

-
草地生态系统约占全球陆地面积的三分之一[1],是地球陆地生态系统的重要组成部分,具有重要的生态和经济价值[2,3]。在保护生物多样性、调节区域气候等方面发挥着重要作用[4]。特别是在干旱和半干旱地区,草地对维护区域生态安全、支持碳循环以及牧区社会经济的可持续发展至关重要。然而,受全球气候变暖和人类活动影响,草地生态系统的生产力持续下降、退化过程加速,生态稳定性明显减弱[5]。长期过度放牧被认为是导致这种变化的重要原因,它不仅改变了植被群落结构,还影响草地的固碳能力与生态承载力[6]。净初级生产力(net primary productivity, NPP)作为反映植被光合固定碳能力的关键生态指标,能够有效表征草地生态系统的生物量积累与生态功能状态[7,8]。在草地生态研究中,NPP既是评估草地健康与退化风险的重要参数,也是量化载畜压力、计算载畜阈值与制定放牧管理策略的基础依据。近年来,随着遥感技术的快速发展,利用MODIS等卫星产品反演NPP成为评估草地生产力动态的重要手段,这为大尺度草地生态承载力评估提供了数据支持[8]。然而,大多数研究都是基于“实际NPP”分析的,容易受到放牧干扰影响从而低估草地未受放牧影响的生产能力,也难以准确识别生态系统在理想状态下的真实承载阈值。因此,建立以未受放牧影响的 NPP 为基础的载畜阈值评估框架,可更准确反映草地在不同放牧强度下的响应过程与退化临界特征。
载畜阈值是指草地在不发生退化的前提下所能承载的最大放牧强度。合理的载畜阈值对于实现草畜平衡、调控草地利用强度、防止草地退化具有重要意义。目前载蓄阈值方面的研究大体可以分为以下几类:第一类是经验模型与静态估算,该类方法大多以单位面积产草量或草地NPP与标准载畜量的比值为基础,结合经验参数推算载畜阈值。Zhang等[9]通过在中国典型草原开展不同放牧强度实验,发现减少放牧压力可同时提升草地产量与环境效益,进而推断存在维持生态与生产平衡的“最佳放牧强度”区间;Qin等[10]利用高时空分辨率遥感数据,估算内蒙古草地的理论载畜量并识别草畜平衡与超载区。这类方法直观且易操作,但主要适用于相对稳定的自然-人为背景,缺乏对生态系统响应过程的动态刻画,难以捕捉临界退化点。第二类是生态-经济耦合模型,这类方法引入了生态学与经济学双重指标,构建放牧效益、草地退化风险与生态承载力之间的多维评估体系。Huang等[11]将草地生产力、牲畜结构、经济效益和退化风险整合为阈值评估指标体系,并构建多目标权衡模型;Zhang等[12]在此基础上进一步引入气候情景分析,讨论载畜阈值在不同气候背景下的变化趋势。这类方法增强了模型的综合性和决策关联性,但多数依赖静态输入参数或多年平均值,并且数据获取成本较,生态系统的非线性响应过程与时序退化机制仍缺乏刻画。第三类是遥感驱动的载畜比-压力模型。随着遥感技术的发展,越来越多的研究者逐渐将遥感反演的 NPP 用作载畜阈值计算的关键指标,并结合实测载畜量进行比值分析,从而识别草地放牧强度的安全与超载状态。Huang等[13]分析了北方草地的饲草-畜群平衡及其保育模式,为基于遥感的放牧压力监测提供了空间化框架。但是该类研究多数仍采用阈值设定,缺乏长期放牧干扰下草地生产力逐步衰减的过程模拟,也未能明确“退化触发点”在时间维度上的表现形式。这一类研究改进了空间尺度的适应性,并具有良好的操作性,但仍然停留在静态指标评估,未能模拟扰动积累下生态系统退化的全过程。第四类是机器学习与多源数据集成方法,此类方法是最新研究尝试融合遥感、气象、地形、土壤和畜牧活动等多源数据,利用机器学习方法识别载畜阈值特征因子和变化模式[14]。虽然在精度和预测能力上有明显提升,但模型解释性较弱,难以揭示退化临界点的生态机制。
尽管近年来载畜阈值研究取得了显著进展,但现有研究仍普遍存在一个核心局限:多数方法以已经受到放牧影响的草地生物量或实际 NPP 作为计算基础,通过样方调查或遥感反演的现状生产力来反推可承载的牲畜数量。这类方法在技术上可行,但在生态意义上存在内在矛盾,即用“已被消耗后的草地状态”去推断“尚未发生退化时能够承载的最大放牧强度”,从而容易低估草地的潜在生产能力,或混淆自然生产力差异与人为放牧干扰的影响。不同研究所采用的样方尺度、调查强度和经验系数差异较大,也使得载畜阈值结果在区域之间缺乏可比性,限制了其在宏观草地管理中的推广应用。因此,有必要引入一种以“未受放牧影响的植被生产力”为统一生态基准,在空间上连续估算草地潜在承载能力的方法,以提高载畜阈值的生态合理性与区域可比性。
本研究以天山北坡草地为研究区域,构建了一种基于未受放牧影响的净初级生产力的过程化载畜阈值识别框架。首先基于 MODIS LAI(MOD15A2H)时间序列数据构建理想LAI 生长曲线,以表征无放牧条件下草地的生长状态,并利用光能利用率(light use efficiency,LUE)模型计算未受放牧影响的 NPP。在此基础上,通过逐年递增载畜压力,动态模拟草地 NPP 在持续放牧干扰下的衰减过程,识别生态系统从稳定到退化的临界点,并量化到达阈值时的载畜量和时间。研究还引入 RCP4.5 和 RCP8.5 两种气候变化情景,评估未来气候变暖背景下NPP 变化对载畜阈值的影响,以揭示气候驱动与人类干扰的交互机制。该方法不仅在理论上完善了基于未受放牧影响的生产力的阈值识别逻辑,也在实践上提供了一种可量化、可视化、可对比的载畜阈值评估新路径。通过将未受放牧影响的生产力与时序退化过程相结合,本研究旨在为干旱与半干旱地区草地的退化预警、适度放牧管理与气候适应性调控提供科学依据与技术支撑。
-
天山北坡位于40°50′—47°60′N和79°42′— 96°36′E之间。西起伊犁河谷,东至准噶尔盆地边缘,主要包括新疆维吾尔自治区的伊犁哈萨克自治州、昌吉回族自治州和乌鲁木齐市等地区(图1)。该区域地势西高东低,自西向东依次由高山草甸、山地草原、典型草原过渡至荒漠草原与戈壁地貌,呈现明显的垂直带谱特征。天山山脉作为中亚重要的生态安全屏障,不仅是干旱区主要的水源涵养地和碳汇中心[15-16],也是新疆畜牧业生产的核心牧区。为避免将依法禁止或严格限制放牧的区域误纳入模拟,本研究团队对研究区内自然保护区范围进行了处理。包括塔城巴尔鲁克山自然保护区、新疆艾比湖湿地自然保护区、新疆西天山自然保护区、天池博格达峰自然保护区,以及新疆罗布泊野骆驼国家级自然保护区在研究区内的覆盖部分,不纳入载畜阈值与退化年限的模拟计算范围。
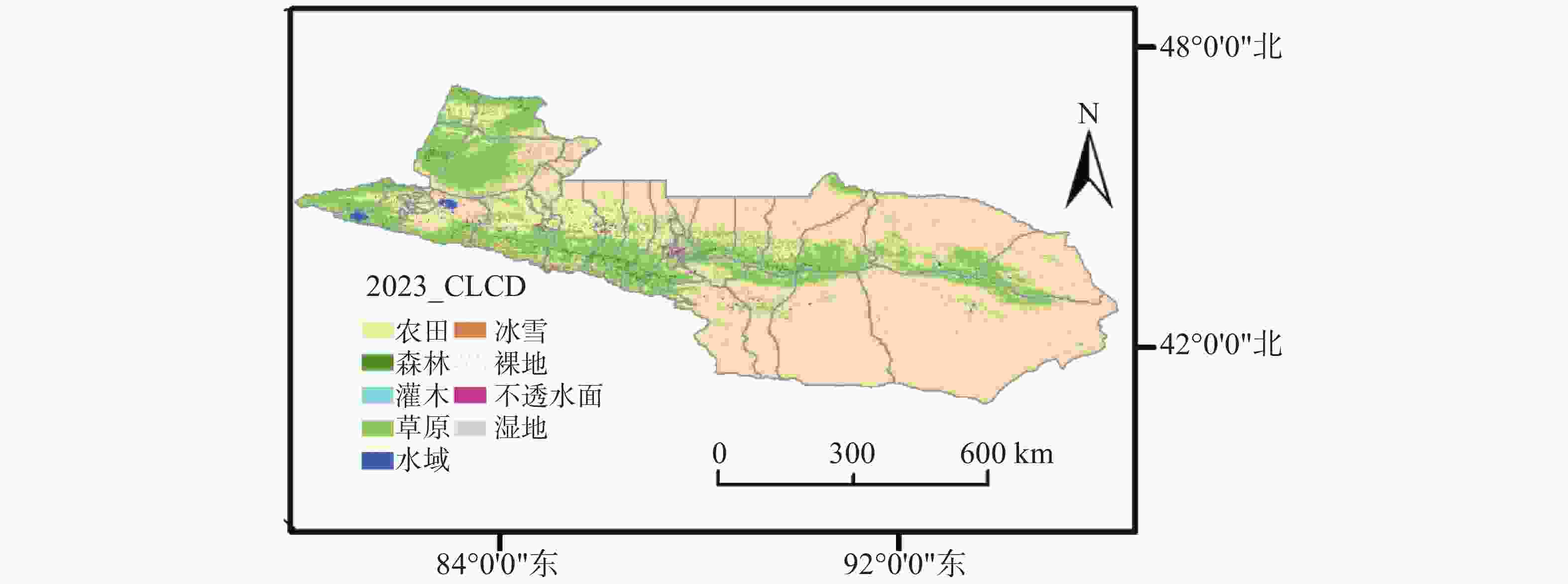
图 1 研究区域
Figure 1. Study area
新疆天山北坡地处亚洲中部干旱区,以其独特的地理环境和丰富的草地资源而闻名,是我国草地生态学和畜牧学研究的重要场所[15,16]。草地在此区域内发挥着至关重要的作用,不仅支撑着当地畜牧业的发展,还维系着生态平衡和生物多样性。该区域内草地类型多样[17],从高山草甸到山地草原,再到低地草原,形成了一个垂直分布的草地生态系统[15]。
-
本研究所用的数据包括遥感植被参数、气象再分析数据、实际载畜量数据及未来气候情景数据。植被参数选取 MODIS 叶面积指数产品 MOD15A2H V6.1[18],其空间分辨率为 500 m,时间分辨率为 8 d,时间范围从2005到2020 年。为保证数据质量,依据产品提供的质量控制层(FparLai_QC)剔除受云影响以及低质量像元。气象数据来源于欧洲中期天气预报中心(european centre for medium-range weather forecasts,ECMWF)发布的 ERA5-Land 气候再分析产品,该数据空间分辨率约为 0.1°,时间范围自 1950年至今。本研究提取其中 2005—2020 年的地表气温与向下短波辐射数据,为了匹配MODIS 产品时间分辨率,对逐日数据进行处理以获得 8 d时间尺度数据。随后,利用双线性插值法将气象数据重采样至 500 m 空间分辨率,实现与 LAI 产品的空间匹配。为确保数据质量与时序一致性,所有遥感数据均在 Google Earth Engine(GEE)平台上进行预处理,包括时间序列拼接、云层掩膜、异常值剔除及空间裁剪等步骤,从而保证输入数据的完整性与可靠性。本研究使用的实际载畜量数据来源于Wang等[2]构建的全球草地畜牧强度数据集(livestock grazing intensity, LHGI)。和现存的其他数据集相比,LHGI 数据集具备全国尺度的空间覆盖能力以及相对较高的空间分辨率,是目前中国范围内较为权威的草地载畜量数据。本研究选取 2020 年的数据作为载畜阈值模拟的初始输入,以反映当前放牧压力下草地系统的稳态水平。为分析未来气候变化情景下的载畜阈值变化趋势,引入 CMIP6 多模式集合数据集中的 RCP4.5 与 RCP8.5 两种情景[19],分别代表中等排放与高排放路径。提取未来气温与辐射数据并驱动光能利用率模型计算未受放牧影响的净初级生产力(NPP),为载畜阈值的情景模拟提供气候输入。
-
本研究以天山北坡草地为研究对象,基于未受放牧影响的草地净初级生产力与实际载畜构建了一个逐年递增放牧强度的过程模拟框架。在模拟过程中,每年增加 0.5 羊单位(SU·hm−2)的载畜压力为步长,逐年叠加放牧扰动,并动态更新草地可利用生产力,以反映放牧累积效应。当放牧导致草地净初级生产力下降至初始水平的 1% 时,认为草地生态系统达到临界退化状态,此时对应的载畜量即定义为载畜阈值。通过提取各像元的临界载畜量及对应的累积放牧年限,获得空间化的草地载畜阈值分布结果,为区域草地可持续利用与放牧强度调控提供科学依据。
-
为消除放牧干扰对草地生产力估算的影响,本研究基于作者此前提出的局部最大值回归算法拟合 MODIS LAI(MOD15A2H,500 m,8 d)时间序列,重建无放牧干扰条件下的理想 LAI 生长曲线。该方法通过自适应滑动窗口提取时间序列中的局部峰值,并采用局部加权回归(LOESS)对极值点进行平滑拟合,从而有效去除由放牧、气候波动及遥感噪声引起的异常波动,获得能代表草地自然生长过程的未受放牧影响的生长曲线。该算法已在相关研究中得到验证,能够较准确地反映草地在理想状态下的时序变化特征。主要包括3个步骤:1)提取LAI时间序列数据。本研究基于MODIS LAI产品(MOD15A2H),提取研究区内每个像元在每8 d时间间隔的LAI值,构建像元级的LAI时间序列。该时间序列代表了草地在放牧干扰下的“实际”生长情况,将用于与理想生长曲线进行对比分析,以便识别因放牧干扰导致的植被生长变化或覆盖度减少的区域。2)提取局部最大值点。采用滑动窗口方法,从LAI时间序列中识别局部最大值作为拟合曲线的关键点,用于表征草地在无放牧干扰条件下的理想生长轨迹。首先,根据草地物候特征,将全年划分为春季返青期、快速生长期和秋季枯黄期[20]。由于返青期和枯黄期的LAI变化较为平缓,使用较短的滑动窗口(3个时间步长)能够更灵敏地捕捉短期的上升或下降趋势;而在快速生长期,LAI变化剧烈,采用较长的窗口(6个时间步长)有助于抑制短期波动对极值识别的影响,提高准确性和稳定性。这种差异化的滑动窗口设置符合草地不同生长阶段的动态特征,同时参考了Cleveland[21]关于LOESS回归中窗口长度选择对趋势提取效果的研究:较短的窗口适合捕捉局部快速变化,但可能引入更多噪声,而较长的窗口则有助于平滑曲线,减少噪声干扰。基于此,本研究在三个生长阶段分别设置不同的滑动窗口长度,以便合理识别理想生长曲线的关键点。接着,采用滑动窗口遍历整个LAI时间序列,在每个窗口内识别局部最大值。在快速生长期,要求某一时间点在连续两个滑动窗口中均为最大值,才能视为有效局部极值,从而提高识别的准确性和稳定性。3)拟合LAI理想生长曲线。完成局部极值点提取后,本研究选取第一个时间点、所有局部最大值对应的时间点及最后一个时间点作为拟合节点,使用局部加权回归(local weighted regression,LOESS)算法对这些节点进行拟合,从而生成连续平滑的LAI理想生长曲线。该曲线模拟了草地在无放牧干扰条件下的生长趋势,作为后续分析放牧干扰和识别被啃食区域的重要依据。
在此基础上,本研究以理想 LAI 作为冠层结构参数输入,结合欧洲中期天气预报中心(european centre for medium-range weather forecasts,ECMWF)提供的 ERA5-Land 气候重分析数据[22],采用光能利用率(light use efficiency,LUE)模型估算未受放牧影响的净初级生产力(NPP)。ERA5–Land 数据包括逐小时的气温与向下短波辐射数据,经逐日平均后汇总为 8 d间隔,以匹配 MODIS 时序分辨率。向下短波辐射(SWrad, J·m-2)转换为光合有效辐射(PAR)的公式[23]:
$$ \mathrm{PAR}={\text{SW}}_{\text{rad}}\times {\mathrm{r}}_{\text{PA}{{\mathrm{R}}_{\text{SW}}}}\times \beta $$ (1) 式中,SWrad( J·m−2)为ERA5数据集中的向下短波辐射;rPAR_SW 取0.45 [24],为PAR与SW的比值[25-26],β取 4.56μmol·J−1,是能量-量子转换因子[27]。
总初级生产力GPP计算为叶绿素吸收光合有效辐射(absorbed photosynthetically active radiation, APAR )与光能利用率( LUE, εg)的乘积[28]。公式如下:
$$ \mathrm{GPP}=\mathrm{APAR}\times {\varepsilon }_{\mathrm{g}} $$ (2) $$ \mathrm{APAR}=\mathrm{PAR}\times \mathrm{fPAR} $$ (3) $$ \mathrm{fPAR}=1-{\mathrm{e}}^{-\mathrm{k}\times \text{Lai}}$$ (4) 式中,APAR为光合有效辐射与叶绿素吸收光合有效辐射的比值( fPAR)的乘积[29]。k为冠层消光系数,表示光在植被冠层中的衰减程度,本研究取0.5。该取值参考了 Zhang 等[30]的研究,该研究通过分析多 个陆地生态系统的数据,得出了草地类型的冠层消光系数 k 平均为 0.50。这一取值已广泛应用于草地生态系统中,表示光在草地冠层中的有效衰减。
光能利用率(εg )与空气温度( Tscalar )和水分( Wscalar )高度相关:
$$ {\varepsilon }_{\mathrm{g}}={\varepsilon }_{0}\times {\mathrm{W}}_{\text{scalar}}\times {\mathrm{T}}_{\text{scalar}} $$ (5) 其中,
$ {\varepsilon }_{0} $ 取0.53( g C mol −1 APAR)为表观量子产率或最大光能利用率[26]。Wscalar计算如下:$$ {\mathrm{W}}_{\text{scalar}}=\dfrac{1+\text{LSWI}}{1+{\text{LSWI}}_{\max }}$$ (6) 其中,LSWImax是一年中生长季内LSWI的最大值。Tscalar计算如下:
$$ {\mathrm{T}}_{\text{scalar}}=\dfrac{\left(\mathrm{T}-{\mathrm{T}}_{\max }\right)\times \left(\mathrm{T}-{\mathrm{T}}_{\min }\right)}{\left(\mathrm{T}-{\mathrm{T}}_{\max }\right)\times \left(\mathrm{T}-{\mathrm{T}}_{\min }\right)-{\left(\mathrm{T}-{\mathrm{T}}_{\text{opt}}\right)}^{2}}$$ (7) 其中,Tmax、Tmin和Topt分别是光合作用活动的最高、最低和最适温度,可以从基于生物组的查找表[31]中获得,本研究中Tmax取40°、Tmin取−1°、Topt取23°。
净初级生产力(NPP)通过 GPP 与碳分配系数 δ 计算得到:
$$ \mathrm{NPP}=\mathrm{GPP}\times \delta $$ (8) 其中,
$ \delta $ 的取值参考了目前发表的文献并经过多次实验验证后确定[32-33]。在此基础上,草地产草量计算公式[34]如下:
$$ {\mathrm{Y}}_{\mathrm{m}}=\frac{\text{NPP}}{\mathrm{t}\times \left(1+\mathrm{R}\right)}$$ (9) 其中,Ym为单位面积草地干草产草量(kg·hm−2),NPP 为草地净初级生产力,R为草地类型根冠比(表1),所有数据来源于文献中的相关研究部分[35-36]。t为生物量到生产力的转换系数,本研究中取值为 0.45[34],表明约 45% 的 NPP 被转化为草食性动物可利用的生物量。
表 1 不同草地类型的根冠比
Table 1. Root-to-shoot ratio of different grassland types
草地类型
Grassland types根冠比
Root/shoot文献
Reference温带禾草、杂类草草甸草原
Temperate grass-forb meadow steppe5.2 [35] 温带丛生禾草典型草原
Temperate needlegrass arid steppe5.6 [35] 温带丛生矮禾草、矮半灌木荒漠草原
Temperate dwarf needlegrass,dwarf semi-shrubby6.4 [35] 高寒禾草、苔草草原
Alpine grass,Carex steppe5.2 [35] 温带禾草、苔草及杂类草沼泽化草甸
Temperate grass,Carex-forbe swamp meadow7.9 [36] 高寒蒿草、杂类草草甸
Alpine Kobresia-forb meadow6.8 [35] 通过上述方法计算得到的未受放牧影响的 NPP 代表了草地在无放牧干扰条件下的最大生态生产潜力。该结果避免了放牧造成的系统性低估,为后续载畜阈值模拟与退化过程动态分析提供了统一的生态基准和空间参照。
-
本研究根据实际载畜量和草地可利用产草量,估算了牲畜对草地生物量的消耗。首先,假定牲畜的日消耗量与其体质量呈固定比例关系,即每日采食量约为体质量的 2%(rIT = 0.02)。考虑到草地生物量与碳含量的转换关系,本研究依据牲畜的碳需求量计算其生物量消耗需求(BioGrzt, kg C·m−2)[37]:
$$ \mathrm{BioGrzt}={\text{LS}}_{\mathrm{w}}\times {\mathrm{r}}_{\text{IT}}\times {\mathrm{C}}_{\text{coe}} $$ (10) 其中,LSw为家畜体质量;Ccoe为单位干物质向碳的转化系数(0.475)。
在放牧过程中,假设夏季放牧期内 95% 的生物量消耗来源于鲜草,5% 来源于枯草。当草地产草量(Ym)大于牲畜需求的 1.9 倍(即 BioGrzt × 0.95 × 2.0)时,认为牧草资源充足,此时牲畜的生物量消耗按 BioGrzt × 0.95 计算;当 Ym 小于 BioGrzt × 0.95 时,表明草地进入低生物量状态,在该阶段大量叶片和地上生物量需要保留以维持植被的基本代谢和再生能力,因此,可供牲畜实际采食的比例将显著低于总生物量。Zhu 等 [38]在草地退化早期预警与可利用生物量研究中指出,在接近退化阈值的低生物量条件下,牲畜可利用的生物量通常不超过总产量的约 20%。据此,本研究在该阶段取0.2×Ym作为可利用生物量。需要说明的是,该 20% 的比例用于表征退化阈值附近“可采食生物量显著受限”的生态状态,属于基于文献的经验参数,而非普适常数。在草地产草量处于中等水平(BioGrzt×0.95 < Ym < BioGrzt ×0.95×2.0)时,参考Shiyomi 等[37]引入指数函数对鲜草供给(BioGrzl,kg·m-2)进行调整,以更好地刻画草地在不同生产力条件下的供牧能力:
$$ {{{\text{BioGrz}}_{\mathrm{l}}}=\mathrm{Y}}_{\mathrm{m}}\times \left(0.08\times {\mathrm{e}}^{\left(1.8326\times \tfrac{{\mathrm{Y}}_{\mathrm{m}}}{{\text{BioGrz}}_{\mathrm{t}}\times 0.95\times 2.0}\right)}\right) $$ (11) 通过这种分段函数设置,本研究能够综合反映草地在不同产草量水平下的供牧能力,从而更真实地模拟牲畜对草地生物量的消耗过程。
-
在本研究中,本研究团队假设草地生产力会随载畜压力的增加而逐步降低,并以每年增加0.5 SU·hm−2的固定步长进行模拟。这一假设旨在揭示草地在长期持续放牧压力下的整体退化趋势,并为生态保护和草地管理提供早期预警参考。固定步长的递增加压方式是一种常见的生态系统压力测试思路,常用于在缺乏长期连续干扰观测数据的情况下[39],分析生态系统对外部扰动的响应边界和稳定性范围。相比于单一载畜水平的静态评估,这种沿放牧强度梯度推进的方式能够系统性地揭示不同压力水平下草地生产力的变化趋势,并避免因直接设定某一“假定合理”的载畜率而掩盖真实阈值所在区间。本研究选择 0.5 SU·hm−2 作为递增步长,主要基于阈值识别分辨率与计算效率之间的平衡考虑。若步长过大,可能跨越退化发生的关键区间,从而使阈值定位过于粗略;而过小的步长虽然可以进一步细化压力梯度,但会显著增加计算量,却难以在区域尺度上带来实质性的阈值精度提升。因此,0.5 SU·hm−2被作为一种在分辨率与可操作性之间折衷的压力扫描尺度,用于定位草地退化的载畜阈值区间。需要指出的是,这种逐年固定增量的模拟方式本身是一种简化。真实草地对放牧压力的响应可能呈现明显的非线性特征,例如,在低强度下具有一定的恢复和缓冲能力,而在高强度下可能出现突变式退化。因此,本研究的递增加压模拟旨在刻画长期趋势和阈值区间,而非精确再现每一阶段的实际退化速率。由此获得的退化年限和载畜阈值应理解为在该压力测试框架下的相对指标,可用于比较不同区域和情景下草地对放牧压力的敏感性与承载潜力,而非对现实放牧路径的直接预测。
本研究假设草地生产力会随载畜压力的增加而逐步降低。在模拟过程中,草地被牲畜消耗的净初级生产力(NPPGrz)由以下公式:
NPP为草地净初级生产力( kg·m - 2),αleaf为总光合产物向叶片的分配比例,BioGrzl为鲜草可提供的生物量。该公式综合反映了草地自身生产力和牲畜采食需求的双重约束[38]。
$$ {\text{NPP}}_{\text{Grz}}=\mathrm{min}\left(\mathrm{NPP}\times {\alpha }_{\text{leaf}},{\text{BioGrz}}_{\mathrm{l}}\right) $$ (12) 本研究以草地未受放牧干扰条件下的未受放牧影响的净初级生产力(NPP)中可供利用的部分作为核心生态指标,构建了像元尺度的持续扰动模拟框架。该未受放牧影响的 NPP 由理想 LAI 拟合得到,反映草地在自然生长状态下的最大生产潜力,可作为评估放牧干扰影响的基准。在模拟过程中,逐步累积放牧压力,并在每一阶段动态更新草地的剩余生产力。当某一像元的实际可利用 NPP 降低至基准水平(2020 年未受放牧影响的 NPP)的 1% 时,判定其进入不可逆退化状态[38],对应的载畜量即界定为该像元的载畜阈值。这一阈值的选择基于生态学理论,表明当 NPP 下降至初始水平的1%时,生态系统的恢复能力已显著降低,可能进入不可逆退化阶段,导致功能性丧失。因此,选择1%作为退化临界点,能够有效反映草地退化的临界状态,并为放牧管理提供警示。对应的载畜量即界定为该像元的载畜阈值。与传统依赖静态经验参数设定的方法相比,本研究以 2020 年实测承载水平为起点,构建逐年增强的放牧情景,系统揭示了长期载畜压力下草地生产力的动态衰退过程。通过这一框架,不仅能够识别不同区域草地达到退化临界点所需的时间尺度,还能量化其最大可承载的载畜水平,从而更全面地刻画草地系统的放牧耐受极限。
-
基于理想 LAI 的拟合结果,并结合光能利用率模型计算得到,天山北坡草地在无放牧干扰条件下的未受放牧影响的净初级生产力(NPP)空间分布如图2所示。结果表明,未受放牧影响的 NPP 呈现出显著的空间梯度特征,整体表现为西部高于东部,山地显著高于平原和荒漠区域。
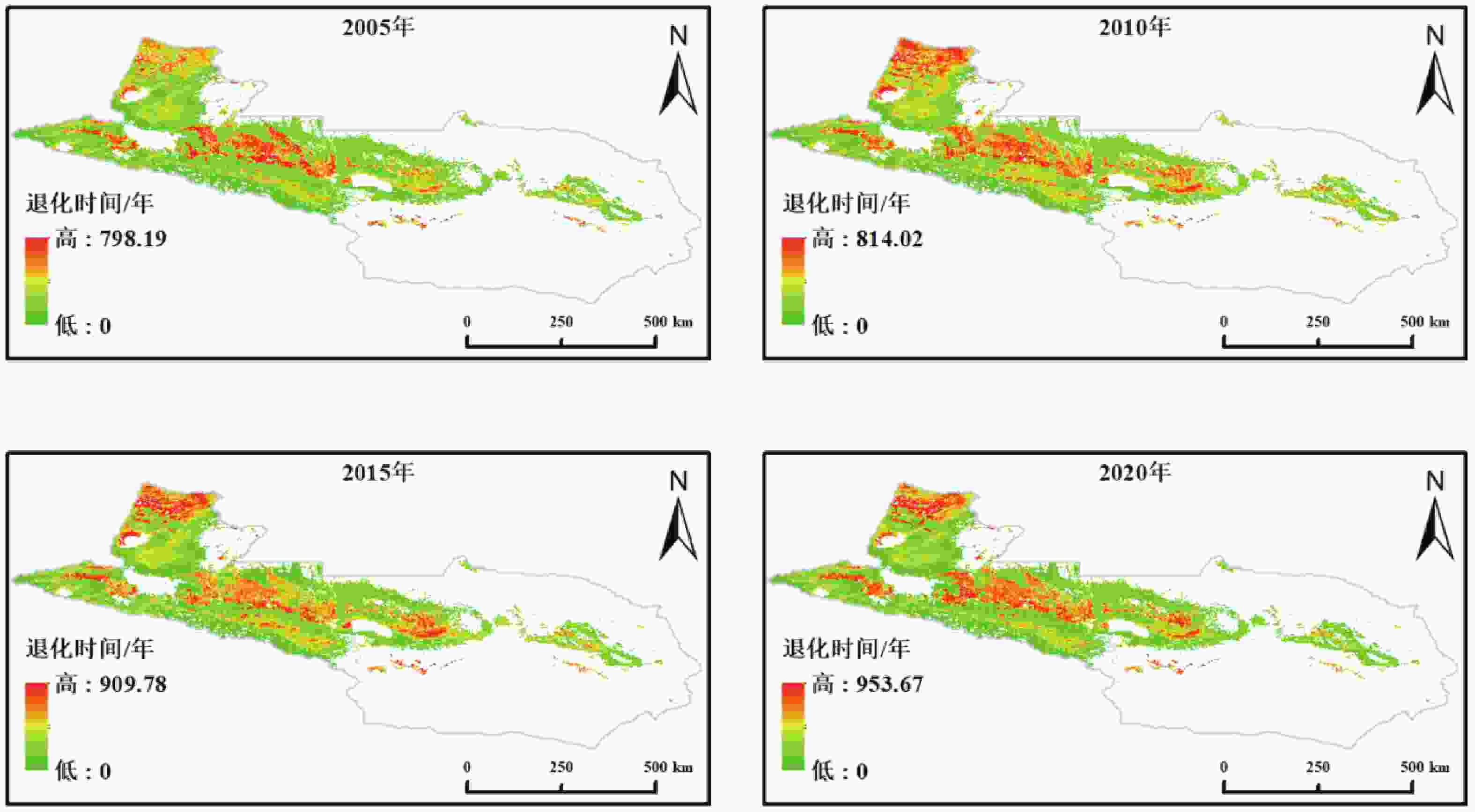
图 2 天山北坡草地未受放牧影响的 NPP 空间分布图
Figure 2. Spatial distribution of ungrazed NPP of grasslands on the northern Tianshan Mountains
在空间分布上,未受放牧影响的 NPP 高值区主要集中于天山中段和伊犁河谷地区,普遍超过 500 g·m−2·a−1,局部草甸草原可达 800 g·m−2·a−1以上。这些区域以高寒草甸和山地草甸为主,受益于高海拔地形带来的充沛降水和适宜热量,植被结构复杂、光合效率高,生产潜力显著[5]。中值区分布于山前草原和丘陵过渡带,未受放牧影响的 NPP 多在 200~400 g·m−2·a−1之间,反映出中等水平的供牧能力。这类区域虽然具备一定的生物量积累潜力,但受限于水热条件和土壤养分,其整体生态耐受性相对有限。低值区主要分布于干旱荒漠草原和低海拔东部地区,未受放牧影响的 NPP 普遍低于 100 g·m−2·a−1,说明水分短缺是制约草地生产力的关键因素。这些生态系统即使在理想条件下也难以形成较高的生产力水平,表现出较强的生态脆弱性。不同草地类型之间也存在显著差异。草甸草原和高寒草甸的未受放牧影响的 NPP 显著高于典型草原和荒漠草原,表明其在自然状态下具有更强的生物量积累与放牧承载潜力。而荒漠草原即便处于无放牧干扰条件下,其未受放牧影响的生产力依然处于较低水平,凸显了该类生态系统的固有脆弱性以及对载畜压力的敏感性。
从时间变化的角度来看,天山北坡草地未受放牧影响的 NPP 在 2005—2020 年间保持了整体“西高东低、山地高于平原”的稳定格局,但不同年份间仍存在一定波动。2005 年未受放牧影响的 NPP 整体水平较高,伊犁河谷与天山中段草甸草原区域形成连续的高值带,部分像元超过 700 g·m−2·a−1。到 2010 年,高值区范围有所收缩,尤其是在东部和部分丘陵过渡带,未受放牧影响的 NPP 较 2005 年出现下降,区域生产力差异加大。2015 年未受放牧影响的 NPP 再度增强,高值区在天山中段重新扩展,但分布呈现出斑块化特征,显示草地生产力在不同生态区间的差异进一步加剧。至 2020 年,整体未受放牧影响的 NPP 水平趋于均衡,整体格局与 2010 年相近,高值区较 2015 年略有减弱,但区域差异性依旧显著。
总之,未受放牧影响的 NPP 的时空格局揭示了天山北坡草地在自然条件下的生产潜力分异,同时反映了水热条件、植被类型及年际气候波动对区域生产力的共同调控作用。基于理想 LAI 所建立的 NPP 基准为后续载畜阈值的过程化模拟奠定了坚实基础,有助于进一步刻画草地在长期载畜压力累积下的退化发生时间尺度及其对应的临界承载水平,从而更全面地认识不同区域草地系统的耐受极限。
-
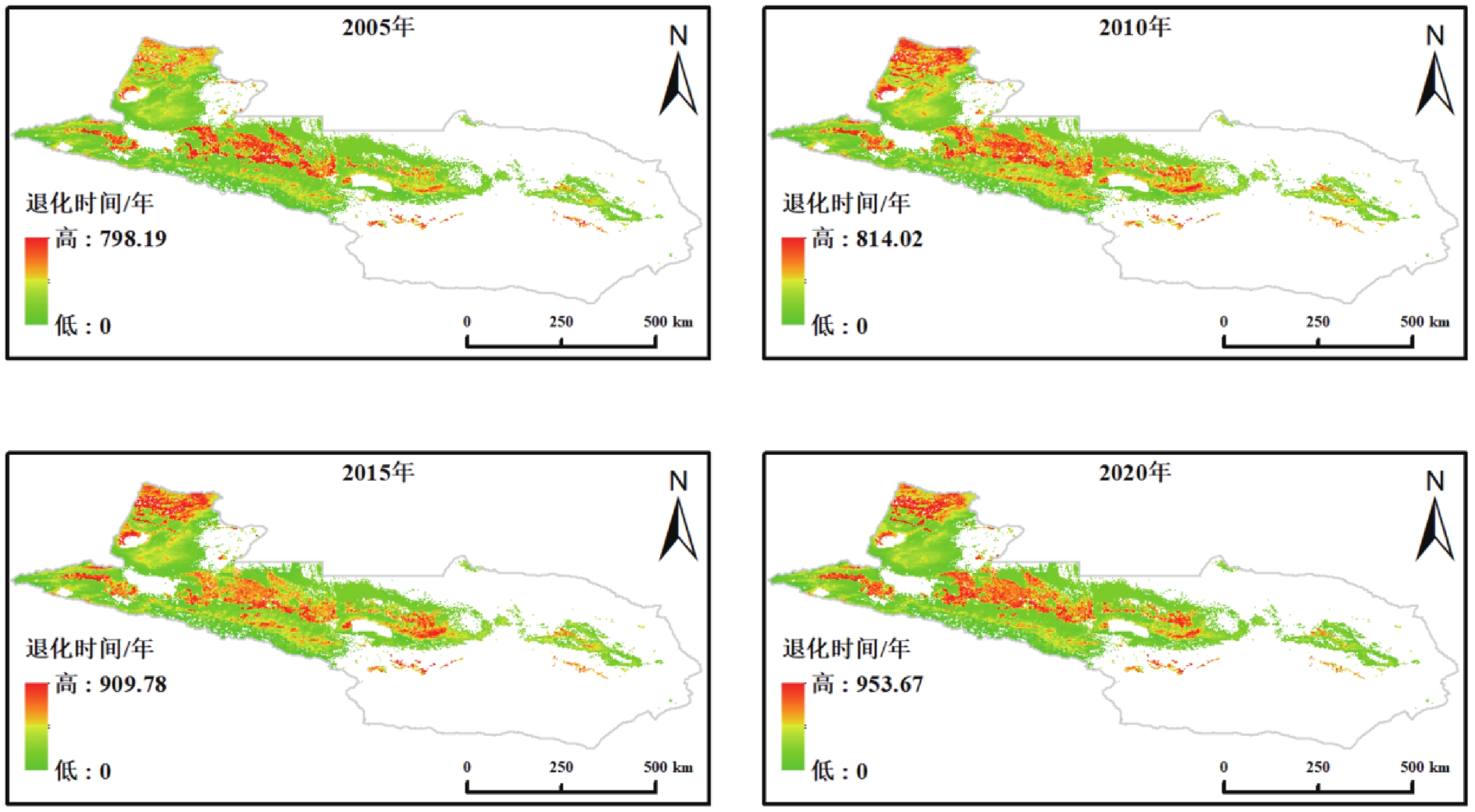
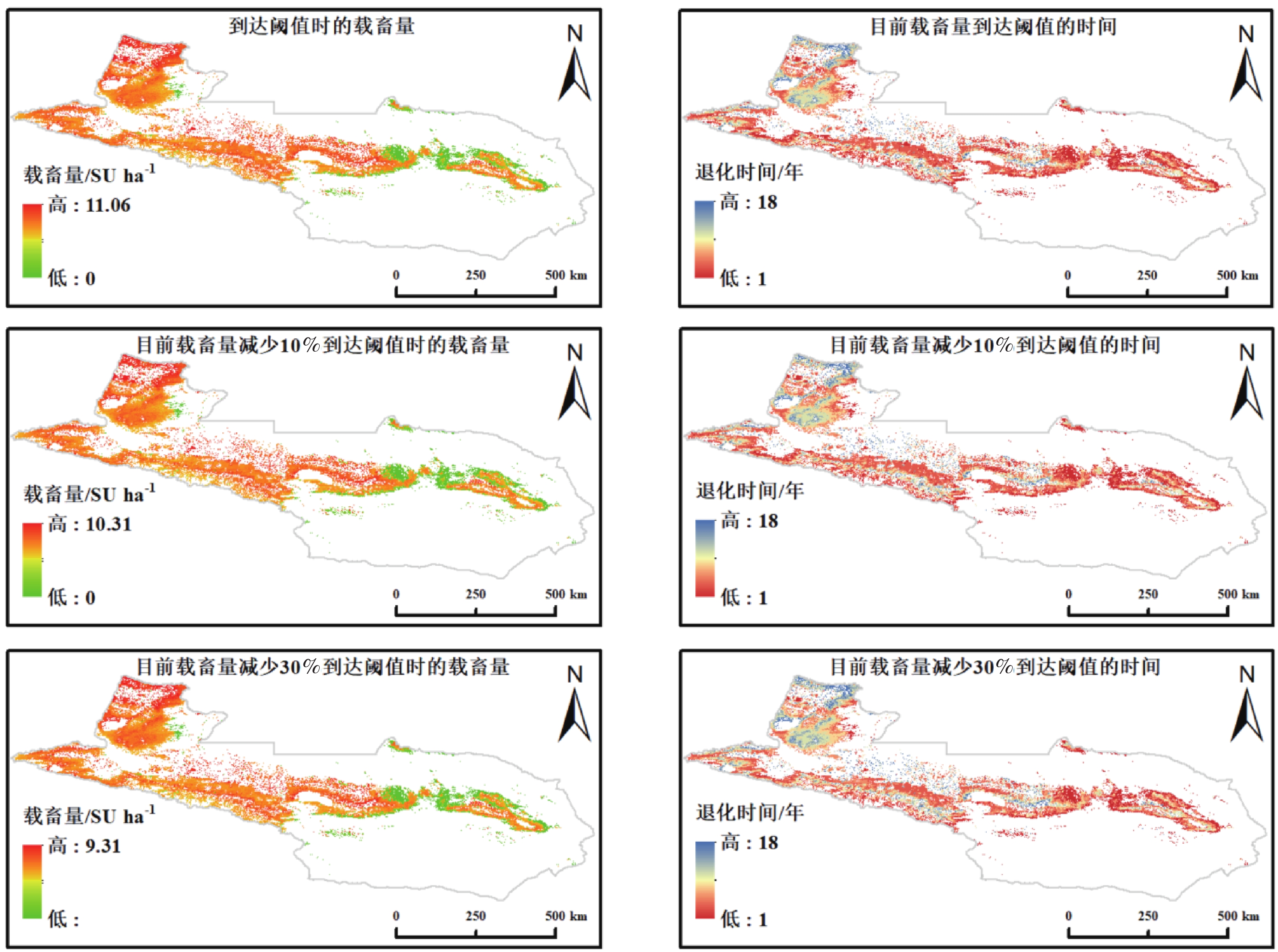
本研究以 2020 年的实际载畜量为起点,构建了逐年递增放牧压力的动态模拟框架,得到草地退化年数和临界载畜量的空间分布格局(图3)。模拟结果显示,草地退化过程在时空上均表现出显著差异,不同区域的耐受性与承载能力存在明显差异。
图 3 天山北坡草地载畜阈值及退化时间空间分布
Figure 3. Spatial distribution of grazing thresholds and degradation time of grasslands on the northern Tianshan Mountains
模拟结果表明,天山北坡草地在不同放牧强度情景下的载畜阈值与退化时间均呈现显著的空间分异特征。整体上,临界载畜量范围为 0~11.06 SU·hm−2,退化年限分布于 1~18 a之间,部分区域的临界载畜量已降至 0,表明这些草地目前处于超载或退化状态。在不减少放牧情景下,临界载畜量介于 0~11.06 SU·hm−2,空间格局呈现“西高东低、山地高于平原”的特征。高值区主要分布在伊犁河谷及天山中段的草甸草原和高寒草甸地带,反映出其较高的未受放牧影响的生产力与生态承载潜力;低值区集中于东部荒漠草原及部分典型草原,受限于水热条件与土壤肥力,其生态系统易在较短时间内接近临界状态。对应退化年限整体分布于 1~18 a之间,高值区集中于高寒草甸与山地草甸,可在较高载畜压力下维持较长的生态稳定期;而荒漠草原及低海拔地带退化年限普遍较短,部分区域在 1~3 a内即可能出现明显退化,表明这些区域在现有放牧强度下已接近生态极限。
在载畜量减少10%的情景下,退化年限略有提升。草地平均退化年限由当前情景下的9.25 a略微增加至9.36 a。当载畜量减少30%时,平均退化年限进一步增加至9.61 a,较当前情景延长0.36 a。然而,进一步统计发现,在减牧10%和减牧30%两种情景下,均不存在退化年限延长超过5 a的像元,表明减牧对退化时间的影响整体较为有限。
-
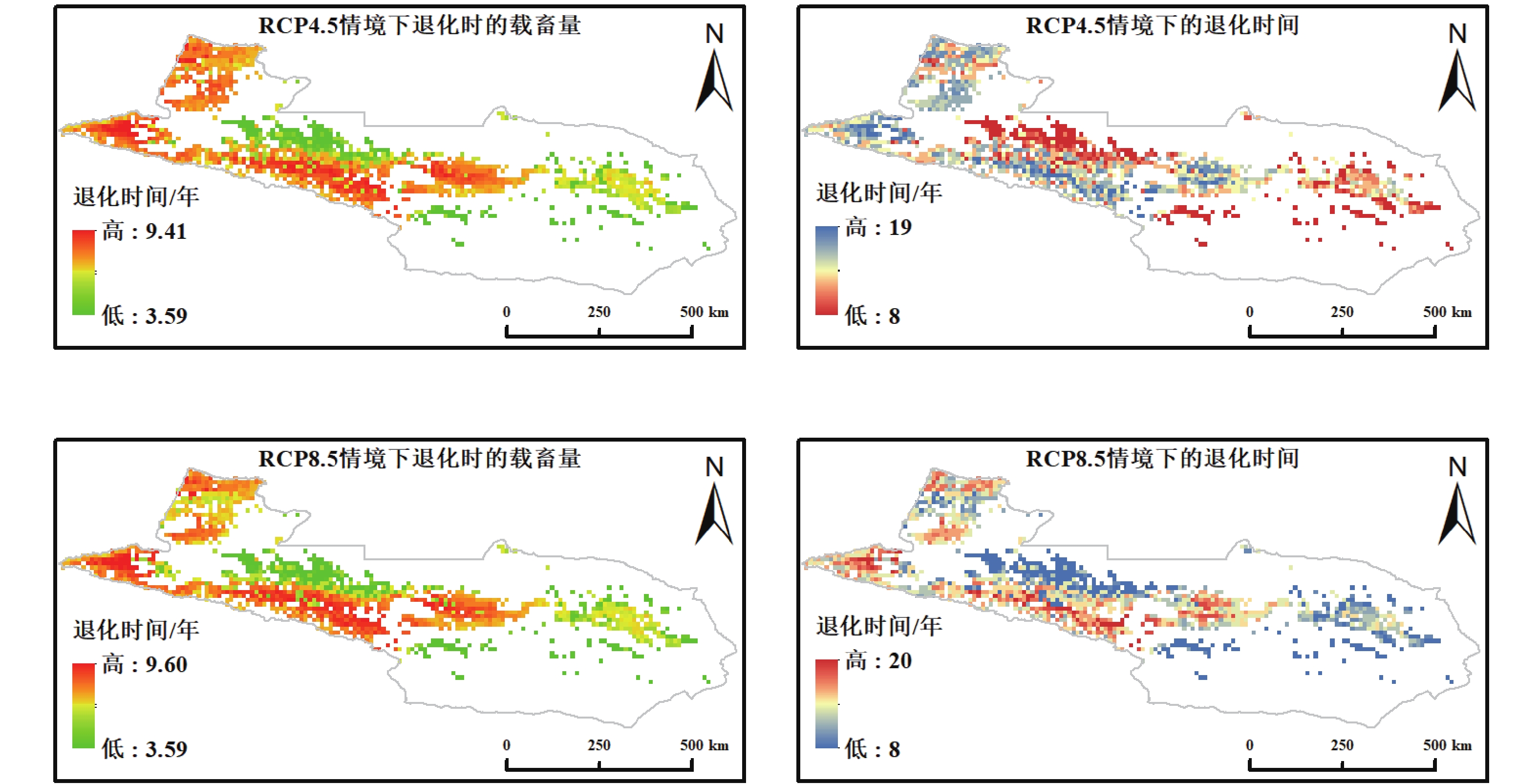
基于 RCP4.5 与 RCP8.5 两种气候情景输入的未受放牧影响的 NPP,本研究对天山北坡草地的临界载畜量与达到退化阈值所需年数进行了空间化模拟(图4)。两种气候情景下临界载畜量范围均较当前情景有所降低,其中 RCP4.5 情景下为 3.49~9.41 SU·hm−2,RCP8.5 情景下为 3.49~9.60 SU·hm−2。退化年限整体分布于 8~20 a 之间,明显高于当前载畜情景的 1~18 a,表明在气候变暖背景下草地的未受放牧影响的恢复期有所延长。
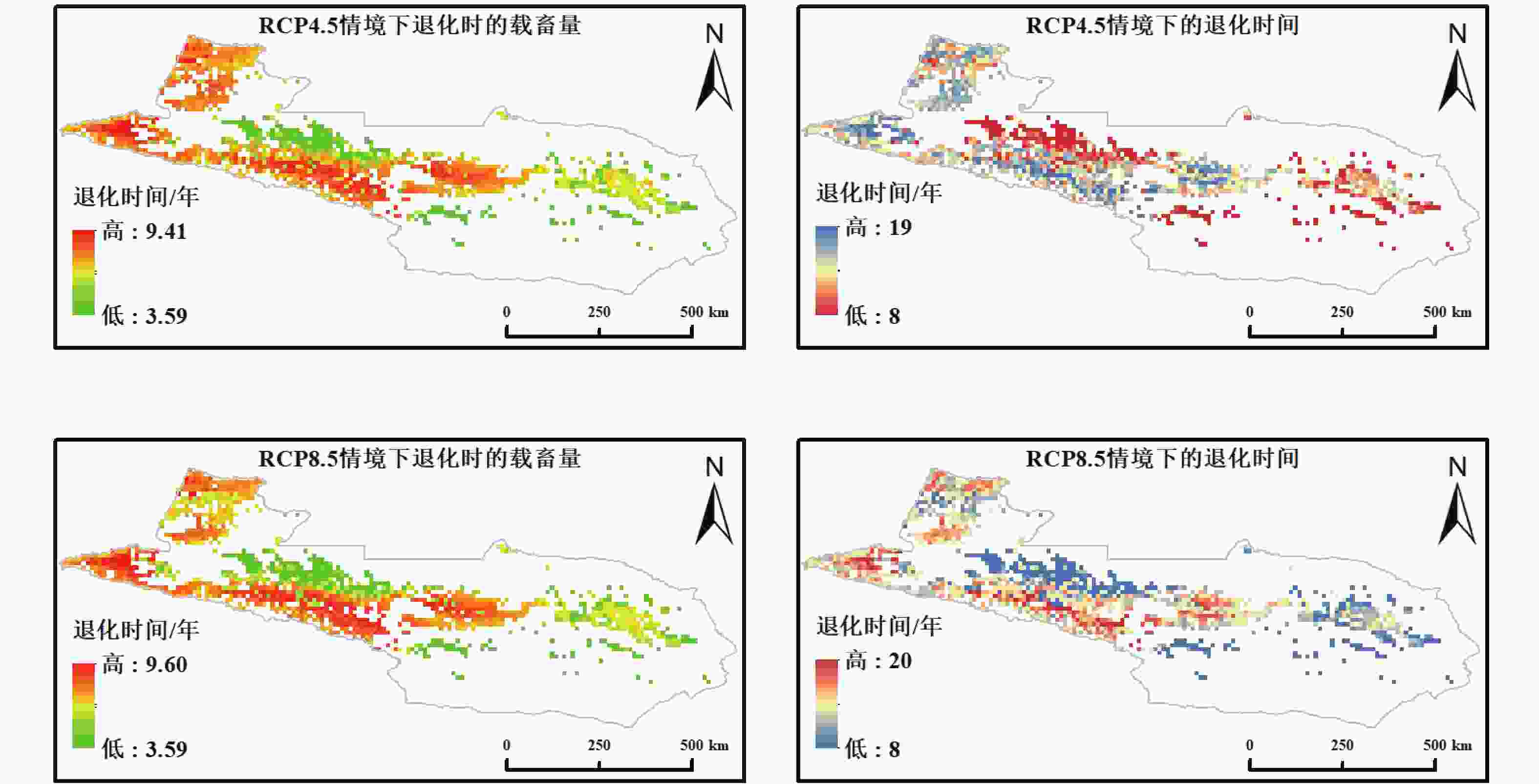
图 4 未来气候情景下天山北坡草地载畜阈值及退化时间空间格局
Figure 4. Spatial patterns of grazing thresholds and degradation time of grasslands on the northern Tianshan Mountains under future climate scenarios
在未来气候情景下,RCP4.5 与 RCP8.5 的载畜阈值和退化年限总体差异有限,甚至在部分区域,RCP8.5 下的退化年限略长于 RCP4.5。其主要原因可能有以下几点:一方面,RCP8.5 高排放情景下大气 CO2 浓度显著上升,增强了植被的光合作用与固碳能力,形成 “CO2 施肥效应”。该效应在水热条件适宜的高寒草甸与山地草甸区表现较为突出,使这些区域在高排放条件下仍能维持较高的未受放牧影响的生产力,从而延长退化时间。另一方面,RCP8.5 下的气候变化虽带来更大的温度升高和未受放牧影响的蒸散增加,但在天山北坡区域,局部地区降水增加与温度升高共同作用,延长了生长季并且改善了水热匹配。这种气候补偿效应在一定程度上缓解了高放牧压力下 NPP 的衰减;此外,本研究采用的生态退化的判定标准,强调的是系统达到极限时的长期累积效应。在这种判定框架下,气候情景之间的差异可能被部分削弱,导致 RCP4.5 与 RCP8.5 的模拟结果相近。
高排放情景并未显著降低草地承载力,反而在局部区域表现出较长的退化年限,说明未来草地生态系统对气候变化的响应具有复杂性和非线性特征。这一现象可能与高排放下增强的 CO2 施肥效应与气候补偿效应有关,例如在藏高原地区的遥感承载力估算中发现,未来情景下气候改善使部分区域载畜能力略有提升[40];在新疆干温带草原,尽管放牧强度加大,CO2 与气候改善在一定程度上减弱了放牧对固碳的负面影响[41]。本研究所采用的光能利用率(LUE)模型虽未将大气 CO2 浓度作为输入变量,但其原始参数化过程中已隐含了 CO2 对植被光合效率的平均响应,并通过辐射、温度和水分条件等气候驱动因子的变化在情景模拟中得到间接体现。由于该模型未针对天山北坡草地进行区域化再校准,在高 CO2 条件下生产力增强的幅度仍存在一定不确定性,这可能对 RCP8.5 情景下退化年限略长的模拟结果产生一定影响。
-
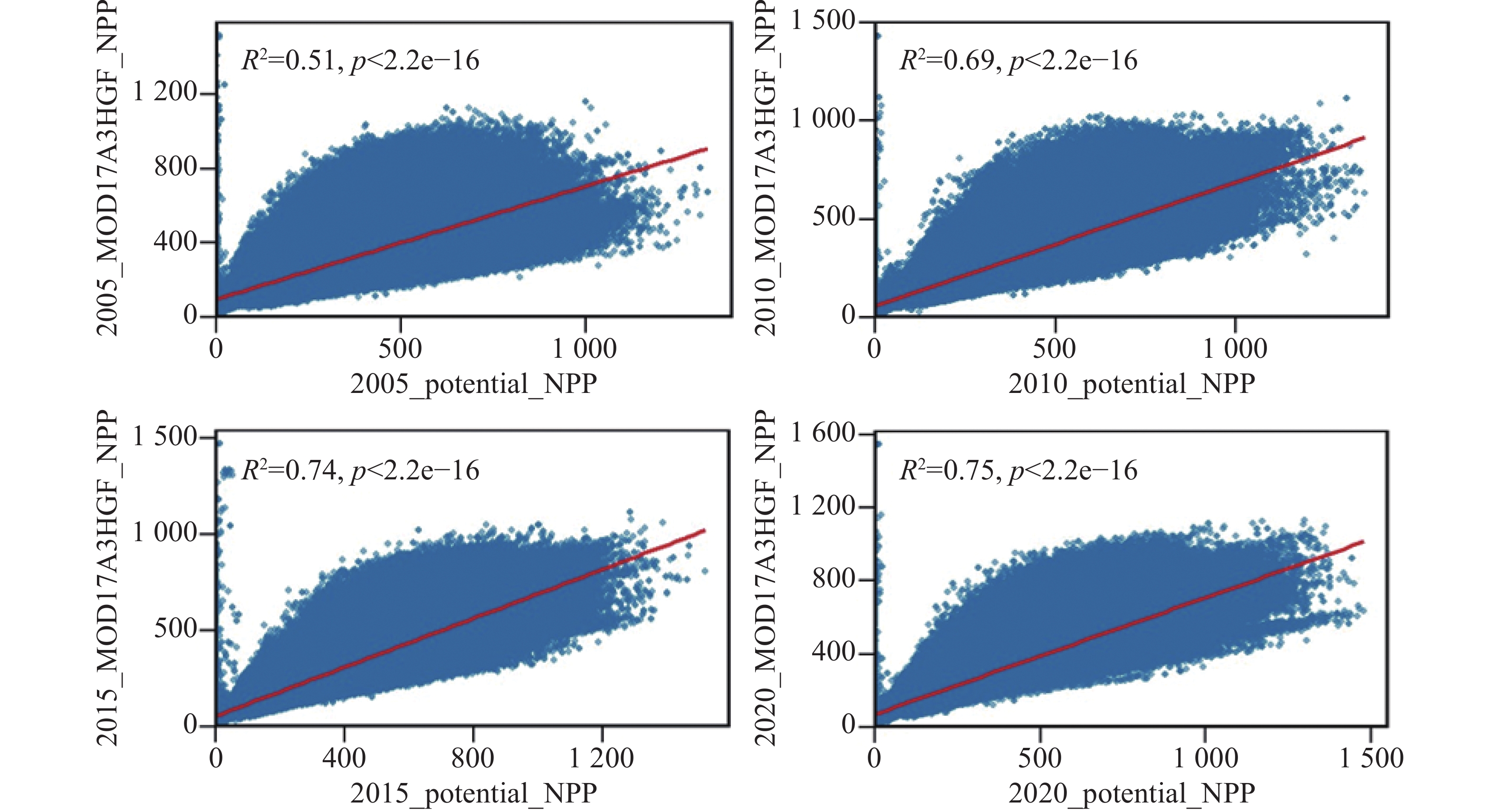
为验证基于理想叶面积指数(LAI)构建的未受放牧影响的净初级生产力(NPP)模拟结果的可靠性,本研究选取2005、2010、2015和2020年4个典型年份,与MOD17A3HGF NPP产品进行像元级对比分析(图5)。结果表明,两者在空间分布与量值变化上总体一致,不同年份的拟合结果均表现出显著正相关关系。未受放牧影响的NPP在空间上与实际NPP表现出良好的耦合关系,验证了本研究方法在区域尺度上表征草地未受放牧影响的生产力的可行性与稳定性。从统计结果来看,未受放牧影响的NPP与MOD17A3HGF NPP之间的决定系数(R2)在2005年为0.51、2010年为0.69、2015年为0.74、2020年为0.75(P<0.01),说明两者在不同年份间保持较高的一致性与稳定性。整体上,未受放牧影响的NPP能够较好地反映草地生产力的空间分异格局与时序变化特征。
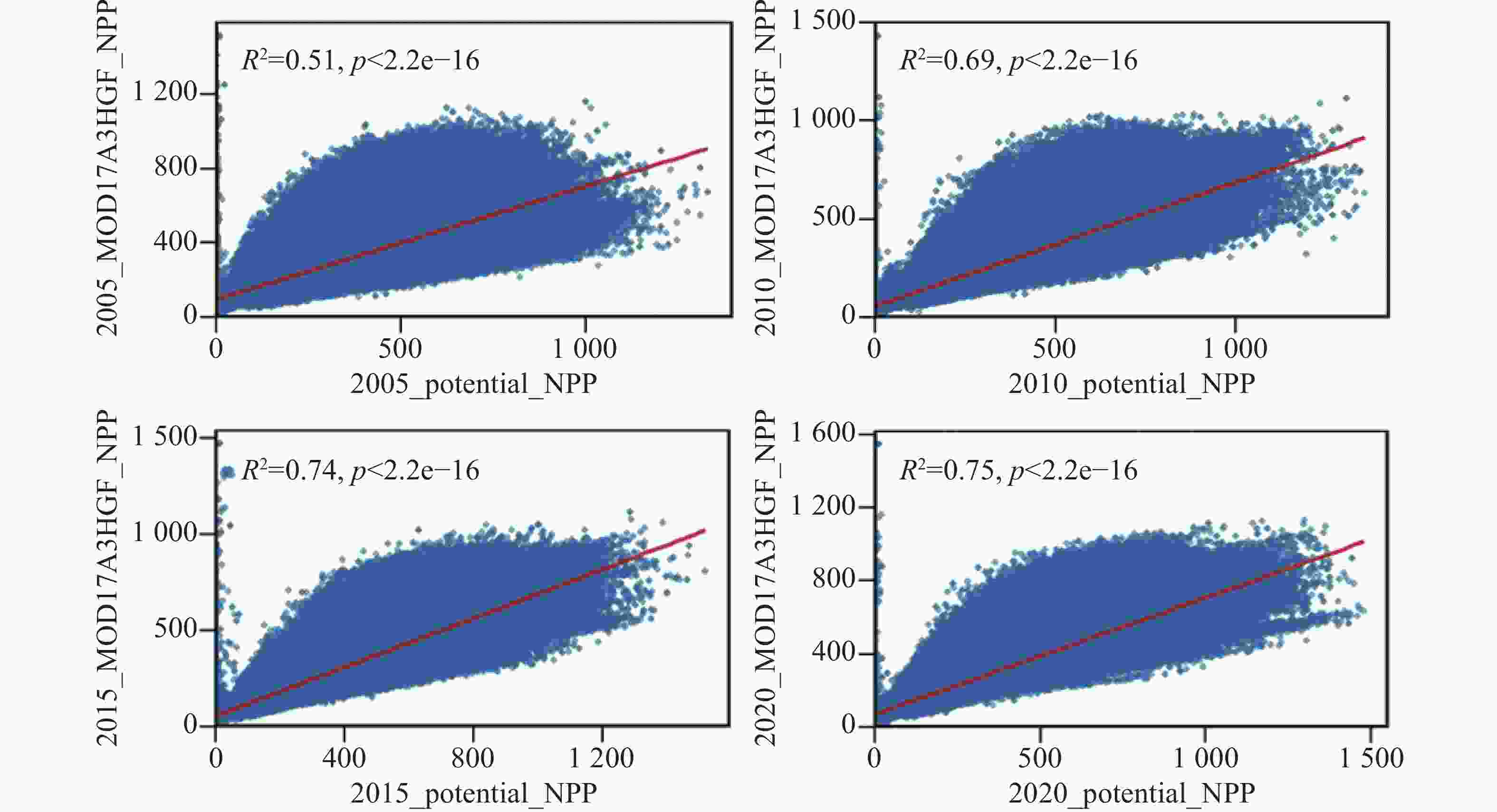
图 5 基于理想LAI构建的未受放牧影响的NPP与MOD17A3HGF NPP的验证
Figure 5. Pixel-level comparison between ungrazed NPP simulated based on ideal LAI and MOD17A3HGF NPP
由于本研究计算的为“未受放牧影响的NPP”,即假设无放牧干扰条件下的草地生产力,而MOD17A3HGF反映的是受放牧、人类干扰及气候约束下的“实际NPP”,二者在生态意义上并非完全对应[42]。因此,部分年份(尤其是2005年)出现较低相关性(R2 = 0.51),可能源于放牧活动导致的NPP下降,使得实际观测值系统性低于未受放牧影响的模拟值。此外,早期MODIS产品存在辐射校准和云掩膜不稳定性问题,以及ERA5-Land气象数据在高山地形区的温度和辐射插值误差,也共同影响了模拟精度。
-
为了量化模型结果的不确定性,本研究团队对关键参数的变动进行了敏感性分析,测试了 LAI拟合算法的参数(如窗口大小、平滑因子等)、LUE模型中的εmax、Topt 等参数在合理范围内的变化,并分析了它们对 “载畜阈值” 和 “退化年限” 空间格局及数值范围的影响程度。通过这些变动的测试,结果表明,LAI拟合算法中的参数(如滑动窗口大小和平滑因子)对 “载畜阈值”和“退化年限”的影响较小,主要体现在空间格局的细微变化。即使在不同的参数设置下,整体趋势保持一致,主要影响局部的细节与变化,但未显著改变模型结果的精度。这表明LAI拟合算法在模型中的角色更偏向于平滑局部数据波动,对于整体模型结构和大范围预测影响有限。相比之下,LUE模型中的关键参数(如最大光能利用率和最适温度)对模拟结果的影响较大,尤其在极端气候条件下。这些参数的波动可能导致GPP和NPP的显著变化,进而影响 “载畜阈值”和“退化年限”的预测结果。在高温和干旱等极端气候下,温度和水分的双重影响可能会放大误差,导致模型预测的不确定性增大。因此,为了提高模型的准确性和鲁棒性,未来的工作应考虑对极端气候条件下的参数敏感性进行更深入的优化,尤其是在应对极端温度和干旱情境时,调节关键参数以减少预测误差。通过敏感性分析,我们明确了模型结果的敏感性来源,为进一步提高模型的可靠性和准确性提供了依据。
未来情景模拟的结果说明在当前气候背景和初始植被状态下,草地退化过程呈现明显的阈值控制特征:适度减少放牧虽可在一定程度上延缓退化发生时间,但难以从根本上改变跨越退化阈值的时间尺度。空间上,高寒草甸和山地草甸对减牧的响应相对更明显,而荒漠草原则受限于水分条件,其恢复潜力依旧有限。综合不同情景的模拟结果表明,天山北坡草地在载畜阈值和退化年限上呈现明显的草地类型差异。草甸草原和高寒草甸因未受放牧影响的生产力较高,退化年数较长,临界载畜量水平显著高于典型草原和荒漠草原,是区域草地可持续利用的关键优势区。而荒漠草原和部分典型草原即使在减少放牧情景下,其退化年数仍相对较短,临界载畜量水平提升有限,显示出较高的生态脆弱性和退化风险。在不同管理方案中,减少放牧能够有效延缓退化进程并提升临界承载力,其中 30% 减少放牧情景在延长退化年限和提高临界水平方面效果最为显著,说明通过阈值模拟可为区域草地的差异化放牧管理与可持续利用提供科学依据。
需要指出的是,在 RCP8.5 情景下退化年限略长于 RCP4.5,主要可归因于 CO2 施肥效应与气候补偿效应共同作用。虽然本研究采用的 LUE 模型未将大气 CO2 浓度作为输入变量,但 CO2 升高对植被生产力的影响已通过辐射、温度等气候驱动因子的变化在模型中得到间接体现。本研究尚未针对“Ym<BioGrzt × 0.95 时可利用比例取 20%”这一分段参数进行专门的敏感性测试。该参数用于表征草地在接近退化阈值时可供牲畜采食生物量的显著下降,其取值依据 Zhu等[38] 中对低生物量草地可利用比例的经验范围。不同草地类型和放牧方式下,该比例可能存在一定变动,这将对阈值和退化年限的绝对数值产生影响,但对不同情景之间的相对比较影响预计较小。未来可通过改变该比例(如 10%~40%)开展系统敏感性分析,以进一步评估模型稳健性。
-
本研究基于未受放牧影响的 NPP 基准与过程化载畜阈值模拟,系统揭示了天山北坡草地的生态耐受性特征及空间差异,反映出不同气候与地形条件下草地对放牧干扰的响应机制与生态调节能力。结果表明,该方法能够较为有效地识别草地生产潜力与实际承载力之间的偏差,为干旱半干旱区载畜阈值的定量模拟提供了新的技术路径与理论参考。但是该研究仍存在一定的局限性,在模型验证、放牧过程刻画、气候情景设定及生态过程表征等方面尚有改进空间。
模型验证仍存在一定不足。尽管本研究通过将模拟得到的未受放牧影响的 NPP 与 MODIS NPP(MOD17A3HGF)产品进行对比,验证了未受放牧影响的生产力模拟结果的合理性,且二者在空间分布上表现出较高一致性,但其验证仍面临方法学上的挑战。由于未受放牧影响的 NPP 表征的是无放牧干扰下的生态系统理论生产上限,缺乏与之直接对应的实测样本,传统的与实测或通量塔观测数据对比的方法难以全面评估其可靠性[43,44]。这种验证方法可能导致模型对 NPP 预测的某些偏差,尤其是在干旱半干旱地区,这些地区的植被生长受限因素较多,模型可能高估了某些区域的生产潜力,从而低估了实际的载畜阈值。
放牧过程的复杂性未被充分刻画。本研究在模拟中假设放牧压力均匀分布,未能充分考虑牲畜的采食偏好、移动行为及不同放牧制度(如转场放牧、季节性轮牧、围栏管理等)对草地退化进程的影响[45]。而这些因素在实际草地利用中往往起到关键作用,可能导致局部地区的退化速度和承载阈值与模型结果存在差异。在假设放牧压力在像元尺度上相对均匀分布的前提下,模型难以刻画牲畜在草种和小尺度斑块上的选择性采食行为。现实中,牲畜往往优先采食营养价值较高、适口性较好的草种,这会使这些斑块的生物量下降速度快于周围区域,从而加速局部退化过程。由于本研究基于 500 m 像元尺度的 LAI 变化来识别放牧影响,这种细尺度的选择性采食效应可能被空间平均所平滑,进而在这些区域倾向于高估其可承载的放牧强度。与此同时,在放牧活动高度集中但空间范围较小的区域(如优势草种斑块或放牧热点),其退化信号可能被周围未受影响区域稀释,从而使模型低估这些区域的实际放牧压力与载畜阈值,尤其在草地类型复杂、空间异质性较强的地区,这种不确定性会更加明显。因此,未来研究有必要将牧场管理措施与牲畜行为过程纳入阈值模拟框架,以提高其适用性和解释力。
气候情景分析仍存在简化。在未来情景评估中,本文主要基于 RCP4.5 和 RCP8.5 情景下的平均 NPP 数据推演载畜阈值。该方法能够反映气候变暖趋势下的整体响应,但难以捕捉极端气候事件(如持续干旱、暴雪灾害)对草地系统的剧烈干扰[45]。由于极端事件在干旱半干旱区发生频率较高,其对生态系统的冲击可能远大于平均气候变化。在干旱半干旱地区,持续干旱等极端气候事件可能导致草地暂时无法恢复,草地承载力急剧下降。若模型未能考虑这些极端气候的影响,可能会高估草地的生产潜力,并因此高估载畜阈值。极端气候事件可能在较短时间内对草地生态造成严重影响,导致草地生产力的剧烈波动。由于本研究采用的情景无法模拟这些波动,可能使得在极端气候条件下的载畜阈值被低估,从而影响放牧管理策略的合理性。另外,当前的放牧压力假设保持不变,未考虑未来由于政策、市场、人口等因素可能导致的系统性变化。这可能导致本研究的载畜阈值模拟与实际情况产生偏差。未来研究应结合社会经济因素和放牧压力的变化,以及极端气候事件模拟和灾害风险评估,进一步改进模型,以全面刻画气候变化背景下的阈值动态。
生态过程的复杂性未被完全纳入。本研究以 NPP 为核心指标来衡量草地生产力及其承载力,但草地退化过程还受到土壤水分[46]、养分循环和植被群落更新等多方面的影响。例如,土壤压实、养分流失或植被群落退化等过程可能在较早阶段限制草地承载力[47],目前的模型中未纳入这些过程,可能导致部分草地的生产潜力和承载力被高估,进而低估其载畜阈值。这些过程可能在草地退化早期就已经显现,加剧了退化进程,而模型未能充分考虑这些因素,导致一些草地退化过程被低估,影响了对载畜阈值和退化年限的准确估算。未来若能结合过程模型(如 BIOME-BGC、CENTURY)或土壤与群落数据,将有助于更全面地解释草地系统的长期演变。
本研究中牲畜可利用生物量的分段比例参数(如低生物量阶段取 20% × Ym)来源于已有文献对退化阈值附近草地采食行为的经验总结[38],用于刻画草地在接近退化状态时“保留生物量以维持植被再生”的生态约束。然而,不同草地类型、植被结构和放牧制度下,该比例可能存在差异。本研究未对该参数进行系统的敏感性分析,因此阈值和退化年限的绝对值仍存在一定不确定性。未来可结合样地调查或多参数扰动实验,对该比例进行优化与校准。
-
本研究基于理想 LAI 拟合构建了天山北坡草地在无放牧干扰条件下的未受放牧影响的生长曲线,并结合光能利用率模型估算未受放牧影响的净初级生产力(NPP),据此建立逐步递增放牧强度的动态模拟框架,系统评估草地退化年限与载畜阈值的时空格局。研究同时结合实际载畜量、减少放牧方案及未来气候情景,揭示了草地系统的生态耐受极限与调控潜力。主要结论如下:
1)天山北坡草地未受放牧影响的 NPP 呈“西高东低、山地高于平原”的空间格局,高寒草甸与草甸草原的生产潜力显著高于典型草原与荒漠草原,表明水热条件与植被类型是控制区域生产力分异的关键因子。
2)在现有载畜水平下,草地退化年限分布于 1–18 年,临界载畜量范围为 0–11.06 SU·ha−1。部分区域出现临界载畜量为零的情况,说明当前已处于或超过生态承载极限。高寒草甸和草甸草原具有较高的生态耐受性与承载潜力,而典型草原与荒漠草原退化速率较快、系统脆弱性较高。
3)适度减少放牧可显著延缓退化进程并提高生态稳定性。载畜量减少 10% 与 30% 的情景下,退化年限均维持在 1–18 年,但在部分像元中表现为约 1–2 年的延长,且高值区范围扩大,系统稳定性明显增强。减少放牧调控效应在高寒草甸和山地草甸区最为显著,而荒漠草原则受限于低水热条件,恢复潜力有限。
4)在未来气候情景下,载畜阈值与退化年限的总体差异有限,但区域差异明显。RCP8.5 情景下部分区域退化时间略长于 RCP4.5,表明高排放条件下 CO2 浓度升高及降水增加共同作用产生了“气候补偿效应”,在一定程度上缓解了放牧压力。总体上,高寒草甸和山地草甸的承载潜力可能增强,而荒漠草原对气候波动更加敏感,生态风险进一步加剧。
Simulation assessment of net primary productivity and grazing thresholds of grasslands on the northern Tianshan Mountains
-
摘要: 以天山北坡草地为例,使用未受放牧影响的 LAI 时间序列来模拟未受放牧影响的生长轨迹,并用光能利用率模型计算未受放牧影响的净初级生产力(net primary productivity, NPP)。在此基础上,建立1个基于未受放牧影响的 NPP 的载畜阈值模型,用于描述不同放牧压力下的退化时间和阈值分布。结果表明,未受放牧影响的 NPP 的空间分布呈现“西高东低、山区高于平原”的特点。在当前载畜水平以及减少放牧 10%、30% 的情景下,草地退化时间总体均处于 1~18 a,部分区域出现临界载畜量为零的情况,说明当前已处于或超过生态承载极限。随着载畜量的降低,部分像元达到退化阈值的时间显著推迟,表明适度减少放牧可在局部区域延缓草地退化的发生。在RCP4.5 和 RCP8.5 情景下,临界载畜量在 3.59~9.60 SU·hm−2之间,载畜阈值与退化年限的总体差异有限,但区域差异明显。本研究提出的基于未受放牧影响的 NPP 的载畜阈值模拟方法,可以为定量识别草地生态的承受能力和实施差异化的放牧管理提供科学依据。Abstract: Grassland ecosystems are an important component of terrestrial ecosystems and play a critical role in maintaining ecological security and supporting livestock production. Due to climate warming and overgrazing, grassland productivity has declined, increasing the risk of ecological degradation. Existing studies on grazing intensity and grazing thresholds largely rely on empirical or static data, making it difficult to capture the dynamic changes of grasslands under grazing pressure. This study takes the grasslands on the northern Tianshan Mountains as an example, using LAI time series unaffected by grazing to simulate ungrazed growth trajectories and applying a light-use efficiency model to calculate ungrazed net primary productivity (NPP). Based on this, a grazing threshold model was established using ungrazed NPP to describe degradation time and threshold distribution under different grazing pressures. The results show that the spatial distribution of ungrazed NPP exhibits a “higher in the west and in mountainous areas, lower in the east and on plains” pattern. Under current stocking rates and scenarios with 10% and 30% reduced grazing, grassland degradation generally occurs within 1–18 years, with some areas reaching a critical stocking rate of zero, indicating that current grazing pressure has reached or exceeded the ecological carrying limit. As stocking rates decrease, the time for some pixels to reach the degradation threshold is significantly delayed, suggesting that moderate grazing reduction can locally postpone grassland degradation. Under the RCP4.5 and RCP8.5 scenarios, the critical stocking rate ranges from 3.59 to 9.60 SU/ha, with limited overall differences in grazing thresholds and degradation time, but notable regional variability. The proposed ungrazed NPP–based grazing threshold simulation approach provides a scientific basis for quantitatively identifying grassland ecological carrying capacity and supporting differentiated grazing management strategies.
-
图 2 天山北坡草地未受放牧影响的 NPP 空间分布图
Fig. 2 Spatial distribution of ungrazed NPP of grasslands on the northern Tianshan Mountains
图 3 天山北坡草地载畜阈值及退化时间空间分布
Fig. 3 Spatial distribution of grazing thresholds and degradation time of grasslands on the northern Tianshan Mountains
图 4 未来气候情景下天山北坡草地载畜阈值及退化时间空间格局
Fig. 4 Spatial patterns of grazing thresholds and degradation time of grasslands on the northern Tianshan Mountains under future climate scenarios
图 5 基于理想LAI构建的未受放牧影响的NPP与MOD17A3HGF NPP的验证
Fig. 5 Pixel-level comparison between ungrazed NPP simulated based on ideal LAI and MOD17A3HGF NPP
表 1 不同草地类型的根冠比
Table 1 Root-to-shoot ratio of different grassland types
草地类型
Grassland types根冠比
Root/shoot文献
Reference温带禾草、杂类草草甸草原
Temperate grass-forb meadow steppe5.2 [35] 温带丛生禾草典型草原
Temperate needlegrass arid steppe5.6 [35] 温带丛生矮禾草、矮半灌木荒漠草原
Temperate dwarf needlegrass,dwarf semi-shrubby6.4 [35] 高寒禾草、苔草草原
Alpine grass,Carex steppe5.2 [35] 温带禾草、苔草及杂类草沼泽化草甸
Temperate grass,Carex-forbe swamp meadow7.9 [36] 高寒蒿草、杂类草草甸
Alpine Kobresia-forb meadow6.8 [35]  下载: 导出CSV
下载: 导出CSV
-
[1] 周伟, 刚成诚, 李建龙, 等. 1982-2010年中国草地覆盖度的时空动态及其对气候变化的响应[J]. 地理学报, 2014, 69(1): 15−30. https://doi.org/10.11821/dlxb201401002 doi: 10.11821/dlxb201401002 [2] Wang D J, Peng Q Y, Li X Q, et al. A long-term high-resolution dataset of grasslands grazing intensity in China [J]. Scientific Data, 2024, 11(1): 1194. https://doi.org/10.1038/s41597-024-04045-x doi: 10.1038/s41597-024-04045-x [3] Yan N N, Zhu W W, Wu B F, et al. Assessment of the grassland carrying capacity for winter-spring period in Mongolia [J]. Ecological Indicators, 2023, 146: 109868. https://doi.org/10.1016/j.ecolind.2023.109868 doi: 10.1016/j.ecolind.2023.109868 [4] 黄季焜, 侯玲玲, 亢楠楠, 等. 草地生态系统服务经济价值评估研究[J]. 中国工程科学, 2023, 25(1): 198−206. https://doi.org/10.15302/J-SSCAE-2023.01.015 doi: 10.15302/J-SSCAE-2023.01.015 [5] Ma L S, Zheng J H, Pen J, et al. Monitoring and influencing factors of grassland livestock overload in Xinjiang from 1982 to 2020 [J]. Frontiers in Plant Science, 2024, 15: 1340566. https://doi.org/10.3389/fpls.2024.1340566 doi: 10.3389/fpls.2024.1340566 [6] 苏日娜, 俎佳星, 金花, 等. 内蒙古草地生产力及载畜量变化分析[J]. 生态环境学报, 2017, 26(4): 605−612. https://doi.org/10.16258/j.cnki.1674-5906.2017.04.009 doi: 10.16258/j.cnki.1674-5906.2017.04.009 [7] Li X Y, Zhao C Z, Kang M P, et al. Responses of net primary productivity to phenological dynamics based on a data fusion algorithm in the northern Qinghai-Tibet Plateau [J]. Ecological Indicators, 2022, 142: 109239. https://doi.org/10.1016/j.ecolind.2022.109239 doi: 10.1016/j.ecolind.2022.109239 [8] Ma R, Xia C L, Liu Y W, et al. Spatiotemporal change of net primary productivity and its response to climate change in temperate grasslands of China [J]. Frontiers in Plant Science, 2022, 13: 899800. https://doi.org/10.3389/fpls.2022.899800 doi: 10.3389/fpls.2022.899800 [9] Zhang Y J, Huang D, Badgery W B, et al. Reduced grazing pressure delivers production and environmental benefits for the typical steppe of north China [J]. Scientific Reports, 2015, 5(1): 16434. https://doi.org/10.1038/srep16434 doi: 10.1038/srep16434 [10] Qin P Y, Sun B, Li Z Y, et al. Estimation of grassland carrying capacity by applying high spatiotemporal remote sensing techniques in Zhenglan Banner, Inner Mongolia, China [J]. Sustainability, 2021, 13(6): 3123. https://doi.org/10.3390/su13063123 doi: 10.3390/su13063123 [11] Huang X T, Luo G P, Ye F P, et al. Effects of grazing on net primary productivity, evapotranspiration and water use efficiency in the grasslands of Xinjiang, China [J]. Journal of Arid Land, 2018, 10(4): 588−600. https://doi.org/10.1007/s40333-018-0093-z doi: 10.1007/s40333-018-0093-z [12] Zhang R R, Li Z H, Yuan Y W, et al. Analyses on the changes of grazing capacity in the three-river headwaters region of China under various climate change scenarios [J]. Advances in Meteorology, 2013, 2013: 951261. https://doi.org/10.1155/2013/951261 doi: 10.1155/2013/951261 [13] Huang L, Ning J, Zhu P, et al. The conservation patterns of grassland ecosystem in response to the forage-livestock balance in North China [J]. Journal of Geographical Sciences, 2021, 31(4): 518−534. https://doi.org/10.1007/s11442-021-1856-6 doi: 10.1007/s11442-021-1856-6 [14] Yan H, Ran Q W, Hu R H, et al. Machine learning-based prediction for grassland degradation using geographic, meteorological, plant and microbial data [J]. Ecological Indicators, 2022, 137: 108738. https://doi.org/10.1016/j.ecolind.2022.108738 doi: 10.1016/j.ecolind.2022.108738 [15] Huang X T, Luo G P, He H L, et al. Ecological effects of grazing in the Northern Tianshan Mountains [J]. Water, 2017, 9(12): 932. https://doi.org/10.3390/w9120932 doi: 10.3390/w9120932 [16] Tang Q H, Liu X C, Zhou Y Y, et al. Climate change and water security in the northern slope of the Tianshan Mountains [J]. Geography and Sustainability, 2022, 3(3): 246−257. https://doi.org/10.1016/j.geosus.2022.08.004 doi: 10.1016/j.geosus.2022.08.004 [17] 江康威, 张青青, 王亚菲, 等. 天山北坡中段草地生态系统多功能性对放牧的响应 [J]. 生态学报, 2024, 44(8): 3440−3456. https://doi.org/10.20103/j.stxb.202308111737 doi: 10.20103/j.stxb.202308111737 [18] Chen J C, Chen Y P, Liu Z, et al. Validation of MODIS LAI product using upscaling sentinel-2 decameter-scale LAI and field measured LAI[C]//IGARSS 2023 - 2023 IEEE international geoscience and remote sensing symposium. Pasadena: IEEE, 2023: 5119−5122. https://doi.org/10.1109/IGARSS52108.2023.10281427 [19] 陈惺, 王军邦, 何启凡, 等. 未来气候情景下中国植被净初级生产力稳定性及气候影响[J]. 地理学报, 2023, 78(3): 694−713. https://doi.org/10.11821/dlxb202303012 doi: 10.11821/dlxb202303012 [20] Griffiths P, Kuemmerle T, Baumann M, et al. Forest disturbances, forest recovery, and changes in forest types across the Carpathian ecoregion from 1985 to 2010 based on Landsat image composites [J]. Remote Sensing of Environment, 2014, 151: 72−88. https://doi.org/10.1016/j.rse.2013.04.022 doi: 10.1016/j.rse.2013.04.022 [21] Cleveland W S. Robust locally weighted regression and smoothing scatterplots [J]. Journal of the American Statistical Association, 1979, 74(368): 829−836. https://doi.org/10.1080/01621459.1979.10481038 doi: 10.1080/01621459.1979.10481038 [22] Zhu J P, Xie A H, Qin X, et al. An assessment of ERA5 reanalysis for Antarctic near-surface air temperature [J]. Atmosphere, 2021, 12(2): 217. https://doi.org/10.3390/atmos12020217 doi: 10.3390/atmos12020217 [23] Ryu Y, Jiang C Y, Kobayashi H, et al. MODIS-derived global land products of shortwave radiation and diffuse and total photosynthetically active radiation at 5km resolution from 2000 [J]. Remote Sensing of Environment, 2018, 204: 812−825. https://doi.org/10.1016/j.rse.2017.09.021 doi: 10.1016/j.rse.2017.09.021 [24] Pan L, Xiao X M, Pan B H, et al. Interannual variations and trends of gross primary production and transpiration of four mature deciduous broadleaf forest sites during 2000–2020 [J]. Remote Sensing of Environment, 2024, 304: 114042. https://doi.org/10.1016/j.rse.2024.114042 doi: 10.1016/j.rse.2024.114042 [25] Ma X L, Huete A, Yu Q, et al. Parameterization of an ecosystem light-use-efficiency model for predicting savanna GPP using MODIS EVI [J]. Remote Sensing of Environment, 2014, 154: 253−271. https://doi.org/10.1016/j.rse.2014.08.025 doi: 10.1016/j.rse.2014.08.025 [26] Xiangming Xiao. Light absorption by leaf chlorophyll and maximum light use efficiency [J]. IEEE Transactions on Geoscience and Remote Sensing, 2006, 44(7): 1933−1935. https://doi.org/10.1109/TGRS.2006.874796 doi: 10.1109/TGRS.2006.874796 [27] Dye D G. Spectral composition and quanta‐to‐energy ratio of diffuse photosynthetically active radiation under diverse cloud conditions [J]. Journal of Geophysical Research: Atmospheres, 2004, 109(D10): D10203. https://doi.org/10.1029/2003JD004251 doi: 10.1029/2003JD004251 [28] Zhang Y, Xiao X, Jin C, et al. Consistency between sun-induced chlorophyll fluorescence and gross primary production of vegetation in North America [J]. Remote Sensing of Environment, 2016, 183: 154−169. https://doi.org/10.1016/j.rse.2016.05.015 doi: 10.1016/j.rse.2016.05.015 [29] Xiao X, Zhang Q, Braswell B, et al. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data [J]. Remote Sensing of Environment, 2004, 91(2): 256−270. https://doi.org/10.1016/j.rse.2004.03.010 doi: 10.1016/j.rse.2004.03.010 [30] Zhang L X, Hu Z M, Fan J W, et al. A meta-analysis of the canopy light extinction coefficient in terrestrial ecosystems [J]. Frontiers of Earth Science, 2014, 8(4): 599−609. https://doi.org/10.1007/s11707-014-0446-7 doi: 10.1007/s11707-014-0446-7 [31] Zhang Y, Xiao X M, Wu X C, et al. A global moderate resolution dataset of gross primary production of vegetation for 2000–2016 [J]. Scientific Data, 2017, 4(1): 170165. https://doi.org/10.1038/sdata.2017.165 doi: 10.1038/sdata.2017.165 [32] Landsberg J J, Waring R H, Williams M. The assessment of NPP/GPP ratio [J]. Tree Physiology, 2020, 40(6): 695−699. https://doi.org/10.1093/treephys/tpaa016 doi: 10.1093/treephys/tpaa016 [33] 王昭生. 1949-2008年东亚区域陆地生态系统NPP和GPP的时空格局演变[D]. 湘潭: 湖南科技大学, 2012. doi: 10.7666/d.D290678 [34] 张千羚. 基于NPP数据的锡林郭勒盟牧区草地资源承载力估算研究[J]. 赤峰学院学报(自然科学版), 2024, 40(2): 1−5. https://doi.org/10.3969/j.issn.1673-260X.2024.02.002 doi: 10.3969/j.issn.1673-260X.2024.02.002 [35] Yang Y H, Fang J Y, Ma W H, et al. Large‐scale pattern of biomass partitioning across China’s grasslands [J]. Global Ecology and Biogeography, 2010, 19(2): 268−277. https://doi.org/10.1111/j.1466-8238.2009.00502.x doi: 10.1111/j.1466-8238.2009.00502.x [36] 莫兴国, 刘文, 孟铖铖, 等. 青藏高原草地产量与草畜平衡变化[J]. 应用生态学报, 2021, 32(7): 2415−2425. https://doi.org/10.13287/j.1001-9332.202107.002 doi: 10.13287/j.1001-9332.202107.002 [37] Shiyomi M, Akiyama T, Wang S P, et al. A grassland ecosystem model of the Xilingol steppe, Inner Mongolia, China [J]. Ecological Modelling, 2011, 222(13): 2073−2083. https://doi.org/10.1016/j.ecolmodel.2011.03.028 doi: 10.1016/j.ecolmodel.2011.03.028 [38] Zhu Q A, Chen H, Peng C H, et al. An early warning signal for grassland degradation on the Qinghai-Tibetan Plateau [J]. Nature Communications, 2023, 14(1): 6406. https://doi.org/10.1038/s41467-023-42099-4 doi: 10.1038/s41467-023-42099-4 [39] Ellis J E, Swift D M. Stability of African pastoral ecosystems: alternate paradigms and implications for development [J]. Journal of Range Management, 1988, 41(6): 450−459. https://doi.org/10.2307/3899515 doi: 10.2307/3899515 [40] Ji F K, Xi G L, Xie Y W, et al. Estimating grassland carrying capacity in the Source Area of Nujiang River and Selinco Lake, Tibetan Plateau (2001–2020) based on multisource remote sensing [J]. Remote Sensing, 2024, 16(20): 3790. https://doi.org/10.3390/rs16203790 doi: 10.3390/rs16203790 [41] Wang Y G, Luo G P, Li C F, et al. Grazing weakens the carbon sequestration capacity of dry temperate grassland ecosystems in Central Asia [J]. CATENA, 2025, 248: 108608. https://doi.org/10.1016/j.catena.2024.108608 doi: 10.1016/j.catena.2024.108608 [42] Zhao M S, Running S W, Nemani R R. Sensitivity of Moderate Resolution Imaging Spectroradiometer (MODIS) terrestrial primary production to the accuracy of meteorological reanalyses [J]. Journal of Geophysical Research: Biogeosciences, 2006, 111(G1): G01002. https://doi.org/10.1029/2004JG000004 doi: 10.1029/2004JG000004 [43] Huang X T, Luo G P, Ma Z, et al. Modeling the effect of grazing on carbon and water use efficiencies in grasslands on the Qinghai–Tibet Plateau [J]. BMC Ecology and Evolution, 2024, 24(1): 26. https://doi.org/10.1186/s12862-024-02215-4 doi: 10.1186/s12862-024-02215-4 [44] Li C F, Han Q F, Xu W Q. Contribution of climate change and grazing on carbon dynamics in central Asian pasturelands [J]. Remote Sensing, 2022, 14(5): 1210. https://doi.org/10.3390/rs14051210 doi: 10.3390/rs14051210 [45] Reinermann S, Asam S, Kuenzer C. Remote sensing of grassland production and management—a review [J]. Remote Sensing, 2020, 12(12): 1949. https://doi.org/10.3390/rs12121949 doi: 10.3390/rs12121949 [46] Lulandala L, Bargués‐Tobella A, Masao C A, et al. Excessive livestock grazing overrides the positive effects of trees on infiltration capacity and modifies preferential flow in dry miombo woodlands [J]. Land Degradation & Development, 2022, 33(4): 581−595. https://doi.org/10.1002/ldr.4149 doi: 10.1002/ldr.4149 [47] Chen H, Zhao X R, Chen X J, et al. Seasonal changes of soil microbial C, N, P and associated nutrient dynamics in a semiarid grassland of north China [J]. Applied Soil Ecology, 2018, 128: 89−97. https://doi.org/10.1016/j.apsoil.2018.04.008 doi: 10.1016/j.apsoil.2018.04.008 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 433
- HTML全文浏览量: 213
- 被引次数: 0














