-
海南岛地处热带,属热带季风气候,除中部山区外,年均气温为23~26 ℃,是天然温室。同时,海南大部分地区年降雨量都超过1 700 mm,良好的水热条件,非常适合各类农作物的种植[1]。海南是我国最早实现“南菜北运”的省份,豇豆(Vigna unguiculata)就是其中尤为重要的一种[2-3],作为海南农民增产提收最重要的蔬菜品种,从2009—2020年,豇豆的种植面积不断增加,并达到了2.25万hm2,总产量为58万t,约占海南省蔬菜总产量的1/10 [4-5]。但随着豇豆种植面积的逐年增加,病虫害的发生也逐年加重。豆大蓟马(Megalurothrips usitatus)是豆科作物的一种毁灭性害虫,严重危害豇豆品质和产量[6-7]。该虫又名豆花蓟马或普通大蓟马,属缨翅目蓟马科,广泛分布于热带和亚热带地区,其寄主植物主要为豆科类作物[6]。豆大蓟马生长周期短、繁殖力强、隐匿性强,世代重叠严重,因此,极易在短时间内大面积爆发[8]。豆大蓟马为锉吸式口器,其在取食过程刺穿植物组织并吸取植物汁液,造成豇豆叶片皱缩和畸形,严重时会造成生长点萎缩,生长缓慢甚至停止,荚果受害果实表面会变得粗糙或者瘢痕,降低豇豆的果实质量[9-10]。此外,该虫还可以传播植物病毒病,从而对豇豆造成间接危害。目前,其防治仍然以化学防治为主[11]。拟除虫菊酯为一类作用于昆虫钠离子通道(Nav)的广谱性杀虫剂,具有易降解等优点,因此在田间已经大面积应用。其通过延长Nav开放时间使动作电位升高,导致神经细胞重复放电,从而实现杀虫的目的[12]。近年来,随着田间用药量的成倍增加,豆大蓟马对拟除虫菊酯类药剂的抗药性不断增强[12]。其产生抗药性的主要原因之一是Nav发生抗性突变,从而影响药剂与Nav的结合能力,导致敏感性降低进而产生抗药性[13-14]。昆虫Nav由α亚基和β亚基组成,α亚基由4个同源结构域组成,每个同源结构域含有6个跨膜片段,其中,S1~S4称为电压传感模块,而S5和S6区域称为P区,β亚基辅助α亚基调控Nav表达[14-16]。
本研究组前期的研究表明,海南省豆大蓟马田间种群对拟除虫菊酯类杀虫剂已经产生了抗性,但对于豆大蓟马钠离子通道结构与功能仍不清楚。因此,本研究对豆大蓟马钠离子通道基因(MuNav)进行克隆和序列分析,旨在为后续解析豆大蓟马对拟除虫菊酯抗性机制奠定基础。
-
豆大蓟马于2018年12月采自海南省海口市美兰区海南大学海甸校区基地,并在实验室内长期饲养,期间未施用任何药剂。饲养方法如下:将豆大蓟马成虫及若虫转移至240 mL养虫罐中,将新鲜豇豆洗净,切成长度约5 cm的豆荚放入20%蜂蜜水中浸泡1 min,晾干,放入养虫罐中,置于人工气候箱(PYX-400Q-A,广东韶关科力实验仪器有限公司)中饲养。成若虫的饲养温度为26 ℃,湿度为60 %~65 %,光周期16 L∶8 D。
