

 下载:
下载:
-
嫁接是一种广泛应用于植物无性繁殖与性状改良的技术,即将接穗(嫩枝)切割后嫁接到砧木(茎或根)上。嫁接后,植物组织经历快速愈合,通过细胞分裂、扩展与分化,形成接穗与砧木间功能性维管连接[1]。嫁接成功与否受植物亲缘关系、组织年龄与类型等因素影响[2]。目前,该技术已广泛应用于农业、园艺及生物学研究[3],嫁接后的植物通常表现出更强的病虫害抗性、逆境适应能力[4]、童期缩短[5]及果实品质提升[6]等优良性状。嫁接在农业与林业中的重要性推动了对其愈合过程中生理、生化和遗传机制的深入研究。嫁接时期不同可能导致亲和性差异。嫁接不亲和性可分为局部不亲和、易位不亲和及病原体介导的不亲和三类[7]。其中,局部不亲和源于形成层与维管组织连续性中断,导致嫁接界面断裂[8]。亲和性嫁接过程包括4个阶段:损伤反应与细胞壁修饰、界面细胞分裂变化、胞间连丝形成与功能建立,以及维管连接重建[9-10]。在此过程中,植物组织的整合不仅受外界环境影响,还受基因表达变化调控[11]。
本氏烟草(Nicotiana benthamiana)是茄科模式植物,原产于澳大利亚北部热带地区,因其基因组信息完备,常被用于嫁接研究[12-13]。通过将其与多种植物进行科间嫁接实验发现,本氏烟草产生的β−1,4−葡聚糖酶可促进细胞间黏连,提高嫁接成活率[14],因此,本氏烟草是开展种间或科间嫁接研究的理想材料。
酚类物质在嫁接愈合中具有复杂功能,既是新组织构建的关键原料,也参与创伤防御,但其过量积累可能抑制愈合[15]。酚类易被过氧化物酶氧化为醌类,阻碍生长素运输,导致砧木与接穗连接受阻[16]。同时,酚类作为木质素合成前体,参与导管与筛管分化,通过木质素沉积增强维管组织机械强度。研究表明,亲和性高的嫁接组合中,嫁接口酚类含量及苯丙氨酸解氨酶活性通常较高[17],提示酚类代谢与嫁接成功率密切相关。此外,嫁接造成的伤口可激活植物防御系统,酚类通过抗氧化作用清除活性氧,减轻氧化损伤,为维管组织分化创造条件。糖类在嫁接愈合中不仅作为能量底物参与呼吸代谢,还参与信号转导及生物大分子合成[18]。糖类通过调控碳水化合物代谢及相关酶活性,影响植株生长发育,从而介导嫁接体的形成与维管重建[19]。外源蔗糖处理可提高接穗伤流液中蔗糖含量,缩小砧木与接穗间的糖分差异,提升嫁接成活率[20]。在以本氏烟草为材料的嫁接愈合过程中,酚类与糖类的动态变化及其对维管重建的调控机制仍有待深入解析。为此,本研究以本氏烟草自体嫁接为模型,系统分析嫁接后不同时间点酚类物质含量、关键酶活性及糖类代谢的动态变化,并结合维管组织解剖学观察,旨在揭示酚类与糖类在嫁接愈合中的协同作用机制,为优化嫁接技术、提高嫁接成活率提供理论依据。
-
供试的植物材料为本氏烟草(Nicotiana benthamiana)
-
将泥炭土、珍珠岩、蛭石按体积比2︰1︰1混合均匀,以兼顾基质的透气性与保肥性,防止幼苗烂根。将上述基质装入耐高温容器,采用121℃高压蒸汽灭菌20 min,以彻底杀灭基质中的杂菌、虫卵及杂草种子。
-
本氏烟草种子的消毒流程如下:先用体积分数75%的乙醇表面处理30 s,再用体积分数5%的次氯酸钠(NaClO)溶液浸泡5 min,最后用无菌水冲洗3次。消毒后的种子播种于含质量分数3%蔗糖和1%琼脂的1/2 MS培养基上,培养基pH调至5.8。培养条件为25℃恒温,光周期为L光照︰D黑暗=16 h︰8 h。幼苗在培养基上生长7天后进行移栽。
-
用含70%乙醇的酒精棉将工作台擦干净,将刀片和镊子浸泡在含无水乙醇的烧杯中。选取7~9 d生长健康,胚轴壮硕垂直的植株作为嫁接材料。砧木选取茎秆直立、的健壮苗。接穗从健康母株上,剪取生长健壮、处于半木质化状态的新梢。从无水乙醇中取出嫁接刀,待乙醇吹干后,将选取好植株放置。对于接穗部分,首先去除烟草植株的多余子叶,然后沿整株植株胚轴的一半处切下来,将根和子叶取出来放到旁边,余下的部分留在培养皿里面作为接穗。而砧木部分,将本生烟草整株植株胚轴的一半处切下来,留下根部分为砧木,子叶那一部分取出来放在旁边。将接穗长斜面向内插入砧木切口,确保一侧形成层对齐,用嫁接夹固定接口。嫁接7 d后,用镊子将接穗轻轻提起,能够提起完整植株,说明嫁接体愈合成功。

图 1 烟草嫁接流程图
Figure 1. Flow chart of tobacco grafting
-
通过羧基荧光素二乙酸盐(Carboxyfluorescein diacetate,CFDA)在嫁接界面的运输情况进行检测。
木质部检测:在嫁接植株的根部下胚轴连接处下方2 cm切断茎段,将5 µL的CFDA(1 mmol·L−1)溶液滴加于切口表面,并用锡箔包裹嫁接部位以避免光照干扰。处理7 h后,使用荧光显微镜(LEICA/ DMi8,ICA)观察嫁接界面上方约1 cm处茎段维管束中的荧光信号,以判断木质部是否建立有效连通。
韧皮部检测:在在嫁接界面上方约2 cm处将茎水平切断,将5 µL的CFDA(1 mmol·L−1)溶液滴加于横截面,并用锡箔遮盖嫁接区域。处理7 h后,使用荧光显微镜观察嫁接界面下方约1 cm处维管束中的荧光信号,以评估韧皮部的连通情况。
CFDA处理后,以嫁接界面为中心截取约1 cm茎段,同时取切口上方或下方约2 cm处的组织。样品使用OCT冷冻包埋剂包埋固定,随后采用冷冻切片机制备纵切和横切切片(厚度约100 µm)。其中,纵切切片需确保清晰暴露维管束结构,以便观察荧光信号在木质部和韧皮部中的分布,从而评估嫁接界面运输通道的重建情况。
-
基于基因的全称在TAIR拟南芥数据库(
http://www.arabidopsis.org )中进行同源检索,获取拟南芥中相关基因的编码序列。随后采用Pfam_scan工具对所得基因序列进行蛋白质结构域鉴定,明确其功能特征结构。基于鉴定获得的关键结构域信息,进一步使用HMMER软件包中的hmmsearch工具,在植物基因组蛋白数据库中开展全基因组范围内的同源蛋白搜索,设置参数保留默认E值阈值以控制结果可靠性。 -
从DDBJ数据库(
https://www.ddbj.nig.ac.jp )获取了登录号为DRA009936的嫁接相关转录组原始数据,用于分析目标基因在嫁接体系中的表达模式。原始数据首先使用SRA Toolkit中的prefetch命令下载,再通过fastq-dump工具解压为fastq格式文件。随后采用Salmon(v1.9.0)进行定量分析:首先基于各物种的参考转录组序列(cds.fa)构建索引,命令为salmon index -t cds.fa -i transcripts_index -k 31;然后进行表达量计算,命令为salmon quant -i transcripts_index -l A -1 paired_reads_1.fq -2 paired_reads_2.fq -o quant_results--validateMappings。最后,通过Python脚本整合各生物学重复的表达量(取均值),利用R语言的tidyverse包对数据进行标准化,并使用pheatmap包(v1.0.12)生成热图,实现可视化分析。 -
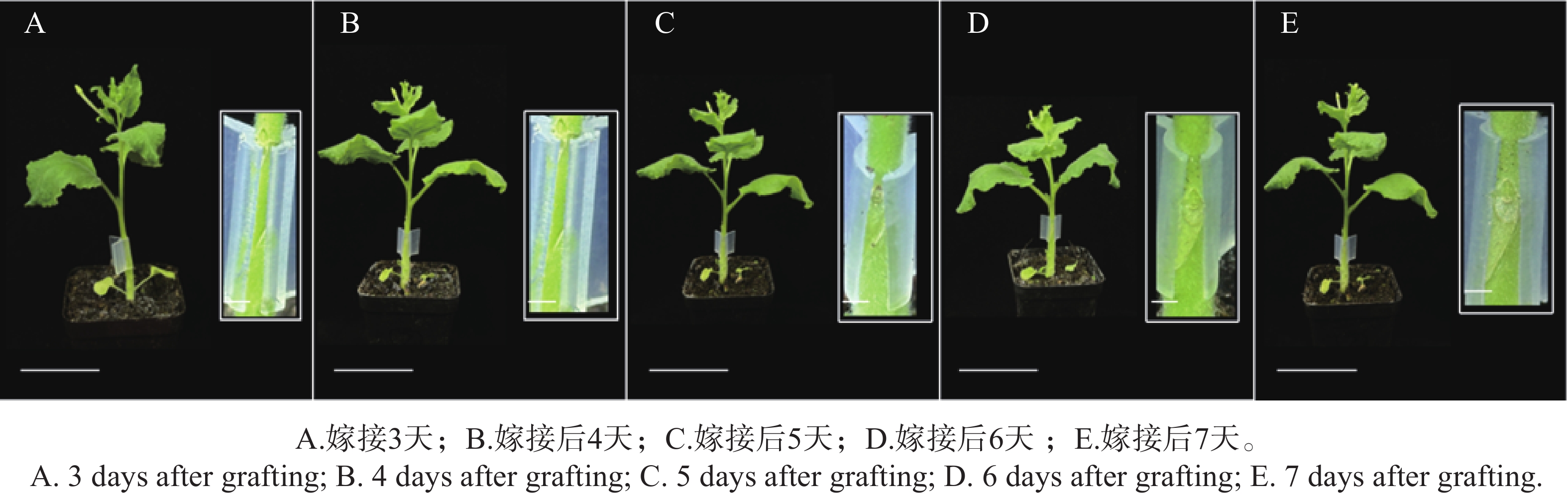
嫁接成功的标志是接穗与砧木牢固附着,且植株根系恢复正常生长。采用本研究建立的本氏烟草嫁接方法,发现嫁接后约7 d接穗与砧木即可实现稳定附着。嫁接后3~7 d的植株生长状态及嫁接口愈合过程如图2所示,A~E依次对应嫁接后第3~7天,嫁接后3~4 d植株保持正常生长,嫁接口切面新鲜,第5天,新生细胞填充嫁接口间隙,切面边界趋于模糊;第6天,愈伤组织明显增生,形成半透明连接桥;第7天,嫁接口被愈伤组织基本包裹,上下茎段连接较为牢固,植株无萎蔫、黄化等胁迫症状,表明嫁接已成功愈合。
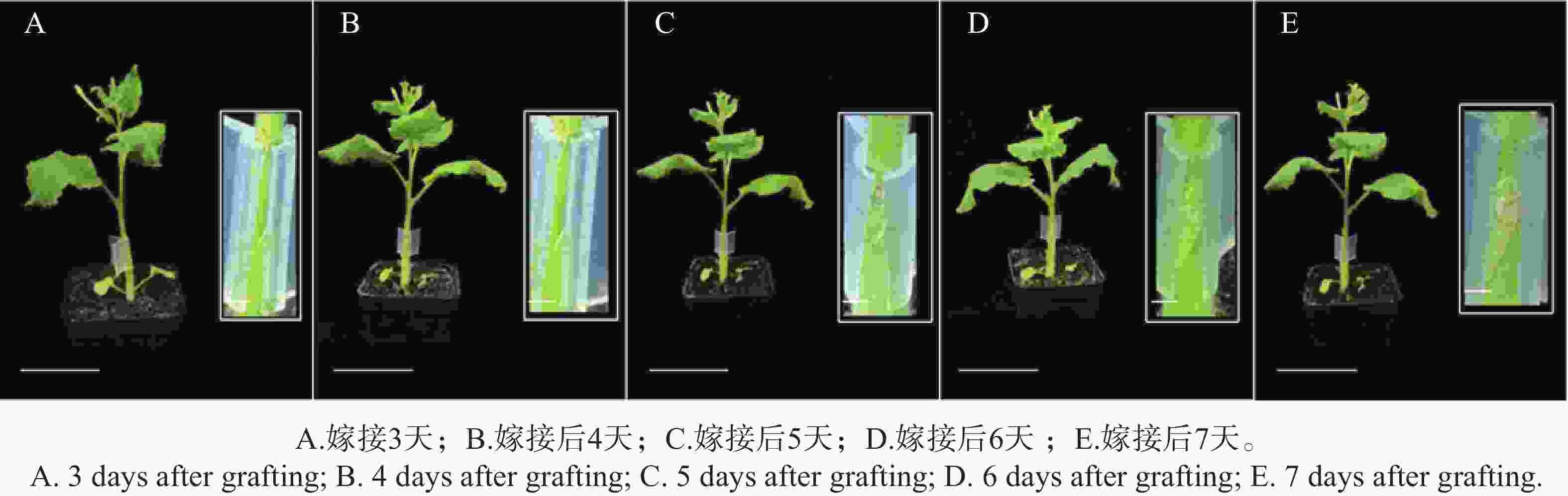
图 2 烟草自嫁接后不同时间生长状态
Figure 2. Growth status of tobacco plants at different time points after self-grafting
-
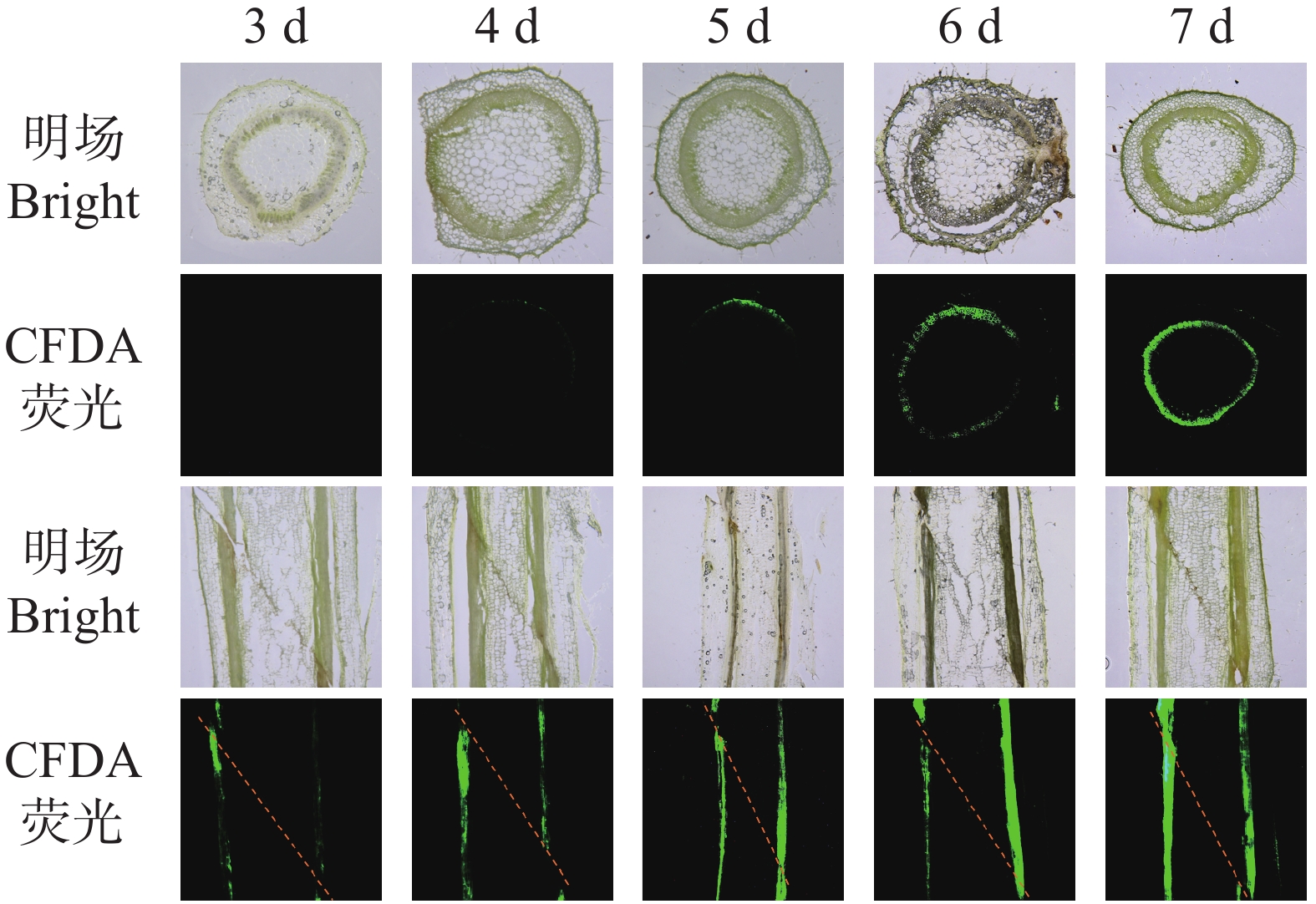
本研究通过CFDA荧光示踪技术,对烟草嫁接后木质部的重新连通过程进行了分析。从图3可知,嫁接后3 d,木质部横切面与纵切面均仅可见微弱或近乎缺失的荧光信号,表明有效输导通道尚未建立;第4天,横切面出现微弱散在的点状或断续条状信号,纵切面荧光呈断续长条形,说明木质部开始初步连接但连通性较差;第5天,横切面荧光呈现为不连续环状,纵切面出现连续但较弱的纵向信号,显示连通性显著提升;而在6~7 d后,横切面呈现连续且明亮的环状荧光,纵切面则为连续、明亮的长条状信号,表明木质部在横向与纵向上基本已实现稳定的连通。上述结果说明,木质部重新连通启动于嫁接后4~5 d,其输导功能在嫁接后6~7 d基本恢复至稳定水平。
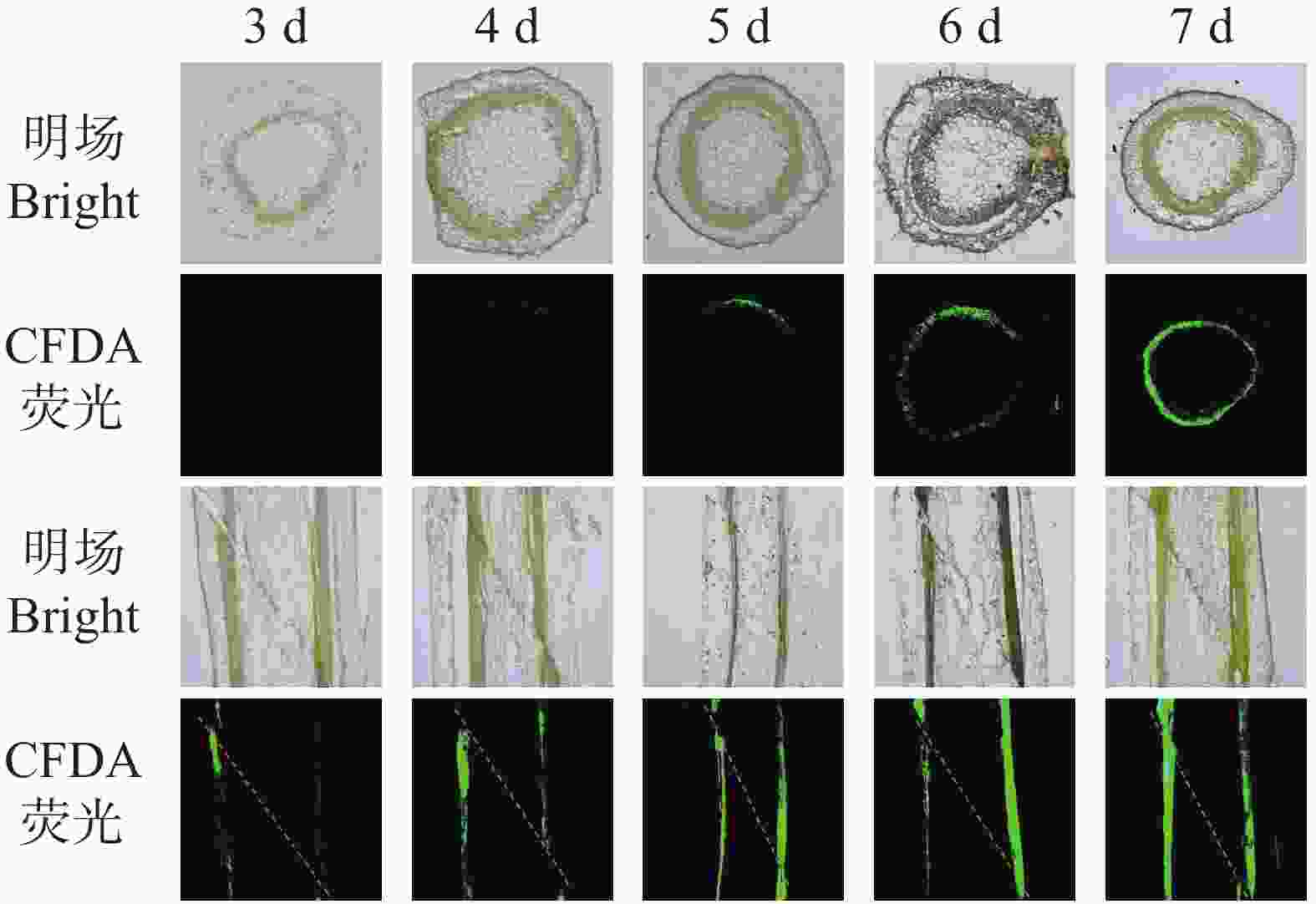
图 3 烟草嫁接中CFDA从砧木到接穗的运输过程。
Figure 3. Transport of CFDA from the rootstock to the scion in tobacco grafts
-
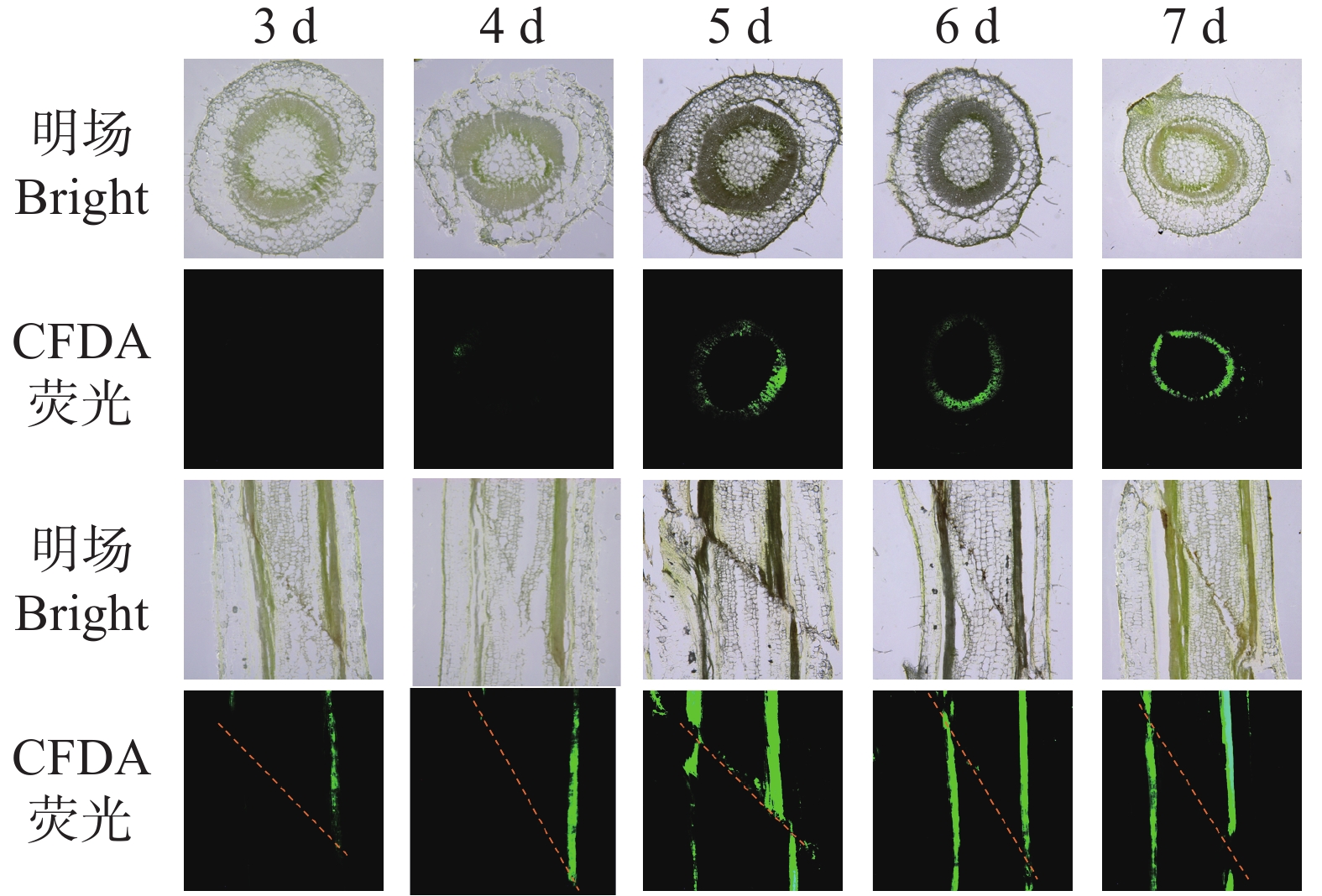
本研究进一步通过CFDA荧光示踪技术,分析了烟草嫁接后韧皮部的重新连通过程。结果显示(图4),嫁接后3 d,横切面仅见零星光点状荧光,纵切面仅呈局部短线条信号,表明筛管尚未建立有效运输通道;第4天,横切面出现微弱散在荧光,纵切面为断续长条形信号,说明筛管开始初步对接但连通仍不完整;第5天,横切面荧光呈现连续环状,纵切面出现连续长条状信号局部断续,显示韧皮部连通性显著提升;嫁接后6~7 d,横切面环状荧光持续增强且均匀,纵切面信号连续、明亮且进一步增强,表明韧皮部在横向与纵向上已实现高效、稳定的连通。
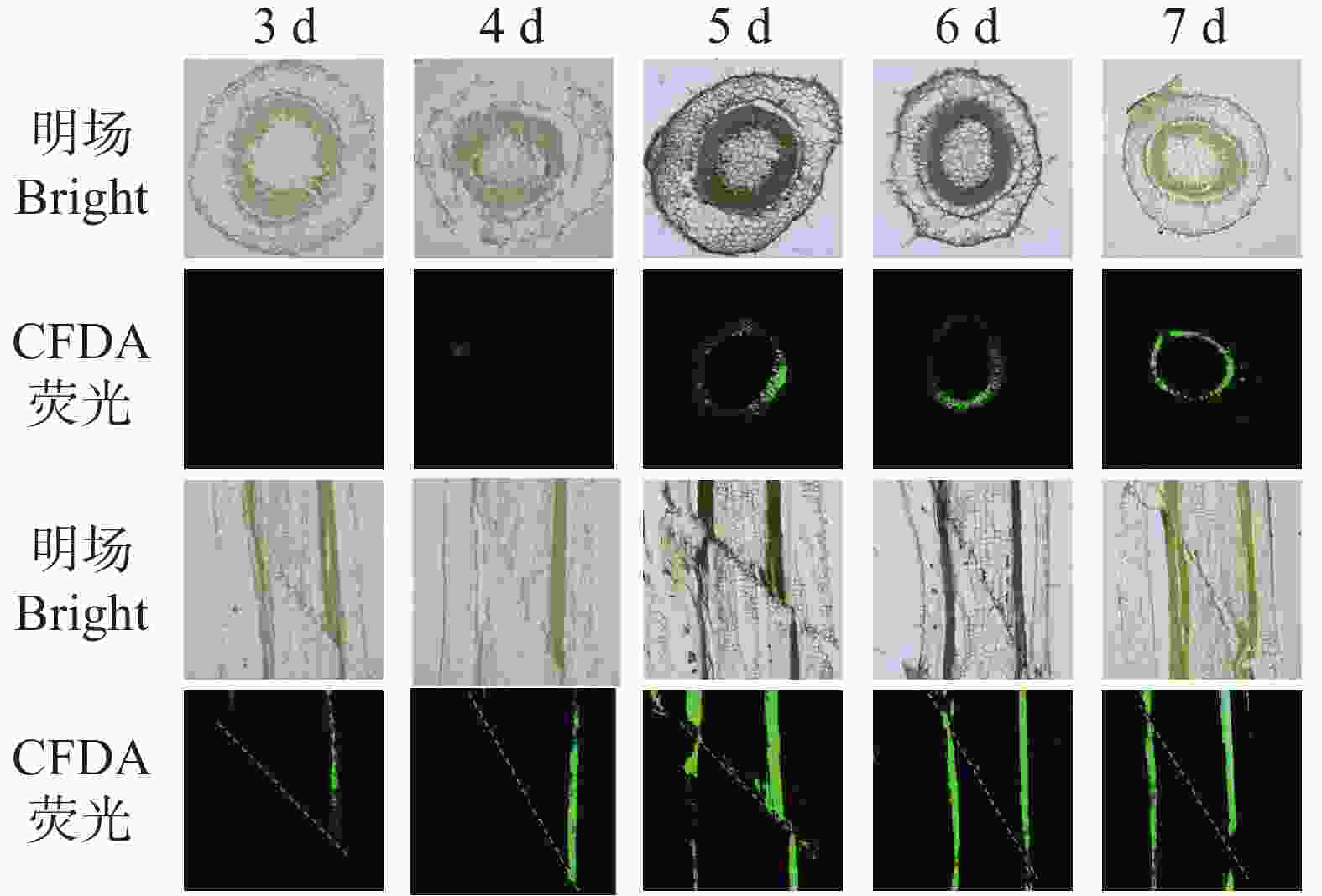
图 4 烟草嫁接中CFDA从接穗到砧木的运输过程
Figure 4. Transport of CFDA from the rootstock to the scion in tobacco grafts
-
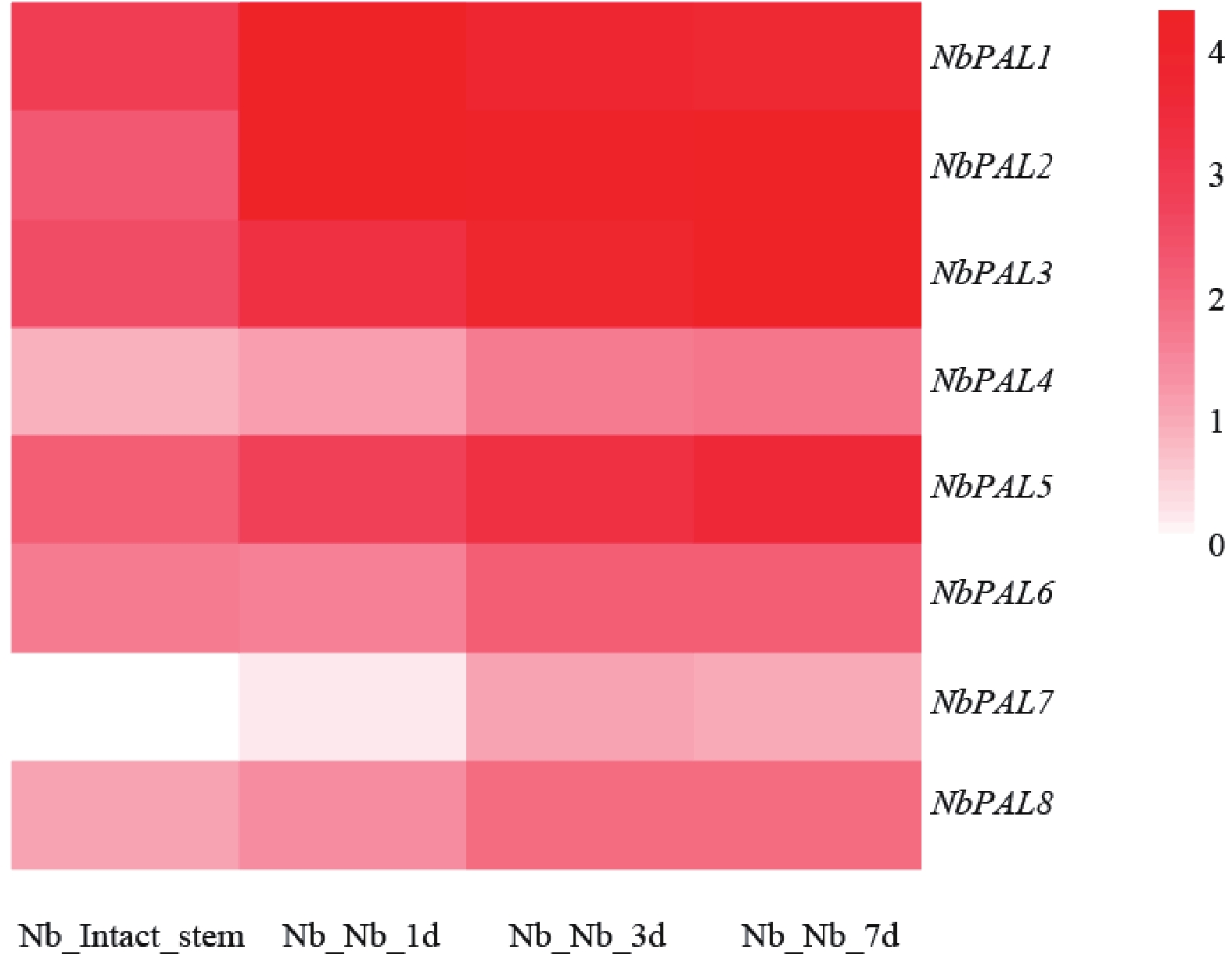
不同嫁接时期PAL基因家族基因的表达模式如图5所示,Nb_Intact_stem为未嫁接对照茎段,Nb_Nb_1d、Nb_Nb_3d、Nb_Nb_7d分别代表嫁接后1、3和7 d的样品。由图可知,个别NbPAL基因如NbPAL1、NbPAL2、NbPAL3 及 NbPAL5等基因在嫁接后各时间点呈现显著高表达,高表达基因的表达峰值出现在嫁接后1 d,3~7 d维持较高水平,与机械损伤嫁接诱导苯丙素代谢通路激活的规律一致。结果表明,烟草嫁接损伤胁迫可特异性诱导部分PAL相关基因的高表达。
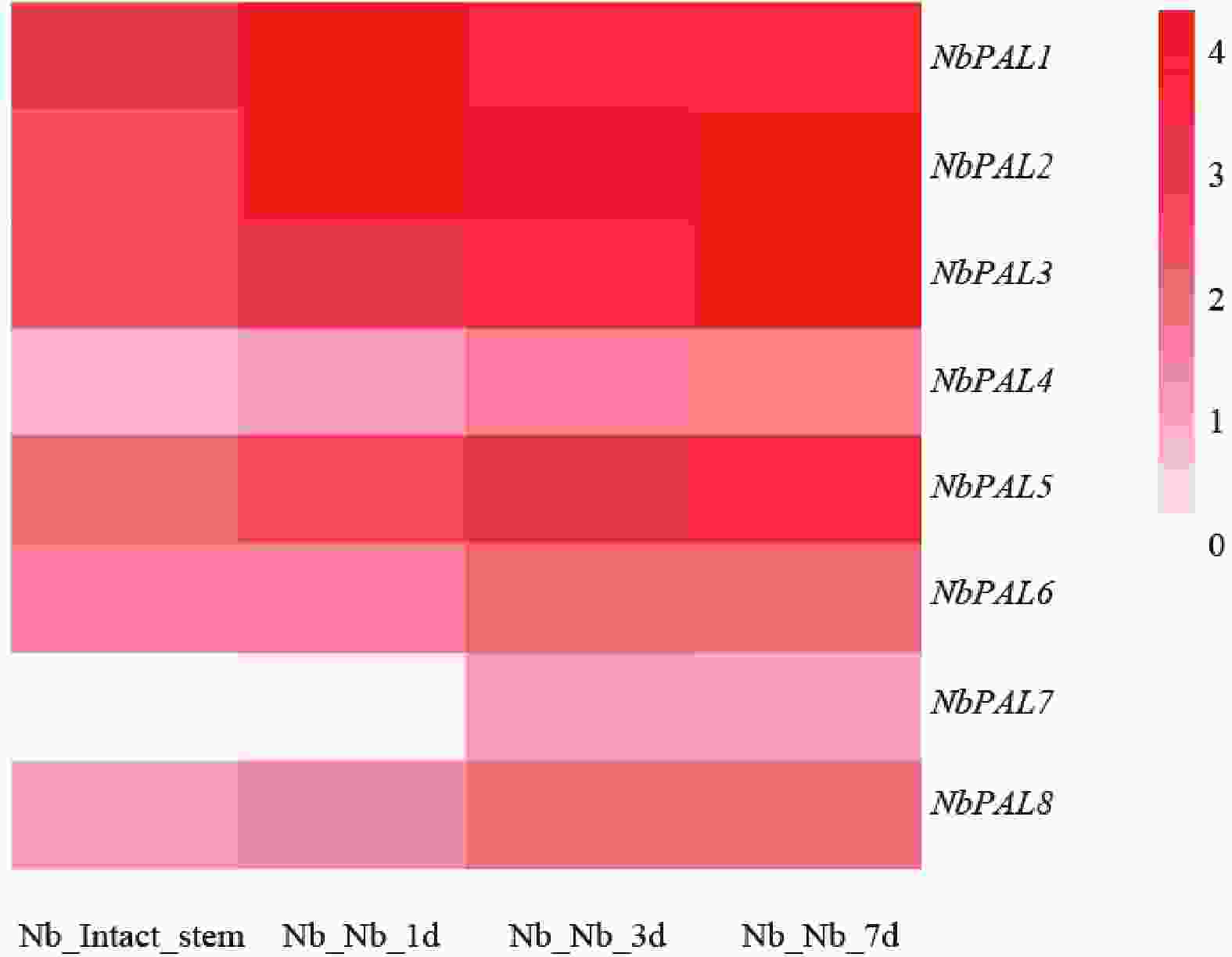
图 5 PAL基因家族在烟草嫁接植株的表达模式
Figure 5. Expression pattern of the PAL gene family in grafted tobacco plants
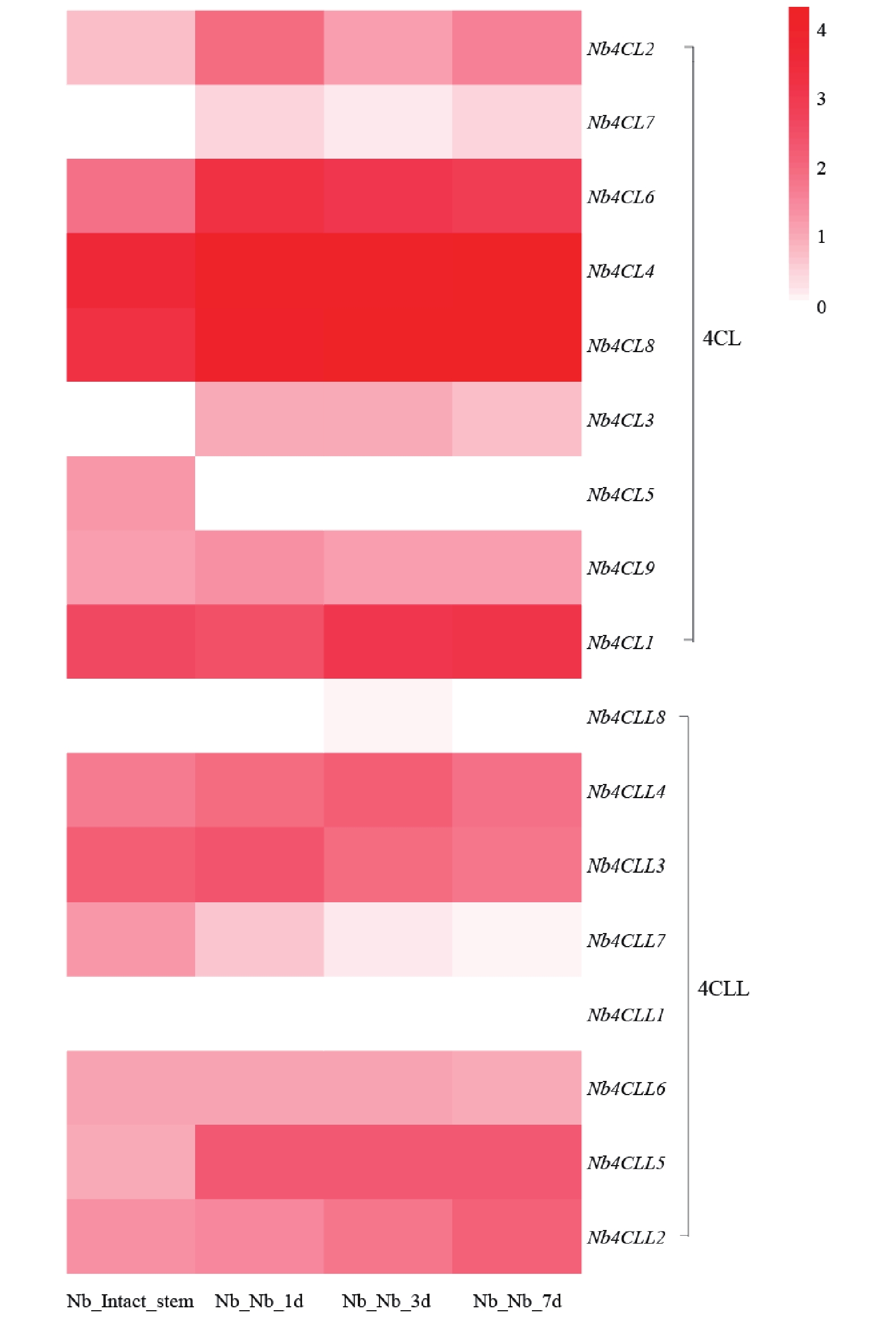
基于转录组数据绘制的4CL相关基因在本氏烟草不同嫁接时期的表达热图显示(图6),其中,Nb4CL6、Nb4CL4和Nb4CL8\等基因在嫁接后各时期均呈现较高表达水平,推测其可能在嫁接愈合全过程发挥基础性调控作用。Nb4CLL8和Nb4CLL1这2个基因在完整茎中表达较低,提示其可能参与嫁接早期的胁迫响应及愈合启动过程。4CL及其同源基因4CLL大部分基因在嫁接后3~7 d表达量逐步上调,该基因家族参与酚类物质代谢与木质素生物合成,推测其可能在嫁接后期维管组织重建及木质化过程中发挥关键作用。
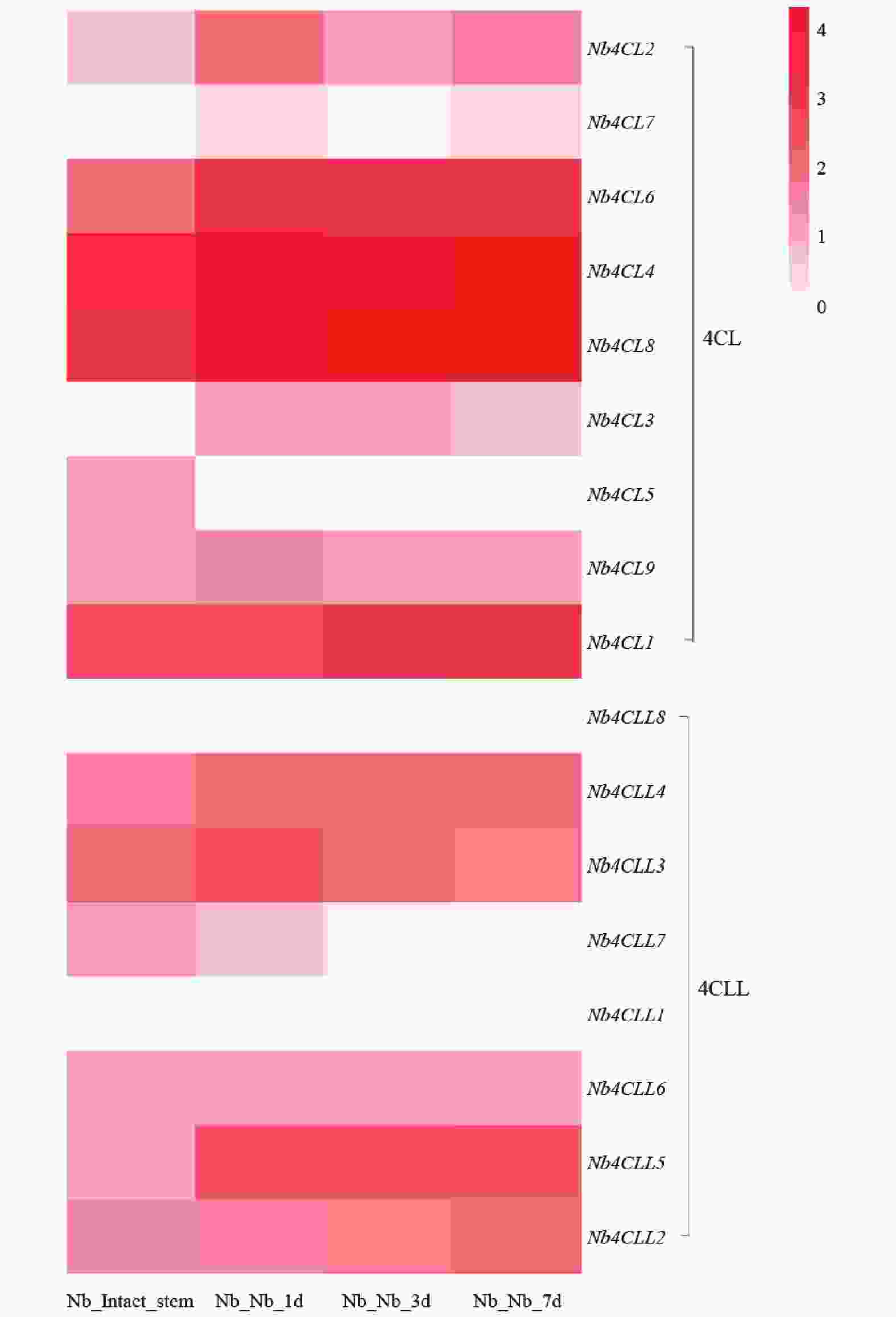
图 6 4CL基因家族在烟草嫁接植株的表达模式
Figure 6. Expression pattern of the 4CL gene family in grafted tobacco plants
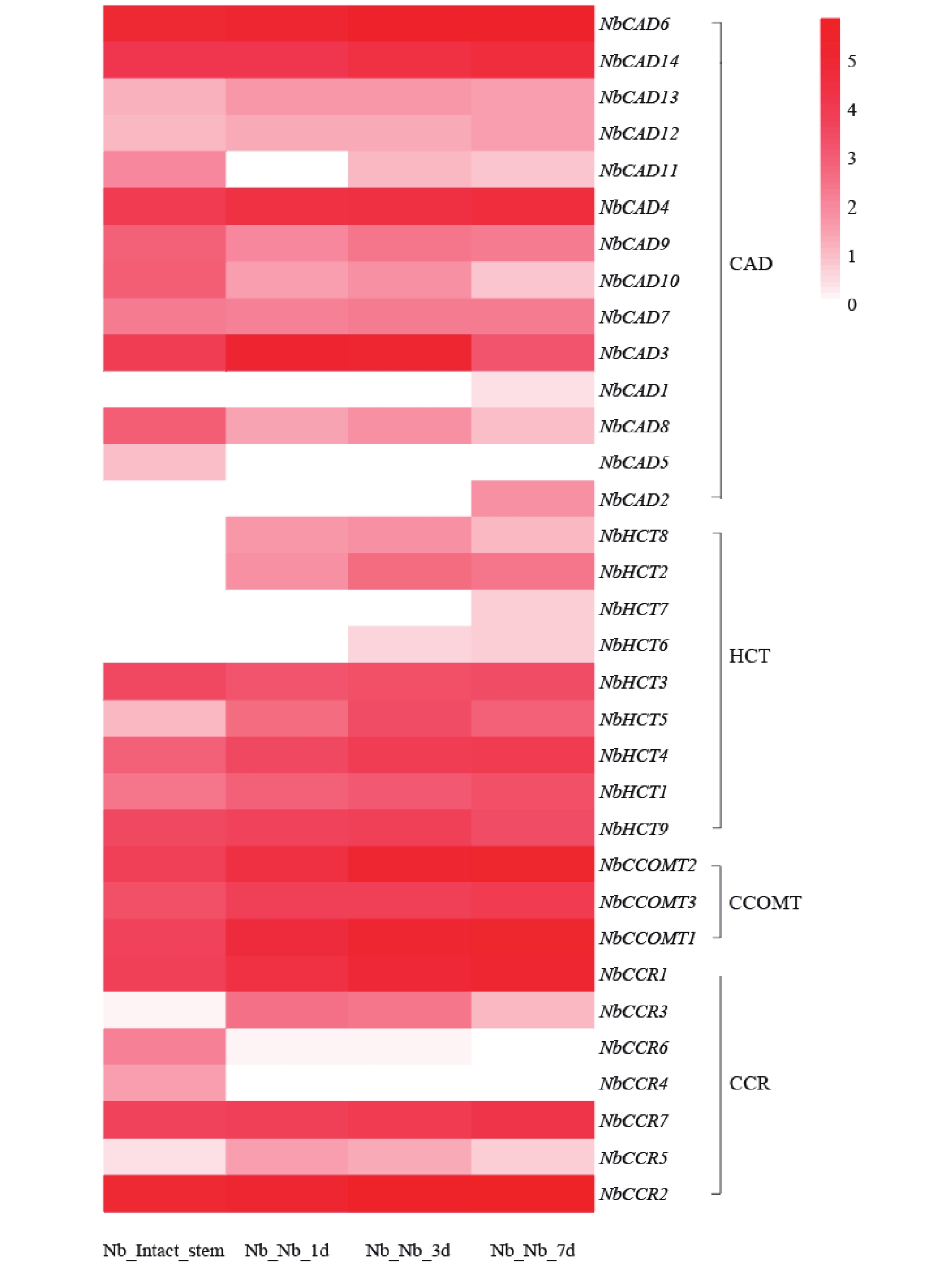
从图7可知,共鉴定到33个参与绿原酸生物合成途径的相关基因。其中,NbCAD6、NbCAD14、NbCAD4等基因在嫁接后1 d即显著上调,并在3~7 d维持较高表达水平,提示其可能参与嫁接早期胁迫响应及绿原酸合成的快速启动。NbCAD7、NbCAD3等基因在嫁接后3 d表达量达到峰值,7 d后仍维持较高水平,推测其在嫁接愈合中期绿原酸积累及防御响应中发挥关键作用。
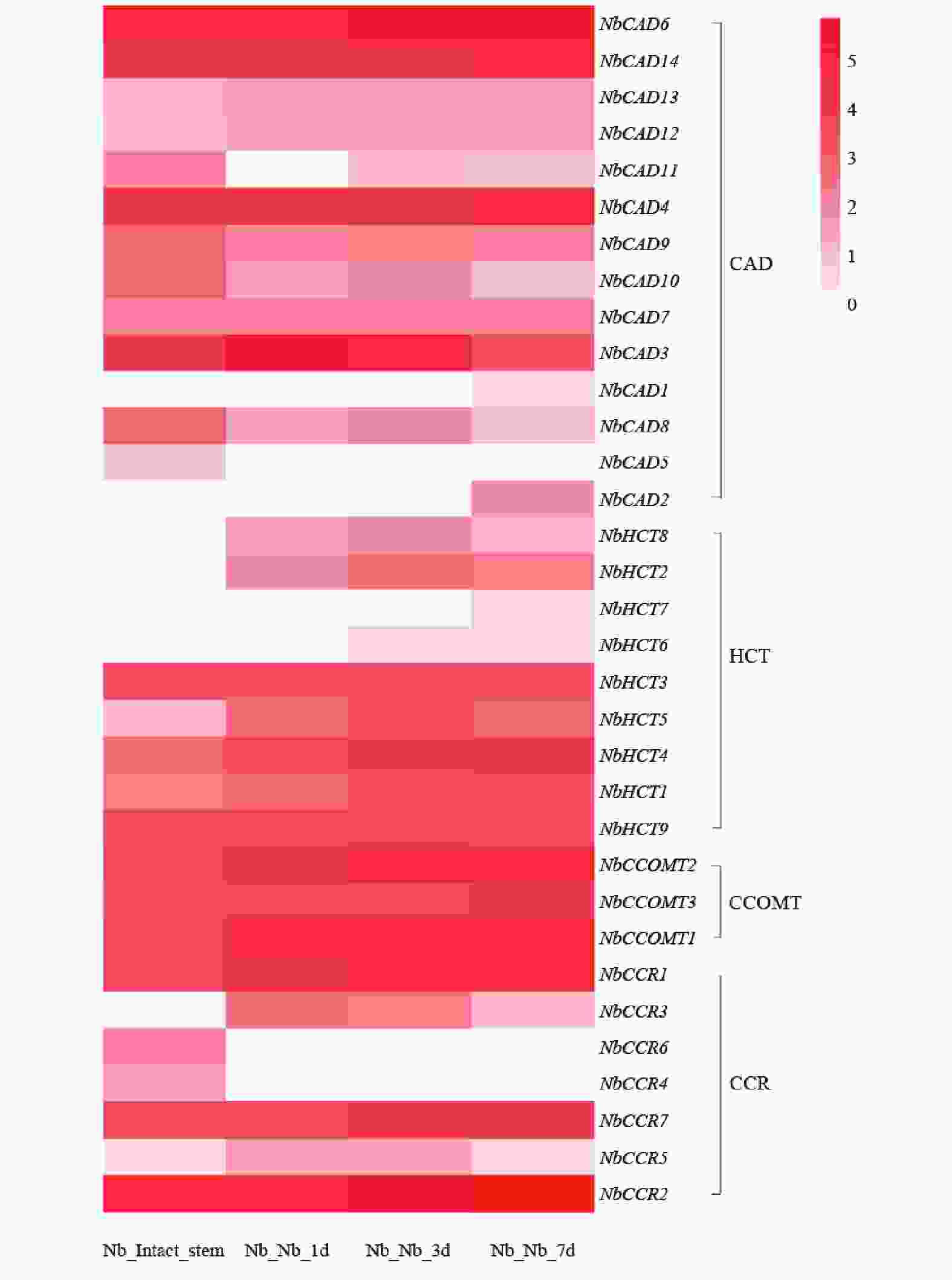
图 7 绿原酸合成相关基因在烟草嫁接植株的表达模式
Figure 7. Expression pattern of genes involved in chlorogenic acid biosynthesis in grafted tobacco plants
HCT其编码羟基肉桂酰转移酶,绿原酸生物合成的关键限速酶及其同源基因在嫁接后各时期均呈现高表达,尤其在嫁接后3~7 d表达水平持续升高,表明HCT基因家族可能在嫁接愈合过程中绿原酸的持续合成与积累中发挥核心调控作用。整体而言,绿原酸合成相关基因在嫁接后时序性上调表达,与嫁接愈合进程中防御响应、界面保护及木质化过程的需求相吻合,提示绿原酸代谢途径在烟草嫁接愈合中具有重要功能。
-
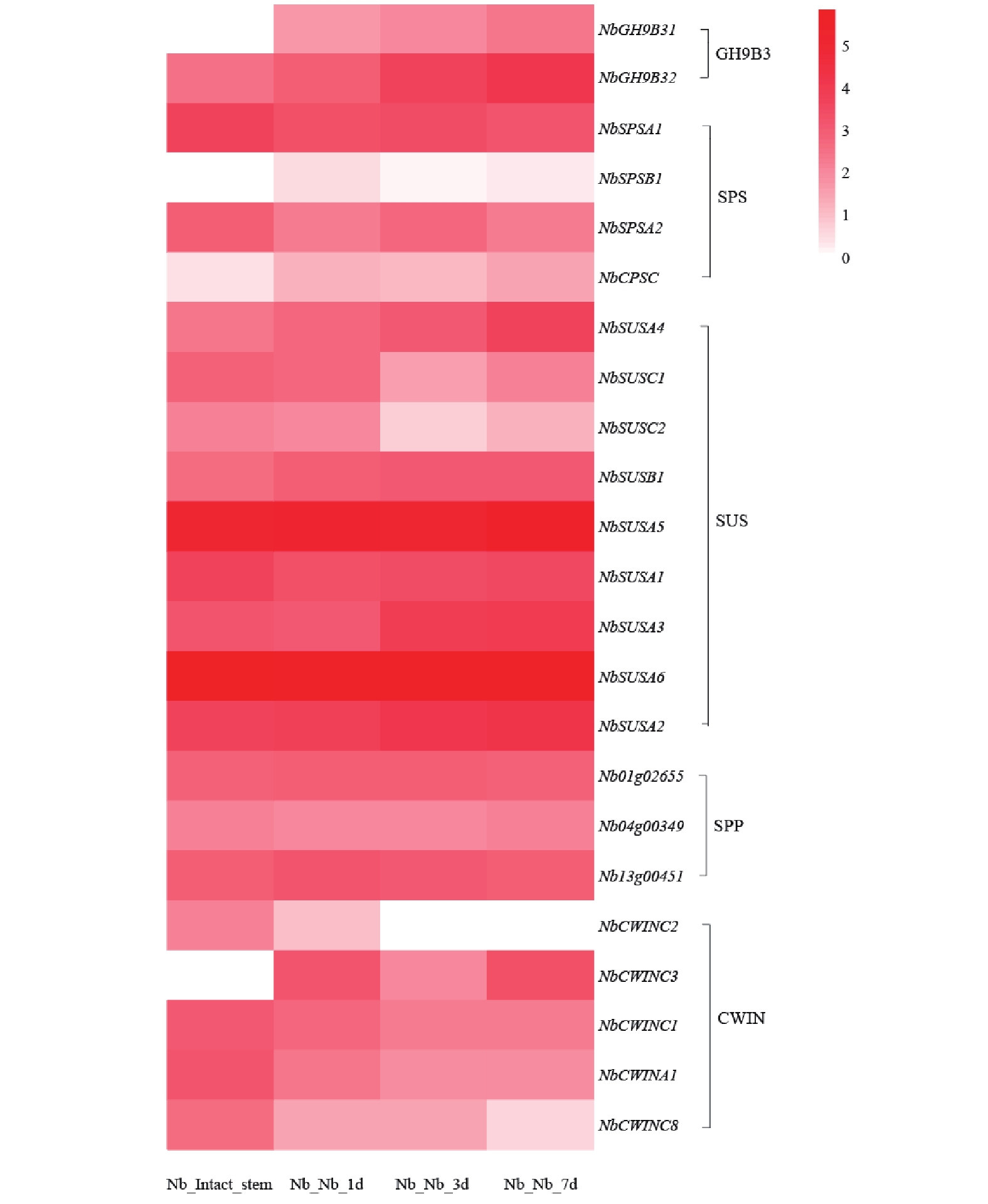
基于转录组数据,绘制了不同嫁接时期蔗糖代谢相关基因的表达热图(图8),SUS 家族中NbSUSA5、NbSUSA6等基因在嫁接后各时间点呈现显著高表达;SPS 家族中NbSPSA1、NbSPSA2也维持较高表达水平,而 SPP、CWIN 基因家族多数基因表达相对温和。结果表明,烟草嫁接可特异性诱导糖代谢核心基因的高表达,其早期且持续的激活可能通过调控糖浓度差异、促进细胞壁重构等途径,参与嫁接界面的维管连接与愈合进程,是嫁接恢复及亲和性建立的重要分子调控特征之一。
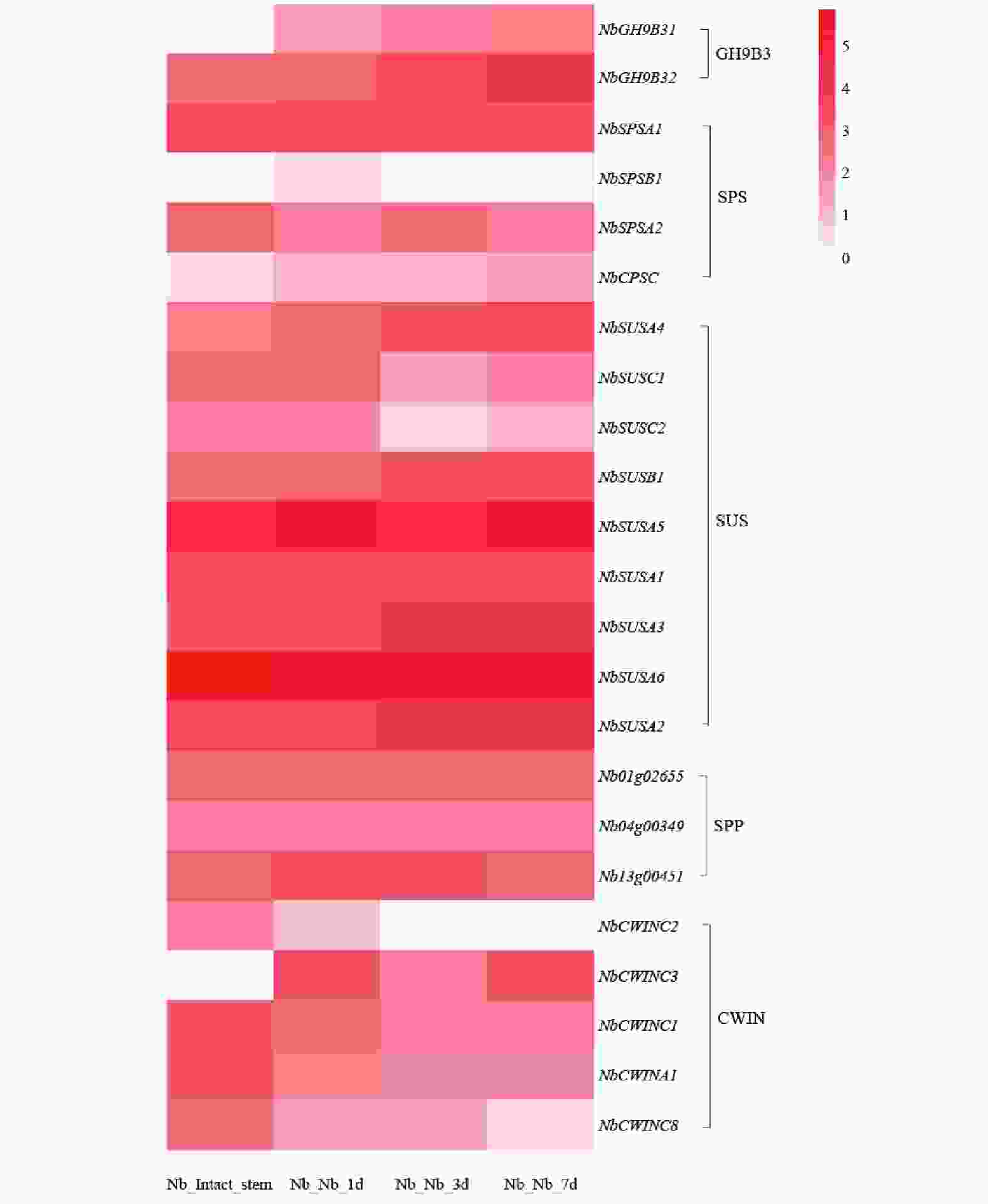
图 8 蔗糖代谢在烟草嫁接植株的表达模式
Figure 8. Expression pattern of sucrose metabolism in grafted tobacco plants
-
本研究通过系统分析烟草嫁接后维管组织重建、酚类代谢及糖代谢的动态变化,初步揭示了嫁接愈合过程的分子与生理调控网络。
植物维管系统(包括韧皮部与木质部)负责地上部与地下部之间的信号交流,以及有机物、水分和养分的长距离运输。因此,维管重新连接是嫁接成功的关键;若连接失败,大多数嫁接组合将难以长期存活[21]。研究表明,拟南芥(Arabidopsis thaliana)韧皮部通常在嫁接后3~4 d完成连接,木质部则在6~7 d完成[22];水稻(Oryza sativa)胚胎下胚轴嫁接也遵循相似的时序规律,韧皮部连接发生于5~7 d,木质部连接在6~10 d[23];番茄(Solanum lycopersicum)嫁接中,木质部连接于4~5 d完成,韧皮部连接则发生在5~6 d[24]。本研究表明,烟草木质部与韧皮部在嫁接后4~5 d开始初步连通,于6~7 d达到稳定运输状态,且韧皮部重建略晚于木质部。综合已有研究可判断,嫁接后3~7 d是维管系统重建的关键时期,但具体进程受植物种类、嫁接方式、苗龄及检测方法等多重因素影响。
嫁接过程会引发与酚类化合物等次生代谢产物合成相关的生化改变[25]。酚类化合物的合成可由机械损伤、病原菌侵染等胁迫条件及植物防御机制所诱导,被视为植物复杂防御系统的一部分[26]。已有研究表明,苯丙烷代谢途径是影响嫁接不亲和、导致生理障碍的重要因素之一[27]。本研究的转录组数据分析发现,嫁接后1 d即出现PAL、4CL等苯丙烷途径关键基因的显著上调,并持续至14 d,提示机械损伤迅速启动了防御与修复相关的次生代谢程序。苯丙烷代谢途径的产物不仅参与儿茶素、木质素等多种酚类物质的合成[23],同时也参与绿原酸的生成。木质素是普遍存在于植物细胞壁中的多酚类聚合物,在维管植物木质部中大量存在;该途径为木质素合成提供前体[28]。绿原酸等酚类物质的积累可发挥抗氧化、抗菌功能[24],保护伤口免受病原侵染及氧化损伤[29];体外实验也证实其对多种病原真菌具有抑菌作用[30]。本研究转录组数据显示,该生物合成途径的相关基因在嫁接后1 d即显著上调,并在3~7 d维持较高表达水平,表明其可能参与愈伤组织木质化、新生维管细胞壁加厚,以及绿原酸代谢途径在烟草嫁接愈合中具有重要功能。
糖代谢的重编程可能为愈合过程提供能量与信号支持。嫁接部位上方接穗糖分积累、下方砧木糖分消耗,可导致糖诱导基因与糖抑制基因在接穗与砧木中呈现不对称表达模式。这种不对称性是嫁接与组织愈合的重要特征,可能在嫁接界面维管组织连接中发挥关键作用[31]。研究表明,苹果(Malus domestica)砧木可通过调控蔗糖代谢关键基因SPS与SUS的表达,参与淀粉与蔗糖代谢通路,从而调控果实糖代谢、营养积累及成熟进程,最终影响果实品质与早熟特性[32]。在嫁接处理下,南瓜(Cucurbita moschata)砧木子叶作为源器官,其CWIN活性显著升高,促进蔗糖降解为可利用的小分子糖,为砧穗生长提供能量与碳源[33]。本研究发现,SUS与SPS家族关键基因在烟草嫁接后显著持续高表达,而SPP与CWIN家族基因表达相对温和,表明糖代谢核心基因的特异性激活可能参与调控嫁接界面维管连接与愈合,是嫁接亲和性形成的重要分子特征。
综上所述,本研究从维管重建、酚类代谢与糖代谢三个维度,系统提供了烟草嫁接愈合过程的实验数据,表明次生代谢与初级代谢在嫁接愈合中具有协同作用:苯丙烷途径通过提供结构物质与防御代谢物,参与伤口封闭与抗逆;糖代谢通过提供能量底物与信号分子,支撑细胞增殖与维管分化。以上结果为深入解析嫁接相容性机制提供了新视角,也为利用代谢调控优化嫁接技术奠定了理论基础。未来可通过多组学整合分析及关键基因功能验证,进一步阐明酚类物质与糖信号互作调控维管再生的分子细节。
本氏烟草自嫁接维管重连与糖、酚代谢相关基因的表达分析
DOI: 10.15886/j.cnki.rdswxb.20260057
 CSTR: 32425.14.j.cnki.rdswxb.20260057
CSTR: 32425.14.j.cnki.rdswxb.20260057
Expression analysis of genes related to vascular reconnection, sugar and phenolic metabolism during self-grafting of Nicotiana benthamiana
-
摘要: 为了揭示酚类与糖类在嫁接愈合中的协同作用机制,以本氏烟草(Nicotiana benthamiana)为材料,研究嫁接后维管重连的动态过程及其与蔗糖代谢、酚类合成相关基因表达的关系。采用CFDA荧光示踪技术观测木质部与韧皮部的再连通过程,并结合转录组数据分析关键代谢通路相关基因的表达变化。结果表明,嫁接后3~4 d为维管组织初步修复期,导管与筛管连接不完整,物质运输受阻;嫁接后5~7 d,木质部与韧皮部在横向与纵向上形成连续畅通的输导通道,荧光信号显著增强,表明运输功能基本恢复。转录组分析显示,蔗糖代谢相关基因在嫁接后普遍上调表达,可能为维管再生提供能量与碳骨架,并参与糖信号调控;酚类合成相关基因同样整体高表达,提示其在界面保护、木质化及抗逆响应中发挥重要作用。综上,本氏烟草嫁接愈合过程中,维管形态重建与关键代谢途径的转录激活呈现时间协同性,为解析嫁接愈合的生理与分子机制提供了依据。Abstract: An attempt was made to reveal the synergistic mechanism of phenolic compounds and sugars during graft healing. Nicotiana benthamiana was selected as the material to investigate the dynamic process of vascular reconnection and its relationship with the expression of genes related to sucrose metabolism and phenolic biosynthesis. The reconnection process of xylem and phloem was obserced by employing CFDA fluorescence tracer, and transcriptome data were analyzed to profile the expression changes of genes involved in key metabolic pathways. The results showed that days 3–4 after grafting were the initial vascular repair stage, during which vessel and sieve element connections were incomplete, resulting in impaired substance transport. At days 5–7, continuous and unobstructed transport channels were formed both transversely and longitudinally in the xylem and phloem, accompanied by significantly enhanced fluorescence signals, indicating that transport function was largely restored. Transcriptome analysis revealed that sucrose metabolism-related genes were generally upregulated after grafting, potentially providing energy and carbon skeletons for vascular regeneration and participating in sugar signaling regulation. Genes involved in phenolic biosynthesis were also highly expressed overall, suggesting their critical roles in interface protection, lignification, and stress responses. In conclusion, during graft healing in N. benthamiana, vascular morphological reconstruction and transcriptional activation of key metabolic pathways exhibited temporal coordination, providing a basis for elucidating the physiological and molecular mechanisms underlying graft healing.
-
Key words:
- Nicotiana benthamiana /
- grafting /
- sucrose metabolism /
- phenolic compounds /
- transcriptome analysis
-
图 2 烟草自嫁接后不同时间生长状态
Fig. 2 Growth status of tobacco plants at different time points after self-grafting
图 3 烟草嫁接中CFDA从砧木到接穗的运输过程。
注:将CFDA施加于砧木,分别在接穗的横切面(上)和嫁接连接处的纵切面(下)监测荧光。
Fig. 3 Transport of CFDA from the rootstock to the scion in tobacco grafts
Note: CFDA was applied to the rootstock, and fluorescence was monitored in transverse sections of the scion (top)and longitudinal sections of the graft junction (bottom).
图 4 烟草嫁接中CFDA从接穗到砧木的运输过程
注:将CFDA应用于接穗,并在根砧横截面(上方)和嫁接接合处纵向段(下端)监测荧光。
Fig. 4 Transport of CFDA from the rootstock to the scion in tobacco grafts
Note: CFDA was applied to the scion and fluorescence was monitored in transverse sections of the rootstock (top)and longitudinal sections of the graft junction (bottom).
图 5 PAL基因家族在烟草嫁接植株的表达模式
Fig. 5 Expression pattern of the PAL gene family in grafted tobacco plants
图 6 4CL基因家族在烟草嫁接植株的表达模式
Fig. 6 Expression pattern of the 4CL gene family in grafted tobacco plants
图 7 绿原酸合成相关基因在烟草嫁接植株的表达模式
Fig. 7 Expression pattern of genes involved in chlorogenic acid biosynthesis in grafted tobacco plants
-
[1] 黄凯波, 徐世福, 桑锦凯, 等. 国内嫁接技术概述[J]. 湖北农机化, 2019(13): 42. https://doi.org/10.3969/j.issn.1009-1440.2019.13.032 doi: 10.3969/j.issn.1009-1440.2019.13.032 [2] 陈晶晶, 李栋梁, 杨倩, 等. 植物嫁接再生机理研究进展[J]. 植物生理学报, 2020, 56(8): 1690−1702.(查阅网上资料,doi打不开,请确认) https://doi.org/10.13592/j.cnki.ppj.2019.0575 [3] 邓竹英. 拟南芥/本生烟草远缘嫁接亲和机理研究[D]. 荆州: 长江大学, 2022. https://doi.org/10.26981/d.cnki.gjhsc.2022.000002 [4] Huang S X, Ding J, Deng D J, et al. Draft genome of the kiwifruit Actinidia chinensis [J]. Nature Communications, 2013, 4: 2640. https://doi.org/10.1038/ncomms3640 doi: 10.1038/ncomms3640 [5] Kotoda N, Iwanami H, Takahashi S, et al. Antisense expression of MdTFL1, a TFL1-like gene, reduces the juvenile phase in apple [J]. Journal of the American Society for Horticultural Science, 2006, 131(1): 74−81. https://doi.org/10.21273/JASHS.131.1.74 doi: 10.21273/JASHS.131.1.74 [6] Oberhuber A, Schwarz A, Hoffmann M H, et al. Influence of different self-expanding stent-graft types on remodeling of the aortic neck after endovascular aneurysm repair [J]. Journal of Endovascular Therapy, 2010, 17(6): 677−684. https://doi.org/10.1583/10-3172.1 doi: 10.1583/10-3172.1 [7] 陈哲. ‘井岗红糯’荔枝嫁接亲和性及其机理研究[D]. 广州: 华南农业大学, 2016. doi: 10.7666/d.D01036724 [8] Wang L X, Liao Y M, Liu J M, et al. Advances in understanding the graft healing mechanism: a review of factors and regulatory pathways [J]. Horticulture Research, 2024, 11(8): uhae175. https://doi.org/10.1093/hr/uhae175 doi: 10.1093/hr/uhae175 [9] Yin H, Yan B, Sun J, et al. Graft-union development: a delicate process that involves cell-cell communication between scion and stock for local auxin accumulation [J]. Journal of Experimental Botany, 2012, 63(11): 4219−4232. https://doi.org/10.1093/jxb/ers109 doi: 10.1093/jxb/ers109 [10] Loupit G, Brocard L, Ollat N, et al. Grafting in plants: recent discoveries and new applications [J]. Journal of Experimental Botany, 2023, 74(8): 2433−2447. https://doi.org/10.1093/jxb/erad061 doi: 10.1093/jxb/erad061 [11] He W, Xie R, Wang Y, et al. Comparative transcriptomic analysis on compatible/incompatible grafts in Citrus [J]. Horticulture Research, 2022, 9: uhab072. https://doi.org/10.1093/hr/uhab072 doi: 10.1093/hr/uhab072 [12] Goodin M M, Zaitlin D, Naidu R A, et al. Nicotiana benthamiana: its history and future as a model for plant-pathogen interactions [J]. Molecular Plant-Microbe Interactions® , 2008, 21(8): 1015−1026. https://doi.org/10.1094/MPMI-21-8-1015 doi: 10.1094/MPMI-21-8-1015 [13] Xia C, Zheng Y, Huang J, et al. Identification of phloem mobile mRNAs using the Solanaceae heterograft system[M]//Liesche J. Phloem: methods and protocols. New York: Humana, 2019: 421-431. doi: 10.1007/978-1-4939-9562-2_32 [14] Notaguchi M, Kurotani K I, Sato Y, et al. Cell-cell adhesion in plant grafting is facilitated by β-1, 4-glucanases [J]. Science, 2020, 369(6504): 698−702. https://doi.org/10.1126/science.abc3710 doi: 10.1126/science.abc3710 [15] Habibi F, Liu T, Folta K, et al. Physiological, biochemical, and molecular aspects of grafting in fruit trees [J]. Horticulture Research, 2022, 9: uhac032. https://doi.org/10.1093/hr/uhac032 doi: 10.1093/hr/uhac032 [16] Pina A, Errea P, Martens H J. Graft union formation and cell-to-cell communication via plasmodesmata in compatible and incompatible stem unions of Prunus spp. [J]. Scientia Horticulturae, 2012, 143: 144−150. https://doi.org/10.1016/j.scienta.2012.06.017 doi: 10.1016/j.scienta.2012.06.017 [17] 刘婧冉, 杜长霞, 樊怀福. 植物嫁接砧穗愈合机制研究进展[J]. 浙江农林大学学报, 2018, 35(3): 552−561. https://doi.org/10.11833/j.issn.2095-0756.2018.03.022 doi: 10.11833/j.issn.2095-0756.2018.03.022 [18] 刘新颖. 光质对黄瓜嫁接苗质量及愈合进程的影响研究[D]. 北京: 中国农业科学院, 2023. https://doi.org/10.27630/d.cnki.gznky.2023.000549 [19] Choudhary A, Kumar A, Kaur N, et al. Molecular cues of sugar signaling in plants [J]. Physiologia Plantarum, 2022, 174(1): e13630. https://doi.org/10.1111/ppl.13630 doi: 10.1111/ppl.13630 [20] Guan C T, Xu Y G, Yue H Z, et al. Difference in sucrose concentration between scion and rootstock influences the incompatibility of cucumber/pumpkin grafted plants [J]. Horticultural Plant Journal, 2025, 11(3): 1166−1180. https://doi.org/10.1016/j.hpj.2024.02.014 doi: 10.1016/j.hpj.2024.02.014 [21] Thomas H, Van den Broeck L, Spurney R, et al. Gene regulatory networks for compatible versus incompatible grafts identify a role for SlWOX4 during junction formation [J]. The Plant Cell, 2022, 34(1): 535−556. https://doi.org/10.1093/plcell/koab246 doi: 10.1093/plcell/koab246 [22] Melnyk C W, Schuster C, Leyser O, et al. A developmental framework for graft formation and vascular reconnection in Arabidopsis thaliana [J]. Current Biology, 2015, 25(10): 1306−1318. https://doi.org/10.1016/j.cub.2015.03.032 doi: 10.1016/j.cub.2015.03.032 [23] Reeves G, Tripathi A, Singh P, et al. Monocotyledonous plants graft at the embryonic root-shoot interface [J]. Nature, 2022, 602(7896): 280−286. https://doi.org/10.1038/s41586-021-04247-y doi: 10.1038/s41586-021-04247-y [24] Cui Q Q, Xie L L, Dong C J, et al. Stage-specific events in tomato graft formation and the regulatory effects of auxin and cytokinin [J]. Plant Science, 2021, 304: 110803. https://doi.org/10.1016/j.plantsci.2020.110803 doi: 10.1016/j.plantsci.2020.110803 [25] Liu Q, Wang X R, Zhao Y, et al. Transcriptome and physiological analyses reveal new insights into delayed incompatibility formed by interspecific grafting [J]. Scientific Reports, 2023, 13(1): 4574. https://doi.org/10.1038/s41598-023-31804-4 doi: 10.1038/s41598-023-31804-4 [26] Pina A, Errea P. A review of new advances in mechanism of graft compatibility–incompatibility [J]. Scientia Horticulturae, 2005, 106(1): 1−11. https://doi.org/10.1016/j.scienta.2005.04.003 doi: 10.1016/j.scienta.2005.04.003 [27] Pina A, Zhebentyayeva T, Errea P, et al. Isolation and molecular characterization of cinnamate 4-hydroxylase from apricot and plum [J]. Biologia Plantarum, 2012, 56(3): 441−450. https://doi.org/10.1007/s10535-012-0114-2 doi: 10.1007/s10535-012-0114-2 [28] Clé C, Hill L M, Niggeweg R, et al. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum;consequences for phenolic accumulation and UV-tolerance [J]. Phytochemistry, 2008, 69(11): 2149−2156. https://doi.org/10.1016/j.phytochem.2008.04.024 doi: 10.1016/j.phytochem.2008.04.024 [29] Martínez G, Regente M, Jacobi S, et al. Chlorogenic acid is a fungicide active against phytopathogenic fungi [J]. Pesticide Biochemistry and Physiology, 2017, 140: 30−35. https://doi.org/10.1016/j.pestbp.2017.05.012 doi: 10.1016/j.pestbp.2017.05.012 [30] Dellero Y, Berardocco S, Bouchereau A. U-13C-glucose incorporation into source leaves of Brassica napus highlights light-dependent regulations of metabolic fluxes within central carbon metabolism [J]. Journal of Plant Physiology, 2024, 292: 154162. https://doi.org/10.1016/j.jplph.2023.154162 doi: 10.1016/j.jplph.2023.154162 [31] Miao L, Li Q, Sun T S, et al. Sugars promote graft union development in the heterograft of cucumber onto pumpkin [J]. Horticulture Research, 2021, 8: 146. https://doi.org/10.1038/s41438-021-00580-5 doi: 10.1038/s41438-021-00580-5 [32] Liu R H, Jia J W, Wang C W, et al. An Transcriptomic and primary metabolic profiles reveal the mechanism of development and maturation of fuji apple grafted onto different dwarfed intermediate rootstocks [J]. Scientia Horticulturae, 2025, 343: 114060. https://doi.org/10.1016/j.scienta.2025.114060 doi: 10.1016/j.scienta.2025.114060 [33] Pu D, Wen Z Y, Sun J B, et al. Unveiling the mechanism of source-sink rebalancing in cucumber-pumpkin heterografts: the buffering roles of rootstock cotyledon [J]. Physiologia Plantarum, 2024, 176(2): e14232. https://doi.org/10.1111/ppl.14232 doi: 10.1111/ppl.14232 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 33
- HTML全文浏览量: 21
- 被引次数: 0
