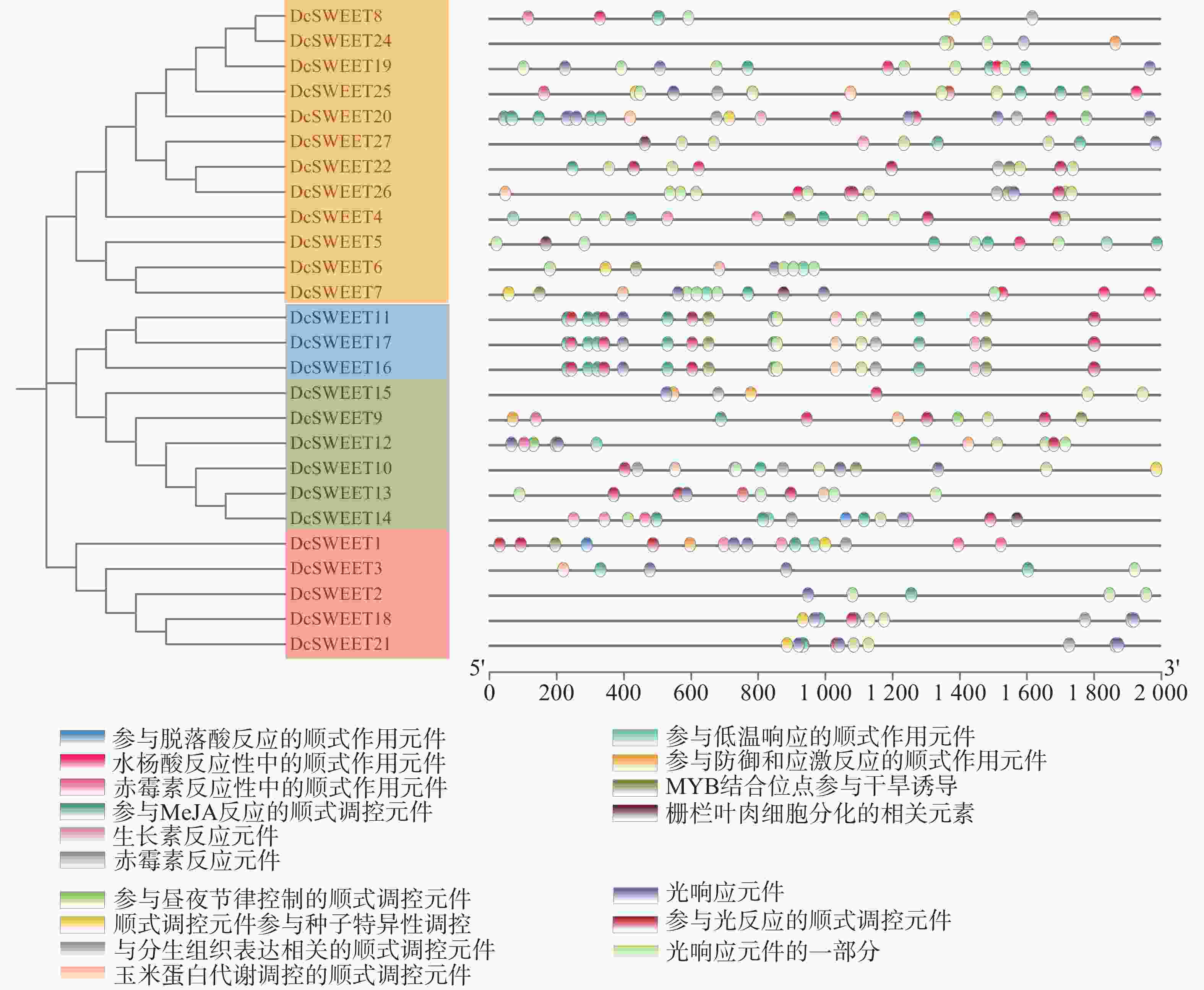
图 3 DcSWEET基因启动子区域的顺式作用元件分析
Figure 3. Analysis of cis-elements in the DcSWEET gene promoter regions
-
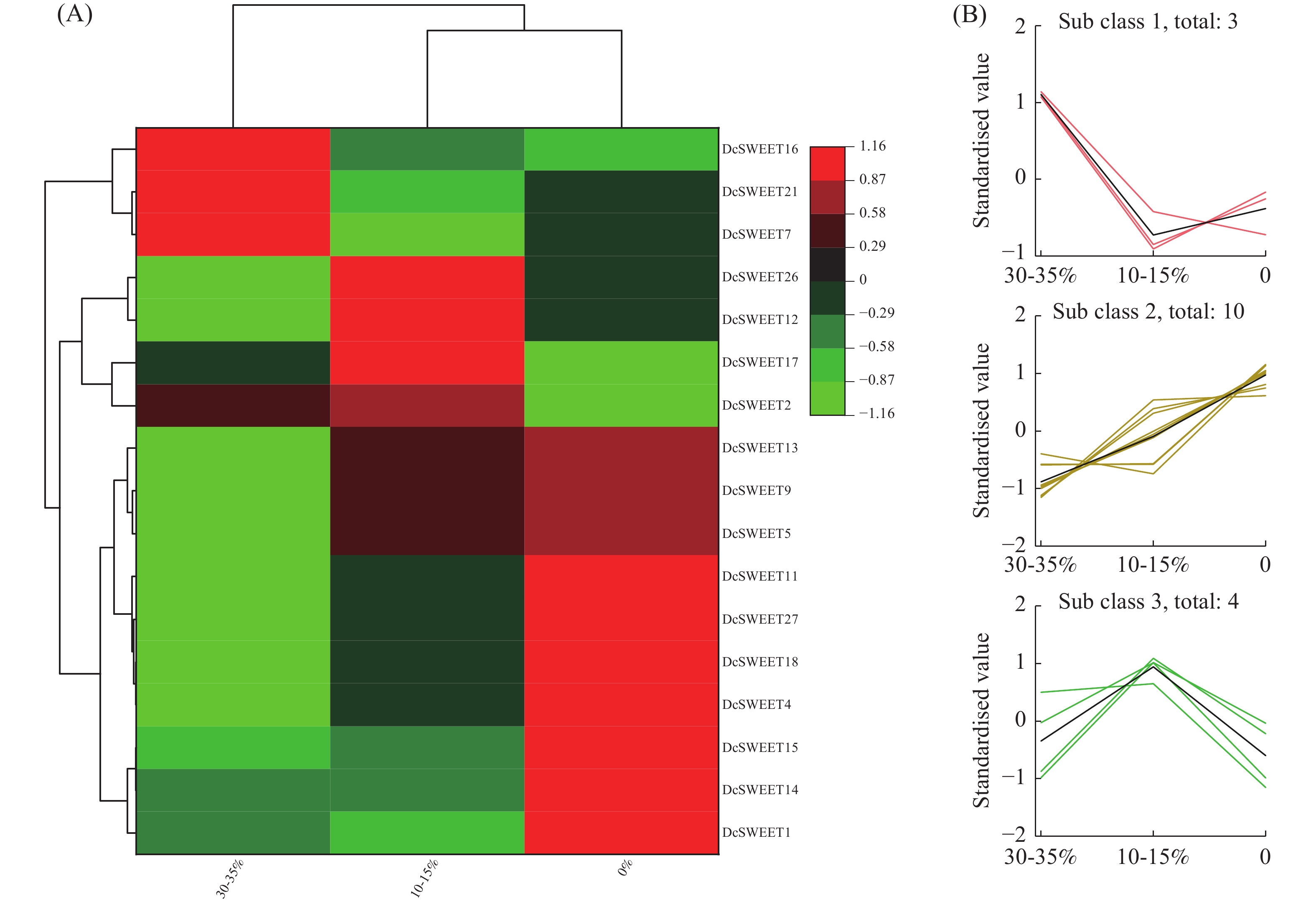
利用SRA数据库中的RNA-seq数据(SRP132541),筛选出17个DcSWEET基因参与响应干旱胁迫(图4−A)。根据基因表达量的趋势分析,可将这些基因按表达量变化分为三组(图4−B)。DcSWEET1、DcSWEET4、DcSWEET5、DcSWEET9、DcSWEET11、DcSWEET13、DcSWEET