


-
微生物生长环境多样,在生物进化过程中形成了自己独特的生长代谢机制。耐热真菌是一类最高生长温度在50℃左右(通常为40℃或40℃以上),最低生长温度在20℃以下的特殊真菌类群[1],极端环境(高温、低温、强酸、强碱、高盐等)微生物有自己特定的代谢方式、独特基因类型及特殊生理机制,耐热真菌是可以在高温条件下生长的适应极端环境的真核微生物,其特殊生长代谢机制使产生特殊代谢产物、获取新生物技术手段成为可能[2]。耐热真菌的分布范围广泛,世界上已描述和报道的耐热真菌有89种,我国报道的有23种[3]。Salar等[4]从印度北部的温带土壤中分离出19种嗜热和耐热真菌。Michele等[5]从番石榴叶中分离得到1株耐热真菌拟青霉(Paecilomyces variotii),并从拟青霉中分离出以55℃为最适反应温度,pH为5.0的热稳定性糖化酶。詹发强等[6]从新疆棉花秸秆高温堆肥中分离得到的耐热真菌Z1、Z2在降解农作物废弃秸秆中具有较高的应用价值。耐热真菌的生理生长特点是耐热性(最高温度、最适温度、最低温度),这是区分耐热真菌与非耐热真菌的重要依据[2]。另外,通过菌株分离经形态学鉴定等手段对耐热真菌的种类鉴别意义重大。目前对于耐热真菌的研究国内外把重心放在菌种鉴定和生理特性研究上,而对产物活性研究报道不多。由于生长环境的不同,来源各异的耐热菌存在特有的生长温度、pH值、盐浓度等的耐受性和其发酵产物的抗菌活性之间会有较大不同,深入研究可丰富相关基础理论。
我国国土面积辽阔,气候类型丰富多样,为耐热真菌提供良好生长环境。湖光岩玛珥湖位于广东省湛江市湖光岩风景区内,是经火山喷发而形成的独特湖泊,其受外界干扰小,特殊的生境为生物进化形成新菌种提供可能。秦青英等[7]研究湖光岩玛珥湖浮游细菌组成时发现其菌落结构较为独特,微生物资源丰富且新颖。刘颖等[8]分离了湖光岩土壤放线菌,并测定了其抗菌活性,分离出23株菌,且其中60.9%有抗菌活性。耐热真菌的来源多种多样,这表明极有可能从湖光岩独特的地理环境中发现耐热真菌。耐热真菌有产酶等方面潜力,故以湖光岩玛珥湖为开发来源的菌株的特殊功能和活性值得探究和开发。本研究以广东省湛江市湖光岩玛珥湖分离得到1株耐热真菌HS1-1为出发菌株,对该菌进行形态学及分子水平鉴定,研究其生理特性,产酶能力及抗菌活性,以期为对该菌的进一步研究与开发提供基础。
-
2020年11月3日从湛江市湖光岩风景区距离湖面1~2 m的岸上5~15 cm深度采集的土壤中分离获得菌株,经44℃初筛和19℃复筛[9-10],确定为耐热真菌。在取样分离中以耐热真菌条件共分离4株耐热真菌, 选择其中的HS1-1菌株为供试菌株。
抗菌活性指示菌:交替假单胞菌属SL-1(Pseudoalteromonas sp.,登录号:FJ404757.1)、吉氏库特氏菌属SL-2(Kurthia gibsonii strain,登录号:EF032685.1)均为笔者所在的课题组从冷冻虾仁中分离鉴定保存的优势腐败菌;大肠杆菌(Escherichia coli)、铜绿假单胞菌(Pseudomonas aeruginosa)、金黄色葡萄球菌(Staphylococcus aureus)均为笔者所在实验室保存的菌。
-
马铃薯葡萄糖琼脂培养基(PDA):广东环凯微生物科技有限公司;马铃薯葡萄糖肉汤(PDB):杭州微生物试剂有限公司;改良马丁琼脂培养基:广东环凯微生物科技有限公司;改良马丁培养基:北京陆桥技术股份有限公司;营养琼脂:北京陆桥技术股份有限公司。淀粉酶培养基[11]、糖苷酶培养基[12]、纤维素酶培养基[13]、壳聚糖酶培养基[14]4种产酶筛选培养基配制方法参照文献并稍作修改,具体配方见表1。
表 1 产酶培养基
产酶培养基 配 方 淀粉酶培养基 可溶性淀粉20.0 g·L−1,KCl 0.5 g·L−1,NaNO3 2.0 g·L−1,K3PO4 1.0 g·L−1,MgSO4·7H2O 0.5 g·L−1,琼脂18.0 g·L−1,海水晶30.0 g·L−1,ddH2O 1000 mL 糖苷酶培养基 乳糖 10.0 g·L−1,K2HPO4 1.0 g·L−1,FeSO4 0.01 g·L−1,NaNO3 3.0 g·L−1,KCl 0.5 g·L−1,MgSO4·7H2O 0.5 g·L−1,琼脂15.0 g·L−1,dd H2O 1000 mL 纤维素酶培养基 羧甲基纤维素8.0 g·L−1,KCl 0.5 g·L−1,NaNO3 3.0 g·L−1,FeSO4 0.01 g·L−1,K2HPO4 1.0 g·L−1,MgSO4·7H2O 0.5 g·L−1,氯霉素0.05 g·L−1,琼脂20.0 g·L−1,ddH2O 1000 mL,pH6.0 壳聚糖酶培养基 壳聚糖10.0 g·L−1,CaCO3 6.0 g·L−1,MgSO4·7H2O 0.25 g·L−1,(NH4)2SO4 2.5 g·L−1,K2HPO4 0.02 g·L−1,NaCl 4.0 g·L−1,琼脂20.0 g·L−1,pH6.0 -
用点植法将菌株HS1-1分别接种于PDA和改良马丁琼脂培养基中,于35℃恒温培养箱中培养2~5 d,观察菌落形态特征。
从培养基挑取少许菌丝,用乳酸石炭酸绵蓝染液染色,压片法镜检,初步鉴定菌株[15]并完成的个体形态观察。
将活化到第三代的菌株HS1-1送至生工生物工程(上海)股份有限公司进行PCR扩增、测序,完成分子鉴定。分子鉴定的具体内容:分子生物学鉴定即采用内部转录间隔区(ITS)测序的方法,选用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′),采用菌落PCR的方法对筛选得到的耐热真菌HS1-1的ITS序列进行扩增。PCR扩增体系:引物ITS1和ITS4各1 µL,10 X PCR Buffer、dNTP (each 10 mmol·L−1)、Taq Plus DNA Polymerase(5 U·µL−1)共12.5 µL,50 mmol·L−1 MgSO4和Template (DNA)各1 µL,ddH2O 9.5 µL,总体积为25 µL;PCR扩增条件:95℃ 5 min;94℃ 30 s,57℃ 30 s,72℃ 90 s,共30次循环;72℃ 10 min。将PCR产物的测序结果用NCBI数据库进行BLAST对比分析,查找亲缘性近、相似性高的菌株基因序列。采用MEGA 7软件中N-J法(neighbor-joining method)自举1000次,确定菌种种属并建立系统发育树。
-
采用点植法将菌株接种于改良马丁琼脂培养基中,倒置于15、20、25、30、35、40、45、50℃恒温培养箱培养7 d,于 HH.S11-2电热恒温水浴锅55℃培养7 d,每个温度条件设置3个平行,采用十字交叉法测量菌落直径并观察菌落形态。
-
在改良马丁琼脂培养基中分别加入0、1%、2%、3%、4%、5%、6%、8%、10%的NaCl,用点植法接种菌株于培养基上,并置于35℃下培养,每个盐浓度设置3个平行,测量菌落直径并观察菌落形态。
-
调节改良马丁琼脂培养基的pH为2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0。用点植法接种菌株于培养基上,并置于35℃下培养,每个梯度设置3个平行重复,测量菌落直径及观察菌落形态。
-
将菌株HS1-1分别接种到4种产酶筛选培养基中,倒置于恒温培养箱中35℃培养3~7 d,每种产酶培养基做3个平行。壳聚糖酶培养基能力的确定是通过观察菌落周围是否产生透明圈[14],淀粉酶结果通过碘液染色观察水解圈的大小来确定;检测糖苷酶以菌落周围有无蓝色产生为依据,有则说明有产糖苷酶的能力,反之,则无。纤维素酶培养基需要用95%乙醇配制的0.1%刚果红染色液染色,再用1 mol·L−1氯化钠脱色,染色和脱色时间均为1 h[16],观察是否出现脱色圈。
-
将活化好的菌株HS1-1挑取一环分别接种到装有100 mL PDB和改良马丁培养基的250 mL三角瓶中,35℃,135 r·min−1摇床(HZQ-F160振荡培养箱)培养7 d,经真空抽滤和微孔滤膜过滤后,取部分发酵液,用N-1100D-WB旋转蒸发仪浓缩5倍,发酵原液和浓缩液均用于抗菌活性测定。
抗菌活性测定: 挑取活化好的指示菌,接种到生理盐水中,振荡均匀并稀释不同倍数,制备成OD600值为0.6的菌悬液[17]。在提前制备好的素琼脂平板中放好无菌牛津杯,倒入含有1∶100比例菌悬液的营养琼脂,待凝固后拔出牛津杯并注入100 µL发酵液或发酵浓缩液,于4℃冰箱中扩散3~4 h[18],置于37℃培养24 h并测量抑菌圈直径,抑菌圈直径算上牛津杯直径(牛津杯直径为7.8 mm),无抑菌圈以牛津杯直径计。
-
对菌落直径、抑菌圈直径等数据均利用JMP Pro软件进行显著性差异分析。取显著性水平α为0.05,通过单因素方差分析,用所有对Tukey HSD来比较均值,P<0.05表示有显著性差异,用不同英文字母表示具有显著性差异,最后采用Origin软件进行绘图。
-
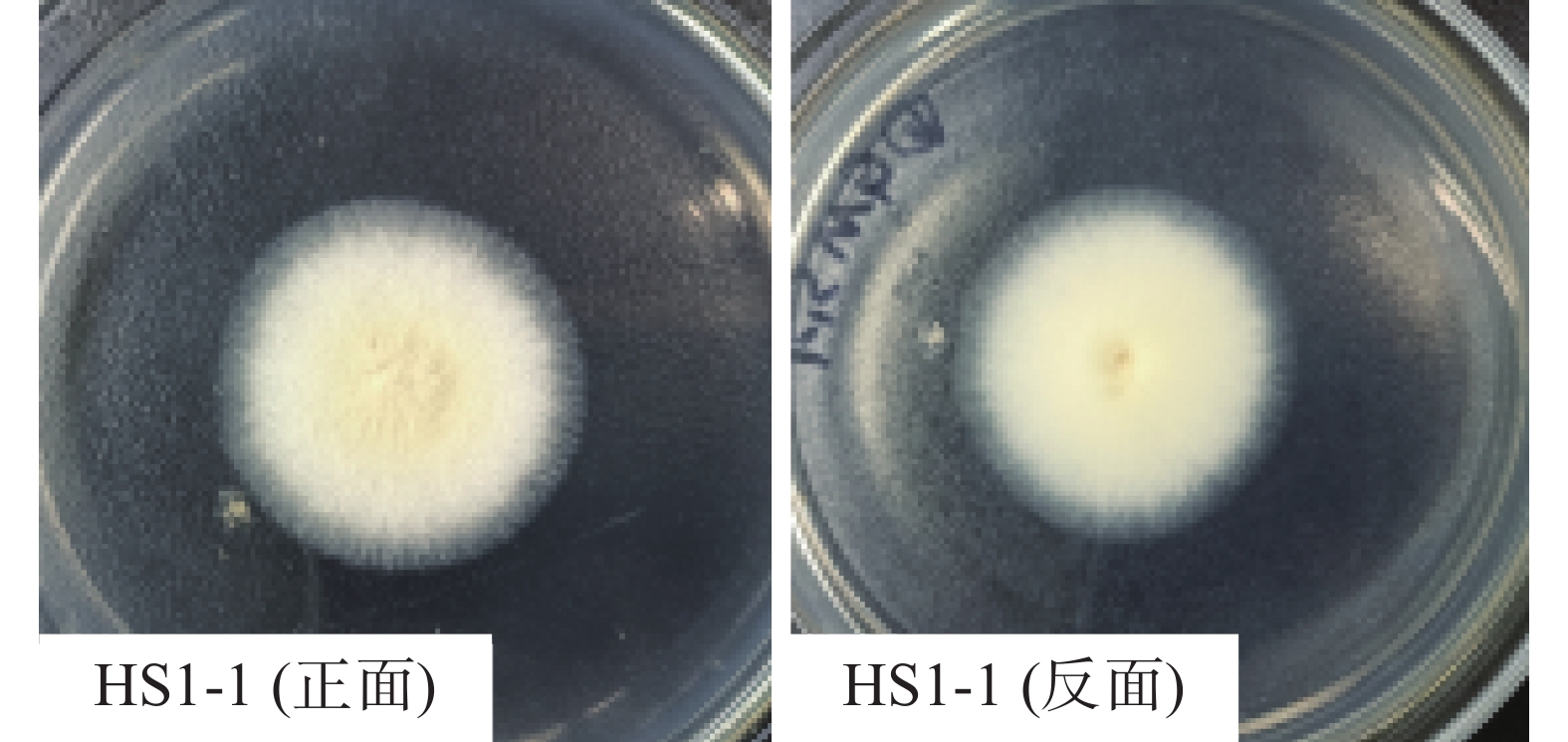
耐热真菌HS1-1在PDA和改良马丁琼脂培养基上均能生长,且在改良马丁琼脂培养基上生长较为良好,因而用改良马丁琼脂培养基作为菌株培养的最适固体培养基。从图1可见,35℃培养2~3 d的菌落形态观察中发现菌落圆形平坦,菌丝自中央向外周延展,由致密到疏松,菌丝发达呈米黄色,有放射性皱褶和晕圈。随着不同培养天数、pH、温度和盐浓度变化,菌落呈现轻微颜色差别,或烟绿色或米黄色。

图 1 改良马丁琼脂培养基2 d菌落形态
-
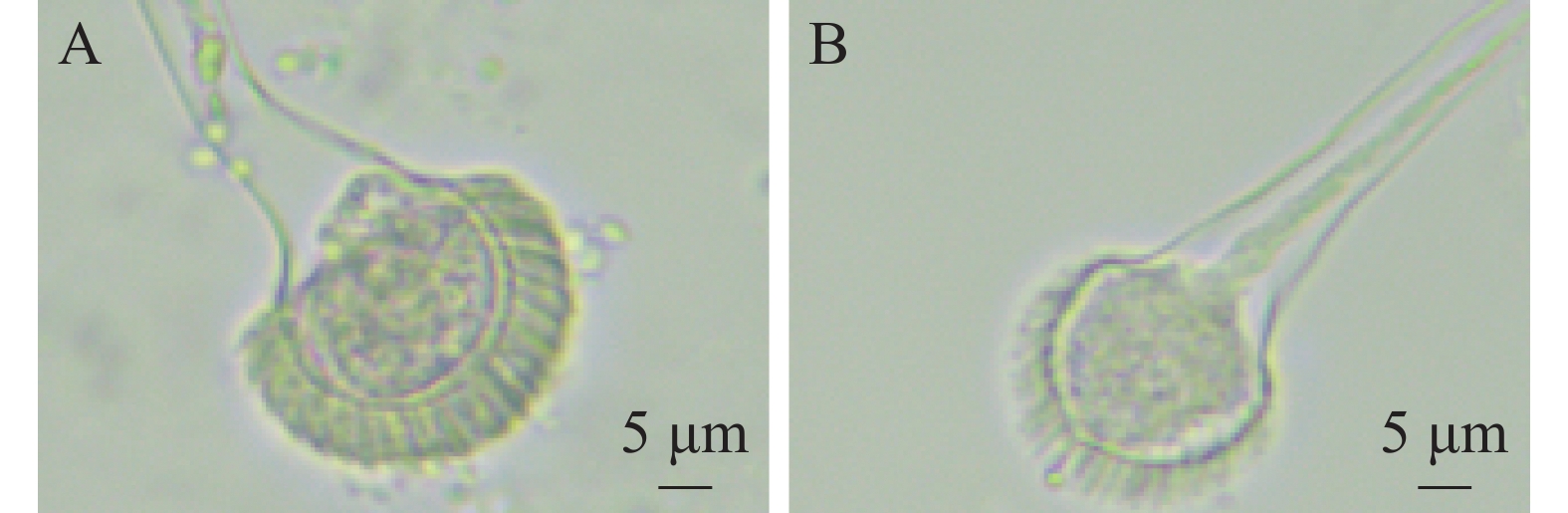
经显微镜观察(图2)发现,菌株HS1-1培养2 d后菌丝透明且显青色,有分生孢子梗和分生孢子头,顶囊成烧瓶状,直径约20 µm,小梗单层排列,成木栅状,孢子为球型,直径约2.5 µm。初步鉴定菌株HS1-1为曲霉属。王世苗等[19]观察烟曲霉呈绒状,颜色为暗烟绿色;电子显微镜结果观察到球形、绿色的分生孢子,分生孢子梗光滑,顶囊呈烧瓶状,与本研究菌株相似。
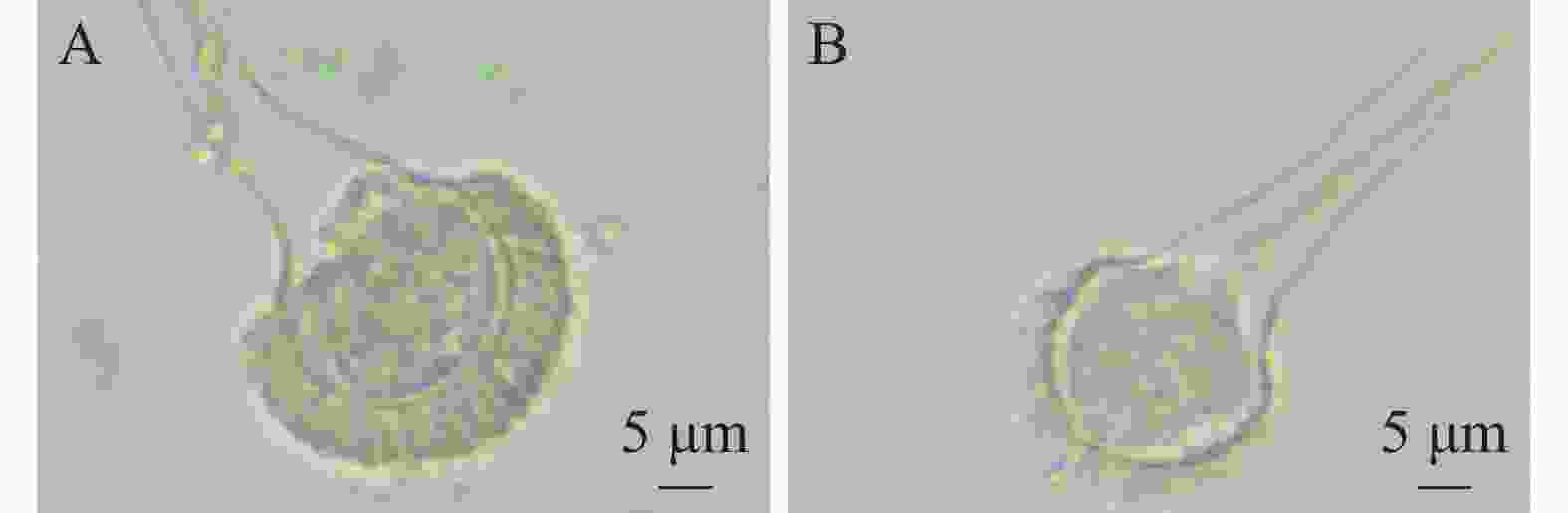
图 2 菌株HS1-1分生孢子头形态(10×40)
-
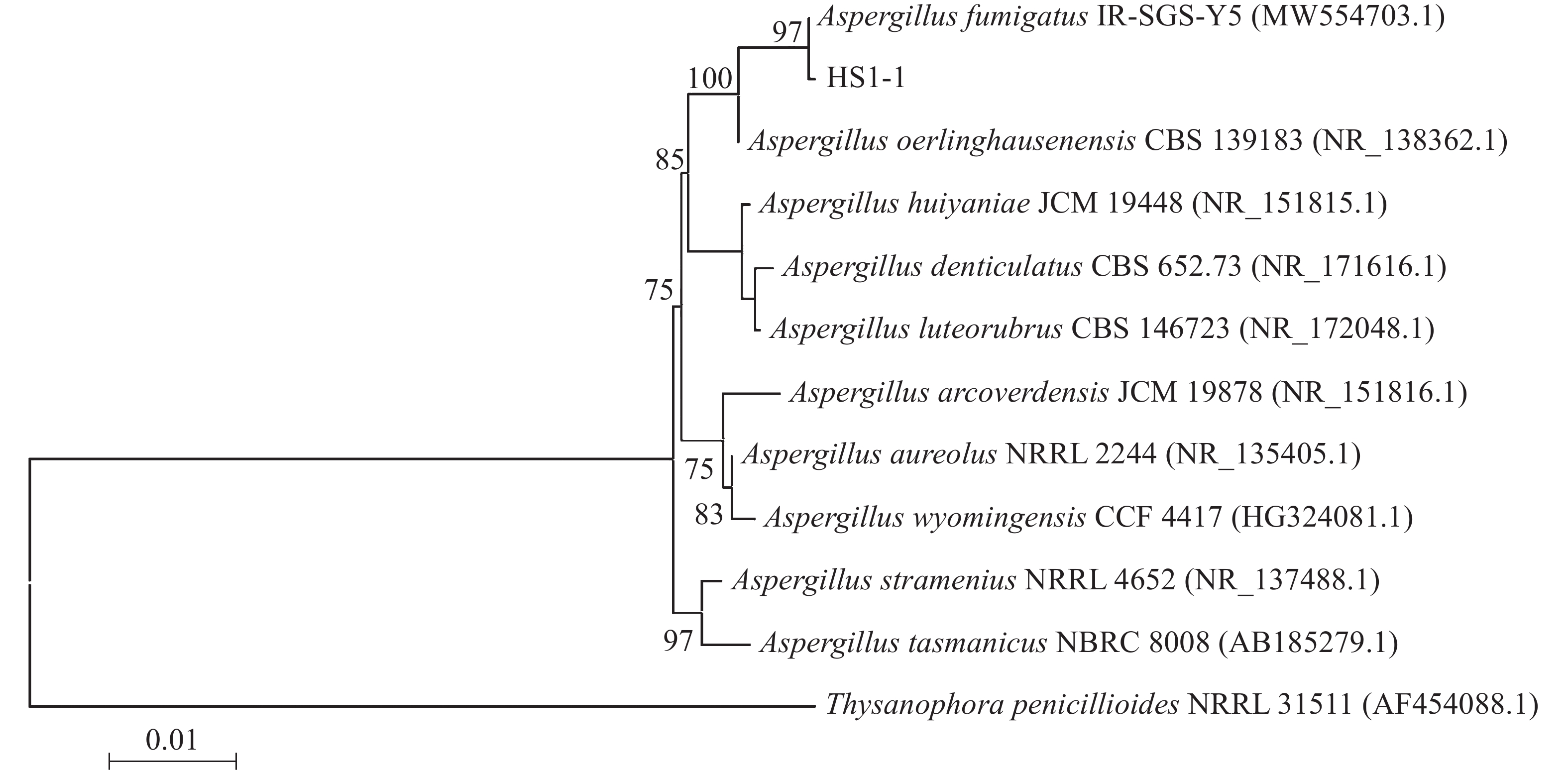
经基因组DNA提取,PCR扩增和凝胶电泳观察,菌株HS1-1在500~600 bp有清晰条带。提交ITS序列至GenBank获得登录号,利用BLAST进行同源性对比分析,构建系统发育树(图3)。HS1-1(登录号为MZ413293)与Aspergillus fumigatus IR-SGS-Y5聚在同一分支上,遗传进化距离相近。结合形态学观察和分子鉴定,确定该耐热真菌为烟曲霉。

图 3 NJ法构建HS1-1的ITS序列系统发育树
-
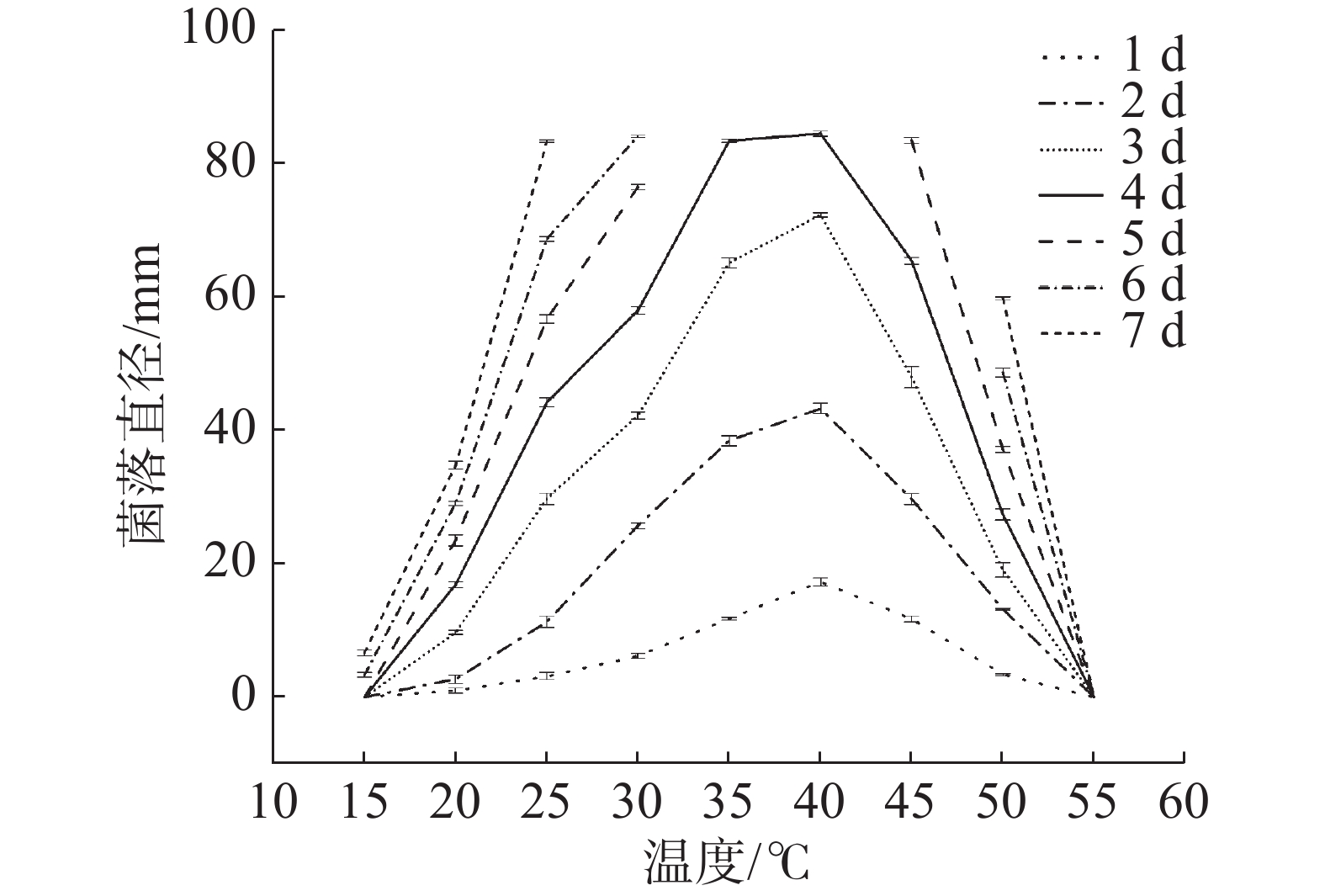
不同温度条件下菌株生长情况见图4。菌株HS1-1在15~50℃的温度范围内均能生长,15℃生长最缓慢,55℃则无法生长。从1~3 d可看出,40℃时生长速度明显高于其他温度,其次是35℃,因此该菌最适生长温度为40℃,该菌在生长温度范围内的生长能力强弱顺序为40℃、35℃、45℃、30℃、25℃、50℃、20℃、15℃。最适温度是40℃,50℃仍能生长,具有明显耐热性,为耐热真菌。烟曲霉最适生长温度一般为37℃[20],有研究发现烟曲霉在37℃培养时菌落直径最大,且在50℃、55℃仍能生长[21],这与本研究生长情况相似。观察不同温度菌落形态,在设置的温度范围内,较高或者较低温度条件下菌丝附着能力减弱,孢子易脱落,菌落沟纹加深,颜色较浅。
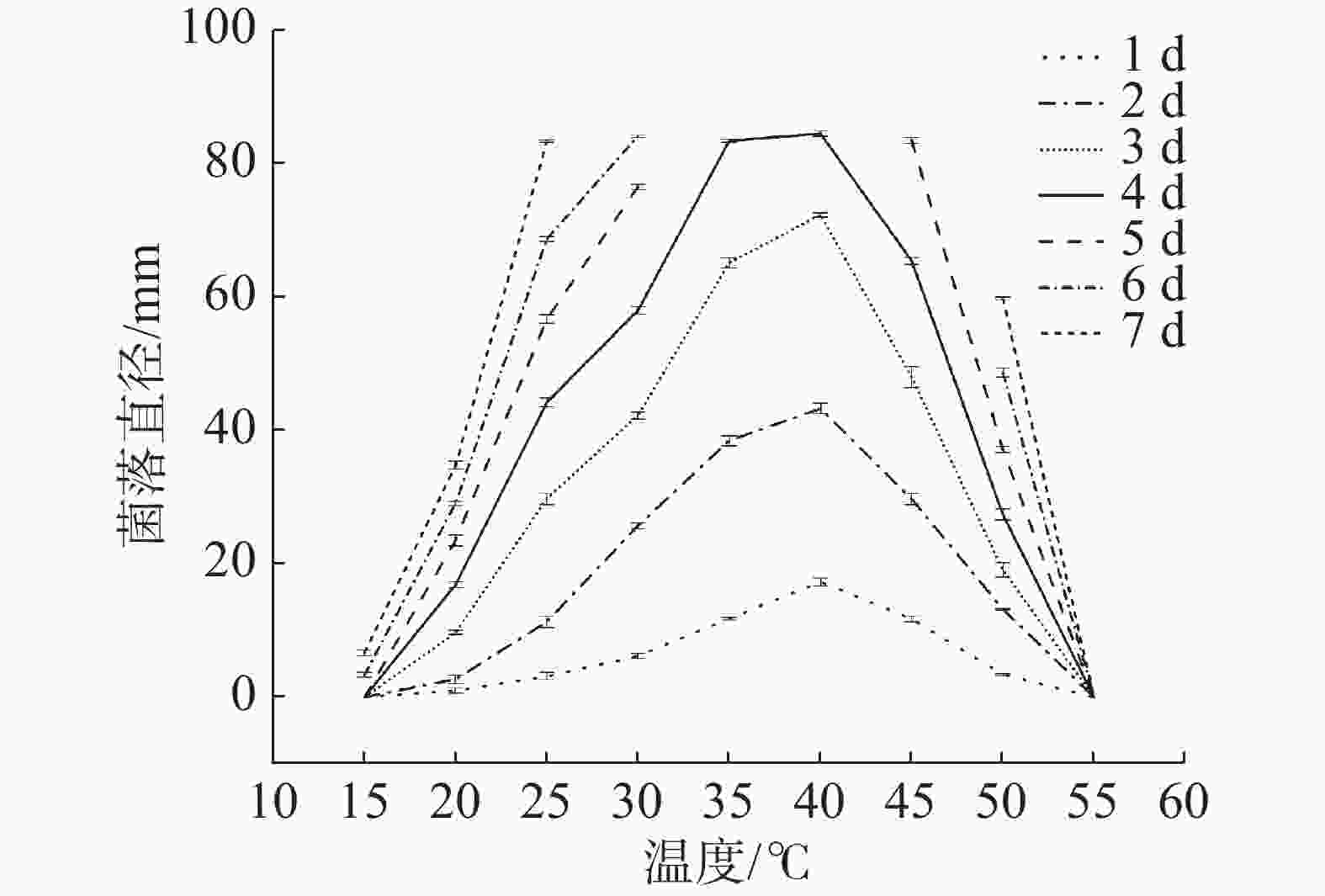
图 4 不同温度条件对菌株HS1-1生长的影响
-
菌株HS1-1在不同盐浓度的生长情况见表2。在35℃时菌株在0~10%的盐浓度范围内均能生长,2~3 d时1%~2%盐浓度明显优于其他组,4 d时盐浓度0~4%的菌落直径均大于84.3 mm,接近平板直径。故该菌适宜盐浓度范围为0~4%,最佳生长盐浓度是1%~2%,具备一定耐盐性。观察菌株第7天的生长形态发现,不同盐浓度的菌落形态差异不大,菌落中央向外周扩散呈现黄色由深变浅。
表 2 不同盐浓度对菌株HS1-1生长的影响
盐浓度/% 菌落直径/ mm 1 d 2 d 3 d 4 d 5 d 6 d 7 d 0 11.60±0.21a 37.75±0.20d 63.50±0.30c 84.56±0.31a 1 10.89±0.45ab 44.91±0.53a 70.36±0.37a 84.92±0.19a 2 10.53±0.73b 43.74±0.03b 69.69±0.76a 84.58±0.40a 3 10.95±0.14ab 41.09±0.30c 65.78±0.19b 84.33±0.11a 4 10.40±0.49b 37.83±0.25d 60.40±0.95d 84.79±0.15a 5 9.05±0.07c 34.31±0.25e 55.98±0.52e 79.38±0.83b 84.30±0.28a 6 7.43±0.04d 27.54±0.46f 49.24±0.19f 70.60±0.52c 81.74±0.58b 83.16±0.19a 8 4.80±0.21e 22.37±0.29g 39.63±0.36g 61.37±0.26d 74.93±0.90c 83.66±0.29a 10 2.07±0.30f 13.93±0.45h 26.18±0.07h 40.31±0.06e 52.31±0.66d 63.38±0.38b 76.20±0.52 注:标记不同字母表示具有显著性差异(P<0.05);空白表示该天数菌株长满平板(下同)。 -
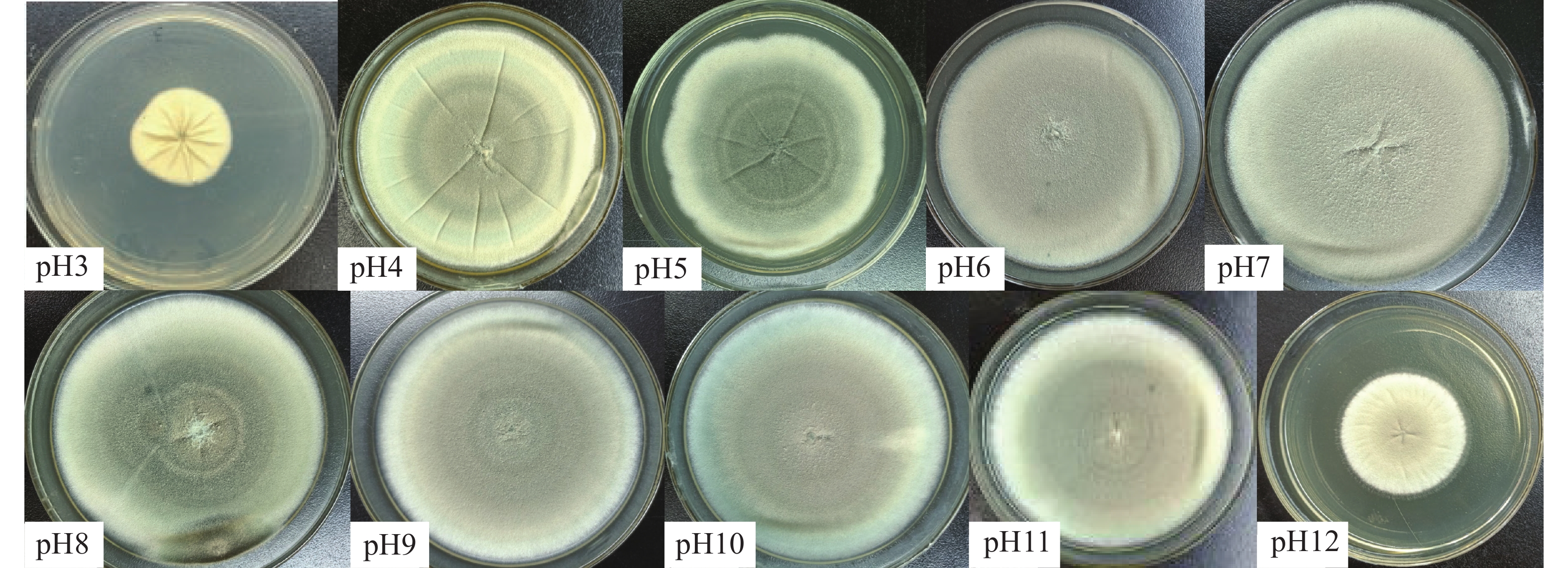
不同pH条件下菌株生长情况见表3。该菌在pH为3~12的范围内均能生长,pH在6~9时长势良好,pH2时菌株不生长。1~4 d时pH为6和7时菌落直径显著大于其他pH(P<0.05),表明该菌最适宜pH范围为6~7,且适应酸碱范围广,具有极强耐酸碱能力。对比酸性条件(pH2~pH3),菌株在较碱性条件(pH11~pH12)生长更具优势,说明耐强碱能力强。不同pH菌落形态各异(图5),pH3倒置培养时观察到菌落中央向下生长,呈突起状。pH为3~5时,菌落颜色较深,褶皱明显,菌落边缘不规整;pH≥6时,菌落颜色较浅,褶皱较少或无褶皱,菌落多呈圆形。
表 3 不同pH条件对菌株HS1-1生长的影响
pH 菌落直径/ mm 1 d 2 d 3 d 4 d 5 d 6 d 7 d 2 0.00±0.00f 0.00±0.00h 0.00±0.00g 0.00±0.00j 0.00±0.00h 0.00±0.00f 0.00±0.00d 3 1.95±0.27e 7.89±0.60f 14.28±0.72f 19.95±0.19i 25.49±0.67g 30.22±0.51e 35.57±0.46d 4 9.67±0.22ab 28.38±0.28c 48.43±0.49c 63.49±0.21d 79.83±0.15b 85.26±0.88a 5 9.27±0.79bc 24.68±0.89d 36.87±0.48d 45.82±0.97f 59.47±0.72d 66.27±0.89c 76.93±0.65b 6 10.45±0.15ab 39.57±0.06a 61.31±0.30a 82.45±0.31a 84.64±0.11a 7 10.84±0.36a 39.62±0.70a 60.96±0.78a 81.11±0.44a 84.69±0.19a 8 8.02±0.95c 30.81±0.58b 52.84±0.78b 76.25±0.79b 84.25±0.10a 9 5.97±0.34d 27.78±0.59c 49.75±0.60c 73.82±0.59c 84.23±0.13a 10 1.92±0.28e 9.69±0.88e 28.00±0.46e 50.98±0.46e 71.29±0.59c 83.50±0.44a 11 1.26±0.15e 2.29±0.35g 15.25±0.72f 36.34±0.5g 56.58±0.28e 76.42±0.83b 83.44±0.24a 12 0.95±0.14ef 1.75±0.19g 6.57±0.31g 25.30±0.15h 41.80±0.23f 46.75±0.67d 53.12±0.54c 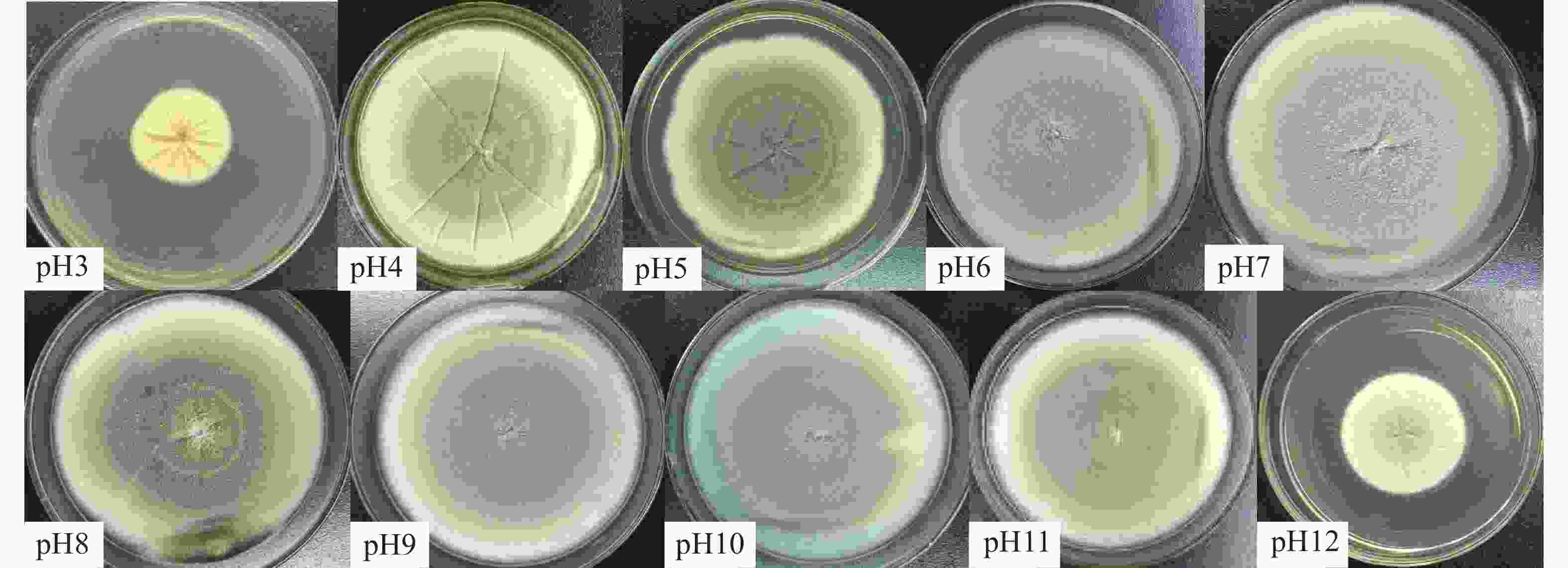
图 5 HS1-1在不同pH条件下第7天菌落形态
-
耐热真菌HS1-1在不同产酶筛选培养基的菌落生长情况见图6。微生物不能直接利用淀粉、蛋白质和脂肪等大分子,必须依靠产生的胞外酶将大分子物质分解后才能利用。菌株在4种产酶培养基上均能生长,说明该菌能产生相应的酶并以可溶性淀粉、乳糖、羧甲基纤维素和壳聚糖作为唯一碳源供自身生长,且利用可溶性淀粉、乳糖和羧甲基纤维素的能力较强,利用壳聚糖的能力较弱。
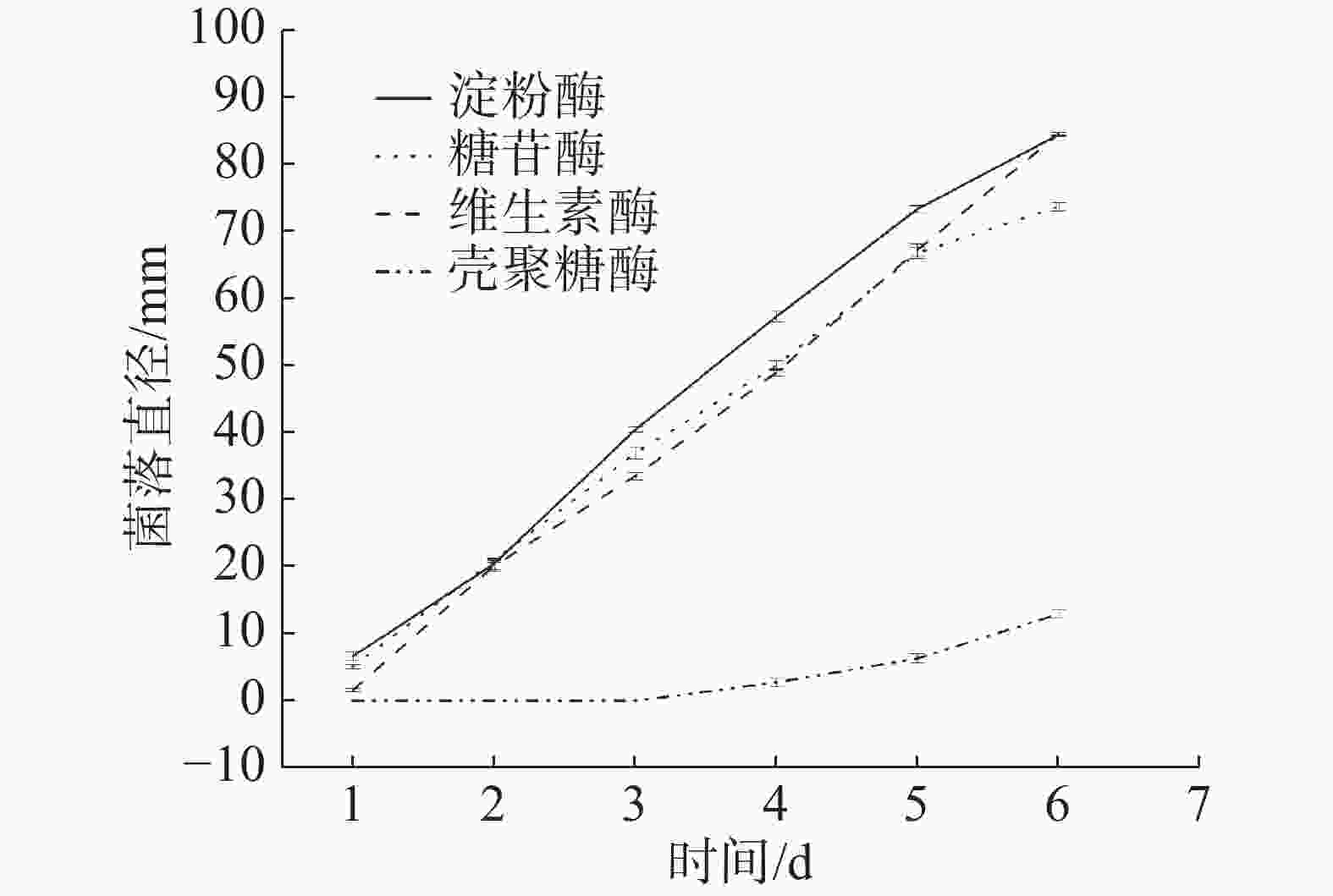
图 6 HS1-1在不同产酶筛选培养基生长情况
-
菌株HS1-1在PDB和改良马丁培养基上所得的发酵液及发酵浓缩液(5倍浓缩)的抗菌情况见表4。该菌的PDB发酵产物对5种革兰氏阴性和革兰氏阳性菌均无明显抑制效果;PDB发酵液经5倍浓缩后均产生抑菌圈,推断可能由于抑菌活性物质富集而出现明显抑菌效果;在改良马丁培养基所得的发酵液对SL-1、大肠杆菌、铜绿假单胞菌、SL-2均有抗菌活性,而对金黄色葡萄球菌无明显抗菌活性;改良马丁培养基发酵液经5倍浓缩后,以SL-1为指示菌时抑菌效果减弱,可能是在旋转蒸发过程中高温(50℃)破坏了部分抑制SL-1指示菌的活性成分,但其他指示菌的结果显示抑菌效果都增强了。
表 4 不同培养基发酵液与发酵浓缩液抗菌活性比较
指示菌 在PDB上的抑菌圈直径/mm 在改良马丁培养基上的抑菌圈直径/mm 发酵液 发酵浓缩液 发酵液 发酵浓缩液 SL-1 7.80±0.00a 15.05±0.99a 14.26±0.63c 12.40±0.40d 大肠杆菌 7.80±0.00a 13.21±0.56a 20.83±0.59a 21.29±0.08b 铜绿假单胞菌 7.80±0.00a 14.15±0.51a 13.51±0.24c 18.23±0.73c SL-2 7.80±0.00a 12.97±0.19a 15.40±0.29b 24.82±0.18a 金黄色葡萄球菌 7.80±0.00a 13.16±0.35a 7.80±0.00d 11.00±0.58e -
本研究以1株湖光岩玛珥湖来源的耐热真菌HS1-1为研究对象,观察其菌落特征,以菌丝形态、孢子、顶囊等特征初步判断该菌为烟曲霉,结合分子水平鉴定确定该菌为烟曲霉。不同温度、盐浓度、pH条件下菌株的生长研究结果表明,耐热烟曲霉HS1-1温度生长范围较广,15~50℃均能生长,有学者研究发现烟曲霉在30~42℃都能较好生长[22],这与本研究结果相符合,后续实验可以继续补充探究15℃以下温度条件以探究该菌的最低生长温度。张晋卿等[23]发现烟曲霉AF293在酵母浸膏培养基中的最适宜生长温度是37℃, 而在改良马丁琼脂培养基中耐热烟曲霉HS1-1的最适生长温度在40℃,说明湖光岩玛珥湖来源的烟曲霉在特定适宜培养基上有更高的最适生长温度,且在高温条件下更具生长优势。菌株HS1-1的最适盐浓度范围在1%~2%,根据其对盐浓度的适应性,说明该菌属于弱嗜盐微生物。吴嘉鑫等[17]对1株海洋耐热真菌C051进行研究,发现菌株C051在pH为3~9范围内均可以生长,而耐热真菌HS1-1在pH为3~12的范围内均能生长,相比之下湖光岩来源菌株HS1-1能适应较广的酸碱环境,有较强抗逆性,并且对碱性环境的抗性更强,这可能是由于湖光岩特殊的环境所造就的,生物的特性更具有多样化。有研究人员从不同碱性生境分离出多株真菌,并从其次生代谢产物中发现多种新颖结构化合物[24],HS1-1在pH为12时仍能生长,说明耐热真菌HS1-1可以适应碱性生境,研究其次级代谢产物将有希望发现新颖结构的化合物。
产酶研究发现该菌株周围无产生水解圈和特定颜色变化,这可能与该菌株酶的分泌能力和生理特性有关。郭潇等[25]在研究湖光岩细菌的纤维素酶降解活性也出现了这一现象。猜测该方法测定下菌株产生水解酶的量不多,在菌株生长的同时培养基的透明圈现象已被菌体生长所覆盖。后续如做进一步研究,可以考虑浓缩粗酶液于特定产酶培养基中,具体参照Chirom等[26]的方法稍加修改来更加明确该菌的产酶特性。
抗菌活性是微生物研究的热点,近年来研究人员致力于为抗菌物质开发提供新手段和新渠道,倪孟祥等[27]从北极海泥中分离得到的其中1株菌株H5被鉴定为烟曲霉属且其发酵液对细菌、真菌和耐药菌有不同抑制活性。本研究也探究了对几种常见革兰氏阳性和阴性菌的拮抗性,耐热真菌具有生长快的特点,下一步拟对菌株HS1-1的其他抗菌活性及生物活性进行研究,为该菌的开发利用提供基础。
Physiological properties and antibacterial activity of a thermotolerant fungus HS1-1
-
摘要: 湖光岩玛珥湖(Huguangyan Maar Lake)地理环境特殊,对于挖掘特殊生境耐热真菌资源具有较大潜力。对1株来源于湖光岩玛珥湖的耐热真菌HS1-1进行了形态观察,分子鉴定和生理特性、产酶能力、抗菌活性的测定。结果表明,该真菌为烟曲霉(Aspergillus fumigatus),菌株在15~50℃和pH3~12均能生长,最适生长条件是温度40℃,盐浓度1%~2%,pH6~7;经4种产酶培养基筛选确定菌株对可溶性淀粉、乳糖、羧甲基纤维素有较强的利用能力,对5种指示菌有一定抑制效果。说明真菌HS1-1具有较强耐热特性、酸碱耐受性和一定的抗菌活性。本研究对深入了解湖光岩玛珥湖耐热真菌并对其进一步开发利用提供理论基础。Abstract: Huguangyan Maar Lake is endowed with a special geographical environment and has a great potential for exploring thermotolerant fungal resources in special habitats. A thermotolerant fungus HS1-1 was collected from Huguangyan Maar Lake, observed morphologically and identified molecularly, and its physiological properties, enzyme production capacity and antibacterial activity were determined. The results showed that the strain of this fungus is identified as Aspergillus fumigatus. This fungus can grow at the temperatures of 15-50℃ and pH3-pH12, and has an optimum growth at the temperature of 40℃, a salt concentration of 1%-2%, and pH6-pH7. The fungus has a strong ability to utilize soluble starch, lactose and carboxymethyl cellulose, and has a certain inhibitory effect on five indicator bacteria, as determined by screening on four enzyme-producing media. This indicates that the fungus HS1-1 is high in thermotolerance and acid and base tolerance with some antibacterial activities. This study provides a theoretical basis for an in-depth understanding of the thermotolerant fungi of Huguangyan Marr Lake and their further development and utilization.
-
Key words:
- thermotolerant fungi /
- molecular identification /
- physiological growth /
- activity research /
- Huguangyan
-
表 1 产酶培养基
产酶培养基 配 方 淀粉酶培养基 可溶性淀粉20.0 g·L−1,KCl 0.5 g·L−1,NaNO3 2.0 g·L−1,K3PO4 1.0 g·L−1,MgSO4·7H2O 0.5 g·L−1,琼脂18.0 g·L−1,海水晶30.0 g·L−1,ddH2O 1000 mL 糖苷酶培养基 乳糖 10.0 g·L−1,K2HPO4 1.0 g·L−1,FeSO4 0.01 g·L−1,NaNO3 3.0 g·L−1,KCl 0.5 g·L−1,MgSO4·7H2O 0.5 g·L−1,琼脂15.0 g·L−1,dd H2O 1000 mL 纤维素酶培养基 羧甲基纤维素8.0 g·L−1,KCl 0.5 g·L−1,NaNO3 3.0 g·L−1,FeSO4 0.01 g·L−1,K2HPO4 1.0 g·L−1,MgSO4·7H2O 0.5 g·L−1,氯霉素0.05 g·L−1,琼脂20.0 g·L−1,ddH2O 1000 mL,pH6.0 壳聚糖酶培养基 壳聚糖10.0 g·L−1,CaCO3 6.0 g·L−1,MgSO4·7H2O 0.25 g·L−1,(NH4)2SO4 2.5 g·L−1,K2HPO4 0.02 g·L−1,NaCl 4.0 g·L−1,琼脂20.0 g·L−1,pH6.0  下载: 导出CSV
下载: 导出CSV
表 2 不同盐浓度对菌株HS1-1生长的影响
盐浓度/% 菌落直径/ mm 1 d 2 d 3 d 4 d 5 d 6 d 7 d 0 11.60±0.21a 37.75±0.20d 63.50±0.30c 84.56±0.31a 1 10.89±0.45ab 44.91±0.53a 70.36±0.37a 84.92±0.19a 2 10.53±0.73b 43.74±0.03b 69.69±0.76a 84.58±0.40a 3 10.95±0.14ab 41.09±0.30c 65.78±0.19b 84.33±0.11a 4 10.40±0.49b 37.83±0.25d 60.40±0.95d 84.79±0.15a 5 9.05±0.07c 34.31±0.25e 55.98±0.52e 79.38±0.83b 84.30±0.28a 6 7.43±0.04d 27.54±0.46f 49.24±0.19f 70.60±0.52c 81.74±0.58b 83.16±0.19a 8 4.80±0.21e 22.37±0.29g 39.63±0.36g 61.37±0.26d 74.93±0.90c 83.66±0.29a 10 2.07±0.30f 13.93±0.45h 26.18±0.07h 40.31±0.06e 52.31±0.66d 63.38±0.38b 76.20±0.52 注:标记不同字母表示具有显著性差异(P<0.05);空白表示该天数菌株长满平板(下同)。
下载: 导出CSV
表 3 不同pH条件对菌株HS1-1生长的影响
pH 菌落直径/ mm 1 d 2 d 3 d 4 d 5 d 6 d 7 d 2 0.00±0.00f 0.00±0.00h 0.00±0.00g 0.00±0.00j 0.00±0.00h 0.00±0.00f 0.00±0.00d 3 1.95±0.27e 7.89±0.60f 14.28±0.72f 19.95±0.19i 25.49±0.67g 30.22±0.51e 35.57±0.46d 4 9.67±0.22ab 28.38±0.28c 48.43±0.49c 63.49±0.21d 79.83±0.15b 85.26±0.88a 5 9.27±0.79bc 24.68±0.89d 36.87±0.48d 45.82±0.97f 59.47±0.72d 66.27±0.89c 76.93±0.65b 6 10.45±0.15ab 39.57±0.06a 61.31±0.30a 82.45±0.31a 84.64±0.11a 7 10.84±0.36a 39.62±0.70a 60.96±0.78a 81.11±0.44a 84.69±0.19a 8 8.02±0.95c 30.81±0.58b 52.84±0.78b 76.25±0.79b 84.25±0.10a 9 5.97±0.34d 27.78±0.59c 49.75±0.60c 73.82±0.59c 84.23±0.13a 10 1.92±0.28e 9.69±0.88e 28.00±0.46e 50.98±0.46e 71.29±0.59c 83.50±0.44a 11 1.26±0.15e 2.29±0.35g 15.25±0.72f 36.34±0.5g 56.58±0.28e 76.42±0.83b 83.44±0.24a 12 0.95±0.14ef 1.75±0.19g 6.57±0.31g 25.30±0.15h 41.80±0.23f 46.75±0.67d 53.12±0.54c
下载: 导出CSV
表 4 不同培养基发酵液与发酵浓缩液抗菌活性比较
指示菌 在PDB上的抑菌圈直径/mm 在改良马丁培养基上的抑菌圈直径/mm 发酵液 发酵浓缩液 发酵液 发酵浓缩液 SL-1 7.80±0.00a 15.05±0.99a 14.26±0.63c 12.40±0.40d 大肠杆菌 7.80±0.00a 13.21±0.56a 20.83±0.59a 21.29±0.08b 铜绿假单胞菌 7.80±0.00a 14.15±0.51a 13.51±0.24c 18.23±0.73c SL-2 7.80±0.00a 12.97±0.19a 15.40±0.29b 24.82±0.18a 金黄色葡萄球菌 7.80±0.00a 13.16±0.35a 7.80±0.00d 11.00±0.58e
下载: 导出CSV
-
[1] 张勇, 李多川. 耐热真菌五个中国新记录种[J]. 菌物学报, 2013, 32(1): 142 − 149. [2] 张勇. 耐热真菌多样性及分子系统学研究[D]. 泰安: 山东农业大学, 2012. [3] 梁茵, 李多川. 耐热真菌的分类及系统学研究[C]// 2010年中国菌物学会学术年会论文摘要集. 湖南: 中国菌物学会, 2010, 228-229. [4] SALAR R K, ANEJA K R. Thermophilous fungi from temperate soils of northern India [J]. Journal of Agriculture Technology, 2006, 2(1): 49 − 58. [5] MICHELE M, ROBERTO R, RICHARD J W, et al. Purification and biochemical characterization of a thermostable extracellular glucoamylase produced by the thermotolerant fungus Paecilomyces variotii [J]. Journal of Industrial Microbiology & Biotechnology, 2008, 35(1): 17 − 25. [6] 詹发强, 崔卫东, 周亚飞, 等. 降解棉秸秆耐热菌株的鉴定及降解条件优化[J]. 微生物学通报, 2012, 39(1): 44 − 54. doi: 10.13344/j.microbiol.china.2012.01.004 [7] 秦青英, 曾永辉, 郭倩茹, 等. 环境16S rDNA和16S rcDNA序列分析湖光岩玛珥湖的浮游细菌组成[J]. 广东海洋大学学报, 2013, 33(3): 1 − 9. doi: 10.3969/j.issn.1673-9159.2013.03.001 [8] 刘颖, 陈倩娜. 湛江湖光岩土壤放线菌分离与抗菌活性测定[J]. 热带农业工程, 2009, 33(1): 1 − 4. [9] COONEY, D G, EMERSON R. Thermophilic fungi: an account of their biology, activities, and classification [J]. The Quarterly Review of Biology, 1965, 40(3): 292. [10] MOUCHACCA J. Thermophilic fungi-biodiversity and taxonomic status [J]. Cryptogamie Mycologie, 1997, 18(1): 19 − 69. [11] 周茂洪, 赵肖为, 周林, 等. 产淀粉酶的海洋曲霉菌的分离及酶学特性初步研究[J]. 海洋学研究, 2007(3): 59 − 65. doi: 10.3969/j.issn.1001-909X.2007.03.007 [12] 李德荣, 郑海艳, 谢庆武. β-半乳糖苷酶产生真菌的筛选及酶学性质研究[J]. 广西轻工业, 2010, 26(5): 3 − 4. doi: 10.3969/j.issn.1003-2673.2010.05.002 [13] 殷亚兰. 新疆吐鲁番地区耐热真菌多样性及其产酶活性初步研究[D]. 乌鲁木齐: 新疆大学, 2016. [14] 朱利平, 黄惠莉. 产壳聚糖酶的海洋青霉菌筛选及其寡营养发酵的探究[J]. 食品工业科技, 2011, 32(5): 212 − 214. doi: 10.13386/j.issn1002-0306.2011.05.047 [15] 魏景超. 真菌鉴定手册[M]. 上海: 上海科学技术出版社, 1979: 487-534. [16] 曲均革, 金甚益, 绕静. 杭州湾海域产纤维素酶海洋真菌的分离筛选及鉴定[J]. 安徽农业科学, 2012, 40(16): 8816 − 8817. doi: 10.3969/j.issn.0517-6611.2012.16.008 [17] 吴嘉鑫, 陈虹, 赵程琳, 等. 近江牡蛎共附生耐热真菌C051的鉴定、生理学特性及抗菌活性研究[J]. 微生物学杂志, 2020, 4(40): 34 − 39. [18] 刘冬梅, 李理, 杨晓泉, 等. 用牛津杯法测定益生菌的抑菌活力[J]. 食品研究与开发, 2006(3): 110 − 111. doi: 10.3969/j.issn.1005-6521.2006.03.044 [19] 王世苗, 陈天意, 冯亚萍, 等. 伊犁野生郁金香内生烟曲霉的分离鉴定及活性[J/OL]. 微生物学通报: 1 − 17[2022−08−11]. DOI: 10.13344/j.microbiol.china.220432. [20] SUGUI J A, KWON-CHUNG K J, JUVVADI P R, et al. Aspergillus fumigatus and related species [J]. Cold Spring Harbor Perspectives in Medicine, Cold Spring Harbor Laboratory Press, 2015, 5(2): a019786. doi: 10.1101/cshperspect.a019786 [21] 王东彧. 功能蛋白质组方法研究Aspergillus fumigatus生长偏好性与环境适应性[D]. 济南: 山东大学, 2016. [22] BEFFA T, STAIB F, LOTT F J, et al. Mycological control and surveillance of biological waste and compost [J]. Med Mycol, 1998, 36(Suppl 1): 137-145. [23] 张晋卿, 刘伟, 谭静文, 等. 烟曲霉在不同温度和营养条件下的生长特性初探[J]. 中国真菌学杂志, 2013, 8(1): 6 − 9. doi: 10.3969/j.issn.1673-3827.2013.01.002 [24] 淳于纬训. 四株碱性生境真菌次生代谢产物及其抑菌活性的研究[D]. 昆明: 云南大学, 2017. [25] 郭潇, 雷晓凌. 湖光岩水域细菌的纤维素降解活性及抑菌活性的研究[J]. 现代食品科技, 2009, 25(10): 1166 − 1169. [26] CHIROM AARTI, AMEER KHUSRO, PAUL AGASTIAN, et al. Molecular diversity and hydrolytic enzymes production abilities of soil bacteria [J]. Saudi Journal of Biological Sciences, 2020, 27: 3235 − 3248. doi: 10.1016/j.sjbs.2020.09.049 [27] 倪孟祥, 胡颖. 北极海泥来源抗菌活性真菌的筛选及菌株H5的初步研究[J]. 化学与生物工程, 2013, 30(5): 61 − 64. doi: 10.3969/j.issn.1672-5425.2013.05.017 -

 点击查看大图
点击查看大图
图(6) / 表 (4)
计量
- 文章访问数: 2365
- HTML全文浏览量: 742
- PDF下载量: 16
- 被引次数: 0
