-
木薯(Manihot esculenta Crantz)为大戟科木薯属灌木状多年生作物,是和马铃薯、甘薯并称的世界三大薯类作物之一,有近五千年的栽培史[1],广泛栽种于热带和亚热带地区的100多个国家和地区[2]。1820年前后,木薯首次传入我国广东地区,目前种植范围已遍及华南地区。木薯在我国主要用作工业原料,可生产2 000多种产品[3],相关产业在当地农业经济中占有重要地位。自2005年以来,我国一直是世界上最大的木薯进口国[4]。由地毯草黄单胞木薯萎蔫致病变种(Xanthomonas axonopodis pv. manihotis,简称Xam)侵染引起的细菌性萎蔫病,也称细菌性枯萎病或细菌性疫病,是世界木薯种植中的重要病害。病原菌从气孔或伤口侵入并在附近细胞的间隙内增殖,通过维管束进行系统性扩散。病原菌主要危害叶片和茎秆,能够产生叶片角斑、萎蔫、枯萎、提前脱落及幼苗萎蔫、枝条回枯、茎干溃疡、维管束坏死等症状。植株受害后长势变弱、严重时死亡,重病田块产量损失可达一半以上。1963年,该病首次在我国台湾地区发生[5],1980年在我国海南省发生,随后扩散到广东、广西等地区[6]。李超萍等[7]的调查表明,该病已在国内木薯主栽区普遍发生,是当前危害我国木薯最严重的病害。
铜基杀菌剂是作物细菌性病害防控中的常用药剂,也是我国木薯细菌性萎蔫病防控中常见的推荐药剂[8]。前期研究发现国内外代表性菌株对铜离子和氢氧化铜、噻菌铜等铜基杀菌剂均具有较高水平的抗性,而国外部分已测序的菌株中均编码有copTAB和XmeRSA两个候选的抗铜基因簇[9],但国内及其他地区的Xam菌株是否具有抗铜性并携带这2个抗铜基因簇尚缺少研究。因此,本研究在继续收集相关菌株并进行抗铜性评价的基础上,根据基因序列特征设计引物,通过 PCR 扩增进行这2个基因簇的克隆和序列分析研究,为进一步开展相关基因的功能验证和病原菌抗铜机理分析提供依据。
-
(1)菌株:供试的39个Xam菌株均由中国热带农业科学院环境与植物保护研究所提供,菌株编号、分离所用样品采集时间和地点见表1。(2)培养基和试剂:Xam菌株的活化和培养均采用YGP 培养基;大肠杆菌的培养采用LB 培养基(参照方中达[10]的方法制备)。DNA片段胶回收试剂盒和pMD18-T载体购买自宝生物工程(大连)有限公司;Taq酶和dNTP购买自天根生化科技(北京)有限公司;所用引物对(Cop1:GTGCTGCATCGCCTGCTC和Cop2:TCAAAACCACACGCGTACAC;Xme1:ATCAGTCGGCCTTGGGC和Xme2:TCACGGCTCAAACCGATACC)由北京六合华大基因科技股份有限公司合成;大肠杆菌T1感受态细胞购自北京全式金生物技术有限公司;其他试剂均为国产分析纯。
序号 菌株 病样采集时间 病样采集地点 重复次数 抑菌最低有效浓度/(mmol·L−1) 1 XamJX02 2018.10 江西省抚州市东乡区 3 1.45 2 XamJX08 2020.08 江西省抚州东乡区圩上桥镇 3 1.25 3 XamJX10 2020.05 江西省抚州东乡区 4 1.30 4 XamGX09 2019.09 广西壮族自治区桂平市 4 1.30 5 XamGX39 2017.11 广西壮族自治区合浦县 3 1.40 6 XamGX42 2017.10 广西壮族自治区桂平市石龙镇 3 1.35 7 XamGX43 2018.09 广西壮族自治区南宁市 5 1.45 8 XamGX44 2019.07 广西壮族自治区隆安县那桐镇大藤村 4 1.25 9 XamGX45 2019.07 广西壮族自治区北海市合浦县石湾镇 3 1.25 10 XamGX46 2019.07 广西壮族自治区南宁市武鸣区里建镇 3 1.25 11 XamGX47 2017.11 广西壮族自治区合浦县 3 1.40 12 XamGX48 2017.11 广西壮族自治区合浦县 3 1.40 13 XamGX49 2017.11 广西壮族自治区南宁市武鸣区 3 1.40 14 XamGX50 2018.11 广西壮族自治区合浦县 4 1.35 15 XamGX51 2019.09 广西壮族自治区桂平市 3 1.25 16 XamGX52 2019.08 广西壮族自治区隆安县 3 1.25 17 XamGX53 2019.09 广西壮族自治区隆安县 3 1.25 18 XamGX68 2019.10 广西壮族自治区平南县 4 1.25 19 XamGX69 2020.07 广西壮族自治区平南县武林镇安塘村 3 1.30 20 XamGX70 2020.07 广西壮族自治区桂平市石龙镇黄塘村 3 1.25 21 XamGD25 2017.04 广东省云浮市 3 1.35 22 XamGD26 2017.09 广东省遂溪县乐民镇 3 1.30 23 XamGD54 2018.09 广东省广州市花都区 4 1.25 24 XamGD55 2019.09 广东省开平市苍城镇 3 1.25 25 XamHN16 2017.09 海南省澄迈县美亭镇美玉村 3 1.30 26 XamHN17 2017.09 海南省儋州市大成镇 4 1.35 27 XamHN54 2018.07 海南省儋州市新盈农场 3 1.35 28 XamHN56 2018.10 海南省儋州市大成镇 3 1.35 29 XamHN57 2019.07 海南省儋州市 4 1.25 30 XamHN58 2019.07 海南省澄迈县美亭镇 3 1.35 31 XamMK1822 2018.08 密克罗尼西亚 3 1.45 32 XamMK1823 2018.08 密克罗尼西亚 4 1.55 33 XamKHM04 2019.09 柬埔寨桔井省 3 1.35 34 XamKHM05 2019.09 柬埔寨上丁省 3 1.25 35 XamML1825 2018.12 马来西亚沙巴市 3 1.25 36 Xam1197 2013.12 哥伦比亚 3 1.65 37 Xam1198 2013.12 哥伦比亚 3 1.35 38 XamUG1 2014.12 乌干达基里扬东戈区 3 1.35 39 XamUG2 2015.02 乌干达马辛迪区 3 1.45 -
各病原菌菌株活化、培养后收集菌体,用SDS法提取基因组DNA[13]。PCR反应体系为:10×PCR缓冲液2.5 µL、10 mmol·L−1 dNTP 1 µL、10 µmol·L−1各引物溶液1 µL、40%甘油3.75 µL、20%DMSO 3.75 µL、DNA模板50~100 ng、Taq DNA聚合酶1.0 U,加无菌水至25 µL。PCR循环参数为:94℃预变性3 min;94℃ 30 s、54℃ 30 s、72℃ 2 min,35个循环;最后72℃延伸10 min。扩增产物用l.0 %琼脂糖凝胶电泳检测。参照DNA片段胶回收试剂盒和pMD18-T载体试剂盒说明书进行扩增产物的回收、连接和克隆,筛选阳性克隆后,各样品随机挑选3个克隆送北京六合华大基因科技股份有限公司进行序列测定,确认序列后在