14、DcSWEET15、DcSWEET18共9个基因的表达量随着干旱胁迫的加重而升高,DcSWEET7、DcSWEET16、DcSWEET21在30−35%的基质含水量条件下表达量最高,表明这3个基因可能响应较轻的干旱胁迫。DcSWEET2、DcSWEET12、DcSWEET17、DcSWEET26在10~15%的基质含水量条件下表达量上调,在完全干旱时(0%的基质含水量)处理时迅速下降,表明这4个DcSWEET基因可能对中度干旱胁迫(10%~15%的基质含水量)起响应作用。
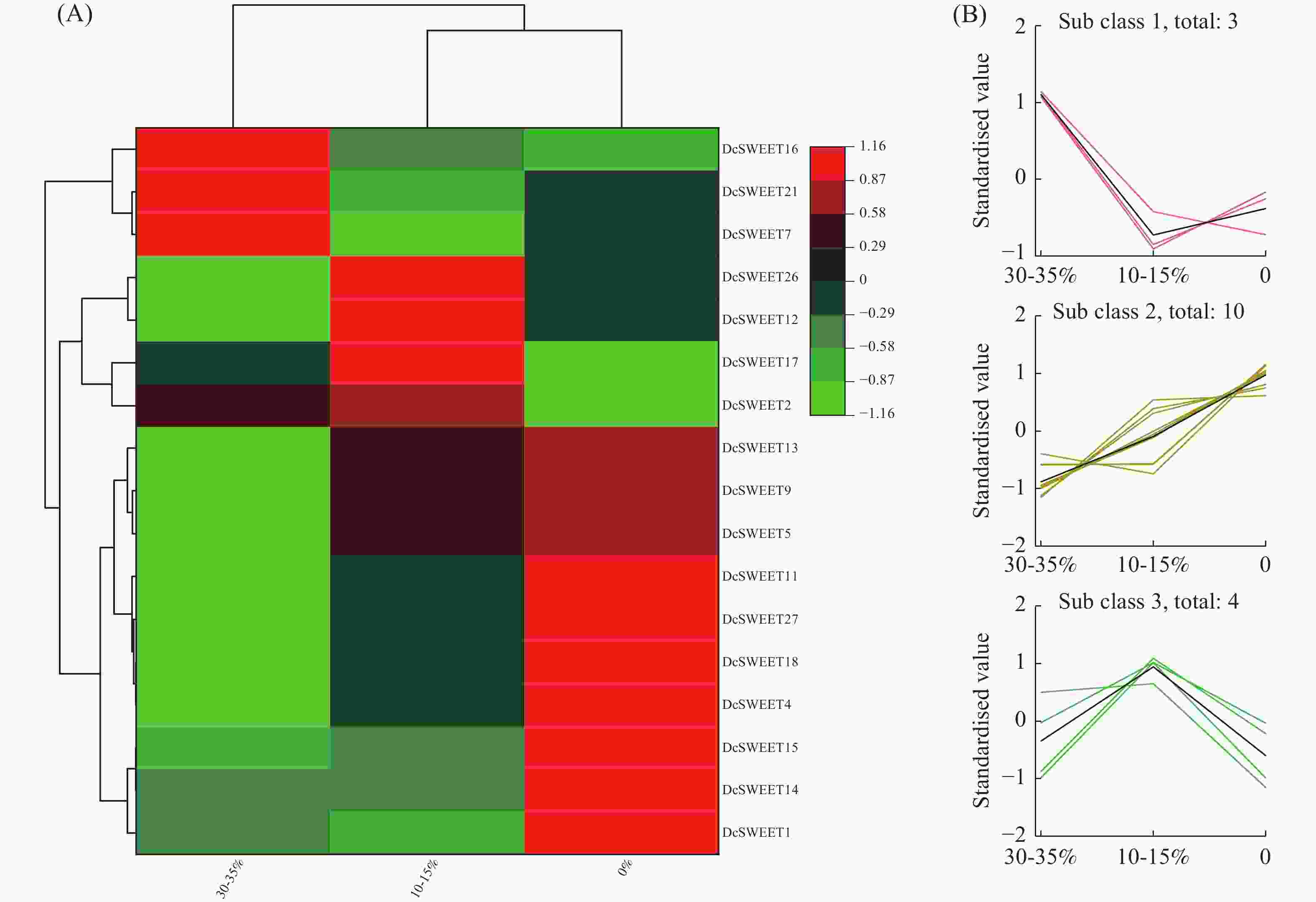
图 4 DcSWEET基因在干旱胁迫下的表达模式
Figure 4. Expressions of DcSWEET genes under drought stress
