

 下载:
下载:
-
红树林湿地广泛分布于全球热带与亚热带海岸带,是全球蓝碳产量最高的生态系统之一,其高效的碳固存能力在减缓气候变化过程中发挥关键作用[1-2]。然而,在气候变化和海岸带开发的双重压力下,红树林湿地受到威胁,严重损害了红树林的生态功能,据统计,1980–2005年间,全球红树林面积减少了近20%,其中约一半的损失归因于养殖围塘等人为开垦活动[3]。近年来,“退塘还林”已成为恢复红树林生态系统的核心措施,其碳汇功能的恢复潜力,很大程度上取决于土壤有机碳库的组分分配格局与结构稳定性[4]。因此,研究退塘种植红树林群落对土壤有机碳(SOC)组分的分布特征及理化性质的影响,对有效提升红树林湿地碳汇功能具有重要的意义。
SOC并不是一个单一均匀的实体,关键组成部分包括颗粒有机碳(POC)、矿物结合有机碳(MAOC)、可溶解有机碳(DOC)和微生物生物量碳(MBC),分别承担着短期碳活性调节、长期碳稳定固存、碳迁移载体和碳转化驱动的核心功能[5]。它们的动态特征是反映退塘还林早期生态系统恢复过程的敏感指标[6]。Cui等[7]的研究发现,红树林SOC随恢复年限增加而升高,但活性有机碳在5年生群落达到峰值后下降,表明恢复早期是活性碳快速响应和重组的阶段。李玲云等[6]进一步发现,EOC、MBC和DOC等活性组分在种植岛中均高于水道和裸岛,且土壤容重、TN和TP是其垂直分布的重要影响因子,说明退塘还林初期土壤碳库整体处于较活跃调整期。除恢复年限外,群落组成和土壤理化环境也是影响土壤碳组分形成与稳定的重要因素。Chen等[8]指出,不同树种间SOC储量差异并不显著,但SOC来源及POC、MAOC组分存在显著差异;其中,MAOC与土壤含水量、盐度呈正相关,与pH呈负相关,表明矿物条件和理化环境共同调控稳定碳的形成。在群落配置方面,现有研究已开始关注多树种配置对碳汇恢复的潜在优势。Chowdhury等[9]的研究发现,4年生混交红树林恢复地的蓝碳库高于周边自然群落与退化泥滩,并据此建议采用混交种植促进退化盐滩生态恢复。Li等[10]的荟萃分析表明,混交恢复红树林较单一树种林更有利于碳储量提升,且这种由物种互补带来的促进作用在恢复7~20年后更为明显且趋于稳定。陈顺洋等[11]的综述也指出,生境条件、物种选择和种植措施是决定红树林生态修复固碳效果的主要因素,但对不同群落配置下土壤碳组分及其理化驱动机制的定量认识仍然不足。因此,系统分析5种退塘红树林群落对SOC组分的特征及理化性质的影响,对准确评价红树林碳汇恢复潜力并优化恢复技术方案具有重要意义。红树林土壤受自身根系和落叶的生物碳输入影响[12]。由于幼龄红树林在生物量生产和化学组成上与成熟林存在显著差异,其SOC输入,尤其是POC可快速响应植被输入变化,是修复早期碳库活性的核心指示指标,有机碳组分的输入也会随红树林生长和生态系统发育而发生变化[13-14]。红树林种植后的早期阶段通常是土壤碳库变化最剧烈且活性最高的时期;因此,识别该阶段土壤碳库特征及其控制因素,对于系统理解红树林演替过程中的碳库演变至关重要[15]。
尽管已有大量研究关注成熟红树林的SOC动态[16-17],但目前针对由养殖池塘转化为红树林的恢复初期阶段,对SOC含量、碳组分变化及其驱动因素的研究仍相对不足。本研究以海南琼海沙美内海退塘还林的红树林群落样地(海桑、木榄+海桑、白骨壤+海桑、海桑+桐花树以及木榄+白骨壤)为研究对象,并以未种植区域光滩为对照,探究塘内不同红树林种植区域土壤碳库特征,比较分析了不同红树林植被群落SOC组分的分布特征及其影响因素,旨在为评估和优化退塘还林生态工程的固碳效益提供科学依据和理论参考。
-
研究区域位于海南岛琼海市博鳌镇的沙美内海(约19°2′17″ N~19°7′53″ N, 110°33′2″ E~110°34′21″ E,图1)。该内海是由玉带滩沙坝屏蔽外海所形成的潟湖,呈狭长的南北走向。潟湖水浅坡缓,其南侧有龙滚河注入,西侧有九曲江注入,北侧经万泉河口与南海连通。区域气候属热带季风气候,四季不明显,旱季和雨季分明,年均气温24℃,1月气温最低,平均18 °C;7月气温最高,平均28.3 °C。降水量丰富,年均降水量约
2072 mm。降水主要集中在夏秋季节,尤其是9月份降水最多,秋季三个月的降水量占全年降水量的50%以上,并且受台风影响频繁。该区域的潮汐性质属不正规半日潮和不正规日潮混合潮,多年平均高潮位为1.85 m,多年平均低潮位为−0.37 m,最高高潮位为2.03 m(1983年7月17日)。样地集中分布在中潮位区域,沙美内海主要分布潮沙泥土,土壤酸性较强,适合红树林等湿地植物的生长。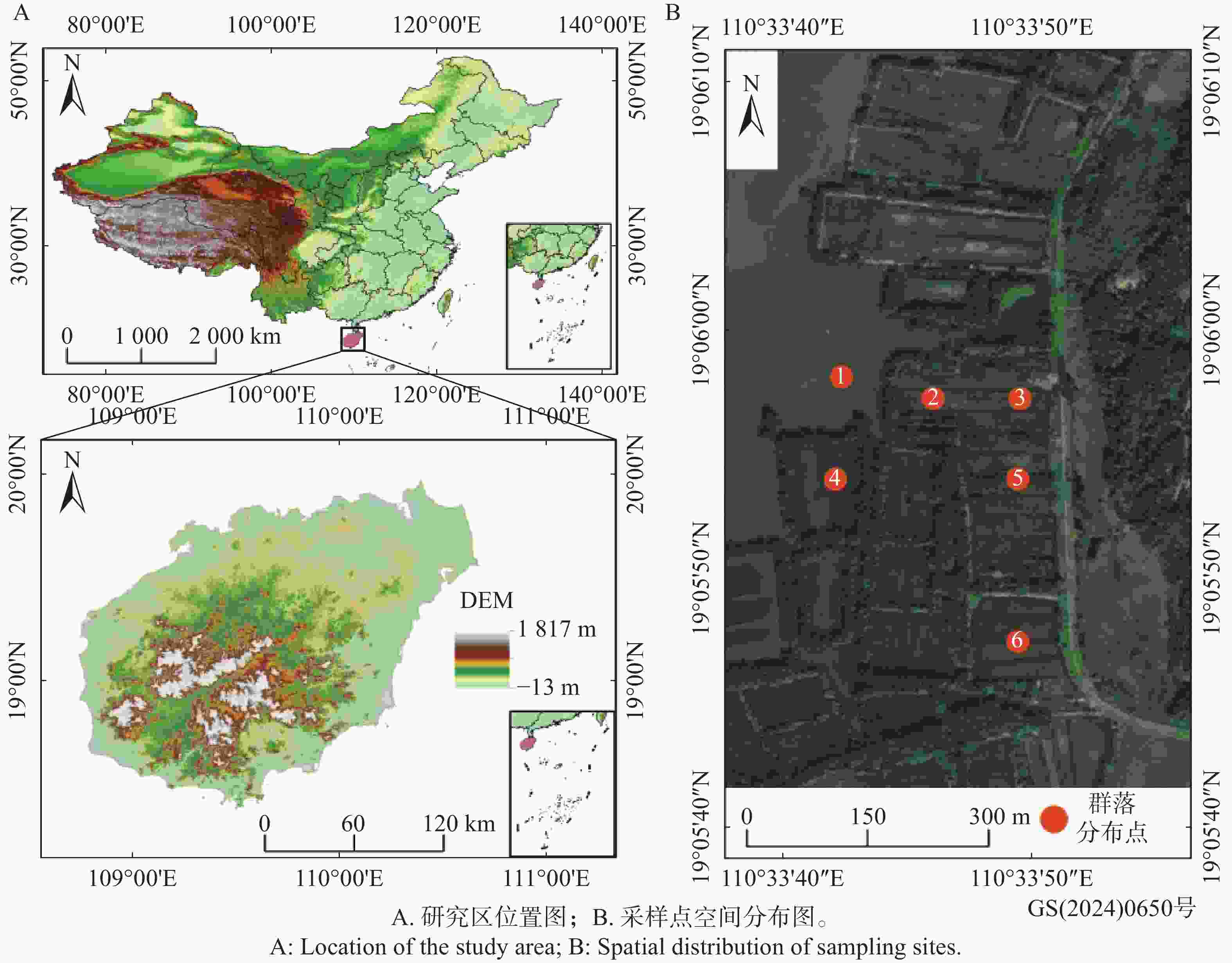
图 1 研究区位置及采样点空间分布
Figure 1. Location of the study area and spatial distribution of sampling sites
-
本研究在海桑、木榄+海桑、白骨壤+海桑、海桑+桐花树和木榄+白骨壤等5个不同红树林群落中各选4个5×5 m典型样地,统计各个样方内的植物树高、基径和密度,所有样地林龄均为4年,所设光滩对照样地位于研究区附近,与恢复样地潮位条件相近,且均属于退塘后未种植红树林的区域。在每个样地内采集4个土样,采样时先去除土壤地表的凋落物,然后用内直径为10 cm的土钻采集土柱,考虑到退塘还林4年仍处于恢复初期,新增有机碳主要来源于地表凋落物输入、浅层根系周转及表层微生物过程,因此本研究重点采集0~20 cm土层,并划分为表层(0~10 cm)和深层(10~20 cm)两层,以表征恢复初期表层SOC组分的垂向分布特征。把采集好的样品用贴有标签纸的自封袋收集好并放入装有冰袋的保温箱内,带回实验室备用。于实验室内剔除碎石、植物残根等杂物后,将一部分鲜土样品于4℃保存,用于土壤MBC和DOC的测定;一部分土壤样品风干处理,用于测定SOC、POC、MAOC及理化性质;另一部分土壤样品将其置于105℃烘箱,以测定其容重。土壤pH采用电极法测定(水土质量比为2.5:1),土壤电导率(EC)采用电导率法测定(水土质量比为5:1),土壤容重(BD)采用环刀法测定;土壤SOC采用重铬酸钾氧化法;土壤POC和MAOC采用53 µm粒径分级湿筛法进行分离。称取10 g过2 mm筛的风干土样,加入30 mL 5 g·L−1六偏磷酸钠溶液,于25℃、90 r·min−1条件下振荡18 h后过53 µm筛网,并以蒸馏水冲洗分离。滞留于筛上的>53 µm组分为POC,过筛的<53 µm组分为MAOC。各组分经烘干、研磨后,采用重铬酸钾外加热法测定有机碳含量。土壤全氮(TN)采用凯氏定氮仪测定;土壤全磷(TP)采用酸熔-钼锑抗比色法测定;土壤MBC采用氯仿熏蒸K2SO4浸提法测定[18]。土壤可溶性有机碳(DOC)采用0.5 mol·L−1 K2SO4浸提法测定。称取8.0 g新鲜土样,按土液比1:5加入40 mL 0.5 mol·L−1 K2SO4溶液,在25℃、250 r·min−1条件下振荡1 h后,以
15000 r·min−1离心10 min,取上清液经0.45 μm微孔滤膜过滤,并设置空白对照,滤液中的有机碳浓度采用TOC分析仪测定。 -
土壤有机碳储量(C-stock,kg·C·m−2),计算公式[19]:
$$ C-stock=SOC\times BD\times H\times 0.01 $$ 式中:SOC为土壤有机碳含量(g·kg−1);BD为土壤容重(g·cm−3);H为土层厚度(cm);0.01为单位转换系数(将g·cm−2转换为kg·m−2)。
本研究采用“碳储量差值-修复年限法”估算SOC积累速率,即根据各红树林样地与对照光滩样地之间的土壤碳储量差值,并结合修复年限,表征不同恢复群落表层SOC的相对积累水平。对照样地为研究区附近潮位条件相近的退塘后未种植红树林区域。
有机碳积累速率(SR),计算公式:
$$ \mathrm{SR}=\frac{\mathrm{C}-{\mathrm{C}}_{0}}{\mathrm{T}} $$ 式中:C为各红树林群落土层碳储量(kg·C·m−2),C0为光滩土层总碳储量(kg·C·m−2);T为退塘还林修复年限(年),本研究中T=4。
应用单因素方差分析法(One-way ANOVA)比较不同样地及不同深度的SOC、EOC、MBC和DOC,MAOC以及活性有机碳各组分占比的差异性,采用Tukey HSD检验进行显著性多重比较。所有数值在统计前先进行正态分布和方差齐性检验,并采用Origin 2021软件进行绘图。采样点空间分布图,采用ArcGIS v10.8绘制。相关性分析及Mantel检验均采用R语言完成,通过Pearson相关系数评估土壤理化因子与有机碳组分间的线性相关性。在此基础上,基于linkET包进行Mantel检验探讨土壤因子对不同植被群落碳库特征的影响。
-
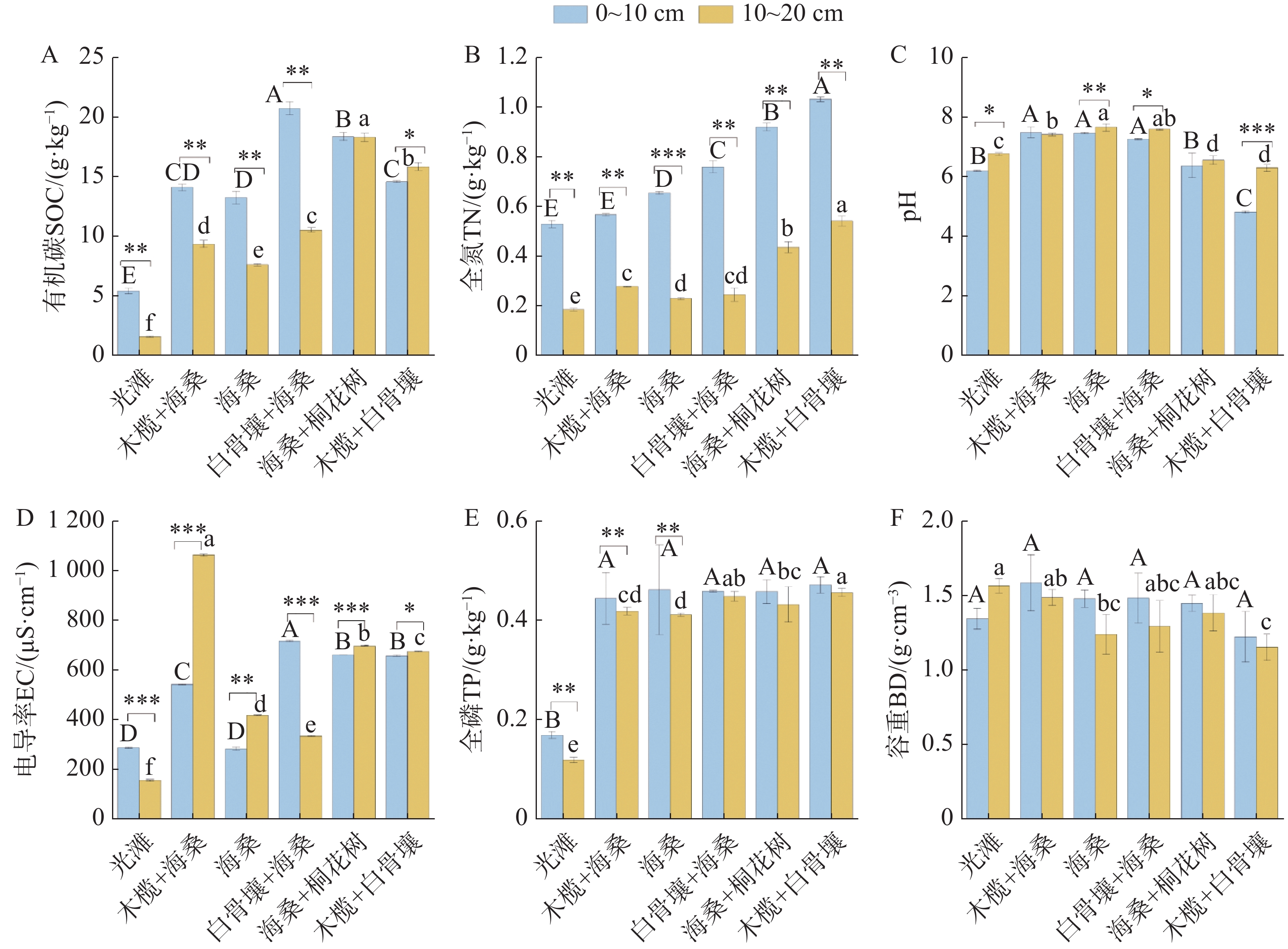
退塘还林初期不同红树林群落及光滩对照样地土壤理化特征如图2所示。在0~20 cm土层,SOC含量介于1.55~20.73 g·kg−1,光滩显著低于红树林群落,白骨壤+海桑群落0~10 cm土层最高,海桑+桐花树在10~20 cm土层维持较高水平;除木榄+白骨壤群落10~20 cm土层略高于0~10 cm外,其余群落均为0~10 cm土层更高,海桑+桐花树群落土层差异极小。TN含量介于0.18~1.03 g·kg−1,红树林群落显著高于光滩,木榄+白骨壤群落最高;所有样地0~10 cm土层均高于10~20 cm土层。土壤pH值介于4.82~7.59,木榄+白骨壤群落最低,多数红树林群落高于光滩,不同群落间差异显著(P<0.05)。土壤EC值介于154.93~
1062.67 μS·cm−1,群落间及土层间差异均显著,木榄+海桑群落10~20 cm土层最高,光滩10~20 cm土层最低;光滩、白骨壤+海桑群落0~10 cm土层高于10~20 cm,其余红树林群落则相反。土壤TP含量介于0.12~0.47 g·kg−1,光滩显著低于各红树林群落,红树林群落间差异较小,多个群落样地两土层间差异不明显。BD介于1.15~1.58 g·cm−3,随土层加深而增大,10~20 cm土层显著高于0~10 cm土层,除光滩外;不同群落间亦存在显著差异(P<0.05)。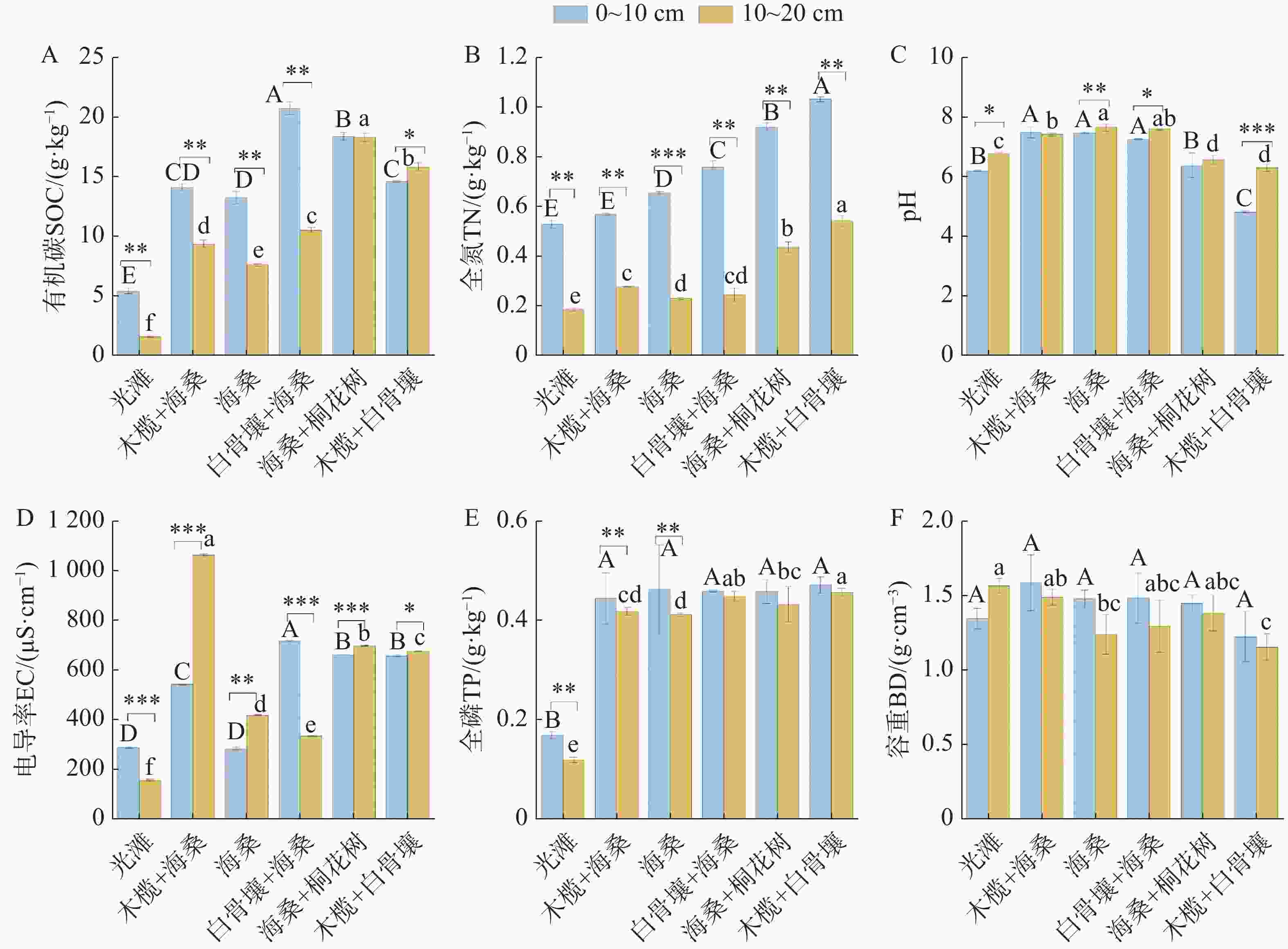
图 2 不同红树林群落土壤理化因子
Figure 2. Soil physicochemical properties of different mangrove communities
-
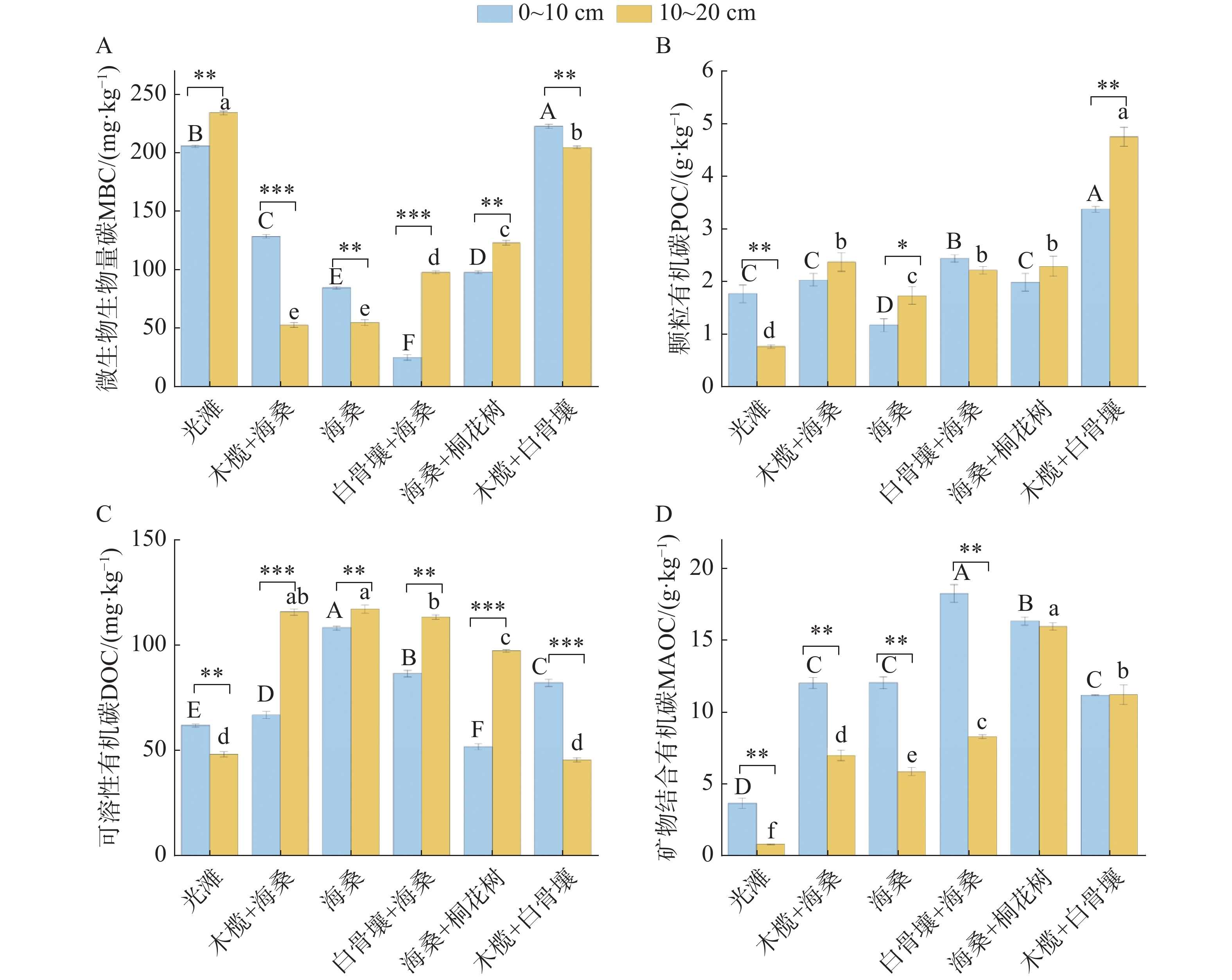
退塘还林初期不同红树林群落及光滩对照样地土壤SOC组分分布特征如图3所示。在0~20 cm土层,MBC含量介于24.57~233.82 mg·kg−1,不同群落样地间差异显著,光滩、木榄+白骨壤群落较高,白骨壤+海桑群落0~10 cm土层最低,木榄+海桑群落10~20 cm土层最低;土层间变化因样地而异,光滩、白骨壤+海桑、海桑+桐花树群落10~20 cm土层MBC含量高于0~10 cm,木榄+海桑、海桑、木榄+白骨壤群落则表现为0~10 cm土层高于10~20 cm。土壤DOC含量介于45.57~117.31 mg·kg−1,样地间差异明显,海桑群落最高,木榄+白骨壤群落及光滩10~20 cm土层最低;土层间变化上,除光滩、木榄+白骨壤群落0~10 cm土层DOC含量高于10~20 cm外,其余红树林群落均表现为10~20 cm土层高于0~10 cm。POC含量介于0.76~4.75 g·kg−1,木榄+白骨壤群落最高,光滩样地最低;土层间变化呈现明显分异,光滩呈现显著的0~10 cm表层富集特征,白骨壤+海桑土层间差异不显著,而木榄+海桑、海桑、海桑+桐花树、木榄+白骨壤等其余红树林群落则表现为10~20 cm土层高于0~10 cm。MAOC含量介于0.79~18.30 g·kg−1,样地间差异显著,红树林群落均显著高于光滩样地,其中白骨壤+海桑在表层含量最高,海桑+桐花树在深层含量最高;土层间变化上,光滩、木榄+海桑、海桑、白骨壤+海桑群落0~10 cm土层MAOC含量高于10~20 cm,海桑+桐花树、木榄+白骨壤群落的两土层间MAOC含量无显著差异。
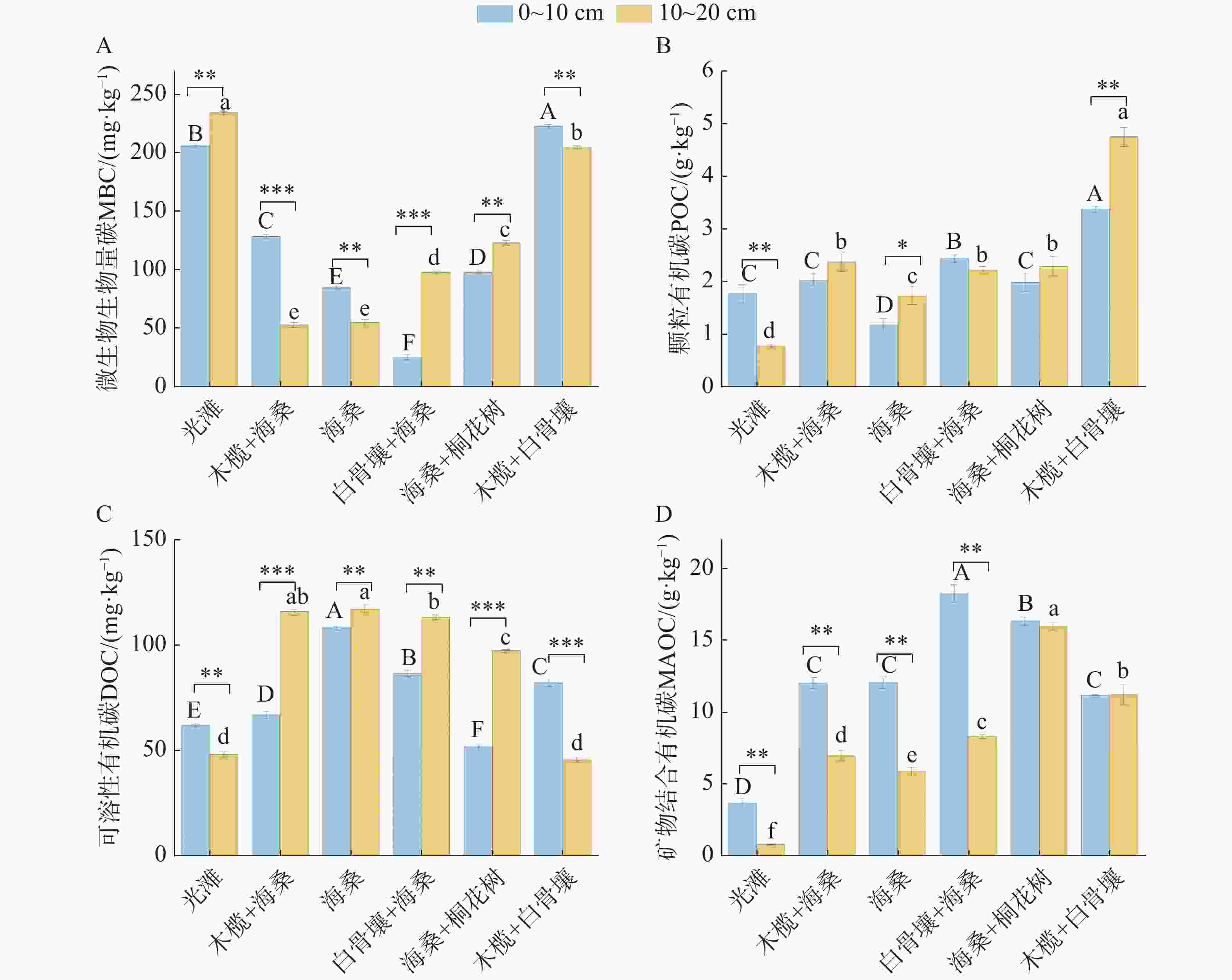
图 3 不同类型样地土壤碳组分特征
Figure 3. Characteristics of soil carbon components across different types of sampling plots
-
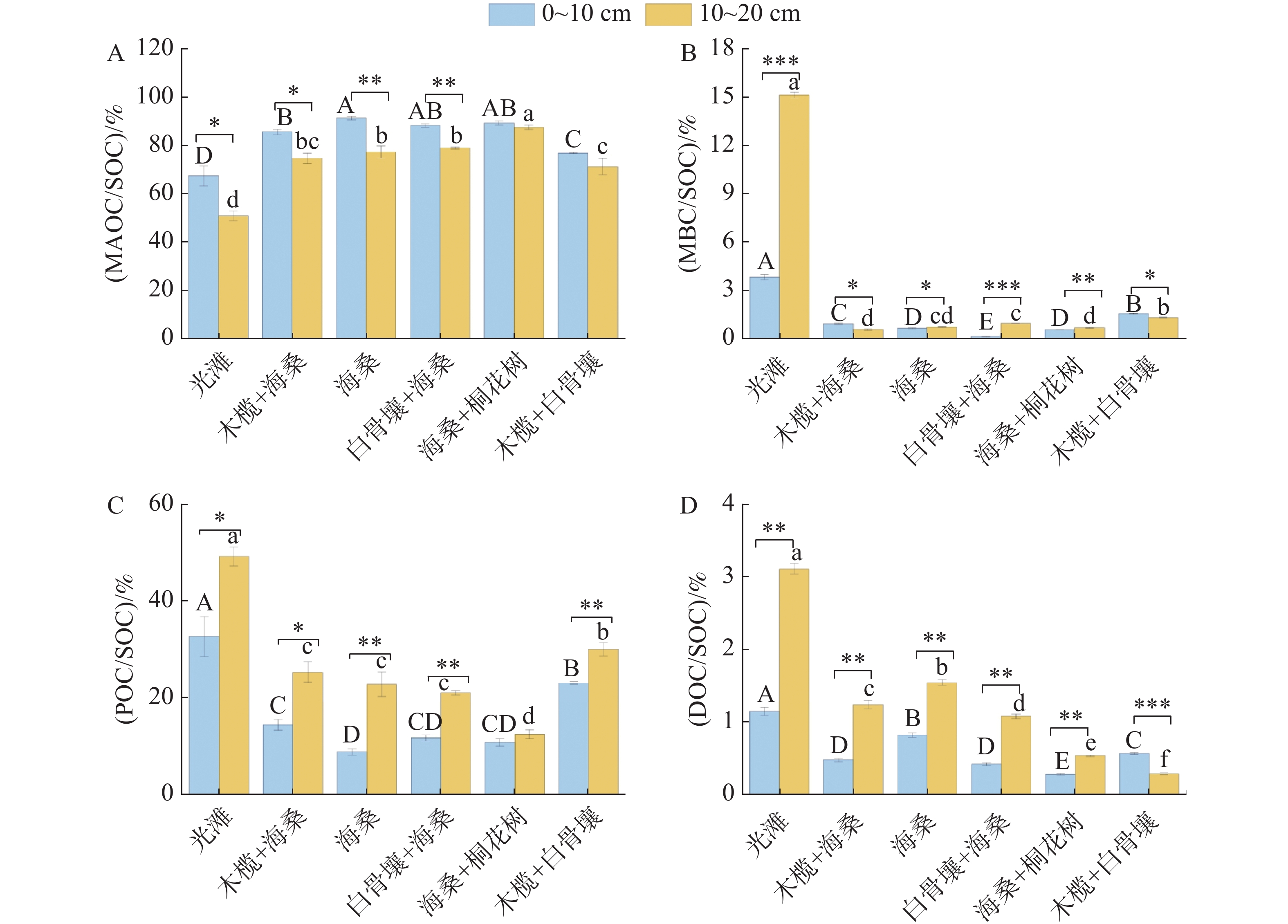
不同红树林群落和土层深度下,SOC各组分相对贡献存在显著差异(图4)。MAOC在各群落与土层中的相对贡献为50.75%~91.19%,为SOC各组分中占比最高的部分,且0~10 cm土层贡献率显著高于10~20 cm土层(P<0.05)。在0~10 cm中,红树林群落区整体高于光滩,最高值为海桑,其中海桑、白骨壤+海桑及海桑+桐花树处理显著高于木榄+白骨壤和光滩处理(P<0.05)。10~20 cm土层比例整体降低,最高值为海桑+桐花树群落,各红树林群落区均显著高于光滩对照区,且各群落间差异显著(P<0.05)。
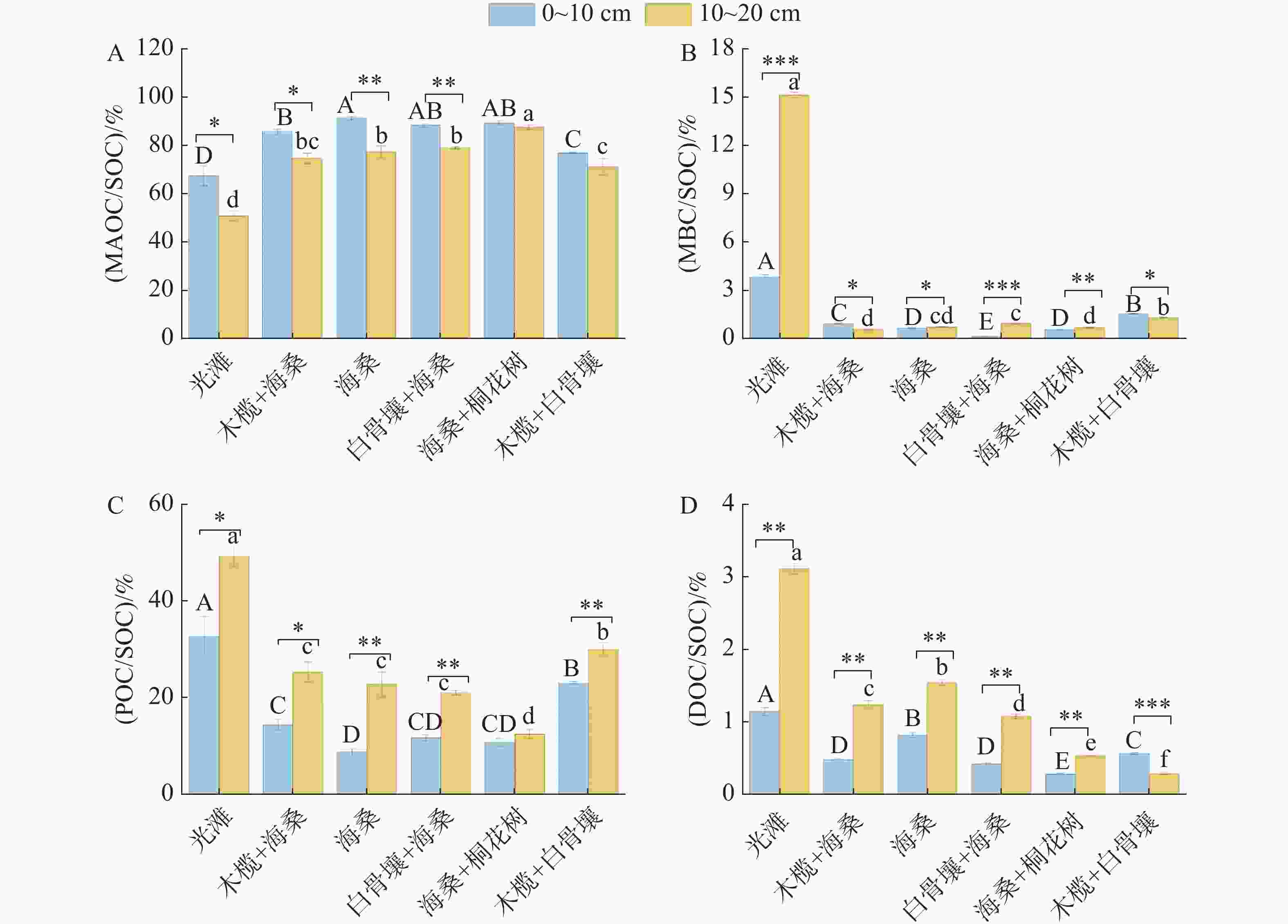
图 4 不同类型样地土壤碳组分比例占比
Figure 4. Proportional composition of soil carbon components across different types of sampling plots
MBC在SOC中的相对贡献整体较低(15.11%±0.19%~0.12%±0.01%),但不同处理和土层间差异显著(P<0.05)。光滩显著高于所有红树林群落区,且表现为10~20 cm土层高于0~10 cm土层。0~10 cm层中,最低值为白骨壤+海桑。10~20 cm土层中,最低值为木榄+海桑,且处理间差异显著(P<0.05)。
POC对SOC的相对贡献在不同处理及土层间差异显著(8.81%±0.66%~49.25%±1.95%)(P<0.05)。除海桑+桐花树外,其余处理均表现为10~20 cm土层比例显著高于0~10 cm土层,呈下层富集特征。光滩始终最高,显著高于所有红树林群落区。0~10 cm土层中,最低值为海桑纯林处理。10~20 cm土层中,最低值为海桑+桐花树,不同红树林群落间分组明显且差异显著(P<0.05)。
DOC在SOC中的占比整体较低(0.28%±0.01%~3.11%±0.07%),但不同群落及土层间差异显著(P<0.05)。光滩土壤DOC在SOC中的占比显著高于所有红树林群落区。除木榄+白骨壤外,其余群落均表现为10~20 cm土层比例显著高于0~10 cm土层,其中光滩和海桑下层土壤比例显著高于其他红树林群落区。0~10 cm土层中,最低值为海桑+桐花树。10~20 cm土层中,最低值为木榄+白骨壤;混交林处理在两个土层中比例均处于较低水平,且在同一土层内,不同群落间差异显著(P<0.05)。
-
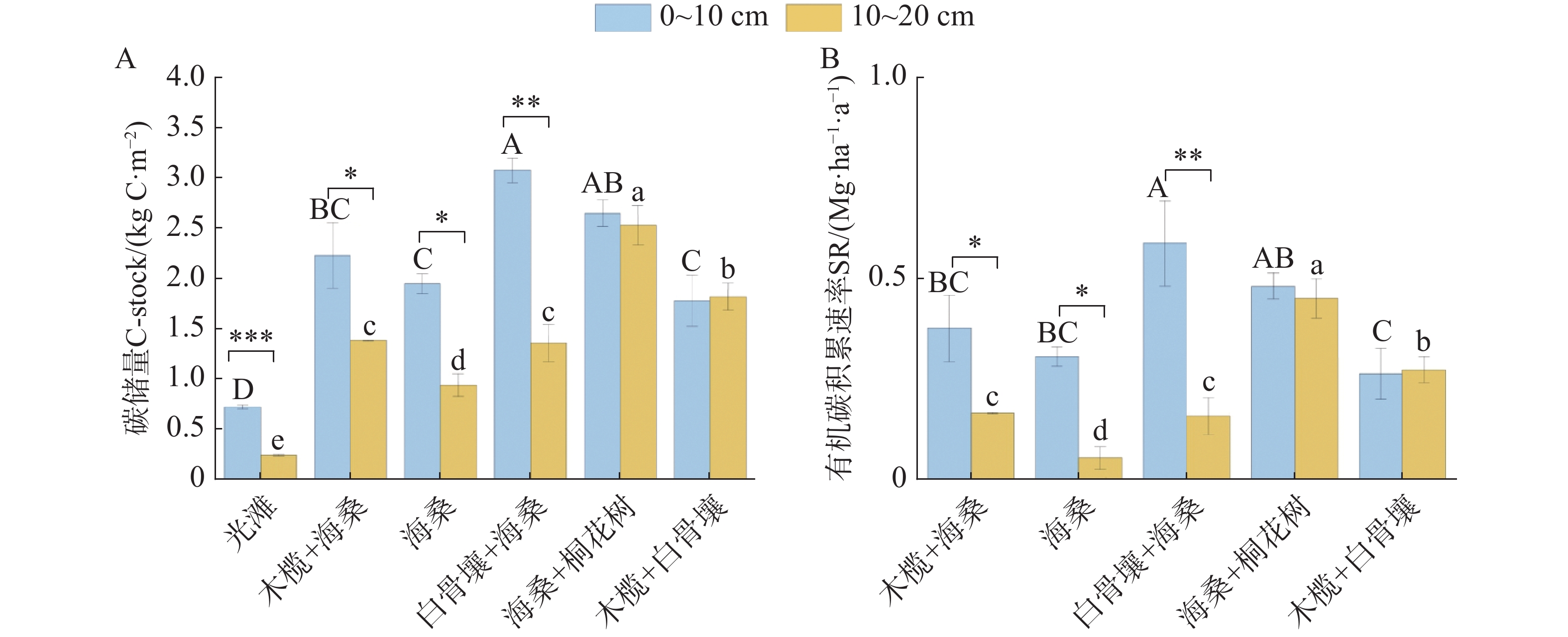
不同植被恢复类型下,SOC储量(C-stock)和有机碳积累速率在0~10 cm与10~20 cm土层中均表现出显著差异(图5)。总体上,红树林群落SOC储量介于1.78 ~ 3.08 kg·C·m−2,多数植被类型在0~10 cm土层中的SOC储量和有机碳积累速率均高于10~20 cm土层。0~10 cm土层中,SOC储量在不同植被恢复类型间差异显著(P<0.05)。白骨壤+海桑的SOC储量最高,其次是海桑+桐花树和木榄+海桑,而光滩和海桑的SOC储量相对较低。在10~20 cm土层中,各植被恢复类型的SOC储量整体低于表层土壤,其中木榄+白骨壤和海桑+桐花树在深层仍维持了较高的碳储量。在有机碳积累速率方面(图5B),不同植被恢复类型在两个土层中均呈现显著差异(P<0.05)。0~10 cm土层中,白骨壤+海桑的有机碳积累速率最高,达到0.59 Mg·ha−1·a−1;海桑+桐花树次之达到0.45 Mg·ha−1·a−1;海桑相对较低。10~20 cm土层中,各群落的有机碳积累速率普遍低于表层,呈现出明显的表层富集特征,其中海桑+桐花树和木榄+白骨壤在深层仍维持了较高的积累速率。总体来看,不同植被恢复类型对SOC储量和有机碳积累速率均具有显著影响,且这种差异在0~10 cm中表现得更为明显。单因素方差分析结果表明,0~20 cm SOC储量在不同群落间存在显著差异(P<0.001)。光滩0~20 cm SOC储量最低0.97±0.02 kg·C·m−2,显著低于所有红树林群落(P<0.05)。在红树林群落中,海桑+桐花树和白骨壤+海桑的0~20 cm SOC储量较高,分别为5.18±0.23和4.44±0.22 kg·C·m−2。0~20 cm SOC积累速率在不同群落间存在显著差异(P<0.001)。各红树林群落的0~20 cm碳积累速率介于0.48~1.05 Mg·ha−1·a−1。Tukey多重比较结果显示,海桑+桐花树处理的0~20 cm有机碳积累速率显著高于海桑纯林处理(P<0.05),与白骨壤+海桑处理差异不显著(P>0.05)。木榄+海桑和木榄+白骨壤处理的0~20 cm有机碳积累速率与海桑纯林处理差异不显著(P>0.05)。
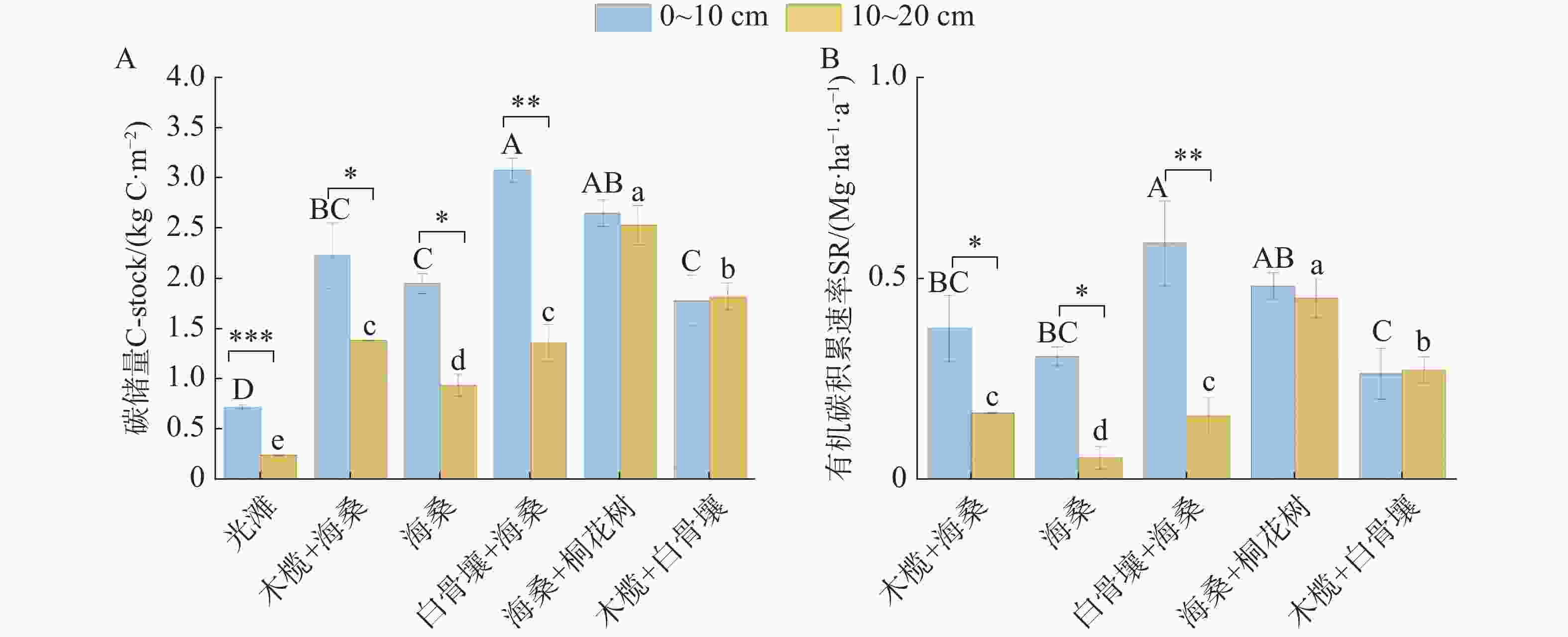
图 5 不同红树林群落SOC储量及有机碳积累速率
Figure 5. Soil organic carbon storage and carbon accumulation rates in different mangrove communities