http://www.ncbi.nlm.nih.gov/BLAST 上进行比对。和近源菌株这2个基因簇的序列进行聚类分析,用MEGA version 4.0中的NJ(Neighbor-Joining)方法生成系统树,用Bootstrap 对系统树进行检验,1 000次重复。 -
完成了39个菌株的抗铜性评价,结果表明,硫酸铜对各菌株的抑菌最低有效浓度在1.25~1.55 mmol·L−1之间。来自哥伦比亚的菌株Xam1197抗铜能力最强,为1.55 mmol·L−1,来自江西的XamJX08,广西的XamGX44、XamGX45、XamGX46、XamGX51、XamGX52、XamGX53、XamGX68和XamGX70,来自广东的XamGD54、XamGD55,来自海南的XamHN57,来自柬埔寨的XamKHM05和马来西亚XamML1825的抗铜性较差,均为1.25 mmol·L−1,其他菌株的抗铜性在1.30~1.45 mmol·L−1之间(表1)。
-
根据2个候选基因簇的序列特征,分别设计了Cop1、Cop2和Xme1、Xme2两对引物。提取各菌株基因组DNA后,分别进行PCR扩增,各菌株用2个引物对均能获得大小约3.4 kb的扩增产物。随机选择国内外17个代表性菌株(XamJX02、XamJX08、XamGX46、XamGX69、XamGX70、XamGD26、XamGD54、XamHN16、XamHN57、XamMK1823、XamKHM04、XamKHM05、XamMLXY1825、Xam1197、Xam1198、XamUG1和XamUG2)的扩增产物,纯化回收、克隆后测序并进行同源基因的比较。
所有菌株均编码有和XamGX11完全相同的CopTAB基因簇,该基因簇由CopT、CopA和CopB等编码方向一致的3个基因组成,3个基因之间的非编码区长度分别为90 nt和454 nt。这些菌株中,仅有XamGD26和其他菌株在copA编码区的第964个nt为“A”,而其他菌株为“C”,对应的氨基酸为异亮氨酸,其他菌株为缬氨酸。以其他菌株共有的基因序列进行了初步分析,CopT编码区全长425 nt,和多个黄单胞菌的预测CopL家族金属结合调节基因(或称铜转运蛋白)仅有2个碱基的差异,同源性最高为99%(登录号CP012063.1、CP012060.1等),该基因编码139个aa,和番茄细菌性斑点病菌(Xanthomonas axonopodis pv. vesicatoria)菌株7882的预测蛋白序列(WP_017157235.1)完全一致[14]。CopA全长1806 nt,和黄单胞菌的预测多铜氧化酶基因同源性最高,为99%(CP012063.1、CP012060.1等),有21个碱基的差异,该基因编码601个aa,和菜豆黄单胞菌(Xanthomonas phaseoli)预测蛋白序列(WP_017157234)一致[15]。CopB全长654 nt,和地毯黄单胞万年青致病变种(Xanthomonas axonopodis pv.dieffenbachiae)菌株LMG 695预测的抗铜相关基因(CP014347.1)仅有16个碱基的差异,同源性为99%,编码369个aa,和菜豆黄单胞菌预测蛋白序列(WP_017157233)一致。
和CopTAB基因簇相同,所有菌株同样编码有XmeRSA基因簇,其中XmeR和XmeS位于染色体负链上,两者编码区有4个碱基的重叠,而XmeA位于正链上,和另外2个基因编码区方向相反并有161 nt的间隔区。这些菌株中,XamGX69和XamGD54与其他菌株均有1nt的序列差异。XamGX69在XmeS编码区的第1 015个nt为“A”,而其他菌株为“C”,对应的氨基酸为苏氨酸,其他菌株为丙氨酸。XamGD54在XmeA编码区的第1 141个nt为“C”,而其他菌株为“G”,对应的氨基酸为天冬氨酸,其他菌株为组氨酸。以其他菌株共有的基因序列进行了分析,XmeR编码区全长684 nt,和多个黄单胞菌预测的DNA结合转录调节因子BaeR基因(CP012063.1、CP012060.1、CP012057.1等)同源性最高,为99%,仅有6个nt的差异,该基因编码227个aa,和菜豆黄单胞菌等病原菌预测蛋白序列(WP_046833221、WP_017157623)一致[16]。XmeS编码区全长1 380 nt,和黄单胞菌预测的组氨酸激酶感受分泌调节基因BaeS同源性为99%,有12个nt的差异,编码459个aa,和菜豆黄单胞菌预测蛋白序列(WP_017157622)完全相同。XmeA编码区全长1 224 nt,和多个黄单胞菌预测的外排RND(resistance-nodulation-cell-division,简称RND,耐药结节细胞分化)转运蛋白亚基基因(CP012063.1、CP012060.1、CP012057.1等)有24个nt的差异,同源性为99%,编码407aa,和菜豆黄单胞菌预测基因蛋白序列(WP_017157621)完全一致。
-
在NCBI数据库中检索和下载近源黄单胞病原菌及其他病原菌中的copTAB和XmeRSA同源基因簇(表2),构建NJ 系统进化树,分析其进化关系,结果表明,所获17个木薯细菌性萎蔫病菌菌株的copTAB基因簇聚为1个分枝,和菜豆黄单胞菜豆变种(Xanthomonas phaseoli pv. phaseoli)菌株PR1同源基因的亲缘关系最近。柑橘溃疡病菌(Xanthomonas axonopodis pv. citri)菌株306、柠檬黄单胞病柠檬致病变种(Xanthomonas citri pv. citri)菌株LH276、柠檬黄单胞菌(Xanthomonas citri pv. punicae)菌株LMG7504和菜豆普通细菌性疫病病菌(Xanthomonas fuscans subsp. fuscans)菌株ISO118C5等聚成1个分枝,和木薯细菌性萎蔫病菌菌株有很高的同源性。细菌性叶斑病菌(Xanthomonas axonopodis pv. vesicatoria)菌株7882、柠檬黄单胞病菌(Xanthomonas citri subsp. citri)菌株A44和野油菜黄单胞菌胡桃致病变种(Xanthomonas campestris pv. juglandis)菌株C5聚为1个分枝,同源性也较高。农杆菌(Agrobacterium)菌株CFBP 6623、丁香假单胞菌丁香致病变种(Pseudomonas syringae pv. syringae)菌株DC3000和木薯细菌性萎蔫病菌菌株的同源性最低(图1)。
病原菌(简写) 菌株名称 序列登录号 基因(基因簇)组成 Agrobacterium(Agr) CFBP 6623 LT009725.1 1个预测的铜输出P型ATP酶基因 Pseudomonas syringae pv. Syringae(Pseu) DC3000 KY362372.1 由copS、copR、copD、copC、copB和copA等基因组成 Xanthomonas citri subsp. citri(Xcc) A44 HM362782 由copL、copA、copB、copM、1个转座酶基因、copG、1个预测蛋白、copC、copD、copF等基因组成[17] Xanthomonas citri