


-
由于大气二氧化碳(CO2)浓度上升导致的全球变暖是人类面临的主要环境问题之一。土壤碳库作为陆地生态系统最大的碳库[1-2],每年由土壤呼吸作用(Soil respiration, Rs)向大气释放的碳是化石燃料燃烧碳释放量的十倍以上[3],所以土壤微小的变化都将显著改变土壤呼吸,进而影响大气二氧化碳浓度和土壤碳库储量,由此导致的气温升高又为加强土壤呼吸提供了一个潜在的积极反馈[4-5]。森林生态系统是全球陆地生态系统的重要组分,全球陆地生态系统中约40%土壤碳库存在于森林生态系统[6]。因此,在全球气候变化背景下研究森林的土壤呼吸碳排放具有重要意义。土壤呼吸主要包括植物根系结合根际微生物的自养呼吸作用和土壤微生物通过分解土壤有机质的异养呼吸作用[7-9],受植物群落、微生物生物量等生物因子和土壤环境、地形等非生物因子的影响[10]。地形是影响森林生态系统土壤呼吸空间变异的一个重要因子,可以通过改变土壤温度和水分[11]、土壤理化性质[12]、地下水再分配[13]、微生物生物量[14]、细根生物量[15]等,从而直接或间接地影响土壤呼吸。前人的研究已经注意到土壤呼吸的空间异质性仅仅用测量样本的平均值对等土壤呼吸值是不可靠的[16],并积极开展了地形对土壤呼吸空间异质性的影响研究。BERRYMAN等[17]对落基山脉半干旱及亚高山森林生态系统进行研究,认为地形属性通过影响土壤水分有效性成为土壤呼吸空间变异的重要预测因子;同样,RIVEROS-IREGUI等[18]在流域森林生态系统的研究表明地形景观通过改变土壤地下水文结构(分配/再分配)导致了土壤呼吸的空间变异。卿明亮等对贺兰山温带森林生态系统的研究表明,处于不同坡位的地形能够显著影响土壤理化性质,进而影响土壤呼吸。Q10值定义了土壤呼吸速率对温度变化10 ℃时的敏感程度,由于生态系统的类型及其地理地形的不同,陆地生态系统土壤呼吸的Q10值主要在1.3~5.6之间变化[19]。大部分的森林中往往存在着复杂的地形,因此,研究地形引起的空间变异对准确计算森林土壤呼吸具有重要意义。本研究以地处热带北缘的吊罗山低地森林为研究区,选择在景观结构上具有显著差异的3种地形(平地、坡地和洼地)为研究对象,通过对土壤呼吸和土壤温湿度的长期监测,分析吊罗山低地森林生态系统不同地形土壤呼吸的季节变化规律及其差异,研究月变化尺度上不同地形对水热因子的响应,旨在为准确评估该区域土壤CO2排放提供基础科学依据。
-
研究区位于海南岛东南部的吊罗山国家森林公园内(DNNR,18°40′N,109°54′E,海拔255 m,图1),受东海洋性热带季风气候的影响,年降雨量高达2 160 mm(台风带来大量降雨);干湿季节划分明显,80%以上的降雨集中在每年5~10月的雨季;每年11月至翌年4月为旱季;年均温24.6 ℃,最热月均温在7月份,为28.4 ℃,最冷月均温在1月份,为16.3 ℃[20]。吊罗山自下而上分布有典型的地带性森林植被,本研究区域可归为热带低地雨林[21]。森林的平均冠层高度为20~25 m(可能是受台风的影响),比处于同一纬度地带的其他内陆热带森林的冠层高度都要低。在建立保护区之前这片森林曾被大面积择伐,主要常见树种为青梅(Vatica mangachapoi)、琼榄(Gonocaryum lobbianum)、九节(Psychotria rubra)、钝叶新木姜子(Neolitsea obtusifolia)、光叶巴豆(Croton laevigatus)、子凌蒲桃(Syzygium tephrodes)、乌心楠(Phoebe tavoyana)等,物种丰富度低于其他热带雨林。土壤类型以由花岗岩或火成岩发育形成的湿润、呈酸性的红壤为主。土壤剖面清晰,平均土层深度大于2 m[22]。其中,表层土壤有机碳(SOC)含量为(9.84 ± 1.97)g·kg−1,总氮(TN)含量为(0.91 ± 0.18)g·kg−1,pH值为5.65 ± 0.17。
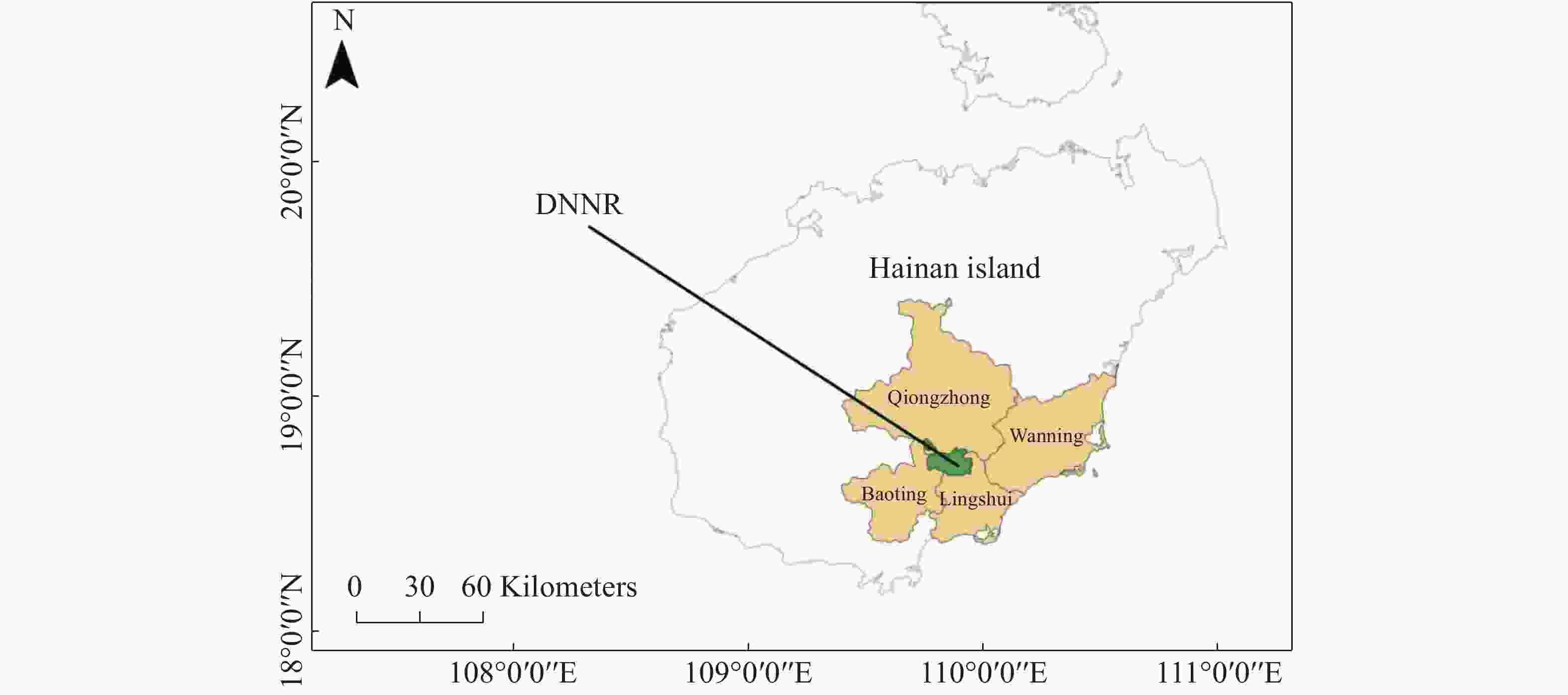
图 1 研究区地理位置
Figure 1. The geographic location of the study site
-
在低地雨林内选取平地、坡地和洼地3种地形,分别设置1块20 m×10 m的样地,其中平地的坡度小于2°,坡地的坡度约为45°,洼地为靠近森林水源地的低洼地。每个地形的样地内划分出8个5 m×5 m的小样方,选取两两相对的4个小样方为重复,分别在样方内沿着对角线布置1~3个土壤呼吸环,呼吸环由内径20 cm、高10 cm的PVC圆环制成,垂直嵌入土壤中3~5 cm,并去除土环内的植物,以避免测定的土壤呼吸中包含植物的呼吸作用,在整个观测过程中保持土环位置不动,保证原位观测。2016年9月至2018年9月,每月的中下旬,采用超便携式温室气体分析仪(UGGA 951-0011, LGR Inc.,USA)和PS-3000土壤气体通量测量系统测定所有位点土壤呼吸通量,其中UGGA测定CO2浓度的频率为1 Hz,浓度值在PS-3000程序中进行滑动平均计算得出土壤CO2的通量值。采用Decagon 5 TM传感器(Decagon Devices Inc.,USA)同步监测土壤5 cm处温度和湿度(体积含水量)。每次的测量时间为9:00~12:00,测定前对分析仪进行足够长的预热(至少5 min),每个土壤环测定每次至少3个重复。
-
采用Microsoft Excel软件进行数据预处理;利用SPSS软件对土壤呼吸速率和温度与水分数据进行相关性分析(皮尔逊相关分析),不同地形及月份的差异显著性采用重复测量方差分析检验,显著性水平设置为α=0.05;采用Origin软件进行绘图。采用指数方程Rs=aebT 拟合不同地形土壤呼吸速率对土壤温度的响应关系;通过方程拟合得到的温度敏感系数b代入方程 Q10=e10b,计算得到温度敏感性指数Q10;用一元线性回归方程Rs=aW+b 表示土壤呼吸速率和土壤湿度之间的回归关系;土壤呼吸速率和土壤温度、土壤湿度的双变量模型采用回归方程Rs=aebTWc 描述(其中Rs为土壤呼吸速率,T为土壤温度,W为土壤湿度,a,b,c均为方程参数)。土壤呼吸年积累量采用插值法计算,即计算相邻两次监测期间每天的平均土壤呼吸积累量,再将每天的积累量累加即可求得土壤呼吸年积累量。
-
2016年9月至2018年9月,笔者在低地雨林内监测了平地、坡地和洼地3种地形的土壤呼吸速率、土壤温度和土壤湿度的月动态变化。结果(图2)表明,在整个观测期间,平地和坡地的土壤呼吸速率均表现出“雨季高、旱季低”的动态变化,雨季土壤呼吸速率升高,6~8月出现排放峰值,接近旱季开始降低;洼地则出现“雨季低、旱季高”的动态变化,进入旱季后土壤呼吸速率升高,3~5月出现排放峰值,进入雨季后降低。土壤呼吸速率的月均值表现为洼地 > 坡地 > 平地;其月均值与范围大小分别为[(2.54 ± 0.85),(1.38~4.58)],[(2.50 ± 0.84),(1.30~3.86)],[(2.16 ± 0.67),(0.88~3.55)] μmol·m−2·s−1,洼地土壤呼吸速率分别高出平地和坡地的17.59%和1.6%,且洼地和平地的土壤呼吸速率月均值之间具有显著差异(P<0.05)。
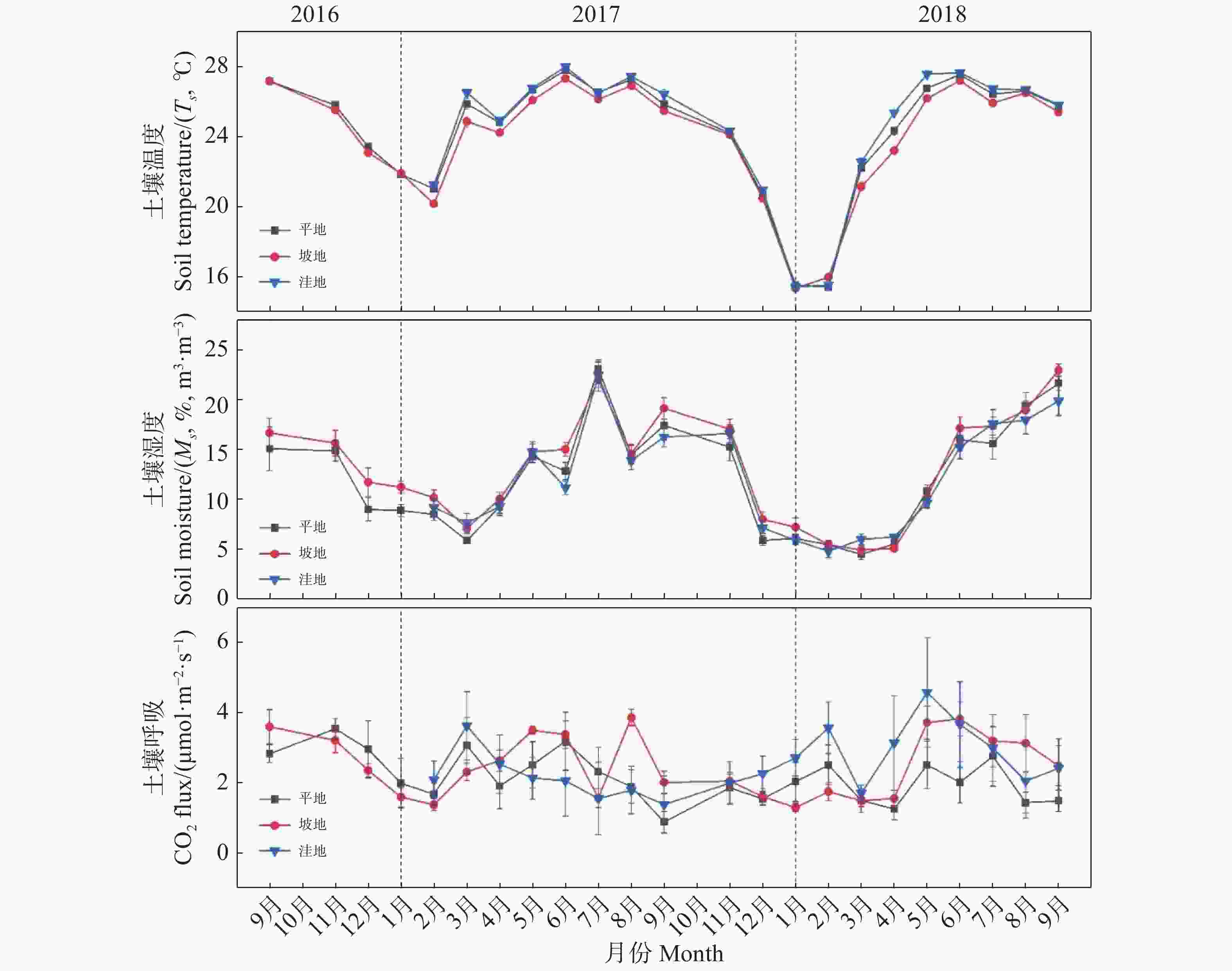
图 2 不同地形土壤呼吸速率、土壤温度和湿度月动态变化
Figure 2. Monthly variation of soil respiration rate, soil temperature and moisture in different terrains
-
在整个观测期间,3种地形的土壤温度及土壤湿度均呈现出相似的单峰型月动态变化,雨季的土壤温度和土壤湿度均明显高于旱季(图2)。旱季土壤温度和湿度同时降低,在2~3月份之间分别达到最小值;接近雨季开始升高,在6~7月份达到最大值。其中土壤温度月均值分别为洼地(24.54 ± 3.80)℃ > 平地(24.32 ± 3.49)℃ > 坡地(23.93 ± 3.38)℃,且土壤温度在各地形间没有显著差异(P>0.05);土壤湿度月均值分别为坡地[(13.15 ± 5.40)%]> 洼地[(12.19 ± 5.37)%]> 平地[(12.16 ± 5.48)%],且各地形的土壤湿度没有显著差异(P>0.05)。总的来看,5 cm处土壤温度与土壤湿度显著相关(P<0.01),高温和高湿在雨季期间同时出现,雨季土壤平均温度高于旱季,符合典型的热带季风性气候特征。
-
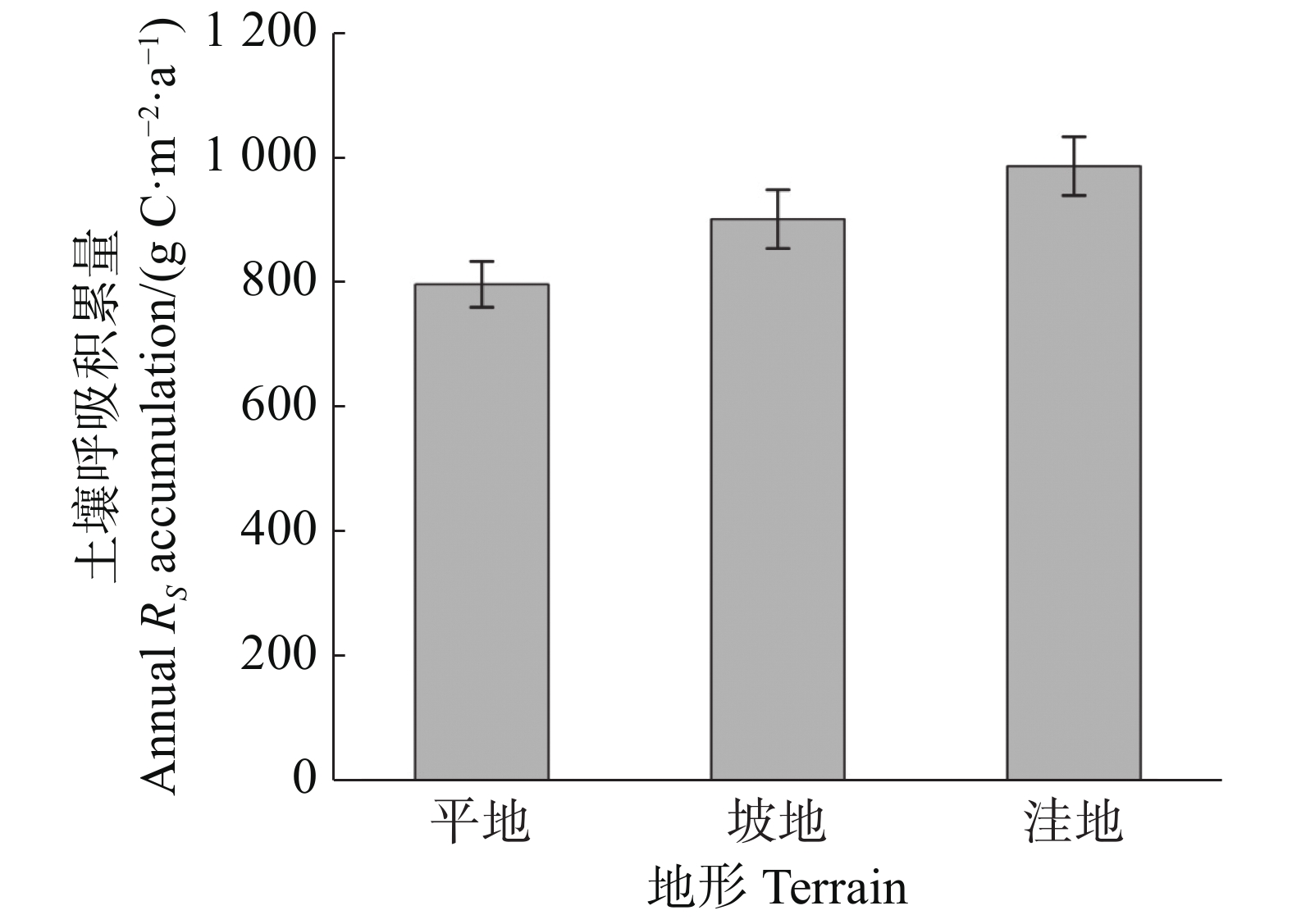
通过插值法计算出平地、坡地和洼地3种不同地形的土壤呼吸年积累量。结果(图3)表明,不同地形的土壤呼吸年积累量具有明显差异,表现为洼地(986.37 ± 46.91) g C·m−2·a−1 > 坡地(901.46 ± 47.00) g C·m−2·a−1 > 平地(796.85 ± 36.86) g C·m−2·a−1,平地的土壤呼吸年积累量分别比坡地和洼地低13.13%和23.78%,且统计分析显示各地形间的土壤呼吸年积累量没有显著差异(P>0.05),说明地形可能不是影响土壤呼吸年积累量的重要因素。
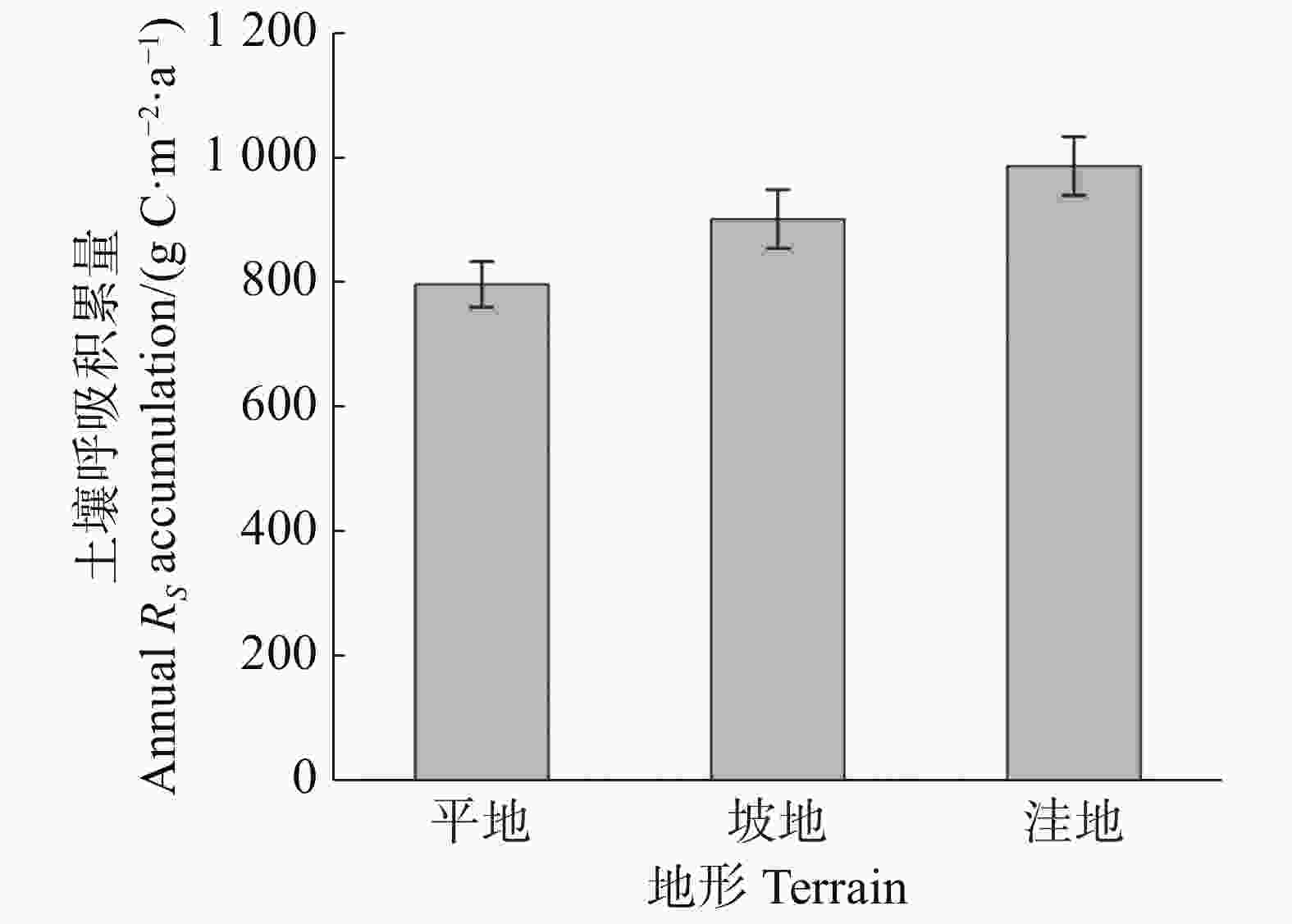
图 3 不同地形土壤呼吸年积累量
Figure 3. Annual accumulation of soil respiration in different terrains
-
土壤温度是影响土壤呼吸速率变化的最重要的因子之一,因此常常被用来解释土壤呼吸速率的大部分变化。以往的研究中常用指数模型来描述土壤呼吸速率与土壤温度之间的关系。回归分析结果(表1)表明,本研究中坡地土壤呼吸月变化与5 cm处土壤温度存在显著正相关关系(P<0.01),而平地和洼地的土壤呼吸月变化与5 cm处土壤温度的相关关系都没有达到显著水平(P>0.05);土壤温度能够解释坡地土壤呼吸月变化的61.0%,通过指数方程计算出Q10值为3.19;而对平地和洼地土壤呼吸的解释率极低,月变化对温度不敏感。因此,本研究中3种地形土壤呼吸月变化对土壤温度的响应并不一致,土壤温度可作为坡地土壤呼吸的限制因子。土壤呼吸和土壤湿度的回归分析结果(表1)表明,坡地和洼地的土壤呼吸月变化与0~5 cm处土壤湿度分别存在显著正相关(P<0.01)和显著负相关(P<0.01)关系,而平地的土壤呼吸月变化与0~5 cm处土壤湿度的相关关系没有达到显著水平(P>0.05);土壤湿度对土壤呼吸的解释率仅为5.2%~16.6%。土壤温度和土壤湿度双变量模型的拟合程度较单变量模型更低,说明土壤呼吸的季节变化与环境因子的关系更为复杂,简单的经验模型无法很好得描述土壤温度和土壤湿度对土壤呼吸的共同作用。
表 1 土壤温度(T)、湿度(W)和呼吸速率(RS)的回归模型与Q10值
Table 1. Regression models of soil respiration rate (RS) using soil temperature (T) and soil moisture (W)as predictor variables, and Q10 values of three terrains
地形 Terrain Rs=aebT Rs=aW+b Rs=aebTWc Q10 a b R2 a b R2 a b c R2 平地 Flat terrain 3.429 −0.025 0.022 −0.044 2.441 0.052 1.342 0.038 −0.205 0.017 0.78 坡地 Sloping terrain 0.145 0.116 0.610** 0.069 1.603 0.166** 0.153 0.125 −0.104 0.244** 3.19 洼地 Low-lying terrain 2.773 −0.004 0.003 −0.076 3.471 0.064** 3.330 0.051 −0.641 0.078 0.96 注:**表示在0.01水平上显著相关(P<0.01)。
Note: ** means significant correlation at the level of 0.01 (P<0.01). -
地形是影响土壤呼吸空间变异的一个重要因子,不同的地形土壤呼吸存在差异[23]。本研究中3种地形的土壤温度和土壤湿度存在相似的单峰型月动态变化,但各地形间没有显著差异(P>0.05)。土壤呼吸月变化均表现出明显的季节规律,平地和坡地的土壤呼吸速率均表现出“雨季高、旱季低”的动态变化,洼地表现出“雨季低、旱季高”的动态变化,洼地和平地的土壤呼吸月变化存在显著差异(P<0.05),且洼地的土壤呼吸年累积量最高。在吊罗山热带森林中,受热带海洋性季风气候的影响,雨季表现出高温高湿的气候,良好的土壤水热条件给土壤微生物及植物根系创造了适宜的生命代谢环境,雨季在总体上往往可以观察比旱季更强的土壤呼吸作用[24-25];而洼地则表现出相反的季节动态,这在热带地区的土壤呼吸研究中是非常罕见的。通常情况下,土壤温度和土壤湿度是影响土壤CO2通量的主导因素[26-27],土壤呼吸随着土壤温度和湿度的升高而升高,而土壤水分的持续增加会导致土壤通透性变差[28-29]。氧气(O2)是土壤微生物和植物根系进行有氧呼吸的必要条件,过高的土壤水分会限制O2的扩散,导致土壤物理结构出现厌氧状态,植物根系和好氧微生物的生命活动受到抑制,土壤的有机碳分解速率变低,CO2排放量减少,所以当土壤湿度超过阈值后,土壤呼吸会受到抑制[30-31]。中等湿度的土壤条件可能更适合土壤呼吸作用,有氧呼吸速率达到峰值[32-34],因此,洼地地形在雨季可能由于土壤水分过多而出现下降的趋势。
本研究中不同地形的土壤呼吸与0~5 cm处土壤温度和湿度的相关关系并不一致。坡地的土壤呼吸与土壤温湿度的相关关系均达到了显著水平(P<0.01),经验模型的解释率达到了61.0%,Q10值为3.19;洼地只与土壤湿度有负相关关系(P<0.01),解释率最高只有7.8%;而平地与土壤温湿度均无相关关系,回归模型的解释率极低。因此,不同地形间的土壤呼吸对土壤温度和水分的响应存在较大差异,坡地地形的土壤呼吸模式与多数研究一致[35-36],而平地与洼地的土壤呼吸模式及主导因素可能与土壤环境的其他因子有关。不同地形如何对土壤呼吸产生影响,尚有待对土壤环境和结构成分进行更详细的分析研究。
本研究证明了在吊罗山低地森林中,地形对土壤呼吸的空间变异有显著影响,所以当要把土壤二氧化碳通量外推到更大尺度时,地形的复杂性则是必须要考虑的因素之一。在土壤呼吸的影响因素上,不同地形间可能受到复杂土壤环境的影响,导致土壤呼吸与水热因子的关系不明显,因此,需要更深入的后续研究来阐明不同地形土壤环境对土壤呼吸作用的影响。同时,本研究选择了不同地形研究土壤呼吸,为准确评估该区域土壤CO2排放提供了基础科学依据。
Soil Respiration of Different Terrains in a Tropical Lowland Rainforest in Diaoluoshan, Hainan, China
-
摘要: 以地处热带北缘的吊罗山低地森林为研究区,选择在景观结构上具有显著差异的3种地形(分别为平地、坡地和洼地)为研究对象,利用LGR便携式温室气体分析仪及土壤气体通量监测系统,测定吊罗山热带低地森林不同地形土壤呼吸速率和土壤温湿度,研究不同森林地形土壤呼吸的季节变化规律及其差异,并探讨土壤呼吸与水热因子的关系。结果表明:(1)平地和坡地的土壤呼吸速率均表现出“雨季高、旱季低”的动态变化,雨季6~8月出现排放峰值;洼地则出现“雨季低、旱季高”的动态变化,进入旱季土壤呼吸速率升高,3~5月出现排放峰值,雨季开始降低;(2)不同地形的土壤呼吸年积累量大小具有明显差异,表现为洼地[(986.37 ± 46.91) g C·m−2·a−1]> 坡地[(901.46 ± 47.00) g C·m−2·a−1] > 平地[796.85 ± 36.86) g C·m−2·a−1];(3)不同地形土壤呼吸与土壤温湿度的关系并不一致,其中坡地的相关关系最显著(P< 0.01),Q10值为3.19。综上所述,吊罗山低地森林中不同地形土壤呼吸存在显著差异,当把土壤二氧化碳通量外推到更大尺度时,必须要考虑地形的复杂性。Abstract: Soil respiration is a frontier topic in the carbon cycle of terrestrial ecosystems. The seasonal variations and differences of soil respiration in different terrains were analyzed by measuring the soil respiration rate and soil temperature and moisture in a tropical lowland forest in Diaoluoshan, Hainan, China, and the relationship between the soil respiration and the hydrothermal factors was also analyzed. The results showed that the soil respiration rate in the flat terrain and sloping terrain had a dynamic change of "higher in rainy season and lower in dry season", with peak emissions from June to August. On the contrary, the soil respiration rate in the low-lying terrain had a dynamic change of "lower in rainy season and higher in dry season", with the peak emissions from March to May. The annual accumulation of soil respiration in different terrains varied significantly, which was in the order of the low-lying terrain (986.37 ± 46.91) g C· m−2·a−1 > the sloping terrain (901.46 ± 47.00) g C·m−2·a−1 > the flat terrain (796.85 ± 36.86) g C·m−2·a−1. The soil respiration in different terrains was not consistently correlated with the soil temperature and moisture, but the soil respiration in the sloping terrain was most significantly correlated with the soil temperature and moisture (P< 0.01), with a Q10 value of 3.19. These results indicated that there were significant differences in soil respiration in different terrains in a tropical lowland forest in Diaoluoshan, Hainan. When the soil CO2 flux is extrapolated to a larger scale, the complexity of the terrain should be taken into account.
-
图 2 不同地形土壤呼吸速率、土壤温度和湿度月动态变化
Fig. 2 Monthly variation of soil respiration rate, soil temperature and moisture in different terrains
表 1 土壤温度(T)、湿度(W)和呼吸速率(RS)的回归模型与Q10值
Table 1 Regression models of soil respiration rate (RS) using soil temperature (T) and soil moisture (W)as predictor variables, and Q10 values of three terrains
地形 Terrain Rs=aebT Rs=aW+b Rs=aebTWc Q10 a b R2 a b R2 a b c R2 平地 Flat terrain 3.429 −0.025 0.022 −0.044 2.441 0.052 1.342 0.038 −0.205 0.017 0.78 坡地 Sloping terrain 0.145 0.116 0.610** 0.069 1.603 0.166** 0.153 0.125 −0.104 0.244** 3.19 洼地 Low-lying terrain 2.773 −0.004 0.003 −0.076 3.471 0.064** 3.330 0.051 −0.641 0.078 0.96 注:**表示在0.01水平上显著相关(P<0.01)。
Note: ** means significant correlation at the level of 0.01 (P<0.01). 下载: 导出CSV
下载: 导出CSV
-
[1] SCHIMEL D S. Terrestrial ecosystems and the carbon cycle [J]. Global change biology, 1995, 1(1): 77 − 91. doi: 10.1111/j.1365-2486.1995.tb00008.x [2] 许文强, 陈曦, 罗格平, 等. 土壤碳循环研究进展及干旱区土壤碳循环研究展望[J]. 干旱区地理, 2011, 34(4): 614 − 620. [3] RAICH J W, POTTER C S, BHAGAWATI D. Interannual variability in global soil respiration, 1980–94 [J]. Global change biology, 2002, 8(8): 800 − 812. doi: 10.1046/j.1365-2486.2002.00511.x [4] SCHLESINGER W H, ANDREWS J A. Soil respiration and the global carbon cycle [J]. Biogeochemistry, 2000, 48(1): 7 − 20. doi: 10.1023/A:1006247623877 [5] RODEGHIERO M, CESCATTI A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps [J]. Global change biology, 2005, 11(7): 1024 − 1041. doi: 10.1111/j.1365-2486.2005.00963.x [6] PRATHER M, EHHALT D, DENTENER F, et al. Atmospheric chemistry and greenhouse gases[C]// Related Information: Climate Change: Working Group I: the Scientific Basis, 2001. [7] HANSON P, EDWARDS N, GARTEN C T, et al. Separating root and soil microbial contributions to soil respiration: a review of methods and observations [J]. Biogeochemistry, 2000, 48(1): 115 − 146. doi: 10.1023/A:1006244819642 [8] HÖGBERG P, READ D J. Towards a more plant physiological perspective on soil ecology [J]. Trends in ecology & evolution, 2006, 21(10): 548 − 554. [9] RYAN M G, LAW B E. Interpreting, measuring, and modeling soil respiration [J]. Biogeochemistry, 2005, 73(1): 3 − 27. doi: 10.1007/s10533-004-5167-7 [10] DE DATO G, LAGOMARSINO A, LELLEI-KOVACS E, et al. The response of soil CO2 efflux to water limitation is not merely a climatic issue: The role of substrate availability [J]. Forests, 2017, 8(7): 241. doi: 10.3390/f8070241 [11] ZHAO Y-L, GOLDBERG S D, XU J-C, et al. Spatial and seasonal variation in soil respiration along a slope in a rubber plantation and a natural forest in Xishuangbanna, Southwest China [J]. Journal of Mountain Science, 2018, 15(4): 695 − 707. doi: 10.1007/s11629-017-4478-9 [12] BURKE I C, LAUENROTH W K, RIGGLE R, et al. Spatial variability of soil properties in the shortgrass steppe: the relative importance of topography, grazing, microsite, and plant species in controlling spatial patterns [J]. Ecosystems, 1999, 2(5): 422 − 438. doi: 10.1007/s100219900091 [13] LUO Y, ZHOU X H. Soil respiration and the environment[M]. San Diego, CA: Academic Press/Elsevier, 2006: 257-305. [14] 成毅, 安韶山, 马云飞. 宁南山区不同坡位土壤微生物生物量和酶活性的分布特征[J]. 水土保持研究, 2010, 17(5): 148 − 153. [15] 黄诚诚, 王迎春, 张渐飞, 等. 东北黑土典型坡耕地土壤呼吸特征的研究[J]. 中国生态农业学报, 2018, 26(1): 1 − 7. [16] 王晓荣, 胡文杰, 潘磊, 等. 森林生态系统土壤呼吸空间异质性研究进展[J]. 西部林业科学, 2017, 46(3): 177 − 186. [17] BERRYMAN E M., BARNARD H R. ADAMS H R, et al Complex terrain alters temperature and moisture limitations of forest soil respiration across a semiarid to subalpine gradient [J]. Journal of Geophysical Research: Biogeosciences, 2015, 120(4): 707 − 723. doi: 10.1002/2014JG002802 [18] RIVEROS-IREGUI D A, MCGLYNN B L, EMANUEL R E, et al. Complex terrain leads to bidirectional responses of soil respiration to inter-annual water availability [J]. Glob Chang Biol, 2012, 18(2): 749 − 756. doi: 10.1111/j.1365-2486.2011.02556.x [19] TANG J, BALDOCCHI D D. Spatial–temporal variation in soil respiration in an oak–grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components [J]. Biogeochemistry, 2005, 73(1): 183 − 207. doi: 10.1007/s10533-004-5889-6 [20] ZHAO J, HE C, QI C, et al. Biomass increment and mortality losses in tropical secondary forests of Hainan, China [J]. Journal of Forestry Research, 2019, 30(2): 647 − 655. doi: 10.1007/s11676-018-0624-7 [21] 杨小波, 林英, 梁淑群. 海南岛五指山的森林植被. 五指山的森林植被类型[J]. 海南大学学报(自然科学版), 1994(3): 220 − 236. [22] YUJIA H. The Dipterocarp forest of Hainan Island, China [J]. Journal of Tropical Forest Science (Malaysia), 1997, 9(4): 477 − 498. [23] HANSON P, WULLSCHLEGER S, BOHLMAN S, et al. Seasonal and topographic patterns of forest floor CO2 efflux from an upland oak forest [J]. Tree Physiology, 1993, 13(1): 1 − 15. doi: 10.1093/treephys/13.1.1 [24] 蔡子良, 邱世平. 西双版纳热带季节雨林土壤呼吸季节动态及驱动因素[J]. 生态环境学报, 2019, 28(2): 283 − 290. [25] JIANG L, MA S, ZHOU Z, et al. Soil respiration and its partitioning in different components in tropical primary and secondary mountain rain forests in Hainan Island, China [J]. Journal of Plant Ecology, 2016, 10(5): 791 − 799. [26] 田琴. 黄土丘陵区典型植被类型土壤微生物及异养呼吸特征[D]. 北京: 中国科学院教育部水土保持与生态环境研究中心, 2017. [27] 禹朴家, 徐海量, 王炜, 等. 沙丘不同部位土壤呼吸对人工降水的响应[J]. 中国沙漠, 2012, 32(2): 437 − 441. [28] 杨开军, 熊莉, 杨万勤, 等. 林窗对川西亚高山云杉人工林土壤呼吸的影响[J]. 应用与环境生物学报, 2016, 22(3): 505 − 509. [29] 陈书涛, 邹建文, 胡正华. 陆地生态系统土壤呼吸的观测与模拟[J]. 生态环境学报, 2017, 26(11): 1985 − 1996. [30] 夏秀雪, 高国强, 王东男, 等. 林下植被去除对水曲柳人工林土壤呼吸、微生物和根系动态的短期影响[J]. 生态学杂志, 2017, 36(5): 1198 − 1207. [31] 付刚, 钟志明. 实验增温对西藏高原玉米田土壤呼吸的影响[J]. 生态环境学报, 2017, 26(1): 49 − 54. [32] LINN D M., DORAN J W Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and non-tilled soils 1 [J]. Soil Science Society of America Journal, 1984, 48(6): 1267 − 1272. doi: 10.2136/sssaj1984.03615995004800060013x [33] SKOPP J, JAWSON M, DORAN J. Steady-state aerobic microbial activity as a function of soil water content [J]. Soil Science Society of America Journal, 1990, 54(6): 1619 − 1625. doi: 10.2136/sssaj1990.03615995005400060018x [34] DAVIDSON E A, SAMANTA S, CARAMORI S S, et al. The Dual Arrhenius and Michaelis–M enten kinetics model for decomposition of soil organic matter at hourly to seasonal time scales [J]. Global Change Biology, 2012, 18(1): 371 − 384. doi: 10.1111/j.1365-2486.2011.02546.x [35] 王家骏, 王传宽, 韩轶. 帽儿山不同年龄森林土壤呼吸速率的影响因子[J]. 生态学报, 2018, 38(4): 1194 − 1202. [36] 刘博奇, 牟长城, 邢亚娟, 等. 小兴安岭典型温带森林土壤呼吸对强降雨的响应[J]. 北京林业大学学报, 2016, 38(4): 77 − 85. -

 点击查看大图
点击查看大图

图(3) / 表 (1)
计量
- 文章访问数: 221
- HTML全文浏览量: 127
- PDF下载量: 56
- 被引次数: 0