-
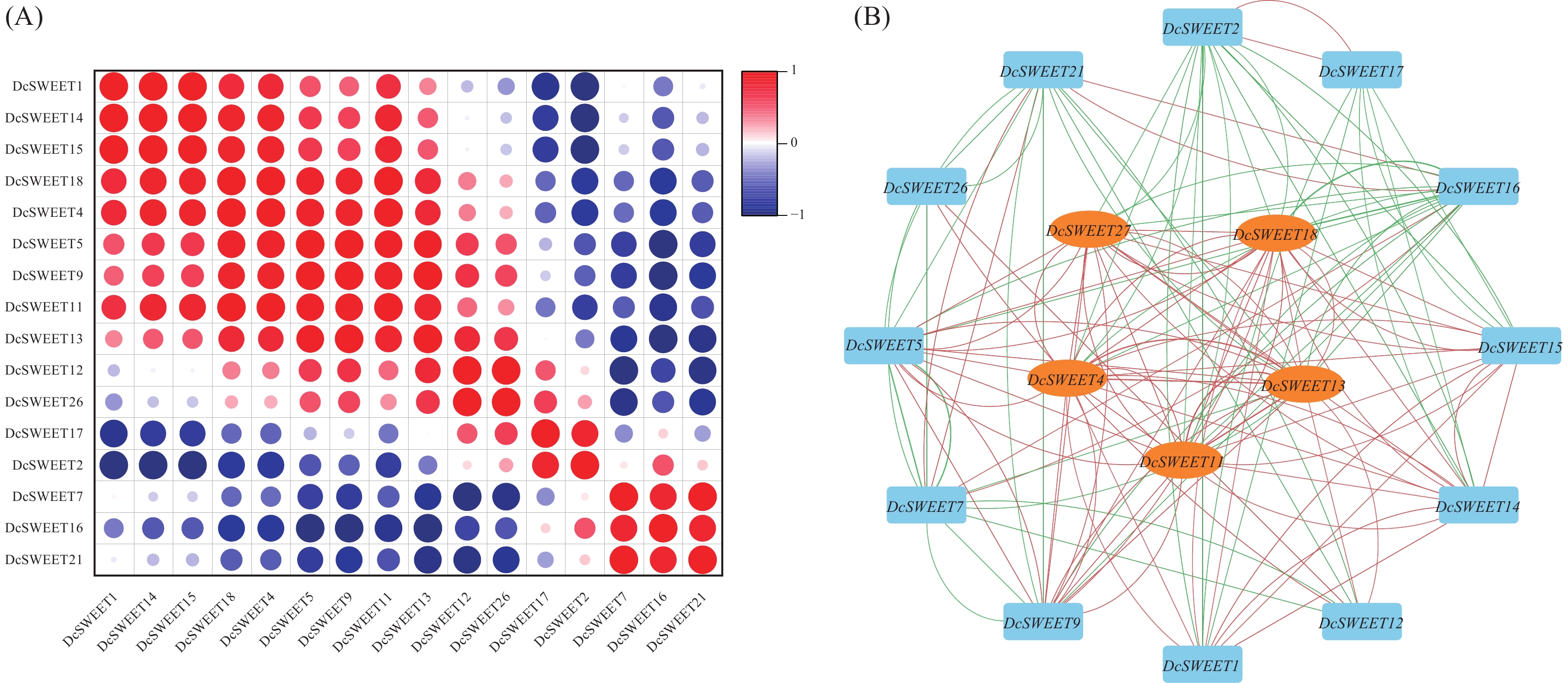
使用Origin软件对干旱胁迫处理下的17个DcSWEET基因TPM值进行相关性分析,选用皮尔逊系数大于0.8的数据,探讨它们之间可能存在的关系。结果表明,80对基因表现为正相关,60对基因表现为负相关(图5−A)。根据皮尔逊相关系数使用Cytoscape软件绘制共表达网络,共鉴定出5个枢纽基因,分别 为DcSWEET4、DcSWEET11、DcSWEET13、DcSWEET18、DcSWEET27(图5−B)。
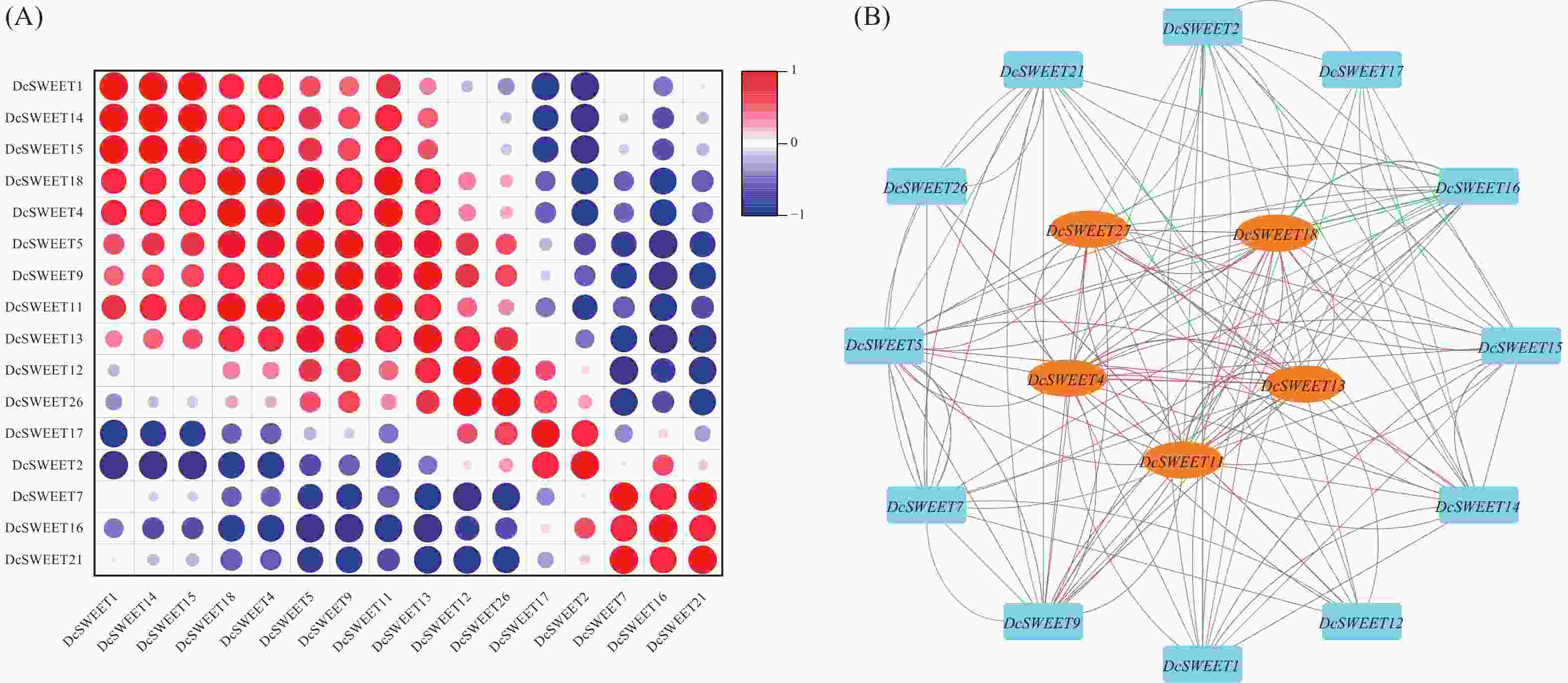
图 5 干旱相关DcSWEET基因的综合分析.