pv. citri(Xcc) LH276 NZ_CP018854.1 由copL、copA和copB等基因组成 Xanthomonas campestris pv. juglandis(Xcj) C5 L19222.1 由4个预测基因组成[18] Xanthomonas axonopodis pv. citri(Xac) 306 AE008923.1 由copL、copA前体和copB等基因组成[19] Xanthomonas axonopodis pv. vesicatoria(Xav) 7882 AY536748 由copL、copA、copB、copM、copG、copF等基因组成[14] Xanthomonas citri pv. punicae(Xcp) LMG7504 CP030166.1 由copL、copA和copB等基因组成 Xanthomonas fuscans subsp. fuscans(Xff) ISO118C5 CP012051 由copL、copA和copB等基因组成 Xanthomonas phaseoli pv. phaseoli (Xpp) PR1 NZ_CP029277.1 由copL、copA和copB等基因组成 Bacillus methylotrophicus(Baci) B25 LN999829.1 由BaeM、BaeN、BaeR和BaeS等4个预测的外排RND转运相关基因组成 Xanthomonas citri pv. citri(Xcc) 29-1 CP023661.1 由BaeS及另外4个外排RND转运相关基因组成 Xanthomonas campestris pv. vesicatoria(Xcv) 85-10 CP017190.1 1个预测的多药运输蛋白基因 Xanthomonas citri pv. mangiferaeindicae(Xcm) GXG07 CP073209.1 由BaeR、BaeS、smeA和1个预测的多药运输蛋白基因组成 Erwinia tasmaniensis(Erw t) ET1/99 CU468135.1 由BaeS和BaeR等2个基因组成[19] Xanthomonas axonopodis pv. phaseoli(Xap) ISO98C12 CP012057.1 由预测的1个RND转运蛋白和2个多药运输蛋白基因组成 Xanthomonas perforans(Xp) 91-118 CP019725.1 由预测的1个RND转运蛋白、1个短链脱氢酶和1个预测的多药运输渗透酶亚基组成[20] Xanthomonas phaseoli pv. phaseoli(Xpp) Xcp25 CP029284.1 由预测的1个运输蛋白、1个多药外排RND转运体渗透酶亚基、1个外排RND转运蛋白细胞质适应亚基和BaeS组成 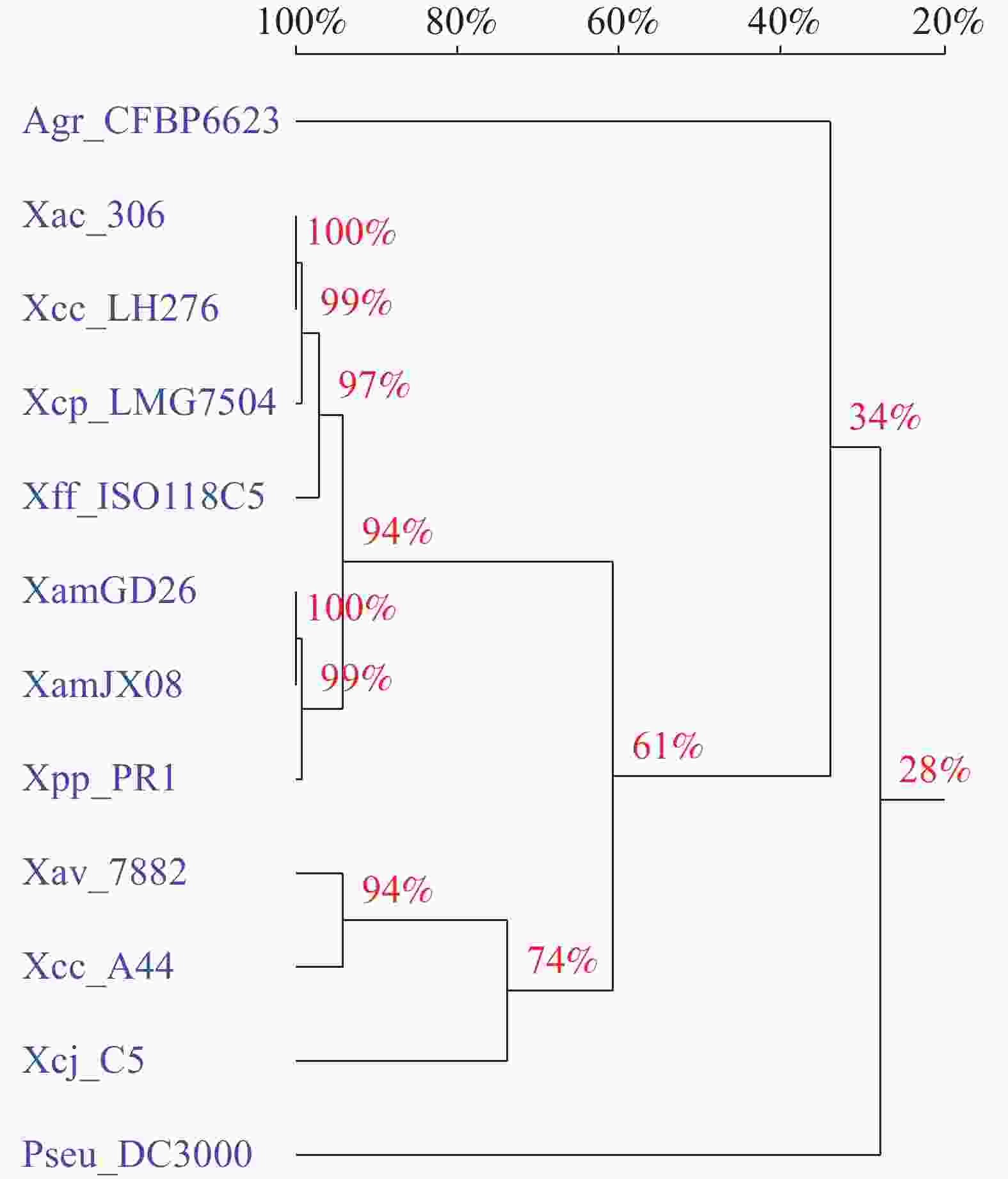
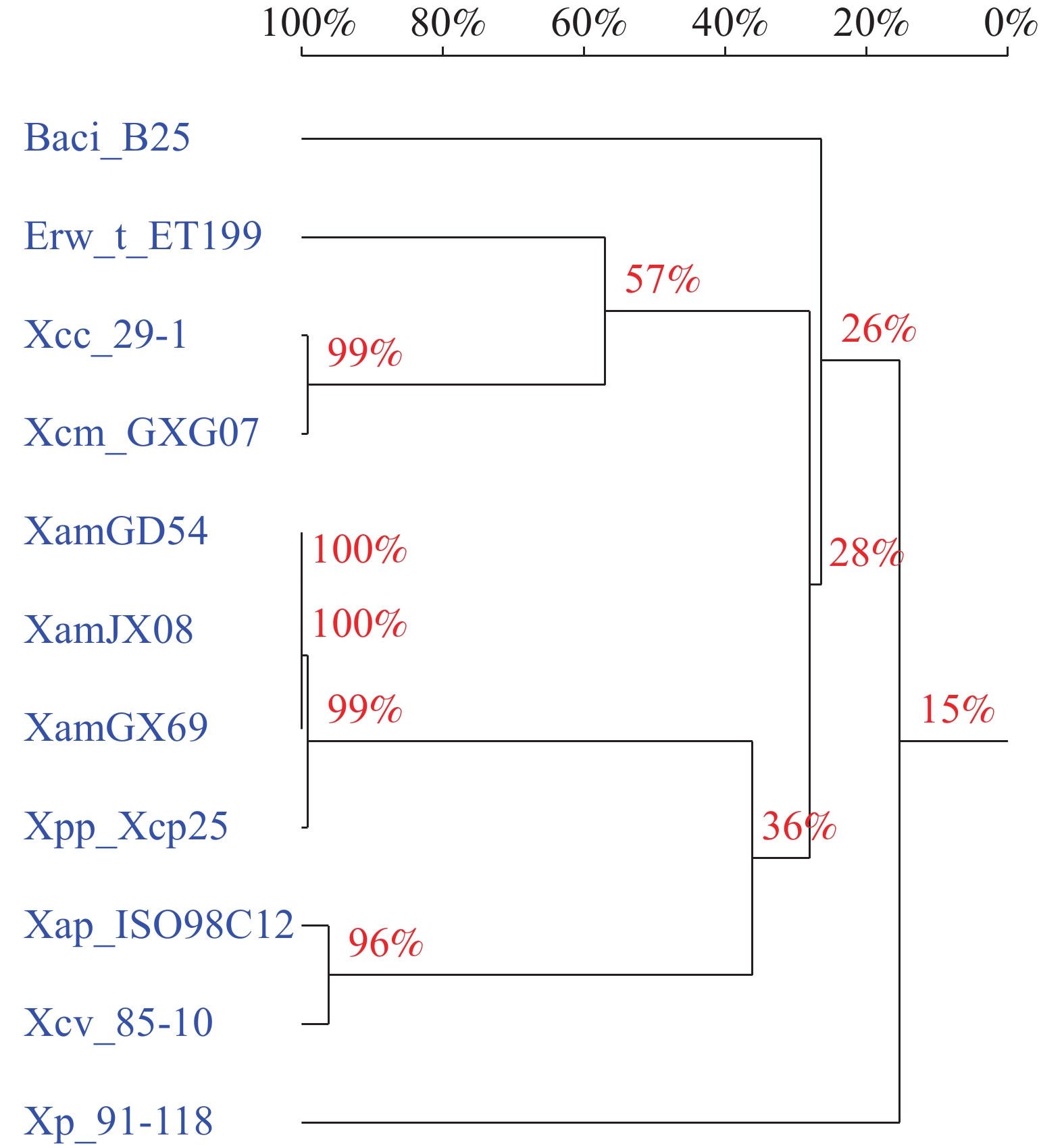
各木薯细菌性萎蔫病菌菌株的XmeRSA基因簇同样聚为1个分枝,菜豆黄单胞菜豆变种菌株Xcp25同源基因的相似性最高。地毯草黄单胞菌菜豆致病菌变种(Xanthomonas axonopodis pv. phaseoli)菌株ISO98C12和野油菜黄单胞菌辣椒致病变种(Xanthomonas campestris pv. vesicatoria)菌株85-10聚为1个分枝,同源性也较高。柠檬黄单胞病菌菌株29-1、柠檬黄单胞病杧果致病变种(Xanthomonas citri pv. mangiferaeindicae)GXG07和 塔斯曼尼亚欧文氏菌(Erwinia tasmaniensis)菌株ET1/99亲缘关系较近,聚为1个分枝,和木薯细菌性萎蔫病菌亲缘关系较远。穿孔黄单胞(Xanthomonas perforans)菌株91-118和甲基营养型芽孢杆菌(Bacillus methylotrophicus)菌株B25与木薯细菌性萎蔫病菌亲缘关系最远(图2)。

-
研究结果表明,来自国内不同种植区以及密克罗尼西亚、马来西亚、乌干达、哥伦比亚等不同国家的木薯细菌性萎蔫病菌均对铜离子具有较高的抗性。分子克隆结果表明,所有供试菌株均携带有copTAB和XmeRSA两个高度保守的抗铜基因簇,其和黄单胞及其他常见病原菌的抗铜相关基因也有较高的同源性,相关基因在病原菌抗铜反应及其他方面的作用机制,将是下一步的研究重点。
不同病原细菌之间的抗铜基因具有保守性,敏感性菌株常通过结合转移获得相关基因并产生抗铜性[21]。国外研究结果表明,随着铜基杀菌剂使用,危害番茄和柑橘的黄单胞病菌中,抗铜菌株的数量和比例也在不断增加[22-23]。国内也有研究结果表明,随着铜基杀菌剂的使用,危害核桃的细菌性疫病病原菌(Xanthomonas arboricola pv. juglandis)菌株种群中出现抗铜能力不同的抗铜性菌株[24];台湾地区的细菌性疮痂病菌(Xanthomonas euvesicatoria)和柑橘溃疡病菌也都具有抗铜性[25]。本研究结果表明,国内外代表性菌株均具备抗铜性,分析菌株之间可能存在广泛的抗铜相关基因转移现象。
除抗铜外,copTAB基因簇还参与黄单胞菌的其他功能。例如番茄疮痂病菌(Xanthomonas gardneri)菌株Xv10的copB基因同时控制着抗铜性和侵染番茄的能力[26],而水稻白叶枯病菌(Xanthomonas oryzae pv. oryzae)菌株PXO99的copAB上产生6个氨基酸突变,导致其对铜离子敏感而且降低了对水稻的侵染能力[27]。笔者团队在国内木薯主栽区及柬埔寨、哥伦比亚、乌干达、密克罗尼西亚、马来西亚等地区收集病原菌,发现所有菌株均具有较高的抗铜性。除国内广西、广东等部分重病区之外,其他国家和地区并无使用铜基杀菌剂的记录,分析这2个基因簇可能和病原菌侵染寄主、适应环境等方面功能相关。
前人研究结果表明,抗铜性黄单胞病菌对铜离子的抗性水平通常在0.6~1.2 mmol·L−1之间[24],本研究结果表明木薯细菌性萎蔫病菌的抗性水平在1.25~1.55 mmol·L−1,整体上其抗铜能力高于前人报道的其他菌株。copTAB基因簇分别编码铜转运蛋白、多铜氧化酶和抗铜相关基因,而XmeRSA基因簇编码DNA结合转录调节因子、组氨酸激酶感受分泌调节因子和外排RND转运蛋白亚基。细菌的抗铜作用包括RND等基因家族的外排作用,阳离子扩散(CDF)基因家族的过滤作用,P型ATP酶的调控作用等不同层次的作用机制,包括铜离子的吸附沉积、隔离、修饰及向外转运等。木薯细菌性萎蔫病菌的抗铜能力相对较高,可能和该病菌同时具备copTAB基因簇的铜离子转运功能和XmeRSA基因簇的外排作用相关。
Cloning and analysis of copTAB and XmeRSA gene clusters from Xanthomonas axonopodis pv. manihotis
DOI: 10.15886/j.cnki.rdswxb.20220009
- Received Date: 2022-03-21
- Accepted Date: 2023-08-27
- Rev Recd Date: 2023-02-20
- Available Online: 2023-09-25
- Publish Date: 2023-09-25
-
Key words:
- c assava bacterial blight /
- copper-resistant gene clusters /
- copTAB /
- XmeRSA
Abstract: Cassava bacterial blight caused by Xanthomonas axonopodis pv. manihotis is one of the important diseases in the world, and it is also the most serious disease in the cassava plantations of China. Some strains in other countries with their whole genomes being sequenced were found to encode two candidate copper resistance gene clusters, copTAB and XmeRSA, and part of these strains were higher in resistance to copper ion. An attempt was made to clone copper resistance gene clusters of more strains from different other countries and analyze their copper resistance level. The results showed all the tested strains from China, Micronesia, Malaysia and other countries were also higher in resistance to copper ion. All the representative strains from these countries also encoded the copTAB and XmeRSA gene clusters. The two gene clusters from these countries were highly homologous, and they were not only involved in copper-resistance metabolism, but might also be related to other conservative functions such as pathogenicity.
| Citation: | XU Chunhua, LI Chaoping, SHI Tao, WANG Guofen, CAI Jimiao, LI Boxun, HUANG Guixiu. Cloning and analysis of copTAB and XmeRSA gene clusters from Xanthomonas axonopodis pv. manihotis[J]. Journal of Tropical Biology, 2023, 14(5): 499-505. doi: 10.15886/j.cnki.rdswxb.20220009

|
 DownLoad:
DownLoad: