


 下载:
下载:
-
尤蠓亚属(Euprojoannisia)隶属于双翅目(Diptera)蠓科(Ceratopogonidae)铗蠓亚科(Forcipomyiinae)铗蠓属(Forcipomyia)[1]。尤蠓亚属物种数量丰富,全世界现已知110种[1]。Brèthes于1914年基于模式种Forcipomyia (Euprojoannisia) platensis Brèthes,建立了尤蠓属[2],后由Wirth和Cavalieri于1975年归入铗蠓属中成为一个亚属。该亚属雌虫为小型棕色无斑种类,触角端部5节延长,基部各节呈短球形或瓶状。触须第3节常有感觉器窝,第4、5节通常部分愈合。翅淡色,无窄鳞。足无鳞片,爪和爪间突发达。雌虫受精囊1~2个,圆形或卵形,颈明显。雄虫尾器第9腹板通常平直,有时稍凸或浅凹;第9背板较短。阳茎中叶呈三角形或方形,阳茎拱几丁质化强。阳基侧突愈合成阳基侧突弓,内陷于第9腹节[1−3]。2005年《中国蠓科昆虫》记载中国尤蠓亚属物种25种[1],近20年来,该类群研究报道较少,中国现已知28种[4−5]。目前海南岛尤蠓亚属共分布8种[5−6]。刘金华等[7]1996年首次报道了海南岛的5种尤蠓亚属昆虫,即附突铗蠓(F. appendicular Liu,Yan and Liu,1996)、小型铗蠓(F. minor Liu,Yan and Liu,1996)、淡色铗蠓(F. pipiens Liu,Yan and Liu,1996)、分离铗蠓(F. separatim Liu,Yan and Liu,1996)及雅浦铗蠓(F. yapensis Tokunaga and Murachi,1959)。王崇财等[8]于2011年记录了落羽铗蠓(F. pennielongata Chan and LeRoux,1921)及具齿铗蠓(F. calamistrata Debenham and Wirth,1984)在海南的分布。2021年,虞以新和王崇财[5]发现并描述了丘茎铗蠓(F. grumula Yu and Wang,2021)。
本研究在对采自海南岛红树林的尤蠓亚属标本的整理过程中,结合形态分类和DNA条形码进行物种鉴定,发现尤蠓亚属1新种,方形铗蠓[F. (E.) quadrata sp. nov.],以及1海南新纪录种,粗野铗蠓[F. (E.) psilonota Kieffer,1911],首次发现并报道了附突铗蠓的雌虫。
-
研究标本为2020—2023年通过灯诱法、网捕法和马氏网法采自海南岛各地红树林内尤蠓亚属成虫乙醇标本。首先使用奥林巴斯SZX16体视显微镜以及其连接的CCD(DP72)拍摄成虫的整体轮廓图,参考吴小香[9]的方法对标本进行无损形态基因组DNA提取、DNA条形码测序和制作永久玻片标本[10]。使用生物显微镜XSP−12CA和CCD(YH5001)对玻片标本的头、胸、翅、尾器等鉴别特征进行显微拍照并测量相关数据,利用Adobe Illustrator CC 2018和Adobe Photoshop CC2020对雄性外生殖器进行绘制和图版制作。
-
物种描述中形态术语及测量数据依据Debenham和Wirth[11]的方法,翅脉和翅室依据Borkent[12]的命名。测量数据表现形式为最小值~最大值(平均值,n=检视标本数量)或[(最大值~最小值)×(最大值~最小值)]。所有模式标本和检视标本均保存于海南大学热带农林学院昆虫标本馆。
-
DNA条形码的物种鉴定以细胞色素C氧化酶亚基Ⅰ(CO Ⅰ)基因作为DNA条形码基因,通用引物为LCO1490:5′−GG TCAACAAATCATAAAGATATTGG−3′和HCO2198:5′−TAAACTTCAGGGTGACCAAAAAATCA−3′,参考吴小香[9]的方法进行PCR扩增和测序。选择并下载NCBI数据库中铗蠓属的台湾蠛蠓F. taiwana Shiraki,1913(GenBank 登录号:KF528689)和F. bipunctata Linnaeus,1767(GenBank 登录号:TTMDI825-10)CO I序列作为外群,利用MEGA 11.0采用Kimura双参数模型(Kimura-2-Parameter,K2P)计算种内和种间遗传距离,构建系统发育树,并进行自举检验(Bootstrap=
2000 )[13]。 -
缩写:触角比(AR),触角鞭节第9~13节总长与鞭节第1~8节总长的比值;翅前缘脉比(CR),前缘脉长与翅长的比值;触须比(PRⅢ),触须第3节长与宽的比值;前足跗节比(TRⅠ)、中足跗节比(TRⅡ)、后足跗节比(TRⅢ),前、中、后足第1跗节与第2跗节的长度比值。
-
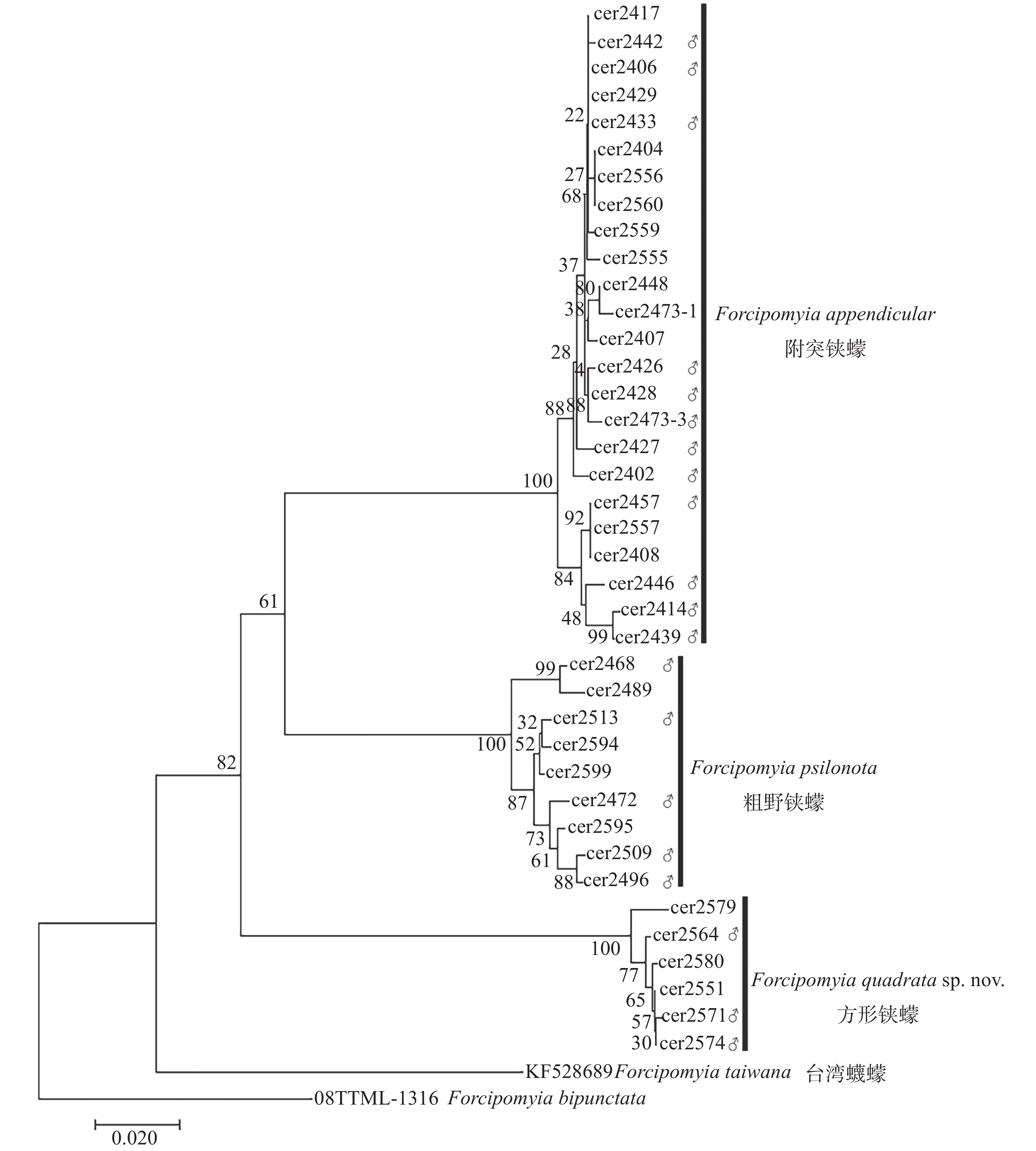
本研究在对雄虫形态分类鉴定的基础上,辅助以DNA条形码技术对方形铗蠓、粗野铗蠓和附突铗蠓的雌虫进行了鉴定,共获得雄性个体CO Ⅰ序列20条,雌性个体CO Ⅰ序列19条,拼接后的序列长度为658 bp。利用邻接法构建的系统发生关系(图1)显示,3种铗蠓的雌虫与雄虫的CO Ⅰ序列各自聚成一支,支持率均为100%。基于K2P模型分别计算种内和种间遗传距离,方形铗蠓种内遗传距离介于0~1.5%,平均遗传距离为0.6%,粗野铗蠓种内遗传距离为0.3%~3.1%,平均遗传距离为1.7%,附突铗蠓种内遗传距离为0~2.5%,平均遗传距离为1%,3种铗蠓的种间遗传距离为13.9%~19.8%。DNA条形码技术分析结果与形态学鉴定结果一致。
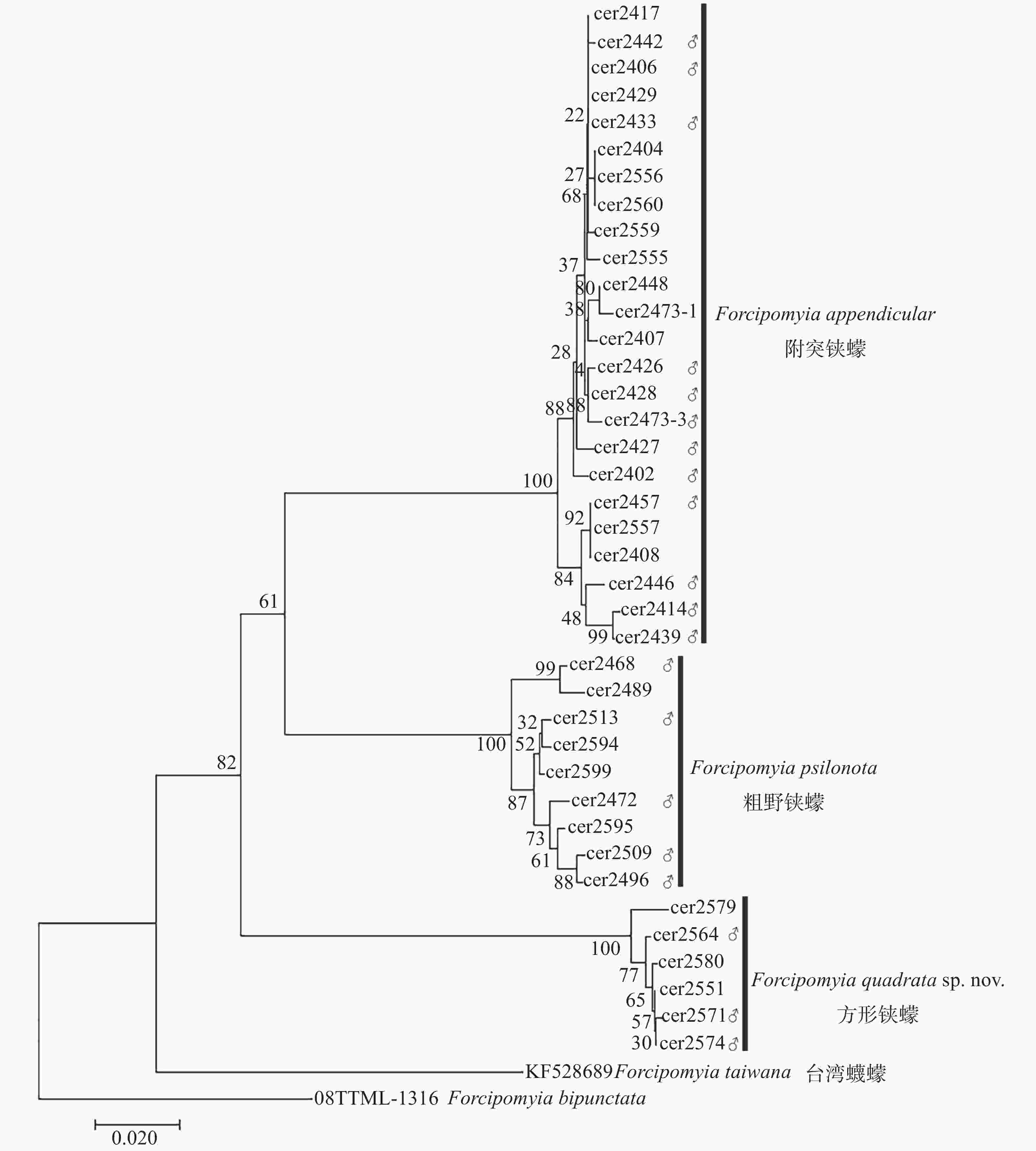
图 1 基于CO I序列利用邻接法构建尤蠓亚属昆虫系统发育树
Figure 1. Neighbor-Joining tree of the subgenus Euprojoannisia based on mitochondrial CO I sequence
-
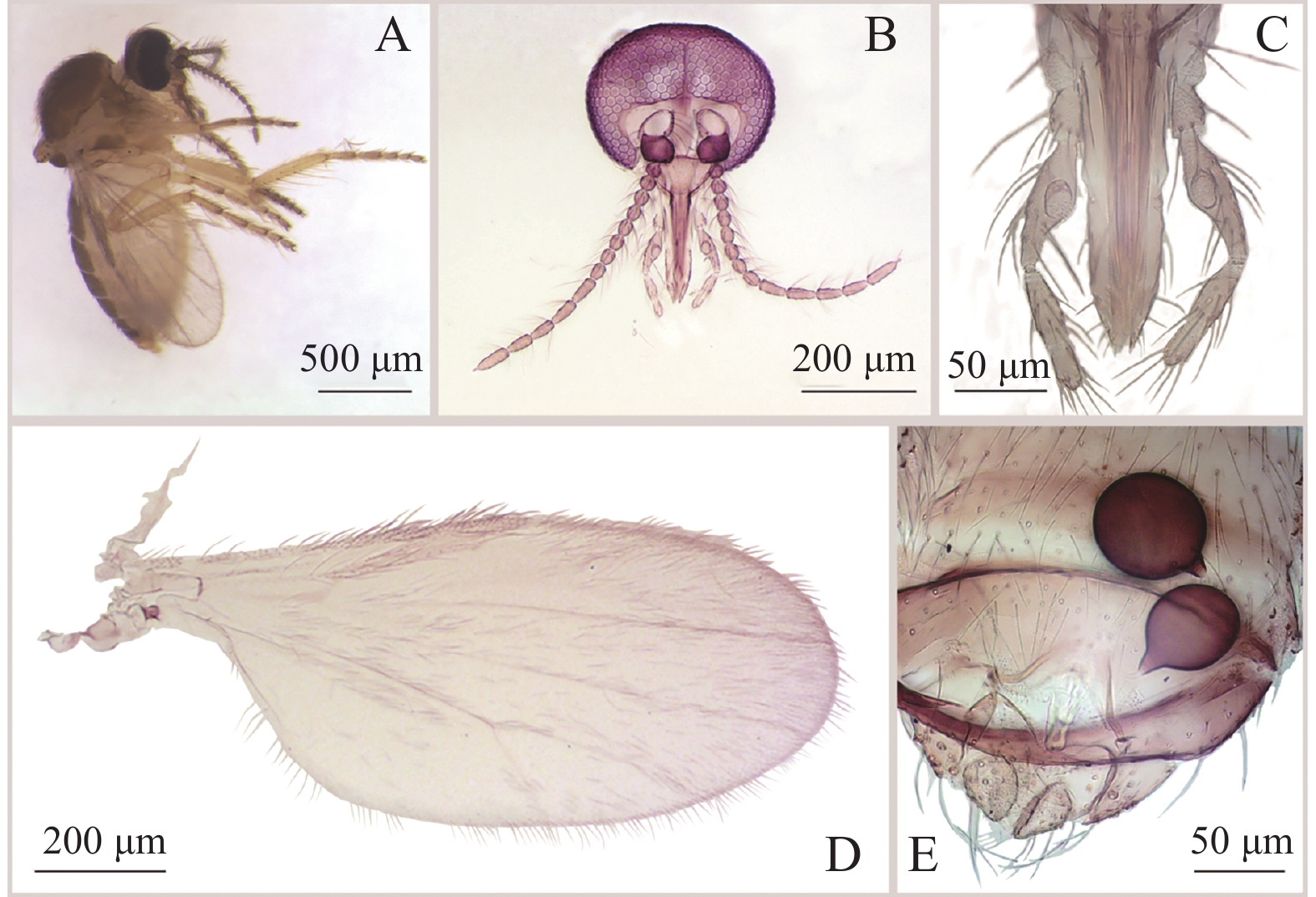
方形铗蠓F. quadrata Meng, Li and Lin sp. nov. (图2、图3)鉴别特征:雄虫第9腹板隆起,后缘中部近方形凸起,密布刚毛;阳茎中叶背侧近端部具2根长刺突;阳基侧突弓底窄平。雌虫触须第3节中部明显膨大,具感觉器窝;大颚密布30余枚细齿,小盾片后缘具粗鬃10~11根;具2个球形不等大的受精囊,其表面无刻点,基部有长颈。
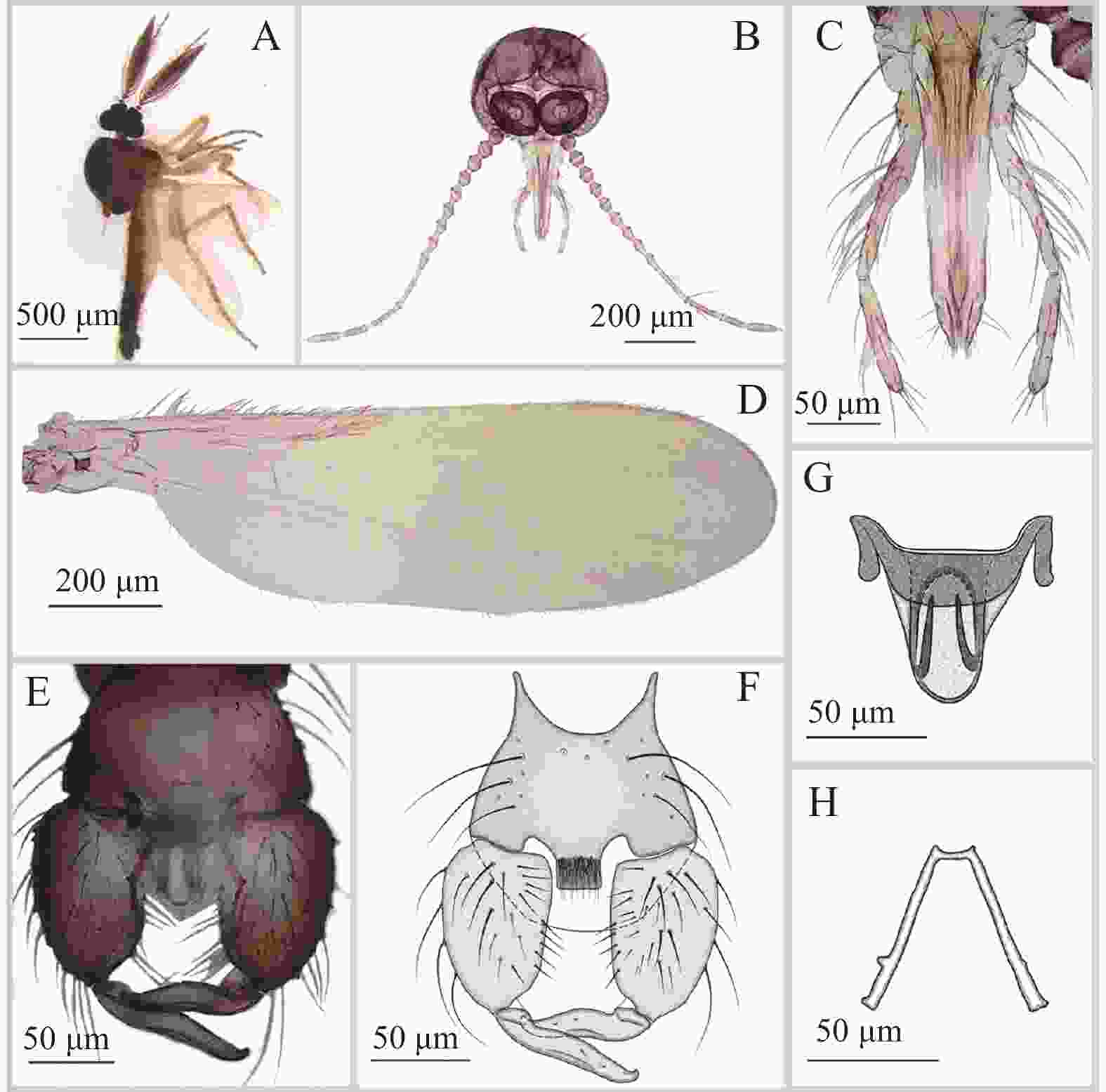
图 2 方形铗蠓Forcipomyia (Euprojoannisia) quadrata Meng,Li and Lin sp. nov.,正模,雄性
Figure 2. Forcipomyia (Euprojoannisia) quadrata Meng, Li and Lin sp. nov., male
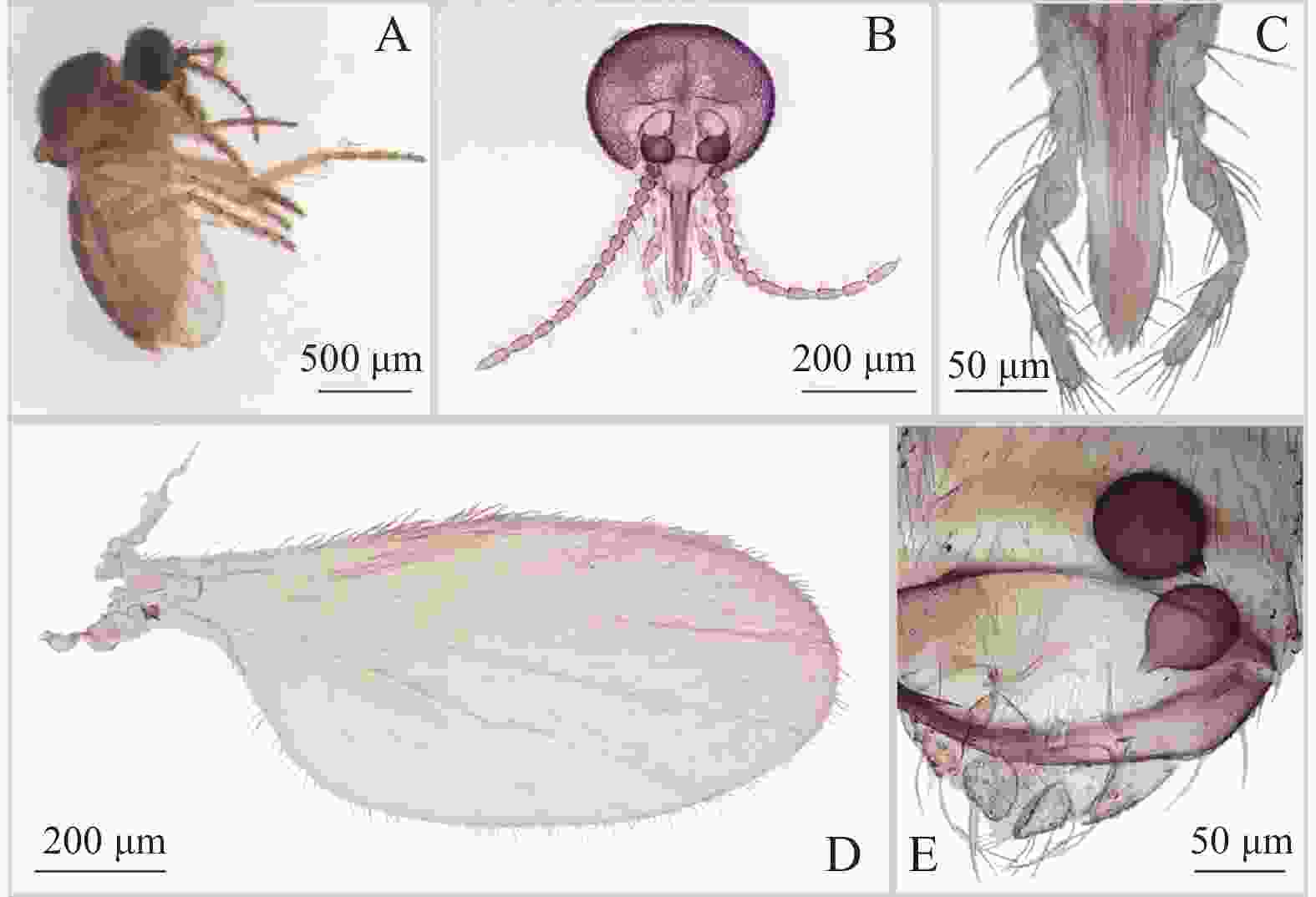
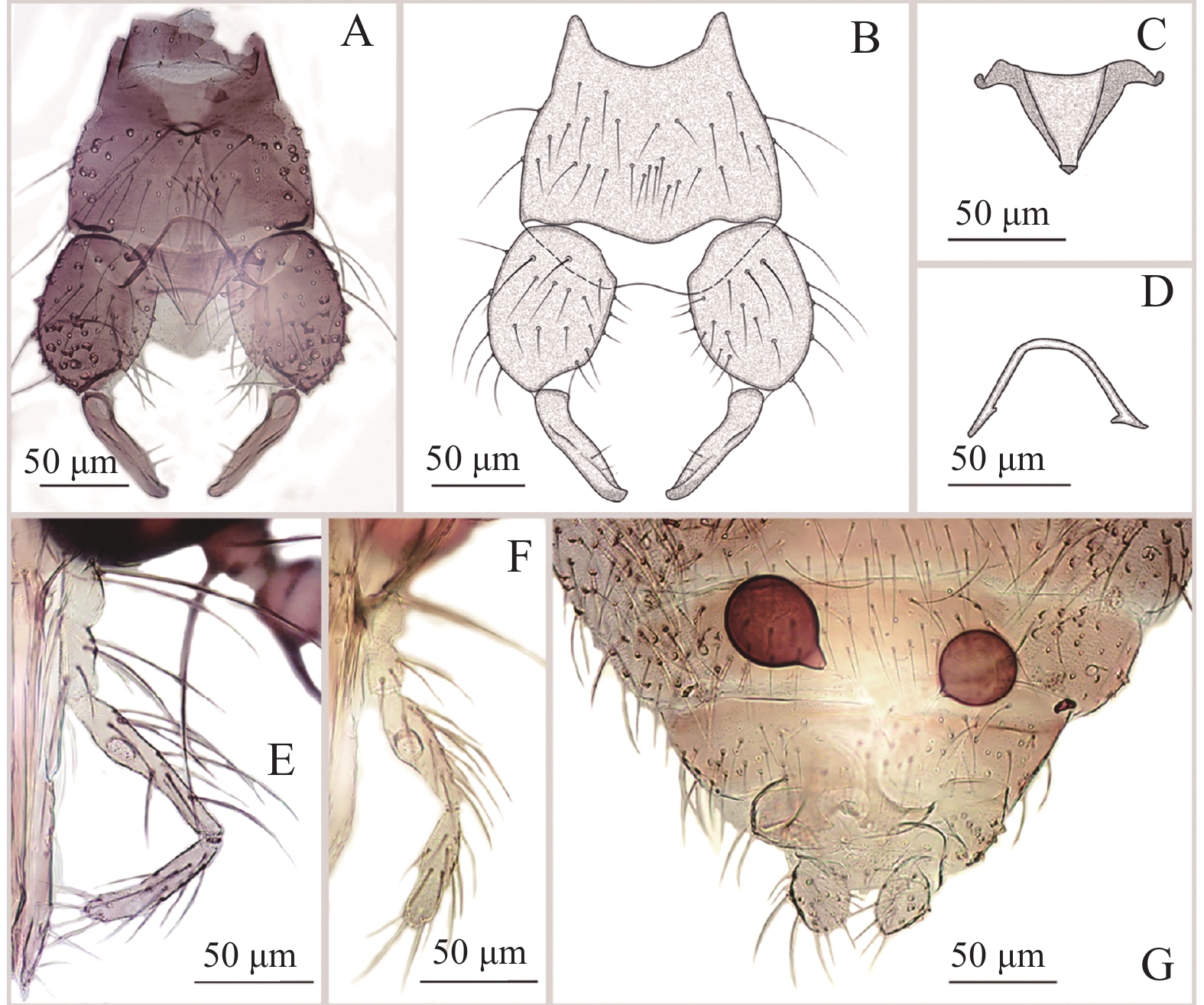
图 3 方形铗蠓Forcipomyia (Euprojoannisia) quadrata Meng,Li and Lin sp. nov.,雌虫
Figure 3. Forcipomyia (Euprojoannisia) quadrata Meng, Li and Lin sp. nov., female
物种描述:
雄成虫(图2−A)翅长1.23~1.40 mm(1.32 mm,n=6),翅宽0.40~0.42 mm(0.41 mm,n=6)。
体色:复眼棕褐色,触角、触须棕色;中胸背板棕色,侧面除侧背片棕色外其余颜色较浅,小盾片颜色较浅;翅面无斑,前缘脉与径分脉颜色明显加深;平衡棒色淡;足浅棕色;腹部背板第6~8节深棕色,其余颜色较浅。
头部:复眼相接,小眼面间无柔毛(图2−B)。触角轮毛发达,鞭节端部4节明显延长,第10节最长,第13节具乳头状端突;AR 1.27~1.30(1.28,n=4)。触须(图2−C)分5节;第3节细长,近基部稍膨大,具明显感觉器窝;第4~5节部分愈合;各节长度比为36∶40∶123∶49∶44,PRⅢ 5.04~5.51(5.27,n=5)。
胸部:前翅(图2−D)径1室封闭,径2室狭长,翅面大毛稀疏,仅见于径3室、中1室及中2室端部,前缘脉与径分脉具少量粗鬃,CR 0.40~0.42(0.41,n=6)。小盾片后缘具粗鬃9~11根。前足股节略长于胫节,中足和后足股节略短于胫节;前足胫节末端具1细长的距;后足胫端鬃6~7根,梳齿11~12枚,后足第1跗节具成排羽状鬃,无鳞片。前足TRⅠ 0.93~1.12(1.06,n=6),中足TRⅡ 0.55~0.60(0.58,n=6),后足TRⅢ 0.60~0.71(0.65,n=6)。爪细长而弯曲,端部不分叉,爪间突发达。
腹部:抱握器(图2−E、F)基节粗壮,端节细短,近基部明显内凹。第9腹板隆起,后缘中部近方形外凸,密被刚毛。阳茎中叶(图2−G)结构特殊,基部强几丁质化,背侧向端部渐窄,端部弧形,近端部具2根反向刺突;阳基侧突弓底窄平(图2−H),近端部外侧具一小凸起。
雌成虫形态特征与雄虫基本一致,不同如下:
雌成虫(图3−A)翅长0.98~1.06 mm(1.02 mm,n=2),翅宽0.47~0.50 mm(0.48 mm,n=2)。
体色:除腹部背板为一致棕色外,雌性体色与雄性别无二致。
头部:触角(图3−B)端部5节明显延长,AR 1.20~1.24(1.22,n=2)。触须(图3−C)第3节中部明显膨大,具感觉器窝;第4、5节几乎完全愈合;各节长度比为27∶29∶83∶40∶29,PR 2.80~3.04(2.92,n=2)。大颚密布30余枚细齿。
胸部:前翅(图3−D)径2室狭小,翅面除臀室外大毛密布,前缘脉及径分脉具粗鬃,CR 0.49~0.52(0.50,n=2)。小盾片后缘具粗鬃10~11根。前足股节与胫节几乎等长;前足股节中部略膨大,后足胫端鬃6根,梳齿10~11枚。前足TRⅠ 1.65~1.66(1.65,n=2),中足TRⅡ 1.18~1.19(1.19,n=2),后足TRⅢ 1.09~1.11(1.10,n=2)。爪细短而弯曲。
腹部:殖下板发达,呈桥拱状。受精囊2个(图3−E),球形,不等大(74.81 μm×71.61 μm,63.42 μm×56.90 μm),表面无刻点,具长颈。
词源:种名来自拉丁词“quadratus”(方形的),依据于雄虫形态特征第9腹板后缘中部呈方形,词性为阴性。
小结:本新种似落羽铗蠓,但本新种阳基侧突弓底窄平,两侧近端部具凸起。
正模:♂,海南省万宁市万城镇周家庄村红树林,
18.8382 °N,110.4419 °E,H 8 m,2021-Ⅺ-20,孟祥瑞、邓斌、杨浩,灯诱,cer2564。副模:2♂2♀,海南省三亚市铁炉港红树林自然保护区,18.2783 °N,109.7077 °E,H 3 m,2022-Ⅰ-09,孟祥瑞、邓斌、杨浩,灯诱,cer2471、cer2574、cer2579、cer2580;3♂1♀,海口塔市红树林,19.9982 °N,110.5407 °E,H 6 m,2022-Ⅳ-20,孟祥瑞、邓斌、杨浩,马氏网,cer2551、cer2714、cer2722、cer2723。国内分布:海南(海口、万宁、三亚)。
-
粗野铗蠓F. psilonota Kieffer, 1911见图4。Ceratopogon psilonotus Kieffer,1911:337[14]. 模式标本产地:塞舌尔。
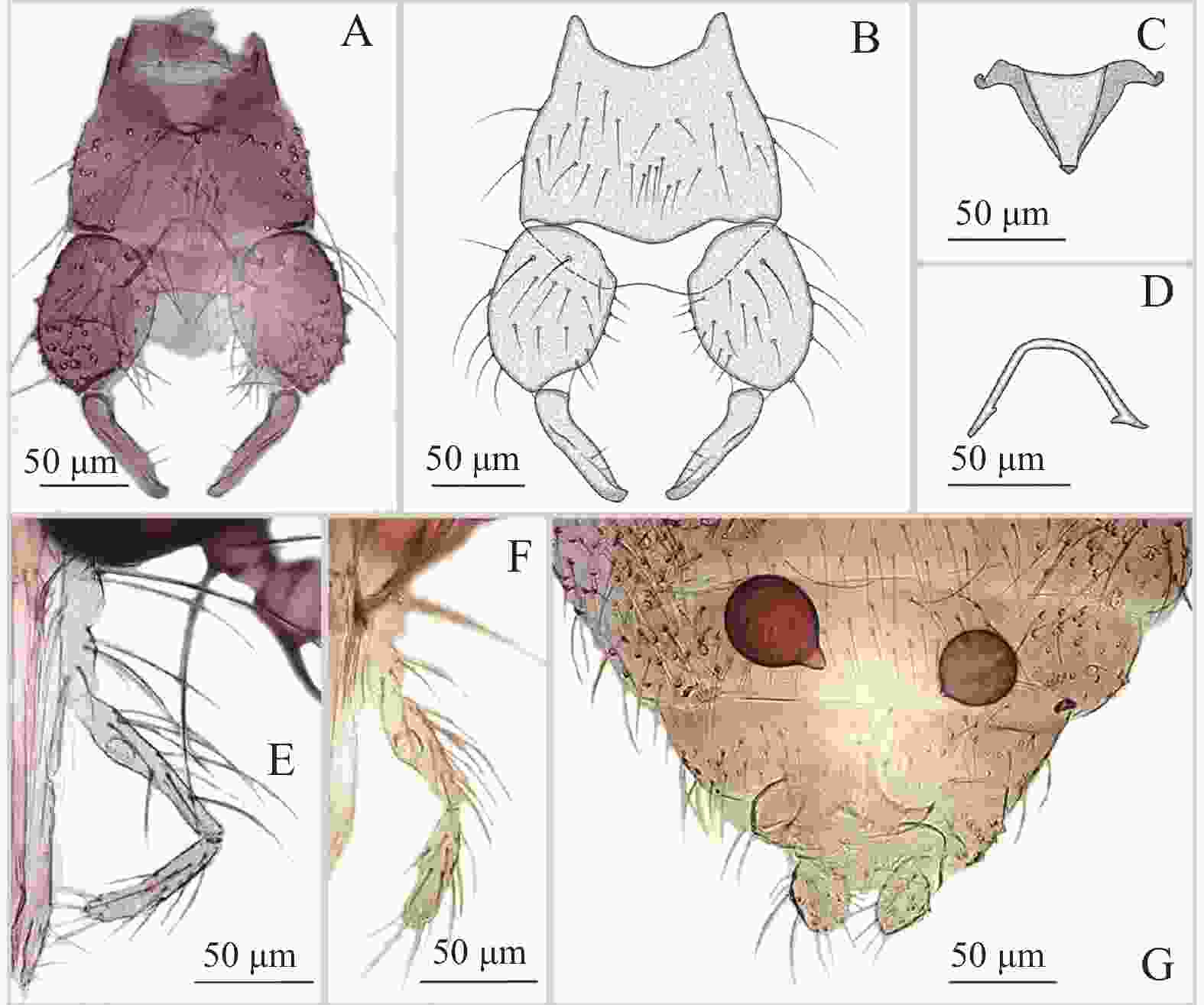
图 4 粗野铗蠓Forcipomyia (Euprojoannisia) psilonota Kieffer,1911,雄性及雌性
Figure 4. Forcipomyia (Euprojoannisia) psilonota Kieffer, 1911, male and female
Ceratopogon aplonota Kieffer,1911:337. 模式标本产地:塞舌尔.
Ceratopogon seychelleana Kieffer,1911:338. 模式标本产地:塞舌尔。
Ceratopogon fulvithorax Kieffer,1911:338. 模式标本产地:塞舌尔。
Forcipomyia psilonota (Kieffer);Ingram and Macfie,1924:546[15];Wirth and Howarth,1982:146;Yu et al. 2005:515[1]. 模式标本产地:埃及。
Forcipomyia indecora Kieffer,1914:269[16];Dessart,1963:73. 模式标本产地:南非。
Forcipomyia litoralis Santos Abreu,1918:277[17]. 模式标本产地:西班牙。
Forcipomyia ingrami Carter,1919:290. 模式标本产地:加纳。
Forcipomyia egypti Macfie,1924:61. 模式标本产地:埃及。
Forcipomyia hathor Kieffer,1925e:247. 模式标本产地:埃及。
Forcipomyia conogensis Goetghebuer,1933:132. 模式标本产地:刚果民主共和国。
Forcipomyia flavipilosella Goetghebuer,1933:130. 模式标本产地:刚果民主共和国。
Forcipomyia lulengaensis Goetghebuer,1935:155. 模式标本产地:刚果民主共和国。
Forcipomyia superata Goetghebuer,1935:160. 模式标本产地:刚果民主共和国。
Forcipomyia griseipluma Goetghebuer,1935:154. 模式标本产地:刚果民主共和国。
Forcipomyia griseolella Goetghebuer,1948:7. 模式标本产地:刚果民主共和国。
Forcipomyia (Euprojoannisia) sauteri Kieffer 1912;Tokunaga and Murachi,1959:190;Wirth and Howarth,1982:147[18]. 模式标本产地:美国。
Forcipomyia (Proforcipomyia) clara Chan et LeRoux,1971:729[19]. 模式标本产地:新加坡.
鉴别特征:雄性抱握器基节粗壮,长约为宽的2/3;阳茎中叶三角形,顶端具1三角形的小端突,两侧几丁质化加厚,阳茎拱较低;阳基侧突呈宽U形。雌性大颚密布30余枚细齿,小颚齿12枚;小盾片后缘具粗鬃8~10根;殖下板发达,两侧较厚;受精囊2个,球形,具长颈。
检视标本:1)3♂,海南临高新盈湿地自然保护区红树林,19.868 5°N,109.549 6°E,H 4 m,2022-Ⅺ-29,孟祥瑞、邓斌、杨浩,灯诱,cer2472、cer2683、cer2686;2)2♂,海南省陵水黎族自治县新村镇桐栖村红树林,18.435 9°N,109.991 3°E,H 13 m,2022-Ⅰ- 09,孟祥瑞、邓斌、杨浩,灯诱,cer2496、cer2509;3)1♂,海南省琼海市沙美红树林湿地公园内红树林,19.109 3°N,110.555 0°,H 11 m,2021-Ⅺ-20,孟祥瑞、邓斌、杨浩,灯诱,cer2513;4)3♀,海南省三亚市铁炉港红树林自然保护区内红树林,
18.2783 °N,109.707 7°E,H 7 m,2022-Ⅰ-09,孟祥瑞、邓斌、杨浩,灯诱,cer2594、cer2595、cer2599。国外分布:塞舌尔,埃及,南非,西班牙,加纳,刚果民主共和国,美国,新加坡。
国内分布:广东(中山),福建(南平),四川(宜宾),陕西(汉中、西安),安徽(淮南),澳门,海南(临高、陵水、琼海、三亚)。
-
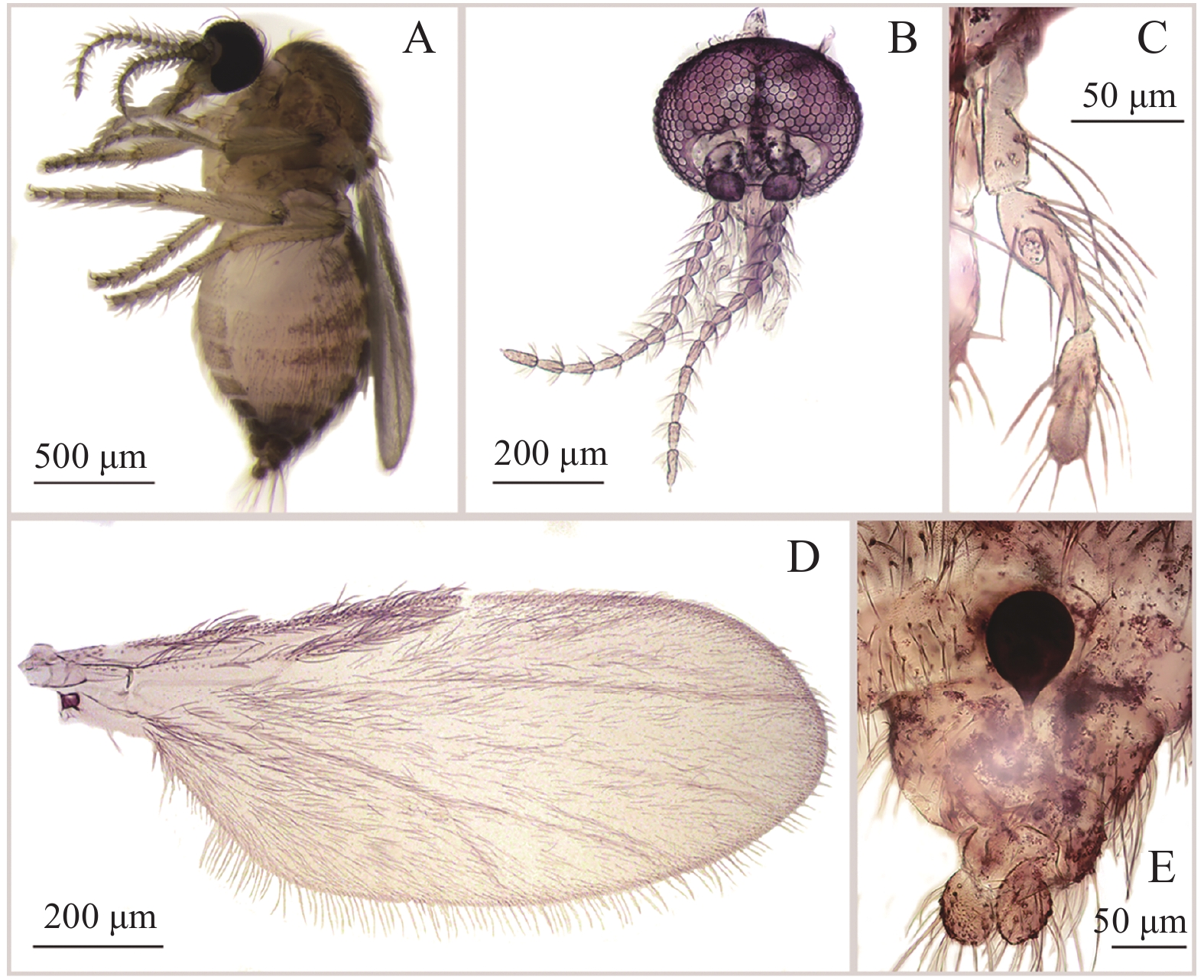
[F. appendicular Liu, Yan and Liu, 1996] 雌虫描述见图5。Forcipomyia (Euprojoannisia) appendicular Liu,Yan and Liu,1996:18[6];Yu et al. 2005:490[1]. 模式标本产地:中国。
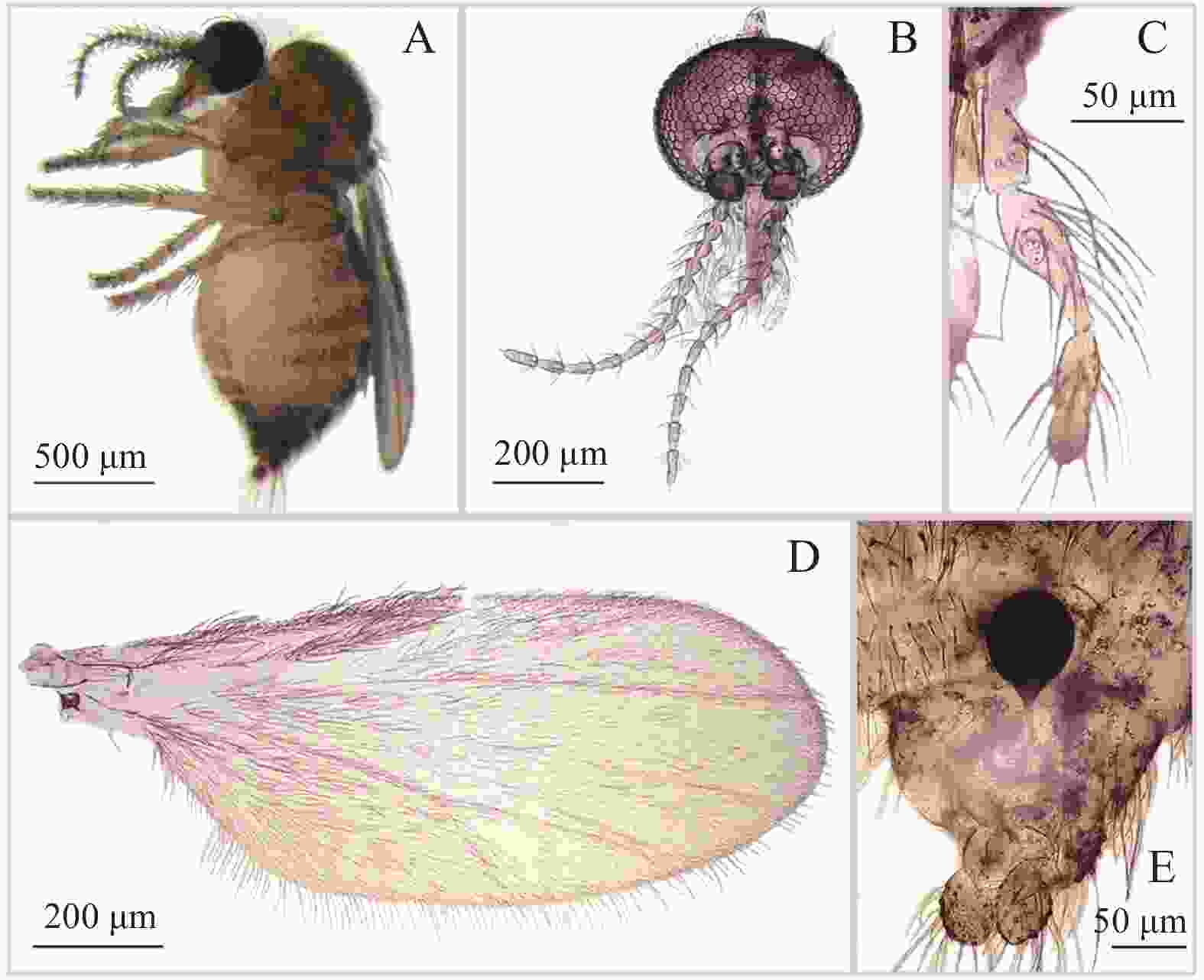
图 5 附突铗蠓Forcipomyia (Euprojoannisia) appendicular Liu,Yan and Liu,1996,雌性
Figure 5. Forcipomyia (Euprojoannisia) appendicular Liu,Yan and Liu,1996,female
雌性鉴别特征:触须第3节近基部膨大,膨大处具明显感觉器窝;大颚具20余枚细齿;小盾片后缘具粗鬃7~9根;殖下板基部不相接,呈不完全的圆环状,两侧加厚;仅1个球形受精囊,具不弯曲的长颈。
物种描述:雌成虫(图5−A)翅长0.99~1.24 mm(1.09 mm,n=6),翅宽0.41~0.55 mm(0.48 mm,n=6)。
体色:复眼棕褐色,触角棕色,触须浅棕色;中胸背板及小盾片棕色,侧背片浅棕色;翅面无色斑;平衡棒淡色;足一致浅棕色;腹部背板棕色。
头部:复眼相接,小眼面间无柔毛(图5−B)。触角鞭节基部各节近瓶形,端部5节明显延长,第13节具乳头状端突,AR 0.92~0.98(0.96,n=5)。触须(图5−C)分5节,第3节近基部膨大,膨大处具明显感觉器窝;第4、5节部分愈合;各节相长度比为33∶43∶81∶34∶25,PRⅢ 2.19~2.96(2.59,n=6)。大颚具20余枚细齿。
胸部:前翅(图5−D)径1室封闭,径2室狭小;翅面和翅外缘密被大毛,前缘脉和径分脉具粗鬃,CR 0.48~0.51(0.49,n=6)。小盾片后缘具粗鬃7~9根。前、中、后足股节与胫节近等长;前足胫节末端具1枚细短的距,被少量鬃毛;后足胫端鬃5~6根,梳齿7~10枚,第1跗节具成排羽状鬃。前足TRⅠ 1.75~1.80(1.78,n=6),中足TRⅡ 1.15~1.18(1.16,n=6),后足TRⅢ 1.31~1.5(1.39,n=6)。爪细长而弯曲,顶端不分叉,爪间突发达。
腹部:殖下板基部不相接,呈不完全圆环状,两侧加厚;受精囊(图5−E)1个,球形[(80.81~63.90)×(69.14~57.43)]μm,有或无刻点,具不弯曲的长颈。
小结:本研究基于形态特征鉴定了附突铗蠓的雄虫,通过DNA条形码鉴定首次发现了附突铗蠓的雌虫,并描述了雌虫的形态特征。尤蠓亚属多数种类具有2个受精囊,中国记录的具有单受精囊的尤蠓亚属共4种,除附突铗蠓外还有巴河铗蠓[F. (E.) bahelea Liu and Yu,2001]、岭南铗蠓[F. (E.) lingnanensis Liu and Yu,1999]和强直铗蠓[F. (E.) tonicus Liu and Yu,2001][20];但附突铗蠓受精囊及殖下板的形状与另外3种区别明显,附突铗蠓为球形受精囊,而巴河铗蠓和强直铗蠓均为葫芦形受精囊;附突铗蠓殖下板基部不相接,呈不完全圆环状,而岭南铗蠓的殖下板为拱桥状。
检视标本:1)9♂5♀,海南省海口市东寨港保护区红树林,19.947 7°N,110.579 6°E,H 5 m,2022-Ⅰ-20,孟祥瑞、邓斌、杨浩,马氏网,cer2402、cer2404、cer2406、cer2407、cer2408、cer2414、cer2417、cer2419、cer2426、cer2427、cer2428、cer2429、cer2433、cer2439;2)2♂6♀,海口塔市红树林,19.998 2°N,110.540 7°E,H 6 m,2022-Ⅳ-20,孟祥瑞、邓斌、杨浩,马氏网,cer2442、cer2446、cer2448、cer2555、cer2556、cer2557、cer2559、cer2560;3)1♂,海南省海口市美兰区北港村红树林,20.022 4°N,110.564 1°E,2022-Ⅴ-19,孟祥瑞、邓斌、杨浩,马氏网,cer2457;4)1♂,海南省海口市秀英区石山火山群国家地质公园,19.959 858°N,110.219 065°E,H 23 m,2022-Ⅶ-22,李加慧、孟祥瑞、杨浩,灯诱,cer2653;5)1♂1♀,海南省文昌市会文镇上圮村红树林,19.451 5°N,110.743 2°E,H 12 m,2021-Ⅺ-18,孟祥瑞、邓斌、杨浩,马氏网,cer2473-1、cer2473-3;6)1♂,海南省琼海市沙美红树林湿地公园内红树林,19.109 3°N,110.555 0°,H 11 m,2021-Ⅺ-20,孟祥瑞、邓斌、杨浩,灯诱,cer2614;7)1♂,海南省三亚市铁炉港红树林自然保护区内红树林,18.278 3°N,109.707 7°E,H 7 m,2022-Ⅰ-09,孟祥瑞、邓斌、杨浩,灯诱,cer2634;8)1♂,海南临高新盈湿地自然保护区红树林,19.868 5°N,109.549 6°E,H 4 m,2022-Ⅺ-29,孟祥瑞、邓斌、杨浩,灯诱,cer2697。
国内分布:海南(琼中、海口、文昌、琼海、临高、三亚),贵州(贵阳),香港,陕西(汉中),江西(南昌、庐山),广东(珠海),安徽(淮南)。
-
尤蠓亚属全世界现已知110种;而海南岛目前仅记录8种,本研究记录了尤蠓亚属一新种和一海南新纪录种以及附突铗蠓雌性的形态特征,为附突铗蠓雌虫的物种鉴定提供了依据,同时进一步丰富了海南岛蠓科昆虫的物种多样性。
-
尤蠓亚属物种体型较小,虫体和翅上由于缺少色斑等便于物种鉴定的关键形态特征,同一物种不同性别的配对较为困难,导致目前世界已记录的110个物种中近70种仅记述了单一性别,仍有大量物种的雌性或雄性亟待发现和描述。鉴于传统形态分类的局限性,DNA条形码技术逐渐被应用于铗蠓亚科这类小型昆虫的物种鉴定中,其中在棕榈裸蠓(Atrichopogon palmatus Tokunaga,1962)雄虫的发现[13]及高飞铗蠓(F. praealtus Liu and Yu,2001)的物种鉴定中发挥了重要作用[21]。本研究结果表明DNA条形码技术与传统形态分类学相结合,有助于尤蠓亚属新种、新纪录种的发现及单性别物种缺失性别的发现。
-
本研究以海南岛红树林内尤蠓亚属昆虫为研究对象,以传统分类学为主,辅助以DNA条形码进行物种鉴定,详细描述新种方形铗蠓[F. (E.) quadrata sp. nov.]、海南新纪录种粗野铗蠓[F. (E.) psilonota Kieffer,1911]、附突铗蠓[F. (E.) appendicular Liu,Yan and Liu,1996]雌虫的鉴别特征,丰富了海南岛蠓科昆虫的物种多样性。
海南岛铗蠓属尤蠓亚属(双翅目:蠓科)一新种及一海南新纪录种
DOI: 10.15886/j.cnki.rdswxb.20240071
 CSTR: 32425.14.j.cnki.rdswxb.20240071
CSTR: 32425.14.j.cnki.rdswxb.20240071
A new species and a newly record species of genus Forcipomyia subgenus Euprojoannisia (Diptera: Ceratopogonidae) from Hainan Island
-
摘要: 尤蠓亚属(Euprojoannisia)是铗蠓属(Forcipomyia)35个亚属中物种数量仅次于铗蠓亚属(Forcipomyia)和蠛蠓亚属(Lasiohelea)的第三大亚属。本研究结合形态鉴定和DNA条形码技术,对海南岛红树林尤蠓亚属昆虫进行鉴定,发现并描述了海南岛尤蠓亚属一新种,方形铗蠓[F.(E.)quadrata sp. nov.]以及一海南新纪录种粗野铗蠓[F.(E.)psilonota Kieffer,1911],首次发现并描述了附突铗蠓[F.(E.)appendicular Liu,Yan and Liu,1996]雌虫特征。Abstract: Euprojoannisia Brèthes is one of the 35 subgenera of genus Forcipomyia Meigen, known as the third largest subgenus in terms of the number of species just behind subgenus Forcipomyia s.str. and subgenus Lasiohelea Kieffer. The specimens of subgenus Euprojoannisia were collected from the mangrove forests in Hainan Province and identified by morphological characters and DNA barcodes. A new species, F. (E.) quadrata sp. nov., and a new record species of Hainan Province, F. (E.) psilonota Kieffer, 1911, were found and described. Besides, the characteristics of the females of F. (E.) appendicular Liu, Yan and Liu, 1996, were first reported.