Figure 5. Comprehensive analysis of drought-related DcSWEET genes
-
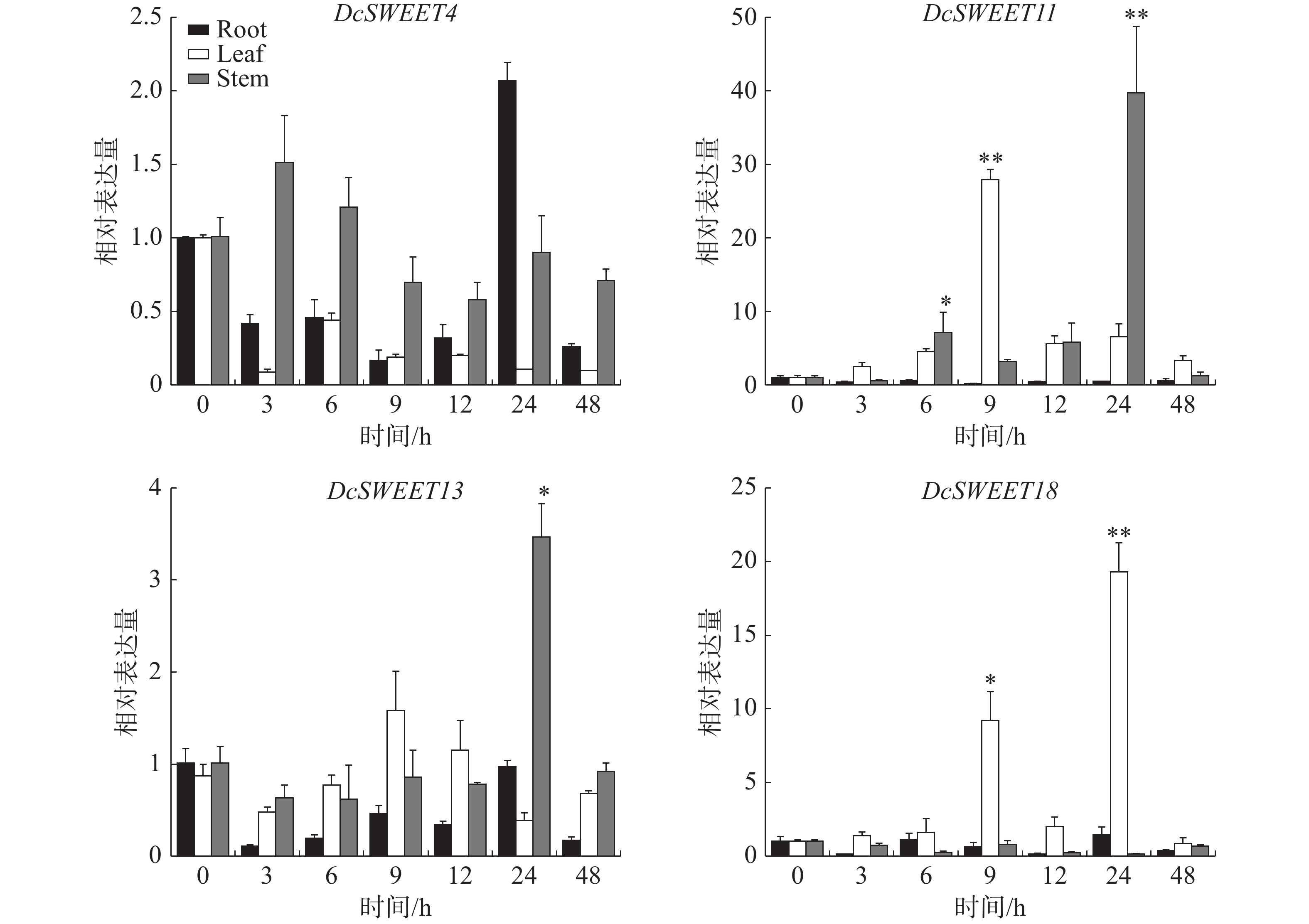
为了进一步探讨它们对干旱胁迫的响应,采用RT-qPCR分析了4个核心DcSWEET基因在20% PEG8000下的表达水平。结果(图6)表明,这些基因在干旱胁迫下均有不同程度的上调。在根中,处理24 h后DcSWEET4基因的表达达到最高值。在叶片中,DcSWEET11、DcSWEET13在干旱处理9 h后达到最高值,DcSWEET18在处理24 h后达到最大值,其中DcSWEET11上调28倍,DcSWEET18在处理24 h后上调19倍。在茎中,DcSWEET11和DcSWEET13处理24 h后达到巅峰值,其中DcSWEET11表达量是0 h表达量的40倍。
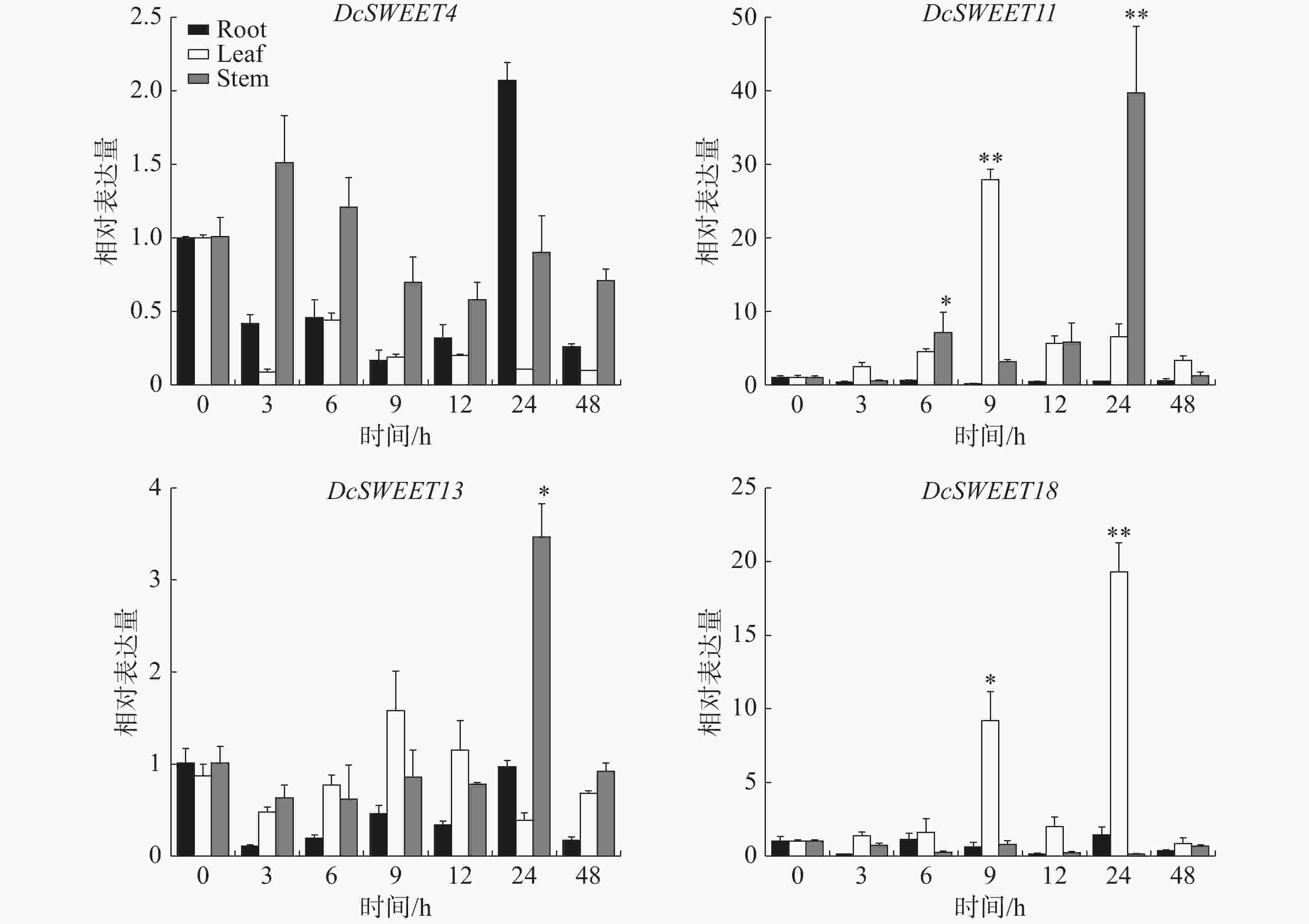
图 6 干旱胁迫下核心DcSWEET基因的RT-PCR分析
Figure 6. RT-PCR analysis of the hub DcSWEET genes under drought stress
-
SWEET蛋白是存在于细菌、真菌、植物和人类中的主要转运蛋白之一,用于促进糖跨细胞膜运输[31]。在高等植物中,已报道的SWEET基因数量从17到108个不等,如在拟南芥中已鉴定出17个SWEET基因[32]、小麦中鉴定到108个[33]、高粱有23个[34]、番茄有29个[35]。本研究鉴定的27个SWEET基因家族成员,编码蛋白长度在201到296个aa。在其他植物中也发现类似的研究结果,如杨桃中的SWEET蛋白的氨基酸数目在229~300[36],葡萄中的SWEET蛋白的氨基酸数目在201~289[37],菜豆在195~304[38]。不同植物中的SWEET蛋白的氨基酸数目相似,说明不同SWEET在进化中具有较高的保守性。根据拟南芥系统发育树的分类方法[39],27个DcSWEETs被划分为4个亚群(图1)。在大多数植物中,SWEET基因家族中以Clade Ⅱ和Clade Ⅲ为主[40]。本研究发现铁皮石斛SWEET蛋白中Clade Ⅱ成员数占比48%,Clade Ⅲ中的成员数占比22%,表明铁皮石斛中Clade Ⅱ和Clade Ⅲ也是主要类群。
基因结构分析显示52%的DcSWEET成员含有5个内含子,22%的成员含有4个内含子,26%的成员含有3个内含子。可以推测,SWEET基因的祖先是由 6 个外显子和 5 个内含子组成的基因基本模型。DcSWEET中有近一半成员中内含子数少于5个,可能是其在进化时发生了内含子的丢失或外显子的融合[41]。MtN3_slv亚家族参与葡萄糖转运,而PQ-loop亚家族参与氨基酸转运[42]。保守结构域分析表明,Clade Ⅲ家族成员包含两个MtN3_slv超家族结构域,Clade Ⅱ家族7个成员包含两个MtN3_slv超家族结构域,6个成员包含1个MtN3_slv超家族结构域和1个PQ-loop超家族结构域(图2−C)。从系统发育、启动子数量和基因结构分析可以看出,铁皮石斛SWEET蛋白家族在进化上是相对保守的。顺式调控元件是位于基因转录起始区的一类DNA序列,相应的转录因子识别顺式作用元件来调节目标基因的mRNA丰度,在非生物胁迫响应中起着至关重要的作用[43]。本研究中,在21个DcSWEET基因启动子中发现茉莉酸甲酯顺式作用元件,部分DcSWEET基因启动子中发现与赤霉素、水杨酸、脱落酸和生长素信号相关的顺式作用元件,在10个、11个和5个DcSWEET基因启动子中分别发现了低温响应元件、干旱诱导元件、防御和胁迫响应元件等非生物胁迫响应元件。这些数据表明,DcSWEET可能通过多种机制参与环境胁迫。
糖不仅充当满足组织代谢需求的能量和信号分子,而且还充当渗透剂,以应对干旱胁迫。在小麦中,SWEET基因在干旱和盐胁迫下的上调率高于热胁迫和热联合干旱胁迫[44]。在脱水和干旱胁迫下, OsSWEET15的表达诱导最为显著,其次是OsSWEET13,OsSWEET12还显示出干旱胁迫的中等诱导作用[45]。研究发现,大豆叶片中的GmSUC2转录水平在干旱胁迫第15 天显著增加(平均 2.18 倍),并上调了种子中GmSUC2、GmSWEET12和GmSWEET21的转录水平[46]。根据茶树干旱胁迫的RNA-seq,7个基因在PEG胁迫下上调[47]。本研究发现,11个含有干旱诱导元件的DcSWEET基因中,DcSWEET1、DcSWEET4、DcSWEET5、DcSWEET9、DcSWEET11、DcSWEET13、DcSWEET14、DcSWEET15、DcSWEET18在基质含水量0%时被诱导,DcSWEET2、DcSWEET12、DcSWEET17、DcSWEET26在基质含水量10−15%时被诱导,DcSWEET7、DcSWEET16、DcSWEET21在基质含水量30-35%时被诱导表达,说明这些基因在不同程度的干旱胁迫下受到胁迫表达,可能参与干旱胁迫的响应。通过共表达网络分析筛选出5个核心基因(图5−B)。RT-qPCR分析进一步表明,DcSWEET11在PEG处理9 h的叶中上调28倍,在处理24 h的茎中表达上调40倍。DcSWEET13在茎和叶中也有不同程度的上调。DcSWEET18在叶中被高度诱导表达,分别在处理9 h和24 h时上调9倍和19倍。DcSWEET4在处理24 h的根中上调2倍。结果显示DcSWEET基因在不同组织中应对干旱胁迫的模式不同,可能参与铁皮石斛不同组织部位应对干旱胁迫,但其功能验证仍需进一步开展。
-
本研究中,共鉴定了27个DcSWEET基因,同一亚组中的基因结构和保守基序具有相似性。在DcSWEETs启动子的上游序列中发现了4种类别的顺式作用元件。利用公开的铁皮石斛干旱胁迫下的RNA-seq数据,根据表达模式将DcSWEET基因划分为3个簇,结合RT-qPCR实验,发现DcSWEET11、DcSWEET13、DcSWEET18基因可作为铁皮石斛耐旱性潜在候选基因。本研究为进一步阐明DcSWEETs基因在干旱胁迫中的作用奠定了基础。
Identification and expression pattern analysis of SWEET gene family from Dendrobium catenatum
-
摘要: 糖转运蛋白SWEET(sugars will eventually be exported transporter)在植物响应生物和非生物胁迫中起重要作用。为探讨铁皮石斛(Dendrobium catenatum)SWEET的基因功能,基于铁皮石斛基因组数据,利用生物信息学方法对SWEET基因家族进行鉴定,并对其在干旱胁迫下的表达模式进行分析。结果显示,铁皮石斛中共有27个DcSWEET基因,理化性质分析表明DcSWEET蛋白质分子量为22.44~33.11 kDa,等电点为8.47~9.63。系统发育分析表明DcSWEET蛋白可以划分为4个亚群,每个亚群具有保守的基序组成和基因结构。顺式作用元件分析表明,大量的生长发育相关元件、激素响应元件以及胁迫响应元件存在于DcSWEET基因启动子序列上。RNA-seq分析发现17个DcSWEET基因受干旱胁迫诱导上调表达。共表达网络分析显示DcSWEET4、DcSWEET11、DcSWEET13、DcSWEET18、DcSWEET27可能在铁皮石斛响应干旱胁迫中起主要作用。实时荧光定量PCR结果表明,茎和叶中的DcSWEET11、DcSWEET13,以及叶中的DcSWEET18基因受干旱胁迫显著上调表达,可作为铁皮石斛耐旱性的候选基因。本研究分析了铁皮石斛中响应干旱胁迫的关键DcSWEET基因,为将来提高铁皮石斛耐旱性提供分子依据。Abstract: The sugar transporter SWEET (Sugars Will Eventually be Exported Transporter) plays a crucial role in plant responses to both biotic and abiotic stresses. To elucidate the function of the SWEET gene family in Dendrobium catenatum, 27 DcSWEET genes were identified and characterized from D. catenatum by using whole-genome data, and their expression patterns under drought stress were analyzed. The molecular weight of the DcSWEET proteins were found to range from 22.44 to 39.10 kDa, with heir isoelectric points being between 8.47 and 9.63. Phylogenetic analysis revealed that the DcSWEET proteins could be classified into four subgroups, each characterized by conserved motif compositions and gene structures. Cis-acting element analysis indicated that the promoters of DcSWEET genes contain various elements associated with hormone signaling, plant growth and development, light responsiveness, and stress response. RNA-seq analysis demonstrated that 17 DcSWEET genes were up-regulated under drought stress. Coexpression network analysis suggested that DcSWEET4, DcSWEET11, DcSWEET13, DcSWEET18, and DcSWEET27 may play pivotal roles in response to drought stress in D. catenatum. RT-qPCR confirmed that DcSWEET11, DcSWEET13, and DcSWEET18 were significantly up-regulated in stems and leaves under drought conditions, making them potential candidate genes for enhancing drought tolerance in D. catenatum. This study provides a comprehensive analysis of key SWEET genes involved in drought stress responses in D. catenatum, offering a molecular foundation for future efforts to improve its drought resistance.
