-
KNOX(KNOTTED1-like homeobox)基因家族属于同源异形盒基因TALE(Three Amino-acid Loop Extension)超家族中的一类亚家族,在植物生长发育中发挥重要作用。KNOX基因家族根据保守结构域的不同可将其分为4种类型:KNOX1、KNOX2、ELK和Homeobox_KN[1]。在结构域N端的KNOX1和KNOX2协同作用,不仅调控靶基因的表达,还促进蛋白质间的同源二聚化。在结构域上游的ELK则具有转录抑制功能,影响基因的表达。而位于结构域C末端的Homeobox_KN能够识别并锁定下游靶基因中的启动子序列,从而启动或调控特定的基因表达过程[2]。根据KNOX基因家族的结构特征和系统发育分析,将该家族分为Class Ⅰ、Class Ⅱ和Class M 3类,不同类别的KNOX基因的表达模式及生物学功能表现出明显差异[3 − 4]。Ⅰ类亚家族对于植物分生组织的形成和维持起着核心作用。这类基因主要在植物的顶端分生组织中特异性表达,通过调控与植物器官形成密切相关的细胞分化过程,最终影响侧生器官的形态建成[5]。相对Ⅰ类亚家族而言,Ⅱ类亚家族的表达部位分布较为广泛,在植物的各个组织器官中均有所表达。Ⅱ类亚家族主要参与调控植物器官的分化过程,并且在次级细胞壁的生物合成中发挥重要作用[6]。M类亚家族主要表达在植物器官原基的近端域和成熟器官的边缘,可能参与感知或产生开花信号[7 − 8]。
在拟南芥(Arabidopsis thaliana)KNOX基因家族中,Ⅰ类亚家族包含STM(SHOOT MERISTEMLESS)、KNAT1、KNAT2和KNAT6,而Ⅱ类亚家族则涵盖KNAT3、KNAT4、KNAT5和KNAT7,这些基因在植物体内广泛分布和表达[9]。STM和KNAT1在SAM(Shoot Apical Meristem,茎尖分生组织)的分化和维持中发挥着重要作用,KNAT6也被证实在SAM和胚胎发生过程中参与维持组织边界的完整性,KNAT2则着重于调节花型特征[10]。Ⅰ类亚家族的基因不仅参与侧向器官的形态发生过程,还维持SAM功能以及塑造叶片形态的多样性[11]。Ⅱ类亚家族的KNAT3基因与拟南芥种子萌发和幼苗生长发育过程中的脱落酸响应过程有关[12],KNAT7基因与植物次生细胞壁合成有关[13]。对M类亚家族研究较少,在拟南芥的研究中发现其与叶片的发育有关[7]。
睡莲属(Nymphaea)是睡莲科(Nymphaeaceae)的多年生水生草本植物,具有极其丰富的遗传多样性[14]。睡莲各部位含有丰富的类黄酮和多糖等活性物质,具有极高的药用价值、食用价值和观赏价值[15]。睡莲的繁殖方式一般分为有性繁殖和无性繁殖,其中,无性繁殖方式较为多样,包括块茎繁殖、球茎繁殖、根状茎繁殖、营养体繁殖和胎生繁殖[16]。睡莲的叶片胎生尤为独特,表现为在叶脐的部位形成胎生点,之后发育形成胎生芽,发育成熟后脱离母体形成新的植株,常见于热带睡莲。‘蓝鸟’睡莲(Nymphaea ‘Blue Bird’)是睡莲中栽培最广泛、商品价值最高的热带品种之一,同时,也是胎生睡莲的代表性品种。本实验室初步的研究表明,KNOX基因家族也可能在‘蓝鸟’睡莲叶片胎生繁殖中发挥重要作用。随着各个物种全基因组数据的发布,目前,在拟南芥、番茄(Solanum lycopersicum)、水稻(Oryza sativa)和玉米(Zea mays)等植物中已完成了KNOX基因家族的鉴定和分析,而睡莲虽然已经公布了代表性种蓝星睡莲(Nymphaea caerulea)的基因组[17],但关于KNOX基因家族的报道尚属空白。因此,本研究首先对蓝星睡莲基因组中的KNOX基因家族成员进行了系统性的分析,成功确定了睡莲15个KNOX基因,并进一步探究了它们的理化性质、亚细胞定位特征,构建了系统进化树以揭示其进化关系,然后分析了这些基因在染色体上的定位情况和种间共线性情况,最后通过转录组数据分析了这些基因在‘蓝鸟’睡莲不同发育时期胎生和非胎生叶片中的表达情况。这些研究成果不仅可以为了解KNOX基因家族的进化特征提供基础的理论支持,同时也为挖掘该家族在睡莲中的潜在功能奠定了坚实的基础。
-
首先从Ensemble Plants数据库(
https://plants.ensembl.org/index.html )下载蓝星睡莲基因组序列、蛋白质序列和注释文件。同时为进行跨物种比较,还需下载拟南芥、番茄、水稻和玉米的KNOX家族成员蛋白序列。为了识别蓝星睡莲中可能含有KNOX家族相关结构域的蛋白,本研究从Pfam数据库(http://pfam-legacy.xfam.org/ )下载了KNOX1(PF03790)、KNOX2(PF03791)、ELK(PF03789)及Homeobox_KN(PF05920)的结构域信息和隐马尔可夫模型(HMM)文件。 -
首先将已获取的保守结构域信息和隐马尔可夫模型(HMM)文件导入TBtools(v2.127)软件[18],并执行HMMER命令,筛选条件设置为e值小于1e−5,在蓝星睡莲的基因组数据库中筛选包含特定结构域的蛋白序列。本研究利用拟南芥、水稻、番茄和玉米的KNOX蛋白序列作为参照,通过TBtools软件的BLASTp命令在蓝星睡莲的蛋白数据库中搜索同源序列,同样以e值小于1e−5为筛选标准,并对HMMER和BLASTp两种方法得到的序列进行交集分析,并利用SMART(
https://smart.embl.de/ )和NCBI CDD数据库(https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml )进行了保守结构域的鉴定。通过MEGA7.0软件对睡莲KNOX家族成员进行多序列比对,再利用TBtools软件的TrimAL Wrapper功能进行蛋白序列修剪和可视化处理。利用ProtParam(
https://web.expasy.org/protparam/ )工具和WoLFPSORT(https://wolfpsort.hgc.jp/ )网站预测每个KNOX蛋白的氨基酸数(aa)、分子质量、等电点(pI)以及亚细胞定位。 -
为了解拟南芥、水稻、番茄、玉米及睡莲中KNOX基因家族成员之间的进化关系,本研究采用MEGA7.0软件的Muscle比对方法对KNOX蛋白序列进行多重比对,选用邻接法(Neighbor-joining)来构建系统发育树,同时运用ITOL(
https://itol.embl.de/ )在线网站来对系统发育树进行美化。 -
基于睡莲KNOX家族成员的蛋白序列和注释信息,本研究通过NCBI Batch CD-search工具(
https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )批量检索并下载保守结构域信息。同时,利用MEME在线工具(https://meme-suite.org/meme/tools/meme )对序列进行motif预测,设定motif数量为10,其余保持默认参数,并下载预测结果。通过TBtools软件的Gene Structure View功能,对睡莲KNOX家族成员的外显子和内含子结构进行分析。 -
利用TBtools软件从睡莲基因组数据库中提取15个NcKNOX基因的CDS序列上游2 000 bp的启动子区域,将获得的启动子区域序列提交到Plant Care数据库(
http:/bioinformatics.psb.ugent.be/webtools/plant care/html/ )来进行顺式作用元件分析。 -
基于睡莲基因组的注释文件获取KNOX基因家族成员的染色体位置信息,利用TBtools软件中的Gene Location Visualize功能进行了可视化处理。
-
基于拟南芥、水稻、番茄、玉米及睡莲的KNOX蛋白序列以及注释文件,用TBtools软件的One Step MC Scan X和Circle Gene View功能来进行多物种间和睡莲种内的共线性分析,获得这几个物种间及睡莲种内KNOX基因的共线关系图。
-
为了解KNOX基因家族成员在睡莲胎生叶片中的表达模式,探究NcKNOX基因在睡莲叶片胎生苗发生过程中的可能作用,选用胎生睡莲的代表性品种‘蓝鸟’睡莲的不同时期胎生叶片和非胎生叶片作为实验材料,研究其表达规律。根据‘蓝鸟’睡莲的胎生(Viviparous, Viv)及非胎生(Non-viviparous, Nvi)叶片的生长状态,将其分为胎生和非胎生1~3期(Viv 1~3和Nvi 1~3)的叶片,并将其胎生点(直径约为1 cm,图1)切下置于−80 ℃保存,送迈维代谢公司进行转录组测序[19]。根据转录组数据,使用TBtools软件中HeatMap对NcKNOX基因的FPKM值进行Log scale处理并进行表达模式可视化分析。

Figure 1. Developmental stages of 'Blue Bird' water lily leaves
-
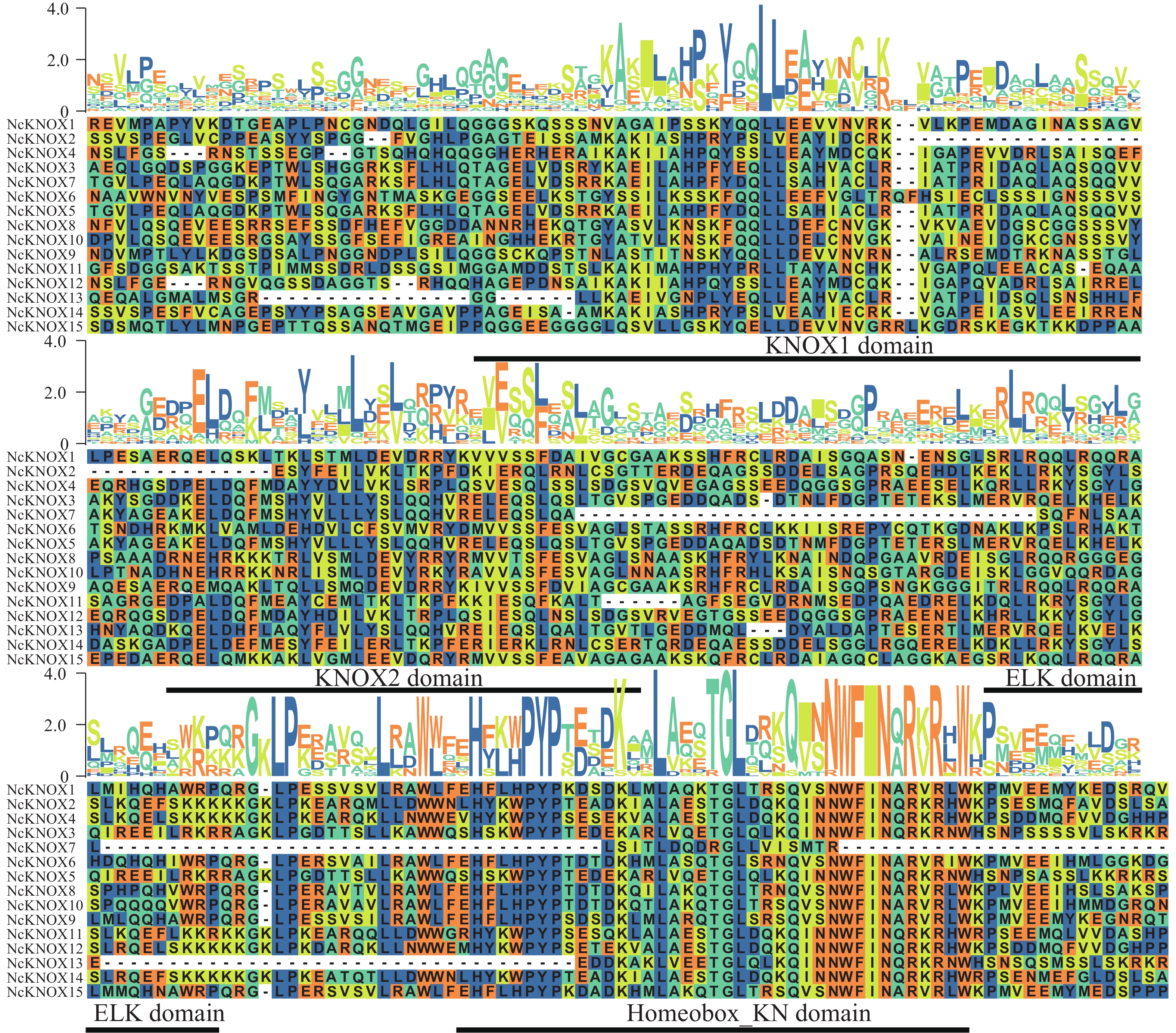
结合BLASTp和HMMER得到的数据取交集并进行人工校正,最终鉴定得到15个睡莲KNOX基因。蛋白序列比对分析(图2)可知,KNOX基因家族的4种保守结构域的保守情况,分别是典型保守序列KNOX1、KNOX2、ELK和Homeobox_KN,其中,Homeobox_KN结构域相较于其他结构域,其保守性更强,表明其在睡莲KNOX基因中更加稳定。
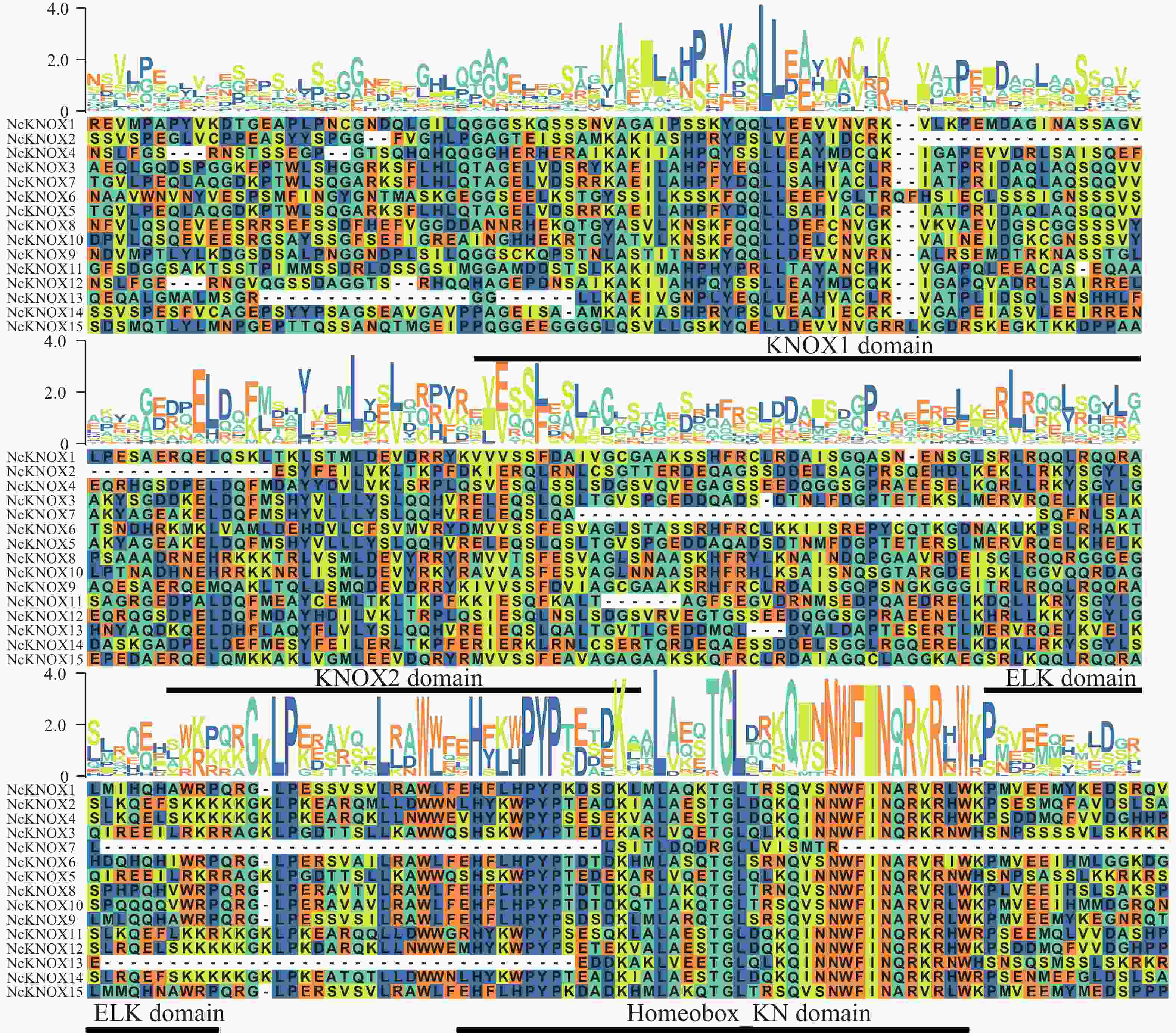
Figure 2. Sequence alignment of NcKNOX proteins
从表1可知,睡莲KNOX基因家族成员的CDS序列长度为909~6 813 bp,氨基酸序列长度为249~2 270 aa,氨基酸序列分子质量在2.84~24.95 kDa之间,等电点(pI)在5.07~8.8之间,其中pI<7的酸性氨基酸有13个,pI>7的碱性氨基酸有2个,富含酸性氨基酸,这与玉米的KNOX基因家族相似[20]。除了NC6G0258750.1(NcKNOX10)以外,其余的蛋白亲疏水性均小于0,且溶脂指数均小于100,则说明睡莲KNOX基因家族成员大都具备显著的亲水性特征。亚细胞定位预测结果显示,NcKNOX基因均位于细胞核内,这也与玉米、谷子等物种的KNOX基因家族成员细胞定位一致,与KNOX基因的转录因子属性特征相符合[21 − 22]。
基因ID
Gene ID基因名称
Gene
Name氨基酸/aa
Amino acid
number/aa分子质量/kDa
Molecular
weight/kDa等电点
PI不稳定指数
Instability
index溶脂指数
Aliphatic
index(AI)亲水性均值
Grand average of
hydrophobicity亚细胞定位预测
Subcellular
localizationNC1G0108910.1 NcKNOX1 1 371 15.20 8.30 49.32 74.49 −0.468 Nucleus NC2G0008460.1 NcKNOX2 296 3.29 5.39 50.65 60.41 −0.698 Nucleus NC3G0221260.1 NcKNOX3 456 5.02 5.67 55.30 67.04 −0.675 Nucleus NC3G0227930.1 NcKNOX4 421 4.82 6.34 48.79 62.11 −0.766 Nucleus NC4G0011790.1 NcKNOX5 443 4.82 5.29 58.34 66.84 −0.667 Nucleus NC4G0020470.1 NcKNOX6 780 8.46 6.03 46.25 71.01 −0.406 Nucleus NC4G0153810.1 NcKNOX7 302 3.21 5.09 58.05 77.35 −0.308 Nucleus NC5G0049120.1 NcKNOX8 800 8.69 6.92 47.53 69.46 −0.554 Nucleus NC6G0041000.1 NcKNOX9 810 8.91 6.12 45.98 71.53 −0.508 Nucleus NC6G0258750.1 NcKNOX10 2 270 24.95 8.80 43.15 89.85 0.022 Nucleus NC7G0308290.1 NcKNOX11 395 4.33 5.67 64.02 57.90 −0.648 Nucleus NC9G0170310.1 NcKNOX12 367 4.12 6.02 47.00 58.53 −0.860 Nucleus NC9G0274690.1 NcKNOX13 249 2.84 5.2 62.95 84.26 −0.602 Nucleus NC11G0123640.1 NcKNOX14 373 4.18 5.17 52.20 68.10 −0.506 Nucleus NC13G0026850.1 NcKNOX15 714 7.68 6.28 52.37 69.41 −0.477 Nucleus Table 1. Basic information of the NcKNOX protein family in water lily
-
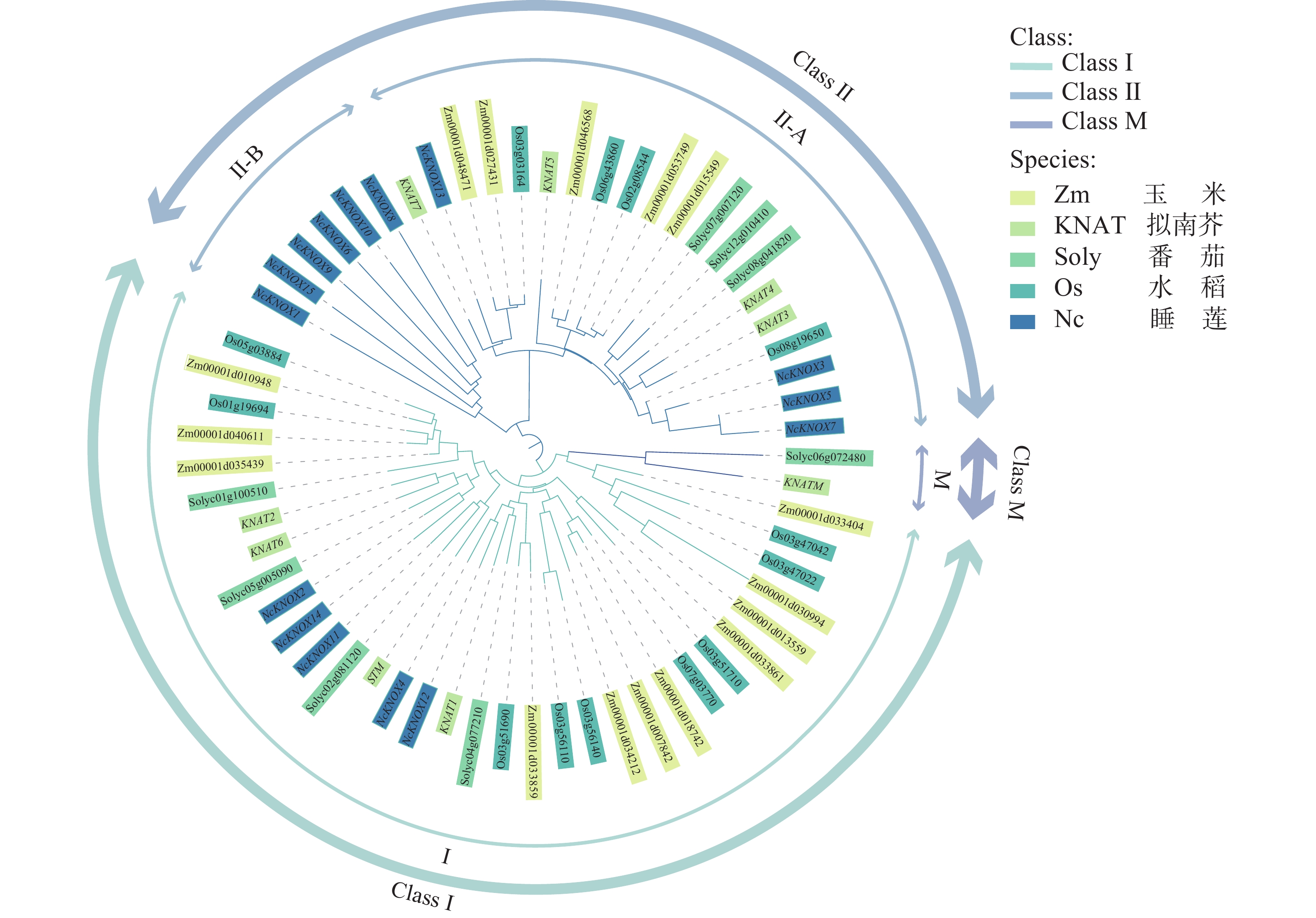
使用MEGA7.0软件利用蛋白序列绘制包含模式植物拟南芥和睡莲等多种植物KNOX基因家族成员的系统发育进化树。从图3可知,KNOX基因被分为3大类:Class Ⅰ、Class Ⅱ和Class M。其中Class Ⅰ类中含有5个NcKNOX成员,分别为NcKNOX2、NcKNOX4、NcKNOX11、NcKNOX12、NcKNOX14,与拟南芥的KNAT1、STM等基因进化关系比较近;Class Ⅱ类中含有10个成员,分别为NcKNOX1、NcKNOX3、NcKNOX5、NcKNOX6、NcKNOX8、NcKNOX9、NcKNOX10、NcKNOX7、NcKNOX13和NcKNOX15,与拟南芥的KNAT3、KNAT7等基因进化关系比较近。
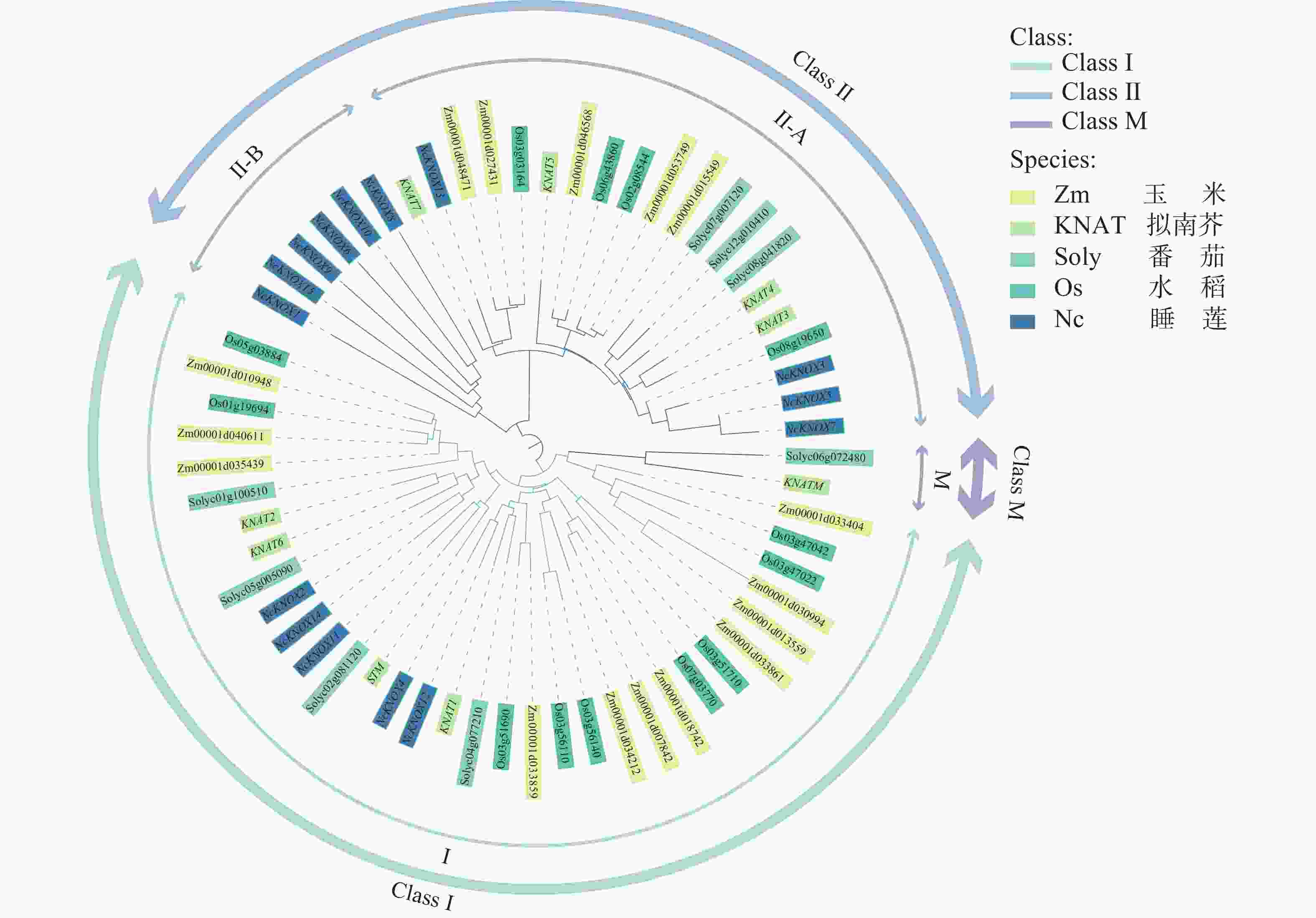
Figure 3. Phylogenetic tree analysis of KNOX proteins from Arabidopsis, tomato, rice, maize, and water lily
睡莲目作为被子植物谱系中的基源分支,其分化进程早于单、双子叶植物,既保留了双子叶植物的某些特征,又兼具单子叶植物的某些属性。而Class M类基因是双子叶植物中一个特殊的KNOX基因亚家族[23],在系统发育树中仅有番茄的Solyc06g072480基因与拟南芥的KNATM基因所属同一分支,与前人研究相符[24],而在睡莲中没有发现与KNATM基因相近的基因。因此,睡莲NcKNOX家族被分为Class Ⅰ类和Class Ⅱ类,没有Class M类基因。值得关注的是,睡莲NcKNOX1、NcKNOX6、NcKNOX8、NcKNOX9、NcKNOX10和NcKNOX15基因虽属于Class Ⅱ类基因,但在进化关系上与其他物种的Class Ⅱ基因相隔较远,独属于一个比较独特的分支,因此,将其命名为Class Ⅱ-B分支,对应的将其他Class Ⅱ基因命名为Class Ⅱ-A。
-
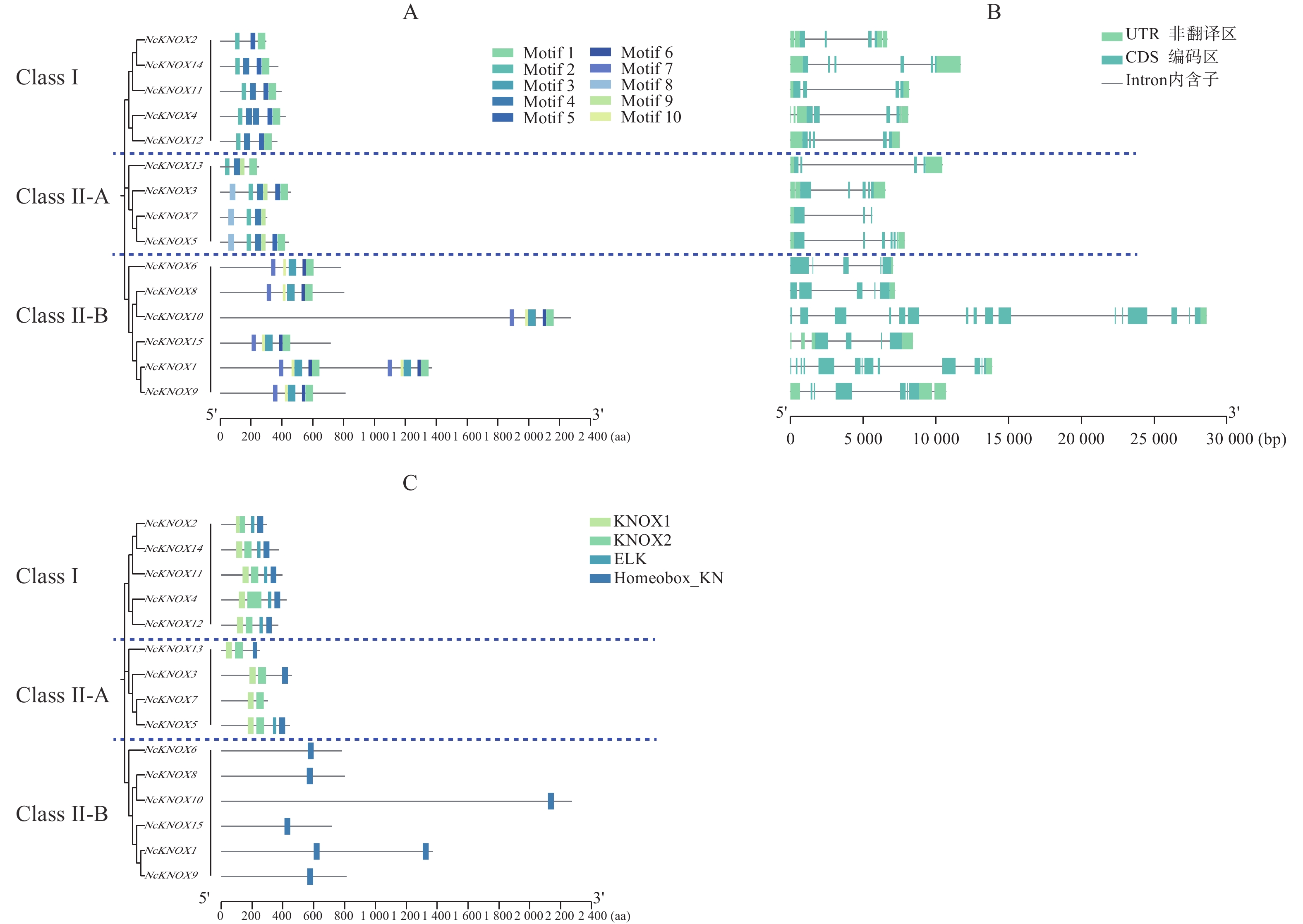
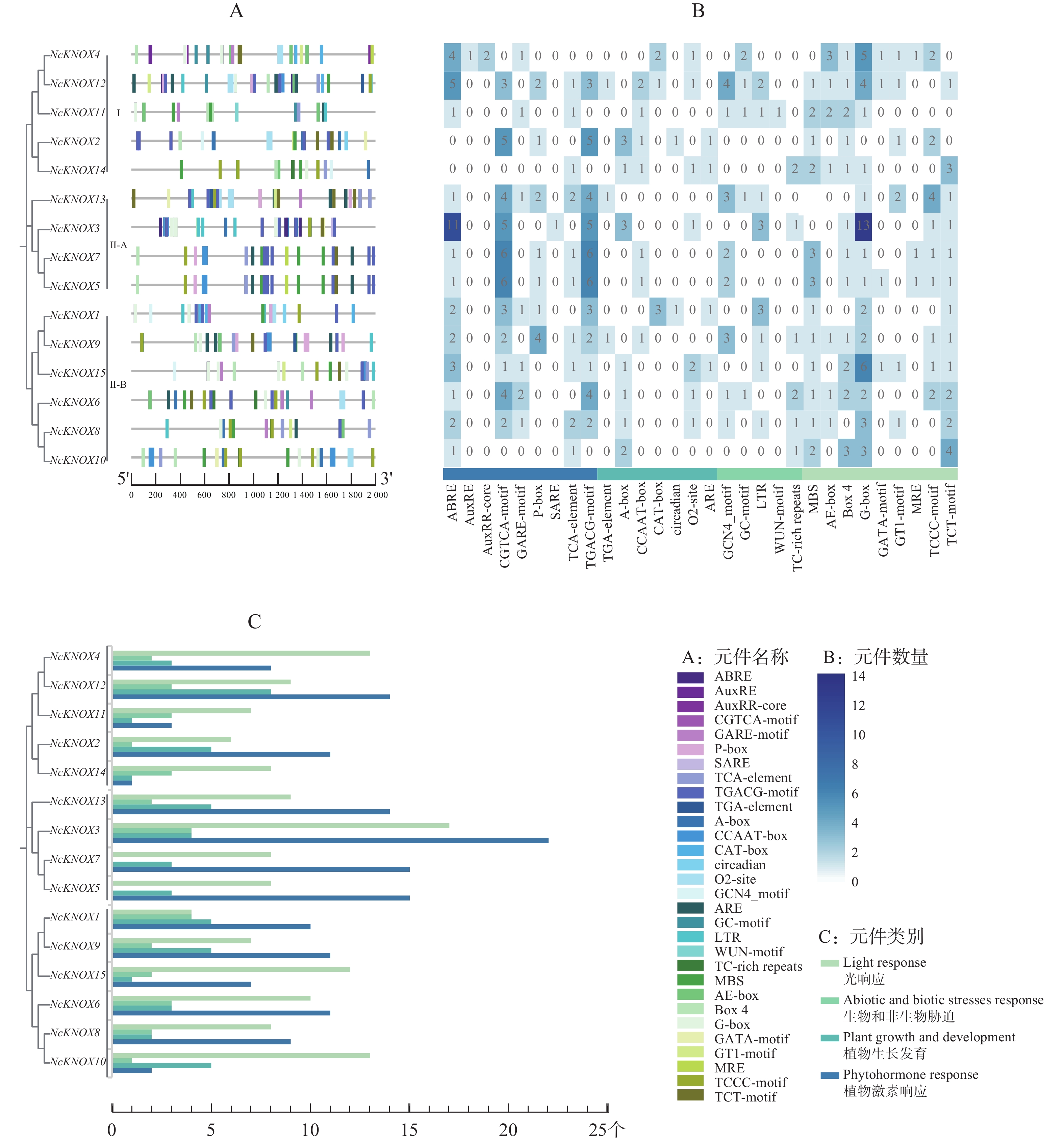
从图4−A可知,睡莲KNOX蛋白家族的大多数motif在蛋白质序列的C末端区域高度富集。某些基序在大多数KNOX蛋白中都有,如motif 1;也有一些基序的分布具有特异性,如Class Ⅰ类亚族没有motif 6、7、8、9、10,而Class Ⅱ类的Ⅱ-B分支则都有motif 2,说明这两类基因在进化上可能保留了某些共同的功能基础但又具有本亚族的独特性。
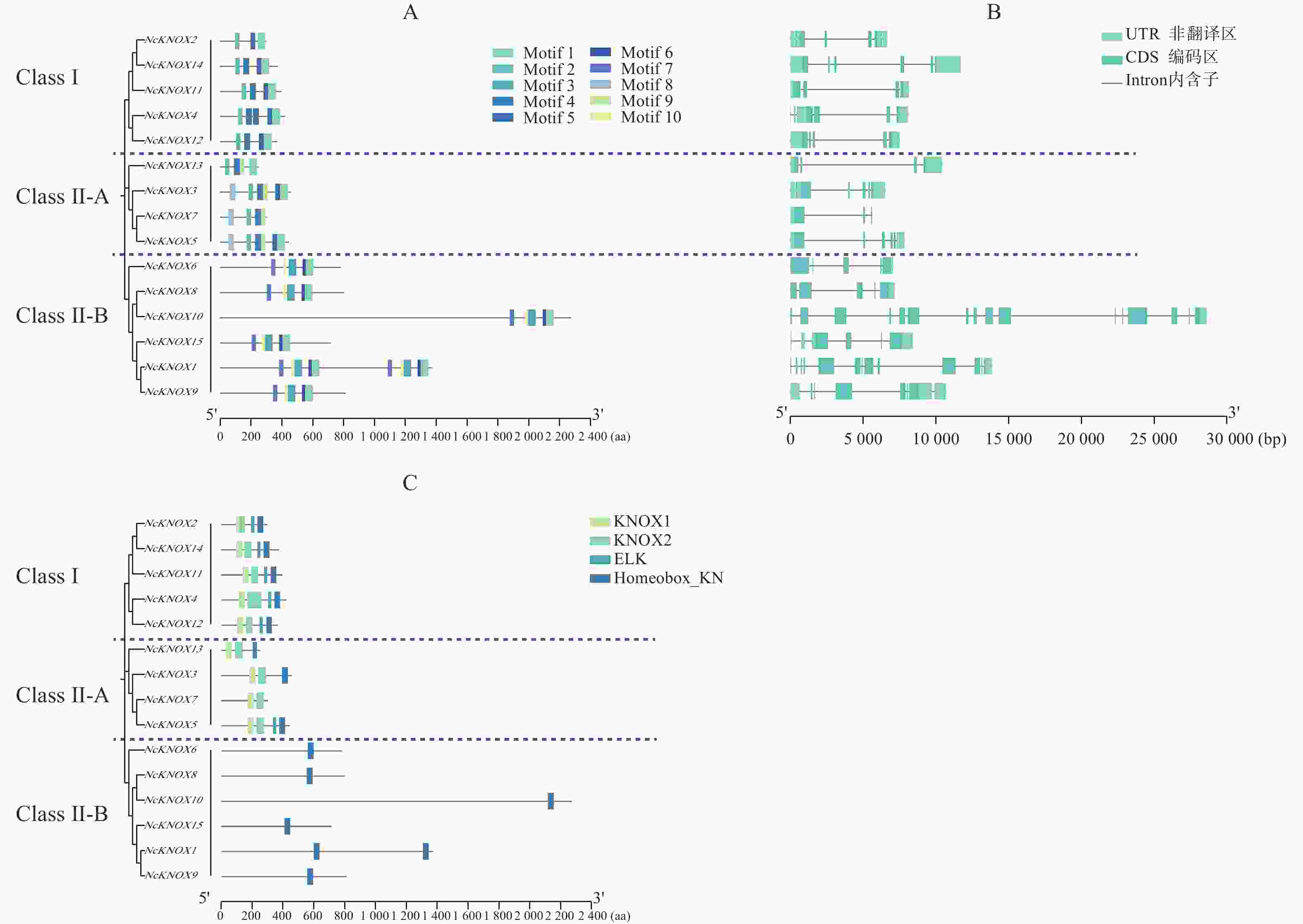
Figure 4. Motif, structure and conserved domain of the NcKNOX gene family in water lily
基因结构分析显示(图4−B),NcKNOX基因家族成员的序列长度相差较大,导致不同成员之间的内含子与外显子数目存在较大差异。NcKNOX基因家族主要由3~16个外显子和2~15个内含子组成,其中,序列最长的NcKNOX10含有16个外显子和15个内含子,序列最短的NcKNOX7含有3个外显子和2个内含子。
保守结构域分析显示(图4−C),除了NcKNOX7之外,所有的NcKNOX基因均编码了Homeobox_KN保守结构域,其中,Class Ⅱ-B分支上的NcKNOX1、NcKNOX6、NcKNOX8、NcKNOX9、NcKNOX10和NcKNOX15等基因比较特殊,仅包含Homeobox_KN结构域,而Class Ⅰ与Class Ⅱ-A的NcKNOX基因还普遍含有KNOX1和KNOX2这两个核心结构域,Class Ⅰ类的全部成员以及Class Ⅱ-A类中的NcKNOX5基因还具有ELK结构域。
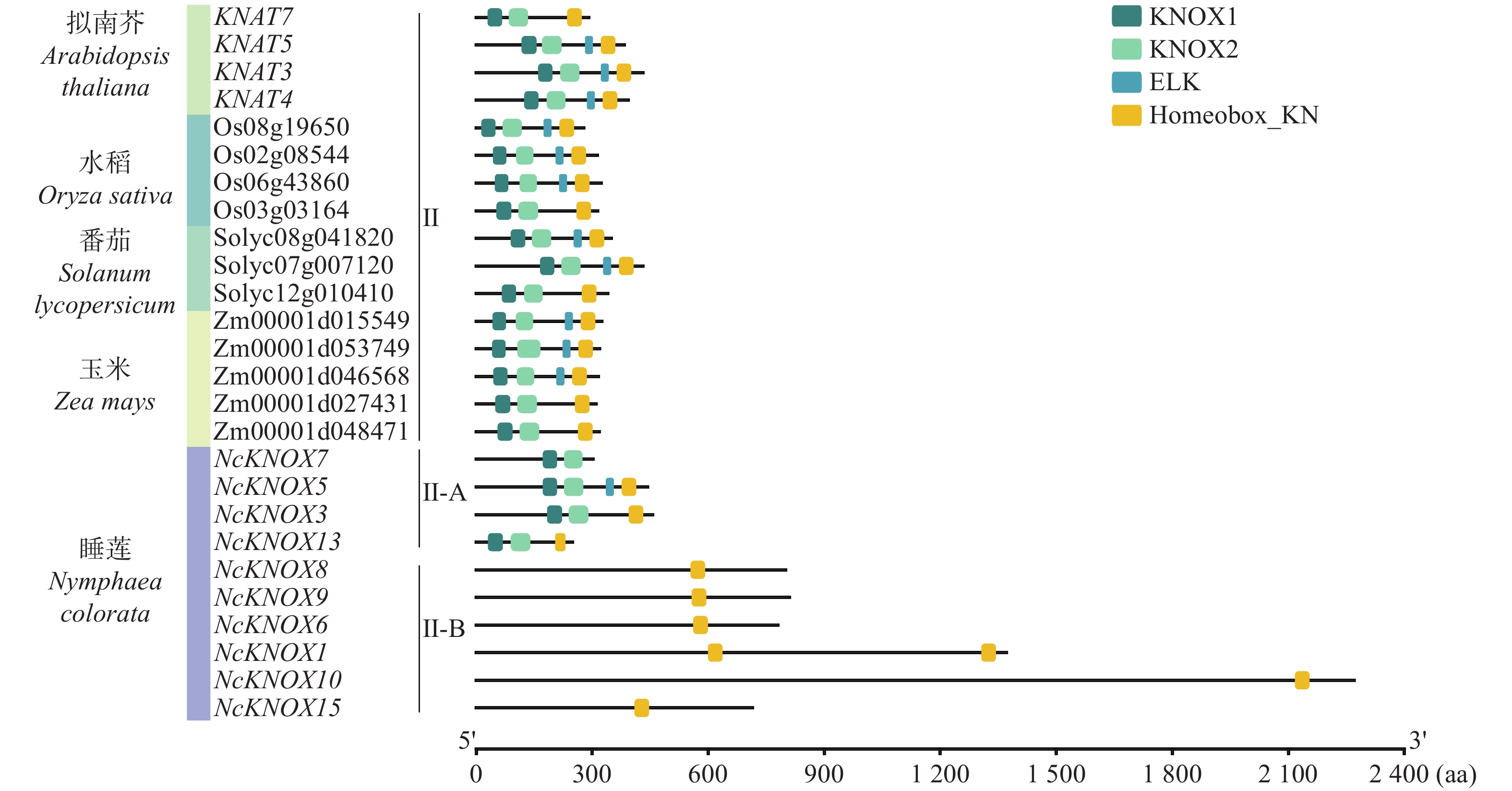
进一步将睡莲与拟南芥、水稻等植物的Class Ⅱ KNOX基因进行比较分析(图5),结果发现,睡莲Class Ⅱ-A基因同其他植物的Class Ⅱ KNOX基因一样,均含有KNOX1和KNOX2结构域,但睡莲Class Ⅱ-B 基因只含有Homeobox_KN。这进一步说明Class Ⅱ-B可能是睡莲KNOX基因家族特有的一个进化分支。
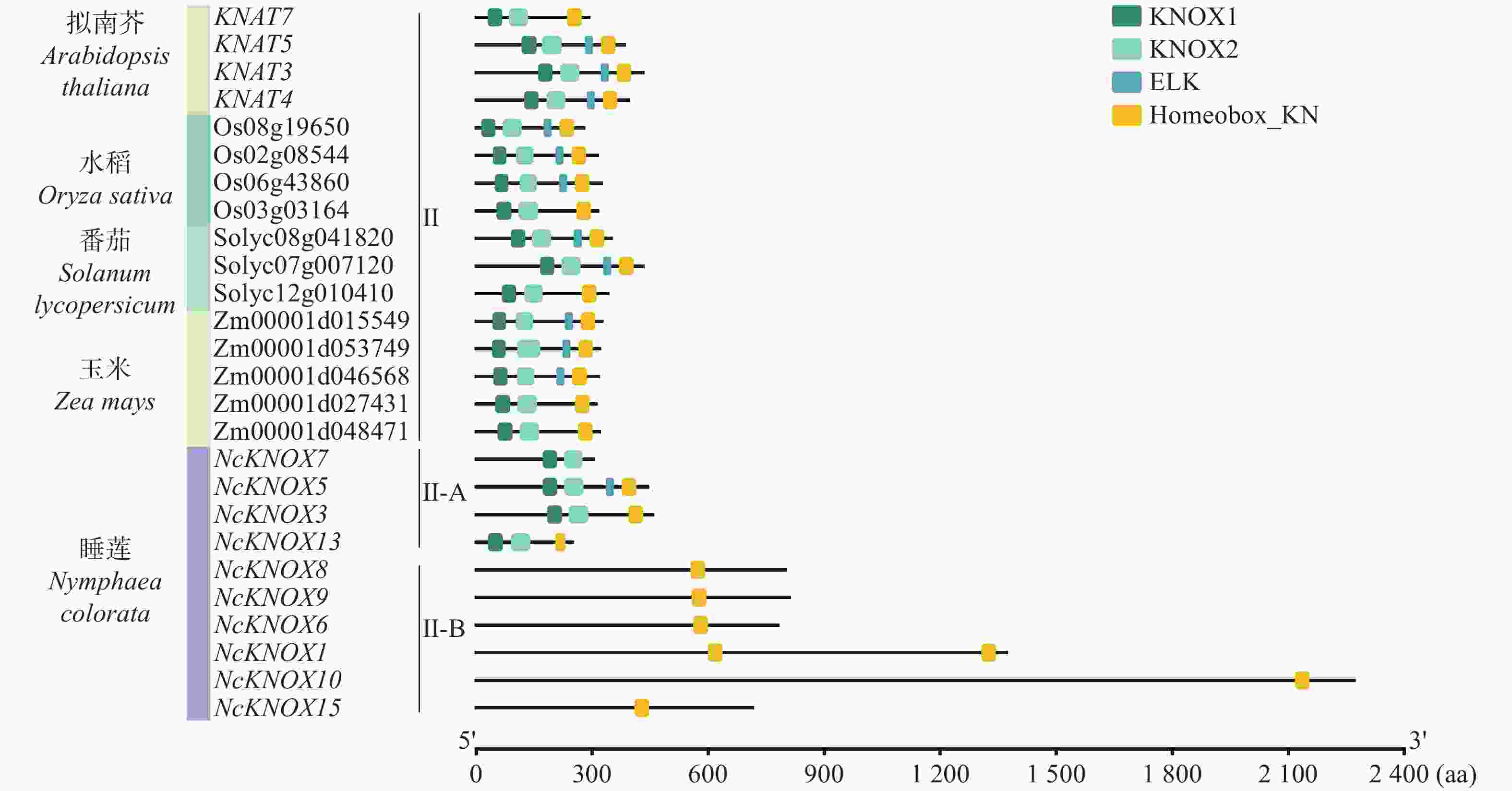
Figure 5. Conserved domain analysis of Class Ⅱ KNOX genes in different species
-
结果表明,睡莲NcKNOX除了真核生物中的基本启动子元件外,还包括植物激素响应(phytohormone response)、植物生长(plant growth and development)、非生物和生物胁迫(abiotic and biotic stress response)和光响应(light response)相关的作用元件。
如图6−A所示,植物激素响应相关的元件有ABRE、AuxRE和AuxRR-core等;植物生长相关的元件有TGA-element、A-box和CCAAT-box等;胁迫反应相关的元件有ARE、GC-motif和LTR等;与光响应相关的元件有MBS、AE-box和Box4等。结合图6−B和图6−C分析,结果发现,在睡莲NcKNOX基因中植物激素响应和光响应元件数量占比更多,其中,NcKNOX3基因较为突出,含有11个ABRE元件(脱落酸响应元件)和13个G-box元件(光响应顺式调节元件)。
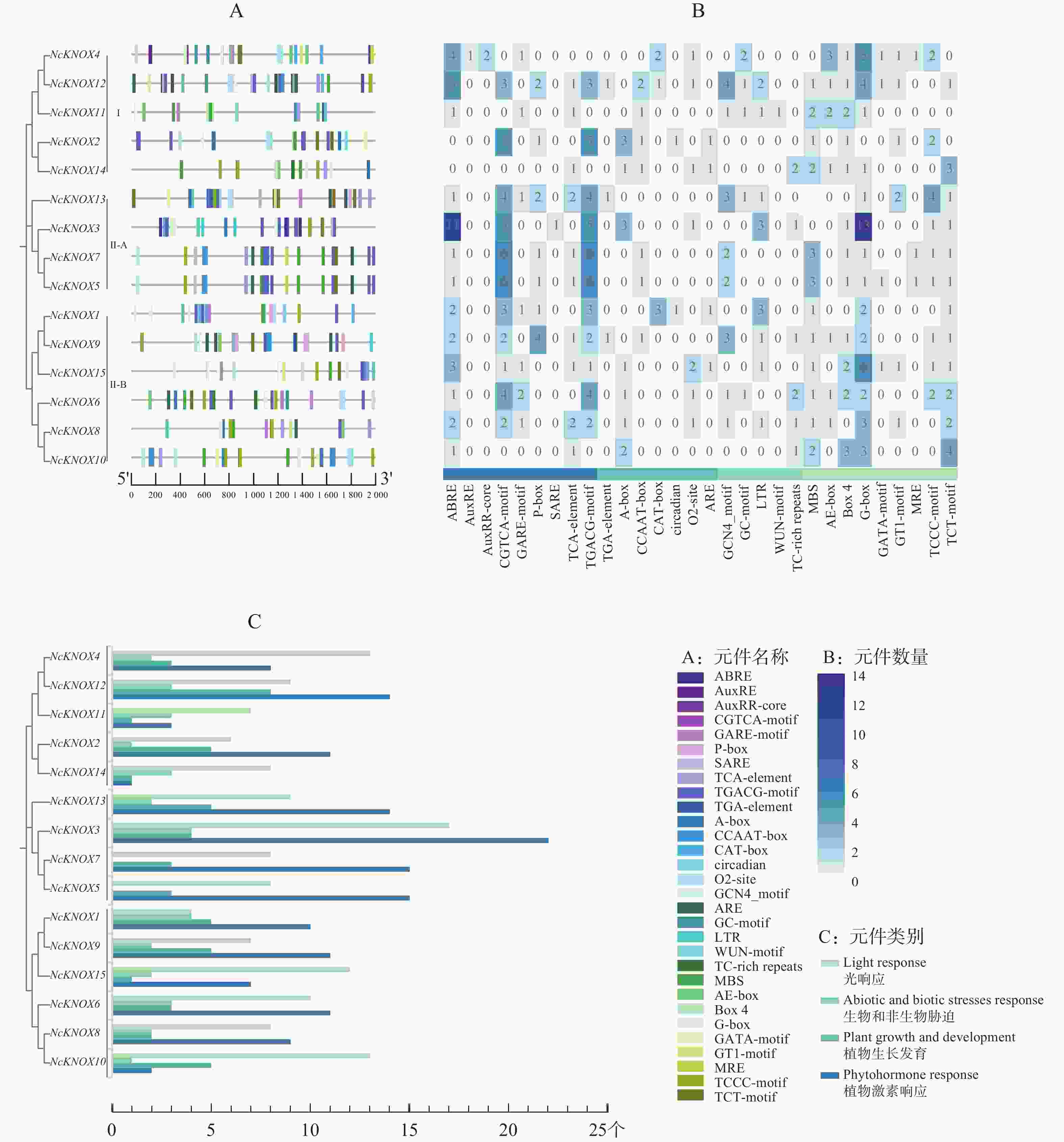
Figure 6. Cis-regulatory element analysis of the promoters of KNOX genes in water lily
-
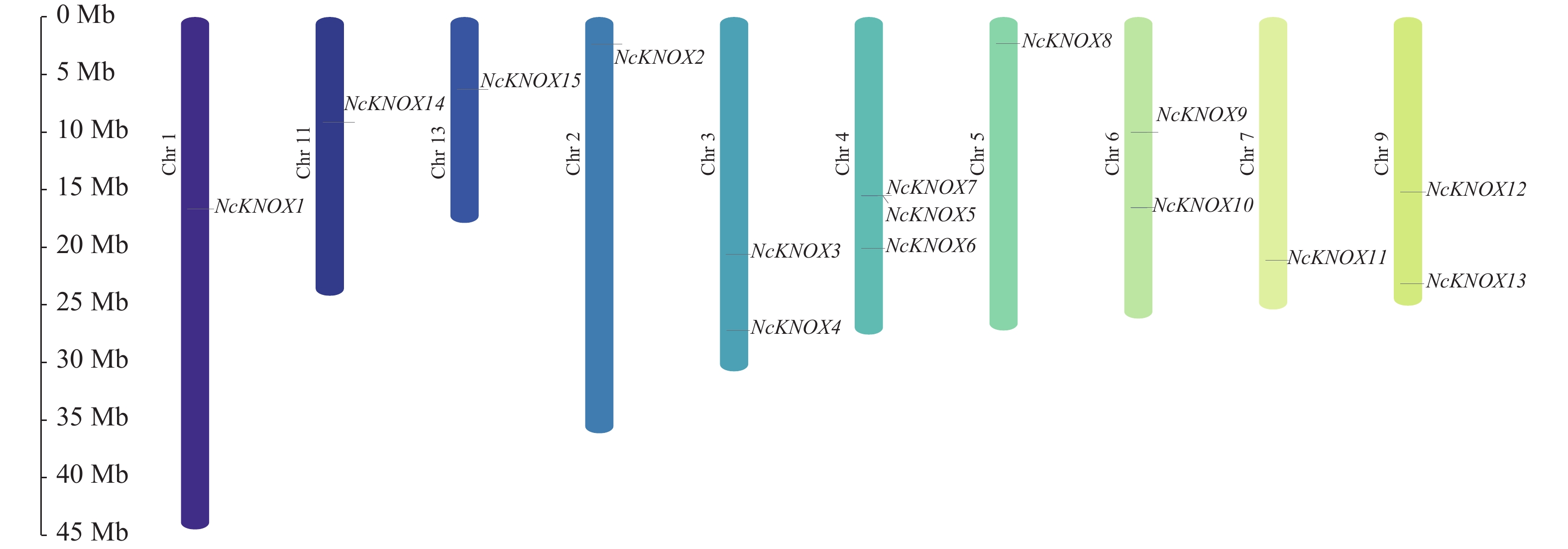
染色体定位结果表明(图7),睡莲NcKNOX基因家族的15个家族成员分布于10条不同染色体上。其中,1号(NcKNOX1)、2号(NcKNOX2)、5号(NcKNOX8)、7号(NcKNOX11)、11号(NcKNOX14)和13号(NcKNOX15)染色体上仅分布1个家族成员,3号(NcKNOX3和NcKNOX4)、6号(NcKNOX9和NcKNOX10)和9号染色体分别有2个家族成员。4号染色体有3个家族成员(NcKNOX5、NcKNOX6和NcKNOX7),其中NcKNOX5与NcKNOX7呈成簇排列,表明两者有着十分紧密的连锁关系。
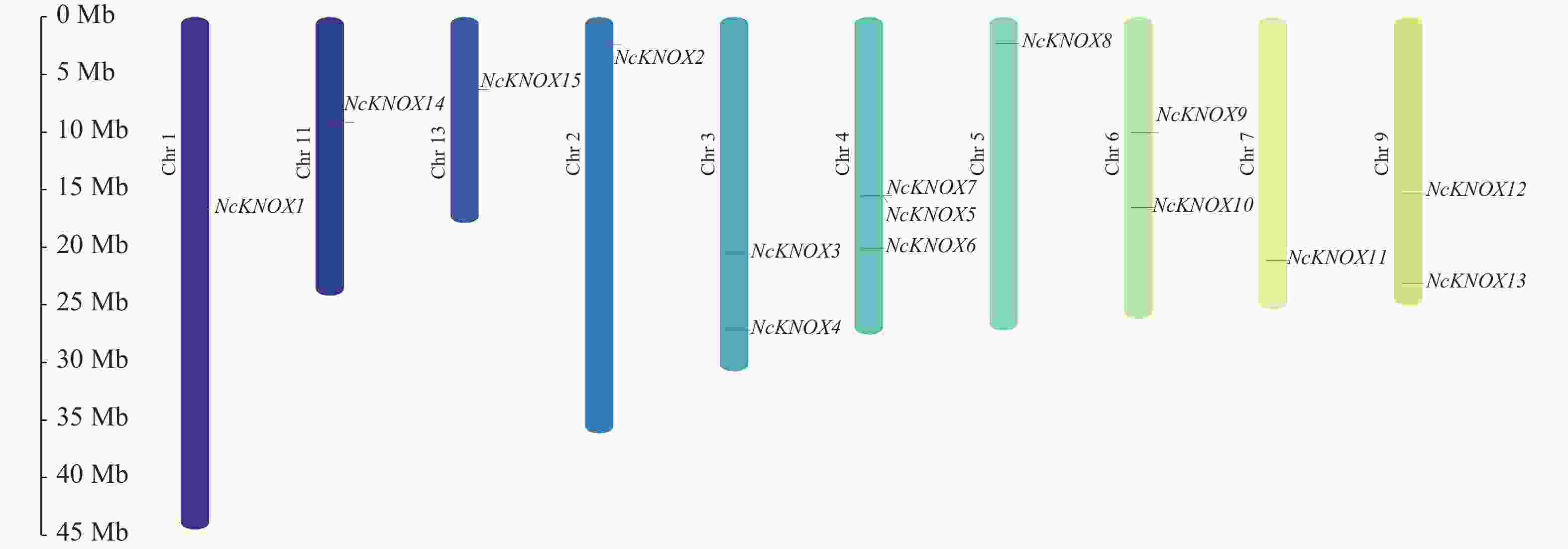
Figure 7. Chromosomal localization analysis of NcKNOX genes in water lily
-
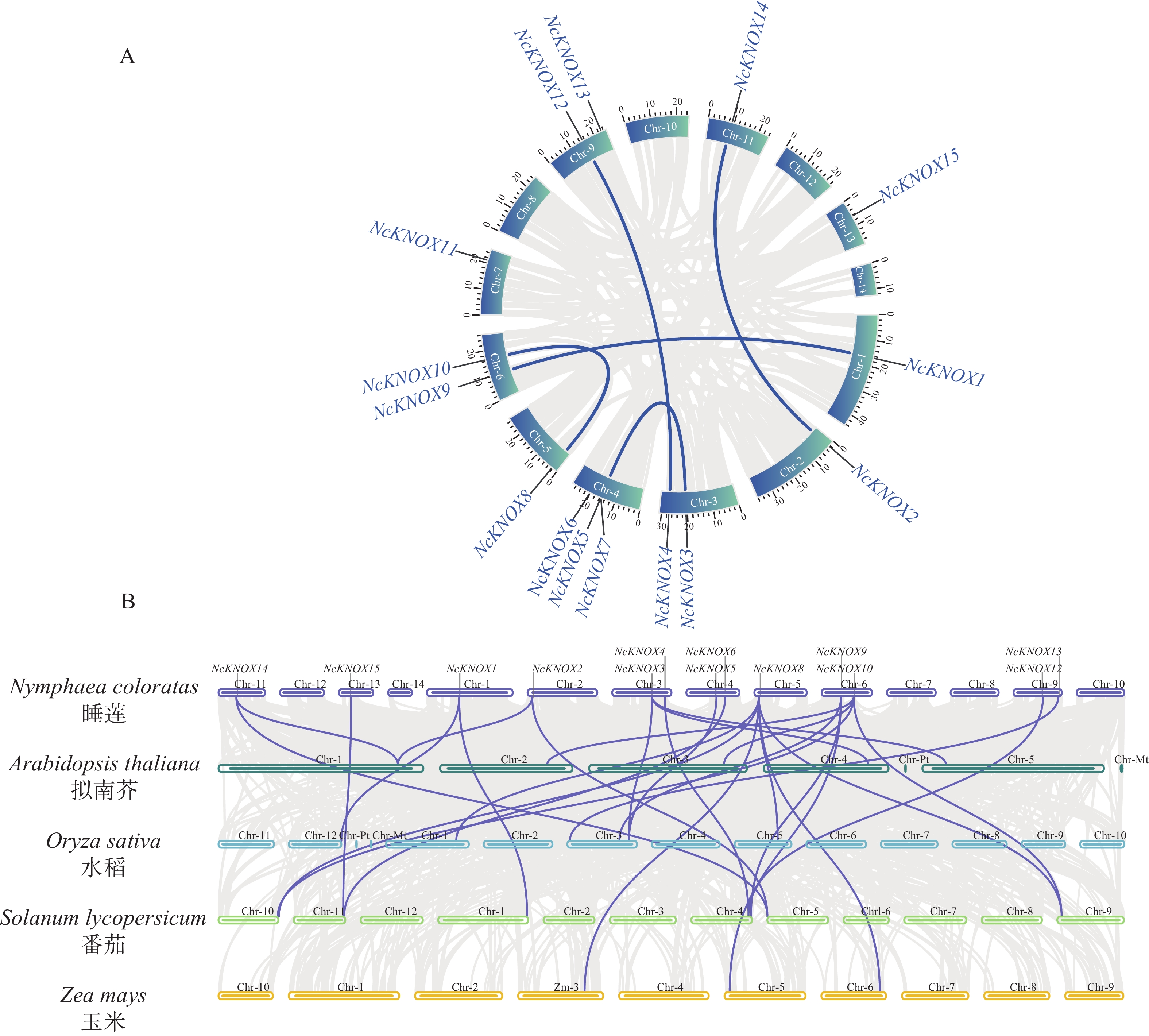
为进一步了解睡莲NcKNOX基因家族进化关系,本研究对其进行种内共线性分析和其他物种的种间共线性分析(图8)。种内共线性分析结果(图8−A)显示,睡莲染色体上存在5组明确的片段复制事件和一组串联重复基因,分别为NcKNOX1与NcKNOX9、NcKNOX2与NcKNOX14、NcKNOX3与NcKNOX5、NcKNOX4与NcKNOX12及NcKNOX8与NcKNOX10之间的复制对,而NcKNOX5和NcKNOX7为串联重复基因。这些发生片段复制的NcKNOX基因对在构建的系统发育树中均归属于同一进化分支,与睡莲KNOX蛋白系统发育树的构建结果相互印证(图3)。
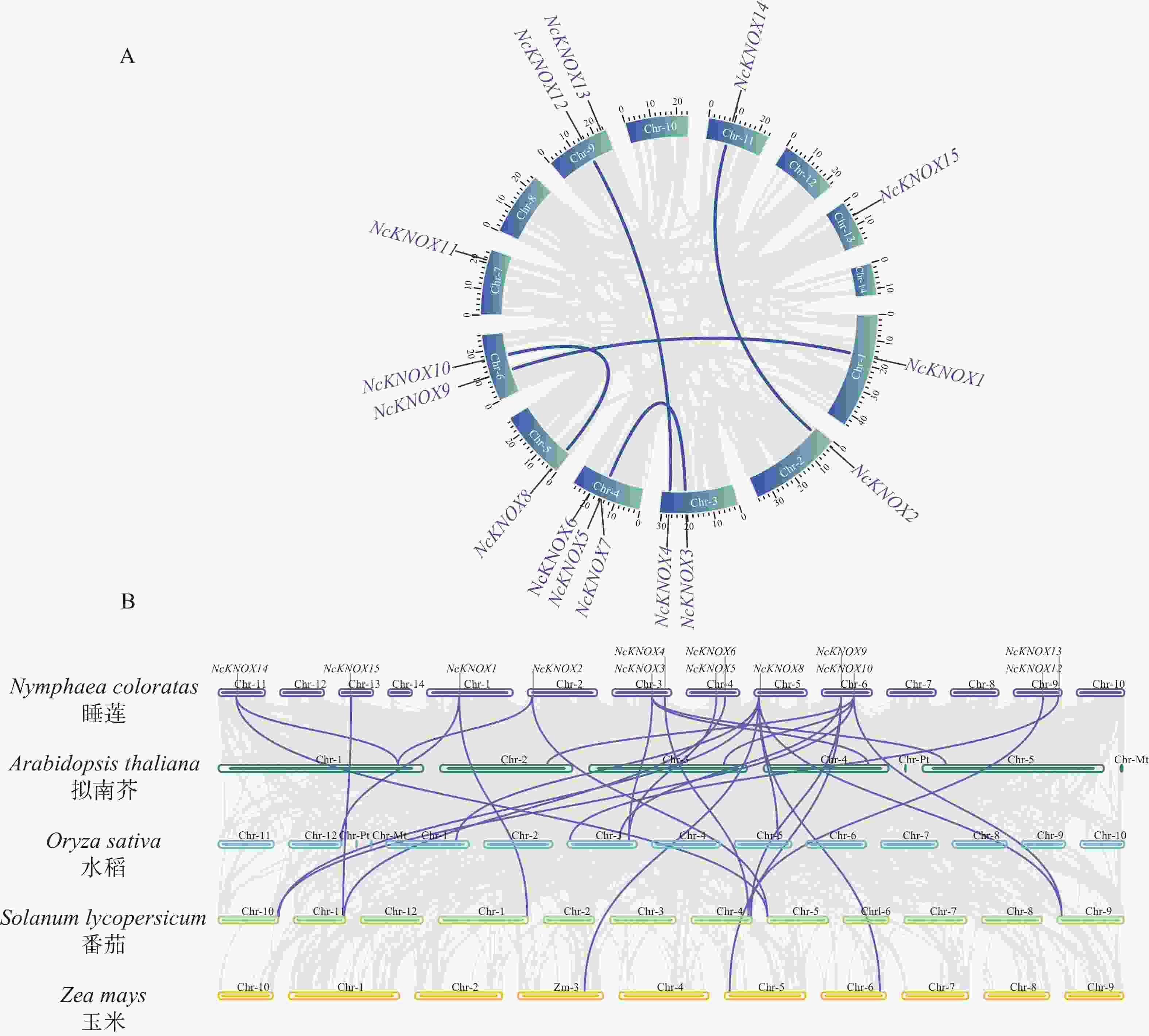
Figure 8. Synteny analysis of KNOX genes
种间共线性分析结果(图8−B)表明,除NcKNOX7和NcKNOX11基因,其余13个NcKNOX基因与拟南芥等模式植物存在一定的共线关系。如表2所示,睡莲分别有4个、6个、10个和2个NcKNOX基因与拟南芥、水稻、番茄和玉米基因存在共线性关系。
基因ID Gene ID Nymphaea colorata
睡莲Arabidopsis thaliana
拟南芥Oryza sativa
水稻Solanum lycopersicum
番茄Zea mays
玉米NcKNOX1 − Os12t0636200 Solyc01g109980 − NcKNOX2 KNAT2 − Solyc05g005090 − NcKNOX3 KNAT4, KNAT5 Os03t0771500 − − NcKNOX4 − − Solyc04g077210 − NcKNOX5 − Os03t0771500 − − NcKNOX6 − − Solyc11g069890 − NcKNOX7 − − − − NcKNOX8 − Os01t0848400
Os05t0455200Solyc09g011380
Solyc10g086640Zm00001eb147970
Zm00001eb289480NcKNOX9 − Os03t0680800 Solyc04g080790 Zm00001eb217350 NcKNOX10 AT2G37900
AT3G53960− Solyc09g011390
Solyc10g084950− NcKNOX11 − − − − NcKNOX12 − − Solyc04g077210 − NcKNOX13 − Os03t0123500 − − NcKNOX14 KNAT2 − Solyc05g005090 − NcKNOX15 − − Solyc11g068950 − Table 2. Summary of NcKNOX genes in water wily with synteny to the genomes of Arabidopsis and other species
-
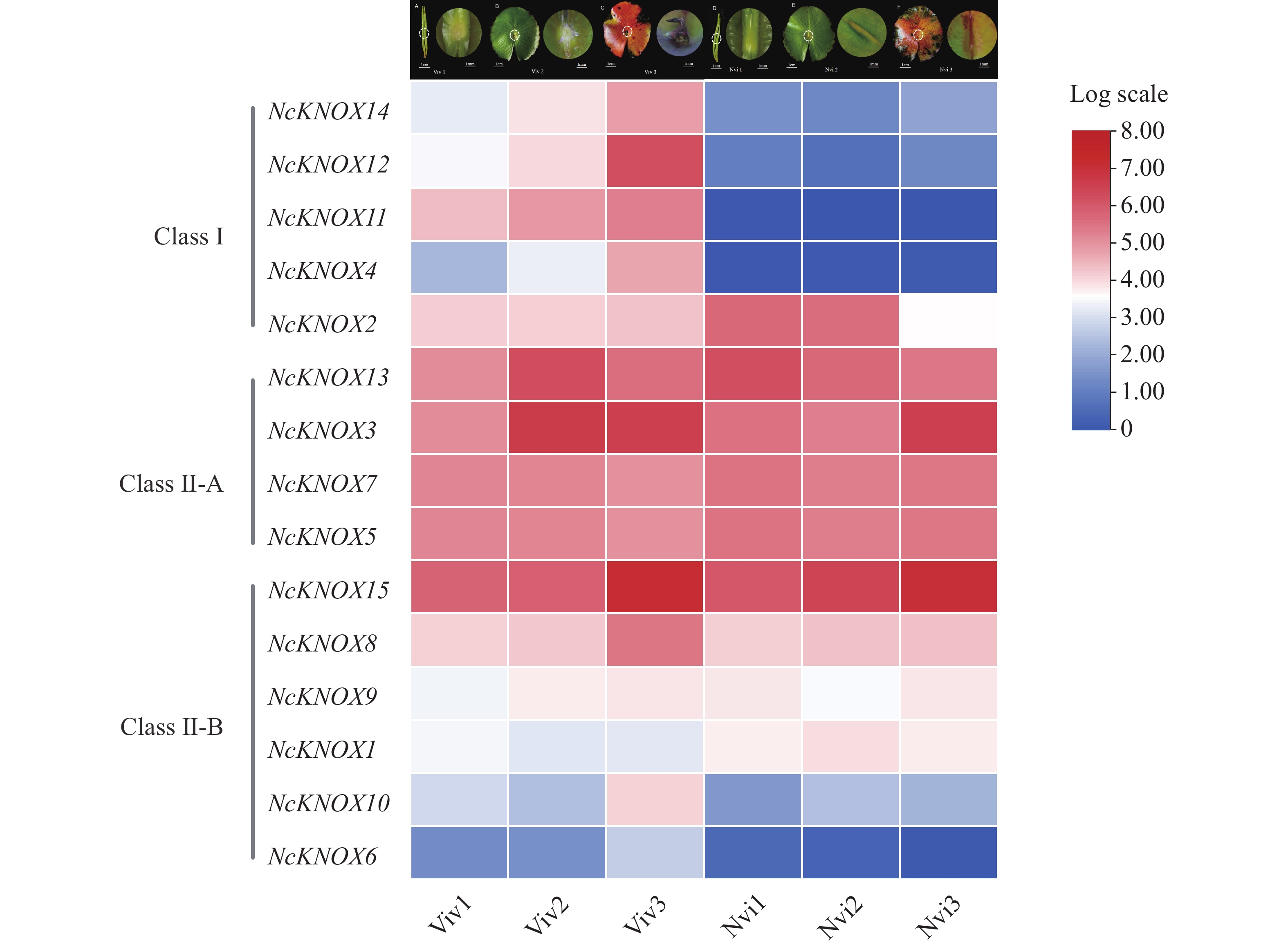
本研究基于蓝鸟睡莲不同发育时期胎生叶片的转录组数据,分析了NcKNOX基因家族成员在不同发育时期叶片的表达模式(图9)。结果显示,Ⅰ类KNOX基因多数在胎生叶片(Viv)时期中高表达,而在非胎生(Nvi)叶片中低表达,如NcKNOX11、NcKNOX12和NcKNOX14等,且这几个基因的表达量均随着胎生苗的发育明显上调;而Ⅱ-A类KNOX基因在所有胎生和非胎生叶片均高表达,如NcKNOX13、NcKNOX7和NcKNOX3等;而Ⅱ-B类KNOX基因中,除了NcKNOX8和NcKNOX15在胎生和非胎生叶片中高表达,其余基因都呈现低表达的趋势。总体上来看,无论是胎生叶片还是非胎生叶片,大多数NcKNOX基因从第1期到第3期叶片的表达量均呈上调的趋势,这就说明NcKNOX在睡莲叶片发育过程中起到重要作用,与拟南芥、番茄的相关研究类似[25 − 26]。
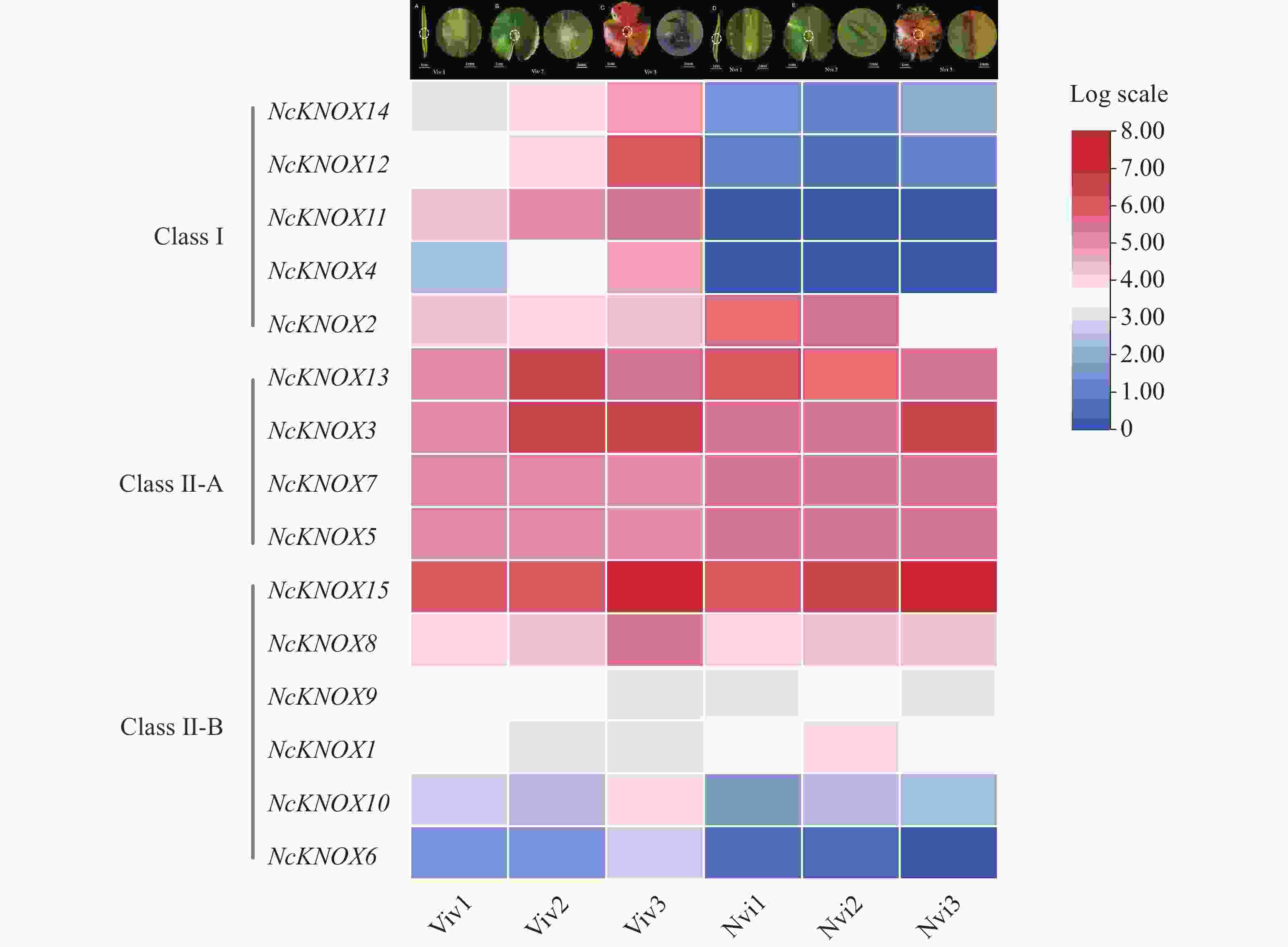
Figure 9. Expression pattern ofNcKNOX genes in viviparous water lily leaves at different developmental stages
-
本研究利用KNOX家族的保守结构域和多种模式植物的KNOX基因序列进行比对分析,在睡莲的基因组中成功鉴定出了15个NcKNOX基因。根据进化关系将睡莲NcKNOX基因分为Ⅰ和Ⅱ类,Ⅱ类亚家族又分为Ⅱ-A和Ⅱ-B类,其中,Ⅱ-B类可能是睡莲中特有的类别,其在KNOX保守结构域上极具特殊性,仅含有Homeobox_KN结构域。根据前人的研究及睡莲NcKNOX的序列保守性分析,Homeobox_KN结构域具有高度保守性[27],使其在复杂的进化过程中稳定存在、不易丢失,这可能是Ⅱ-B类NcKNOX基因仅含有Homeobox_KN结构域的重要因素之一。
KNOX基因普遍含有KNOX1、KNOX2、ELK和Homeobox_KN 4种结构域,但在睡莲NcKNOX基因家族中,ELK结构域仅存在于Ⅰ类NcKNOX基因和Ⅱ-A类的NcKNOX5。在拟南芥中,ELK结构域通过调节STM基因的核定位在其翻译后的调控阶段发挥重要作用[28],说明在睡莲中具有该结构域的Class Ⅰ及Class Ⅱ-A的基因成员可能参与STM转录网络的特殊调控和特异性响应。同时在系统进化树分析中,Ⅱ类亚家族基因NcKNOX3、NcKNOX5和NcKNOX7与番茄的LeT12(Solyc07g007120)、拟南芥的KNAT3和KNAT4在进化关系上较为相近,且LeT12多数在成熟组织中表达[29],KNAT3和KNAT4在花序中表达,调控拟南芥珠被发育[30]。由此推测,Ⅱ类NcKNOX基因多数会在成熟部位表达,可能参与叶片发育后期及衰老过程的调控[25]。
启动子顺式作用元件分析显示,NcKNOX基因家族成员主要受到植物激素和光响应类元件的调控,与金鱼草的研究结果相似[31],可能参与光反应和胁迫反应,还与植物体内的脱落酸、赤霉素等多种激素存在关联。在苹果中,MdKNOX19基因通过调节脱落酸(ABA)的敏感性来抑制种子的萌发和器官的发育过程[32]。在拟南芥中,STM基因可以激活SAM中的细胞分裂素(CK)生物合成的相关元件,促进分生组织的形成[33]。因此,睡莲NcKNOX基因也可能会通过响应不同激素的顺式作用元件,作为激素信号转导途径中的关键节点,调节植物生长发育,参与细胞伸长、分裂和组织分化等过程。同样地,光响应相关的元件在睡莲KNOX基因中数量也较高,说明睡莲KNOX基因也可能通过光响应元件参与调控植物的光合作用、叶片形态和生长方向等光形态建成过程。
基因复制在物种进化中具有重要的推动作用,主要包括全基因组复制/片段复制和串联复制2种机制[34]。染色体定位分析发现,NcKNOX基因家族成员分别分布于10条染色体,可能反映了睡莲进化历史上发生的全基因组复制事件[35],从而产生大量冗余基因,随后通过功能分化或亚功能化适应新环境;4号染色体上NcKNOX5和NcKNOX7成簇排列,则说明存在串联重复,它们可能共享调控元件(如启动子或增强子),形成协同表达模块,在特定发育阶段(如器官边界形成或逆境响应)中发挥叠加或互补作用。此外,15个NcKNOX基因在10条不同染色体上的分布是不均匀的,种内共线性分析结果显示睡莲染色体上存在5组明确的片段复制对和一组串联重复现象,说明小规模重复(small scale duplication, SSD)可能在睡莲KNOX基因家族扩增中发挥关键作用[36]。
从睡莲叶片不同发育时期的NcKNOX基因的表达热图分析得知,Ⅱ类NcKNOX基因的整体表达水平高于Ⅰ类NcKNOX基因,这表明Ⅱ类中的基因可能对睡莲叶片的整个生长周期更为重要。但是多数Ⅰ类KNOX基因在胎生和非胎生叶片的表达量差异较大,表现为在非胎生叶片的表达量较低,在胎生叶片的表达量较高,且随着胎生芽的发育表达量呈明显上升的趋势。在拟南芥中,Ⅰ类KNOX基因如STM、KNAT1、KNAT6对分生组织分化、胚胎发生等过程中发挥关键作用,还可以维持分生组织功能以及塑造叶片形态的多样性[10]。因此,推测睡莲中Ⅰ类KNOX基因可能对胎生叶片中的胎生芽的发生具有重要的意义,其中,NcKNOX4和NcKNOX11的表达差异最大,可能对睡莲叶片胎生现象的产生起到主要作用。
-
本研究基于睡莲基因组数据,通过系统分析鉴定出15个NcKNOX家族成员,分为Class Ⅰ和Class Ⅱ两类,其中,Class Ⅱ-B可能是睡莲独有的类型。睡莲KNOX基因存在大量的CGTCA-motif、AuxRE及ABRE等激素元件,显示其对生长素、脱落酸等关键植物激素具有高度敏感性,基因共线性分析显示其染色体上存在五组明确的片段复制事件。结构域分析表明,睡莲KNOX家族基因可能具有和拟南芥、水稻同源基因类似的功能,在睡莲生长发育调控网络中的核心地位,特别是在响应激素信号、调控分生组织活性以及促进器官分化等关键生物学过程中发挥着不可或缺的作用。睡莲叶片的表达模式表示Ⅰ类和Ⅱ类KNOX基因对睡莲叶片的整个生长周期起到重要作用,其中Ⅰ类KNOX基因可能对睡莲叶片胎生芽的产生和发育起到关键作用。
Identification and bioinformatics analysis of KNOX gene family in water lily (Nymphaea)
DOI: 10.15886/j.cnki.rdswxb.20250024
- Received Date: 2025-01-21
- Accepted Date: 2025-03-21
- Rev Recd Date: 2025-03-20
- Available Online: 2025-09-20
- Publish Date: 2025-11-12
-
Key words:
- Nymphaea /
- KNOX gene family /
- leaf vivipary
Abstract: The KNOX gene family encodes homeobox proteins that function as transcription factors and are integral to regulating plant growth and development. To investigate the bioinformatic characteristics of the KNOX gene family and its potential roles during viviparous seedling development in water lily (Nymphaea), this study identified 15 KNOX genes from the Nymphaea genome using bioinformatics approaches. These genes were distributed across chromosomes 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, and 13. Detailed analyses were performed to assess their physicochemical properties, cis-regulatory elements, and interspecies collinearity. The physicochemical characterization revealed that the KNOX proteins ranged from 249 to
| Citation: | XU Huixian, LI Jie, LI Meier, ZHANG Ruxin, WANG Tongxin, LI Tingge, ZHAO Ying, WANG Jian. Identification and bioinformatics analysis of KNOX gene family in water lily (Nymphaea)[J]. Journal of Tropical Biology, 2025, 16(6): 842-854. doi: 10.15886/j.cnki.rdswxb.20250024

|
 DownLoad:
DownLoad: