

 下载:
下载:
-
文心兰(Oncidium)是兰科(Orchidaceae)重要观赏植物,因花形独特、花色丰富、花期较长而广受青睐,是国际花卉市场主要的切花与盆花种类之一[1-3]。中国自20世纪90年代引种文心兰,以切花为主,在海南、广东、福建等地形成规模化种植。其适宜生长温度为15~30℃,最适营养生长温度为15~20℃,最适开花温度为20~25℃[4]。随着全球气候变暖,夏季极端高温频发,海南等主产区常出现持续35℃以上的高温天气,严重制约文心兰的生长与开花品质[5]。高温胁迫主要通过抑制光合作用、加剧膜脂过氧化、扰乱抗氧化酶系统等途径影响植物[6]。光合作用是对高温最敏感的生理过程之一,高温可导致叶绿素降解、光系统II活性下降及气孔关闭,从而抑制碳同化,造成植株整体光合性能下降[7-8]。同时,高温胁迫破坏细胞内活性氧(ROS)代谢平衡,过量ROS引发膜脂过氧化、蛋白质变性与DNA损伤等氧化胁迫[9]。为清除ROS,植物进化出超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)等抗氧化酶系统[10]。
目前,兰科植物耐热性研究主要集中于蝴蝶兰(Phalaenopsis)、石斛兰(Dendhobium)等种类,发现高温可致其叶绿素含量下降、膜透性增加、抗氧化酶活性改变[11-13]。在文心兰中,高温胁迫会抑制花芽分化与开花,降低叶绿素含量[14]。但现有研究多关注单一温度或单一时间点的生理响应,对胁迫进程中光合系统与抗氧化系统的动态协同演变,尤其是SOD、POD、CAT在时序上的差异化响应特征,尚缺乏系统报道,限制了对文心兰耐热生理基础的深入理解。‘小蜜蜂’是华南热区栽培的文心兰商业切花品种之一,具有花色亮丽、花期长、瓶插期较长等优良性状。文心兰环境温度高于35℃时生长受抑制,而海南夏季棚内温度常超过35℃甚至接近42℃。因此,本研究以文心兰品种‘小蜜蜂’为材料,在人工气候室中设置35℃(温和高温)和42℃(极端高温)2种胁迫条件,分别处理0 h(基线)、24 h和48 h,系统测定叶片表型、光合参数(SPAD值、Pn、Gs、Tr、WUE)、抗氧化酶(SOD、POD、CAT)活性及丙二醛(MDA)含量。旨在揭示不同强度高温胁迫下文心兰光合系统与抗氧化防御系统的协同响应及时序动态特征,阐明其对短时极端高温的生理适应机制,为耐热性评价、栽培管理优化及耐热品种选育提供理论依据。
-
供试的植物材料为文心兰品种‘小蜜蜂’(Oncidium ‘Honey Bee’),由海南博大兰花科技有限公司东山基地种质资源圃(N 19°47′29.7″,E 110°14′33.1″ )提供。选取生长状态一致、无病虫害、长势健康的2年生成熟植株共50株作为实验材料,所有植株均处于营养生长期(假鳞茎已成熟、未抽花梗),植株高度、叶片数及假鳞茎大小基本一致。所有植株均采用相同基质(V树皮:V碎石=3:1,pH5.5~6.5,EC为0.3~0.5 mS·cm−1)栽培于直径15 cm的塑料营养钵中。栽培期间每7 d浇灌1次,每14 d喷施1次稀释2 000倍的花宝2号(Hyponex No.2,N-P-K = 20-20-20)平衡复合肥水溶液,其余时间保持基质适度湿润。试验开始前24 h对所有植株统一浇透水,使基质达到田间持水量,以保证试验起始时各处理组的水分状态一致。
主要试剂:SOD活性测定试剂盒(苏州梦犀生物医药科技有限公司),POD活性测定试剂盒(苏州梦犀生物医药科技有限公司),CAT活性测定试剂盒(苏州梦犀生物医药科技有限公司),MDA含量测定试剂盒(苏州梦犀生物医药科技有限公司)。
-
采用人工气候培养箱(RXZ-500B,宁波江南仪器厂)模拟高温胁迫环境。设置2个温度处理:35℃(温和高温)和42℃(极端高温),所有植株在正式处理前于28℃条件下预培养3 d以适应培养箱环境 ,预培养结束时(0 h)取样作为胁迫前基线,随后将植株分别转入35℃和42℃培养箱进行胁迫处理,光照强度统一设置为12 000 lx,光周期为14 h光照/10 h黑暗,相对湿度控制在75%±5%。每个温度处理使用独立的人工气候培养箱。将长势一致的文心兰植株随机分为3组,分别置于35℃和42℃的培养箱中进行处理。处理期间保持土壤水分充足,避免干旱胁迫的交互影响,试验开始前24 h对所有植株统一浇透水,试验期间(0~48 h)保持基质表层湿润,每24 h检查基质水分状况,如基质表面干燥则用喷壶补充等量去离子水(35℃与42℃处理组同步进行,以保持两组水分条件一致)。处理时间设置为0 h(对照,即预培养结束未开始高温处理的时间点)、24 h和48 h。每个温度处理每个时间点设置3个生物学重复,每个重复包含5株植株。
-
分别于高温胁迫处理的0、24和48 h 3个时间点,对35℃和42℃处理组的文心兰植株进行叶片样品采集。每个时间点每个处理随机选取5株长势一致的植株,每株选取植株中部完全展开的功能叶(自顶端向下第3~4片叶)用于各项生理指标的测定。用于生理指标测定的新鲜叶片采集后立即装入自封袋中,置于冰盒内暂存,迅速带回实验室进行各项生理指标的测定。所有生理指标测定均在样品采集后24 h内完成,以保证酶活性和代谢物的原始状态。每个处理每个时间点设置3次重复。
-
于各取样时间点,对处理组和对照组的文心兰植株进行拍照记录,观察叶片颜色变化、萎蔫程度和整体生长状态。使用数码相机(EOS 80D,Canon,Japan)在相同光照条件和拍摄参数下进行拍摄,拍摄距离为50 cm,角度为俯视45°。
-
采用便携式叶绿素计(SPAD-502,Konica Minolta,日本)对各处理组文心兰叶片的叶绿素相对含量进行活体测定。每株选取与表型观察相同部位的完全展开功能叶(自顶端向下第3~4片叶),每片叶测定时避开主叶脉,在叶片上、中、下3个部位分别读取SPAD值,取平均值作为该叶片的叶绿素相对含量。每株测定3片叶,每个处理共测定15片叶,结果以平均值±标准差表示。所有测定均在上午9:00—11:00完成,以保证光照条件一致。
-
采用便携式光合测定系统(LI-
6800 ,LI-COR Biosciences,USA)对各处理组文心兰叶片的气体交换参数进行活体测定。测定时光合有效辐射(PAR)设置为1 000 μmol·m−2·s−1(饱和光强),参比室CO2浓度设定为400 μmol·L−1,叶室温度为28℃(与对照温度一致),相对湿度控制在60%~70%。于各取样时间点的上午9:00—11:00,每个处理随机选取5株长势一致的植株,每株选取与SPAD测定相同部位的完全展开功能叶进行测定。待仪器显示读数稳定后,记录以下参数:净光合速率(Pn,μmol ·m−2·s−1)、气孔导度(Gs,mol ·m−2·s−1)、蒸腾速率(Tr,mmol ·m−2·s−1)和胞间CO2浓度(Ci,μmol ·mol−1)。每株测定1片叶,5次生物学重,每个处理共测定5片叶。水分利用效率(WUE)(WUE=Pn / Tr,μmol CO2·mmol−1 H2O)。所有光合参数的测定均在稳定的环境条件下进行,测定前对仪器进行充分预热和校准,每次更换CO2和H2O吸收剂后均进行系统自检,确保数据准确性。 -
采用苏州梦犀生物医药科技有限公司生产的相应试剂盒,对文心兰叶片中超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性及丙二醛(MDA)含量进行测定。具体测定方法严格按照各试剂盒说明书操作,简述如下。
1)酶液提取:称取新鲜文心兰叶片0.1 g,剪碎后置于预冷的研钵中,加入1.0 mL预冷的相应提取缓冲液(SOD、POD、CAT、MDA分别使用各自试剂盒提供的提取液),冰浴条件下充分研磨至匀浆。将匀浆液转移至2.0 mL离心管中,于4℃、
8000 ×g离心10 min,取上清液即为粗酶液,置于冰上备用。2)超氧化物歧化酶(SOD)活性测定:采用氮蓝四唑(NBT)光化还原法测定SOD活性。反应体系包含SOD测定缓冲液2.0 mL、NBT溶液0.2 mL、核黄素溶液0.1 mL、酶液0.1 mL,用蒸馏水补足至3.0 mL。同时设置对照管(不加酶液,以等体积提取液代替)。混匀后置于4 000 lx光下反应20 min,反应结束后立即避光终止反应,于560 nm波长处测定吸光度值。SOD活性以每克鲜质量样品抑制NBT光化还原50%所需的酶量为1个酶活性单位(U·g−1)表示。计算公式如下:
$$ \mathrm{SOD}=\frac{{A}_{C}-{A}_{S}}{{A}_{C}\times 0.5}\times \frac{{V}_{T}}{{V}_{S}\timesW}\text{,} $$ (1) 式中,SOD为超氧化物歧化酶活性(U·g−1 );$ {A}_{C} $为对照管吸光度;$ {A}_{S} $为测定管吸光度;$ {V}_{T} $为反应体系总体积(mL);$ {V}_{S} $为加入酶液体积(mL);$ W $为样品鲜质量(g)。
3)过氧化物酶(POD)活性测定:采用愈创木酚法测定POD活性。反应体系包含POD测定缓冲液2.0 mL、0.5%愈创木酚溶液1.0 mL、体积分数0.5% 的H2O2溶液0.5 mL、酶液0.1 mL,用蒸馏水定容至4.0 mL。加入酶液后立即混匀并计时,于470 nm波长处测定1 min内吸光度值的变化。以每克鲜质量样品每分钟吸光度变化0.01为1个酶活性单位(U·g−1·min−1)表示POD活性。计算公式如下:
$$ \mathrm{POD}=\dfrac{\mathit{\Delta }{A}_{470}}{0.01}\times \frac{{V}_{T}}{{V}_{S}\times W\times t}\text{,} $$ (2) 式中,POD 为过氧化物酶活性(U·g-1·min-1);$ \text{Δ}{A}_{470} $为470 nm处1 min内吸光度变化值;$ {V}_{T} $为反应体系总体积(mL);$ {V}_{S} $为加入酶液体积(mL);$ W $为样品鲜质量(g);$ t $为反应时间(min)。
4)过氧化氢酶(CAT)活性测定:采用紫外吸收法测定CAT活性。反应体系包含CAT测定缓冲液2.0 mL、0.1 mol·L−1 H2O2溶液1.0 mL、酶液0.1 mL,用蒸馏水定容至3.5 mL。加入酶液后迅速混匀,于240 nm波长处测定1 min内吸光度值的变化。以每克鲜质量样品每分钟吸光度变化0.01为1个酶活性单位(U·g−1·min−1 FW)表示CAT活性。计算公式如下:
$$ \mathrm{CAT}=\dfrac{\mathit{\Delta }{A}_{240}}{0.01}\times \frac{{V}_{T}}{{V}_{S}\times W\times t}\text{,} $$ (3) 式中,CAT为过氧化氢酶活性(U·g−1·min−1 FW);$ \text{Δ}{A}_{240} $为240 nm处1 min内吸光度变化值;$ {V}_{T} $为反应体系总体积(mL);$ {V}_{S} $为加入酶液体积(mL);$ W $为样品鲜质量(g);$ t $为反应时间(min)。
5)丙二醛(MDA)含量测定:采用硫代巴比妥酸(TBA)比色法测定MDA含量。取酶液1.0 mL,加入2.0 mL 0.6% TBA溶液(溶于10%三氯乙酸),混匀后于沸水浴中反应30 min,迅速冷却后于4℃、10 000×g离心10 min。取上清液于532 nm和600 nm波长处测定吸光度值。考虑到0 h时各处理组MDA测定值存在一定基线差异(可能源于TBA显色法的批次效应),而其他抗氧化指标(SOD、CAT、POD)在0 h时各组间均无显著差异(P>0.05),提示该差异源于测定环节而非材料本身。为更准确反映高温胁迫诱导的膜脂过氧化动态,本研究中MDA 数据以增量形式(ΔMDA)进行分析,其余指标以绝对值呈现。计算公式如下:
$$ \mathrm{MDA}=\dfrac{({A}_{532}-{A}_{600})×{V}_{T}×1{0}^{6}}{\varepsilon \times W\times{V}_{S}}\text{,} $$ (4) 式中,$ {V}_{T} $为提取液总体积(mL),$ {V}_{S} $为测定时所取提取液体积(mL),$ W $为样品鲜重(g),$ \varepsilon $为MDA的摩尔消光系数(155 L·mol−1·cm−1)。
以上所有生理指标测定均设置3次生物学重复,每个样品进行3次技术重复取平均值。
-
所有数据采用Microsoft Excel 2021进行整理,使用GraphPad Prism 9.5软件进行统计分析和图表绘制。采用双因素方差分析(Two-way ANOVA)检验温度(35℃、42℃)和处理时间(0 h、24 h、48 h)及其交互作用对各生理指标的显著性影响。采用Tukey‘s HSD多重比较进行事后检验,以P<0.05作为差异显著性阈值。结果以平均值±标准差(Mean ± SD)表示。所有图表均使用GraphPad Prism 9.5软件绘制完成。
-
为明确高温胁迫对文心兰造成的可见损伤,对处理前后的植株进行了表型观察。从图1可知,35℃温和高温处理24 h后,文心兰植株未表现出明显的表型异常,叶片仍保持舒展和翠绿,与0 h对照组基本一致;处理48 h后,部分植株叶片出现轻微的失绿和黄化迹象,但整体形态仍较为正常。在42℃极端高温处理下,文心兰在24 h后即表现出明显的胁迫症状,叶片出现黄化并伴随轻微萎蔫;处理48 h后,黄化程度急剧加重,大部分叶片变为黄绿色或鲜黄色,叶片严重萎蔫、失水下垂,呈现典型的热害症状。上述表型变化表明,42℃极端高温在48 h内即可对文心兰造成严重的生理损伤,而35℃温和高温处理的影响相对较轻。

图 1 高温胁迫下文心兰表型变化
Figure 1. Phenotypic changes of Oncidium under heat stress
-
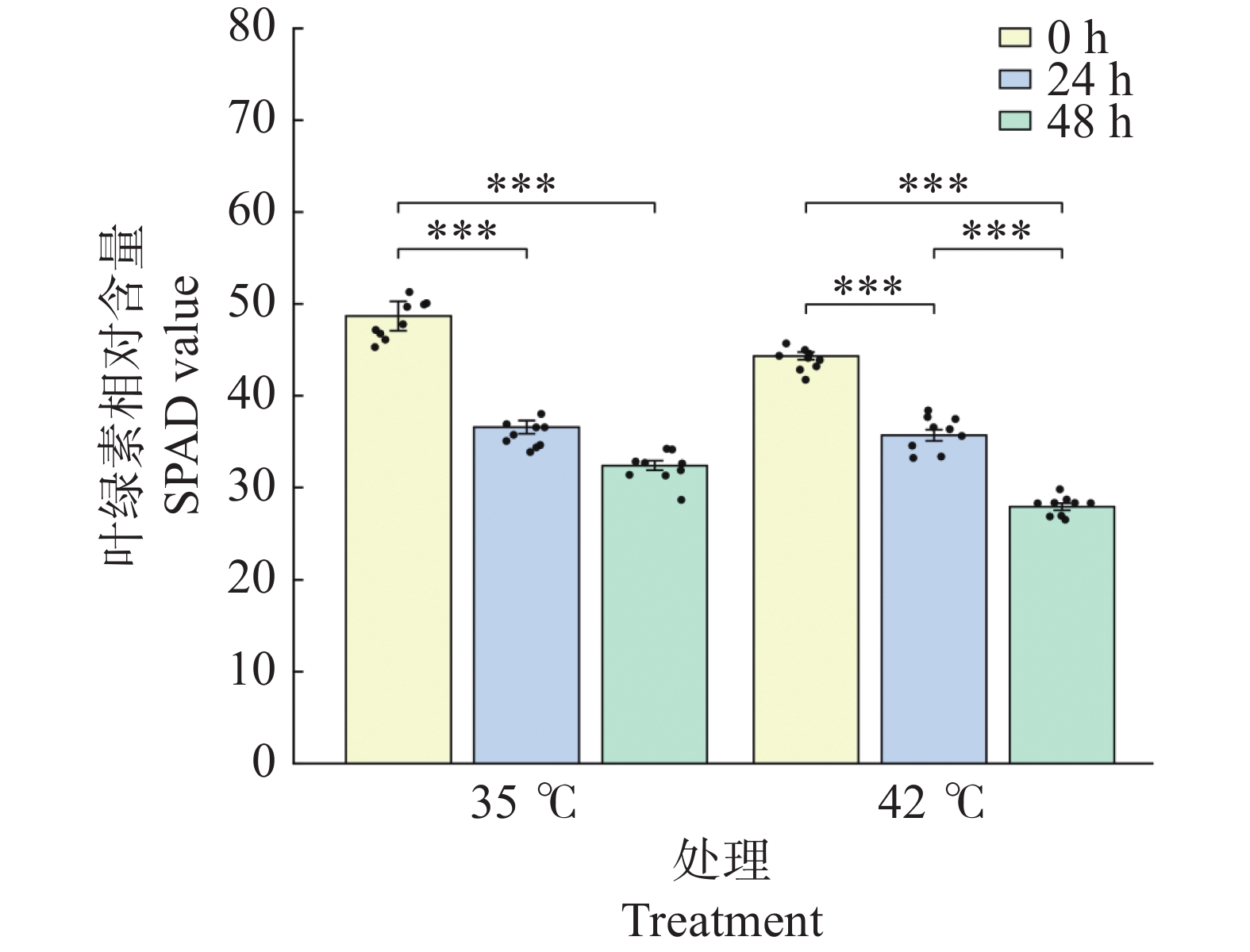
文心兰叶片SPAD值(反映叶绿素相对含量)在2种温度处理下均呈显著下降趋势(图2)。35℃处理下,SPAD值由0 h的48.69显著下降至24 h的36.59(降幅24.9%,P<0.001),48 h进一步降至32.42(较0 h下降33.4%,P<0.001),48 h与24 h间差异不显著(P>0.05),提示35℃处理下叶绿素降解主要发生在前24 h;42℃处理下,SPAD值由0 h的44.34持续显著下降至24 h的35.69(降幅19.5%,P<0.001)和48 h的27.93(较0 h下降37.0%,P<0.001),且48 h显著低于24 h(P<0.001),表明42℃高温对叶绿素的降解作用持续存在。48 h时42℃处理组SPAD值显著低于同期35℃处理组(P<0.001),提示极端高温对光合色素系统的损害程度更深。
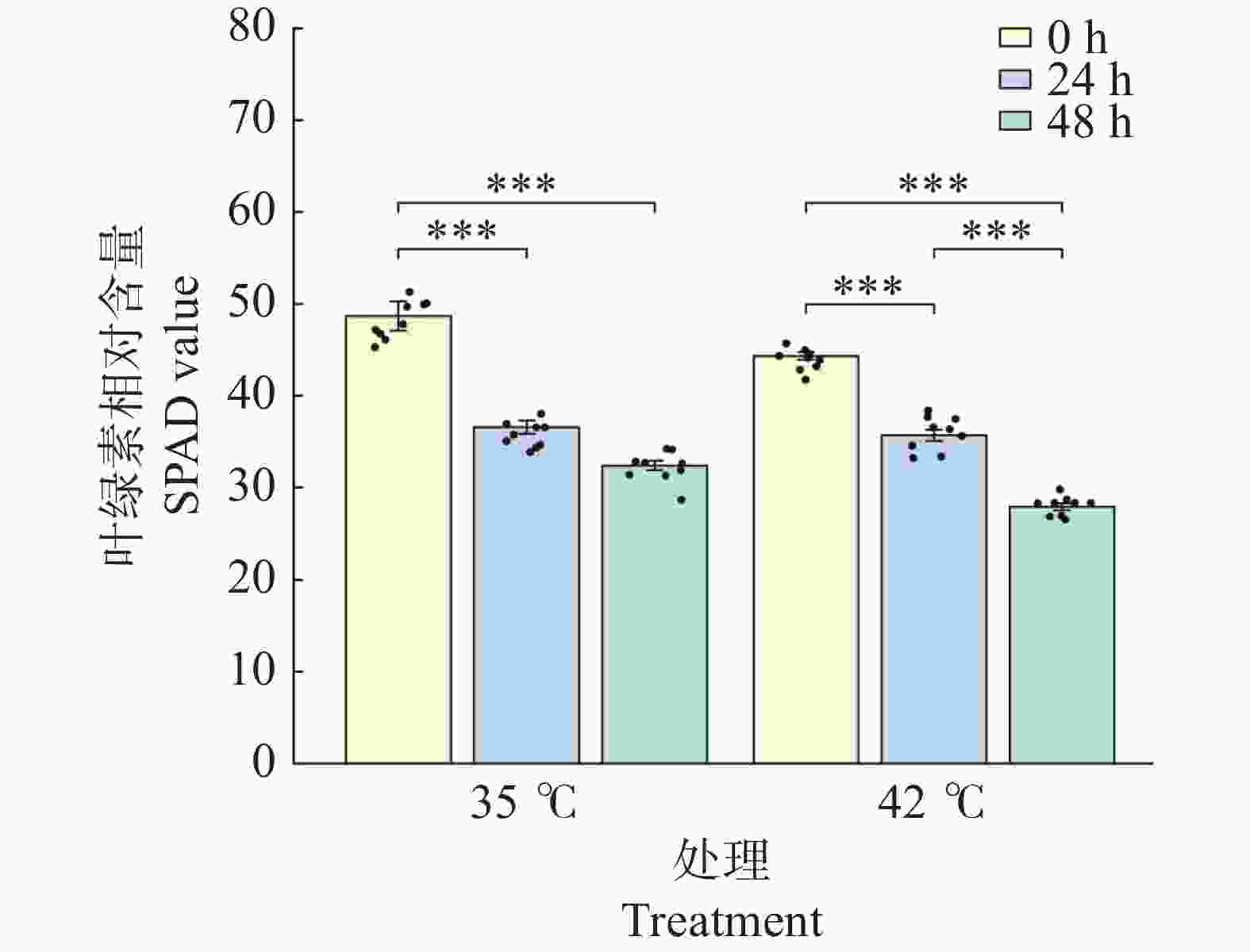
图 2 高温胁迫对文心兰叶绿素含量的影响
Figure 2. Effects of heat stress on chlorophyll content (SPAD value)in Oncidium leaves
-
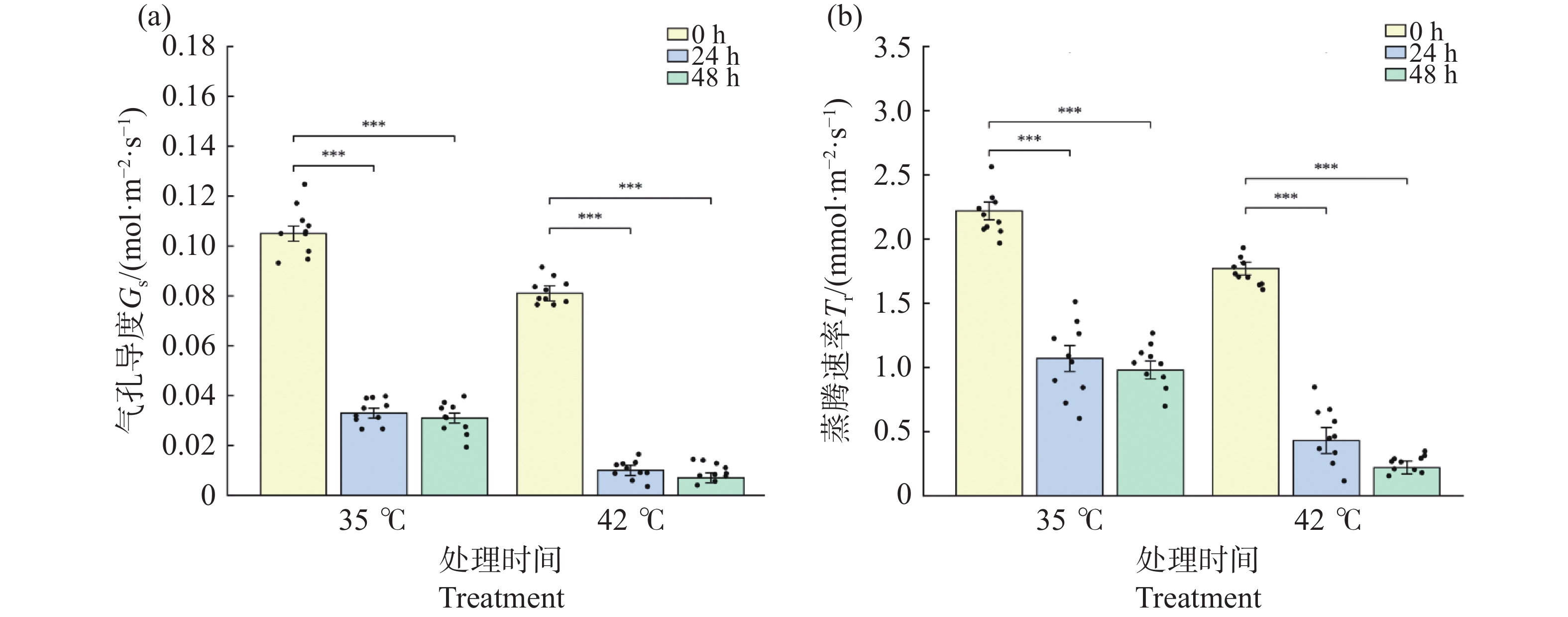
气孔导度(Gs)和蒸腾速率(Tr)在2种温度处理下均急剧下降。35℃处理下,Gs从0 h的0.105 mol·m−2·s−1极显著下降至24 h的0.033(降幅68.4%,P<0.001),48 h维持在0.031(较0 h下降70.6%,P<0.001),24 h与48 h间无显著差异(P>0.05),表明35℃处理下气孔在24 h内迅速部分关闭并维持稳定(图3-a)。同期Tr表现出相似变化模式(图3-b),从0 h的2.22 mmol·m−2·s−1降至24 h的1.07(降幅 52.0%)和48 h的0.98(降幅 56.0%);42℃处理下气孔响应更为剧烈:Gs从0 h的0.081 mol·m−2·s−1极显著降至24 h的0.010(降幅87.9%,P<0.001),48 h进一步降至0.007(降幅 91.4%);Tr从0 h的1.77 mmol·m−2·s−1降至 24 h的0.43(降幅75.6%)和48 h的0.22(降幅87.4%)。24 h与48 h时42℃处理组的Gs和Tr均极显著低于同期35℃处理组(P<0.001)。结果表明,极端高温触发气孔近乎完全关闭,蒸腾几乎停止,体现了植物在极端胁迫下减少水分散失的应急保护机制。
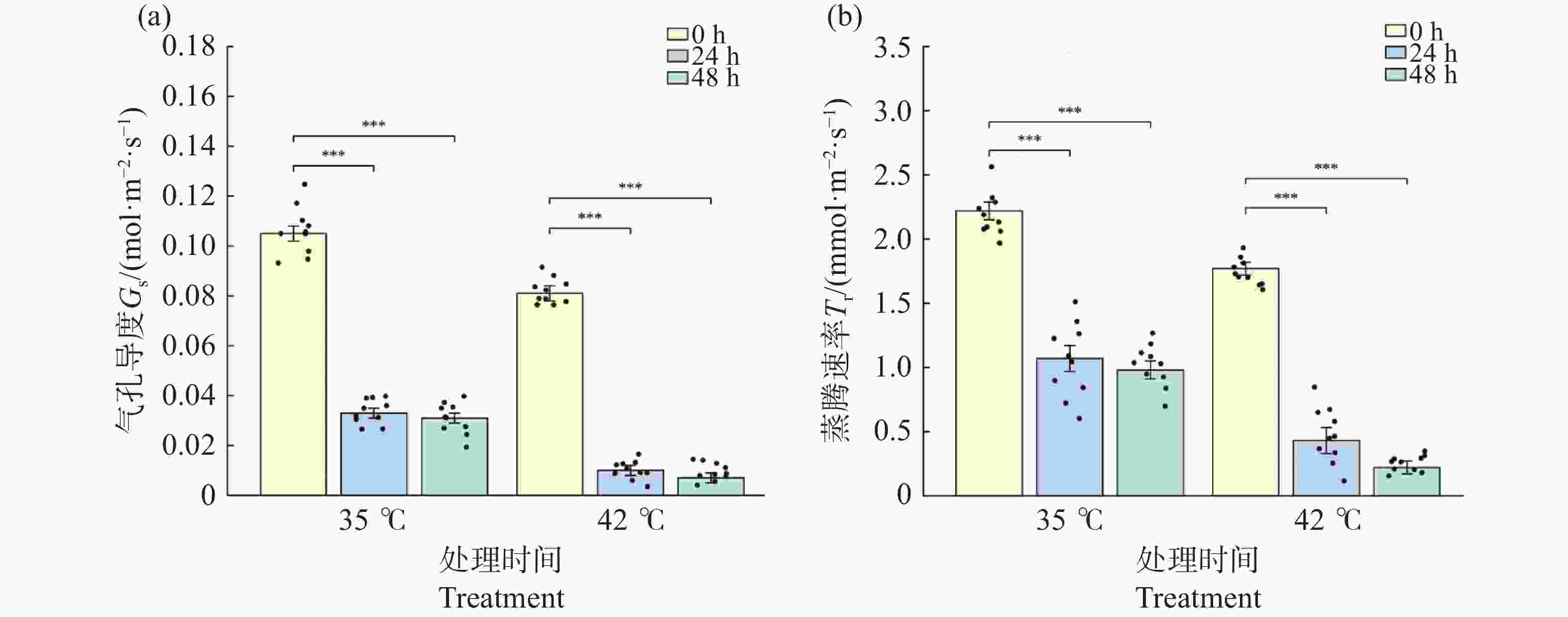
图 3 高温胁迫对文心兰气孔导度(GS)与蒸腾速率(Tr)的影响
Figure 3. Effects of heat stress on stomatal conductance (Gs)and transpiration rate (Tr)in Oncidium leaves
-
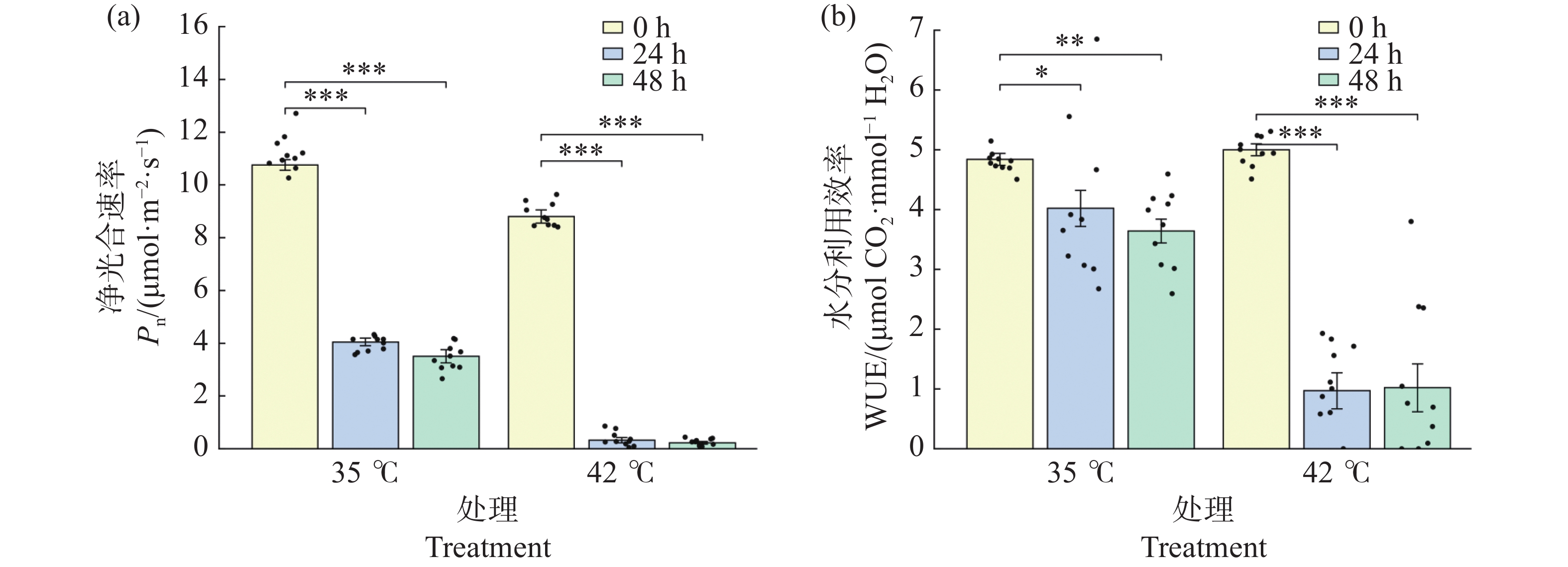
净光合速率(Pn)在高温胁迫下显著下降(图4-a)。35℃处理下,Pn 从0 h的10.76 μmol·m−2·s−1极显著下降至24 h的4.05 μmol·m−2·s−1(降幅62.3%,P<0.001),48 h进一步降至3.51 μmol·m−2·s−1(降幅67.3%,P<0.001),24 h与48 h之间差异不显著(P>0.05)。42℃处理下,Pn下降更为剧烈:24 h时已极显著降至0.32 μmol·m−2·s−1(降幅96.3%,P<0.001),48 h时进一步降至0.23 μmol·m−2·s−1(降幅97.4%),光合作用基本停止。24 h与48 h时,42℃处理组的Pn均极显著低于同期35℃处理组(P<0.001),表明42℃持续高温对光合作用的抑制作用远强于35℃。
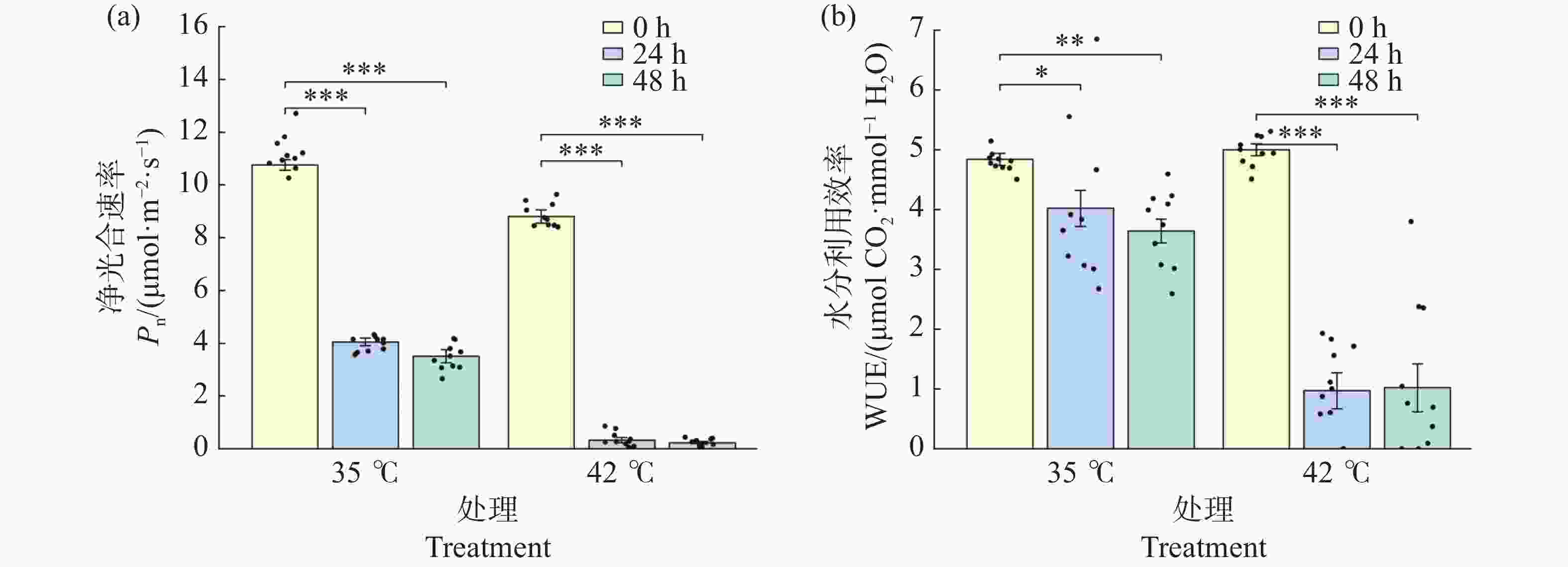
图 4 高温胁迫对文心兰净光合速率(Pn)与水分利用效率(WUE)的变化
Figure 4. Effects of heat stress on net photosynthetic rate (Pn)and water use efficiency (WUE)in Oncidium leaves
水分利用效率(WUE)反映了光合碳同化与水分消耗之间的平衡(图4-b)。0 h时,两组WUE接近(35℃:4.84,42℃:5.00 mmol·mol-1,P > 0.05)。35℃处理下,WUE从0 h的4.84缓慢下降至24 h的4.02(P<0.05)和48 h的3.64(P<0.01),表明在35℃下光合-蒸腾仍维持一定平衡。42℃处理下,WUE急剧下降至24 h的0.97(降幅80.6%,P < 0.001)和48 h的1.02(降幅79.5%)。24 h与48 h时,42℃处理组的WUE均极显著低于同期35℃处理组(P<0.001),反映出极端高温下气孔关闭和光合系统受损的双重作用,导致碳同化效率与水分利用效率同步崩溃。
-
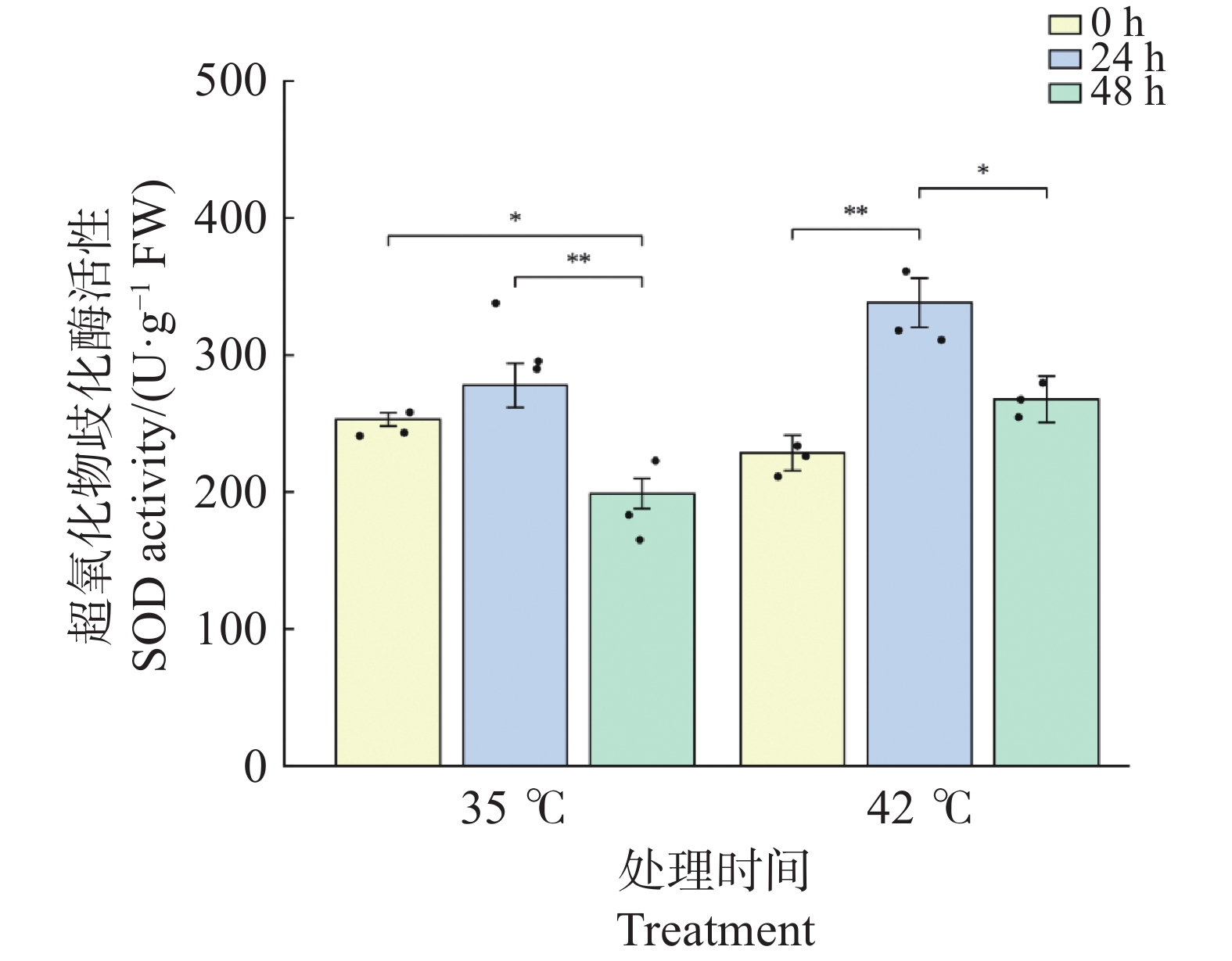
文心兰叶片SOD活性在2种温度处理下均呈先升后降的变化趋势(图5)。35℃处理下,SOD活性在24 h较0 h略有上升(由252.94升至277.65 U·g−1 ,升幅9.8%),但差异不显著(P>0.05);48 h时显著下降至198.72 U·g−1 (较0 h下降21.4%,P<0.05),且较24 h极显著下降(P<0.01),表明35℃处理下SOD活性在胁迫后期出现明显衰减。42℃处理下,SOD响应更为剧烈:24 h时活性极显著上升至338.02 U·g−1 (较0 h升高47.8%,P<0.01),增幅约为同期35℃处理的5倍;48 h时虽较24 h显著回落至267.58 U·g−1(P<0.05),但仍维持在略高于0 h的水平(+17.0%,P>0.05),且显著高于同期35℃处理组(P<0.05)。这表明42℃高温在胁迫早期更强烈地激活SOD系统,且其高活性维持时间较35℃更长,反映了文心兰对极端高温的应激响应能力。
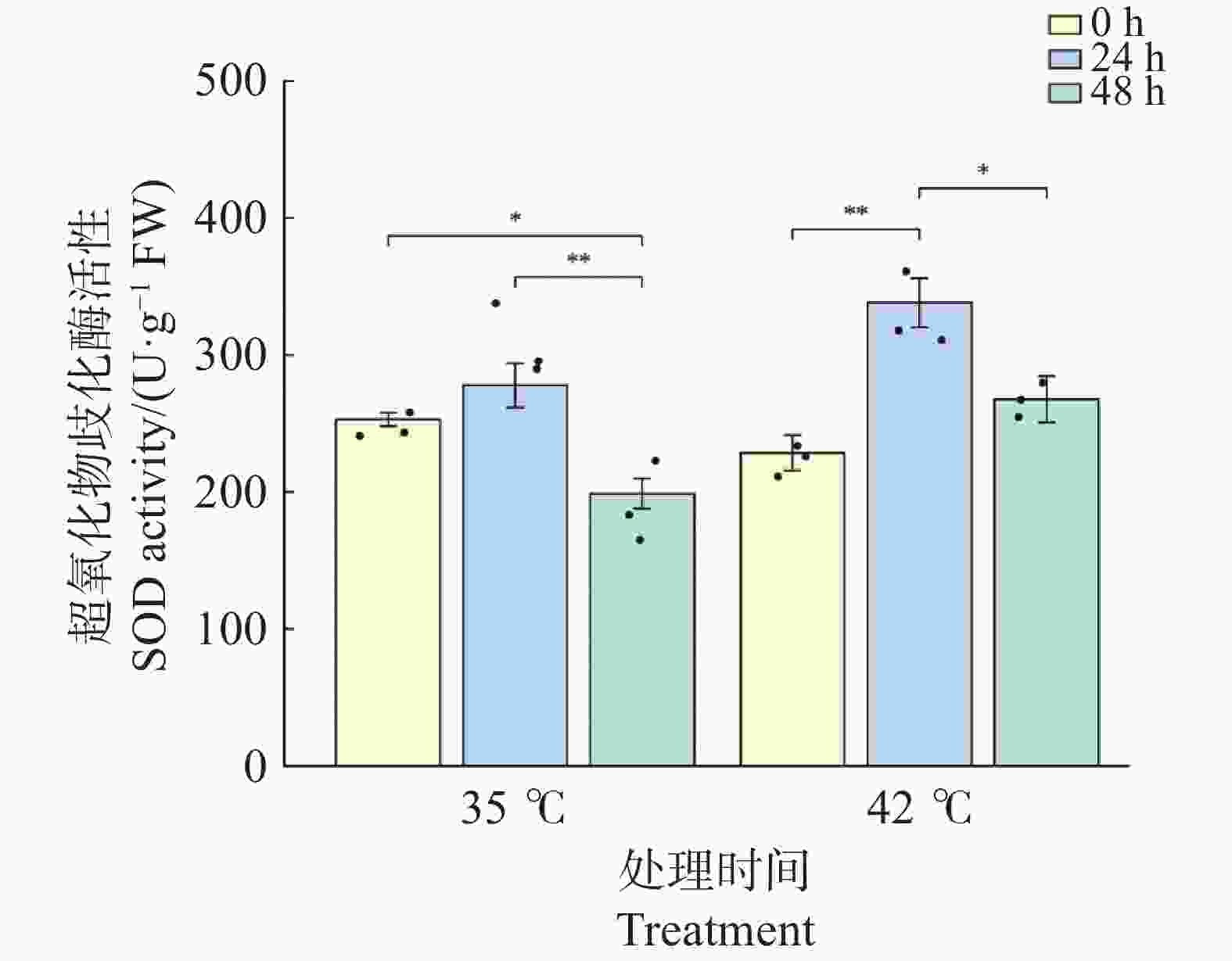
图 5 高温胁迫下文心兰超氧化物歧化酶(SOD)活性
Figure 5. Effects of heat stress on superoxide dismutase (SOD)activity in Oncidium leaves
-
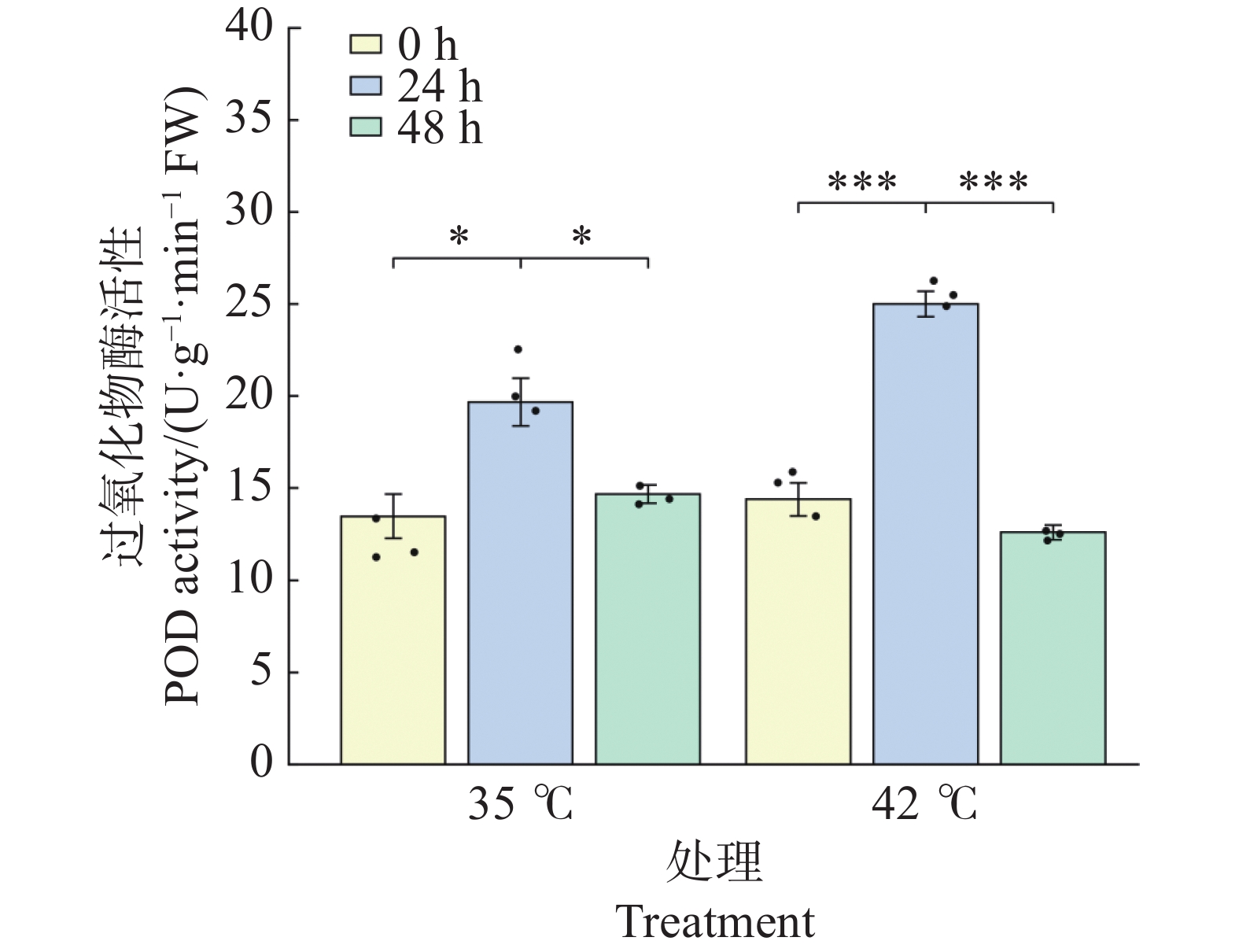
POD活性的变化趋势与SOD相似,但响应幅度更大且时间更集中(图6)。35℃处理下,POD活性在24 h较0 h显著升高46.0%(由13.47升至19.67 U·g−1·min−1,P<0.05);48 h时回落至14.67 U·g−1·min−1 ,与0 h相比无显著差异(P>0.05),提示35℃下POD仅在胁迫早期短暂激活。42℃处理下,POD响应极为剧烈:24 h时活性极显著上升至25.00 U·g−1·min−1 (较0 h升高73.6%,P<0.001),且显著高于同期35℃处理组(P<0.05);48 h时活性极显著下降至12.60 U·g−1·min−1 (较24 h下降49.6%,P<0.001),与0 h相比无显著差异(P>0.05),但显著低于同期35℃处理组(P<0.05)。结果表明,42℃高温在24 h时启动了比35℃更强烈的POD抗氧化反应,但在48 h持续胁迫后POD系统出现明显衰减,提示极端高温下POD的代偿能力存在时间限制。
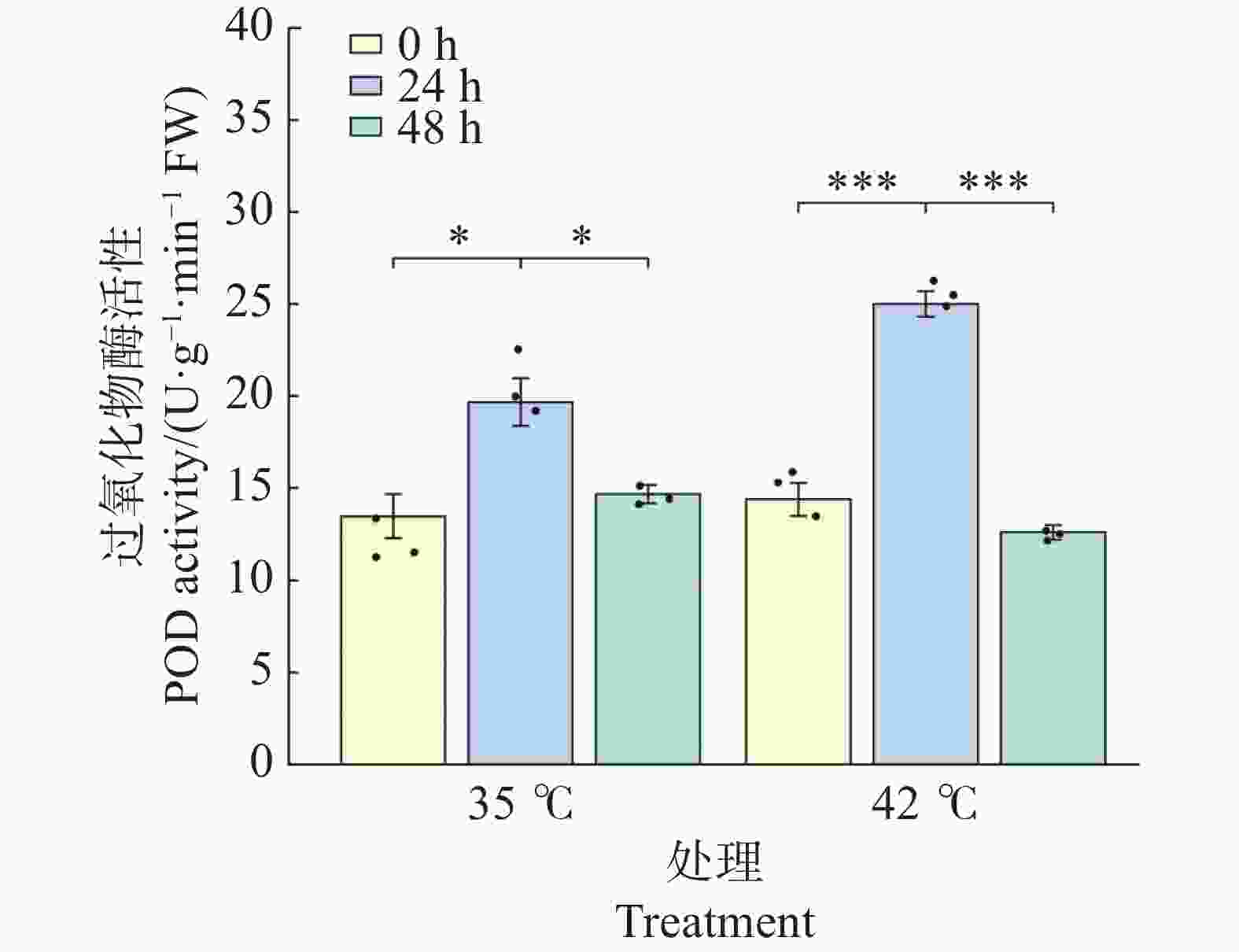
图 6 高温胁迫下文心兰过氧化物酶(POD)活性
Figure 6. Effects of heat stress on peroxidase (POD)activity in Oncidium leaves
-
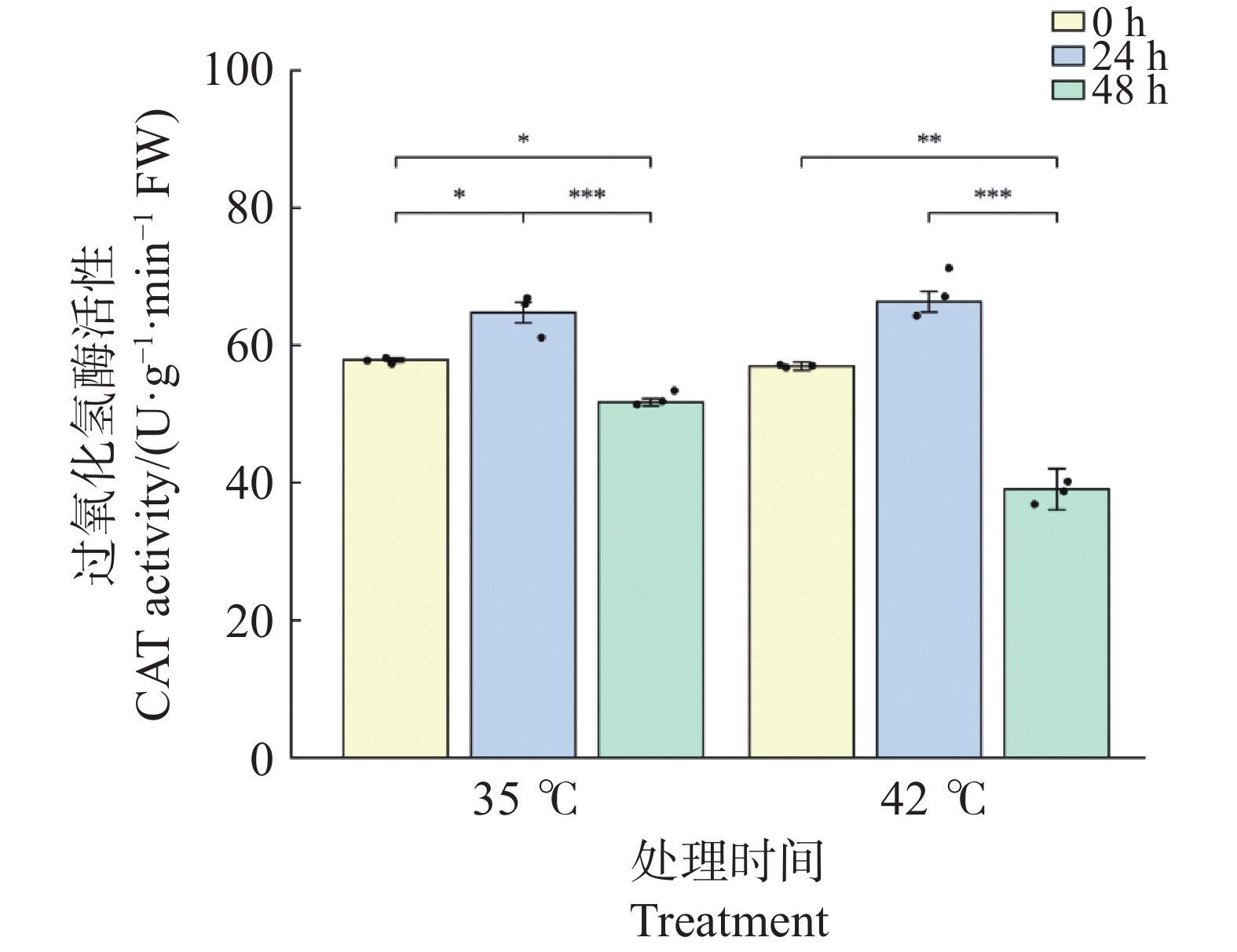
CAT活性在2种温度处理下均呈现“先升后降”的变化模式,且在胁迫后期下降幅度显著(图7)。35℃处理下,CAT活性在24 h较0 h显著升高至64.73 U·g−1·min−1(升幅11.9%,P<0.05);48 h时显著下降至51.73 U·g−1·min−1(较0 h下降10.6%,P<0.05),且较24 h极显著下降(P<0.001)。42℃处理下,CAT活性在24 h时升至66.31 U·g−1·min−1(较0 h升高16.4%,P>0.05);48 h时急剧下降至39.03 U·g−1·min−1(较0 h下降31.5%,P<0.01;较24 h下降41.2%,P<0.001),且显著低于同期35℃处理组(P<0.05)。这表明在42℃持续胁迫下,CAT活性下降幅度显著大于35℃处理,提示极端高温对CAT系统的损害在48 h时已超出植物自身的代偿范围。
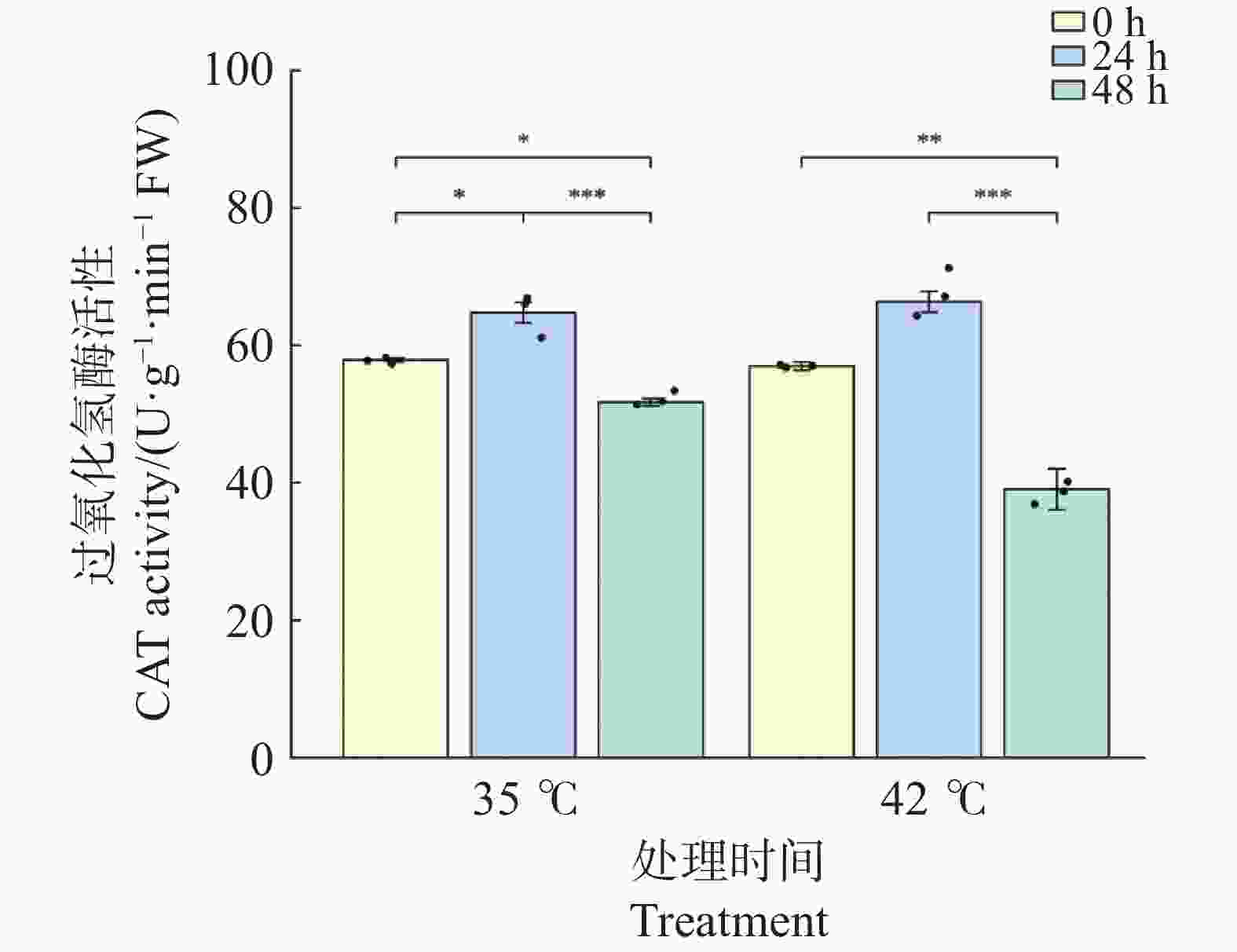
图 7 高温胁迫下文心兰过氧化氢酶(CAT)活性
Figure 7. Effects of heat stress on catalase (CAT)activity in Oncidium leaves
-
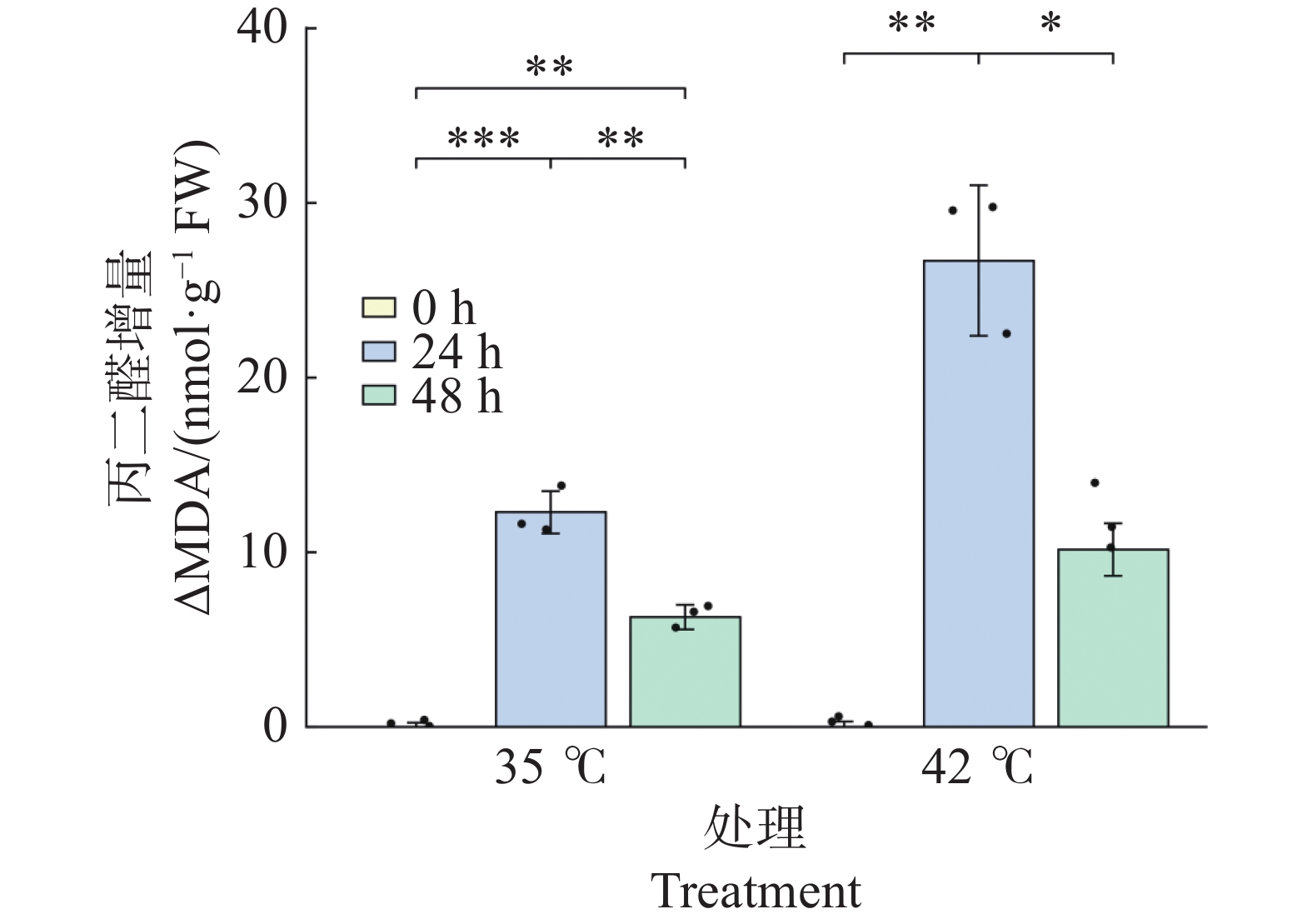
胁迫处理诱导文心兰叶片MDA水平显著上升(图8)。35℃ 处理下,24 h 时ΔMDA较0 h极显著升高(+12.28 nmol·g−1 ,P<0.001),48h时较24 h显著回落(P<0.01),但仍极显著高于0 h(P<0.01),表明35℃处理引起持续的轻度膜脂过氧化。42℃处理下,24 h时ΔMDA急剧升高至26.69 nmol·g−1 ,为同期35℃处理组的2.17倍(P<0.05),表明42℃高温对细胞膜系统的损伤显著强于 35℃;48 h时ΔMDA显著回落至10.15(较24 h下降61.9%,P<0.05),与0 h无显著差异(P>0.05),提示在持续高温下抗氧化系统介导的修复过程开始发挥作用。
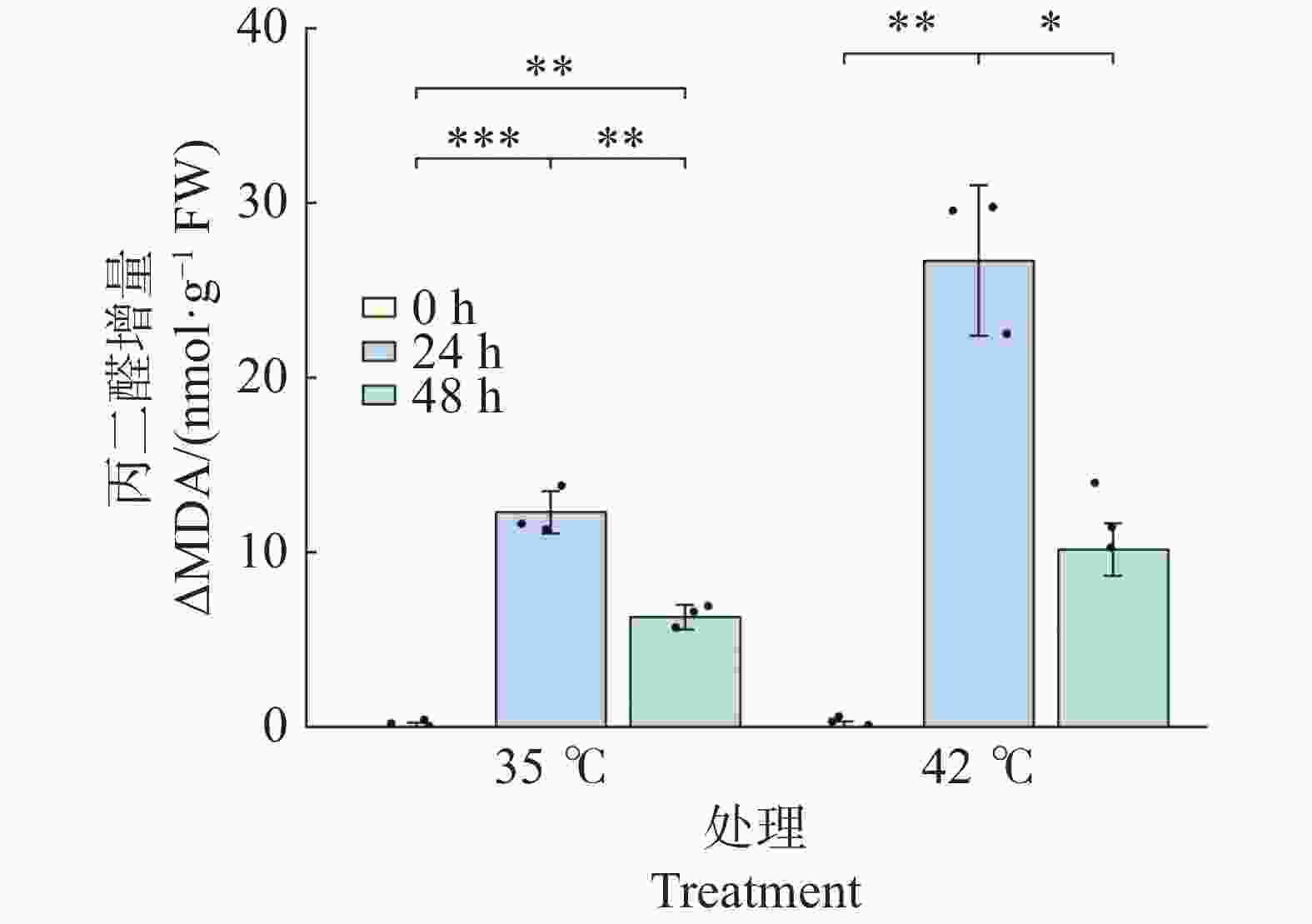
图 8 高温胁迫下文心兰丙二醛(ΔMDA)含量
Figure 8. Effects of heat stress on malondialdehyde (MDA)content in Oncidium leaves
-
高温胁迫导致文心兰叶片叶绿素相对含量(SPAD)、气孔导度(Gs)、蒸腾速率(Tr)、净光合速率(Pn)和水分利用效率(WUE)均显著下降,且损伤程度随温度升高而加剧。35℃处理48 h后,SPAD值下降33.4%,Gs下降 70.6%,Pn下降67.3%,WUE下降24.9%,各指标虽下降明显但仍保持一定的代谢活性(图2)。42℃处理下光合系统几近崩溃:48 h后Gs降至 0.007 mol·m−2·s−1(较0 h下降91.4%)(图3),Pn降至0.23 μmol·m−2·s−1(较0 h下降97.4%),WUE降至1.02 μmol CO2·mmol−1 H2O(较0 h下降79.5%),且24 h与48 h时各光合参数均极显著低于同期35℃处理组(P<0.001)(图4)。结果表明,文心兰对35℃温和高温具有一定的耐受能力,而42℃极端高温在24 h内即触发气孔几乎完全关闭,导致光合作用近乎停止。
-
文心兰抗氧化酶系统在高温胁迫下呈现出酶组分间差异化的时序响应模式(图5~图7)。在胁迫早期(24 h), 3种抗氧化酶在两种温度下均出现不同程度的活化;42℃处理下响应更为剧烈:SOD活性较0 h升高47.8%(P<0.01))(图5),POD活性升高73.6%(P<0.001)(图6),CAT 活性升高16.4%,均显著高于或与同期35℃处理组相当(图7)。在胁迫后期(48 h),3种抗氧化酶呈现分化的衰减模式:在42℃处理下,POD活性较24 h极显著下降49.6%(P<0.001),CAT活性较24 h极显著下降41.2%(P<0.001),并显著低于同期35℃处理组(P<0.05);而SOD活性虽较24 h显著回落(P<0.05),但仍维持在略高于0 h的水平(+17.0%),且仍显著高于同期35℃处理组(P<0.05)。这表明在42℃持续胁迫下,SOD的高活性维持时间长于POD和CAT,反映出三种抗氧化酶对极端高温的代偿能力存在时间差异。
-
ΔMDA 增量(以0 h为基线)的变化反映了膜脂过氧化的动态(图8)。35℃处理下,ΔMDA在24 h时升至12.28 nmol·g−1(P<0.001),48 h时显著回落至6.28 nmol/g(P<0.01)。42℃处理下,ΔMDA在24 h时急剧升至26.69 nmol·g−1,为同期35℃处理组的2.17倍(P<0.05),表明42℃对膜系统的损伤显著强于35℃;48 h时ΔMDA显著回落至10.15 nmol/g(较24 h下降61.9%,P<0.05),与0 h无显著差异(P > 0.05)。膜脂过氧化的动态变化与抗氧化酶活性的时序响应在时间上相吻合—胁迫早期MDA急剧升高对应抗氧化酶的快速激活,胁迫后期MDA部分回落则与抗氧化系统的持续作用有关。
-
综合各生理指标的变化可知,温度强度与胁迫持续时间共同决定文心兰的生理响应:35℃温和高温下,文心兰的光合作用与抗氧化系统均能维持一定的代偿能力;42℃极端高温下,虽然抗氧化酶在24 h时启动了更强烈的应答(SOD、POD升幅显著高于35℃),但48 h后POD与CAT活性显著衰减,光合作用已近乎停止(Pn下降97.4%),提示长时间(≥48 h)极端高温胁迫已超出文心兰生理代偿的时间窗口。这种“温度强度与时间”的协同损伤模式,为文心兰夏季栽培管理(如降温措施的启动时机与持续时间)提供了重要的生理参考。
-
本研究以文心兰主栽品种‘小蜜蜂’(Oncidium ‘Honey Bee’)为材料,系统探究了35℃(温和高温)和42℃(极端高温)胁迫下的生理响应机制。结果表明,高温胁迫对文心兰的光合系统和抗氧化防御系统均产生显著影响,且效应呈现温度依赖性和时间依赖性。
在光合作用方面,高温胁迫导致文心兰叶片黄化、萎蔫,叶绿素相对含量显著下降,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)及水分利用效率(WUE)均降低。42℃极端高温处理24 h时,Pn已极显著降至0.32 μmol·m−2·s−1(降幅96.3%,P<0.001),Gs降至0.010 mol·m−2·s−1(降幅87.9%);处理48 h后,Pn进一步降至0.23 μmol·m−2·s−1,光合作用近乎停止,WUE降至1.02 mmol·mol−1 ,表明极端高温对光合系统造成了不可逆损伤。35℃温和高温处理48 h后,SPAD值、Pn、Gs、Tr和WUE分别下降33.4%、67.3%、70.6%、56.0%和24.9%,各指标虽显著下降但仍维持基本代谢活性,说明文心兰对温和高温具有一定的耐受能力。这一结果与赵森等[15]在水稻(Oryza sativa)中的研究一致,该研究发现高温胁迫下净光合速率下降与叶绿素降解密切相关。本研究中,胁迫早期(24 h)Gs急剧下降,光合抑制以气孔限制为主;胁迫后期(48 h)Gs趋于稳定但Pn进一步下降,且SPAD值持续降低,表明非气孔限制(如叶绿素降解、光合机构损伤)逐渐占据主导地位,这与薛伟等[16]在胡杨(Populus euphratica)中的研究结论相吻合,即短期高温胁迫下气孔限制是光合下降的主要因素,而长期胁迫下非气孔限制逐渐占主导。
在抗氧化防御系统方面,文心兰3种抗氧化酶呈现差异化的时序响应模式。胁迫早期(24 h),SOD、POD和CAT活性均被显著激活,42℃处理下分别较0 h升高47.8%(P<0.01)、73.6%(P<0.001)和16.4%,且SOD和POD的升幅显著高于35℃处理组,反映了文心兰对极端高温的快速应答能力。胁迫后期(48 h),三种抗氧化酶呈现明显分化的衰减模式:42℃处理下,POD与CAT活性较24 h分别极显著下降49.6%和41.2%(P<0.001),而SOD活性虽较24 h显著回落,但仍维持在略高于0 h的水平(+17.0%),且显著高于同期35℃处理组(P<0.05),表明SOD在极端高温下的代偿维持时间长于POD和CAT。这一酶组分间的差异化时序响应,与崔明等[17]在豇豆(Vigna unguiculata)叶片中的研究结果一致,该研究发现,短期高温胁迫下SOD和POD活性快速上升,而CAT在长期胁迫中变化显著。同时,SOD在持续高温下维持较高活性的特征,与王弋博等[18]在苜蓿(Medicago sativa)中的研究结论相印证,即抗氧化酶在应对长期高温胁迫时存在组分特异性,部分酶在持续胁迫中具有独特的重要性。膜脂过氧化程度(以ΔMDA表示)在胁迫早期急剧加剧:42℃处理24 h时,ΔMDA升至26.69 nmol·g−1 ,为同期35℃处理组(12.28 nmol·g−1 )的2.17倍(P<0.05),表明42℃高温对细胞膜系统的损伤显著强于35℃。胁迫后期(48 h),ΔMDA显著回落:42℃处理组降至10.15 nmol·g−1 (较24 h下降61.9%,P<0.05),与0 h无显著差异(P>0.05);35℃处理组也由12.28 nmol·g−1显著回落至6.28 nmol·g−1 (P<0.01)。值得注意的是,ΔMDA的回落与SOD活性的持续维持在时间上相吻合,而非与CAT相关,提示SOD介导的超氧阴离子清除可能在胁迫后期的膜系统修复中发挥了关键作用。MDA先升后降的动态规律与前人研究结果相符[19],印证了抗氧化系统在胁迫后期对膜损伤的修复作用。
综合各生理指标的变化可知,高温胁迫对文心兰的生理影响既依赖于胁迫强度,也依赖于胁迫持续时间:35℃温和高温下,文心兰的光合作用与抗氧化系统均能维持一定的代偿能力;42℃极端高温下,尽管抗氧化酶在24 h时启动了更强烈的响应,但48 h后POD和CAT已显著衰减,光合作用近乎停止,提示长时间(≥ 48 h)极端高温胁迫已超出文心兰的生理代偿能力。这一“温度强度 × 时间”的协同损伤特征,与Yeh等[20]关于植物热胁迫耐受性研究的结论一致,即热胁迫的影响程度取决于胁迫的强度、持续时间及植物自身的适应能力。
与其他兰科植物的比较进一步支持上述结论。本研究表明,文心兰对35℃温和高温表现出一定的耐受能力,但对42℃极端高温较为敏感,24 h内即发生气孔近乎完全关闭与光合作用近乎停止。这一结果与罗远华等[4]在文心兰中的研究一致,该研究发现文心兰在高温下叶绿素含量下降、抗氧化酶活性发生显著变化;同时与孙映波等[21]的研究结果相印证,即文心兰在逆境胁迫下通过调节抗氧化酶活性来应对环境胁迫。此外,本研究揭示的抗氧化酶组分间差异化的时序响应模式(SOD维持时间长于POD和CAT),与Huang等[22]在蝴蝶兰中的研究结果相印证,该研究发现不同兰科植物对高温的响应存在差异,但抗氧化酶系统的激活及其组分间的协同是其共同的防御机制。
综上所述,本研究系统揭示了文心兰应对不同强度高温胁迫的生理响应特征,明确了其光合系统对极端高温的快速崩溃机制以及抗氧化酶组分间差异化时序响应的防御策略。42℃极端高温在24 h内即触发文心兰光合系统近乎崩溃,而抗氧化酶系统对极端高温的代偿存在明确的时间窗口与组分差异。这一发现不仅丰富了兰科植物逆境生理学的理论内涵,也为文心兰耐热性评价、夏季栽培管理优化(如降温措施的启动时机与持续时间)以及耐热品种选育提供了理论依据。建议在文心兰夏季栽培管理中,当棚温接近42℃时应在24 h内启动降温措施(如喷雾、加大通风、补充遮阴),以避免不可逆的光合损伤。
由于试验材料的限制,本研究未能直接观察高温胁迫下气孔开度、密度及保卫细胞形态的变化,气孔导度(Gs)的下降仅作为气孔关闭的间接指标。未来研究应进一步结合转录组学和代谢组学手段,深入解析抗氧化酶系统差异调控的分子机制,并借助扫描电镜与光学显微镜直接观察气孔形态,从而完善高温胁迫下文心兰光合作用与抗氧化防御协同调控的机制研究。
Physiological response mechanisms of Oncidium under heat stress
-
摘要: 为探明‘小蜜蜂’文心兰( Oncidium‘Honey Bee’)对高温胁迫的生理响应机制,以35℃(温和高温)和42℃(极端高温)处理0、24、48 h,系统测定了其叶片表型变化、叶绿素相对含量(SPAD值)、光合参数(净光合速率Pn、气孔导度Gs、蒸腾速率Tr、水分利用效率WUE)以及抗氧化酶(超氧化物歧化酶SOD、过氧化物酶POD、过氧化氢酶CAT)活性和丙二醛(MDA)含量等生理指标。结果表明,1) 高温胁迫显著抑制文心兰光合作用,且抑制程度具温度依赖性。35℃处理48 h后,SPAD 值、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和水分利用效率(WUE)分别下降33.4%、67.3%、70.6%、56.0%和24.9%;42℃ 处理24 h后,Pn即降至0.32 μmol·m−2·s−1(降幅 96.3%,P<0.001),Gs降至0.010 mol·m−2·s−1(降幅 87.9%),48 h后,Pn几近停止,光合系统接近崩溃,42℃处理组各光合参数均极显著低于同期 35℃处理组(P<0.001)。2) 3种抗氧化酶在胁迫早期(24 h)被显著激活,42℃处理下 SOD、POD、CAT活性较0 h分别升高47.8%(P<0.01)、73.6%(P<0.001)和 16.4%;胁迫后期(48 h) 3种抗氧化酶呈差异化衰减:POD和CAT活性较24 h分别极显著下降49.6%和41.2%(P<0.001),而SOD活性仍维持在略高于0 h的水平(+17.0%),且显著高于同期35℃处理组(P<0.05),表明SOD对极端高温的代偿维持时间长于POD 和CAT。3) 膜脂过氧化(以ΔMDA表示)在24 h时急剧升高,42℃处理组ΔMDA(26.69 nmol·g−1 FW)为同期35℃处理组的2.17倍(P<0.05);48 h时ΔMDA 显著回落,与0 h 无显著差异(P>0.05),提示抗氧化系统在中后期发挥了一定的修复作用。研究显示,42℃极端高温在24 h内即可触发文心兰气孔近乎完全关闭、光合作用近乎停止,而抗氧化酶系统对极端高温的代偿存在明确的时间窗口与组分差异,长时间(≥ 48 h)极端高温胁迫已超出其生理代偿能力。Abstract: To investigate the physiological responses of Oncidium 'Honey Bee' to heat stress, plants of Oncidium 'Honey Bee' were exposed to 35℃ (moderate heat) and 42℃ (extreme heat) for 0, 24, and 48 h, and phenotypic changes, changes in chlorophyll content (SPAD value), photosynthetic gas exchange parameters, antioxidant enzyme activities, and lipid peroxidation level were systematically determined. Results showed that heat stress significantly suppressed photosynthesis in a temperature-dependent manner. After 48 h of treatment at 35℃, SPAD value, net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and water-use efficiency (WUE) of the plants decreased by 33.4%, 67.3%, 70.6%, 56.0%, and 24.9%, respectively. Under 42℃ stress, Pn dropped to 0.32 μmol·m−2·s−1 within 24 h (a 96.3% decrease, P<0.001) and Gs to 0.010 mol·m−2·s−1 (a 87.9% decrease); by 48 h, photosynthesis was nearly halted, with all photosynthetic parameters in the 42℃ group being significantly lower than those in the 35℃ group at corresponding time points (P<0.001). The three antioxidant enzymes were significantly activated during the early phase (24 h). Under 42℃ stress, SOD, POD, and CAT activities increased by 47.8% (P<0.01), 73.6% (P<0.001), and 16.4%, respectively at the early phase as against those at 0 h. During the late phase (48 h), the three enzymes exhibited differential attenuation patterns: POD and CAT activities decreased by 49.6% and 41.2% as against those at 24 h (P<0.001), while SOD activity remained 17.0% above the 0 h baseline and was significantly higher than that in the 35℃ group (P<0.05), indicating that SOD sustained its compensatory activity longer than POD and CAT under extreme heat. Lipid peroxidation, expressed as ΔMDA, increased sharply at 24 h, with the 42℃ group (26.69 nmol·g−1) being 2.17-fold higher than the 35℃ group (P<0.05). By 48 h, ΔMDA decreased significantly and was not significantly different from that at 0 h (P>0.05), suggesting partial repair by the antioxidant system. In conclusion, extreme heat at 42℃ triggers near-complete stomatal closure and cessation of photosynthesis within 24 h in Oncidium, while the antioxidant system shows a clear temporal window and component-specific differentiation in compensation for extreme heat, with prolonged (≥48 h) extreme heat stress exceeding the plant's physiological compensatory capacity. These findings provide a physiological basis for determining the timing and duration of cooling measures in summer cultivation of Oncidium.
-
Key words:
- Oncidium /
- heat stress /
- photosynthesis /
- antioxidant enzymes /
- malondialdehyde
-
图 2 高温胁迫对文心兰叶绿素含量的影响
注:柱形为均值,误差线为标准误(SE),散点为各重复实测值;SPAD 的样本量n=9。* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 2 Effects of heat stress on chlorophyll content (SPAD value)in Oncidium leaves
Note: Bars represent means; error bars represent standard errors (SE); dots represent individual replicate values; sample size of SPAD was n=9. * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 3 高温胁迫对文心兰气孔导度(GS)与蒸腾速率(Tr)的影响
注:(a)气孔导度 Gs;(b)蒸腾速率 Tr。柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=8~12)。* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 3 Effects of heat stress on stomatal conductance (Gs)and transpiration rate (Tr)in Oncidium leaves
Note: (a): Stomatal conductance Gs; (b): Transpiration rate Tr. Bars represent means; error bars represent SE; dots represent individual replicate values (n=8–12). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 4 高温胁迫对文心兰净光合速率(Pn)与水分利用效率(WUE)的变化
注:(a)净光合速率 Pn;(b)水分利用效率 WUE。柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=8~12)。* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 4 Effects of heat stress on net photosynthetic rate (Pn)and water use efficiency (WUE)in Oncidium leaves
Note: (a): Net photosynthetic rate Pn; (b): Water-use efficiency WUE. Bars represent means; error bars represent SE; dots represent individual replicate values (n=8–12). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 5 高温胁迫下文心兰超氧化物歧化酶(SOD)活性
注:柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=3);* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 5 Effects of heat stress on superoxide dismutase (SOD)activity in Oncidium leaves
Note: Bars represent means; error bars represent SE; dots represent individual replicate values (n=3). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 6 高温胁迫下文心兰过氧化物酶(POD)活性
注:柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=3);* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 6 Effects of heat stress on peroxidase (POD)activity in Oncidium leaves
Note: Bars represent means; error bars represent SE; dots represent individual replicate values (n=3). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 7 高温胁迫下文心兰过氧化氢酶(CAT)活性
注:柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=3);* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 7 Effects of heat stress on catalase (CAT)activity in Oncidium leaves
Note: Bars represent means; error bars represent SE; dots represent individual replicate values (n=3). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
图 8 高温胁迫下文心兰丙二醛(ΔMDA)含量
注:ΔMDA = MDA_t − MDA_0(以各处理组0 h均值为基线)。柱形为均值,误差线为标准误(SE),散点为各重复实测值(n=3)。* P<0.05,** P<0.01,*** P<0.001(Tukey HSD)。
Fig. 8 Effects of heat stress on malondialdehyde (MDA)content in Oncidium leaves
Note: ΔMDA = MDA_t − MDA_0 (the mean of each treatment group at 0 h was used as the baseline). Bars represent means; error bars represent SE; dots represent individual replicate values (n=3). * P<0.05; ** P<0.01; *** P<0.001 (Tukey HSD).
-
[1] 陈俊玲, 杨光穗, 陈金花. 新形势下文昌花卉产业发展策略研究[J]. 热带农业科学, 2021, 41(7): 119−124. https://doi.org/10.12008/j.issn.1009-2196.2021.07.021 doi: 10.12008/j.issn.1009-2196.2021.07.021 [2] 罗远华, 张元法, 林兵. 福建省文心兰产业现状与发展对策[J]. 农业与技术, 2025, 45(14): 115−117. https://doi.org/10.19754/j.nyyjs.20250730024 doi: 10.19754/j.nyyjs.20250730024 [3] 张叶, 叶蓓蕾, 邬静, 等. 77份文心兰种质资源表型性状遗传多样性分析[J]. 热带作物学报, 2021, 42(8): 2183−2190. https://doi.org/10.3969/j.issn.1000-2561.2021.08.009 doi: 10.3969/j.issn.1000-2561.2021.08.009 [4] 罗远华, 方能炎, 林榕燕, 等. 温度对文心兰生长特性和生理指标的影响[J]. 福建农业学报, 2018, 33(7): 702−707. https://doi.org/10.19303/j.issn.1008-0384.2018.07.010 doi: 10.19303/j.issn.1008-0384.2018.07.010 [5] 罗远华, 王振波, 黄敏玲, 等. 高温胁迫对文心兰顶叶若干生理指标的影响[J]. 福建农业学报, 2017, 32(6): 625−629. https://doi.org/10.19303/j.issn.1008-0384.2017.06.010 doi: 10.19303/j.issn.1008-0384.2017.06.010 [6] Awasthi R, Bhandari K, Nayyar H. Temperature stress and redox homeostasis in agricultural crops [J]. Frontiers in Environmental Science, 2015, 3: 11. https://doi.org/10.3389/fenvs.2015.00011 doi: 10.3389/fenvs.2015.00011 [7] Zhang Y C, Li C N, Wang Y. Characterization of a salt-tolerant plant growth-promoting bacterial isolate and its effects on oat seedlings under salt stress [J]. Agronomy, 2026, 16(1): 135. https://doi.org/10.3390/agronomy16010135 doi: 10.3390/agronomy16010135 [8] 穆心愿, 宋淼, 马智艳, 等. 夏玉米光合特性及产量对花期高温胁迫的响应[J]. 南方农业学报, 2025, 56(5): 1361−1378. https://doi.org/10.3969/j.issn.2095-1191.2025.05.002 doi: 10.3969/j.issn.2095-1191.2025.05.002 [9] 胡玥, 申建双, 潘向东, 等. 高温胁迫对杜鹃光合作用及叶绿素荧光的影响[J]. 分子植物育种, 2026, 24(5): 1608−1615. https://doi.org/10.13271/j.mpb.024.001608 doi: 10.13271/j.mpb.024.001608 [10] 刘子强, 姜文海, 毛琳俐, 等. 短时高温胁迫对毛竹叶片生理特性的影响[J]. 浙江林业科技, 2025, 45(5): 26−34. https://doi.org/10.3969/j.issn.1001-3776.2025.05.004 doi: 10.3969/j.issn.1001-3776.2025.05.004 [11] 司宝华, 吕明皓, 马文舒, 等. 六种蝴蝶兰新品种高温生理响应及耐热性评价[J]. 北方园艺, 2025(12): 82−88. https://doi.org/10.11937/bfyy.20243571 doi: 10.11937/bfyy.20243571 [12] 张沥曼玲. 高温胁迫下金钗石斛生理特征与分子调控基础分析[D]. 林芝: 西藏农牧学院, 2025. https://doi.org/10.27979/d.cnki.gadce.2025.000097 [13] 罗小燕, 易双双, 李崇晖, 等. 兰花热胁迫响应机制及耐热性研究进展[J]. 分子植物育种, 2023, 21(19): 6406−6412. https://doi.org/10.13271/j.mpb.021.006406 doi: 10.13271/j.mpb.021.006406 [14] 罗小燕, 罗远华, 易双双, 等. 不同文心兰品种耐热性指标筛选与评价体系构建[J]. 热带作物学报, 2024, 45(7): 1348−1358. https://doi.org/10.3969/j.issn.1000-2561.2024.07.006 doi: 10.3969/j.issn.1000-2561.2024.07.006 [15] 赵森, 于江辉, 肖国樱. 高温胁迫对爪哇稻剑叶光合特性和渗透调节物质的影响[J]. 生态环境学报, 2013, 22(1): 110−115. https://doi.org/10.16258/j.cnki.1674-5906.2013.01.022 doi: 10.16258/j.cnki.1674-5906.2013.01.022 [16] 薛伟, 李向义, 林丽莎, 等. 短时间热胁迫对疏叶骆驼刺光系统Ⅱ、Rubisco活性和活性氧化剂的影响[J]. 植物生态学报, 2011, 35(4): 441−451. https://doi.org/10.3724/SP.J.1258.2011.00441 doi: 10.3724/SP.J.1258.2011.00441 [17] 崔萌, 魏娟娟, 苏晓星, 等. 高温对豇豆叶片细胞膜脂过氧化和蛋白质表达的影响[J]. 西南大学学报(自然科学版), 2016, 38(4): 21−27. https://doi.org/10.13718/j.cnki.xdzk.2016.04.003 doi: 10.13718/j.cnki.xdzk.2016.04.003 [18] 王仪明, 李伟民, 雷艳芳, 等. 高温胁迫对淮阴苜蓿差异蛋白表达及生理的影响[J]. 中国生态农业学报, 2015, 23(4): 441−446. https://doi.org/10.13930/j.cnki.cjea.141117 doi: 10.13930/j.cnki.cjea.141117 [19] 周文豪, 郑永润, 傅强, 等. 五个杜鹃品种耐热性评价及大鸳鸯锦杜鹃对高温胁迫的生理响应[J]. 湖北农业科学, 2025, 64(9): 107−113. https://doi.org/10.14088/j.cnki.issn0439-8114.2025.09.018 doi: 10.14088/j.cnki.issn0439-8114.2025.09.018 [20] Yeh C H, Kaplinsky N J, Hu C, et al. Some like it hot, some like it warm: phenotyping to explore thermotolerance diversity [J]. Plant Science, 2012, 195: 10−23. https://doi.org/10.1016/j.plantsci.2012.06.004 doi: 10.1016/j.plantsci.2012.06.004 [21] 孙映波, 尤毅, 朱根发, 等. 干旱胁迫对文心兰抗氧化酶活性和渗透调节物质含量的影响[J]. 生态环境学报, 2011, 20(11): 1675−1680. https://doi.org/10.3969/j.issn.1674-5906.2011.11.015 doi: 10.3969/j.issn.1674-5906.2011.11.015 [22] Huang L C, Lin C J, Kuo C I. Effects of high temperature on growth and flowering of Oncidium orchids [J]. Horticultural Science, 2016, 43(2): 45−52. (查阅网上资料, 未找到本条文献信息, 请确认) -

 点击查看大图
点击查看大图
计量
- 文章访问数: 155
- HTML全文浏览量: 98
- 被引次数: 0