-
Key words:
- morphological characters /
- taxonomy /
- DNA barcoding /
- molecular identification /
- mangrove forest
-
图 2 方形铗蠓Forcipomyia (Euprojoannisia) quadrata Meng,Li and Lin sp. nov.,正模,雄性
注:A. 整体侧面观;B. 头;C.触须;D.前翅;E. 尾器,腹面观;F.尾器(不含阳茎中叶和阳基侧突),腹面观;G. 阳茎中叶;H. 阳基侧突。
Fig. 2 Forcipomyia (Euprojoannisia) quadrata Meng, Li and Lin sp. nov., male
Note: A. Habitus in lateral view;B. Head;C. Palpi;D. Forewing;E. Hypopygium,in ventral view;F. Hypopygium exclude aedeagus and parameres,in ventral view;G. Aedeagus;H. Parameres.
图 3 方形铗蠓Forcipomyia (Euprojoannisia) quadrata Meng,Li and Lin sp. nov.,雌虫
注:A. 整体侧面观;B. 头;C. 触须;D. 前翅;E. 受精囊。
Fig. 3 Forcipomyia (Euprojoannisia) quadrata Meng, Li and Lin sp. nov., female
Note: A. Habitus in lateral view;B. Head;C. Palpi;D. Forewing;E:Spermathecae.
图 4 粗野铗蠓Forcipomyia (Euprojoannisia) psilonota Kieffer,1911,雄性及雌性
注: A. 雄虫尾器,腹面观;B. 雄虫尾器(不含阳茎中叶和阳基侧突),腹面观;C. 阳茎中叶;D. 阳基侧突;E. 雄虫触须;F. 雌虫触须;G.雌虫受精囊。
Fig. 4 Forcipomyia (Euprojoannisia) psilonota Kieffer, 1911, male and female
Note: A. Male hypopygium,in ventral view;B. Male hypopygium exclude aedeagus and parameres,in ventral view;C. aedeagus;D. parameres;E. Male palpi;F. Female palpi;G. Female spermathecae.
-
[1] 虞以新. 中国蠓科昆虫名录及其检索表(昆虫纲, 双翅目)[M]. 北京: 军事医学科学出版社, 2005. [2] BRÈTHES J. Descripcion de six Cécidomyidae (Dipt.) de Buenos Aires[J]. Anales del Museo Nacional de Buenos Aires, 1914, 26: 151 − 156. [3] HAN X J, LI X F, CHANG Q Q, et al. Description of a new species of Forcipomyia (Euprojoannisia) Brèthes, 1914 (Diptera: Ceratopogonidae) and a key to Chinese species of the subgenus[J]. Entomologica Fennica, 2017, 28(2): 107 − 112. doi: 10.33338/ef.84680 [4] BORKENT A, DOMINIAK P. Catalog of the biting midges of the world (Diptera: Ceratopogonidae)[J]. Zootaxa, 2020, 4787(1): 1 − 377. doi: 10.11646/zootaxa.4787.1.1 [5] 虞以新, 王崇财. 铗蠓属二新种的描述(双翅目: 蠓科)[J]. 中华卫生杀虫药械, 2021, 27(3): 261 − 262. doi: 10.19821/j.1671-2781.2021.03.019 [6] 王崇财. 海南省蠓类鉴定集[M]. 北京: 中国质检出版社, 2018: 1−118. [7] 刘金华, 严格, 刘国平. 海南岛的蠓类[M]. 北京: 军事医学科学出版社, 1996: 1-184. [8] 王崇财, 陈星恒, 谭润谦, 等. 海南蠓科昆虫名录(双翅目: 蠓科)[J]. 中国国境卫生检疫杂志, 2011, 34(1): 24 − 27. doi: 10.16408/j.1004-9770.2011.01.001 [9] 吴小香, 杨浩, 孟祥瑞, 等. 阿蠓属一新种及一中国新记录种(昆虫纲: 双翅目: 蠓科)[J]. 环境昆虫学报, 2022, 44(4): 1043 − 1052. doi: 10.3969/j.issn.1674-0858.2022.04.30 [10] 邓斌, 林晓丹, 李加慧. 海南省红树林生境库蠓属一新种及一中国新记录种记述(双翅目: 蠓科)[J]. 中国媒介生物学及控制杂志, 2023, 34(4): 552 − 558. doi: 10.11853/j.issn.1003.8280.2023.04.020 [11] DEBENHAM M L, WIRTH W W. Australian and new guinea species of the Forcipomyia subgenus Euprojoannisia (Diptera: Ceratopogonidae)[J]. Australian Journal of Zoology, 1984, 32(6): 851 − 889. doi: 10.1071/ZO9840851 [12] BORKENT A. Ceratopogonidae (biting midges)[M]//KIRK-SPRIGGS A H, SINCLAIR B J. Manual of afrotropical diptera. Volume 2. Nematocerous diptera and lower brachycera. Suricata 5. Pretoria: South African National Biodiversity Institute, 2017: 733-812. [13] 何泽华, 李加慧, 林晓丹. 基于DNA条形码的棕榈裸蠓雄虫的发现与描述[J]. 热带生物学报, 2024, 15(1): 94 − 99. doi: 10.15886/j.cnki.rdswxb.20230012 [14] KIEFFER J J. Les chironomides (Tendipedidae) de l'Himalaya et d'Assam[J]. Records of the Indian Museum, 1911, 6: 319 − 349. [15] INGRAM A, MACFIE J W S. Notes on some African Ceratopogoninae-species of the genus Forcipomyia[J]. Annals of Tropical Medicine & Parasitology, 1924, 18(4): 533 − 593. doi: 10.1080/00034983.1924.11684432 [16] KIEFFER J J. South African Chironomidae (Diptera)[J]. Annals of the South African Museum. Annale van die Suid-Afrikaanse Museum, 1914, 10: 259 − 270. doi: 10.5962/bhl.part.9317 [17] ABREU E S. Ensayo de una monografía de los tendipedidos de las islas canarias[J]. Memorias de la Real Academia de Cienciasy Artes de Barcelona, 1918, 14(2): 159 − 326. doi: 10.5962/bhl.title.8537 [18] WIRTH W W, HOWARTH F G. The "Forcipomyia ingrami" complex in Hawaii (Diptera: Ceratopogonidae)[J]. Proceedings of the Hawaiian Entomological Society, 1982, 24(1): 127 − 151. [19] CHAN K L, LEROUX E J. Nine new species of Forcipomyia (Diptera: Ceratopogonidae) described in all stages[J]. The Canadian Entomologist, 1971, 103(5): 729 − 762. doi: 10.4039/Ent103729-5 [20] LIU J H, YAN G, LIU G P, et al. Forcipomyiinae of China (Diptera: Ceratopogonidae) Ⅱ. The genus Forcipomyia meigen[J]. Fauna of China, 2001, 3: 1 − 256. [21] 孟锦昕, 李楠, 何于雯, 等. 高飞铗蠓的形态学和分子生物学鉴定[J]. 中国国境卫生检疫杂志, 2022, 45(6): 447 − 449. doi: 10.16408/j.1004-9770.2022.06.004 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1226
- HTML全文浏览量: 681
- PDF下载量: 27
- 被引次数: 0