-
取豆大蓟马成虫30~40头,通过Trizol法提取RNA,并用1 %的琼脂糖进行凝胶电泳。随后进行反转录,方法参照Prime ScriptTM 1st Strand cDNA Synthesis Kit(东洋纺生物科技有限公司,上海)说明书。根据NCBI西花蓟马(Frankliniella occidentalis)(XM_026422864.1)和棕榈蓟马(Thrips palmi)(XM_034385472.1)Nav序列,使用Primer 5软件对MuNav进行通用引物设计,正向引物为:ATGCCGAGTGTCCGGGAGTCG,反向引物为:TCAGACATCCGCAAGGGCCGGAG。以上述cDNA为模板,进行PCR扩增,反应体系如下:cDNA 1 μL、dNTP Mix 1 μL、上下游引物各2 μL、ddH2O 18 μL、2×Phanta Max Buffer (Mg2+plus) 25 μL、Phanta Max Super-Fidelity DNA Ploymerase 1 μL。PCR反应条件如下:95 ℃预变性3 min;95 ℃ 15 s,56 ℃ 15 s,72 ℃延伸6.5 min,共35个循环;72 ℃反应后延伸8 min。
-
用Cycle-Pure Kit(OMEGA, America)将上述PCR产物进行纯化回收,用NanoDrop 2000分光光度计(赛默飞世尔科技(中国)有限公司,上海,中国)对回收产物进行浓度测定,并置于−20 ℃备用。分别用HindⅢ-HF和X maⅠ对PGH 19进行双酶切,反应时间为2 h,温度为37 ℃,将纯化回收产物和PGH 19链接,反应条件为:37 ℃、45 min,取出后在冰上冷却5 min,将连接好的产物导入感受态细胞stbl 2中并涂板,方法参照stbl 2感受态细胞(吐露港生物科技有限公司,上海)说明书,将平板倒置在22 ℃的保温箱中培养。在96孔板每孔中加入200 μL LA培养基,并挑取单克隆置于其中,密封后180 r·min−1,30 ℃摇菌12 h。取50 μL菌液加入5 mL LA培养基中于30 ℃、180 r·min−1 2次摇菌16 h,使用Plasmid mini Kit(OMEGA, America)试剂盒进行质粒提取,并进行测序。
-
克隆得到MuNav全长,并上传至NCBI(GenBank accession number: MZ043856),用Simple Modular Architecture Research Tool预测其保守性结构域。同时,从NCBI上下载得到其他58种昆虫的Nav全长(表1)。随后,用Clustal Omega软件(
https://www.ebi.ac.uk/Tools/msa/clustalo/ )进行多序列比对,并利用Mega 7 (DNASTAR, America)软件,采用最大似然法构建系统发育树,自举值设置为1 000。昆虫拉丁文 NCBI序列号 昆虫拉丁文 NCBI序列号 Acromyrmex echinatior XP_011061306.1 Aedes aegypti ACB37024.1 Anopheles gambiae CAM12801.1 Apis cerana XP_016917417.1 Apis dorsata XP_006613060.1 Apis florea XP_012347662.1 Apis mellifera NP_001159377.1 Apolygus lucorum ALF41049.1 Athalia rosae XP_025602777.1 Atta cephalotes XP_012055204.1 Bactrocera dorsalis XP_029407928.1 Bactrocera oleae XP_036228508.1 Blattella germanica AAC47483.1 Bombus impatiens ARH02610.1 Bombus vancouverensis nearcticus XP_033202238.1 Bombyx mori NP_001136084.1 Brassicogethes aeneus AJM87404.1 Camponotus floridanus XP_025269364.1 Ceratitis capitate XP_020717222.1 Cimex lectularius NP_001303632.1 Copidosoma floridanum XP_023247004.1 Diachasma alloeum XP_015109194.1 Dinoponera quadriceps XP_014472738.1 Drosophila melanogaster AAB59195.1 Dufourea novaeangliae XP_015436344.1 Eufriesea Mexicana XP_017755817.1 Fopius arisanus XP_011306701.1 Habropoda laboriosa XP_017797362.1 Halyomorpha halys XP_024214489.1 Helicoverpa zea ADF80418.1 Heliothis virescens AAC26513.1 Liposcelis bostrychophila AGL91669.1 Megachile rotundata XP_012144116.1 Microplitis demolitor XP_014298584.1 Monomorium pharaonis XP_012540554.1 Musca domestica NP_001273814.1 Nasonia vitripennis NP_001128390.1 Odontomachus brunneus XP_032685128.1 Ooceraea biroi RLU19677.1 Orussus abietinus XP_012272834.1 Papilio machaon XP_014357465.1 Papilio xuthus XP_013169662.1 Pediculus humanus capitis AAP20108.1 Plutella xylostella BAF37093.2 Pogonomyrmex barbatus XP_011636355.1 Polistes canadensis XP_014608498.1 Polistes dominula XP_015185094.1 Solenopsis invicta XP_039304541.1 Stomoxys calcitrans XP_013100826.1 Thrips plami XP_034241363.1 Tribolium castaneum EFA11577.2 Trichogramma pretiosum XP_014230252.1 Varroa destructor AAP13992.1 Vollenhovia emeryi XP_011881693.1 Wasmannia auropunctata XP_011689749.1 Zeugodacus cucurbitae XP_028896326.1 -
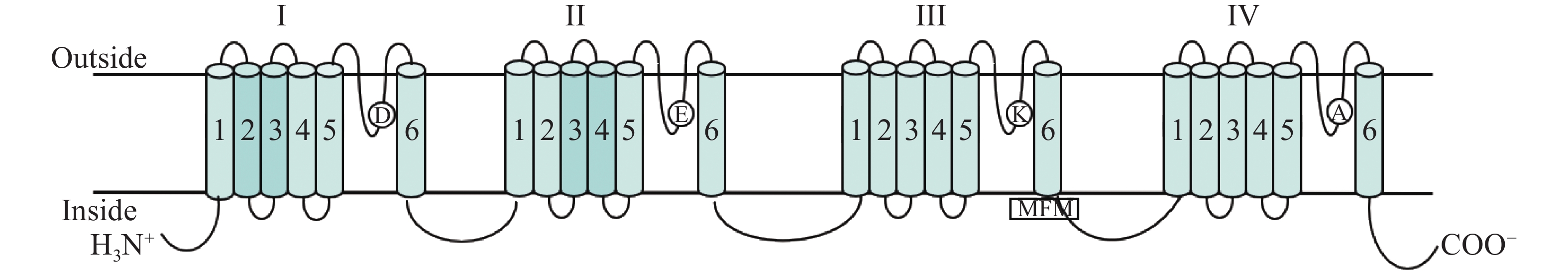
克隆得到MuNav,其全长为6279 bp,编码2093 aa (GenBank:MZ043856) (图1),含有4个同源结构域,结构域 Ⅰ~Ⅳ,每个同源结构域含有6个跨膜片段,第3和第4结构域之间含有“MFM”疏水性残基,符合昆虫Nav的典型特征(图2)。


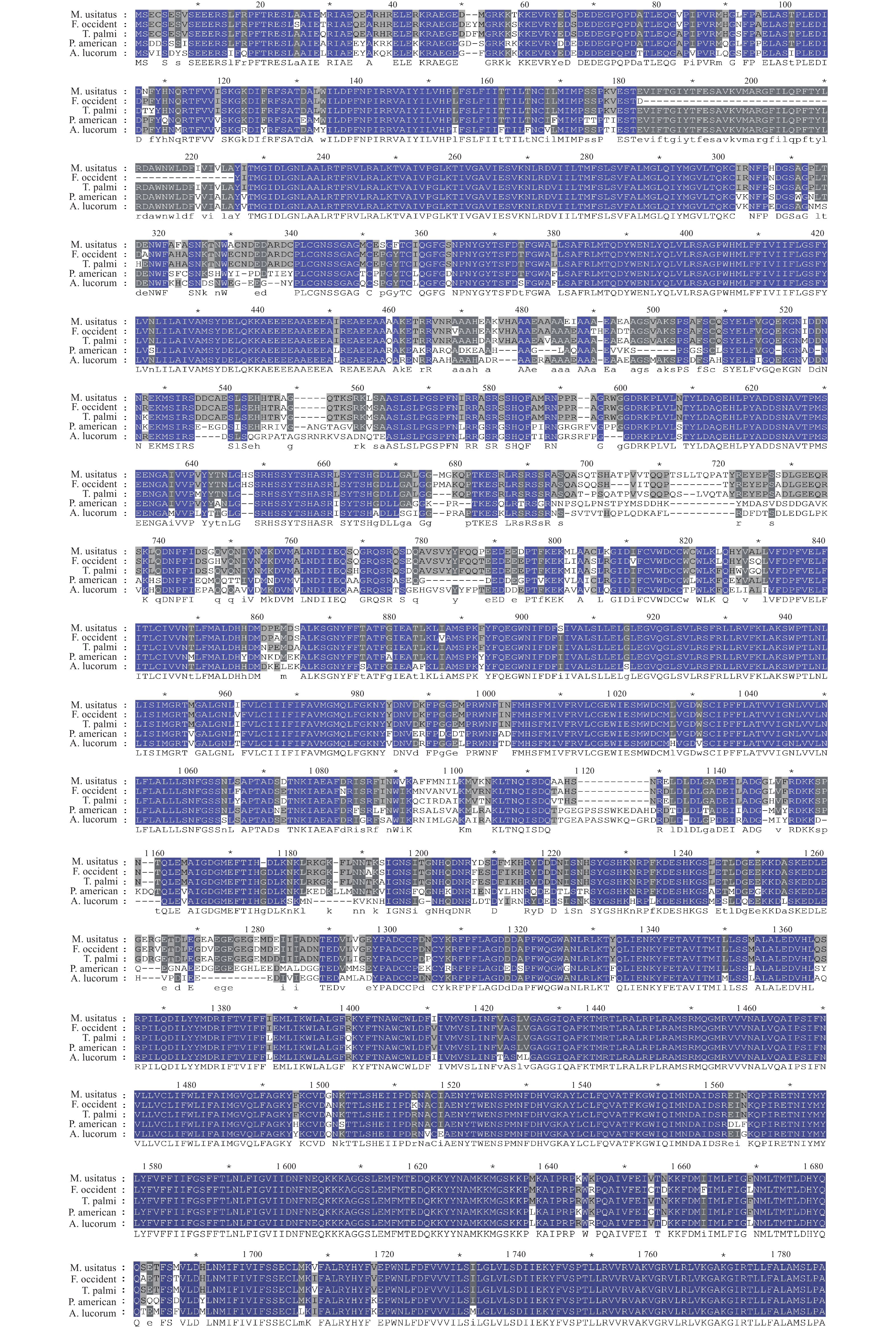
将MuNav的氨基酸序列与西花蓟马、棕榈蓟马、绿盲蝽(Apolygus lucorum)、美洲大蠊(Periplaneta americana)Nav进行同源性比对,发现其与棕榈蓟马Nav序列的相似度高达94.88%,与西花蓟马Nav序列的相似度同样高达94.68%,而与美洲大蠊和绿盲蝽Nav的相似度分别仅为80.74%和80.19%,表明豆大蓟马与蓟马类昆虫的亲缘关系较近,而与其他目昆虫则发生了分化(图3)。
-
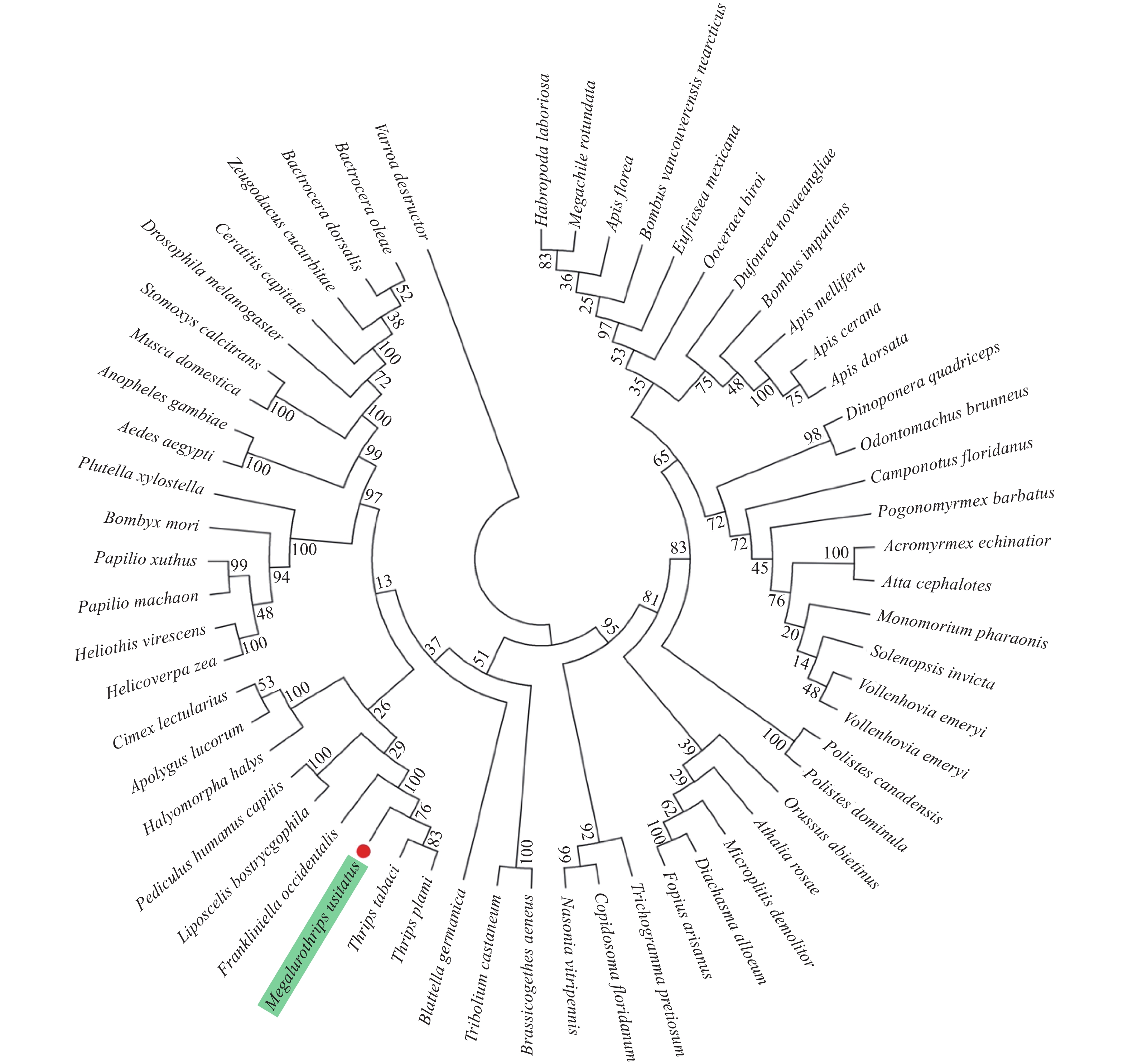
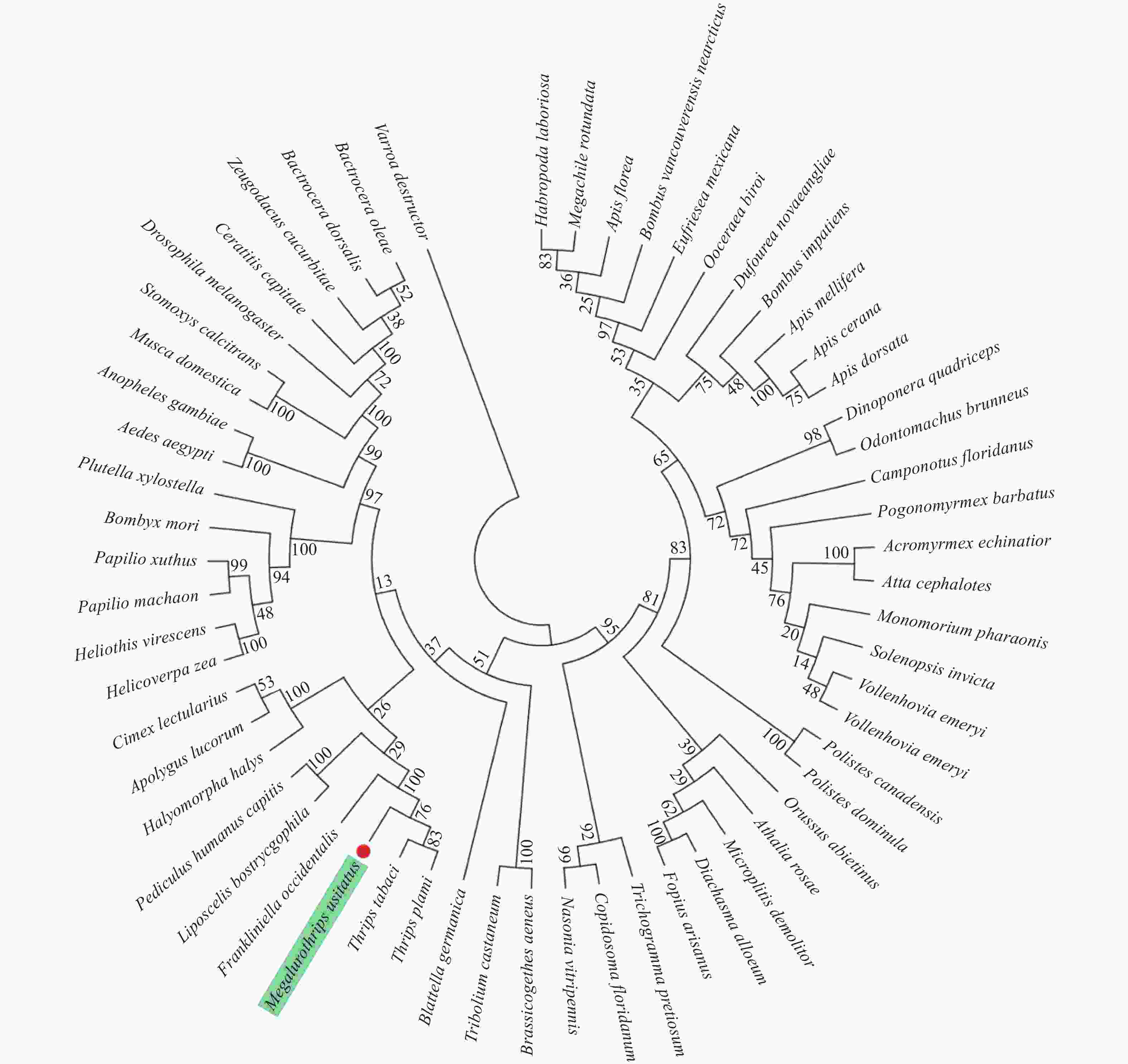
进化树的结果表明(图4),MuNav与其他昆虫均存在一定的亲缘关系。其中,豆大蓟马与棕榈蓟马(T. palmi)、烟蓟马(Thrips tabaci) 和西花蓟马聚为一类,显示蓟马类昆虫Nav符合进化过程,即同一类昆虫聚类在相近位置。反之,豆大蓟马与其他目的昆虫,如黑腹果蝇(Drosophila melanogaster)、德国小蠊(Blattella germanica)、绿盲蝽等则发生了较大程度的分化,亲缘关系相对较远。
-
大部分昆虫仅编码1条或2条Nav,昆虫Nav由1个分子量约为260 kD的α亚基和5个辅助小亚基组成,α亚基具有Nav活性,而辅助小亚基则起到调节Nav表达的作用。本研究通过PCR的方法克隆得到了MuNav全长序列,发现其与其他昆虫Nav结构类似,有4个同源性极高的跨膜结构域,即结构域I、结构域II、结构域III和结构域IV组成,且每个结构域均包含6个疏水性跨膜螺旋体(S1~S6)。在MuNav的P-LOOP环上同样鉴定到与其他昆虫类似的4个保守性氨基酸残基D、 E、 K、A和1个关键失活阀门MFM,这些结果均表明,MuNav具有昆虫Nav特有的全部结构特征,这为后续MuNav功能研究奠定了基础。
本研究的序列比对结果表明,豆大蓟马与其他目昆虫,如蜚蠊目美洲大蠊和半翅目绿盲蝽Nav的序列相似性均高达80%以上,这为MuNav功能研究提供了重要参考。值得一提的是,豆大蓟马与其他蓟马类昆虫序列的相似性极高,如西花蓟马和棕榈蓟马的序列相似性高达94%以上,表明缨翅目蓟马类昆虫Nav的结构与功能极为保守。另外,基于NCBI上获取得到的59种昆虫Nav序列,构建了系统发育树。与上述序列比对结果类似,进化树结果表明虽然昆虫Nav具有一定的保守性,但豆大蓟马与其他蓟马类昆虫聚为一类,表明且它们在进化过程中极为保守,即发生的变异较小,而与其他昆虫则形成了不同的分支,表明它们发生了一定程度的分化。
昆虫Nav可通过转录后修饰,即选择性剪切和RNA编辑实现其功能的多样性[17-18]。RNA编辑是mRNA在转录水平上通过碱基插入、缺失或替换,从而引起氨基酸改变,扩大了生物的遗传信息,增加Nav结构与功能的多样性,从而帮助其迅速适应外界环境的变化,其研究在过去的几年里逐渐得到重视[17,19-20]。昆虫Nav RNA编辑存在2种形式:一种是腺苷A去氨基转变为次黄嘌呤I,即A至I编辑,另外一种是U至C编辑。其中A至I的RNA编辑在昆虫中尤为常见,可导致蛋白质的结构和功能发生改变。如在黑腹果蝇、德国小蠊和绿盲蝽中均被鉴定到,并可引起昆虫神经性兴奋[21-23]。昆虫RNA编辑存在组织特异性,A-I RNA编辑主要发生在编码离子通道、神经递质受体或G蛋白偶联受体的神经系统基因转录本中[24-25]。例如,在黑腹果蝇钠通道转录本中发现了11个A-I编辑[26],在拟果蝇中发现了3个A-I的RNA编辑,分别是Q52R、C189Y和N1260D[27],而在德国小蠊和绿盲蝽Nav中也分别发现了2个和1个A-I RNA编辑[21,23]。此外,昆虫Nav还存在U-C编辑,该类型的RNA编辑一般存在卵巢和肠道中,如黑腹果蝇和德国小蠊体内F1919S位点的U-C编辑可产生持续电流[20]。
近年来,随着田间用药量的增加,昆虫对拟除虫菊酯的抗药性不断增加,Nav突变也越来越频繁,在很多高抗药性种群中发现多个突变位点。大部分昆虫中最常见的突变发生在 1 014位点,该位点在家蝇中最早被发现,由L突变为F,使家蝇对氯菊酯的敏感性降低了至少10倍,并且增加了拟除虫菊酯诱导的钠尾电流的衰减率[28]。除了家蝇外,在冈比亚按蚊(Anopheles gambiae)、人蚤(Pulex irritans)、中华按蚊(Anopheles sinensis)、致倦库蚊Culex quinquefasciatus)等昆虫中也发现了L1014突变[29-32]。除了1 014位点外,918位点氨基酸发生改变也会影响拟除虫菊酯药剂的敏感性。1999年,LEE等[33]对家蝇钠通道918位点进行定点修饰,用苏氨酸代替甲硫氨酸,通过双电极电压钳进行验证,发现其对高浓度的氯氰菊酯完全不敏感。在白纹伊蚊体内发现了I1532T和F1534S/L,当这2个突变单独出现时显著降低了对I型拟除虫菊酯类药剂氯菊酯和联苯菊酯的敏感性,但是对Ⅱ型拟除虫菊酯类药剂敏感性没有显著影响[34]。本研究未在MuNav中发现918和1 014位点突变,但在对序列分析时发现了T929I突变,其在对拟除虫菊酯杀虫剂产生抗性过程中的具体作用还需进一步实验探究。
在高抗种群中,突变点往往都不是单个出现的,它们会同时出现2个突变点,大部分情况下该类型突往往比单点突变更高的抗药性。在蚊子中,Nav中单独的N1575Y突变不会改变昆虫对拟除虫菊的敏感性,但N1575Y和L1014F双突变的种群对氯氰菊酯的抗性是野生型的80倍,对溴氰菊酯的抗性是野生型的53倍,相较于单突变分别增加了3.4倍和9.8倍[35]。在烟蓟马中,T929I的单突变会导致烟蓟马对氯氰菊酯产生中等水平抗性,当T929I和K1774N双突变共同出现会导致烟蓟马对氯氰菊酯产生高等水平抗性(RR=2 700)[36]。反之,某些Nav双突变时,并不会降低钠通道的抗性,例如在埃及伊蚊体内,无论是V1023G/S996P还是V1023G/D1794Y对拟除虫菊酯的抗性都没有单一突变体高[37]。此外,有些位点单独突变不会改变钠通道的敏感性,但会显著降低其他位点对拟除虫菊酯杀虫剂的敏感性。例如,蟑螂体内E434K和C764R单独出现不会改变钠通道对溴氰菊酯的敏感性,但是E434K或C764R与L993F双突变时钠通道对溴氰菊酯的敏感性会降低100倍[38]。除单突变,双突变外,昆虫钠通道有时还会出现三突变,例如在蟑螂Nav中引入V409M突变,可使其对溴氰菊酯的敏感性降低了10倍,而当V409M、E434K、C764R 3个突变同时出现时,蟑螂对溴氰菊酯的敏感性降低了100倍[39]。除上述突变类型,昆虫体内Nav突变还会出现连锁反应,在致倦库蚊中,M943V和I973T就存在着明显的连锁反应,但是其生理意义还需进一步探索[40]。虽然在MuNav中未发现918和1 014位点突变,但豆大蓟马钠离子通道出现众多潜在的抗性位点(未发表),仍需用双电极电压钳技术进一步验证。
本研究克隆了豆大蓟马钠离子通道全长序列,不仅扩展了昆虫钠离子通道的遗传信息,也为后续豆大蓟马抗性位点的鉴定,kdr抗性突变位点分子诊断技术的研发和探明化学杀虫剂与MuNav的结合能力奠定了基础,同时为延缓豆大蓟马抗药性的发展及抗药性治理指明了方向。
Cloning and analysis of sodium channel of Megalurothrips usitatus
doi: 10.15886/j.cnki.rdswxb.2022.03.007
- Received Date: 2022-01-15
- Accepted Date: 2022-02-28
- Rev Recd Date: 2022-02-26
- Available Online: 2022-03-09
- Publish Date: 2022-05-23
-
Key words:
- Megalurothrips usitatus /
- sodium channel /
- gene cloning /
- sequence analysis
Abstract: The strong concealment and difficult control of Megalurothrips usitatus lead to the extensive use of pyrethroids, which results in continuously increase in pesiticidal resistance of M. usitatus population in the field. The target of pyrethroid to pest insects is the sodium channel. Therefore, it is of great significance to explore the characteristics of M. usitatus sodium channel (MuNav) and analyze its resistance molecular mechanism. MuNav (Accession number: MZ043856) was cloned by PCR with a full-length of 6 279 bp, which encodes 2093aa. MuNav is embedded with four homologous domains containing six transmembrane fragments. Homologous comparison showed that the similarity of sodium channel between M. usitatus and Thrips palmi was 94.88%, indicating their close relationship.
| Citation: | PAN Xuelian, YANG Lei, YUAN Linlin, CHEN Weilong, WU Shaoying. Cloning and analysis of sodium channel of Megalurothrips usitatus[J]. Journal of Tropical Biology, 2022, 13(3): 249-258. doi: 10.15886/j.cnki.rdswxb.2022.03.007

|
 DownLoad:
DownLoad: