

-
热带原始森林拥有最丰富的生物多样性,具备调节气候、水源涵养、水土保持等生态系统功能,还蕴藏着巨大的资源潜力与文化意义。因此,保护热带雨林对减缓全球气候变化至关重要。海南岛拥有中国最广阔的热带森林资源,是全球三大热带雨林群系中印度-马来群系的重要组成环节,其岛屿型热带雨林生态系统具有国家代表性和全球保护意义[1]。由于人为干扰,不同恢复程度的次生林已成为海南热带雨林的主要森林类型之一[1]。土壤持水能力是水源涵养的核心指标,而水源涵养又是热带雨林重要的生态系统服务功能,因此研究不同恢复阶段热带次生林土壤持水能力的变异特征及其调控因素,对于指导热带雨林生态系统的恢复与管理,提升水源涵养功能具有重要意义。
目前,国内外学者从不同角度研究了不同恢复阶段森林土壤持水能力变异特征[2]。研究表明,温带和亚热带地区森林恢复过程中土壤持水能力逐渐改善,随着林分发育会改善其水源涵养功能,具体表现为土壤变得更疏松(孔隙度增大、容重减小),从而能够容纳更多的水分(最大持水量和非毛管持水量增加)[3-5]。对华南热带区不同恢复阶段人工林土壤持水能力研究发现,最大持水量、毛管持水量和最小持水量均为村边次生林>阔叶混交林>光裸地>桉树林,随着恢复阶段的推进,土壤持水能力呈现向好的趋势[4]。通过分析安徽老山亚热带常绿阔叶林不同林龄阶段土壤特性及其水源涵养功能的变化发现,该区域随着林分的发育,其水源涵养功能不断提高,不同林分的水源涵养功能依次为:65年生>48年生>36年生>幼龄林[5]。总体而言,以往大部分学者研究的是温带和亚热带地区森林恢复过程土壤持水能力特征,但对于热带地区森林恢复过程土壤持水能力演变特征仍不清楚。
基于此,本研究以不同恢复阶段的热带次生林为研究对象,通过对测定热带次生林土壤理化性质和土壤持水特性,探讨不同恢复阶段土壤水源涵养功能的变异特征以及调控因素,该研究为预测热带地区森林土壤持水能力是否随着恢复阶段的推进趋势向好甚至趋近于原始林提供一定的理论依据。
-
研究区域位于海南岛保亭县(18°32′~18°58′ N,109°21′~109°56′ E),属热带季风气候,该气候类型四季更替不显著,终年高温,但夏季少有极端高温天气,冬季也罕有低温寒冷,气温年际变化幅度小;降水呈现鲜明的干湿季交替,雨量集中于夏秋两季,冬春则较为干旱;热带气旋活动频繁。光热条件充足,水资源总量丰富,然而台风、干旱及暴雨等气象灾害亦常发生[6]。年均温23℃,降水与风向有密切关系,冬季盛行源于大陆的东北季风,降水稀少,夏季4月至10月,占全年降水量的80%以上[7]。盛行源于印度洋的西南季风,降水丰沛,年降水1900 mm。土壤以铁铝土和原始土为主[8]。选取3个恢复阶段样地,详细信息见表1和表2:
表 1 不同恢复阶段次生林群落特征
Table 1. Characteristics of secondary habitat communities at different restoration stages
恢复阶段
Recovery stages高度/m
Height胸径/cm
DHB郁闭度
Canopy density主要树种
Dominant tree species早期
Early stage5.64±0.15 8.49±0.32 0.73 厚皮树Lannea coromandelica
细基丸Huberantha cerasoides
黄牛木Cratoxylum cochinchinense
银柴Aporosa dioica中期
Middle stage10.98±0.75 15.84±1.72 0.80 无患子Sapindus saponaria
杂色榕Ficus variegate
锈毛野桐Mallotus anomalus
楹树Albizia chinensis
倒吊笔Wrightia pubescens后期
Late stage15.28±1.17 20.74±1.81 0.88 重阳木Bischofia polycarpa
调羹树Heliciopsis lobata
海南黄檀Dalbergia hainanensis
杂色榕Ficus variegate表 2 不同恢复阶段次生林根和凋落物的生物量
Table 2. Biomass of secondary forest roots and litter at different stages of restoration
恢复阶段
Recovery stages根生物量/(g·m−2)
Root biomass凋落物生物量/(t·hm−2)
Litter biomass早期 Early stage 83.21 1.68 中期 Middle stage 158.29 2.83 后期 Late stage 249.75 4.45 -
本试验于2023年12月选取海南岛保亭县不同恢复阶段(10~15年为恢复阶段早期,20~30年为恢复阶段中期,40~60年为恢复阶段后期)的次生林,每个恢复阶段样地设置20 m × 20 m的5块重复样地,并在每个重复样地随机设置1 m × 1 m的四个小重复样地进行取样。
-
在不同恢复阶段样地的每个小重复样地内,按照10 cm间隔,用环刀分别采集0~10 cm、10~20 cm、20~30 cm、30~40 cm、40~50 cm、50~60 cm六种土层深度土壤样品,挑去石砾和植物残体,装入自封袋保存,土壤样品采集结束后带回实验室备用。
-
将采集土壤样品封装后备用,将环刀上下盖子替换成垫有滤纸的带孔盖。称取空环刀质量(M0,g),其次放入平底盘中,在平盘中注入刚好没过环刀的水,注意避免各种原因引起水分减少,使其吸水达24 h以上,这个状态环刀土壤中所有孔隙主要是非毛管孔隙及毛管孔隙已经充分吸收了水分,水平取出,立即称量(M1,g)。为了排出环刀中土壤的非毛管水分,将其置于铺有干砂的容器中,静置2 h,此时已全部流出,但环刀中土壤的毛细管仍充满水分,立即称量(M2,g)。为了测量环刀中土壤的毛管悬着水,将环刀置于铺有全新干砂的容器中,静置24 h,立即称量(M3,g)。最后将土样放入105℃烘箱中烘24 h至恒重,称烘干后的土样环刀质量($ \text{M}_{\text{0}}^{\text{'}} $,g),将干土去除并洗净环刀和盖子,称烘干后的环刀和盖子质量(M4,g)。具体计算公式如下:
$$ \text{SWHC=}\frac{{\text{M}}_{\text{1}}-\text{M}_{\text{0}}^{{'}}}{\text{M}_{\text{0}}^{{'}}-{\text{M}}_{\text{4}}}\times{100\%} $$ (1) $$ \text{CWC=}\frac{{\text{M}}_{\text{2}}-\text{M}_{\text{0}}^{{'}}}{\text{M}_{\text{0}}^{'}-{\text{M}}_{\text{4}}}\times{100\%} $$ (2) $$ \text{FC=}\frac{{\text{M}}_{\text{3}}-\text{M}_{\text{0}}^{'}}{\text{M}_{\text{0}}^{'}\text-{\text{M}}_{\text{4}}}\times{100\%} $$ (3) $$ \text{SWC=}\frac{{\text{M}}_{\text{0}}-\text{M}_{\text{0}}^{'}}{\text{M}_{\text{0}}^{'}-{\text{M}}_{\text{4}}}\times{100\%} $$ (4) $$ \text{BD=}\frac{\text{M}_{\text{0}}^{'}-{\text{M}}_{\text{4}}}{\text{V}} $$ (5) $$ {\text{K}}_{\text{s}}=\frac{{\text{M}}_{\text{1}}\text-{\text{M}}_{\text{2}}}{\text{t}\cdot{\mathrm{A}}}\times{100\%} $$ 式中,$\text{M}_{\text{0}}^{'} $为烘干后土壤环刀重量(g);M1为浸泡24 h土壤环刀重量(g);M2为控水2 h土壤环刀重量(g);M3为控水二至三昼夜土壤环刀重量(g);M4为空环刀重量(g)。其中BD、SWHC、CWC、FC和SWC分别为土壤容重(g·cm−3)、土壤饱和持水量(%)、毛管持水量(%)、田间持水量(%)和自然含水量(%)、V为环刀体积(cm3)、$ {K}_{\mathrm{s}} $为饱和导水率(cm3·min−1)、t为控水时间,为120 min(2 h)、A为环刀横截面积(cm2)。
-
部分土壤样品在实验室自然风干,人工去除植物根系并过筛,制备成2 mm,1 mm,0.25 mm的待测土样,用于测定土壤机械组成和土壤有机质等理化性质。按照国际分类标准土壤粒径划分为砂粒(0.02~2 mm)、粉粒(0.002~0.02 mm)和黏粒(0~0.002 mm)。土壤全碳(TC)、全氮(TN)、C:N比值采用德国elementar公司的vario EL cube型元素分析仪测定。土壤总孔隙度(TP,%)、毛管孔隙度(CP,%)和非毛管孔隙度(NCP,%)采用如下公式进行计算:
$$ \text{TP}=\left(1-\frac{\text{BD}}{{\text{d}}_{\text{s}}}\right)\times 100\% $$ $$ {\mathrm{CP}}={\mathrm{CWC}}\times{\mathrm{BD}} $$ $$ \text{NCP}={\mathrm{TP}}-{\mathrm{CP}} $$ 式中,土壤颗粒密度(ds)为2.65 g·cm−3,CWC的维度转换与体积-质量换算假设(ρwater=1 g·cm−3)。
-
本研究采用单因素方差分析(ANOVA)检验了四个恢复阶段相同土壤深度下土壤和水文特性之间的差异,当方差分析结果在P<0.05时显著,则进行最小显著差异检验以分析差异。然后采用双因素方差分析(ANOVA)探究水文和土壤特性受恢复阶段和土壤深度及其交互作用的影响。采用冗余分析(RDA)来确定土壤特性与土壤水文特性之间的关系。利用“rdacca.hp”和“vegan”包定量评估土壤特性对水文特性相对贡献。采用主成分分析(PCA)计算土壤持水特性的PC1。所有数据均以平均值±标准误差(SE)表示。数据处理使用R软件(版本4.4.3)完成;制图运用Origin 2021完成。
-
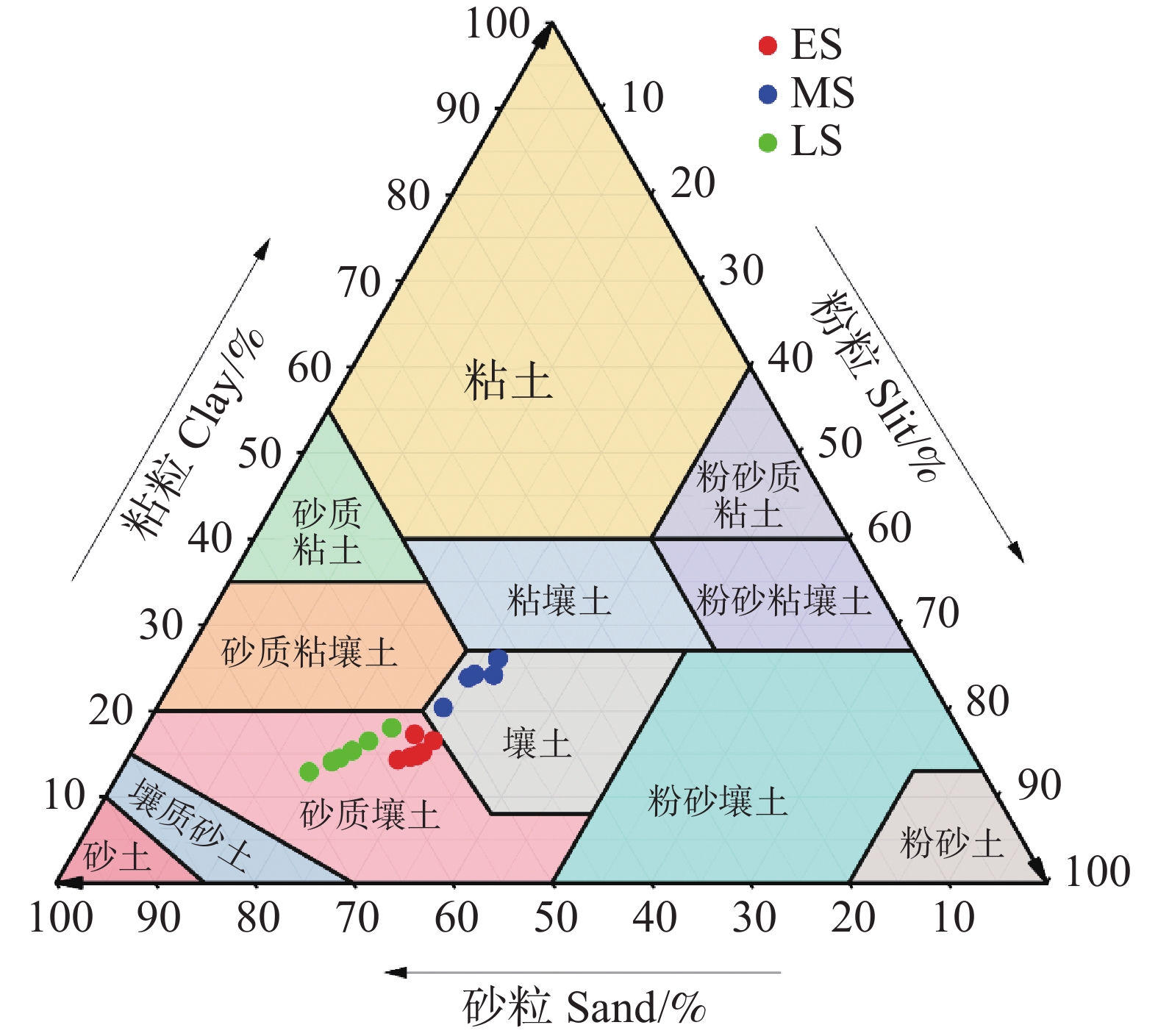
不同恢复阶段土壤机械组成存在显著差异(图1)。在三个恢复阶段的(0~60 cm)土层中,土壤粒径分布以砂粒为主(平均54.63%),粉粒(平均27.10%)和粘粒(平均18.27%)次之(图1)。恢复早期和后期土壤质地为砂壤土,中期土壤质地为壤土(图2)。而恢复中期(0~60 cm)土壤粘粒含量高于早期和后期,砂粒含量低于其它两个阶段,其中(0~20 cm)土层更为显著(P<0.05),恢复后期砂粒略高于早期。恢复早期和中期(0~60 cm)粉粒含量显著高于恢复后期,其中(10~20 cm)土层更为显著(P<0.05)。由此说明,次生林阶段对土壤粒径分布有显著影响,而土壤深度以及土壤深度与恢复阶段之间的交互作用对土壤粒径分布无显著影响。
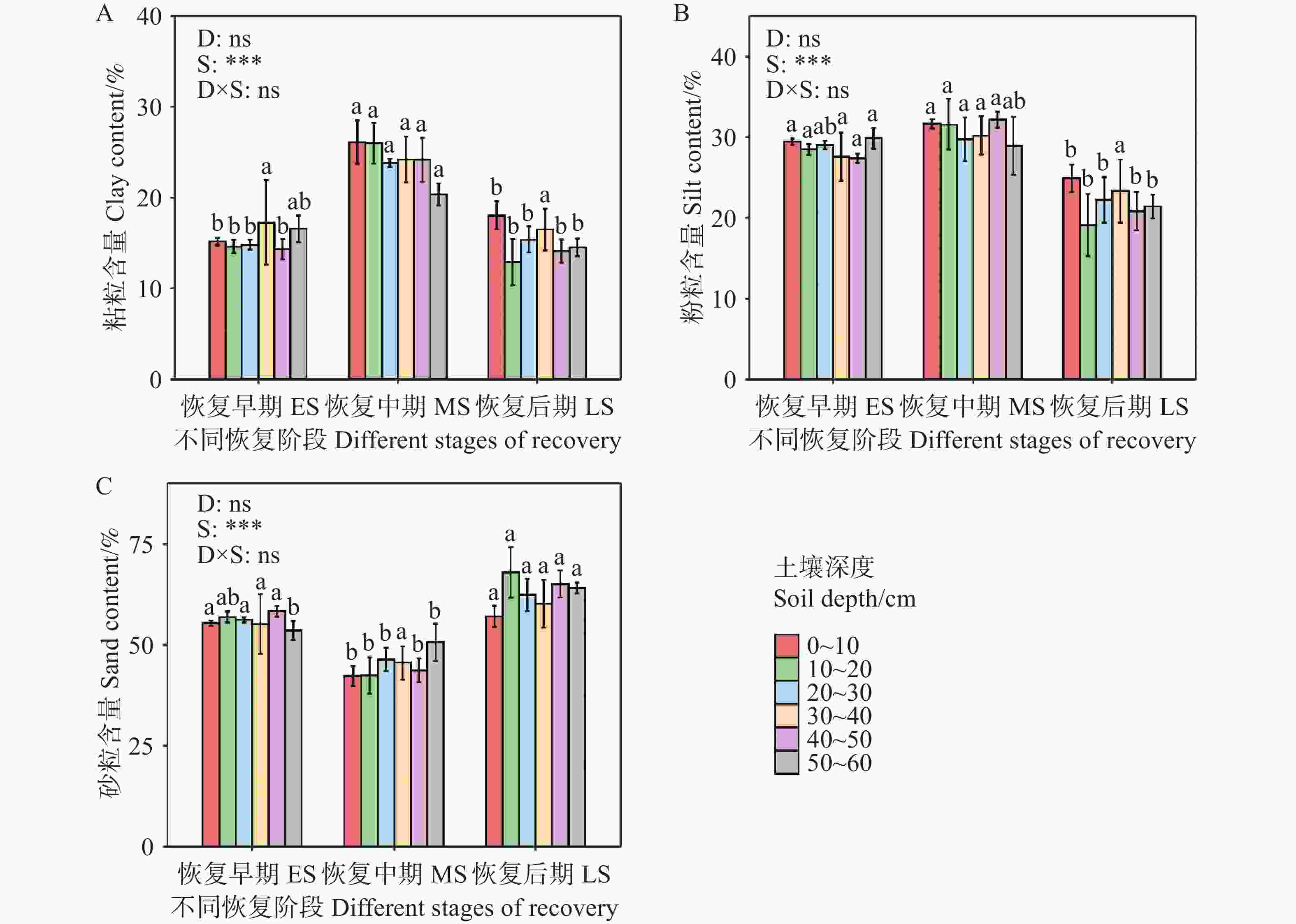
图 1 不同恢复阶段次生林土壤粒径分布
Figure 1. Distribution of soil particle size of secondary forests at different stages of recovery
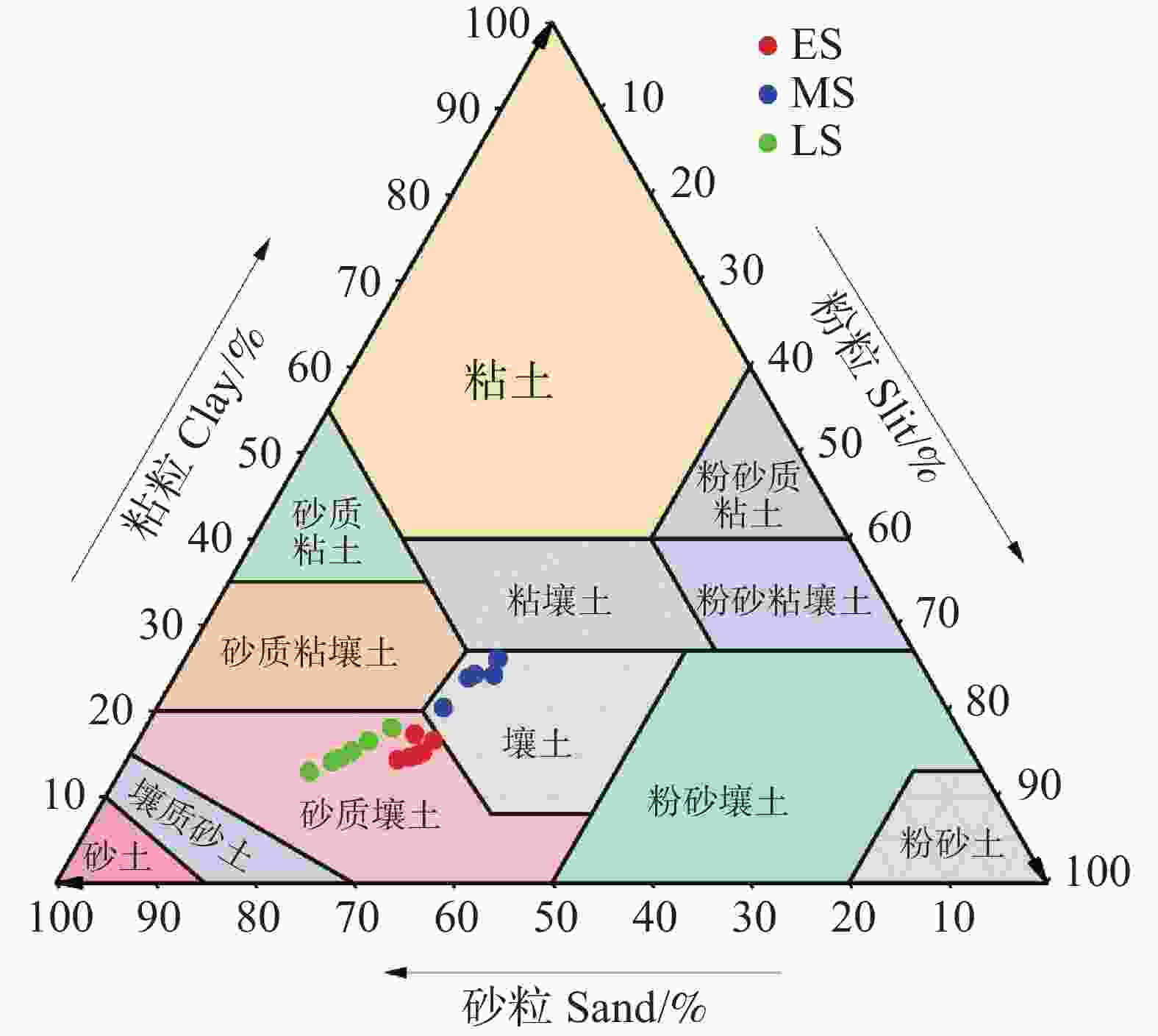
图 2 不同恢复阶段次生林的土壤质地
Figure 2. Soil particle size distribution of secondary forests at different stages of recovery
-
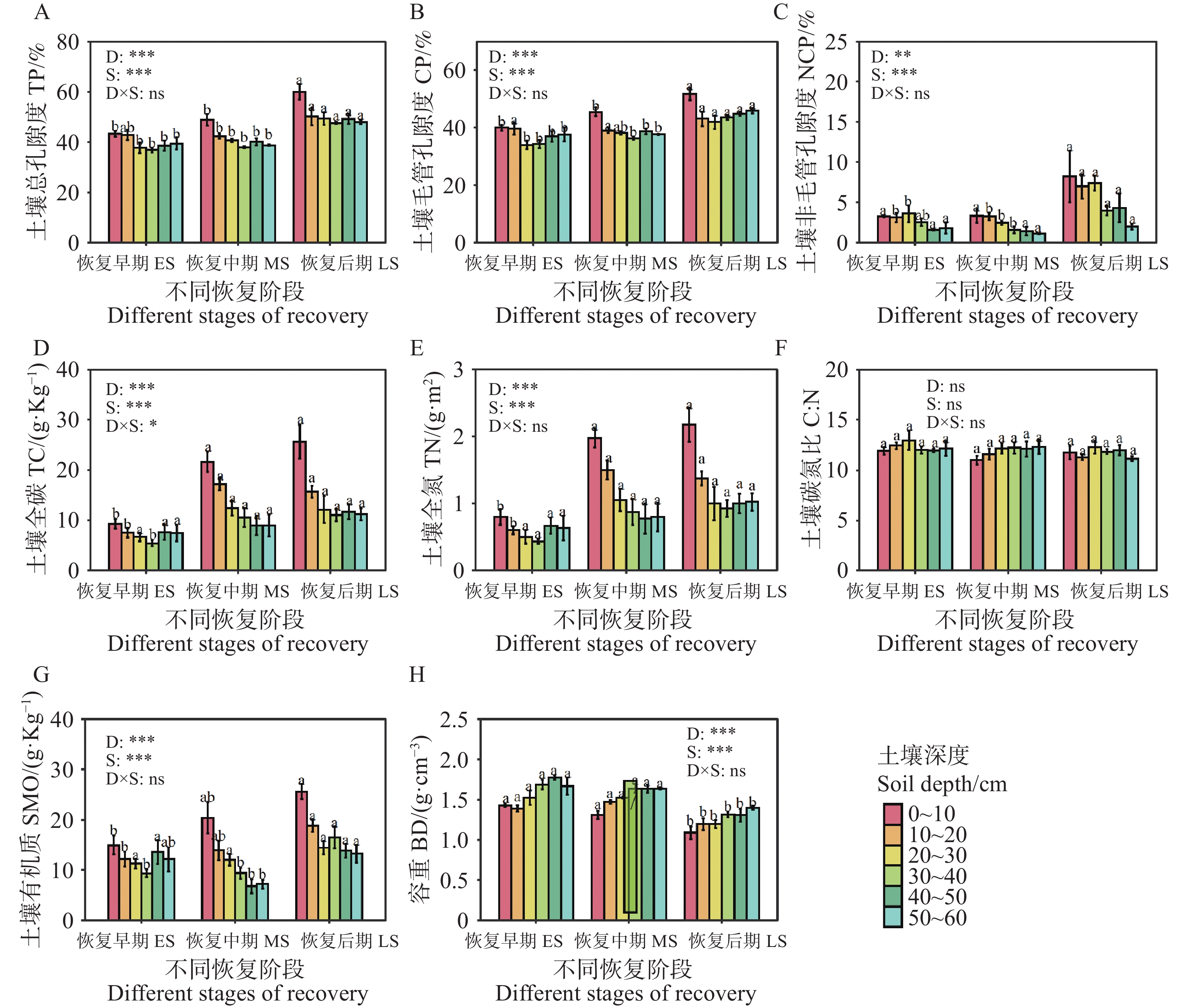
不同恢复阶段土壤理化属性存在显著差异,次生林(0~60 cm)的土壤有机质、土壤总孔隙度、土壤毛管孔隙度和非毛管孔隙度随恢复进程的推进总体呈增加趋势,而土壤容重则呈降低趋势(图3)。恢复后期(0~10 cm)层总孔隙度和毛管孔隙度显著提升,(P<0.05,图3-A—B),而次生林(0~60 cm)土壤容重在恢复后期显著低于恢复早期和中期(P<0.05,图3-H)。次生林(0~60 cm)的土壤全碳和土壤全氮随恢复阶段而增加,其恢复早期(0~20 cm)显著低于其它两个恢复阶段(P<0.05,图3-G—H)。土壤碳氮比在同一土层不同恢复阶段没有存在显著差异(P<0.05,图3-F)。
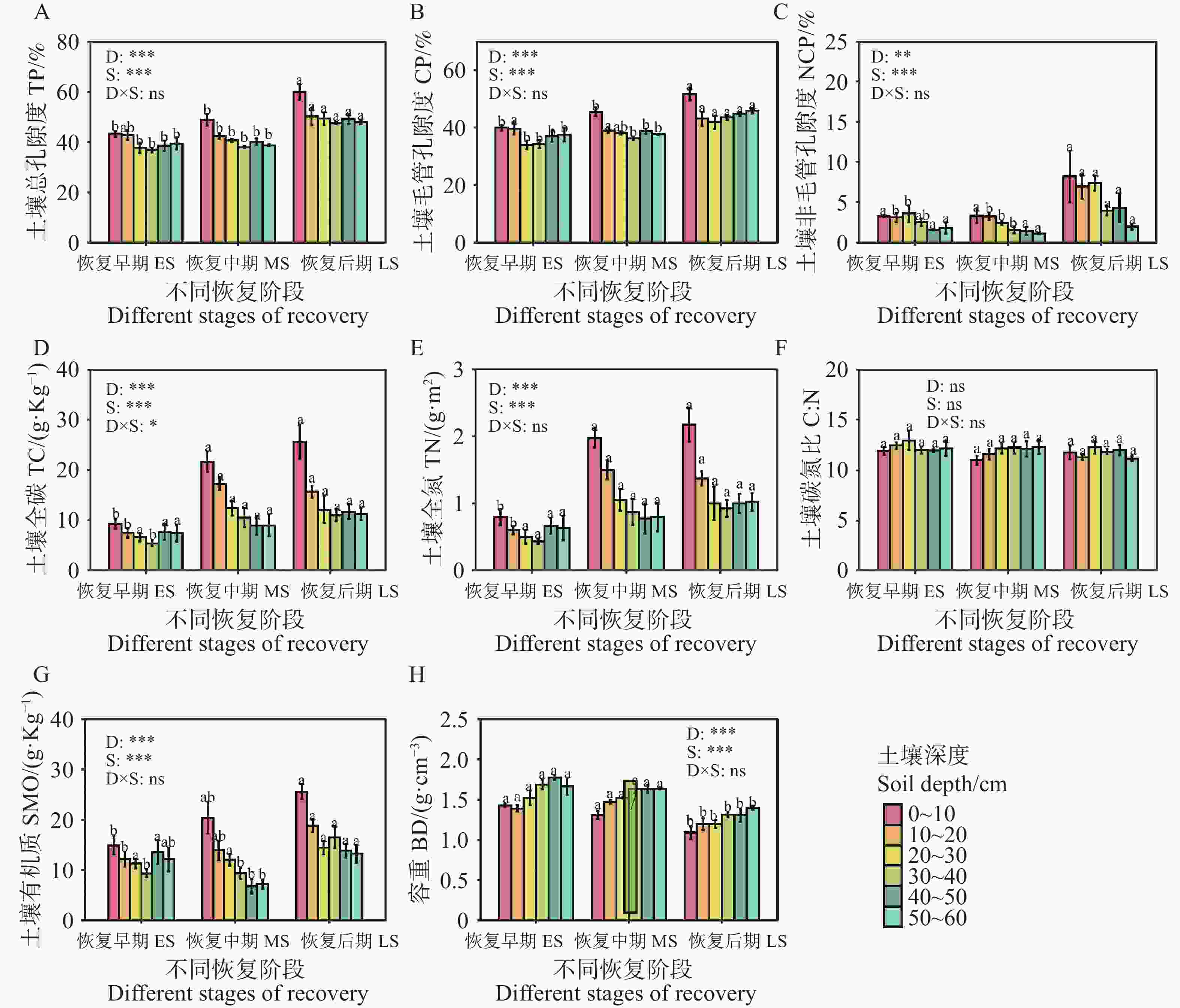
图 3 不同恢复阶段次生林的土壤理化性质
Figure 3. Soil physicochemical properties of secondary forests at different stages of recovery
-
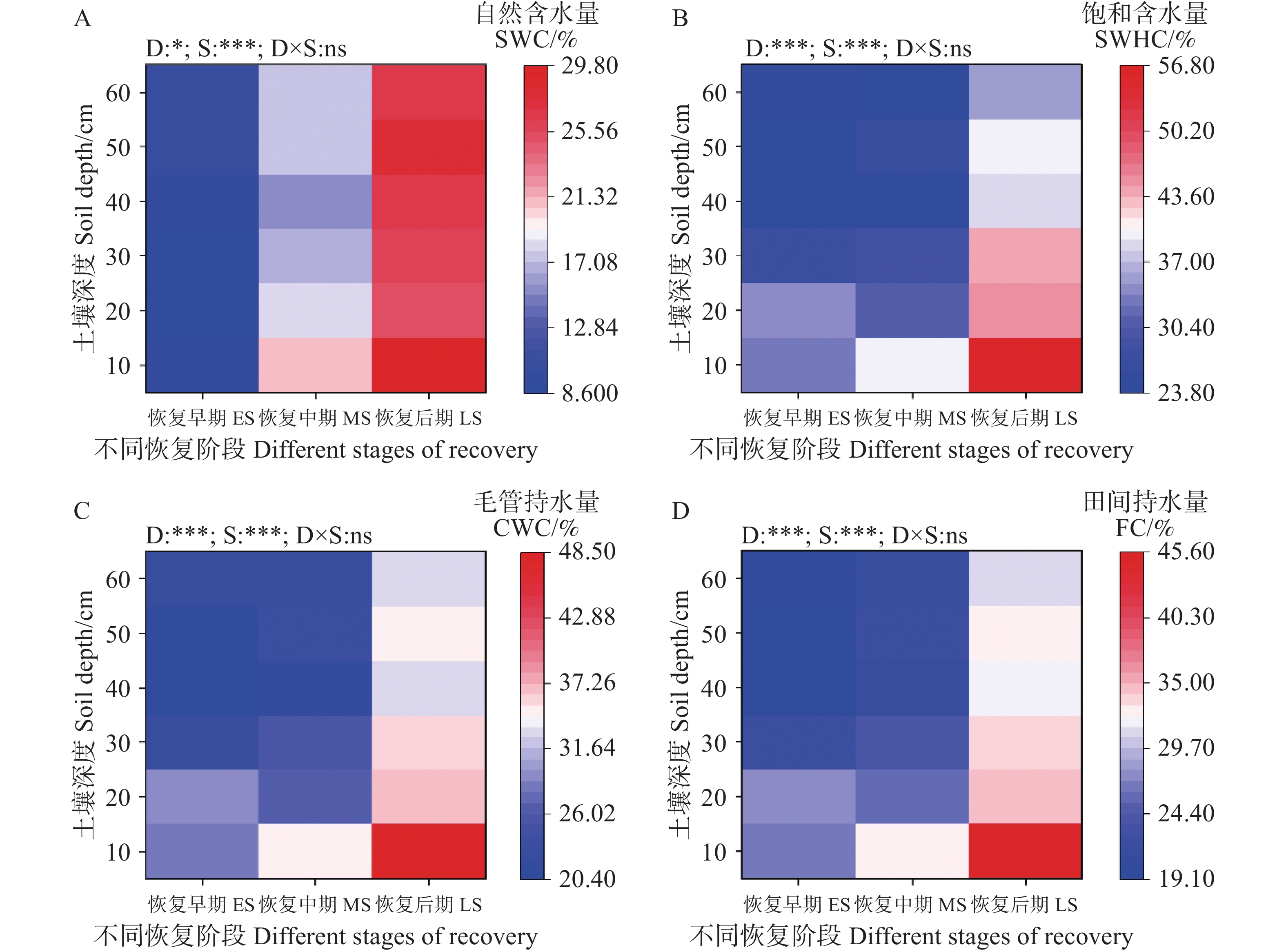
次生林(0~60 cm)土壤含水量随恢复进程的推进呈增加趋势(图4)。其中,(0~10 cm)土层次生林土壤自然含水量、饱和持水量、毛管持水量和田间持水量均随恢复阶段的进行而增加(图4)。双因素方差分析表明,土壤自然含水量、饱和持水量、毛管持水量和田间持水量受土壤深度和恢复阶段的影响显著(P<0.001),而土壤深度与恢复阶段之间的交互作用对土壤持水能力没有显著影响(图4)。饱和导水率随恢复阶段逐渐增加(表3)。
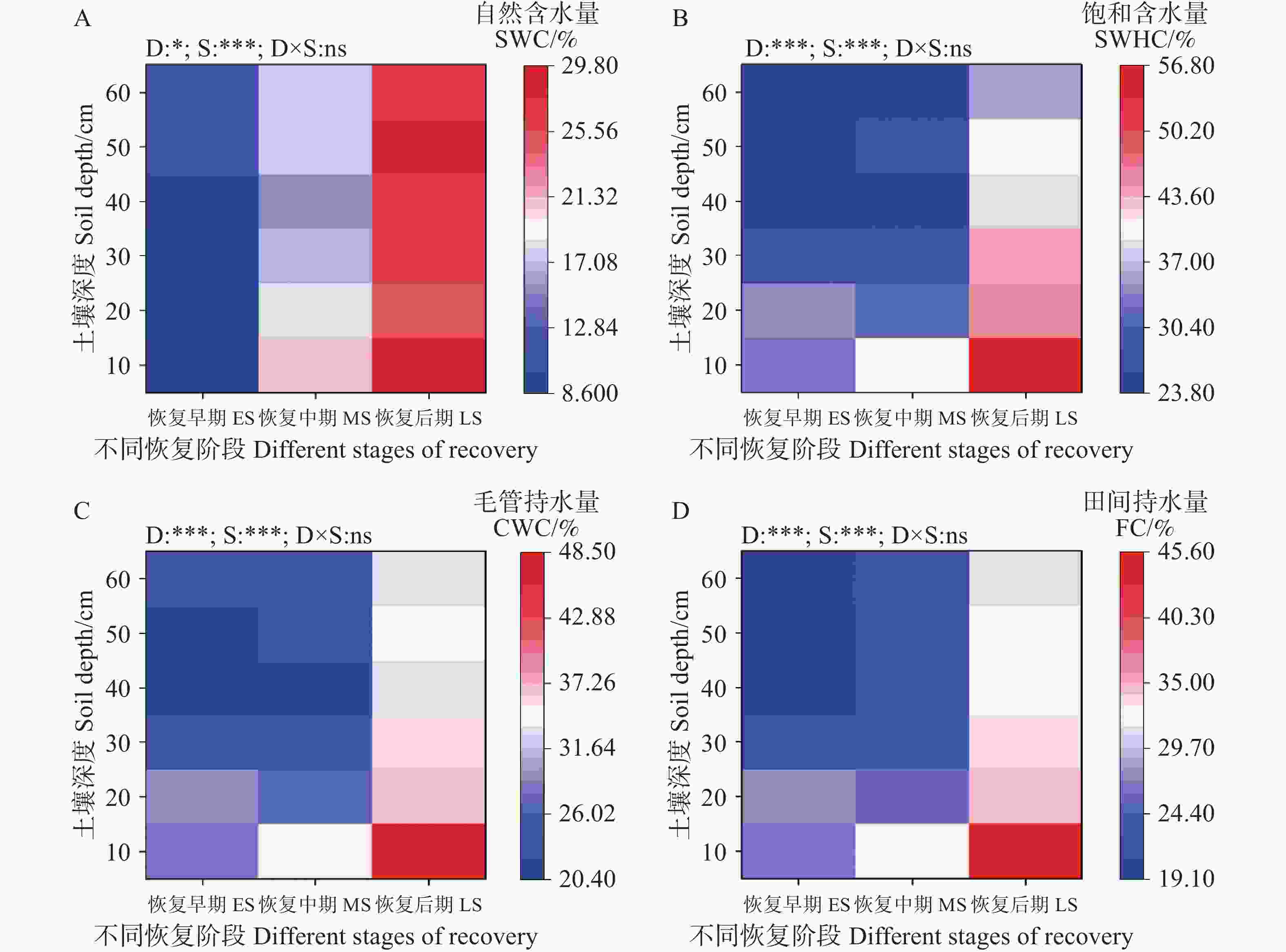
图 4 不同恢复阶段次生林的土壤含水量
Figure 4. Soil water content in secondary forests at different restoration stages
表 3 不同恢复阶段次生林表层土壤(0~10 cm)的饱和导水率
Table 3. Topsoil (0~10 cm)saturated hydraulic conductivity of secondary forests at different stages of recovery
恢复阶段Recovery stages 早期 Early stage 中期 Middle stage 后期 Late stage 饱和导水率/(cm·min−1)
Saturated hydraulic conductivity16.99 17.38 18.23 -
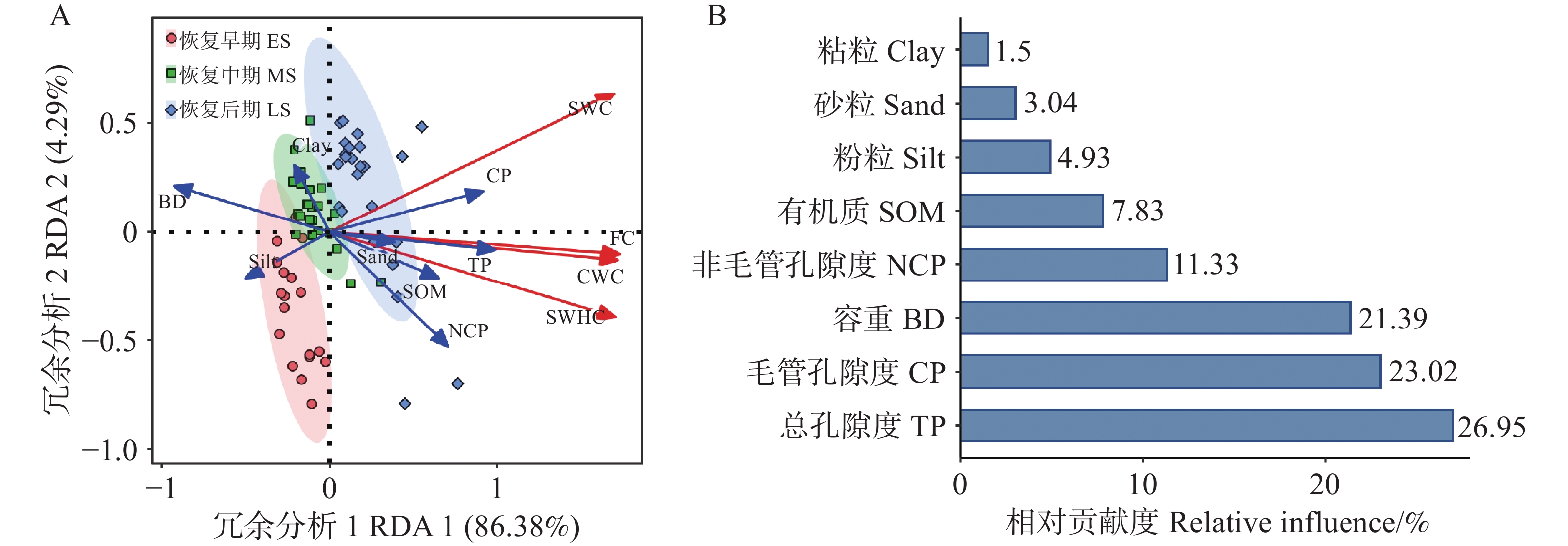
冗余分析表明,土壤持水能力与总孔隙度、毛细孔隙度、非毛细孔隙度和土壤有机质呈正相关,但与土壤容重呈负相关(图5)。RDA1和RDA2共同解释了土壤持水能力总变异的90.67%(图5A)。在土壤性质中,总孔隙度是影响土壤持水能力的最重要因素,能够解释总变异的26.95%,其次是毛管孔隙度(23.02%)、土壤容重(21.39%)、非毛细孔隙度(11.33%)和土壤有机质(7.83%),而土壤粒径粘粒、粉粒、砂粒含量对土壤持水能力的影响较弱(图5B,P<0.05)。
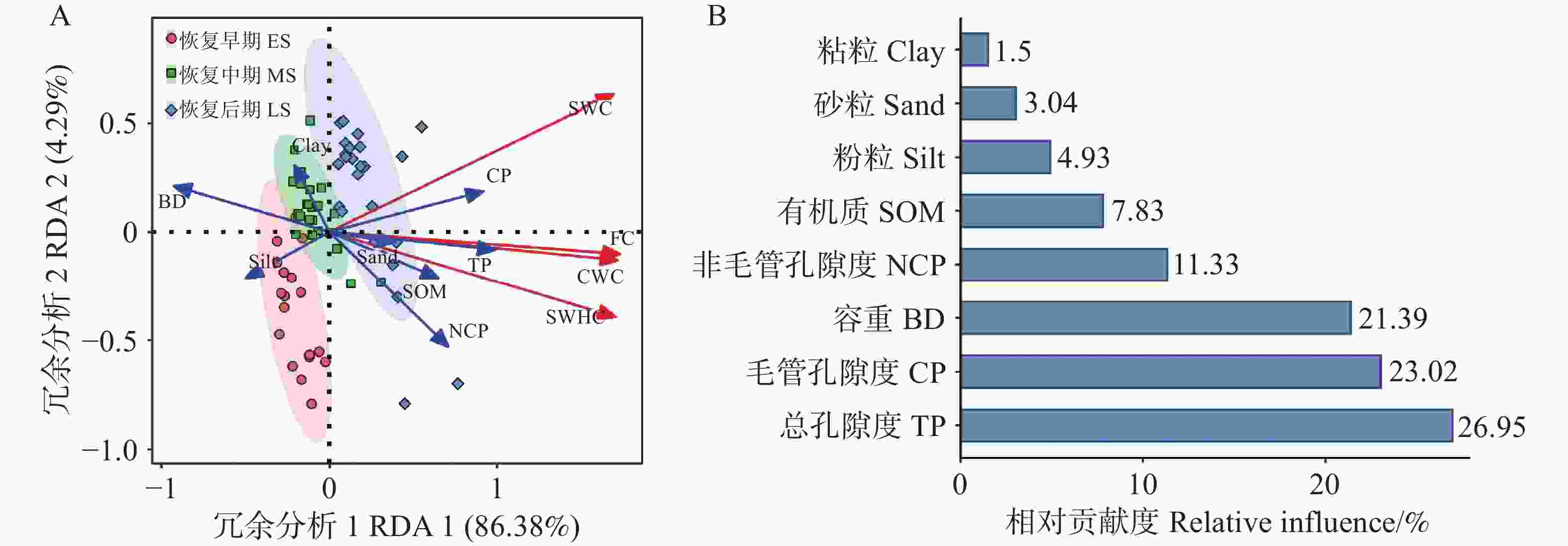
图 5 不同恢复阶段次生林土壤理化性质对土壤持水能力的冗余分析
Figure 5. Redundant analysis of soil water-holding capacity of secondary forests at different recovery stages
-
森林植被的恢复过程,无论是直接还是间接途径,均会引发生物因素(例如物种丰富度)的改变,进而对土壤的理化性质产生深远且显著的影响[9]。不同类别的植被恢复模式,通过各自独特的方式作用于土壤,从而对土壤质量予以改善,然而,用以衡量土壤性质的具体指标却因植被恢复类型的差异有所不同[10]。土壤容重与土壤孔隙度作为土壤物理性质的关键基础指标,能够直观地反映森林土壤的结构状况以及持水能力。本研究中,随着恢复进程的逐步推进,土壤容重呈现下降趋势,这一结果与先前研究的结论相契合,即在恢复后期,土壤容重达到最低值,尤其在(0~10 cm)的土层中,容重显著降低[11]。分析其原因,浅层土壤中分布着数量庞大的根系,这些根系在生长扩展过程中,犹如天然的“改良剂”,有效地改善了土壤结构,使得土壤孔隙度得以增加,进而导致土壤容重相应降低[12]。此外,凋落物的输入在改善土壤物理性质方面亦发挥着重要作用,它能够降低土壤容重,同时增加土壤孔隙度[10]。正如预期,在恢复后期,凋落物生物量明显高于其他两个恢复阶段(表2),与之相伴的是,土壤容重低于其他两个阶段(图4-H)。
不同的森林群落由于受到不同植被类型凋落物分解以及养分释放特性的影响,在土壤理化性质方面展现出显著的差异[13]。随着恢复阶段的不断推进,土壤有机质含量呈现出显著的增加态势(图4),这一现象极有可能与凋落物输入量的增多以及微生物活动的增强存在密切关联。在森林生态系统中,土壤中的凋落物以及根系的类型和数量,共同为土壤养分的积累提供了有力支持[14]。值得注意的是,土壤有机质含量在恢复进程中的显著提升,并非个例[15-16]。本研究结果表明,次生林土壤全碳和全氮含量随着森林恢复过程的持续推进而逐渐增加,这一发现与大多数先前的研究结果一致[17]。凋落物和根系在增强土壤有机质积累的过程中扮演着重要角色[18],它们不仅促进了土壤团聚体的形成,还提升了其稳定性。而团聚体形成和稳定性的增强,能够显著优化土壤的物理性质,例如进一步降低土壤容重,增加土壤孔隙度[19]。土壤物理性质的改善,显著增强了阔叶林土壤碳氮库的稳定性,再加上阔叶林本身养分返回速度较快,这对于通过提升土壤养分水平来提高土壤肥力具有至关重要的意义[20]。这些不断增加的养分,为植物的生长提供了不可或缺的营养支持,同时也对改善土壤性质起到了积极作用。
-
热带次生林的恢复过程,对土壤持水能力产生了极为显著的提升作用,在很大程度上重塑了土壤的水文特征,尤其是在表层土壤中表现尤为突出。本研究中,随着恢复阶段的逐步推进,土壤持水能力以及表层土壤饱和导水率均呈现出明显的提高趋势。土壤孔隙度的增加,为水分的储存和运输创造了更为广阔的空间;有机质含量的提升,则进一步增强了土壤的保水能力;而容重的降低,有效减少了土壤的紧实度,这些因素共同作用,进一步优化了土壤的水文特性。土壤的物理特性和化学特性相互交织,共同对土壤持水能力产生影响[21],因此可以合理推测,土壤持水能力的差异主要是由三个恢复阶段中土壤特性的不同所造成的。近期的研究也表明,土壤持水能力与土壤孔隙度、土壤容重以及土壤有机质之间存在着紧密的联系[22]。在本研究中,土壤特性可以解释超过90%以上的土壤持水能力变化(图5),这一数据充分表明,土壤持水能力在很大程度上取决于土壤特性。此外,还进一步揭示出,毛管孔隙度和总孔隙度是控制土壤持水能力的关键因素,这一发现与之前关于高寒山区田间持水量受土壤孔隙度影响的研究结果相一致[23]。
土壤孔隙结构在决定水分进入和流经土壤的速度方面起着决定性作用[24],随着次生林恢复进程的推进,土壤容重逐渐降低,土壤孔隙度相应增加,这一系列变化最终导致了土壤持水能力的显著提高。与此同时,随着恢复阶段的不断深入,根系生物量的增加在调控土壤持水能力方面也发挥着不可或缺的重要作用。研究指出,土壤有机质含量的增加能够激活各类根际微生物,而这一过程又依赖于凋落物的丰度和分解情况。随着森林年龄的增长,土壤含水量呈现出上升趋势[25]。当土壤中有机质丰富时,微生物活动也愈发旺盛,这对林地土壤的改良效果显著,能够促使土壤团粒结构增加,土壤孔隙增多,从而为水分的传输和储存创造有利条件[26]。此外,根系的分布呈现出明显的层次性特征,主要集中在表层土壤中,深层土壤中的根系活动相对较少,这一分布特点也在一定程度上限制了深层土壤持水能力的改善。不同恢复阶段下,土壤的凋落物输入情况、土壤有机质含量以及地下生物的分布和数量等方面的变化,会对土壤机械结构产生影响,进而影响到土壤持水能力[27]。
需要强调的是,次生林不同恢复阶段土壤特性的研究,受到时间和环境的干扰较为明显。本次研究仅对该区域土壤进行了一次取样分析,因此对于时间和环境因素所导致的影响尚不明确。然而,深入研究次生林不同恢复阶段土壤理化性质与土壤持水量之间的关系,对于全面理解森林恢复的生态效应具有积极的推动作用。为了更全面、深入地了解热带次生林土壤持水能力的变化规律及其调控因素,有必要开展长期定位观测和研究分析工作。通过对该区域次生林演替过程中土壤各项性质与特性的持续监测,并将其与原始林群落土壤进行系统、细致的对比分析,能够更精确地揭示不同恢复阶段土壤性质的变化特征,从而为深入理解热带次生林恢复过程中土壤持水能力的演变规律与内在机制提供坚实的数据支撑与理论依据。
-
热带次生林的恢复对土壤水文过程及理化性质产生了显著影响。本研究揭示,随着次生林恢复进程的深入,土壤持水能力得到明显提升。本研究表明,森林恢复是增强表层土壤持水能力的有效策略。然而,随着恢复阶段的推进,深层土壤持水能力的恢复可能面临一定挑战。土壤持水能力的变化与土壤性质及根系生物量的动态变化紧密相关。随着恢复阶段的推进,土壤结构得到优化,土壤养分含量增加,而土壤容重则呈现下降趋势。这些变化共同作用于土壤持水能力,其中土壤孔隙度和容重成为决定土壤持水能力的关键因素。根系生物量通过调节土壤有机质含量和容重,在控制土壤持水能力方面发挥了重要作用。综上所述,次生林恢复后期较高的土壤持水能力主要得益于其较高的土壤孔隙度。这一结论不仅为热带原始森林的可持续管理和保护提供了科学依据,同时也为次生林的恢复实践提供了有益启示。通过深入了解次生林恢复过程中土壤持水能力的变化规律及其影响因素,可以更加精准地制定森林恢复策略,以促进土壤水资源的合理利用和森林生态系统的健康发展。
Effects of tropical secondary forest restoration on soil water-holding capacity and its driving factors
-
摘要: 近年来,受大规模采伐和刀耕火种等人类活动的影响,热带原始森林面积持续减少,逐渐形成一系列处于不同恢复阶段的热带次生林。然而,目前关于不同恢复阶段次生林土壤持水能力的演变规律及其主导调控因素,仍缺乏清晰认识。为此,本研究以海南热带次生林的三个恢复阶段(早期、中期和后期)为研究对象,采用“空间替代时间”的方法,通过测定土壤理化性质与持水性能,系统探讨不同恢复阶段对土壤持水能力的影响及其内在调控机制。研究结果表明,随恢复进程推进,(0~60 cm)土层土壤持水能力显著增强,其中(0~10 cm)土层的自然含水量、饱和持水量、毛管持水量和田间持水量均呈逐步上升趋势。同时,恢复后期(0~10 cm)土层的总孔隙度和毛管孔隙度显著高于早期和中期(P<0.05),而(0~60 cm)土层的土壤容重在恢复后期显著降低。土壤持水能力与总孔隙度、毛管孔隙度、非毛管孔隙度及有机质含量呈正相关,与容重呈负相关。其中,总孔隙度的影响最为显著,贡献率达26.95%,其次为毛管孔隙度(23.02%)和容重(21.39%);非毛管孔隙度与有机质也有一定影响,土壤颗粒组成的影响则相对较弱。综上,次生林恢复通过改善孔隙结构及提升有机质含量显著增强了土壤持水能力,且在恢复后期效果最为显著。Abstract: In recent years, due to the impact of human activities such as large-scale logging and slash-and-burn cultivation, the area of tropical primeval forests has continued to decrease, gradually forming a series of tropical secondary forests at different stages of recovery. However, there is still a lack of clear understanding of the evolution of soil water holding capacity of secondary forests and its dominant regulatory factors at different recovery stages. In this context, soil physical and chemical properties and water-holding performance of tropical secondary forests at three restoration stages (early, middle and late) in Hainan were determined to systematically analyze the effects of different restoration stages on soil water-holding capacity and its internal regulatory mechanism. The results showed that with the progress of restoration, the soil water holding capacity of the (0~60 cm) soil layer increased significantly, and that the natural water content, saturated water holding capacity, capillary water holding capacity and field water holding capacity of the (0~10 cm) soil layer gave a gradual upward trend. At the same time, the total porosity and capillary porosity of the (0~10 cm) soil layer at the late stage of restoration were significantly higher than those at the early and middle stages (P<0.05), while the soil bulk density of the (0~60 cm) soil layer decreased significantly at the late stage of restoration. Soil water holding capacity was positively correlated with total porosity, capillary porosity, non-capillary porosity and organic matter content, and negatively correlated with bulk density. The influence of total porosity was the most significant, with a contribution rate of 26.95%, followed by capillary porosity (23.02%) and bulk density (21.39%). Non-capillary porosity and organic matter also had a certain effect, while the influence of soil particle composition was relatively weak. In conclusion, the restoration of secondary forests significantly enhanced the soil water-holding capacity by improving the pore structure and increasing the content of organic matter, and the effect was most significant at the late stage of restoration.
-
图 2 不同恢复阶段次生林的土壤质地
注:ES、MS、LS分别为恢复阶段早期、中期、后期。
Fig. 2 Soil particle size distribution of secondary forests at different stages of recovery
Note: ES, MS, and LS represent the early, middle, and late stages of recovery, respectively.
图 3 不同恢复阶段次生林的土壤理化性质
注:不同小写字母表示土壤性质在同一土层不同恢复阶段的差异显著(P<0.05)。
Fig. 3 Soil physicochemical properties of secondary forests at different stages of recovery
Note: Different lowercase letters denote significant differences (P < 0.05)in soil properties among recovery stages within the same soil layer.
图 4 不同恢复阶段次生林的土壤含水量
Fig. 4 Soil water content in secondary forests at different restoration stages
图 5 不同恢复阶段次生林土壤理化性质对土壤持水能力的冗余分析
Fig. 5 Redundant analysis of soil water-holding capacity of secondary forests at different recovery stages
表 1 不同恢复阶段次生林群落特征
Table 1 Characteristics of secondary habitat communities at different restoration stages
恢复阶段
Recovery stages高度/m
Height胸径/cm
DHB郁闭度
Canopy density主要树种
Dominant tree species早期
Early stage5.64±0.15 8.49±0.32 0.73 厚皮树Lannea coromandelica
细基丸Huberantha cerasoides
黄牛木Cratoxylum cochinchinense
银柴Aporosa dioica中期
Middle stage10.98±0.75 15.84±1.72 0.80 无患子Sapindus saponaria
杂色榕Ficus variegate
锈毛野桐Mallotus anomalus
楹树Albizia chinensis
倒吊笔Wrightia pubescens后期
Late stage15.28±1.17 20.74±1.81 0.88 重阳木Bischofia polycarpa
调羹树Heliciopsis lobata
海南黄檀Dalbergia hainanensis
杂色榕Ficus variegate 下载: 导出CSV
下载: 导出CSV
表 2 不同恢复阶段次生林根和凋落物的生物量
Table 2 Biomass of secondary forest roots and litter at different stages of restoration
恢复阶段
Recovery stages根生物量/(g·m−2)
Root biomass凋落物生物量/(t·hm−2)
Litter biomass早期 Early stage 83.21 1.68 中期 Middle stage 158.29 2.83 后期 Late stage 249.75 4.45
下载: 导出CSV
表 3 不同恢复阶段次生林表层土壤(0~10 cm)的饱和导水率
Table 3 Topsoil (0~10 cm)saturated hydraulic conductivity of secondary forests at different stages of recovery
恢复阶段Recovery stages 早期 Early stage 中期 Middle stage 后期 Late stage 饱和导水率/(cm·min−1)
Saturated hydraulic conductivity16.99 17.38 18.23
下载: 导出CSV
-
[1] 丁易, 臧润国. 海南岛霸王岭热带低地雨林植被恢复动态[J]. 植物生态学报, 2011, 35(5): 577−586. https://doi.org/10.3724/SP.J.1258.2011.00577 doi: 10.3724/SP.J.1258.2011.00577 [2] Ayala-Orozco B, Gavito M E, Mora F, et al. Resilience of soil properties to land-use change in a tropical dry forest ecosystem [J]. Land Degradation & Development, 2018, 29(2): 315−325. https://doi.org/10.1002/ldr.2686 doi: 10.1002/ldr.2686 [3] Yuan Y, Li X Y, Xiong D H, et al. Effects of restoration age on water conservation function and soil fertility quality of restored woodlands in phosphate mined-out areas [J]. Environmental Earth Sciences, 2019, 78(23): 653. https://doi.org/10.1007/s12665-019-8671-8 doi: 10.1007/s12665-019-8671-8 [4] 邹碧, 王刚, 杨富权, 等. 华南热带区不同恢复阶段人工林土壤持水能力研究[J]. 热带亚热带植物学报, 2010, 18(4): 343−349. https://doi.org/10.3969/j.issn.1005-3395.2010.04.001 doi: 10.3969/j.issn.1005-3395.2010.04.001 [5] 徐小牛, 邓文鑫, 张赟齐, 等. 安徽老山亚热带常绿阔叶林不同林龄阶段土壤特性及其水源涵养功能的变化[J]. 水土保持学报, 2009, 23(1): 177−181. https://doi.org/10.3321/j.issn:1009-2242.2009.01.038 doi: 10.3321/j.issn:1009-2242.2009.01.038 [6] 吴胜安, 邢彩盈, 朱晶晶. 海南岛气候特征分析[J]. 热带生物学报, 2022, 13(4): 315−323. https://doi.org/10.15886/j.cnki.rdswxb.2022.04.001 doi: 10.15886/j.cnki.rdswxb.2022.04.001 [7] 程思源, 陈俏艳, 乔栋, 等. 海南热带雨林不同林分凋落物储量及其持水特性[J]. 热带地理, 2024, 44(4): 700−708. https://doi.org/10.13284/j.cnki.rddl.003853 doi: 10.13284/j.cnki.rddl.003853 [8] Sun L, Yang L, Chen L D, et al. Tracing the soil water response to autumn rainfall in different land uses at multi-day timescale in a subtropical zone [J]. CATENA, 2019, 180: 355−364. https://doi.org/10.1016/j.catena.2019.05.013 doi: 10.1016/j.catena.2019.05.013 [9] Zhang B, Hu G, Xu C H, et al. Effects of natural vegetation restoration on soil physicochemical properties in tropical karst areas, southwestern China [J]. Forests, 2024, 15(7): 1270. https://doi.org/10.3390/f15071270 doi: 10.3390/f15071270 [10] Li P, Qi S, Zhang L, et al. The effect of different vegetation restoration types on soil quality in mountainous areas of Beijing [J]. Forests, 2023, 14(12): 2374. https://doi.org/10.3390/f14122374 doi: 10.3390/f14122374 [11] Zhang Y H, Xu X L, Li Z W, et al. Improvements in soil quality with vegetation succession in subtropical China karst [J]. Science of the Total Environment, 2021, 775: 145876. https://doi.org/10.1016/j.scitotenv.2021.145876 doi: 10.1016/j.scitotenv.2021.145876 [12] Jiao F, Wen Zm, An S S. Changes in soil properties across a chronosequence of vegetation restoration on the Loess Plateau of China [J]. CATENA, 2011, 86(2): 110−116. https://doi.org/10.1016/j.catena.2011.03.001 doi: 10.1016/j.catena.2011.03.001 [13] Danise T, Innangi M, Curcio E, et al. Covariation between plant biodiversity and soil systems in a European beech forest and a black pine plantation: the case of Mount Faito, (Campania, southern Italy) [J]. Journal of Forestry Research, 2022, 33(1): 239−252. https://doi.org/10.1007/s11676-021-01339-3 doi: 10.1007/s11676-021-01339-3 [14] Morrison I K, Foster N W. Fifteen-year change in forest floor organic and element content and cycling at the Turkey Lakes watershed [J]. Ecosystems, 2001, 4(6): 545−554. https://doi.org/10.1007/s10021-001-0028-x doi: 10.1007/s10021-001-0028-x [15] Chen S T, Feng X, Lin Q M, et al. Pool complexity and molecular diversity shaped topsoil organic matter accumulation following decadal forest restoration in a karst terrain [J]. Soil Biology and Biochemistry, 2022, 166: 108553. https://doi.org/10.1016/j.soilbio.2022.108553 doi: 10.1016/j.soilbio.2022.108553 [16] Bárcena T G, Kiær L P, Vesterdal L, et al. Soil carbon stock change following afforestation in northern Europe: a meta-analysis [J]. Global Change Biology, 2014, 20(8): 2393−2405. https://doi.org/10.1111/gcb.12576 doi: 10.1111/gcb.12576 [17] 袁荣利, 龙翠玲, 何庆, 等. 茂兰喀斯特森林不同恢复阶段植物叶片-土壤碳氮磷生态化学计量特征[J]. 应用与环境生物学报, 2025, 31(3): 518−526. https://doi.org/10.19675/j.cnki.1006-687x.2024.12008 doi: 10.19675/j.cnki.1006-687x.2024.12008 [18] Zagyvai-Kiss K A, Kalicz P, Szilágyi J, et al. On the specific water holding capacity of litter for three forest ecosystems in the eastern foothills of the Alps [J]. Agricultural and Forest Meteorology, 2019, 278: 107656. https://doi.org/10.1016/j.agrformet.2019.107656 doi: 10.1016/j.agrformet.2019.107656 [19] Chen C F, Liu W J, Jiang X J, et al. Effects of rubber-based agroforestry systems on soil aggregation and associated soil organic carbon: implications for land use [J]. Geoderma, 2017, 299: 13−24. https://doi.org/10.1016/j.geoderma.2017.03.021 doi: 10.1016/j.geoderma.2017.03.021 [20] Shao G D, Ai J J, Sun Q W, et al. Soil quality assessment under different forest types in the Mount Tai, central eastern China [J]. Ecological Indicators, 2020, 115: 106439. https://doi.org/10.1016/j.ecolind.2020.106439 doi: 10.1016/j.ecolind.2020.106439 [21] Zimmermann B, Elsenbeer H. Spatial and temporal variability of soil saturated hydraulic conductivity in gradients of disturbance [J]. Journal of Hydrology, 2008, 361(1/2): 78−95. https://doi.org/10.1016/j.jhydrol.2008.07.027 doi: 10.1016/j.jhydrol.2008.07.027 [22] Qin D X, Xu R R, Wang C X, et al. Vegetation restoration improves soil hydrological properties by regulating soil physicochemical properties in the Loess Plateau, China [J]. Journal of Hydrology, 2022, 609: 127730. https://doi.org/10.1016/j.jhydrol.2022.127730 doi: 10.1016/j.jhydrol.2022.127730 [23] Pan T, Hou S, Wu S H, et al. Variation of soil hydraulic properties with alpine grassland degradation in the eastern Tibetan Plateau [J]. Hydrology and Earth System Sciences, 2017, 21(4): 2249−2261. https://doi.org/10.5194/hess-21-2249-2017 doi: 10.5194/hess-21-2249-2017 [24] Uteau D, Pagenkemper S K, Peth S, et al. Root and time dependent soil structure formation and its influence on gas transport in the subsoil [J]. Soil and Tillage Research, 2013, 132: 69−76. https://doi.org/10.1016/j.still.2013.05.001 doi: 10.1016/j.still.2013.05.001 [25] Li Y L, Yang F F, Ou Y X, et al. Changes in forest soil properties in different successional stages in lower tropical China [J]. PLoS One, 2013, 8(11): e81359. https://doi.org/10.1371/journal.pone.0081359 doi: 10.1371/journal.pone.0081359 [26] 朱万泽, 盛哲良, 舒树淼. 川西亚高山次生林恢复过程中土壤物理性质及水源涵养效应[J]. 水土保持学报, 2019, 33(6): 205−212. https://doi.org/10.13870/j.cnki.stbcxb.2019.06.029 doi: 10.13870/j.cnki.stbcxb.2019.06.029 [27] 祁凯斌, 黄俊胜, 杨婷惠, 等. 亚高山森林自然与人工恢复对土壤涵水能力的影响[J]. 生态学报, 2018, 38(22): 8118−8128. https://doi.org/10.5846/stxb201711302159 doi: 10.5846/stxb201711302159 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 225
- HTML全文浏览量: 130
- 被引次数: 0