-
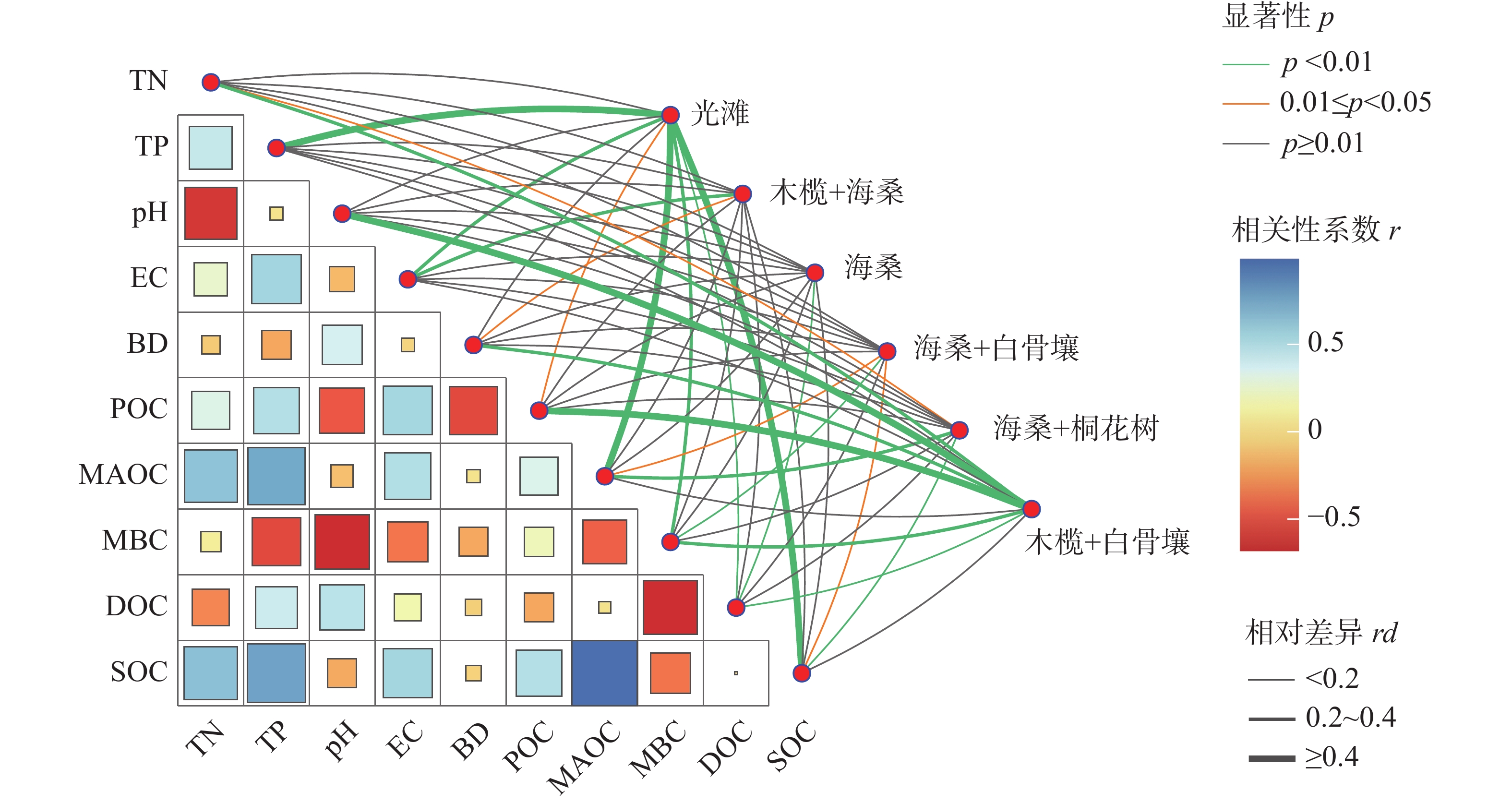
相关性分析结果(图6)表明,SOC与TN、TP和EC均呈显著正相关,与pH呈显著负相关。其中,MAOC与TN、TP和EC均呈显著正相关;POC与TN、TP和EC呈显著正相关,而与pH和BD呈显著负相关;DOC与TN呈显著负相关,而与pH呈显著正相关;MBC与pH和EC均呈显著负相关。
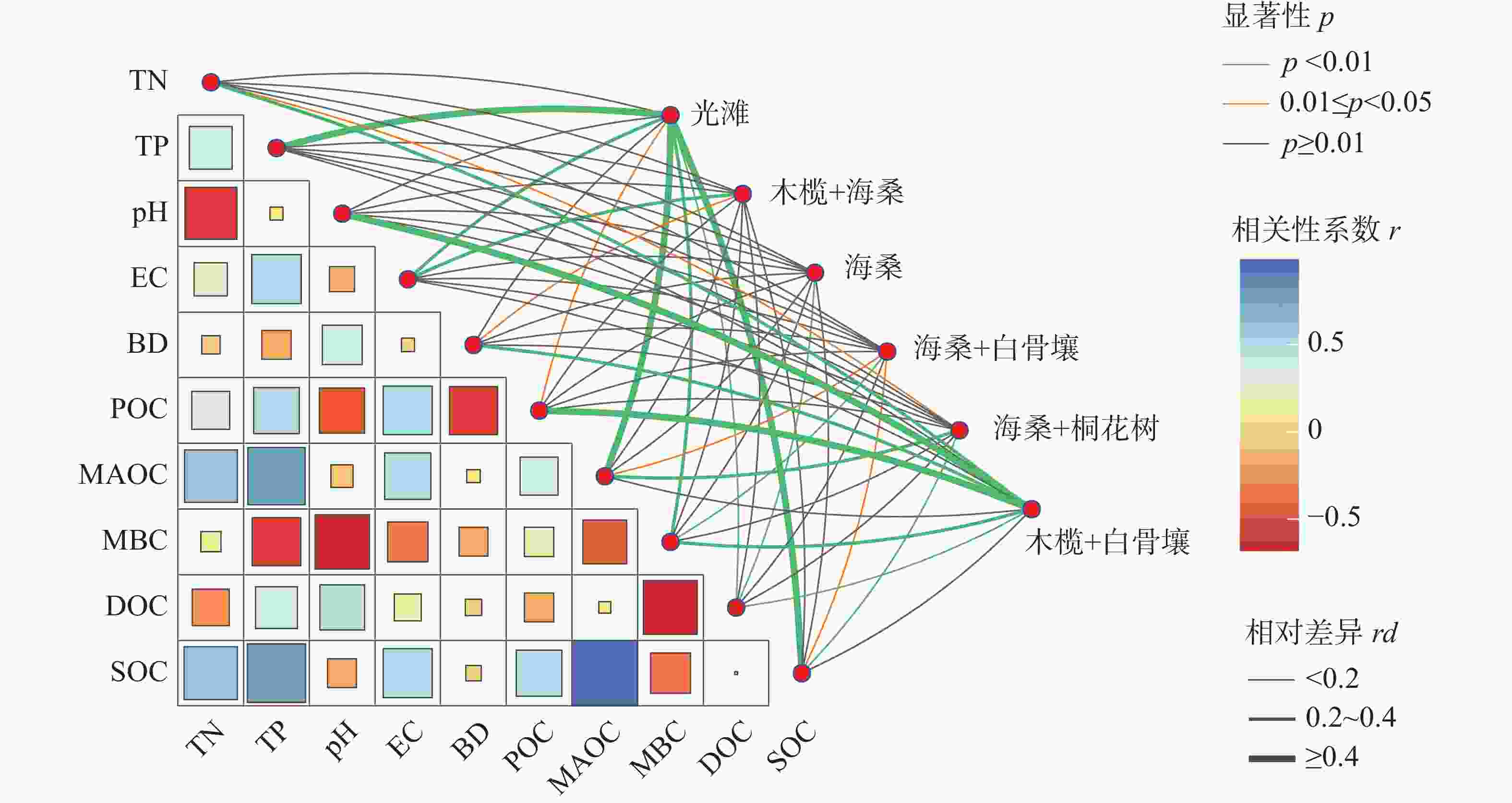
图 6 土壤碳组分与土壤理化性质之间的相关性热图及显著性网络
Figure 6. Correlation heatmap and significance network between soil carbon components and soil physicochemical properties
显著性网络结果进一步表明,不同红树林群落样地中SOC组分与土壤理化因子之间的关联存在差异。Mantel检验显示,不同群落样地与SOC组分及土壤因子之间的相关强度并不一致,其中光滩、木榄+白骨壤和白骨壤+海桑样地与SOC、MAOC、TN和TP之间的关联相对较强,而海桑单一种植样地的网络连接相对较少。相比之下,DOC和MBC等活性碳组分与pH等理化因子的关联更为明显。
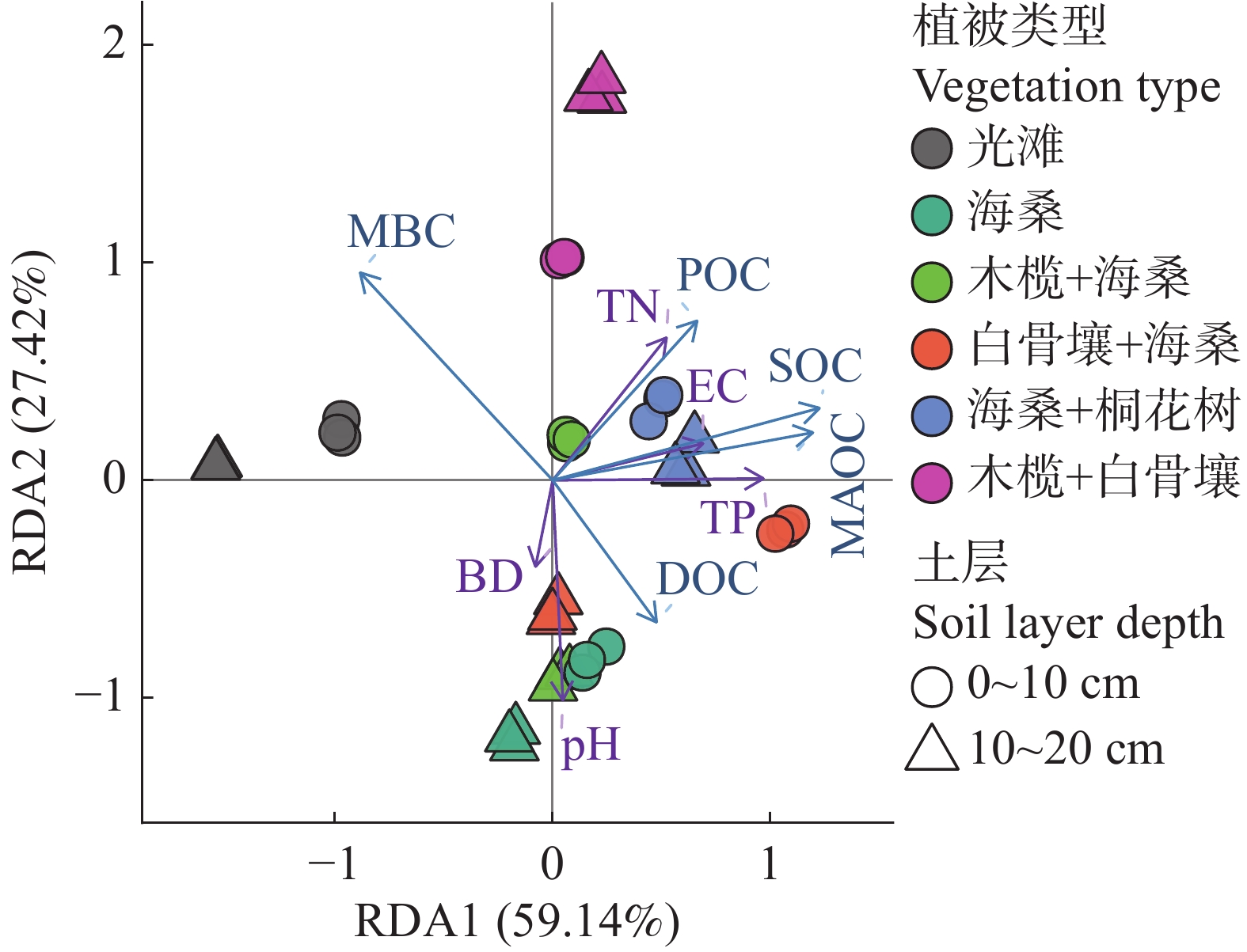
进一步地,RDA排序结果(图7)表明,SOC、MAOC、POC、DOC和MBC与pH、EC、TN、TP及BD等因子在排序空间中表现出明显的相关性,不同群落样地及不同土层样品沿环境梯度呈现一定的变化规律。RDA1和RDA2分别解释了59.14%和27.42%的变异,总解释率为86.56%。其中,SOC、MAOC和POC整体与EC、TN和TP方向较为一致,而DOC和MBC与pH、BD方向的关系相对更为明显。
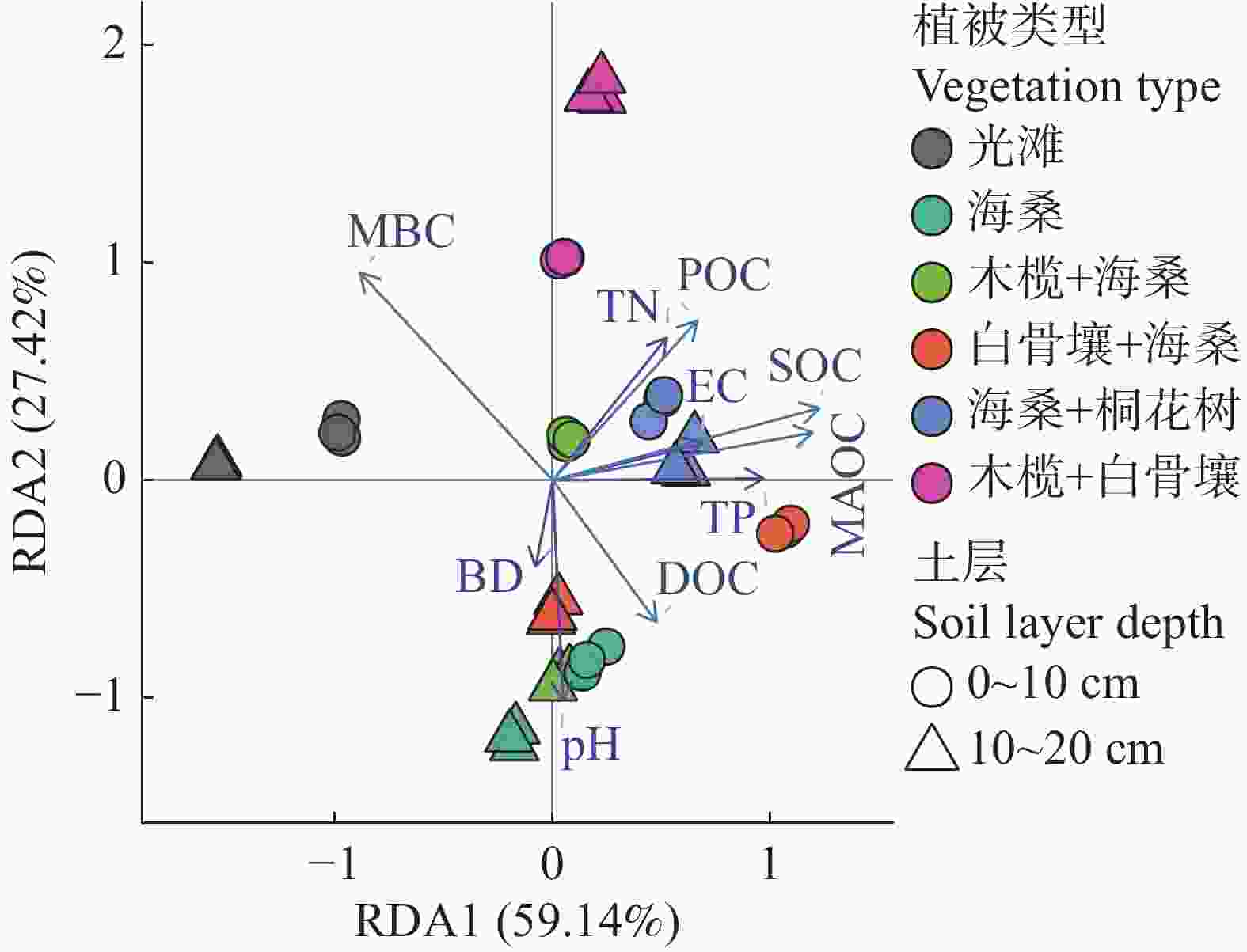
图 7 不同植被类型与土层土壤碳组分及理化因子的冗余分析(RDA)排序图
Figure 7. Redundancy analysis (RDA)ordination of soil carbon components and physicochemical properties across different vegetation types and soil depths
-
退塘还林初期,不同红树林群落与光滩土壤理化特征的显著差异,反映了红树林种植对退塘土壤的早期改造作用。已有研究表明,退塘还林初期种植红树林明显提升SOC及其活性组分,并在土壤理化特征上表现出显著空间异质性[6]。SOC和TN是土壤肥力核心指标,本研究中红树林群落两者含量均显著高于光滩,且多数样地表层(0~10 cm)高于深层(10~20 cm),表明红树林对土壤碳氮库的影响效应[20]。本研究表明,退塘还林初期的植被恢复显著改变了SOC及其组分的积累与分配格局。红树林即使在恢复初期也已表现出较强的有机碳固持能力,这种碳增益可能与红树林较高的初级生产力、地上与地下生物量输入增加,以及其对潮汐携带的细颗粒沉积物和有机碎屑的拦截作用有关[21]。然而,不同红树林群落间的SOC积累效应表现出显著的物种依赖作用。白骨壤+海桑混交群落和海桑+桐花树群落在表层SOC含量较高,说明不同树种组合会影响表层SOC的积累过程,这可能与海桑有机质输入较多有关[22-23]。相比之下,木榄+白骨壤群落的SOC积累相对较低,表明不同群落类型在有机质输入、沉积物拦截及土壤环境调节方面可能存在差异,从而影响SOC空间异质性。已有研究表明,在海南东寨港的多种红树林群落中,不同群落的SOC储量存在明显差异,表明群落类型是影响SOC的重要驱动因子[24]。在本研究中,海桑+桐花树群落在深层仍维持了较高的SOC含量,这可能与地下根系输入、沉积环境及有机质向下迁移和保存过程有关[25]。需要指出的是,基于样地比较、相关性分析和RDA排序结果,尚无法明确区分pH和EC在退塘还林初期演替过程中究竟是驱动因素还是结果反馈。一方面,不同红树林群落可能通过根系分泌、凋落物归还、离子吸收以及对沉积环境的调节作用,改变土壤的酸碱度和盐分条件[26];另一方面,样地初始的盐度、酸碱度及微地形差异也可能对红树林定植、生长及有机质输入产生作用。因此,pH和EC与SOC组分之间的关系更可能是初始微环境与植被恢复过程共同作用下形成的耦合结果,而非单一方向的因果作用。海桑+桐花树群落表现出中性偏碱性的pH和较高的EC(图2),可能与该群落的离子调节、沉积物淤积及有机质输入有关,同时这一环境特征也可能进一步影响SOC的积累与稳定[27]。
在碳组分层面,研究揭示了MAOC在SOC固持中的主导地位。MAOC在各红树林群落中的相对贡献率高达50.75%~91.19%(图4),显著高于光滩。MAOC通常被视为一种受到矿物物理保护、周转时间较长的稳定碳[28]。红树林群落显著提高了MAOC的含量及比例,表明退塘还林不仅增加了土壤碳的数量,更提升了土壤碳的质量及稳定性[15]。相反,光滩土壤SOC总量低,但其活性组分(MBC、POC、DOC)的相对占比却显著高于红树林区域(图4),且容重较大(图2),表明有机碳主要以未受保护的颗粒形态存在,极易受潮汐扰动和微生物矿化作用的影响而流失[29]。因此,从光滩到红树林的演替过程实质上是SOC从低储量与高周转性向高储量与高稳定性转变的过程。
此外,SOC组分的变异性受土壤因子与植被类型的共同驱动。Mantel检验和网络分析显示(图6),相比单一的海桑群落,混交群落木榄+白骨壤、白骨壤+海桑等组合与TN、TP和SOC之间形成了更强且更复杂的网络关系,说明混交群落中SOC积累与养分状态之间具有更紧密的耦合关系。结合MAOC在红树林土壤中的较高贡献率,推测混交群落较高的SOC含量可能不仅与有机质输入增加有关,还与其更有利于有机碳向稳定组分转化和保存有关[30]。不同树种组合可能通过改变土壤养分状况及理化环境,增强土壤碳与养分的耦合关系,进而影响SOC的积累与稳定[31]。
综上所述,退塘还林初期红树林恢复显著提升土壤SOC含量,并促进有机碳向以MAOC为主的较稳定组分转化,表明植被恢复对退塘土壤碳库的数量和稳定性均有积极作用。不同群落间SOC及其组分的差异表明,树种配置会影响土壤碳积累过程;根据本研究结果,这种差异更可能与树种组合引起的土壤环境调节、碳与养分耦合增强及碳稳定化过程的差异有关。因此,在退塘还林实践中,混交配置可作为提升土壤碳汇潜力的重要恢复方向。
-
本研究表明,不同植被恢复类型显著影响SOC储量及有机碳积累速率,且这种影响在表层(0~10 cm)中表现显著。总体上,各红树林群落的SOC储量显著高于光滩(图5),说明植被通过增加有机质输入和改善土壤环境条件来促进SOC恢复,这一结果与前文SOC含量及其稳定组分显著增加的结论相一致。红树林群落的SOC储量与中国其他恢复期红树林的SOC储量范围相一致。根据已有研究[24],海南东寨港红树林不同群落的0~120 cm土层SOC储量范围在2.74~10.18 kg·C·m−2之间。此外,Zou等[32]研究发现恢复期为4年的红树林的SOC密度范围为2.99~11.41 kg·C·m−2,在广东省惠东县考洲洋海域红树林种植岛的0~100 cm土层,SOC密度为7.2 kg·C·m−2[6]。因此,本研究结果与已有研究相符,与其他恢复期红树林湿地的SOC储量一致,表明本研究区的SOC储量与一般恢复期红树林湿地的碳储量水平相当,符合恢复过程中的趋势。然而,在本研究观测的碳储量均处于已有报道范围的低值区间,主要归因于两点:第一,研究区尚处于恢复初期,植被凋落物和根系的积累时间较短,尚未达到成熟林的土壤碳饱和状态[33];第二,退塘还林前的长期水产养殖活动,导致了原有SOC的流失。因此,本研究观测到的碳储量主要代表了恢复后的新生碳。值得注意的是,尽管总量尚不及成熟林,但海桑+桐花树群落的储量已接近部分次生林水平,这表明通过优化物种配置,可以有效加速碳储量的补充过程。与此同时,本研究仅分析0~20 cm土层,因此,相关结果主要代表恢复初期表层碳库特征,而不能直接等同于整个土壤剖面的蓝碳储量。
在碳动态方面,本研究的碳积累速率0.48~1.05 Mg·C·ha−1·a−1与中国华南沿海其他区域的研究结果高度一致。已有研究表明,中国沿海红树林的土壤碳积累速率具有明显的地域与生境差异。在海南岛东北部的实测研究中,河口型红树林沉积物碳埋藏速率约为141.5 g C·m−2·a−1(约1.42 Mg·C·ha−1·a−1),而开放海域型红树林的埋藏速率约为61.7 g C·m−2·a−1(约0.62 Mg ·C·ha−1·a−1)[34-35]。本研究结果恰好介于两者之间,可以看出,沙美内海半封闭的潟湖环境,其沉积物输入虽不及大型河口,但缓和的水动力条件优于开放海域,更有利于细颗粒有机质的沉降。Lin等[36]在广东省研究发现秋茄的土壤碳埋藏为0.25~1.55 Mg·ha−1·a−1,白骨壤的碳积累速率为0.36~1.00 Mg·C·ha−1·yr−1,与本研究在不同混交群落中的碳积累速率一致。这些数据进一步验证了红树林恢复在不同区域内对碳固存功能的增强作用[36]。此外,与邻近的广东省红树林相比,本研究中白骨壤+海桑混交群落的碳积累速率为0.59 Mg·ha−1·a−1,高于广东省区域研究的白骨壤单一种植林0.36~1.00 Mg·ha−1·a−1,这进一步验证了前文关于混交林协同增汇效应的结论。
研究表明,合理配置红树林物种组成,尤其是优先采用混种恢复模式,有助于在较短修复年限内显著提升土壤碳储量和碳积累速率。同时,10~20 cm土层在特定混交群落中表现出的高碳汇维持能力,未来在评估红树林修复碳汇潜力时,应充分考虑土壤垂向尺度的碳动态,以避免低估生态系统整体碳固存能力。本研究采样深度为0~20 cm,主要用于反映退塘还林初期不同恢复群落表层SOC组分及其对植被恢复的响应。表层土壤对红树林恢复具有较高的敏感性,但为长期监测土壤碳组分对红树林恢复的响应,未来可进一步增加采样土层深度,如覆盖50 cm甚至1 m,以更全面揭示碳组分的垂向变化特征。未来还可结合更深层土壤剖面采样与沉积柱分析,利用210Pb放射性核素测年建立沉积年代序列,重建近百年尺度的沉积物加积与SOC埋藏过程,从而更精准地揭示退塘还林碳汇形成过程及碳累积速率。
-
退塘还林初期(4年),所有红树林群落SOC、TN、TP含量均显著高于光滩,表明红树林恢复已对表层土壤碳、氮、磷库产生显著影响。其中白骨壤+海桑群落在表层(0~10 cm)的SOC含量最高,木榄+白骨壤群落TN含量最高。MAOC是土壤碳库的主要组分,红树林群落MAOC含量显著高于光滩,说明退塘还林有助于促进较稳定碳组分积累;相比之下,光滩中POC、DOC和MBC等活性有机碳组分相对贡献较高,其碳库稳定性相对较弱。土壤碳组分与TN、TP、EC和pH等环境因子密切相关,不同碳组分对环境因子的响应存在差异。其中,MAOC与TN、TP和EC的关联较强,而DOC、MBC等活性碳组分与pH等因子的关系更为显著。不同红树林群落样地土壤碳储量均高于光滩,部分混交群落在表层土壤碳储量及以光滩为参照的SOC积累速率高于单一树种群落,其中0~10 cm土层以白骨壤+海桑样地最高。总体而言,退塘还林有助于提升恢复初期土壤碳汇潜力,其中海桑+桐花树和白骨壤+海桑群落的0~20 cm SOC储量较高,且其SOC积累速率也处于较高水平,表明这两种混交群落具有较高的土壤碳汇提升潜力。
红树林群落对土壤有机碳组分及理化性质的影响
DOI: 10.15886/j.cnki.rdswxb.20260032
 CSTR: 32425.14.j.cnki.rdswxb.20260032
CSTR: 32425.14.j.cnki.rdswxb.20260032
Effects of mangrove communities on soil organic carbon fractions and physicochemical properties
-
摘要: 退塘还林是恢复红树林湿地生态系统、增加碳汇功能的重要途径。以海南琼海沙美内海退塘还林(4年)的红树林湿地为研究对象,采集光滩、海桑、木榄+海桑、白骨壤+海桑、海桑+桐花树以及木榄+白骨壤样地0~20 cm土壤样品(0~10 cm、10~20 cm),分析不同红树群落土壤有机碳(SOC)及其组分的分布特征,并探讨与土壤理化因子的关系。结果表明,研究区0~20 cm SOC含量为1.55~20.73 g·kg−1,5种红树群落样地SOC显著高于光滩,其中白骨壤+海桑与海桑+桐花树SOC含量最高。矿质结合有机碳(MAOC)为有机碳库的主要组成成分(50.75%~91.19%),退塘种植红树样地整体提高MAOC含量及比例;对照样地光滩SOC中颗粒有机碳(POC)、可溶性有机碳(DOC)和微生物生物量碳(MBC)占比较高,表明碳库稳定性较弱。退塘种植红树样地土壤碳储量显著高于光滩,且混交种植模式的土壤碳储量及以光滩为参照估算的SOC积累速率整体高于单一树种群落。有机碳积累速率在不同土层间差异显著,其中白骨壤+海桑样地0~10 cm的积累速率最高(0.59 Mg·ha−1·a−1)。Mantel检验与相关性分析结果表明,SOC与MAOC、TN、TP均为正相关(p<0.05),表明SOC组分分布与植被类型及养分条件密切相关。研究表明,退塘还林能够提高恢复初期表层土壤碳储量,并促进SOC向较稳定组分积累;其中海桑+桐花树和白骨壤+海桑群落的0~20 cm SOC储量较高,且其SOC积累速率也处于较高水平,可能具有较高的表层碳汇提升潜力,可为退塘还林生态工程中的树种配置优化与碳汇功能评估提供科学参考。Abstract: Pond-to-mangrove restoration is an important approach for restoring mangrove wetland ecosystems and enhancing their carbon sequestration function. A 4-year-old pond-to-mangrove restoration area in Shamei Inland Sea, Qionghai, Hainan, was selected as the study site. Soil samples from 0–20 cm depth (0–10 cm and 10–20 cm) were collected from mudflat, Sonneratia apetala, Bruguiera gymnorhiza + S. apetala, Avicennia marina + S. apetala, S. apetala + Aegiceras corniculatum, and B. gymnorhiza + A. marina stands. The distribution characteristics of soil organic carbon (SOC) and its fractions in different mangrove communities were analyzed, and their relationships with soil physicochemical factors were explored. The results showed that SOC content in the 0–20 cm soil layer ranged from 1.55 to 20.73 g·kg−1. SOC contents were significantly higher in the five mangrove communities than in the mudflat, with the highest values being observed in the A. marina + S. apetala and S. apetala + A. corniculatum stands. Mineral-associated organic carbon (MAOC) was the dominant component of the soil organic carbon pool, accounting for 50.75%–91.19%, and mangrove planting in restored ponds generally increased both the content and proportion of MAOC. In contrast, particulate organic carbon (POC), dissolved organic carbon (DOC), and microbial biomass carbon (MBC) accounted for relatively higher proportions of SOC in the mudflat, indicating weaker carbon pool stability. Soil carbon storage was significantly higher in mangrove-planted restored ponds than in the mudflat, and mixed-species plantations generally showed higher soil carbon storage and soil organic carbon accumulation rates, estimated using the mudflat as a reference, than monospecific stands. Significant differences in organic carbon accumulation rates were found among soil layers, with the highest accumulation rate occurring in the 0–10 cm layer of the A. marina + S. apetala stand (0.59 Mg·ha−1·a−1). Mantel tests and correlation analyses showed that SOC was positively correlated with MAOC, total nitrogen (TN), and total phosphorus (TP) (p < 0.05), indicating that the distribution of SOC fractions was closely related to vegetation type and nutrient conditions. These findings suggest that pond-to-mangrove restoration can increase surface soil carbon storage during the early restoration stage and promote the accumulation of soil organic carbon in more stable fractions. Among the restored mangrove communities, the S. apetala + A. corniculatum and A. marina + S. apetala stands exhibited relatively higher SOC storage in the 0–20 cm soil layer and maintained comparatively high SOC accumulation rates, suggesting a greater potential for enhancing surface soil carbon sequestration during the early restoration stage. These results may provide a scientific reference for optimizing species configuration and evaluating carbon sequestration functions in pond-to-mangrove restoration projects.
-
图 2 不同红树林群落土壤理化因子
注:不同大写字母表示0~10 cm土层不同样地间存在差异显著;不同小写字母表示10~20 cm土层不同样地间存在差异显著(P<0.05);* 表示同一样地不同土层间存在差异显著(P<0.05)。下同。
Fig. 2 Soil physicochemical properties of different mangrove communities
Note: Different uppercase letters indicate significant differences among the sites for the 0–10 cm soil layer; different lowercase letters indicate significant differences among the sites for the 10–20 cm soil layer (p<0.05), and * indicate significant differences between soil layers. Similarly hereinafter.
图 3 不同类型样地土壤碳组分特征
Fig. 3 Characteristics of soil carbon components across different types of sampling plots
图 4 不同类型样地土壤碳组分比例占比
Fig. 4 Proportional composition of soil carbon components across different types of sampling plots
图 5 不同红树林群落SOC储量及有机碳积累速率
Fig. 5 Soil organic carbon storage and carbon accumulation rates in different mangrove communities
图 6 土壤碳组分与土壤理化性质之间的相关性热图及显著性网络
注:颜色表示相关系数(r),连线颜色与粗细分别反映相关性的显著性水平和强度。
Fig. 6 Correlation heatmap and significance network between soil carbon components and soil physicochemical properties
Note: Heatmap and correlation network illustrating relationships between soil carbon fractions and soil physicochemical properties. Color gradients represent correlation coefficients (r), while line color and thickness indicate significance level and correlation strength, respectively.
图 7 不同植被类型与土层土壤碳组分及理化因子的冗余分析(RDA)排序图
注:点表示土壤样品,颜色表示植被类型,符号表示土层深度(0~10 cm和10~20 cm)。箭头表示土壤碳组分及理化因子,包括SOC、POC、MAOC、DOC、MBC、TN、TP、EC、pH和BD。箭头方向表示变量变化的梯度方向,箭头长度表示其与排序轴的相关强度;RDA1 和 RDA2 分别解释59.14%和27.42%的变异。
Fig. 7 Redundancy analysis (RDA)ordination of soil carbon components and physicochemical properties across different vegetation types and soil depths
Note: Points represent soil samples; colors indicate vegetation types, and symbols indicate soil depth intervals (0–10 cm and 10–20 cm). Arrows represent soil carbon fractions and physicochemical factors, including SOC, POC, MAOC, DOC, MBC, TN, TP, EC, pH, and BD. Arrow direction indicates the gradient of variable change, and arrow length indicates the strength of correlation with the ordination axes. RDA1 and RDA2 explain 59.14% and 27.42% of the variance, respectively.
-
[1] 张骁栋, 朱建华, 张小全, 等. 中国湿地碳汇功能的提升途径[J]. 自然保护地, 2022, 2(3): 17−23. https://doi.org/10.12335/2096-8981.2022072804 doi: 10.12335/2096-8981.2022072804 [2] 林鹏. 中国红树林湿地与生态工程的几个问题[J]. 中国工程科学, 2003, 5(6): 33−38. https://doi.org/10.3969/j.issn.1009-1742.2003.06.005 doi: 10.3969/j.issn.1009-1742.2003.06.005 [3] Peng Y S, Chen G Z, Li S Y, et al. Use of degraded coastal wetland in an integrated mangrove-aquaculture system: a case study from the South China Sea [J]. Ocean & Coastal Management, 2013, 85: 209−213. https://doi.org/10.1016/j.ocecoaman.2013.04.008 doi: 10.1016/j.ocecoaman.2013.04.008 [4] 徐彩瑶, 濮励杰, 朱明. 沿海滩涂围垦对生态环境的影响研究进展[J]. 生态学报, 2018, 38(3): 1148−1162. https://doi.org/10.5846/stxb201611142316 doi: 10.5846/stxb201611142316 [5] 罗庆, 何清, 吴慧秋, 等. 辽河口湿地土壤有机碳组分特征及其影响因素[J]. 生态环境学报, 2024, 33(3): 333−340. https://doi.org/10.16258/j.cnki.1674-5906.2024.03.001 doi: 10.16258/j.cnki.1674-5906.2024.03.001 [6] 李玲云, 韩婷婷, 张黄琛, 等. 养殖池塘退塘还林初期土壤有机碳及活性组分特征[J]. 南方水产科学, 2024, 20(4): 88−97. https://doi.org/10.12131/20240094 doi: 10.12131/20240094 [7] Cui X W, Liang J, Lu W Z, et al. Stronger ecosystem carbon sequestration potential of mangrove wetlands with respect to terrestrial forests in subtropical China [J]. Agricultural and Forest Meteorology, 2018, 249: 71−80. https://doi.org/10.1016/j.agrformet.2017.11.019 doi: 10.1016/j.agrformet.2017.11.019 [8] Chen G Y, Zhang M X, Yao X Y, et al. Soil organic carbon sequestration after 20-year afforestation of mangrove plantations on Qi'ao Island, southern China [J]. Agronomy, 2023, 13(9): 2389. https://doi.org/10.3390/agronomy13092389 doi: 10.3390/agronomy13092389 [9] Chowdhury A, Naz A, Maiti S K. Variations in soil blue carbon sequestration between natural mangrove metapopulations and a mixed mangrove plantation: a case study from the world's largest contiguous mangrove forest [J]. Life, 2023, 13(2): 271. https://doi.org/10.3390/life13020271 doi: 10.3390/life13020271 [10] Li Z, Li Y F, Fan B X. Diverse restored mangrove stands enhance carbon storage compared to monospecific plantation: a meta-analysis [J]. Forest Ecology and Management, 2025, 578: 122446. https://doi.org/10.1016/j.foreco.2024.122446 doi: 10.1016/j.foreco.2024.122446 [11] 陈顺洋, 安文硕, 陈彬, 等. 红树林生态修复固碳效果的主要影响因素分析[J]. 应用海洋学学报, 2021, 40(1): 34−42. https://doi.org/10.3969/J.ISSN.2095-4972.2021.01.004 doi: 10.3969/J.ISSN.2095-4972.2021.01.004 [12] 赵泽阳, 赵志忠, 付博, 等. 海南岛北部地区红树林湿地土壤有机碳分布规律及影响因素[J]. 广东农业科学, 2018, 45(12): 49−55. https://doi.org/10.16768/j.issn.1004-874X.2018.12.009 doi: 10.16768/j.issn.1004-874X.2018.12.009 [13] 罗娜娜, 盛茂银, 王霖娇, 等. 长期植被恢复对中国西南喀斯特石漠化土壤活性有机碳组分含量和酶活性的影响[J]. 植物生态学报, 2023, 47(6): 867−881. https://doi.org/10.17521/cjpe.2022.0216 doi: 10.17521/cjpe.2022.0216 [14] Zhang Y S, Xiao L, Guan D S, et al. The role of mangrove fine root production and decomposition on soil organic carbon component ratios [J]. Ecological Indicators, 2021, 125: 107525. https://doi.org/10.1016/j.ecolind.2021.107525 doi: 10.1016/j.ecolind.2021.107525 [15] Huang X Y, Li W S, Qin G M, et al. Mangrove restoration enhances blue carbon sequestration and its stability in a subtropical tidal wetland [J]. Functional Ecology, 2025, 39(9): 2437−2451. https://doi.org/10.1111/1365-2435.70121 doi: 10.1111/1365-2435.70121 [16] Kristensen E, Bouillon S, Dittmar T, et al. Organic carbon dynamics in mangrove ecosystems: a review [J]. Aquatic Botany, 2008, 89(2): 201−219. https://doi.org/10.1016/j.aquabot.2007.12.005 doi: 10.1016/j.aquabot.2007.12.005 [17] Zhang J F, Gan S C, Yang P J, et al. A global assessment of mangrove soil organic carbon sources and implications for blue carbon credit [J]. Nature Communications, 2024, 15(1): 8994. https://doi.org/10.1038/s41467-024-53413-z doi: 10.1038/s41467-024-53413-z [18] 陈小花, 陈宗铸, 雷金睿, 等. 东寨港红树林中不同群落区表层土壤有机碳及其活性组分含量[J]. 湿地科学, 2022, 20(4): 499−506. https://doi.org/10.13248/j.cnki.wetlandsci.2022.04.006 doi: 10.13248/j.cnki.wetlandsci.2022.04.006 [19] . Chen G Y, Zhang M X, Yao X Y, et al. Soil organic carbon stock, source, and stability after 20-year mangrove afforestation in southern China [EB/OL]. (2023-04-17). https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4421376.(查阅网上资料,未找到本条文献引用日期信息,请补充) [20] Jiang D L, Nie T, Yan J, et al. Restoration type determines synchronic recovery of soil carbon, nitrogen, and phosphorus in mangrove wetlands [J]. Journal of Environmental Management, 2025, 385: 125658. https://doi.org/10.1016/j.jenvman.2025.125658 doi: 10.1016/j.jenvman.2025.125658 [21] 苏娟, 王凡, 王艺杰, 等. 中国红树林生态系统的碳源汇特征及核算[J]. 地球科学进展, 2024, 39(9): 902−914. https://doi.org/10.11867/j.issn.1001-8166.2024.068 doi: 10.11867/j.issn.1001-8166.2024.068 [22] Yang Z Y, Song W M, Zhao Y, et al. Differential responses of litter decomposition to regional excessive nitrogen input and global warming between two mangrove species [J]. Estuarine, Coastal and Shelf Science, 2018, 214: 141−148. https://doi.org/10.1016/j.ecss.2018.09.018 doi: 10.1016/j.ecss.2018.09.018 [23] Shaltout K H, Ahmed M T, Alrumman S A, et al. Standing crop biomass and carbon content of mangrove Avicennia marina (Forssk. ) Vierh. along the Red Sea coast of Saudi Arabia [J]. Sustainability, 2021, 13(24): 13996. https://doi.org/10.3390/su132413996 doi: 10.3390/su132413996 [24] 詹绍芬, 黄勃, 陈玉军, 等. 不同红树林群落土壤环境有机碳比较[J]. 热带生物学报, 2015, 6(4): 397−402. https://doi.org/10.15886/j.cnki.rdswxb.2015.04.007 doi: 10.15886/j.cnki.rdswxb.2015.04.007 [25] Conroy B M, Kelleway J J, Rogers K. Root productivity contributes to carbon storage and surface elevation adjustments in coastal wetlands [J]. Plant and Soil, 2025, 513(1): 605−631. https://doi.org/10.1007/s11104-025-07204-0 doi: 10.1007/s11104-025-07204-0 [26] Panchal P, Preece C, Peñuelas J, et al. Soil carbon sequestration by root exudates [J]. Trends in Plant Science, 2022, 27(8): 749−757. https://doi.org/10.1016/j.tplants.2022.04.009 doi: 10.1016/j.tplants.2022.04.009 [27] Teng J L, Tian J, Yu G R, et al. Soil properties and root traits jointly shape fine-scale spatial patterns of bacterial community and metabolic functions within a Korean pine forest [J]. PeerJ, 2021, 9: e10902. https://doi.org/10.7717/peerj.10902 doi: 10.7717/peerj.10902 [28] Lavallee J M, Soong J L, Cotrufo M F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century [J]. Global Change Biology, 2020, 26(1): 261−273. https://doi.org/10.1111/gcb.14859 doi: 10.1111/gcb.14859 [29] Cooray I G, Chalmers G, Chittleborough D, et al. Soil carbon fractionation as a tool to monitor coastal wetland rehabilitation [J]. Journal of Environmental Management, 2025, 373: 123950. https://doi.org/10.1016/j.jenvman.2024.123950 doi: 10.1016/j.jenvman.2024.123950 [30] Wu L L, Song Z L, Wu Y T, et al. Organic matter composition and stability in estuarine wetlands depending on soil salinity [J]. Science of the Total Environment, 2024, 945: 173861. https://doi.org/10.1016/j.scitotenv.2024.173861 doi: 10.1016/j.scitotenv.2024.173861 [31] Guo J H, Kneeshaw D, Peng C H, et al. Positive effects of species mixing on biodiversity of understory plant communities and soil health in forest plantations [J]. Proceedings of the National Academy of Sciences of the United States of America, 2025, 122(11): e2418090122. https://doi.org/10.1073/pnas.2418090122 doi: 10.1073/pnas.2418090122 [32] Zou H M, Li X L, Li S, et al. Soil organic carbon stocks increased across the tide-induced salinity transect in restored mangrove region [J]. Scientific Reports, 2023, 13(1): 19758. https://doi.org/10.1038/s41598-023-45411-w doi: 10.1038/s41598-023-45411-w [33] Zuo H L, Xu W S, Liu Z Y, et al. Long-term plant diversity increases soil extractable organic carbon and nitrogen contents in a subtropical forest [J]. Science of the Total Environment, 2023, 878: 163118. https://doi.org/10.1016/j.scitotenv.2023.163118 doi: 10.1016/j.scitotenv.2023.163118 [34] Thura K, Serrano O, Gu J L, et al. Mangrove restoration built soil organic carbon stocks over six decades: a chronosequence study [J]. Journal of Soils and Sediments, 2023, 23(3): 1193−1203. https://doi.org/10.1007/s11368-022-03418-2 doi: 10.1007/s11368-022-03418-2 [35] Jiang Z M, Sanders C J, Xin K, et al. Increasing carbon and nutrient burial rates in mangroves coincided with coastal aquaculture development and water eutrophication in NE Hainan, China [J]. Marine Pollution Bulletin, 2024, 199: 115934. https://doi.org/10.1016/j.marpolbul.2023.115934 doi: 10.1016/j.marpolbul.2023.115934 [36] Lin W J, Lin C W, Wu H H, et al. Mangrove carbon budgets suggest the estimation of net production and carbon burial by quantifying litterfall [J]. CATENA, 2023, 232: 107421. https://doi.org/10.1016/j.catena.2023.107421 doi: 10.1016/j.catena.2023.107421 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 162
- HTML全文浏览量: 94
- 被引次数: 0