-
Key words:
- Dendrobium catenatum /
- SWEET gene family /
- drought stress /
- expression pattern
-
图 2 DcSWEET家族成员的基序、结构及系统发育关系
A:DcSWEET蛋白的系统发育树;B:DcSWEET蛋白的保守基序;C:DcSWEET蛋白的保守结构域;D:DcSWEET基因的外显子/内含子结构。
Fig. 2 Motifs, structures and phylogenetic relationships of DcSWEET family members
A: Phylogenetic tree of DcSWEET protein; B: Conserved motifs of DcSWEET proteins; C: Conserved domains of DcSWEET proteins; D: Exon/intron structures of DcSWEET genes.
图 3 DcSWEET基因启动子区域的顺式作用元件分析
Fig. 3 Analysis of cis-elements in the DcSWEET gene promoter regions
图 4 DcSWEET基因在干旱胁迫下的表达模式
A:采用RNA-seq分析DcSWEET基因的表达模式;B:K-means均值分析。
Fig. 4 Expressions of DcSWEET genes under drought stress
A: Expression patterns of DcSWEET genes analyzed by RNA-seq; B: K-means analysis of DcSWEET genes.
图 5 干旱相关DcSWEET基因的综合分析.
A:DcSWEET基因相关性分析;B:共表达网络分析。
Fig. 5 Comprehensive analysis of drought-related DcSWEET genes
A: Correlation analysis of DcSWEET genes; B: Co-expression network analysis.
图 6 干旱胁迫下核心DcSWEET基因的RT-PCR分析
Fig. 6 RT-PCR analysis of the hub DcSWEET genes under drought stress
表 1 本试验中实时荧光定量PCR引物序列
Table 1 Primer sequences of quantitative real time PCR in this experiment
基因ID
Gene ID上游引物(5′→3′)
Forward primers(5′→3′)下游引物(5′→3′)
Reversed primers(5′→3′)DcSWEET1 TGGTGAAGACAAAAAGTGTGGAG AGAGCACTCCCACAACCATTT DcSWEET2 GTCTTCCCTACTTGTATTCCCTTCT CATTCCACACATTTTGAGTTTCTTA DcSWEET4 ATTTCTCTCTCTTGCTTCCTTCTTT CGATCTCTGTGGTCTTTTTACGT DcSWEET5 TGGAGAAAGAAGGCAGTAGAACA GATAAGCACCCTCAAGCACAATA DcSWEET7 CGGTGGGTCATAGTCACGAGA GCATACGCCATTCAGAAAGCT DcSWEET9 CCGAATGTATTGGGTTTCCTGT ATCTTATCTTTCACCTGGTTTGTCC DcSWEET11 GGTGATGGGATCTTTGTGTGC CTCTCTATGTCTGTTTCCTTGGCT DcSWEET12 GCCCCAAAGAAGGTCAAGATAC GCGAAGACGCAAACAGAGAAG DcSWEET13 TAGGATTCACATTTGGGGCG TCATACAGGGCTCATCTCATTTTTA DcSWEET14 ATCATGTATCTCCTCTATGCCCCT AGAAAGCAACGCAAATCCAAC DcSWEET15 GTGTTACCTTCTCTGTGAGCGTCT GAGTCCGCAAGTATTATTTCAGTCA DcSWEET16 ACTGCCGAATGCTAAACCAAC AACTCATCTCCAGCCGAAAAA DcSWEET17 GGATTTGGAGCATCTACTCTGTG CTCTCTATGTCTGTTTCCTTGGC DcSWEET18 GTGTCGCCAATACCAACTTTTAGA CCAAAAATACACCATAGGAGACACA DcSWEET21 GTGTCGCCAATACCAACTTTTAGA CCAAAAATACACCATAGGAGACACA DcSWEET26 ACTTCTCATCACAGACATCACCTTC CACACTCTTTGTTCTTATCACCCTT DcSWEET27 TTGTGTTGGATTCTGGTGATGGTGT TTTCCCGTTCTGCTGTTGTTGTGAA DcActin TTGTGTTGGATTCTGGTGATGGTGT TTTCCCGTTCTGCTGTTGTTGTGAA  下载: 导出CSV
下载: 导出CSV
表 2 铁皮石斛DcSWEET蛋白的理化性质
Table 2 Physiochemical properties of D. catenatum SWEET proteins
基因名
Gene Name位置
Locus长度
Length分子量/kDa
MW/kDa等电点
pI亚细胞定位
Subcellular localization组
GroupDcSWEET1 NW_021319548.1:92365...96183 211 23.2 8.95 plasma membrane Ⅰ DcSWEET2 NW_021320155.1:178296..180115 237 26.62 8.87 plasma membrane Ⅰ DcSWEET3 NW_021319455.1:1525477..1538903 277 30.99 9.25 plasma membrane Ⅰ DcSWEET4 NW_021319579.1:2296589..2300708 248 27.45 9.63 plasma membrane Ⅱ DcSWEET5 NW_021320057.1:1532934..1534265 201 22.44 9.34 plasma membrane Ⅱ DcSWEET6 NW_021318787.1:354097..355486 235 26.27 9.03 plasma membrane Ⅱ DcSWEET7 NW_021318787.1:348952..350461 236 26.37 9.14 plasma membrane Ⅱ DcSWEET8 NW_021320141.1:81668..104479 283 31.65 9.39 plasma membrane Ⅱ DcSWEET9 NW_021319221.1:10426..13058 262 29.22 9.49 plasma membrane Ⅲ DcSWEET10 NW_021319083.1:1983587..1985446 271 30.5 9.25 plasma membrane Ⅲ DcSWEET11 NW_021415225.1:997761..1000664 296 33.11 9.52 chloroplast Ⅳ DcSWEET12 NW_021319195.1:493114..496184 266 29.94 8.87 plasma membrane Ⅲ DcSWEET13 NW_021318711.1:2482437..2484846 265 29.55 9.27 plasma membrane Ⅲ DcSWEET14 NW_021318711.1:2495228..2496861 265 29.63 9.00 plasma membrane Ⅲ DcSWEET15 NW_021319091.1:54815..57648 249 27.59 9.13 plasma membrane Ⅲ DcSWEET16 NW_021415225.1:997761..1000664 254 28.71 9.91 chloroplast Ⅳ DcSWEET17 NW_021415225.1:997761..1000664 283 31.54 9.50 chloroplast Ⅳ DcSWEET18 NW_021320019.1 :9075754..9077812 237 26.58 8.99 plasma membrane Ⅰ DcSWEET19 NW_021320141.1:137711..138871 259 28.92 8.99 plasma membrane Ⅱ DcSWEET20 NW_021320141.1:73730..79633 285 32 9.24 plasma membrane Ⅱ DcSWEET21 NW_021320019.1:9075754..9077812 223 25.5 9.30 plasma membrane Ⅰ DcSWEET22 NW_021319959.1:1593464..1596798 237 26.78 9.18 plasma membrane Ⅱ DcSWEET23 NW_021594465.1:9716..14348 237 26.51 8.92 plasma membrane Ⅱ DcSWEET24 NW_021320141.1:124587..125761 259 28.72 9.11 plasma membrane Ⅱ DcSWEET25 NW_021320141.1:170130..171297 259 28.93 8.47 plasma membrane Ⅱ DcSWEET26 NW_021319959.1:1778270..1781665 237 26.59 9.32 plasma membrane Ⅱ DcSWEET27 NW_021319959.1:1514664..1518903 295 33.08 8.53 plasma membrane Ⅱ
下载: 导出CSV
-
[1] LASTDRAGER J, HANSON J, SMEEKENS S. Sugar signals and the control of plant growth and development[J]. Journal of Experimental Botany, 2014, 65(3): 799 − 807. doi: 10.1093/jxb/ert474 [2] ROLLAND F, BAENA-GONZALEZ E, SHEEN J. Sugar sensing and signaling in plants: conserved and novel mechanisms[J]. Annual Review of Plant Biology, 2006, 57: 675 − 709. doi: 10.1146/annurev.arplant.57.032905.105441 [3] GIBSON S I. Control of plant development and gene expression by sugar signaling[J]. Current Opinion in Plant Biology, 2005, 8(1): 93 − 102. doi: 10.1016/j.pbi.2004.11.003 [4] LUKASZUK E, RYS M, MOŻDŻEŃ K, et al. Photosynthesis and sucrose metabolism in leaves of Arabidopsis thaliana aos, ein4 and rcd1 mutants as affected by wounding[J]. Acta Physiologiae Plantarum, 2017, 39(1): 17. doi: 10.1007/s11738-016-2309-1 [5] HUANG H, JIAO Y X, TONG Y, et al. Comparative analysis of drought-responsive biochemical and transcriptomic mechanisms in two Dendrobium officinale genotypes[J]. Industrial Crops and Products, 2023, 199: 116766. doi: 10.1016/j.indcrop.2023.116766 [6] KHAN M I R, PALAKOLANU S R, CHOPRA P, et al. Improving drought tolerance in rice: ensuring food security through multi-dimensional approaches[J]. Physiologia Plantarum, 2021, 172(2): 645 − 668. doi: 10.1111/ppl.13223 [7] EOM J S, CHEN L Q, SOSSO D, et al. SWEETs, transporters for intracellular and intercellular sugar translocation[J]. Current Opinion in Plant Biology, 2015, 25: 53 − 62. doi: 10.1016/j.pbi.2015.04.005 [8] XUAN Y H, HU Y B, CHEN L Q, et al. Functional role of oligomerization for bacterial and plant SWEET sugar transporter family[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(39): E3685 − E3694. doi: 10.1073/pnas.1311244110 [9] FRANK BAKER R, LEACH K A, BRAUN D M. SWEET as sugar: new sucrose effluxers in plants[J]. Molecular Plant, 2012, 5(4): 766 − 768. doi: 10.1093/mp/SSS054 [10] LEE Y, NISHIZAWA T, YAMASHITA K, et al. Structural basis for the facilitative diffusion mechanism by SemiSWEET transporter[J]. Nature Communications, 2015, 6(1): 6112. doi: 10.1038/ncomms7112 [11] SCHNEIDER A, GODIN C, BOUDON F, et al. Light regulation of axillary bud outgrowth along plant axes: an overview of the roles of sugars and hormones[J]. Frontiers in Plant Science, 2019, 10: 1296. doi: 10.3389/fpls.2019.01296 [12] ANJALI A, FATIMA U, MANU M S, et al. Structure and regulation of SWEET transporters in plants: an update[J]. Plant Physiology and Biochemistry, 2020, 156: 1 − 6. doi: 10.1016/j.plaphy.2020.08.043 [13] JI J L, YANG L M, FANG Z Y, et al. Plant SWEET family of sugar transporters: structure, evolution and biological functions[J]. Biomolecules, 2022, 12(2): 205. doi: 10.3390/biom12020205 [14] NINAN A S, GRANT J, SONG J C, et al. Expression of genes related to sugar and amino acid transport and cytokinin metabolism during leaf development and senescence in Pisum sativum L.[J]. Plants, 2019, 8(3): 76. doi: 10.3390/plants8030076 [15] KLEMENS P A W, PATZKE K, DEITMER J, et al. Overexpression of the vacuolar sugar carrier AtSWEET16 modifies germination, growth, and stress tolerance in Arabidopsis[J]. Plant Physiology, 2013, 163(3): 1338 − 1352. doi: 10.1104/pp.113.224972 [16] IQBAL J, ZHANG W H, FAN Y D, et al. Genome-wide bioinformatics analysis of SWEET gene family and expression verification of candidate PaSWEET genes in Potentilla anserina[J]. Plants, 2024, 13(3): 406. doi: 10.3390/plants13030406 [17] TIAN R R, XU J Y, XU Z C, et al. Genome-wide identification and expression analysis of SWEET gene family in strawberry[J]. Horticulturae, 2024, 10(2): 191. doi: 10.3390/horticulturae10020191 [18] YUE W H, CAI K F, XIA X, et al. Genome-wide identification, expression pattern and genetic variation analysis of SWEET gene family in barley reveal the artificial selection of HvSWEET1a during domestication and improvement[J]. Frontiers in Plant Science, 2023, 14: 1137434. doi: 10.3389/fpls.2023.1137434 [19] HAN X W, HAN S, ZHU Y X, et al. Genome-wide identification and expression analysis of the SWEET gene family in Capsicum annuum L.[J]. International Journal of Molecular Sciences, 2023, 24(24): 17408. doi: 10.3390/ijms242417408 [20] CAO L H, WANG J Y, WANG L X, et al. Genome-wide analysis of the SWEET gene family in Hemerocallis citrina and functional characterization of HcSWEET4a in response to salt stress[J]. BMC Plant Biology, 2024, 24(1): 661. doi: 10.1186/s12870-024-05376-y [21] WU Y D, DI T M, WU Z J, et al. CsLHY positively regulates cold tolerance by activating CsSWEET17 in tea plants[J]. Plant Physiology and Biochemistry, 2024, 207: 108341. doi: 10.1016/j.plaphy.2024.108341 [22] WANG L, YAO L N, HAO X Y, et al. Tea plant SWEET transporters: expression profiling, sugar transport, and the involvement of CsSWEET16 in modifying cold tolerance in Arabidopsis[J]. Plant Molecular Biology, 2018, 96(6): 577 − 592. doi: 10.1007/s11103-018-0716-y [23] VALIFARD M, LE HIR R, MÜLLER J, et al. Vacuolar fructose transporter SWEET17 is critical for root development and drought tolerance[J]. Plant Physiology, 2021, 187(4): 2716 − 2730. doi: 10.1093/plphys/kiab436 [24] XU X Y, ZHANG C, WANG N, et al. Bioactivities and mechanism of actions of Dendrobium officinale: a comprehensive review[J]. Oxidative Medicine and Cellular Longevity, 2022, 2022(1): 6293355. doi: 10.1155/2022/6293355 [25] LI Y X, ZHANG T T, XING W T, et al. Comprehensive genomic characterization of the NAC transcription factors and their response to drought stress in Dendrobium catenatum[J]. Agronomy, 2022, 12(11): 2753. doi: 10.3390/agronomy12112753 [26] ZHANG G Q, XU Q, BIAN C, et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution[J]. Scientific Reports, 2016, 6(1): 19029. doi: 10.1038/srep19029 [27] HAO L, SHI X, QIN S W, et al. Genome-wide identification, characterization and transcriptional profile of the SWEET gene family in Dendrobium officinale[J]. BMC Genomics, 2023, 24(1): 378. doi: 10.1186/s12864-023-09419-w [28] CHEN C J, WU Y, LI J W, et al. TBtools-Ⅱ: a “one for all, all for one” bioinformatics platform for biological big-data mining[J]. Molecular Plant, 2023, 16(11): 1733 − 1742. doi: 10.1016/j.molp.2023.09.010 [29] SHANNON P, MARKIEL A, OZIER O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks[J]. Genome Research, 2003, 13(11): 2498 − 2504. doi: 10.1101/gr.1239303 [30] ZHANG T T, CUI Z, LI Y X, et al. Genome-wide identification and expression analysis of MYB transcription factor superfamily in Dendrobium catenatum[J]. Frontiers in Genetics, 2021, 12: 714696. doi: 10.3389/fgene.2021.714696 [31] JEENA G S, KUMAR S, SHUKLA R K. Structure, evolution and diverse physiological roles of SWEET sugar transporters in plants[J]. Plant Molecular Biology, 2019, 100(4/5): 351 − 365. doi: 10.1007/s11103-019-00872-4 [32] CHEN L Q, HOU B H, LALONDE S, et al. Sugar transporters for intercellular exchange and nutrition of pathogens[J]. Nature, 2010, 468(7323): 527 − 532. doi: 10.1038/nature09606 [33] GAUTAM T, SARIPALLI G, GAHLAUT V, et al. Further studies on sugar transporter (SWEET) genes in wheat (Triticum aestivum L.)[J]. Molecular Biology Reports, 2019, 46(2): 2327 − 2353. doi: 10.1007/s11033-019-04691-0 [34] MIZUNO H, KASUGA S, KAWAHIGASHI H. The sorghum SWEET gene family: stem sucrose accumulation as revealed through transcriptome profiling[J]. Biotechnology for Biofuels, 2016, 9(1): 127. doi: 10.1186/s13068-016-0546-6 [35] FENG C Y, HAN J X, HAN X X, et al. Genome-wide identification, phylogeny, and expression analysis of the SWEET gene family in tomato[J]. Gene, 2015, 573(2): 261 − 272. doi: 10.1016/j.gene.2015.07.055 [36] LIN Q H, ZHONG Q Z, ZHANG Z H. Identification and functional analysis of SWEET gene family in Averrhoa carambola L. fruits during ripening[J]. PeerJ, 2021, 9: e11404. doi: 10.7717/peerj.11404 [37] ZHONG L J, XU S Y, SONG C C, et al. Genome-wide identification, characterization, and expression profile of SWEETs gene family in grapevine (Vitis vinifera L. )[J]. Horticulturae, 2024, 10(5): 428. doi: 10.3390/horticulturae10050428 [38] DU Y L, LI W J, GENG J, et al. Genome-wide identification of the SWEET gene family in Phaseolus vulgaris L. and their patterns of expression under abiotic stress[J]. Journal of Plant Interactions, 2022, 17(1): 390 − 403. doi: 10.1080/17429145.2022.2044079 [39] CHEN L Q, HOU B H, LALONDE S, et al. Sugar transporters for intercellular exchange and nutrition of pathogens[J]. Nature, 2010, 468(7323): 527−532. doi:10.1038/nature09606 (查阅网上资料,本条文献与第32条文献重复,请确认) [40] SUI J L, XIAO X H, QI J Y, et al. The SWEET gene family in Hevea brasiliensis–its evolution and expression compared with four other plant species[J]. FEBS Open Bio, 2017, 7(12): 1943 − 1959. doi: 10.1002/2211-5463.12332 [41] HU W C, HUA X T, ZHANG Q, et al. New insights into the evolution and functional divergence of the SWEET family in Saccharum based on comparative genomics[J]. BMC Plant Biology, 2018, 18(1): 270. doi: 10.1186/s12870-018-1495-y [42] LIU B, DU H W, RUTKOWSKI R, et al. LAAT-1 is the lysosomal lysine/arginine transporter that maintains amino acid homeostasis[J]. Science, 2012, 337(6092): 351 − 354. doi: 10.1126/science.1220281 [43] LIU J H, PENG T, DAI W S. Critical cis-acting elements and interacting transcription factors: key players associated with abiotic stress responses in plants[J]. Plant Molecular Biology Reporter, 2014, 32(2): 303 − 317. doi: 10.1007/s11105-013-0667-z [44] QIN J X, JIANG Y J, LU Y Z, et al. Genome-wide identification and transcriptome profiling reveal great expansion of SWEET gene family and their wide-spread responses to abiotic stress in wheat (Triticum aestivum L.)[J]. Journal of Integrative Agriculture, 2020, 19(7): 1704 − 1720. doi: 10.1016/S2095-3119(19)62761-9 [45] MATHAN J, SINGH A, RANJAN A. Sucrose transport in response to drought and salt stress involves ABA-mediated induction of OsSWEET13 and OsSWEET15 in rice[J]. Physiologia Plantarum, 2021, 171(4): 620 − 637. doi: 10.1111/ppl.13210 [46] DU Y L, ZHAO Q, CHEN L R, et al. Effect of drought stress during soybean R2-R6 growth stages on sucrose metabolism in leaf and seed[J]. International Journal of Molecular Sciences, 2020, 21(2): 618. doi: 10.3390/ijms21020618 [47] JIANG L, SONG C, ZHU X, et al. SWEET transporters and the potential functions of these sequences in tea (Camellia sinensis)[J]. Frontiers in Genetics, 2021, 12: 655843. doi: 10.3389/fgene.2021.655843 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 641
- HTML全文浏览量: 593
- 被引次数: 0
