


 下载:
下载:



-
堇菜属Viola L.是堇菜科Violaceae Batsch中最大的属,全球共有525~600种[1-4]。东亚山区是堇菜属形态与分类多样性的中心之一[5]。该属被划分为14个组,其中物种最丰富的堇菜组sect. Viola下进一步细分为17个亚组[6]。而匍匐堇菜组sect. Diffusae原由Becker(1925)归为堇菜组下的一个亚组subsect. Diffusae分出来,包含约4种,分布于南亚和东南亚。其鉴别特征为:花期具匍匐茎,茎顶端莲座状着生叶片,柱头微2裂[7]。山柑属Capparis Tourn. ex L.隶属于山柑科Capparaceae Juss.包含约146种植物,广泛分布于旧大陆热带及亚热带地区[8-10],少数种类延伸至中亚和地中海区域[11]。该属的形态特征包括:单叶互生,全缘,少见戟形;花两侧对称,排列成簇生、近伞形、总状或腋上生花序,稀单生;萼片4枚排成2轮;花瓣4枚背瓣通常不对称且基部合生;雄蕊6 ~ 200;浆果[12]。广义报春苣苔属Primulina s.l.[13]作为苦苣苔科Gesneriaceae的一个重要类群,在2011年经过重新界定,整合了小花苣苔属Chiritopsis W.T.Wang、文采苣苔属Wentsaiboea D.Fang & D.H.Qin(除天等文采苣苔W. tiandengensis Yan Liu & B. Pan外)以及唇柱苣苔属Chirita唇柱苣苔组sect. Gibbosaccus C.B.Clarke的大量物种,修订后的报春苣苔属已成为中国苦苣苔科中物种多样性最丰富的类群,包含约123种8变种[14-15]。该属的形态特征包括:多为多年生草本,常具根状茎或块茎;叶片通常基生,呈莲座状排列,形态多样,部分种类叶面具有银白色斑纹或绒毛;花冠二唇形(上唇2裂,下唇3裂),花色丰富,常见紫色、白色、黄色等;花萼5裂;雄蕊2枚,具退化雄蕊(部分种类无);子房上位,柱头2裂或头状;果实为蒴果,成熟时纵裂,种子细小;报春苣苔属主要分布在中国华南至西南地区,生境为特殊的喀斯特石灰岩地区[16]。魔芋属Amorphophallus Blume ex Decaisne共包含239个物种[17],是天南星科Araceae Juss.中物种最丰富的属之一[18],该属的形态特征包括:拉长的块茎、中等尺寸的花序、内表面具细长乳突的佛焰苞基部、佛焰苞基部与檐部无明显缢缩区分、以及具显著花柱的柱头[17-18]。黄杞属Engelhardia Lesch. ex Bl.[19-23]隶属于胡桃科Juglandaceae DC. ex Perleb,主要分布于东亚的热带和亚热带地区、中南半岛及马来群岛[20]。
海南尖峰岭位于海南省西南部,隶属乐东黎族自治县,与东方市交界(18°20'—18°57'N,108°41'—109°12'E),位于热带北缘,气候类型为低纬度热带岛屿季风气候,总面积约640 km2,保护区由未经破坏的原始热带雨林与生态恢复区域的次生林组成,其生境异质性较高,丰富的热带、亚热带物种组成独特的热带雨林景观[24-25]。2023年10月至2025年6月在海南热带雨林国家公园尖峰岭片区进行植物调查,笔者对园内植物拍摄照片及科学用途采集的标本进行整理及鉴定过程中,查阅中国植物志[7,12,16,21]、海南植物名录[26]、中国热带雨林地区植物图鉴—海南植物(1~3卷)[27],以及相关植物类群分类文献[28],发现并采集到的有4个海南省新记录种,在查阅物种名录与标本时,发现云南黄杞Engelhardia spicata Lesch. ex Blume在海南有分布,现予以报道。
-
本研究收集了南岭堇菜Viola nanlingensis J. S. Zhou & F. W. Xing、薄叶山柑Capparis tenera Dalz.、黄斑报春苣苔Primulina flavimaculata(W. T. Wang)Mich. Möller & A. Weber、东京魔芋Amorphophallus tonkinensis Engl. & Gehrm(图1−4)4个种的原始文献、性状数据、植物精细解剖图和云南黄杞的原始文献、性状数据、模式标本。

图 1 南岭堇菜
Figure 1. Viola nanlingensis
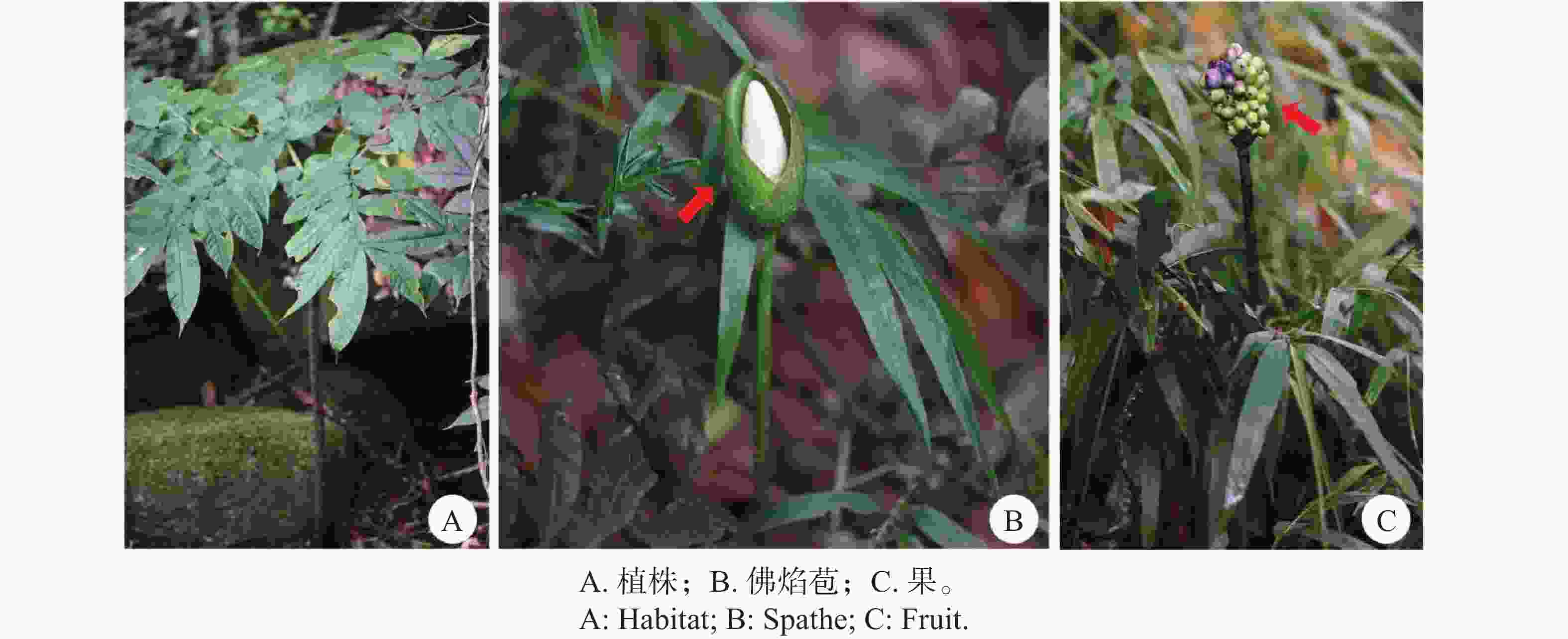
图 4 东京魔芋
Figure 4. Amorphophallus tonkinensis
-
本研究采集的南岭堇菜、薄叶山柑、黄斑报春苣苔、东京魔芋标本及植物照片均来自海南热带雨林国家公园尖峰岭片区内,基于其原始文献描述和模式,每种植物选择3份标本用于性状指标观察、测量、描述。此外,本研究观察、测量、描述与分析的南岭堇菜标本材料来源于华南国家植物园的标本馆(IBSC)。薄叶山柑、黄斑报春苣苔、云南黄杞的标本照片来自JSTOR(图2-A; 图3-D;
https://www.jstor.org/ )。
图 2 薄叶山柑
Figure 2. Capparis tenera

图 3 黄斑报春苣苔
Figure 3. Primulina flavimaculata
-
研究对南岭堇菜、薄叶山柑、黄斑报春苣苔、东京魔芋和云南黄杞的植株整体形态、叶片、花、果等与模式进行了对比,然后基于5者的原白(protologue)[4-5,8]、期刊文献[6,9-10]、植物志[7,11,28]等,确认南岭堇菜、薄叶山柑、黄斑报春苣苔、东京魔芋为海南省新记录,此外,对尖峰的海南黄杞与云南黄杞进行形态学的比对、观察、测量与描述。标本鉴定主要依据对其关键形态性状的观察和测量,包括但不限于:营养器官性状(叶片形状、长宽、毛被情况、叶尖与叶基形态、叶柄长度);生殖器官性状(花序类型、花萼与花瓣的数量与形态、长宽、雄蕊的数目与着生方式等)。对叶片、花、果等关键结构,使用数显游标卡尺(精度0.01 mm)进行至少10次重复测量,结果以性状差异的最小值与最大值表示。
-
Viola nanlingensis J. S. Zhou & F. W. Xing in Ann. Bot. Fennici. 45:233–236. 2008.——HOLOTYPE:China. Guangdong Province: Shaoguan City, Nanling National Nature Reserve, Ruyang administration, moist places along roadsides, alt.500 m,25.III.2004 J.S. Zhou 117(IBSC!).——PARATYPE: China. Guangdong Province: Shaoguan City, NanlingNational Nature Reserve, Ruyang administration, alt.500 m, 20. VII.2004 J.S. Zhou697(IBSC!).(图1-F—G)
多年生草本。无地上茎,具匍匐枝,在匍匐枝顶端常生出新的植株;根状茎垂直。叶基生;托叶披针形,长3 ~ 4 cm,宽2 ~ 3 mm,基部与叶柄贴生,边缘具疏流苏,先端渐尖;叶片卵形或椭圆形,长3 ~ 7 cm,宽2 ~ 4 cm,两面沿叶脉及叶缘被短硬毛,基部心形,边缘具圆齿,先端急尖。花生于叶腋;花梗长4 ~ 17 cm;小苞片条形,2,生于花梗中上部;萼片条状披针形,长5 ~ 8 mm,边缘具疏流苏,先端尖;花浅紫色,直径2 ~ 3.5 cm,下方1瓣短,具有紫色条纹,先端尖,距长2 ~ 2.5 mm,子房无毛,花柱长条型,基部膝曲,柱头顶端浅二裂,先端具喙。蒴果卵球形,长5 ~ 7 mm。种子卵圆形,褐色带斑点,长1 ~ 1.2 mm。花期:3 ~ 5月。果期:7 ~ 10月。
分布:中国特有种,产广东、香港、澳门、海南、广西、江西、湖南和湖北。海南为首次记录,产于海南热带雨林国家公园尖峰岭片区,生于林缘溪水边。
凭证标本:海南热带雨林国家公园尖峰岭片区,18.43°N,108.50°E,海拔:800 m,2025年3月22日,XMY20250326(CSH!)。
-
Capparis tenera Dalz. in Hook., Journ. Bot. Kew Misc. 2: 41. 1850; Hook. f. et Thoms. in Hook. f., Fl. Brit. Ind. 1: 179. 1872; Gagn., Fl. Gen. Indo–Chin 1: 183. 1908; Jacobs in Blumea 12(3): 497. 1965; 云南植物志2: 54. 1979; 中国高等植物图鉴补编1: 701. 1982, in davi.——ISOTYPE(G!).(图2-A)
大乔木或小乔木,幼时为灌木或藤本状,高8 m。新生枝无毛或被短绒毛立即变无毛;刺长3 ~ 4 mm,尖利,外弯,但在小枝上细小常上举或微内弯。叶椭圆形、卵形或倒卵形,草质至近革质,长4 ~ 7 cm,宽1.8 ~ 4.0 cm,长约为宽的1.8 ~ 2.3倍,基部急尖至近圆形,顶端锐尖或亚渐尖,尖头长3 ~ 7( ~ 10)mm,中脉较细,表面中部以下浅凹,侧脉4 ~ 6对,网状脉稍明显;叶柄短,长3 ~ 5 mm。花(1 ~ )2 ~ 3朵排成一短纵列,生于叶腋上;花梗长1.5 ~ 2.5 cm,纤细,无毛;萼片外面无毛,内面及边缘有时被毛,长4 ~ 5 mm,宽2 ~ 3 mm,外轮稍宽,卵形,稍内凹,内轮略狭,椭圆形,顶端急尖或钝形;花瓣白色,上面1对稍宽略短,近基部红色,下面1对稍长略狭长4 ~ 7 mm,宽约2 mm,内外均被绒毛;雄蕊14–18( ~ 21),有时更少;雌蕊柄长1.5 ~ 2.5 cm,丝状,无毛;子房梨形,长1 ~ 1.5 mm,1室,胎座2,胚珠数个;花柱与柱头不分明,小丘状。浆果球形,光滑,直径7 ~ 10 mm,很少具短尖头,成熟时黑色,果时花梗与雌蕊柄均不增粗;种子1 ~ 2( ~ 4)粒,种皮褐色。花期2 ~ 4月,果期6 ~ 7月。
分布:国内产于云南、西藏;国外见于安达曼群岛、阿萨姆、印度、老挝、缅甸、斯里兰卡、泰国等地。海南为首次记录,产于海南热带雨林国家公园尖峰岭片区,生于林缘溪水边。
凭证标本:海南热带雨林国家公园尖峰岭片区,18.38°N,108.54°E,海拔:346 m,2025年3月10日,XMY20250310(IBSC!)。
-
Primulina flavimaculata(W. T. Wang)Mich. Möller & A. Weber in Bull. Bot. Res. 5(3): 40(1985).(图3-D)
多年生草本。圆柱形根状茎,直立或匍匐状,直径可达2.2 cm。叶生于根状茎顶端;叶片纸质,长圆状椭圆形,长8 ~ 26 cm,宽4.5 ~ 14.0 cm,顶端微尖,基部宽楔形,边缘有锯齿或近全缘,两面被短柔毛,侧脉每侧4 ~ 6条。花序腋生于叶腋处,长7 ~ 12 cm,2 ~ 4回分枝,有7 ~ 28花;花序梗长20 ~ 30 cm,被短柔毛;苞片对生,三角形,长10 ~ 14mm,宽3 ~ 4 mm,被柔毛;花梗长1.2 ~ 3.0 cm,被短柔毛。花萼紫红色,长约1.2 cm,宽约2 mm,披针形,深裂至基部;花冠蓝紫色,被短柔毛,在上唇之下有1黄斑;筒长约2.4cm,筒口直径约1 cm;上唇2浅裂,裂片半圆形,下唇3裂近中部,裂片卵形。可育雄蕊2,着生于距花冠基部1.4 cm处,连接处有1黄斑,长10 mm,顶部有短柔毛,花药长2.2 mm,无毛;退化雄蕊3,侧生的长5 ~ 6 mm,中央的长0.8 mm。雌蕊长2.9 ~ 3.2 cm,密被短柔毛,子房长2.0 ~ 2.3 cm;柱头狭椭圆形,长2.2 ~ 3.0 mm,不分裂。花果期全年。
分布:中国特有种,产广西。海南为首次记录,产于海南热带雨林国家公园尖峰岭片区,生于山体岩壁间。
凭证标本:海南热带雨林国家公园尖峰岭片区,18.43°N,108.50°E,海拔:800 m,2025年3月10日,XMY20250330(IBSC!)。
-
Amorphophallus tonkinensis Engler & Gehrmann, Pflanzenr. 48(IV. 23C): 87. 1911.
块茎棕色,略具光泽,扁球形,直径至少9 cm,根痕微隆起。叶单生;叶柄均匀淡绿色或基部为淡灰绿色,近基部及地下部分具明显的蓝晕,并具白色斑点,这些斑点通常宽大于长,形状不规则,散生于整个表面,具短纵生的黑色小点,这些点在近基部常隆起,叶柄长80–200 cm,粗2.5 ~ 7.0 cm,无毛,肿胀;叶片深裂,直径80 ~ 200 cm;叶轴仅自基部主要分枝处起向远端具翅,有时部分节膨大并形成腋生珠芽;小叶椭圆状披针形至披针形,长8.5 ~ 24.0 cm,宽3.5 ~ 6.0 cm,长渐尖。花序单生,具长花序梗;花序梗颜色同叶柄,长约22 ~ 50 cm,直径约1 cm。佛焰苞基部短旋卷,顶部直立或弯拱覆盖肉穗花序,外部绿色,具少量横向白色斑点,内部白绿色,具细小点状或略伸长的疣状凸起,卵形,长8 ~ 20 cm,宽5 ~ 17 cm,强烈凹形;外部檐部暗绿色,带不明显的黑紫色晕及少量散生的小白点,内部亮绿色,具少量小白斑。肉穗花序无柄,短于或近等长于佛焰苞,长7.5 ~ 17.0cm;雌花序区圆柱形或略倒圆锥形,长1.0 ~ 1.5 cm,粗1 ~ 2 cm,花密生;子房亮淡绿色,扁球形,高约2 mm,直径2.5 ~ 3.0 mm,2室;花柱白色,长0.8 ~ 1.0 mm,直径约0.8 mm;柱头浅黄棕色,下凹,大,高约1.5 mm,直径2.0 ~ 2.5 mm,横截面圆形或卵形,微具疣,全缘,具一向基的浅凹,或浅至明显的2裂,裂片圆形;雄花序区略呈或明显呈倒圆锥形,长1.5 ~ 4.0 cm,基部直径1.5 ~ 2.0 cm,顶部直径1.6 ~ 3.0 cm,花密生;雄花由4或5枚雄蕊组成;雄蕊长3.0 ~ 3.5 mm,宽1 ~ 1.5 mm;花丝长约1.5 mm,合生;花药长1.5 ~ 2.0 mm,截形;孔顶生或近顶生,椭圆形,小,每孔具一沟延伸至花药边缘;药隔大,隆起。附属器圆锥形、卵状椭圆形或狭纺锤形,长4.5 ~ 12.0 cm,粗2.2 ~ 5.0 cm,钝,白色,具多数浅的、常纵向汇合的凹穴,有或无散生的微小点状乳突,基部具不育雄蕊,逐渐融合形成附属器壁。花期5月,果期7月。
分布:国内产广西、云南;国外见于越南。海南为首次记录,产于海南热带雨林国家公园尖峰岭片区,生于山体岩壁间。
凭证标本:海南热带雨林国家公园尖峰岭片区,18.43°N,108.50°E,海拔:800 m,2025年3月10日,XMY20250330(IBSC!)。
-
Engelhardia spicata Lesch. ex Bl., Bijdr. 10: 528. 1825.(图5)——ISOTYPE(图5-C1—C3)

图 5 云南黄杞与海南黄杞
Figure 5. Engelhardia spicata & E. hainanensis
大乔木,高达15 ~ 20 m;小枝后来无毛,有腺体,褐色,具皮孔。叶为偶数或稀奇数羽状复叶,长25 ~ 35 cm,叶柄及叶轴最后变为无毛;小叶4 ~ 7对,对生或几乎互生,具0.5 ~ 1.0 cm长的小叶柄,长成后薄革质,长椭圆形至长椭圆状披针形,长7 ~ 15 cm,宽2 ~ 5 cm,顶端短渐尖,基部阔楔形,全缘,上面无毛而仅散生腺体,下面中脉及小叶柄有疏短柔毛,最后变无毛,侧脉每边10 ~ 13条。雄性葇荑花序常集为圆锥状花序,自叶痕腋内无叶的侧枝上生出。雄花较密集,几乎无柄,苞片3裂,有柔毛,花被片4枚,花药具毛,药隔具1凸头伸出于花药顶端。雌性葇荑花序单独生于侧枝顶端或生于雄性圆锥状花序束的顶端。雌花近于无柄,苞片及小苞片基部有毛,花柱短,柱头2 ~ 4裂。果序长可达30 ~ 45( ~ 60)cm,俯垂;果实球状,直径3.5 mm左右,上部被刚毛,苞片及小苞片基部被有刚毛,贴生至近果实中部;苞片的裂片倒披针状矩圆形,向上端略扩大,顶端钝,中间裂片长2.5 ~ 3.5 cm,宽0.7 ~ 1.0 cm,侧裂片长约1.5 cm。11月开花,1 ~ 2月果成熟。
与海南黄杞E. hainanensis P. Y. Chen相似,但后者叶片革质,边缘具锯齿。
分布:国内分布于贵州、云南、西藏、广东、广西、海南;国外产阿萨姆、孟加拉国、婆罗洲、柬埔寨、印度、爪哇、老挝、小巽他群岛、马来西亚、缅甸、尼泊尔、巴基斯坦、菲律宾、苏门答腊、泰国、越南等地。
凭证标本:海南热带雨林国家公园尖峰岭片区,18.43°N,108.50°E,海拔:800 m,2025年3月10日,XMY20250330(IBSC!)。
-
中国是世界上野生保护植物多样性最为丰富的地区之一,其热带区域主要分布于西藏东南部、云南西南到东南部、广西西南部、广东雷州半岛、台湾南部以及海南岛,这些地区的植物区系主要属于热带亚洲植物区系的北缘类型[29]。其中,海南由于在地史上逐渐南移,形成典型的热带气候,从而孕育出丰富且典型的热带植物区系。该地区热带植物成分比例最高,并以泛热带分布型占据主导地位[30],海南省处于热带北缘,在地质时期自古生代起即与广东陆地相连,直至第四纪初期才与大陆分离[31],因此植物区系组成与广东、广西有不可分割的联系,均呈现出以热带分布为主[32],其独特的热带岛屿属性,使当地的植物多样性研究成为全球生物多样性保护的关键地区,深入探究海南的植物资源,不仅能推动海南省的植物区系研究,也对保护其植物多样性与实现资源可持续利用具有至关重要的意义[33]。近年来,在海南持续开展植物资源调查中,陆续发现并报道了多个新记录物种,如条纹双唇兰(Didymoplexis striata J. J. Sm.)[34],距隔菊(Koyamasia curtisii(Craib & Hutch.)Bunwong, Chantar. & S. C. Keeley)和婴带菊(Struchium sparganophorum(L.)Kuntze)[35],这些新记录进一步丰富了海南植物区系资料,也反映出该地区仍存在大量未被充分认知的植物种类。南岭堇菜、黄斑报春苣苔和东京魔芋的花具有较高的观赏价值,而薄叶山柑树形优美,花量繁茂,是一种优质的乡土树种,可以进一步开发供园林绿化观赏使用;同时,针对薄叶山柑在尖峰岭仅发现单株,南岭堇菜、黄斑报春苣苔和东京魔芋为罕见小种群分布,发现地点均为保护区内,人为干扰较少的区域,但仍然存在濒危风险。文章对海南植物新分布资料予以报道,进一步丰富了海南植物多样性,同时也为海南与广东、广西、云南地区植物区系的紧密联系提供了依据,对推动海南热带雨林国家公园的生物多样性调查、保护和研究等具有重要意义。
-
本次报道的4个新记录种均来自海南热带雨林国家公园尖峰岭片区内,本文提供的资料丰富了海南维管植物多样性。南岭堇菜主要产于广东省南岭山脉,与广东紧邻的香港、澳门、江西、广西、湖南和湖北均有记载[28],目前该种在尖峰岭仅发现一处种群,有大概100株;薄叶山柑在我国仅分布于云南,目前该种在尖峰岭仅发现一株成年大树带2株萌生小树,根据其花量繁茂,预估有更多的种群有待发现;黄斑报春苣苔在原始文献中描述其花期为11月,作者根据此研究在海南尖峰岭的观测为黄斑报春苣苔可在海南全年开花,黄斑报春苣苔在此之前也仅分布于广西,广东华南植物园与仙湖植物园有人工栽培,在海南尖峰岭的3个管护站附近发现3个种群,估计有200株;在尖峰岭仅发现一处集中分布的东京魔芋种群,有25株。在海南植物区系中,仅有7个特有属和约10%的特有种,极低的特有性显示了它具有明显的大陆起源特征,通过比较研究发现中国海南植物区系与越南植物区系关系最密切,其次是广西植物区系,根据中国海南岛可能曾经连接着越南和中国广西的这一理论,在海南可能还会发现更多的分布在云南南部、广西西南部的植物[36],此次的海南省植物新记录发现也进一步证实了这一理论并丰富了海南植物名录的物种组成,也为海南热带雨林国家公园生物多样性评估提供关键基础数据。
此前,本研究团队人员对黄杞、少叶黄杞与双叶黄杞分类地位进行重新评估时,在华南国家植物园标本馆(IBSC)查阅黄杞属的标本时发现有秦新生、张荣京(
14868 !)与邢福武等(14807 !)在海南采集的云南黄杞的标本与陈邦余1981年在植物分类学报中发表的海南黄杞属植物二新种论文中提及“云南黄杞不产海南,原误定为云南黄杞的植物应是一新种海南黄杞”给予纠正,即云南黄杞在海南存在分布,并提供在华南国家植物园标本馆(IBSC)拍摄的云南黄杞与其模式标本,及拍摄海南尖峰岭的海南黄杞的照片进行比对,即云南黄杞与海南黄杞相似,但后者叶片革质,边缘具锯齿是与云南黄杞区分开的关键识别特征。致谢:感谢辰山植物园标本馆(CANT)与华南国家植物园标本馆(IBSC)等标本馆在标本查阅中给予的帮助,感谢辰山植物园李晓晨老师提供南岭堇菜模式标本图片及其在堇菜科植物鉴定中给予的帮助!
New records and literature revision of plant distribution in Hainan Province
-
摘要: 在海南热带雨林国家公园尖峰岭片区调查时,发现了海南省植物新记录4种,分别为:南岭堇菜Viola nanlingensis J. S. Zhou & F. W. Xing、薄叶山柑Capparis tenera Dalz.、黄斑报春苣苔Primulina flavimaculata (W. T. Wang) Mich. Möller & A. Weber、东京魔芋Amorphophallus tonkinensis Engl. & Gehrm.。每种均给出实地拍摄的照片并进行了物种形态特征描述,补充了薄叶山柑在原始文献和中国植物志中果、种子的空白。凭证标本保存于上海辰山植物标本馆(CSH)与华南国家植物园标本馆(IBSC)。同时,对陈邦余1981年在植物分类学报中发表的海南黄杞属Engelhardia植物二新种论文中提及“云南黄杞Engelhardia spicata Lesch. ex Blume不产海南,原误定为云南黄杞的植物应是一新种海南黄杞Engelhardia hainanensis P. Y. Chen”给予纠正,即云南黄杞在海南存在分布,凭证标本存于华南植物园标本馆(IBSC)。Abstract: During the field survey in Jianfengling district of Hainan Tropical Rainforest National Park, four plant species newly recorded in Hainan Province were reported, including: Viola nanlingensis J. S. Zhou & F. W. Xing, Capparis tenera Dalz and Primulina flavimaculata (W. T. Wang) Mich. Möller & A. Weber, Amorphophallus tonkinensis Engl. & Gehrm. For each species, field photographs were provided along with descriptions of their morphological characteristics. Additionally, the gaps in the original literature and the Flora of China regarding the fruit and seed descriptions of Capparis tenera were supplemented. Voucher specimens are deposited in the Shanghai Chenshan National Botanical Garden Herbarium (CSH) and the South China Botanical Garden Herbarium (IBSC). Furthermore, a correction was made regarding the 1981 publication by Chen Bangyu in the Acta Phytotaxonomica Sinica, which proposed two new species of Engelhardia from Hainan and stated that "Engelhardia spicata does not occur in Hainan, and that the plants previously misidentified as Engelhardia spicata should be recognized as a new species, Engelhardia hainanensis." It has now been confirmed that Engelhardia spicata does indeed exist in Hainan, with voucher specimens deposited in the South China Botanical Garden Herbarium (IBSC).
-
Key words:
- new records /
- Violaceae /
- Capparaceae /
- Gesneriaceae /
- Araceae /
- Juglandaceae
-
[1] Linnaeus C. Species plantarum [M]. Stockholm: Impensis Laurentii Salvii, 1753: 933−937. [2] Batsch A J G K. Tabula affinitatum regni vegetabilis [M]. 2nd ed. Weimar: Landes-Industrie-Comptoir, 1802: 57. (查阅网上资料, 未找到对应的出版地信息, 请确认) [3] Clausen J. Cytotaxonomy and distributional ecology of western North American violets [J]. Madroñ o, 1964, 17(6): 173−197. [4] Ballard H E Jr. Phylogenetic relationships and infrageneric groups in Viola (Violaceae) based on morphology, chromosome numbers, natural hybridization and Internal Transcribed Spacer (ITS) sequences [D]. Madison: University of Wisconsin, 1996. [5] Ballard H E Jr, Sytsma K J, Kowal R R. Shrinking the violets: phylogenetic relationships of infrageneric groups in Viola (Violaceae) based on Internal Transcribed Spacer DNA sequences [J]. Systematic Botany, 1999, 23(4): 439−458. https://doi.org/10.2307/2419376 doi: 10.2307/2419376 [6] Engler A. Die natürlichen pflanzenfamilien [M]. 2nd ed. Leipzig: Engelmann, 1925: 363−376. [7] 中国科学院中国植物志编辑委员会. 中国植物志: 第51卷 被子植物门 双子叶植物纲 堇菜科[M]. 北京: 科学出版, 1991: 8-129. [8] Linnaeus C. Species plantarum [M]. Stockholm: Impensis Laurentii Salvii, 1753: 1, 503. [9] de Jussieu A L. Genera plantarum [M]. Paris: Apud viduam Herissant, 1789: 242−243. [10] Plants of the World Online. Plants of the world online [EB/OL]. [2025-10-27]. https://powo.science.kew.org/. [11] Souvannakhoummane K, Fici S, Lanorsavanh S, et al. Capparis macrantha sp. nov. (Capparaceae, Brassicales), a new shrub species from a deciduous forest of the Nam Kading National Protected Area (central Lao PDR) [J]. European Journal of Taxonomy, 2020, 656: 1−12. https://doi.org/10.5852/ejt.2020.656 doi: 10.5852/ejt.2020.656 [12] 中国科学院中国植物编辑委员会. 中国植物志-第三十二卷-被子植物门 双子叶植物纲 罂粟科 山柑科[M]. 北京: 科学出版, 1999: 500. [13] 杨文光. 广义报春苣苔属(苦苣苔科)系统学研究[D]. 郑州: 河南农业大学, 2016. [14] Wang Y Z, Mao R B, Liu Y, et al. Phylogenetic reconstruction of Chirita and allies (Gesneriaceae) with taxonomic treatments [J]. Journal of Systematics and Evolution, 2011, 49(1): 50−64. https://doi.org/10.1111/j.1759-6831.2010.00113.x doi: 10.1111/j.1759-6831.2010.00113.x [15] Weber A, Middleton D J, Forrest A, et al. Molecular systematics and remodeling of Chirita and associated genera (Gesneriaceae) [J]. Taxon, 2011, 60(3): 767−790. https://doi.org/10.1002/tax.603012 doi: 10.1002/tax.603012 [16] 中国科学院中国植物志编辑委员会. 中国植物志: 第69卷[M]. 北京: 科学出版社, 1990: 333−352. [17] Scholten J. An updated identification guide to the species of Amorphophallus (Araceae): new synonyms and a set of global dichotomous keys [J]. Blumea-Biodiversity, Evolution and Biogeography of Plants, 2023, 68(2): 139−161. https://doi.org/10.3767/blumea.2023.68.02.03 doi: 10.3767/blumea.2023.68.02.03 [18] Croat T B, Boyce P. Überlist - April 2023 [EB/OL]. [2023-06-10]. https://www.researchgate.net/publication/369741356_Uberlist_-_April_2023. [19] Blume C L. Bijdragen tot de flora van Nederlandsch Indië [M]. Batavia: Lands Drukkerij, 1826: 528. [20] 路安民. 论胡桃科植物的地理分布[J]. 植物分类学报, 1982, 20(3): 257−274. (查阅网上资料, 未找到本条文献信息, 请确认) [21] 中国科学院中国植物志编辑委员会. 中国植物志: 第21卷[M]. 北京: 科学出版社, 1979: 11−18. [22] 广东省植物研究所. 海南植物志: 第三卷[M]. 北京: 科学出版社, 1974: 114−115. [23] 陈邦余. 海南黄杞属植物二新种[J]. 植物分类学报, 1981, 19(2): 250−251. (查阅网上资料, 未找到本条文献信息, 请确认) [24] 李意德, 许涵, 骆土寿, 等. 中国生态系统定位观测与研究数据集: 森林生态系统卷: 海南尖峰岭站(生物物种数据集)[M]. 北京: 中国农业出版社, 2012: 10−35. [25] 许涵, 李意德, 骆土寿, 等. 海南尖峰岭不同热带雨林类型与物种多样性变化关联的环境因子[J]. 植物生态学报, 2013, 37(1): 26−36. https://doi.org/10.3724/SP.J.1258.2013.00003 doi: 10.3724/SP.J.1258.2013.00003 [26] 杨小波, 陈玉凯, 李东海, 等. 海南植物名录[M]. 北京: 科学出版社, 2013: 65−278. (查阅网上资料, 未找到对应的标黄作者信息, 请确认) [27] 邢福武, 陈红锋, 秦新生, 等. 中国热带雨林地区植物图鉴—海南植物(1−3卷)[M]. 武汉: 华中科技大学出版社, 2014. (查阅网上资料, 未找到对应的页码信息, 请确认) [28] 李沛琼. 深圳植物志: 第2卷[M]. 北京: 中国林业出版社, 2010: 64. [29] 朱华. 中国南部热带植物区系[J]. 生物多样性, 2017, 25(2): 204−217. https://doi.org/10.17520/biods.2016055 doi: 10.17520/biods.2016055 [30] Zhu H. Biogeographical evidences help revealing the origin of Hainan Island [J]. PLoS One, 2016, 11(4): e0151941. https://doi.org/10.1371/journal.pone.0151941 doi: 10.1371/journal.pone.0151941 [31] 张宏达. 海南植物区系的多样性[J]. 生态科学, 2001, 20(1/2): 1−10. https://doi.org/10.3969/j.issn.1008-8873.2001.01.001 doi: 10.3969/j.issn.1008-8873.2001.01.001 [32] 林泽钦, 杨小波, 陈玉凯, 等. 海南本地野生维管植物区系研究[J]. 热带作物学报, 2016, 37(2): 351−358. https://doi.org/10.3969/j.issn.1000-2561.2016.02.022 doi: 10.3969/j.issn.1000-2561.2016.02.022 [33] 梁彩群, 陈玉凯, 杨小波, 等. 海南省野生维管植物编目和分布数据集[J]. 生物多样性, 2023, 31(6): 23067. https://doi.org/10.17520/biods.2023067 doi: 10.17520/biods.2023067 [34] 张培春, 杨小波, 陈琳, 等. 条纹双唇兰, 中国兰科双唇兰属一新记录种(英文)[J]. 热带亚热带植物学报, 2022, 30(1): 132−134. https://doi.org/10.11926/jtsb.4445 doi: 10.11926/jtsb.4445 [35] 陈又生, 王清隆, 廖俊杰, 等. 中国菊科二新记录属(英文)[J]. 热带亚热带植物学报, 2022, 30(2): 257−266. https://doi.org/10.11926/jtsb.4453 doi: 10.11926/jtsb.4453 [36] 朱华. 论中国海南岛的生物地理起源[J]. 植物科学学报, 2020, 38(6): 839−843. https://doi.org/10.11913/PSJ.2095-0837.2020.60839 doi: 10.11913/PSJ.2095-0837.2020.60839 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 306
- HTML全文浏览量: 212
- 被引次数: 0
