
-
巴西橡胶树(Hevea brasiliensis)是热带和亚热带地区的重要经济作物,是重要工业原料天然橡胶的主要来源[1 − 2]。白粉病菌(Erisiphe quercicola)引起的橡胶树白粉病严重影响天然橡胶产量[3 − 4],作为一类专性寄生真菌,还可侵染多种粮食和经济作物。白粉菌在侵染后形成吸器,以获得寄主营养并释放毒性因子效应蛋白[5],对效应蛋白功能的研究有利于阐明白粉菌致病机理,为抗病育种及相关防控工作提供理论依据。
通过分析白粉病菌全基因组或蛋白质组,研究人员从橡胶树白粉菌(Erisiphe quercicola)、大麦白粉菌(Blumeria graminis f. sp. hordei, Bgh)、小麦白粉菌(Blumeria graminis f. sp. tritici, Bgt)、葡萄白粉菌(Erysiphe necator)等病原中预测出一系列候选效应蛋白(Candidate Secreted Effector Proteins,CSEPs)[6 − 8]。这些CSEPs含有N端信号肽、无跨膜结构域,在其他非白粉菌物种中无同源序列[9]。已有研究报道了一些小麦(及大麦)白粉菌效应蛋白功能,例如效应蛋白CSEP0055能够抑制植物病程相关蛋白PR17活性[10],CSEP0264可以抑制病原菌诱导的细胞死亡[11],CSEP0105和CSEP0162与大麦热激蛋白Hsp16.9和Hsp17.5互作[12]。
本研究团队前期在橡胶树白粉菌效应蛋白功能研究上也取得了一定进展,已在该菌中鉴定出133个CSEPs类型的候选效应蛋白[13]。部分效应蛋白的功能已得到了鉴定,发现效应蛋白EqCSEP01276可以干扰脱落酸合成所需的9-顺式-环氧类胡萝卜素双加氧酶(NCED)在叶绿体中聚集,抑制了脱落酸介导的免疫反应[14]。另外,鉴定效应蛋白EqCmu具分支酸变位酶活性,可作用于水杨酸合成前体(分支酸),从而抑制植物中水杨酸介导的免疫反应[15]。但前期一直缺乏橡胶树白粉菌基因研究方法,比如未开发基因沉默的技术,导致这些效应蛋白在致病过程中所扮演的角色尚未被完全了解。但最近研究表明,使用喷施体外合成的双链RNA(double-stranded RNA,dsRNA)分子可以有效诱导基因沉默(spray induced gene silencing,SIGS)[16]。目前,有关橡胶树白粉菌致病分子机理,特别是效应蛋白作用机理还缺少研究。在本研究中,通过利用烟草和拟南芥异源表达待测CSEPs和免疫抑制试验,从两个高保守性的效应蛋白中筛选到效应蛋白CSEP00565。通过致病性测定、侵染过程中表达量分析和信号肽的活性验证,初步分析了该效应蛋白在侵染中的作用机理,为深入解析致病机理提供了依据。
-
橡胶树(Hevea brasiliensis)品种:‘热研7-33-97’(嫁接苗),培养条件为温度22 ℃,光照16 h黑暗8 h;
烟草(Nicotiana benthamiana)品种:本氏烟草,培养条件为温度25 ℃,光照16 h,黑暗8 h;拟南芥(Arabidopsis thaliana)品种:‘哥伦比亚’野生型,培养条件为温度25 ℃,光照16 h,黑暗8 h。
-
橡胶树白粉菌(Erisiphe quercicola)(菌株HO-73),酵母YTK12菌株,由热带农林生物灾害绿色防控教育部重点实验室提供。大肠杆菌(DH5α)、农杆菌(GV3101),购于天根生化科技有限公司。
pBIN-GFP载体,pSUC2载体均由热带农林生物灾害绿色防控教育部重点实验室提供。
-
CMD-W培养基、YPRAA培养基均按照常规方法配制[17]。LB培养基、YPD培养基按常规方法配制[18 − 19]。
Taq Pro Universal SYBR qPCR Master Mix、RNA isolater Total RNA Extraction Reagent、T7 RNAi Transcription Ki试剂盒、DNA纯化试剂盒、同源重组酶、高保真酶均购买自诺唯赞生物公司;质粒提取试剂盒购买自Omage公司;苯胺蓝、3,3′-二氨基联苯胺(DAB)、几丁质类似物(GlcNAc)7购买自生工生物有限公司;其他试剂购自TaKaRa公司及北京索莱宝科技有限公司。
-
使用候选效应蛋白CSEPs的氨基酸序列,在NCBI(www.ncbi.nlm.nih.gov)BlastP在线系统搜索同源蛋白,高于30%相似度即认为具有同源性,并统计含同源蛋白的物种数目。
使用SignalP(
http://www.cbs.dtu.dk/services/SignalP/ )在线系统预测效应蛋白CSEP00565信号肽并确定信号肽序列位置。使用单因素方差分析和Duncan法进行数据分析,确定显著性差异。数据至少来自3个生物学重复。
-
选取橡胶树古铜期叶片,使用毛笔刷接种新鲜橡胶树白粉菌孢子于叶片表面,培养14 d后,收集新鲜的橡胶树白粉菌孢子进行后续实验。使用RNA提取试剂盒提取橡胶树白粉菌HO-73的RNA,反转录合成cDNA。利用Primer 5.0设计引物(表1),以橡胶树白粉菌cDNA为模板,进行目的片段的扩增。在获得目的片段之后,通过同源重组将片段重组到相关表达载体中。使用质粒提取试剂盒进行质粒提取,最终获得重组载体。
引物
Primer引物序列(5′-3′)
Primer sequence(5′-3′)限制性内切酶
Restriction
endonucleaseCSEP00565-F ATGTTTTTCAGTTCTGTCTTC CSEP00565-R TTCGGTTTCTTTGTCAGACTC CSEP01106-F ATGCATTTTATGAAGATATCG CSEP01106-R CATGTTTTGAAGGATCCAACC CSEP04728-F ATGAATACTGTTCCAGTCGAA CSEP04728-R AGCCCCATAAGCTAATAGGAC CSEP01329-F ATGCGTACTTCATTACTCTCG CSEP01329-R CAAAGCCAGTGCTAAACCAAG CSEP02231-F ATGCGGCTCAATATCTTAACT CSEP02231-R CCATATTGCTAAGGCTGCCAT CSEP00565-GFP-F acccccggggtcgacggatccATGTTTTTCAGTTCTGTCTTC BamHI CSEP00565-GFP-R gcccttgctcaccatggatccTTCGGTTTCTTTGTCAGACTC CSEP02231-GFP-F acccccggggtcgacggatccATGCGGCTCAATATCTTAAC BamHI CSEP02231-GFP-R gcccttgctcaccatggatccCCATATTGCTAAGGCTGCCAT CSEP01106-GFP-F acccccggggtcgacggatccATGCATTTTATGAAGATATCG BamHI CSEP01106-GFP-R gcccttgctcaccatggatccCATGTTTTGAAGGATCCAACC CSEP01329-GFP-F acccccggggtcgacggatccATGCGTACTTCATTACTCTC BamHI CSEP01329-GFP-R gcccttgctcaccatggatccCAAAGCCAGTGCTAAACCAAG CSEP04728-GFP-F acccccggggtcgacggatccATGAATACTGTTCCAGTCGAA BamHI CSEP04728-GFP-R gcccttgctcaccatggatccAGCCCCATAAGCTAATAGGAC CSEP00565Δsp-GFP-F acccccggggtcgacggatccATGATTCCACTTAATATCAAC BamHI CSEP00565nbsp-GFP-F cccggggtcgacggatccATGAATTTTACTGGCTATTCTCGA
TTTTTAATCGTCTTTGTAGCTCTTGTAGGTGCTTTT
TTCAGTTCTGTCBamHI CSEP00565sp-pSUC2-F CGGAATTCCGATGTTTTTCAGTTCTGTC EcoRI CSEP00565sp-pSUC2-R CCCTCGAGGGTGAGATTTCACCATCACC XhoI CSEP00565-Pet28a-F CGGGATCCCGATGTTTTTCAGTTCTGTC BamHI CSEP00565-Pet28a-R CGGAATTCCGTTCGGTTTCTTTGTCAGA EcoRI Pet28a-F TAATACGACTCACTATAGGG Pet28a-R GCTAGTTATTGCTCAGCGG dsRNA-CSEP00565-F GATCACTAATACGACTCACTATAGGGATGAGAGGGAAG GAAGAGGTA dsRNA-CSEP00565-R TAATACGACTCACTATAGGGTGTCAATGAATCAGAGGC dsRNA-GFP-F GATCACTAATACGACTCACTATAGGGATGCAGTGCTTC AGCCGCTAC dsRNA-GFP-R TAATACGACTCACTATAGGGTTCACCTTGATGCCGTTC CSEP00565-RT-F GGGAAGATTATTACTCCAA CSEP00565-RT-R TTCACCCTTACACCAACA GFP-RT-F TGGGGCACAAGCTGGAGT GFP-RT-R TCTGCTGGTAGTGGTCGG EF1a-RT-F GAACCTTCATCTAACTGC EF1a-RT-R GTCGTAGTGGTTTGTCAG AtAct2-RT-F ACTACGAGCAGGAGATGGAA AtAct2-RT-R TTAATCTTCATGCTGCTTGG CSEP00565-MCS-F GAGCTCGGTACCCGGGGATCCATGTTTTTCAGTTCTGTCTTC BamHI CSEP00565-MCS-R CATGTCGACTCTAGAGGATCCTTCGGTTTCTTTGTCAGACTC 注:小写字母表示用于融合线性载体的序列。 Note: The sequences used in the fusion with linearized vectors were marked by lowercase letters. Table 1. Primers used in the experiment
-
利用限制性内切酶BamHI对pBIN-GFP载体进行单酶切,并使用DNA纯化试剂盒回收载体。用表1中的5个效应蛋白引物扩增目的片段,并将目的片段与酶切后的载体通过同源重组连接并转入大肠杆菌DH5α中,最终获得重组的表达载体质粒,将构建好的载体导入农杆菌GV3101中。将长出的单菌落转移至新的固体培养基,并进行PCR验证,获得阳性转化子。将阳性转化子加入质量浓度为50 mg·L−1利福平和50 mg·L−1硫酸卡那霉素的LB培养基中培养过夜。离心(3 500 r·min−1,2 min)收集菌体,用1mL 10 mmol·L−1氯化镁重悬。悬浮液浓度被调整到OD600值为0.5[20]。将含有效应蛋白载体的农杆菌用针孔注射器注入烟草叶片,24 h后,共表达含有致病疫霉激发子(INF1)基因的农杆菌,3~4 d后,观察注射区域的坏死情况。INF1具有诱导烟草叶片产生免疫反应过敏性坏死(Hypersensitive response)的作用[21 − 22]。
-
将1.5.2构建完成的CSEP00565-GFP阳性转化子在含有50 mg·L−1利福平和50 mg·L−1硫酸卡那霉素的LB培养基中28 ℃培养48 h。离心(3 500 r·min−1,2 min)收集菌体,使用悬浮液(菌体悬浮包含50 g·L−1 Sucrose、MS Salt 2.2 g·L−1和MES 0.5 g·L−1)对菌体进行悬浮,并调整OD600值为0.8~1.0。对培养四周的拟南芥进行果荚修剪,然后将其上部浸泡在菌体悬浮液中持续5 min,用保鲜膜将拟南芥的花序包裹,在黑暗环境中进行48 h的保湿培养,然后正常培育拟南芥并收集其种子[23]。
-
烟草或拟南芥在接种农杆菌48 h后,将注射叶片剪下放于脱色液(V乙醇∶V乙酸=3∶1)中脱色至透明,将脱色叶片表面脱色液洗净,使用0.01%苯胺蓝溶液(苯胺蓝溶于0.067 mol·L−1K2HPO4)进行避光染色4 h,通过荧光显微镜检测胼胝质积累情况,使用ImageJ软件在1 mm2范围内计算胼胝质积累情况[19]。
-
使用T7 RNAi Transcription Kit试剂盒合成dsRNA,以pBin-GFP质粒中的GFP基因为模板,扩增 GFP-dsRNA;使用含有CSEP00565编码序列的质粒,扩增CSEP00565-dsRNA。设计用于dsRNA扩增的正向和反向引物,5′端为T7启动子序列(表1)。
1)dsRNA的合成
① 根据T7试剂盒说明书配置反应体系
② PCR仪里37 ℃反应2 h
③ 37 ℃反应结束后,当双链长度超过800 bp,需经过72 ℃反应10 min,再自然冷却退火形成dsRNA
④ 用RNase T1 Dilution Buffer将100 μmol·min−1 RNase T1稀释成10 μmol·min−1。
⑤ 配制双酶消化体系
⑥ 37 ℃孵育30 min,电泳检测转录产物。
2)产物纯化
按照T7试剂盒说明书,将试剂RNA Clean Beads加入RNA转录产物中,置于磁力架上弃去上清。干燥后,加入无酶水,获得纯化后的产物,保存于−20 ℃。
-
使用dsRNA处理橡胶树白粉菌:选择生长周期为10 d左右的橡胶树叶片,进行白粉孢子悬浮液的接种(1×105个·mL−1)后,将dsRNA用无酶水稀释,调整至最终质量浓度为20 μg·L−1,喷洒于叶片表面接种孢子处,将处理后的幼苗在24 ℃、每天12 h光照,10 h黑暗的室内。3 d后对吸器数量统计分析,7 d后观察发病情况并对病斑面积进行统计分析。
吸器数量:使用ImageJ软件对每1 mm2的吸器数量计数,数据来自3次生物重复。
病斑面积:使用ImageJ软件对叶片上病斑面积进行计算,数据来自3次生物重复。
-
将野生型橡胶树白粉菌接种在叶龄基本一致的古铜期叶片上,并在接种后4、12、24、48 h对叶片进行样本采集。用RNA isolater Total RNA Extraction Reagent试剂盒提取总RNA。荧光定量PCR(quantitative real-time PCR,qRt-PCR)采用SYBR GreenⅠ荧光染料法进行检测,PCR扩增引物如表1所示。以EF1a作为内参基因。每个样本进行3次独立重复。反应完成后,使用2−ΔΔCT法分析CSEP00565基因在橡胶树白粉菌侵染橡胶树不同阶段的相对表达水平。
-
将CSEP00565的信号肽序列构建到含有色氨酸合成基因但缺失蔗糖转化酶基因的载体pSUC2中,将构建好的载体转化至酵母菌株YTK12中[24]并涂布于CMD-W培养基上,挑取阳性克隆在YPRAA培养基上观察菌落生长情况。
通过将2,3,5-三苯基氯化四氮唑(2,3,5-Triphenyltetrazolium chloride, TTC)还原为不溶的红色化合物,检测信号肽分泌活性。挑取转化成功的酵母转化子于CMD-W液体培养基中培养,将YTK12和转入pSUC2空载体的YTK12酵母菌株加入YPD液体培养基中,28 ℃培养48 h。12 000 r·min−1收集菌体,ddH2O清洗2次,用750 μL的ddH2O对菌体进行重悬,加入500 μL的10%(质量分数)蔗糖溶液和250 μL的醋酸-醋酸钠缓冲液,在37 ℃孵育10 min。12 000 r·min−1离心收集上清液,放入含有0.1% TTC溶液的试管中。室温下静置15 min,以观察其颜色变化情况。
-
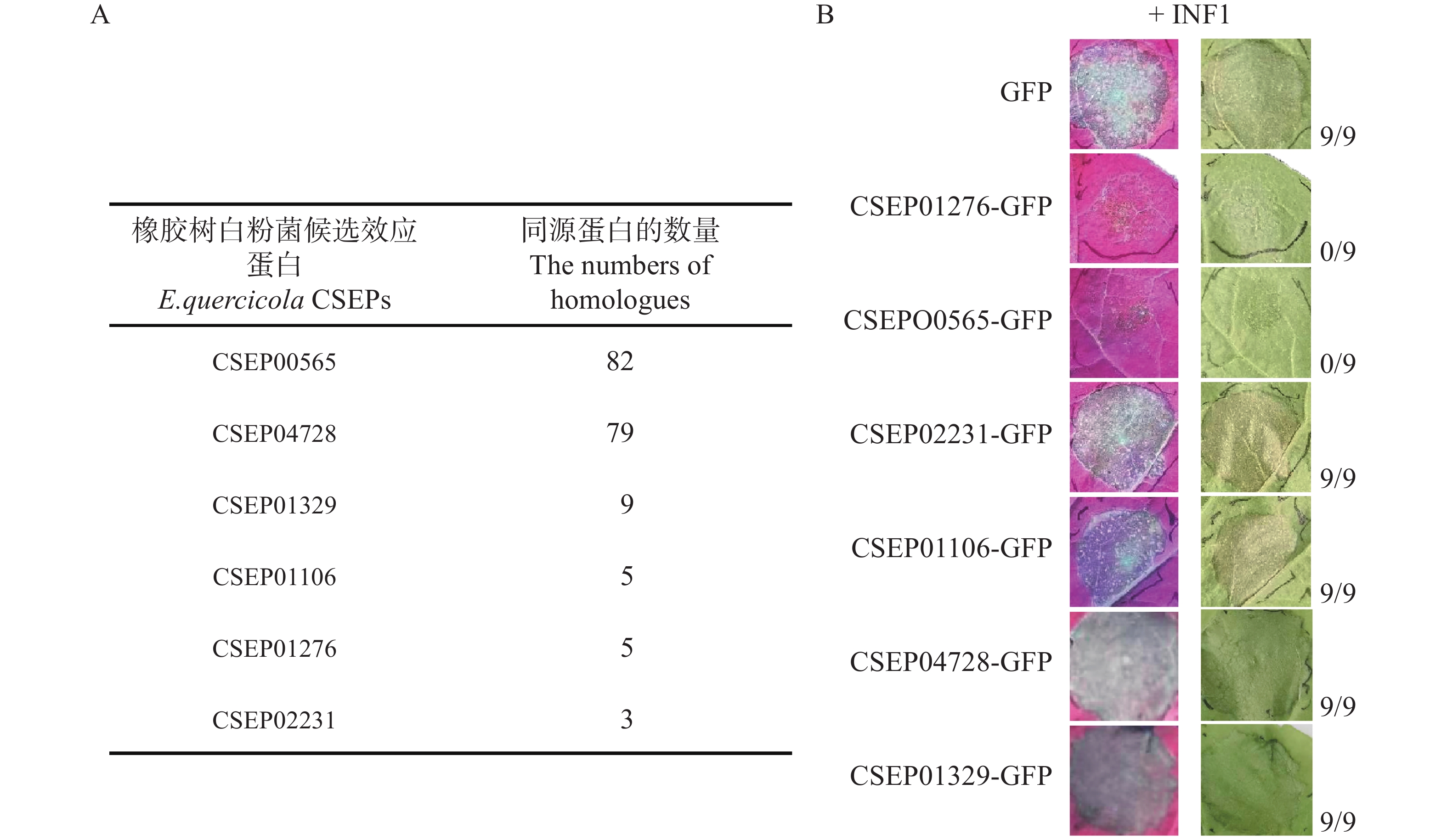
在很多种白粉菌中都具有同源的保守效应蛋白,可能在致病过程中扮演重要角色,因而遗传中被保留下来。因此,通过将橡胶树白粉菌CSEPs效应蛋白氨基酸序列与其他白粉菌的进行对比,找到了该菌中的24个高保守性的蛋白(在75个及以上的白粉菌菌种中存在同源蛋白)。本次研究选取其中2个蛋白CSEP00565和CSEP04728在本氏烟草上表达和测定抑制植物过敏性反应的功能,并同时选取4个低保守的CSEPs(低于10个白粉菌种中存在同源蛋白)作为对照(图1−A)。通过农杆菌介导转化在烟草叶片细胞表达待测效应蛋白(GFP标签),同时共表达卵菌激发子INF1[21 − 22]。并于3~4 d后检测叶片状况,发现CSEP00565抑制了INF1诱导的过敏性坏死(图1−B)。低保守性的CSEP01276[14]也产生了同样的效果,与前期报道一致。因此,通过该烟草上的功能测定,筛选到了潜在的致病因子CSEP00565。
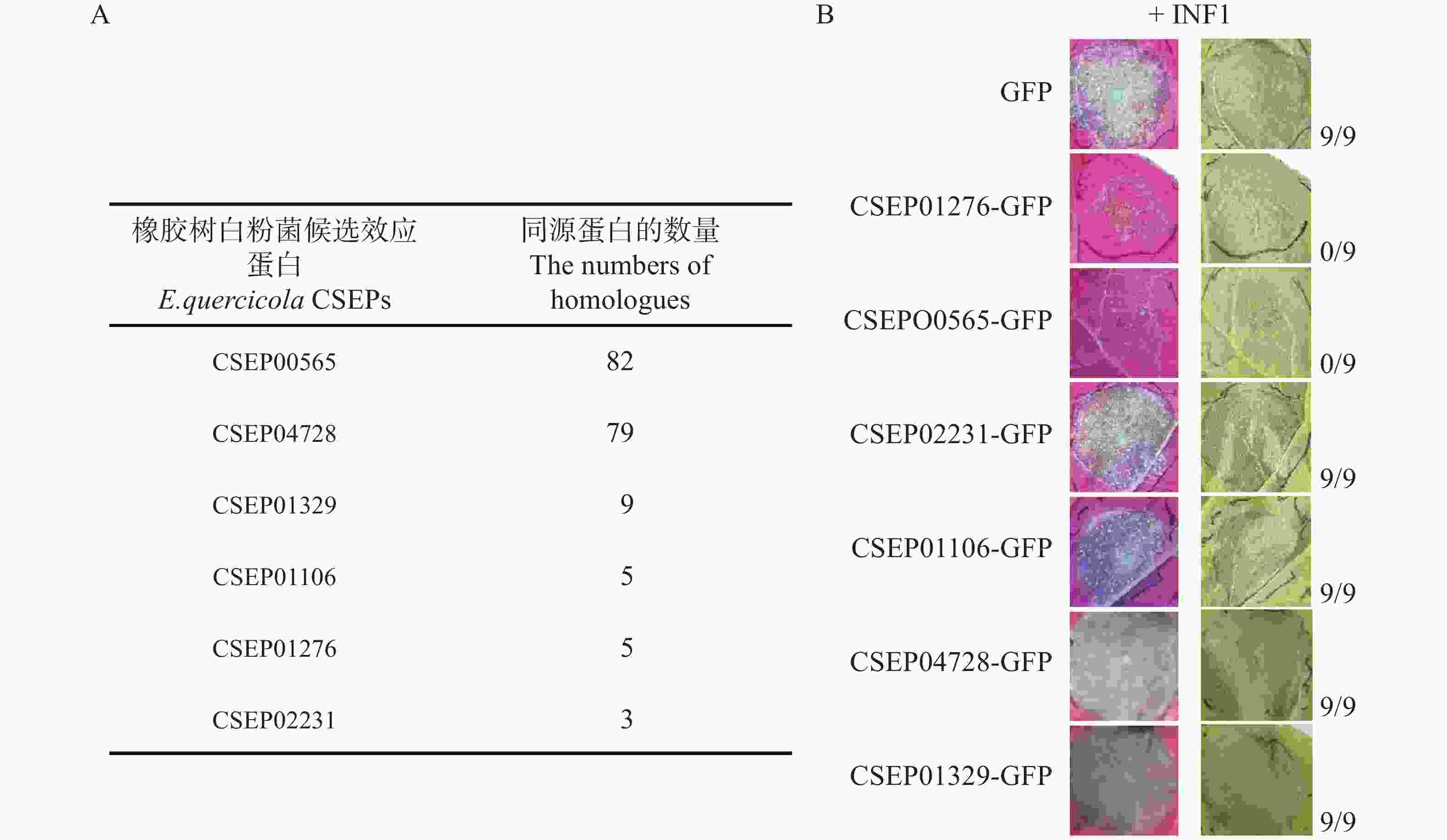
Figure 1. The activity of E. quercicola CSEPs in suppressing INF1-induced hypersensitive response was tested in Nicotiana benthamiana
-
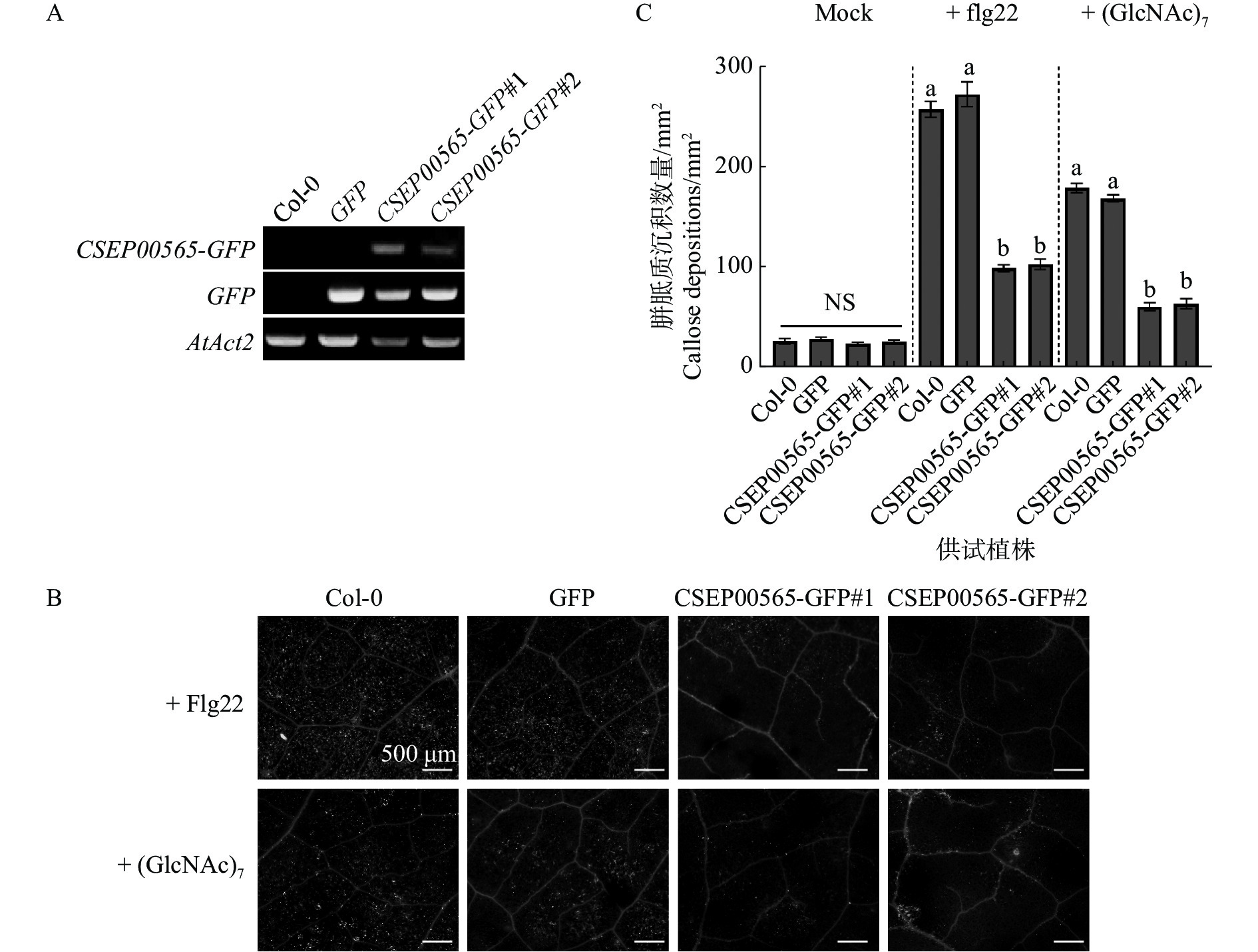
为了确认CSEP00565具有植物免疫反应抑制的功能,利用农杆菌转化获得稳定CSEP00565转基因拟南芥植株,并测试CSEP00565对病原物相关分子模式(pathogen-associated molecular patterns)诱导免疫是否具有抑制作用。通过qRT-PCR分析验证了植物中转化基因的表达(图2−A)。病原物相关分子模式如细菌鞭毛蛋白(flagellin)和真菌几丁质(chitin)[25 − 26]可触发拟南芥产生胼胝质沉积,使用20 μmol·L−1的几丁质单体(GlcNAc)7和flagellin肽段flg22处理拟南芥叶片,观察不同植株胼胝质的沉积情况。试验结果表明,与对照组野生型植株Col-0及阴性对照GFP转基因植株相比,CSEP00565-GFP转基因植株(CSEP00565-GFP#1、CSEP00565-GFP#2)叶片中的胼胝质数量下降了60%~64%,以上结果表明CSEP00565可抑制拟南芥产生的免疫反应(图2−B,C)。
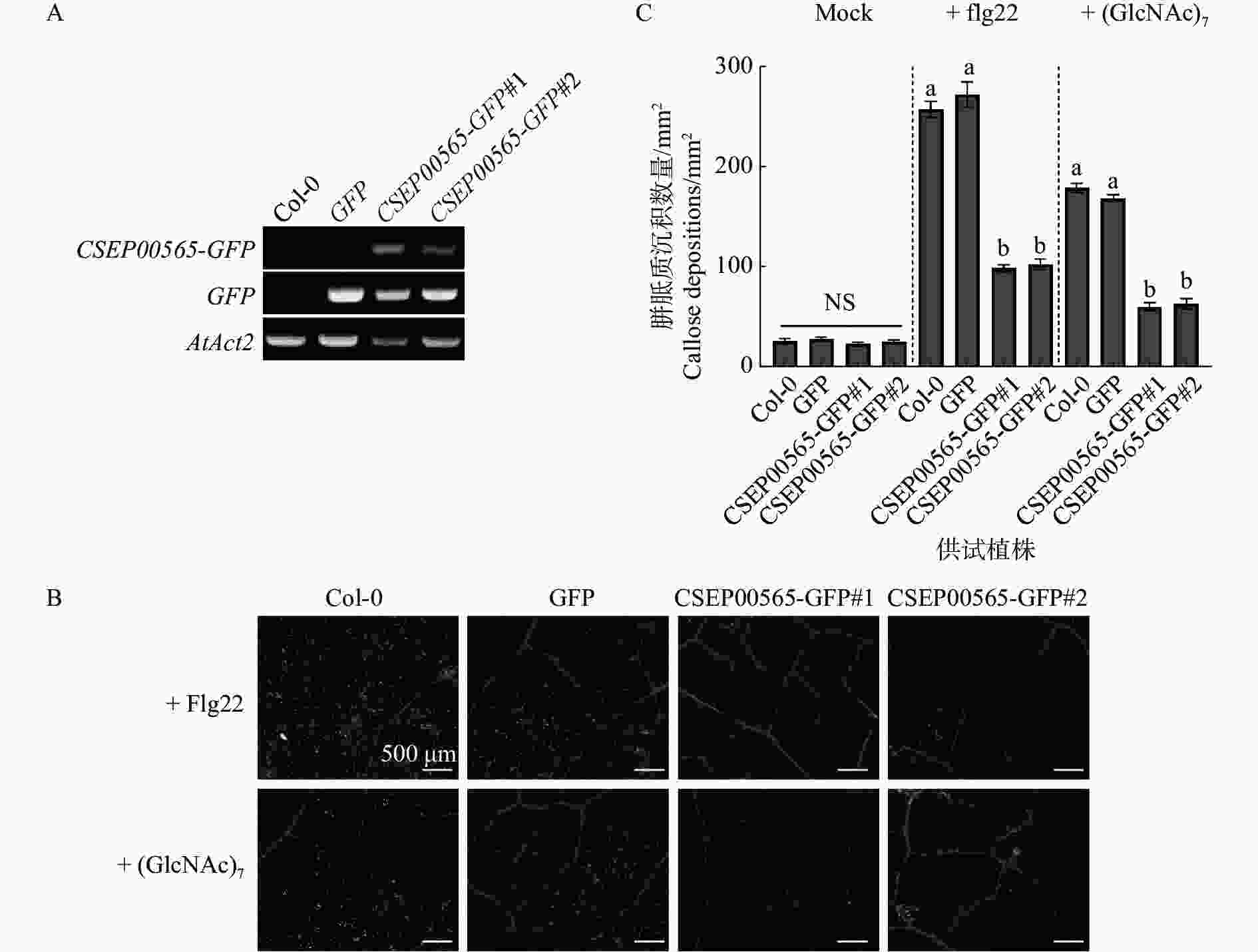
Figure 2. The CESP00565 expression inhibited callose deposition in Arabidopsis thaliana Col-0 plant
-
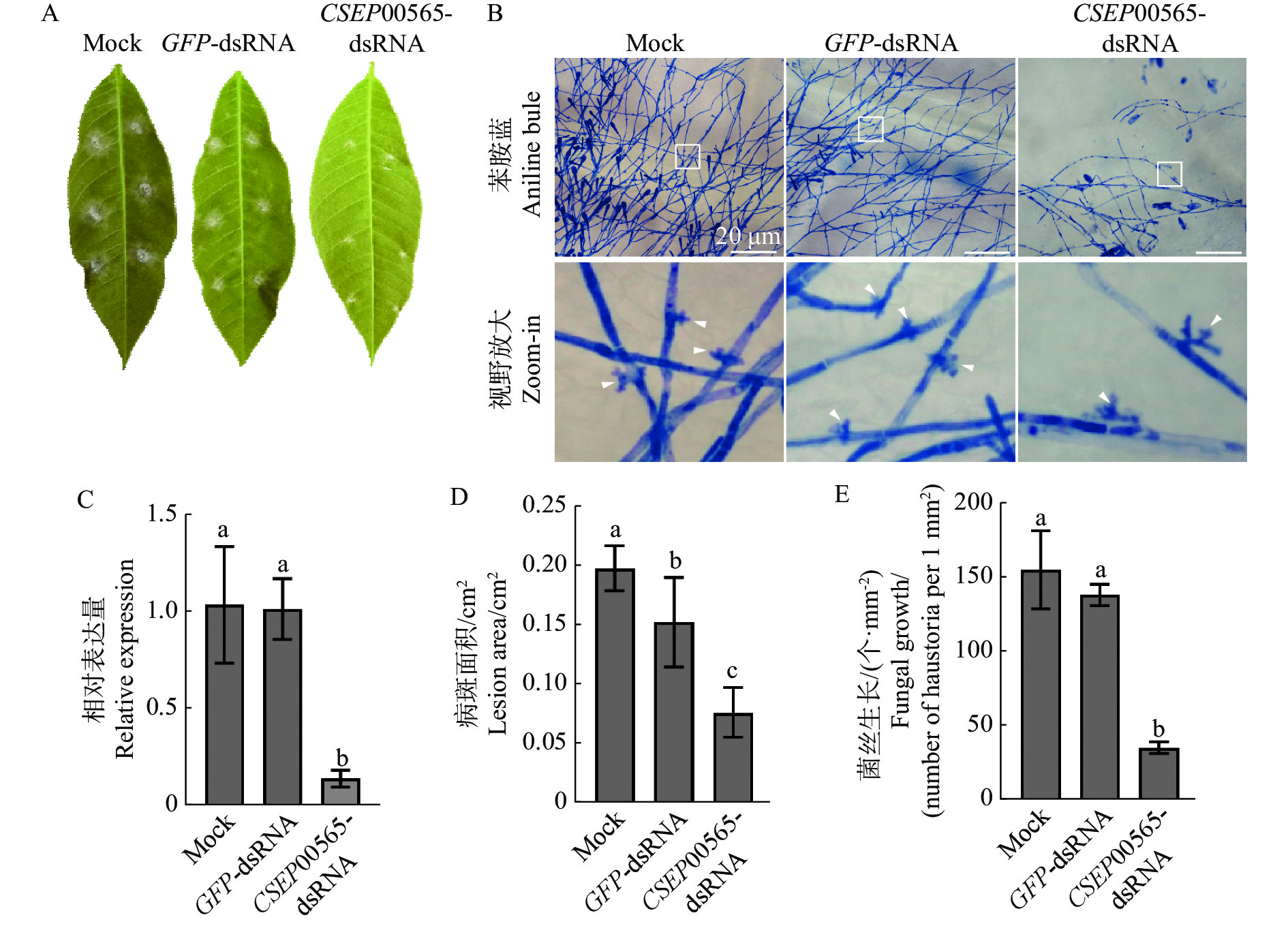
为了证明CSEP00565是致病关键的,将通过SIGS诱导橡胶树白粉菌中靶基因沉默并测定菌株致病性。以CSEP00565和GFP的编码序列为模板,利用T7启动子进行体外转录,合成质量浓度为500 ng·μL−1的CSEP00565-dsRNA和GFP-dsRNA产物。在古铜期橡胶树叶片表面接种孢子悬浮液(1×105个·mL−1)后,使用dsRNA(100 ng·μL−1)溶液进行喷洒处理。3 d后,和对照(无酶水Mock)、GFP-dsRNA处理相比,CSEP00565P-dsRNA处理导致吸器数量减少了82.7%(图3−C,E);7 d后,CSEP00565P-dsRNA处理导致病斑面积降低了58.1%(图3−A,D),qRT-PCR检测发现,CSEP00565-dsRNA处理后的叶片中CSEP00565基因表达量下调约87.0%(图3−B)。结果表明CSEP00565沉默病菌致病力显著下降。
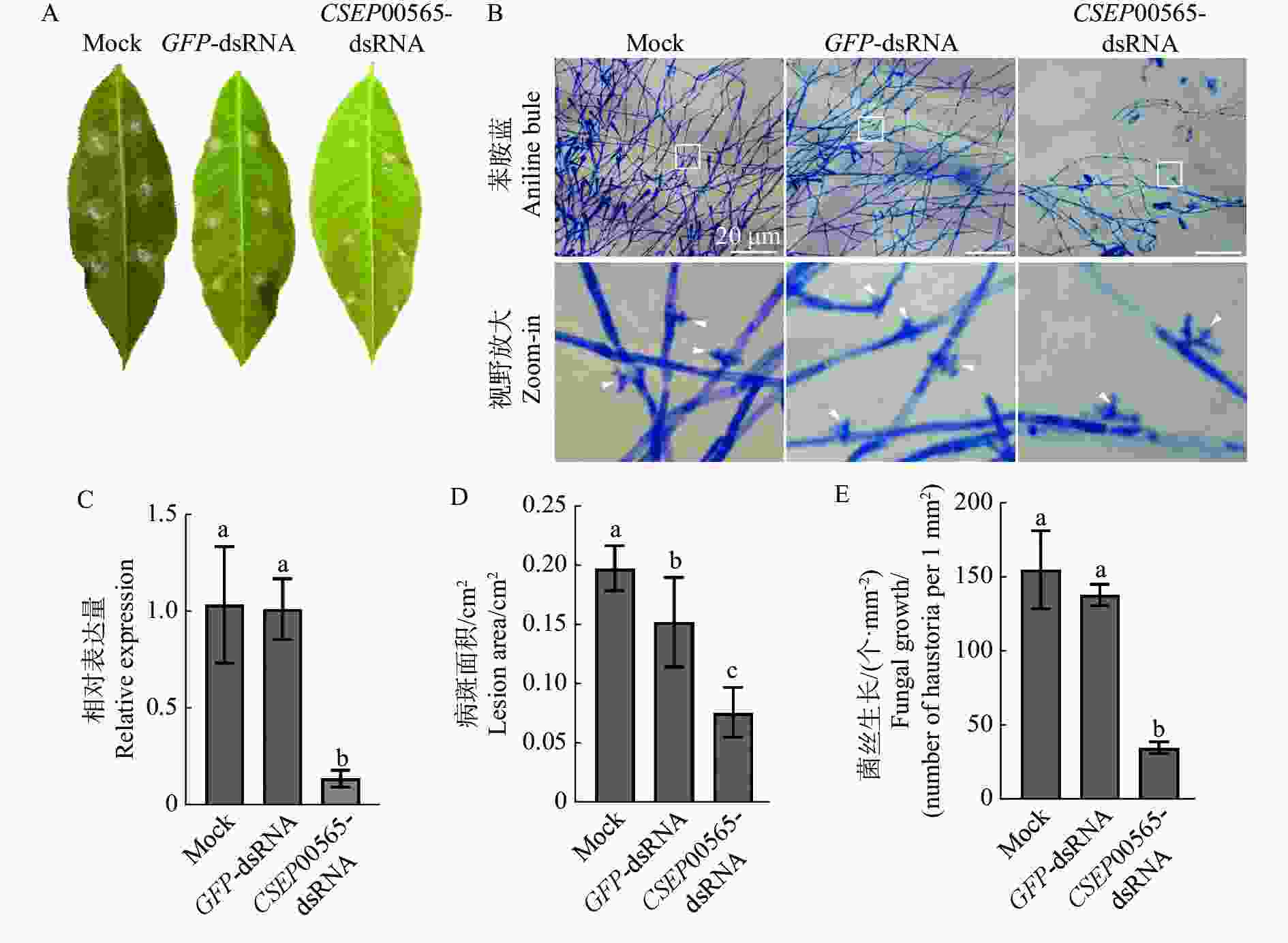
Figure 3. CSEP00565 silencing inhibited hyphal extension after the pathogen of the powdery mildew was penetrated into host
-
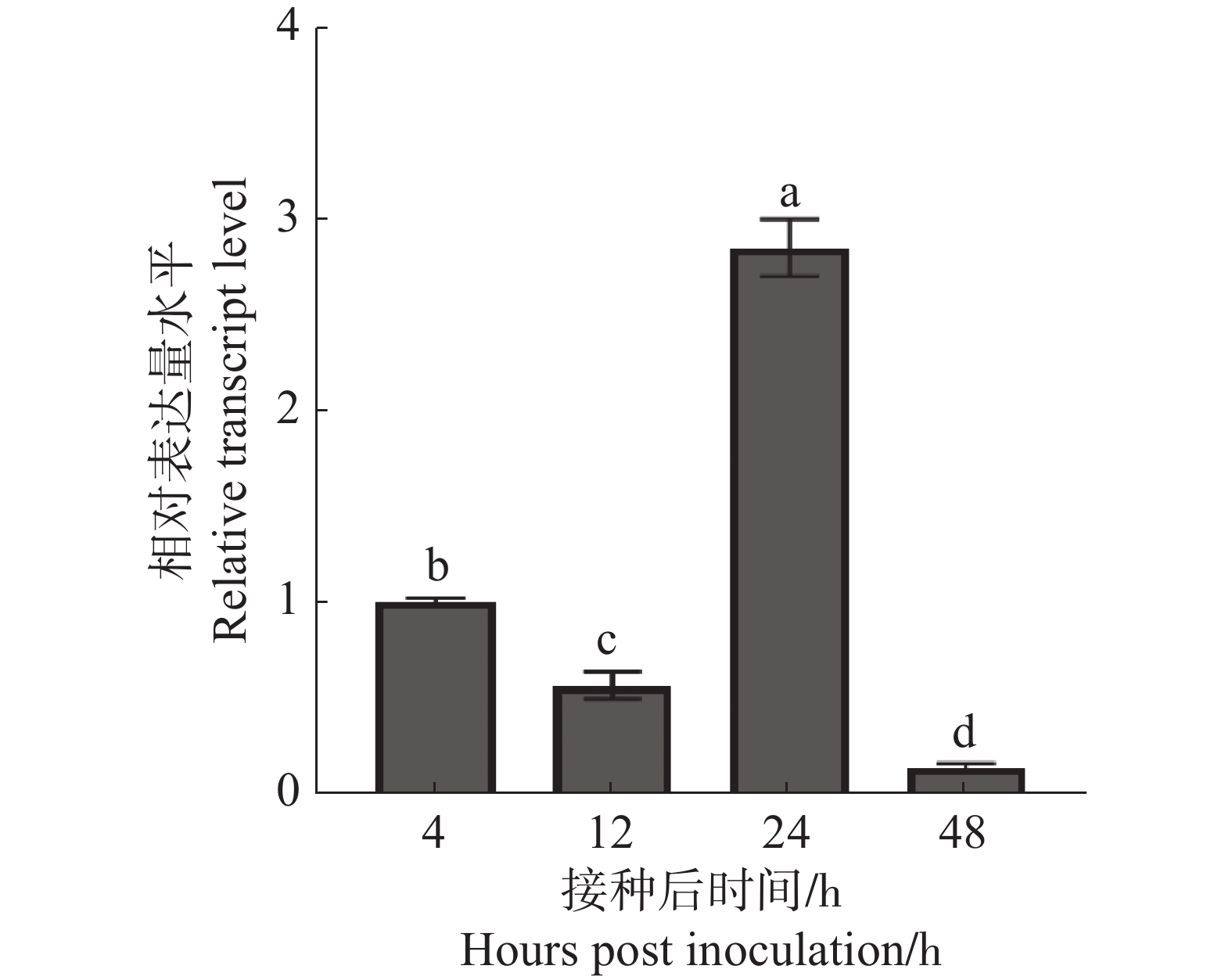
根据前期研究报导的橡胶树白粉菌HO-73菌株在接种后侵染几个关键时间节点(包括4 h分生孢子萌发的高峰期、12 h侵染的高峰期、24 h吸器的形成期、48 h菌丝的扩展阶段及二次侵染的时期)[15],分析CSEP00565基因在与寄主互作过程中表达特性。分别从整个感染橡胶树白粉菌的橡胶树叶片中提取接种后4、12、24和48 h的橡胶树白粉菌的总RNA。通过qRT-PCR分析橡胶树白粉菌侵染过程中CSEP00565基因表达水平。结果显示:与4 h CSEP00565基因表达量水平相比,CSEP00565在白粉菌侵染12 h时表达量降低,但在24 h升高,是4 h的2.8倍,而48 h又大幅度降低(图4),说明CSEP00565可能主要在侵染过程中吸器形成时期发挥作用。
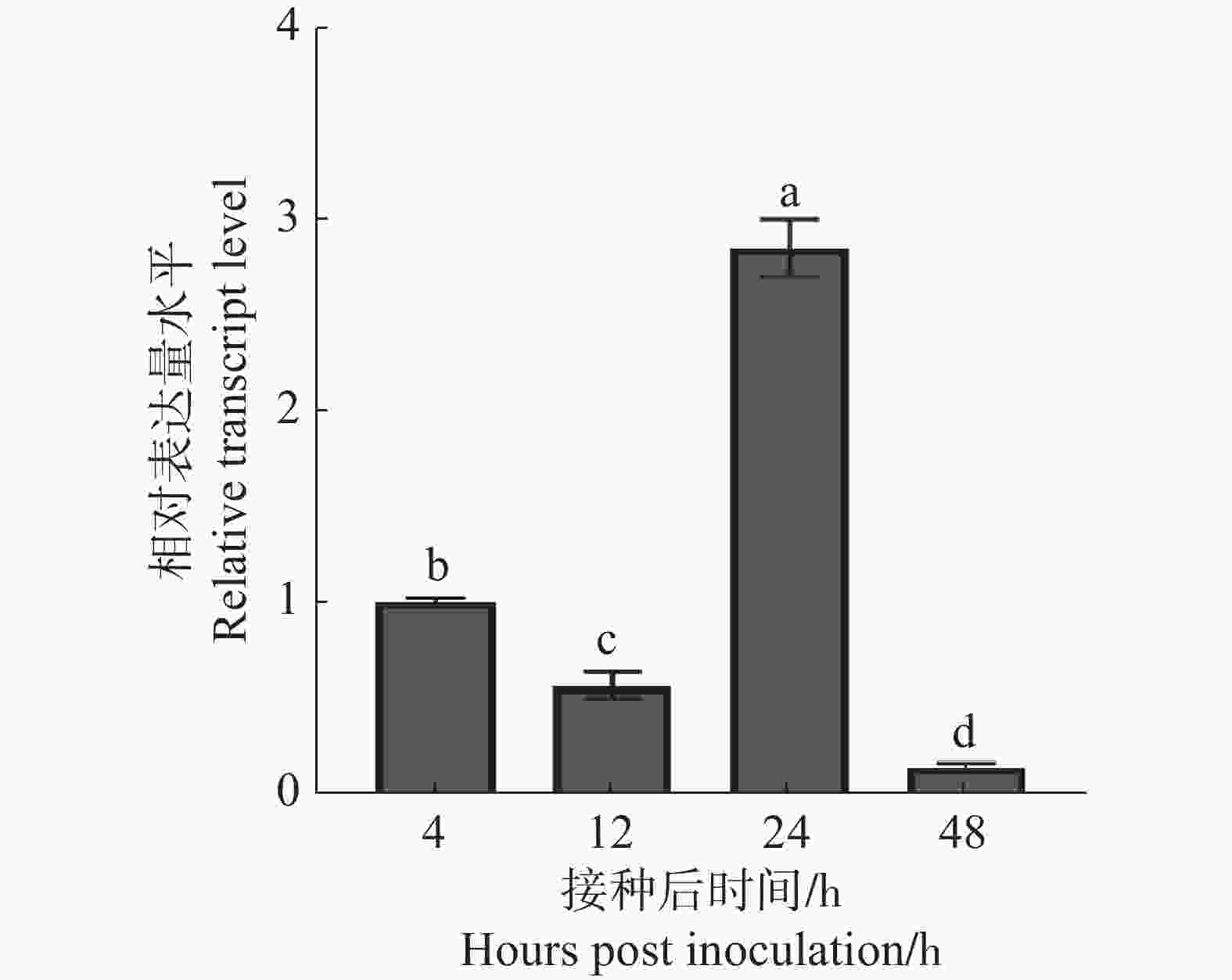
Figure 4. The relative expression level of CSEP00565 at the infection stage
-
SignalP在线服务系统(
http://www.cbs.dtu.dk/services/SignalP/ )对CSEP00565的蛋白氨基酸序列分析表明,CSEP00565携带含有18个氨基酸的N端信号肽。本研究中使用酵母蔗糖转化酶分泌试验检测了该信号肽的分泌活性。将CSEP00565信号肽序列片段插入pSUC2载体的EcoRI、XhoI酶切位点(pSUC2-CSEP00565-SP),并将pSUC2-CSEP00565-SP和pSUC2转入YTK12菌株。以已知分泌蛋白Avr1b信号肽作为阳性对照,而未转化的YTK12和转化了空载pSUC2的YTK12作为阴性对照[17]。与Avr1b信号肽相似,pSUC2-CSEP00565-SP转化的YTK12可以在YPRAA培养基上生长(图5)。同时CSEP00565信号肽转化菌株具有蔗糖转化酶活性,可将2,3,5-三苯基氯化四氮唑(TTC)还原为红色化合物(图5)。实验结果表明CSEP00565是分泌蛋白,且其信号肽序列能够有效地引导蛋白的分泌。
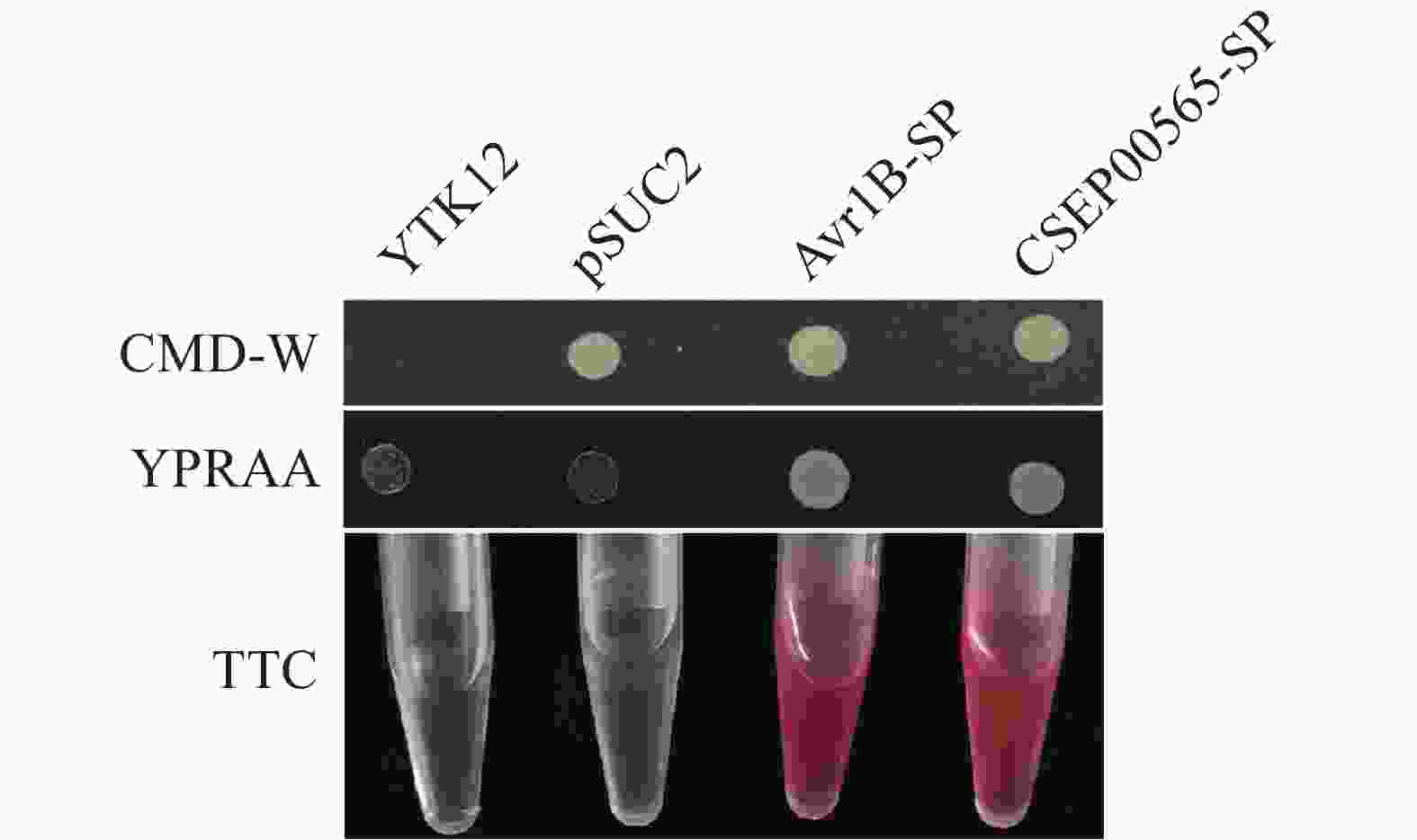
Figure 5. Yeast invertase secretion assay to determine the function of CSEP00565 signal peptide
-
在多个白粉菌中都有同源的CSEPs效应蛋白可能源自共同的祖先[27],并可能是致病或适应寄主所需的因子被继承下来。橡胶树白粉菌含有24个高保守性的CSEPs,它们在其他75种以上白粉菌都有同源。本研究通过烟草表达CSEPs效应蛋白抑制INF1诱导过敏性坏死的测试,筛选到CSEP00565表现出了抑制坏死反应的活性。INF1被烟草细胞表面受体NbEIR1识别,激活下游BAK1和BSK1受体,传导过敏性反应信号[28]。其他一些研究中,也将病原真菌效应蛋白在本氏烟草上异源表达并测试其对过敏性坏死的抑制水平,据此鉴定出具有促进致病功能的毒性效应蛋白。比如,小麦条锈菌效应蛋白Pst_12806、Pst_8713、小麦镰刀菌的效应蛋白OsP24在烟草中瞬时表达可以抑制BAX或INF1诱导的过敏性坏死[29 − 31]。在拟南芥表达CSEP00565也使flg22和(GlcNAc)7诱导的胼胝质沉积被抑制,进一步验证了CSEP00565具有抑制植物免疫的功能。细菌鞭毛蛋白flg22被拟南芥细胞表面受体激酶FLS2识别,FLS2可对类受体胞内蛋白激酶BIK1进行磷酸化,激活NADPH氧化酶及其他信号通路,从而产生免疫反应[32 − 33];而几丁质被细胞表面受体CERKI识别,激活了丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号传递途径,从而触发免疫反应[34 − 35]。也有研究表明FLS2和CERKI下游途径都与BIK1受体有关[36]。根据本文研究结果,推测CSEP00565可能影响了烟草中NbEIR1下游信号通路,或拟南芥中FLS2和CERK1下游信号通路的共同节点,从而干扰了植物免疫。
为了进一步研究效应蛋白CSEP00565是否是橡胶树白粉菌侵染所必需,通过SIGS对CSEP00565进行沉默,发现沉默菌株吸器数量减少,菌丝扩展率大幅度下降,表明CSEP00565功能对橡胶树白粉菌侵染非常重要。寄主诱导基因沉默(Host-induced gene silencing)[37]技术被应用于评估专性寄生菌(包括白粉菌和小麦条锈菌)的基因功能[11,38],然而,该方法并不适用于橡胶树白粉菌,因为橡胶树难以转化,不适于表达基因沉默载体。目前研究发现,使用SIGS技术诱导基因沉默也是有效的[16]。SIGS已被应用于核盘菌(Sclerotinia sclerotiorum)、葡萄孢菌(Botrytis cinerea)、禾谷镰刀菌(Fusarium graminearum)及专性寄生菌瓜类白粉菌(Podosphaera xanthii)[39 − 40]。最近研究表明使用SIGS可以沉默橡胶树白粉菌CYP5基因,将体外合成荧光素标记的β-tubulin基因dsRNA与白粉菌分生孢子孵育,检测荧光发现孢子中出现荧光,说明橡胶树白粉菌吸收了外源dsRNA。将CYP5基因的dsRNA喷施到发病橡胶树叶片中,发现基因表达量也下降了40.3%−59.7%[41],表明SIGS也可用于研究橡胶树白粉菌基因功能。本研究中CSEP00565基因的表达量也下降了87%,说明SIGS干扰橡胶树白粉菌基因表达效果显著。
通过qRT-PCR分析,发现CSEP00565在接种后24 h上调表达,说明该蛋白可能在吸器形成阶段影响白粉菌和寄主的互作。类似地,通过对小麦白粉菌侵染阶段的转录组分析,发现吸器形成阶段有大量CSEPs效应蛋白基因上调[42];利用寄主诱导基因沉默技术干扰这些CSEPs基因,导致病菌吸器形成率和扩展能力显著降低。在麦白粉菌中,通过分离吸器组分,也鉴定到其中富集许多效应蛋白[43]。这些结果也揭示白粉菌效应蛋白对于侵入和定殖具有重要作用。
效应蛋白CSEP00565被预测含有N端信号肽,通过酵母蔗糖转化酶分泌实验验证该信号肽具有引导蛋白分泌功能,因此CSEP00565属于经典泌出型蛋白(依赖内质网—高尔基体分泌途径)。此外,有一些分泌蛋白不含有信号肽,可能通过非经典途径分泌。有的非经典分泌型效应蛋白,也在植物与病原互作中发挥重要作用。如大豆疫霉(Phytophthora sojae)和大丽轮枝菌(Verticillium dahliae)的ISC类型效应蛋白[5]、橡胶树白粉菌EqCmu[15]。
在下一步研究中,将分析效应蛋白转运至植物中的定位和所干扰的免疫信号途径,开发将CSEP00565作为靶点的SIGS技术,为发展病害防控策略提供理论支撑。
Screening of effector protein CSEP00565 of Erysiphe quercicola and preliminary analysis of its mechanism
DOI: 10.15886/j.cnki.rdswxb.20230143
- Received Date: 2023-12-29
- Accepted Date: 2025-01-03
- Rev Recd Date: 2024-03-12
- Available Online: 2025-05-23
- Publish Date: 2025-05-25
-
Key words:
- powdery mildew /
- effector proteins /
- plant immunity
Abstract: The rubber tree (Hevea brasiliensis) is an important economic crop in tropical areas of China, and its related industries play an important role in the local agricultural economy. The powdery mildew fungus Erysiphe quercicola is the most severe disease infecting H. brasiliensis. The effector proteins secreted by phytopahogenic microbes play important roles in the pathogenesis, but how effector proteins promote the infection by E. quercicola is less documented, which has limited disease resistance breeding programme and disease control strategies. The E. quercicola has 24 highly conserved candidate effector proteins, which has homologous in many other powdery mildews. It is possible that these proteins are required for the pathogenicity of powdery mildew. In order to find the important virulence factor among the 24 CSEPs, two conserved effector proteins were selected for functional analysis and found that only the effector CSEP00565 suppressed hypersensitive responses induced by Phytophthora infestans INF1 when these effectors were expressed in Nicotiana benthamiana. The effector CSEP00565 suppressed callose deposition induced by fungal chitin and bacteria flg22 when this effector was expressed in Arabidopsis thaliana, further supporting that CSEP00565 has a function to inhibit plant immunity. When CSEP00565 was further silenced by using spray-induced gene, the pathogenicity of the fungus significantly decreased. And the expression level of CSEP00565 was up-regulated at the haustorium formation stage (24 h) after inoculation with E. quercicola. Further research has discovered that the signal peptide of CSEP00565 possesses the activity of direct protein secretion. All these results indicate that CSEP00565 is a key virulence factor in promoting the infection by E. quercicola, which is conducive to understanding of the molecular mechanisms underlying H. brasiliensis-E. quercicola interaction.
| Citation: | LIU Lijuan, LIU Yuhan, FANG Yuxiao, SHAN Jiaxin, ZUO Minghao, MIAO Weiguo, LI Xiao. Screening of effector protein CSEP00565 of Erysiphe quercicola and preliminary analysis of its mechanism[J]. Journal of Tropical Biology, 2025, 16(3): 389-399. doi: 10.15886/j.cnki.rdswxb.20230143

|
 DownLoad:
DownLoad: