
-
热带雨林分布在赤道附近降雨充沛的热带地区,是1种具有独特外貌和群落结构,物种多样性很高的森林生态系统[1-2]。热带雨林在调节局部与全球气候、维持生物多样性和生态平衡等方面发挥着重要作用[3]。我国热带地区位于亚洲热带北缘,是东南亚热带雨林向北延伸的边界,主要包括西藏东南部、云南和广西南部、台湾南部和海南岛,其中,海南岛具有我国分布最集中、连片面积最大的热带雨林[4-6]。龙脑香科(Dipterocarpaceae)是海南岛热带雨林的标志性树种,广泛分布在海南岛700 m以下的低海拔山地[7]。胡玉佳[7]根据群落结构和生态外貌特征,将海南岛的龙脑香群落分为混合群落与单优群落,后者包括单优青皮林与单优无翼坡垒林。与原生演替形成的单优青皮林不同,单优无翼坡垒林是经过多次人为干扰破坏后,通过次生演替形成的[8]。自20世纪70年代在甘什岭发现无翼坡垒以来,许多学者围绕单优无翼坡垒林开展了种群、群落和区系研究。胡玉佳[7]在1983年调查了无翼坡垒林的物种组成,发现群落的物种数较少,森林尚处于更新阶段。最近,邢福武等[9]和漆良华等[10]再次调查了无翼坡垒群落,发现物种数有所增长,表明甘什岭保护区在物种保护上发挥了积极的作用。杨小波等[11-13]研究了无翼坡垒的种群结构,分布格局,物种多样性和空间配置,确定无翼坡垒为增长型种群,但自疏作用强烈,生长到一定阶段后个体数量减少,种群的集群程度变小。胡璇等[14]针对无翼坡垒种群结构与动态的最新研究显示,老龄树的比例较25年前有所增长,种群结构在向好的方向发展,但无翼坡垒的幼龄树对病害敏感,且生长受磷元素限制[15-17]。光合生理特性研究显示,虽然无翼坡垒为耐阴树种,但到一定时期后需要充足的光照才能生长更好,否则将进入长时间的蹲苗期[18]。林中无翼坡垒的密度大,光照和养分条件差,种内竞争激烈,群落总体上仍处在不稳定阶段[8]。加上分布范围局限,适生区面积狭小[19],国际自然保护联盟(IUCN)将无翼坡垒评级为极度濒危种(
https://www.iucnredlist.org/species/33393/9776515 ),所以海南岛无翼坡垒的生存状况仍然不容乐观。现有报道主要集中在无翼坡垒种群与群落生态的研究上,缺乏遗传多样性的评估与分析。种群遗传变异是阐明物种进化历史和生态适应的基础[20],是评价物种存活潜力、解释濒危原因的重要依据[21-22]。因此,研究濒危物种的遗传多样性和遗传结构是制定其保护策略的前提。简单重复序列(simple sequence repeats,SSR) [23],也称为微卫星(microsatellite),是基因组中以1~6个核苷酸为单位的串联重复序列,长度一般在100~200 bp之间。因其多态性高、易于检测等优点[24],广泛应用于濒危物种的保护研究[25-26]。龙脑香科濒危树种常用微卫星标记评估遗传多样性,用以指导保护管理。LEE等[27]发现娑罗双属的极度濒危种Shorea lumutensis仍具有较高水平的微卫星变异,但因个体数量不足500,建议通过就地保护与迁地保护相结合的方式实施物种保护。坡垒属的极度濒危种狭叶坡垒(Hopea chinensis)仅在越南广宁和中国广西发现,TRANG等[28]使用SSR标记检测到种群瓶颈和高水平近交,揭示种群规模缩小是狭叶坡垒濒危的主要原因。WANG等[29]利用12对SSR标记评估了海南岛濒危植物坡垒(H. hainanensis)的遗传多样性,发现坡垒的微卫星变异水平明显低于同属的非濒危种H. dryobalanoides,推断种群瓶颈是坡垒遗传变异丧失的可能原因。综上所述,SSR标记是研究龙脑香科濒危物种保护的有效分子标记。本研究利用11个微卫星标记度量海南甘什岭无翼坡垒种群的遗传多样性,并分析种群遗传结构,评估无翼坡垒遗传变异下降的程度,旨在为制定无翼坡垒合理的保护措施提供遗传学依据。
HTML
-
研究区域位于海南省甘什岭省级自然保护区(109°34′~109°42′ E, 18°21′~18°26′N)。甘什岭属低山丘陵地貌,海拔约50~681 m,坡度<50°,土壤母质以花岗岩为主,岩层裸露率10%,属热带海洋季风气候,干湿季分明,雨季集中在5~10 月,年降雨量约为1 800 mm。年平均气温25.4 ℃,地带性森林类型是以无翼坡垒为优势种的热带低地雨林[9-10]。甘什岭保护区建立于1985年,主要保护目标是单优无翼坡垒林。本研究根据胸径将无翼坡垒划分为3个龄级:幼龄(Ⅰ)(胸径 ≤ 2.5 cm),中龄(Ⅱ)(2.5 cm < 胸径 ≤ 7.5 cm)和成熟龄(Ⅲ)(胸径 >7.5 cm)。在面积2 500 m2的样方内,随机选取59株空间上均匀分布的无翼坡垒,3种龄级分别有21(Ⅰ)、20(Ⅱ)和18(Ⅲ)株。选择无病害的嫩叶,采下后立即用硅胶干燥,保存备用。并记录采样个体的树高和胸径。
-
采用改良CTAB法提取基因组DNA[30]。基因组DNA的浓度和质量用NanoDrop 2000分光光度计测量。本实验利用WANG等[31]针对坡垒设计的35对微卫星引物,从中筛选出在无翼坡垒中能稳定扩增,且多态的11对引物用于后续试验。PCR扩增在Eppendorf的热循环仪中进行。总反应体积30 μL,由1 μL的模板DNA(50 μg·mL−1),1 μL正反向引物(10 μmol·L−1),15 μL 2× Taq PCR MasterMix(TIANGEN生物技术,北京)和12 μL ddH2O。循环程序如下:首先94 ˚C预变性4 min,然后循环30次,其中94 ˚C变性30 s,56~63 ˚C退火30 s,72 ˚C延伸1 min,循环结束后再72 ˚C延伸12 min。使用ABI 3730XL分析仪(Applied Biosystems)进行片段分析,使用GeneMarker软件进行微卫星位点的基因分型(SoftGenetics, State College, Pennsylvania, USA)。
-
无翼坡垒为同源四倍体,因此本研究采用MAC-PR方法,根据峰值强度比值确定等位基因剂量[32]。由于MAC-PR方法不能完全解决等位基因剂量未知导致基因分型的不确定性,同时考虑到多倍体物种在减数分裂时可能发生的双减数,使标准群体遗传分析结果出现偏差[33]。因此,笔者用GENDIVE version3.04[34]和POLYGENE version1.2b[35]2个软件解决因剂量信息缺失及减数分裂时发生双减数等产生的问题。在POLYGENE version 1.2b软件实现了4种多体遗传模型,根据贝叶斯信息准则(Bayesian information criterion, BIC)选择最优模型,基于最优模型计算遗传多样性的各项指标,如等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)和期望杂合度(He)。
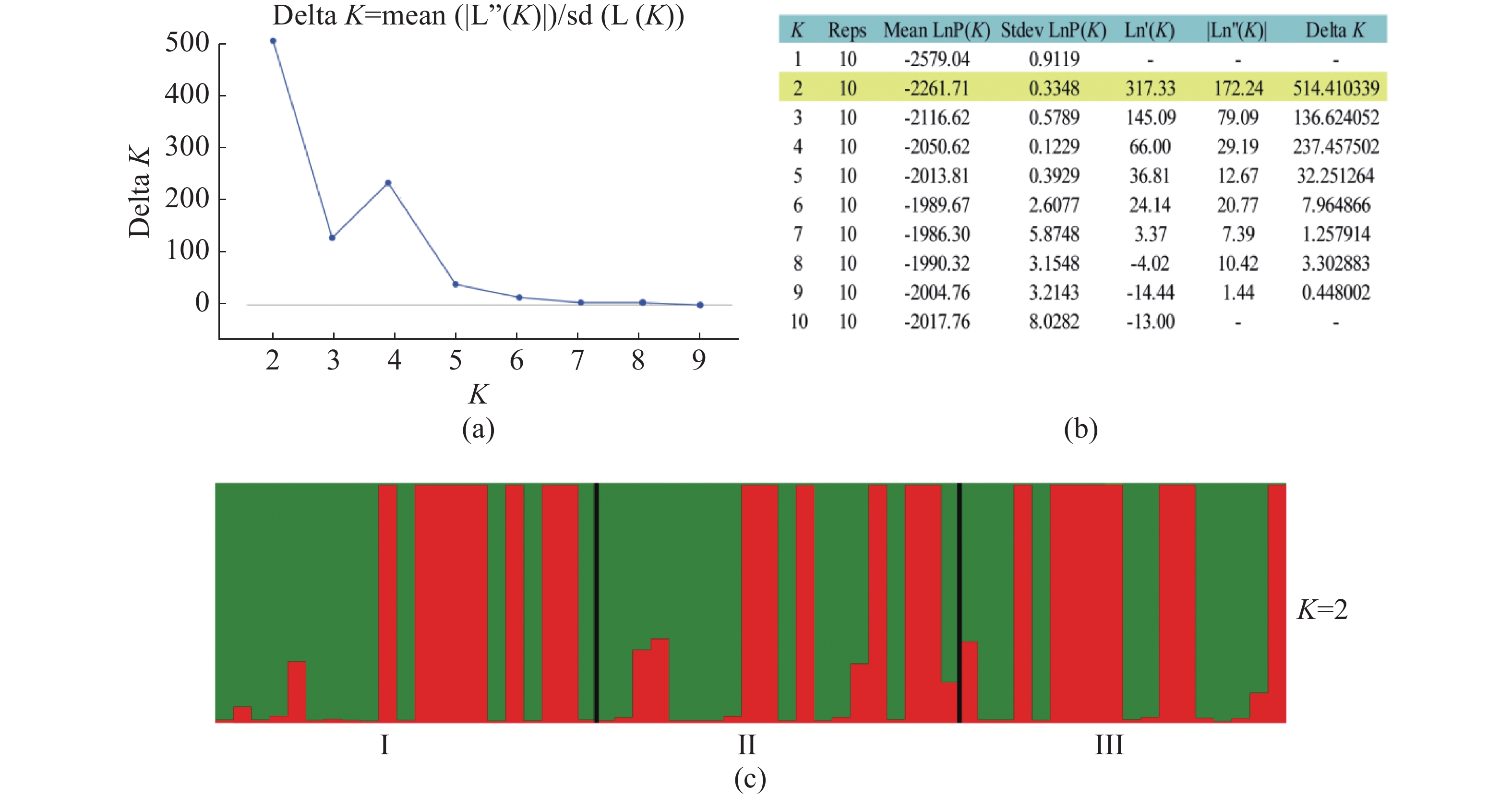
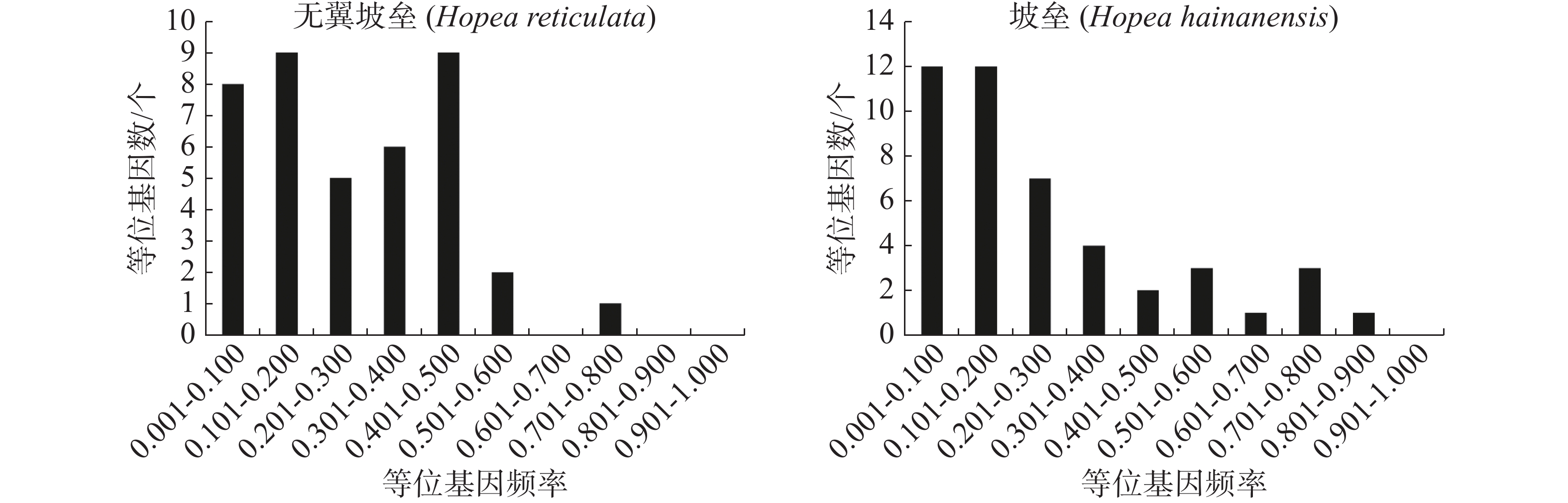
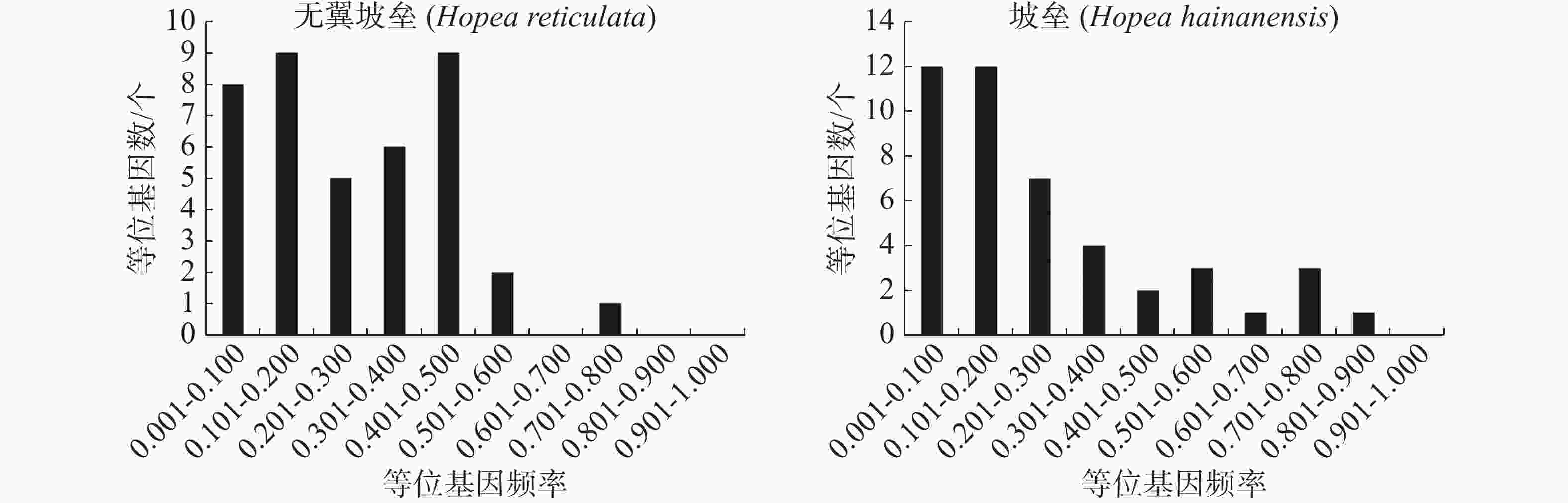
使用Structure version 2.3.4[36]分析种群的遗传结构。K值从1到10,每个K值进行10次独立运算,burn-in设置为100 000次,然后是1 000 000次MCMC(马尔科夫链蒙特卡罗)迭代。最佳K值使用STRUCTURE Harvester程序推断[37]。使用Clumpp version 1.1.2[38]的Greedy算法重排最优K值的10次重复结果,最后由Distruct version 1.1[39]生成种群结构的图形表示。基于Cavalli-Sforza[40]的遗传距离,使用MEGA 5.0软件包[41]构建无翼坡垒个体的邻接树。根据Cavalli-Sforza的弦距离做主坐标分析(PCoA),当等位基因的剂量未知时,弦距离是偏差最小的距离测度[42]。将微卫星等位基因频率划分为10个等级(0~0.1, 0.1~0.2, ..., 0.9~1.0),使用POLYGENE的最佳多体遗传模型估计各个等位基因的频率,绘制微卫星等位基因频率的直方分布图。采用定性的图形法确定无翼坡垒是否经历了种群瓶颈[43]。图解法的基本原理是,与普通等位基因相比,稀有等位基因会在瓶颈期间迅速丢失。因此,无论突变率和模型如何,在出现瓶颈后,低频( < 0.1)的等位基因都会比中频(0.101~0.200)的等位基因少。
1.1. 无翼坡垒种群样本采集
1.2. DNA提取和PCR扩增
1.3. 数据分析
-
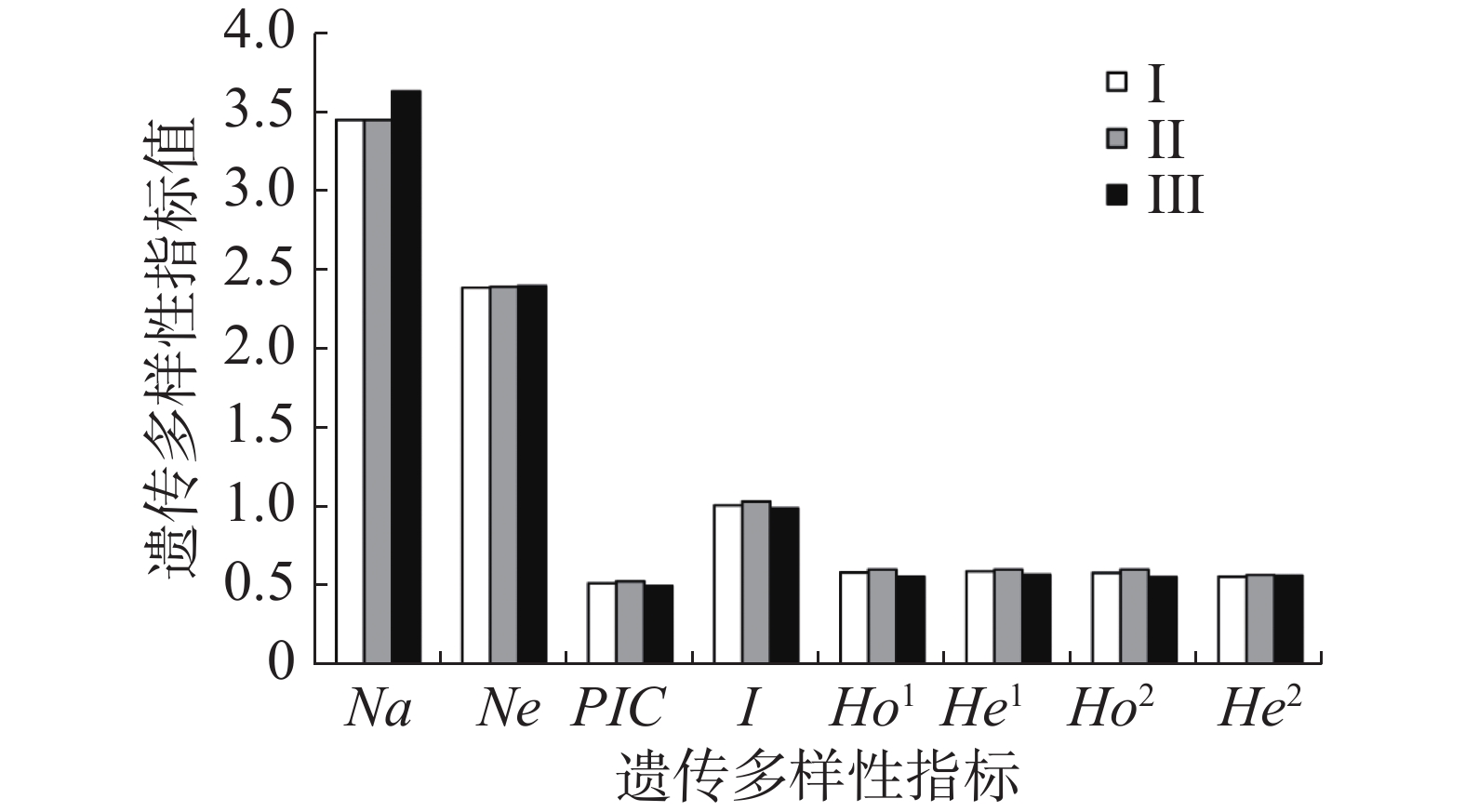
针对坡垒开发的35个微卫星标记中,11个能在无翼坡垒中稳定扩增且有多态性,用于无翼坡垒遗传多样性分析,其中,扩增产物长度在75~190 bp之间,最小重复单元为15,最大重复单元为30(表1)。POLYGENE软件首先确定最优多体遗传模型,根据BIC信息标准,选择BIC值最小的为最优遗传模型,结果显示,CES(Complete equational segregation,完全等分式分离)为无翼坡垒的最优多体遗传模型,非自交无负PCR为最佳参数,后续种群遗传参数的计算将基于CES模型进行(表2)。由POLYGENE估计的等位基因个数在2(Hre9)至7(Hre11)之间,平均3.636个;有效等位基因的个数在1.428(Hre9)至3.282(Hre11)之间,平均2.380个。多态信息含量0.289(Hre9)至0.716(Hre11),平均值0.526;Shannon信息指数0.535(Hre9)至1.589(Hre11),平均值1.035。观测杂合度最小0.343(Hre9),最大0.698(Hre6、Hre7),平均值0.585。期望杂合度最小0.350(Hre9),最大0.751(Hre11),平均值0.599。GENODIVE计算的观测杂合度在0.342(Hre9)至0.697(Hre7)之间,平均值为0.583;期望杂合度在0.303(Hre9)至0.709(Hre11)之间,平均值为0.567。POLYGENE和GENODIVE估算的遗传多样性很接近,显示多样性的估计值是可靠的。11个位点中有9个(除Hre2和Hre11)显著偏离Hardy-Weinberg平衡,可能是多体遗传或近交导致的。通过POLYGENE和GENODIVE估算了无翼坡垒3个龄级的遗传多样性,结果显示,无翼坡垒在不同龄级的有效等位基因个数、多态信息含量、香农指数、观测杂合度以及期望杂合度上没有显著差异,遗传多样性水平基本相同(图1)。
位点 引物序列 (5′–3′) 重复单元 产物长度/
bp等位基因
数(Na)/个POLYGENE GENODIVE 观测杂
合度
(Ho)期望杂
合度
(He)多态信息
含量
(PIC)香农
指数
(I)有效等位
基因数
(Ne)观测杂
合度
(Ho)期望杂
合度
(He)近郊
系数
(Gis)哈温
平衡Hre1 F:ACATGGTCTTTGTTATCTGCTTA TTCT(4*7) 140~180 3 0.664 0.628 0.550 1.035 2.792 0.663 0.650 −0.020 −0.048 R:CCATGGTGCTACAACCTTTCTTG Hre2 F:TTCATGGTCATTGAGTCATAGGT AT(2*10) 120~170 4 0.525 0.613 0.545 1.059 2.207 0.525 0.555 0.053 0.144 R:GCCTCTACCTAGTGTATGAAGGC Hre3 F:TGCTATTCTACCCTAAAAACCCA AAAATA(6*4) 135~170 3 0.601 0.540 0.433 0.839 2.039 0.600 0.515 −0.166 −0.144 R:TGGTTGATGCTCTCACAAGCTAT Hre4 F:GATGAGGGATAATGGTGCGTTTG AAG(3*5) 100~150 3 0.561 0.538 0.436 0.845 1.901 0.555 0.480 −0.157 −0.085 R:CAACTCACGCCTCTGTGTTATTG Hre5 F:AAGTCACCTCCATCCATTTCTCC CTG(3*5) 120~150 3 0.622 0.586 0.497 0.952 2.226 0.618 0.557 −0.108 −0.131 R:ATGCGGTGAAGAATCATTGGAAC Hre6 F:CCTGGAAATCAAAGATGAATCAGCT AC(2*8) 140~170 4 0.698 0.666 0.604 1.208 2.879 0.694 0.661 −0.050 −0.096 R:TTGGGGAGGGTAAATAGCAGATG Hre7 F:CCGTGGGAGATAGGACATCATTA GA(2*7) 140~170 4 0.698 0.616 0.540 1.061 2.539 0.697 0.614 −0.134 −0.230 R:GTTGTGCTTTCACTTATTCATCCCT Hre8 F:ATTGTGGATGTAGCTATGGTGAC TTA(3*5) 140~195 3 0.630 0.646 0.573 1.068 2.607 0.629 0.625 −0.007 0.020 R:TGTACGGACATTTGGTGGAATCA Hre9 F:AGTTGGAGATTAAAGAAAGTGGCT TTTTA(5*6) 75~115 2 0.343 0.350 0.289 0.535 1.428 0.342 0.303 −0.128 −0.030 R:TTCAATTTAGACCCGTGGACCTC Hre10 F:TGAGATTCACATGGTTACTGGAA AT(2*8) 80~135 4 0.669 0.653 0.598 1.192 2.285 0.671 0.569 −0.181 −0.033 R:ACCATAATTCAAGAAGCATACGCA Hre11 F:GCTTTCTGCATTTCCTTGAGAGA AT(2*9) 135~170 7 0.427 0.751 0.716 1.589 3.282 0.420 0.709 0.407 0.402 R:TGATTAGCTGCTGAATTTGGCTG 参数 多体遗传模型 染色体
随机
分离染色单体
随机
分离完全
等分
分离部分等
分分离,
rs=0.25部分等
分分离单,
rs=0.50自交与
负PCR3017.62 2960.57 2957.34 2992.52 2975.30 仅自交 2803.93 2746.88 2743.65 2778.83 2761.61 仅负PCR 2997.00 2940.00 2937.33 2972.35 2955.37 非自交
无负PCR2783.31 2726.97 2723.64 2758.66 2741.68 注:rs为单染色单体重组率。 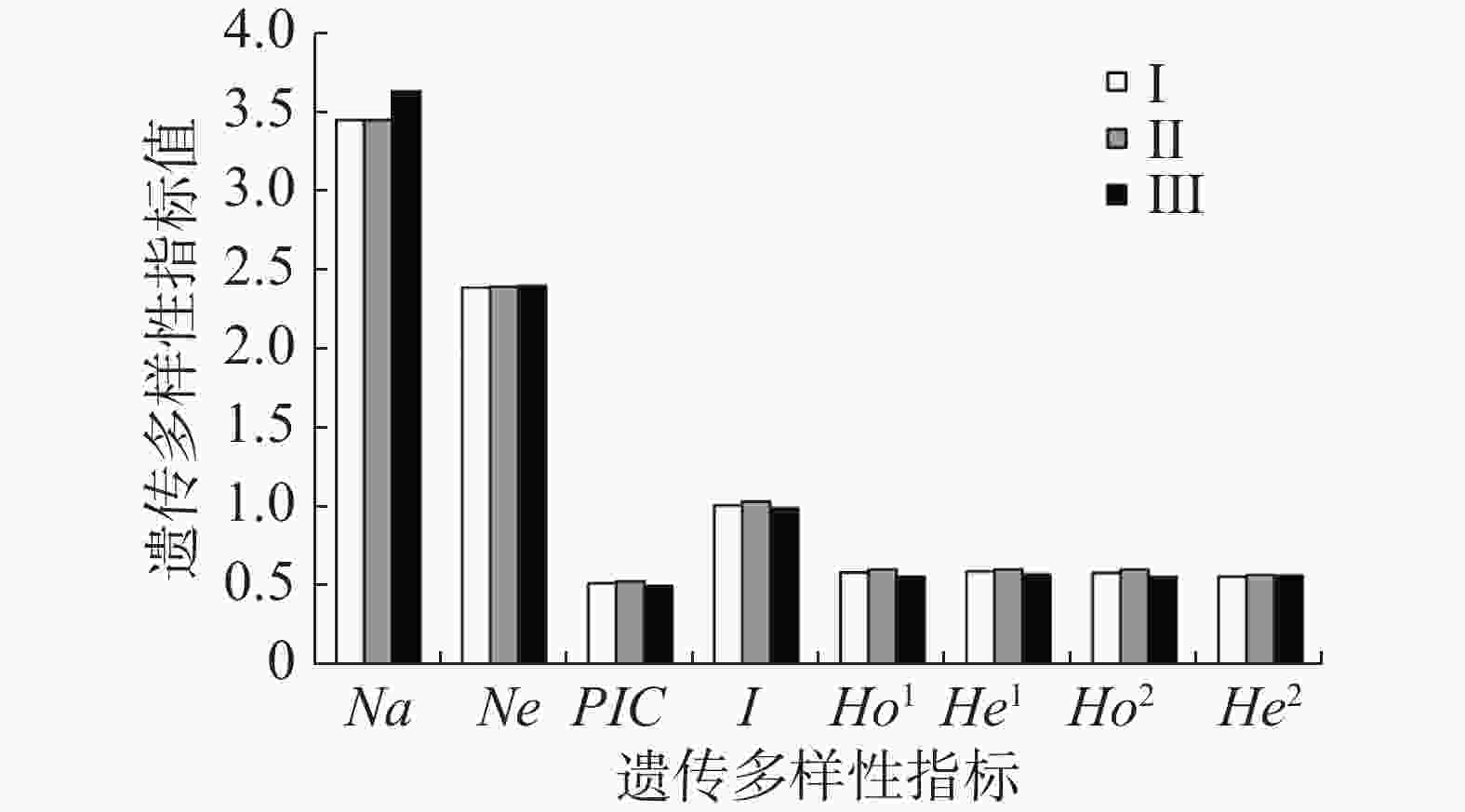
-
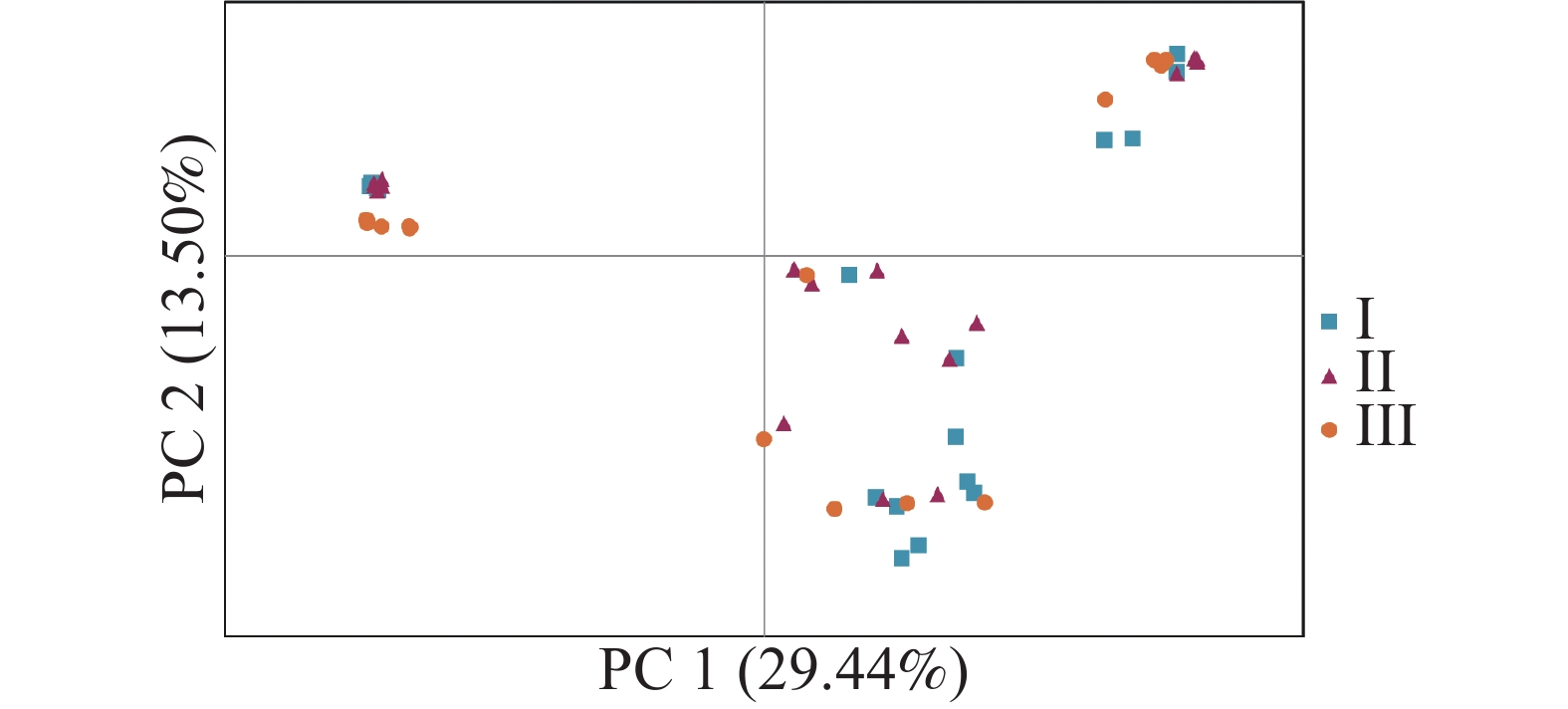
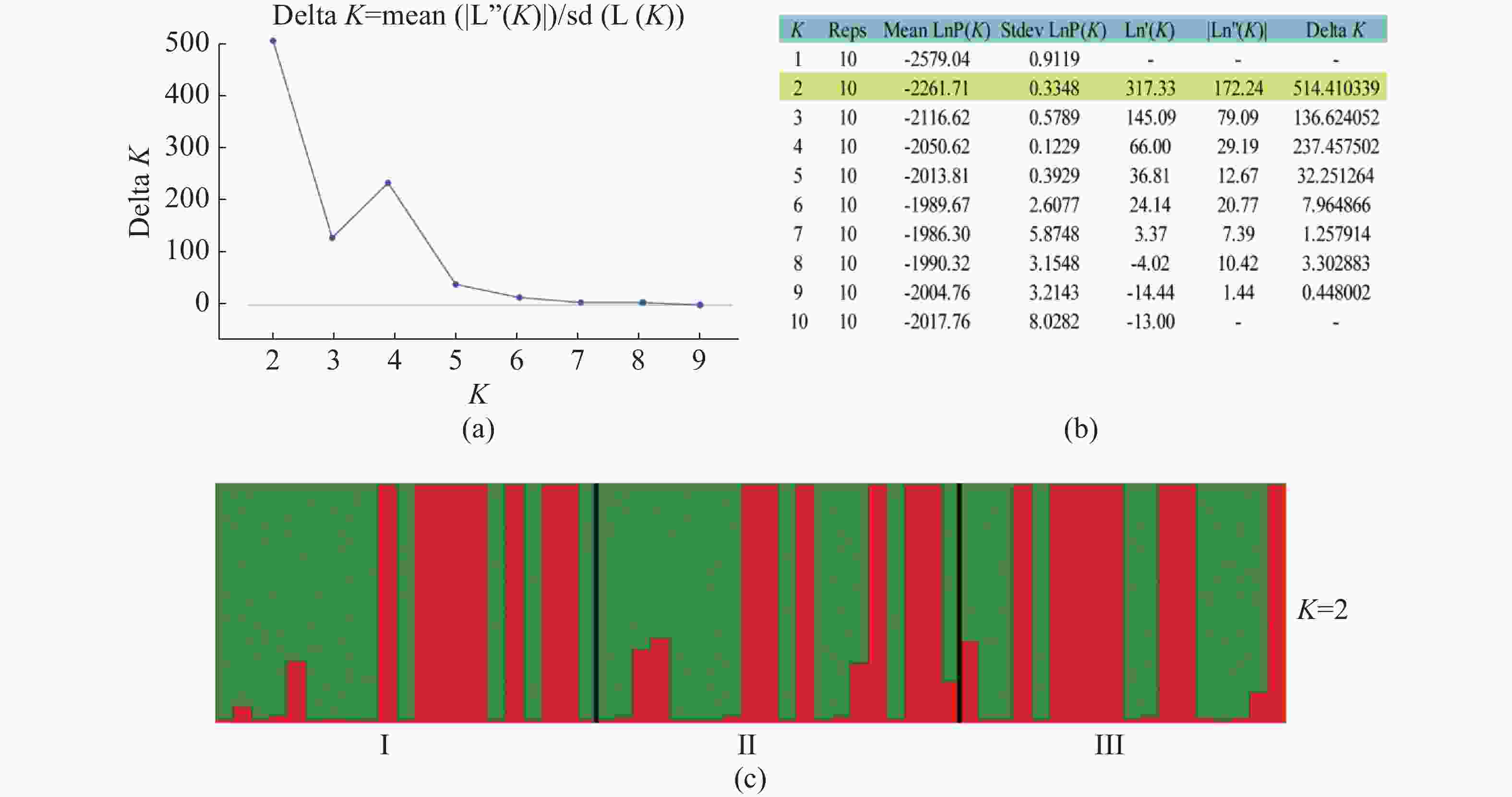
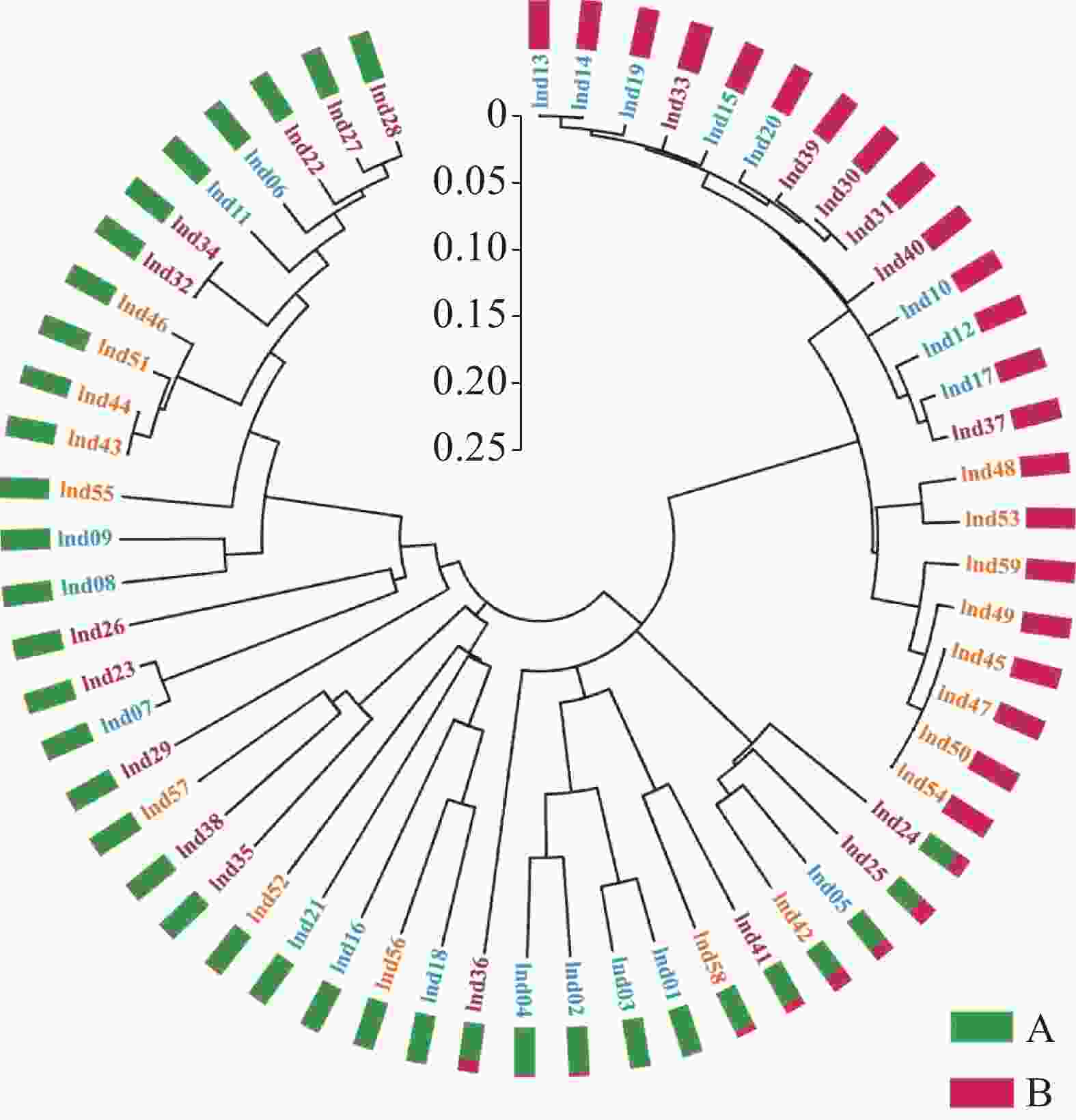
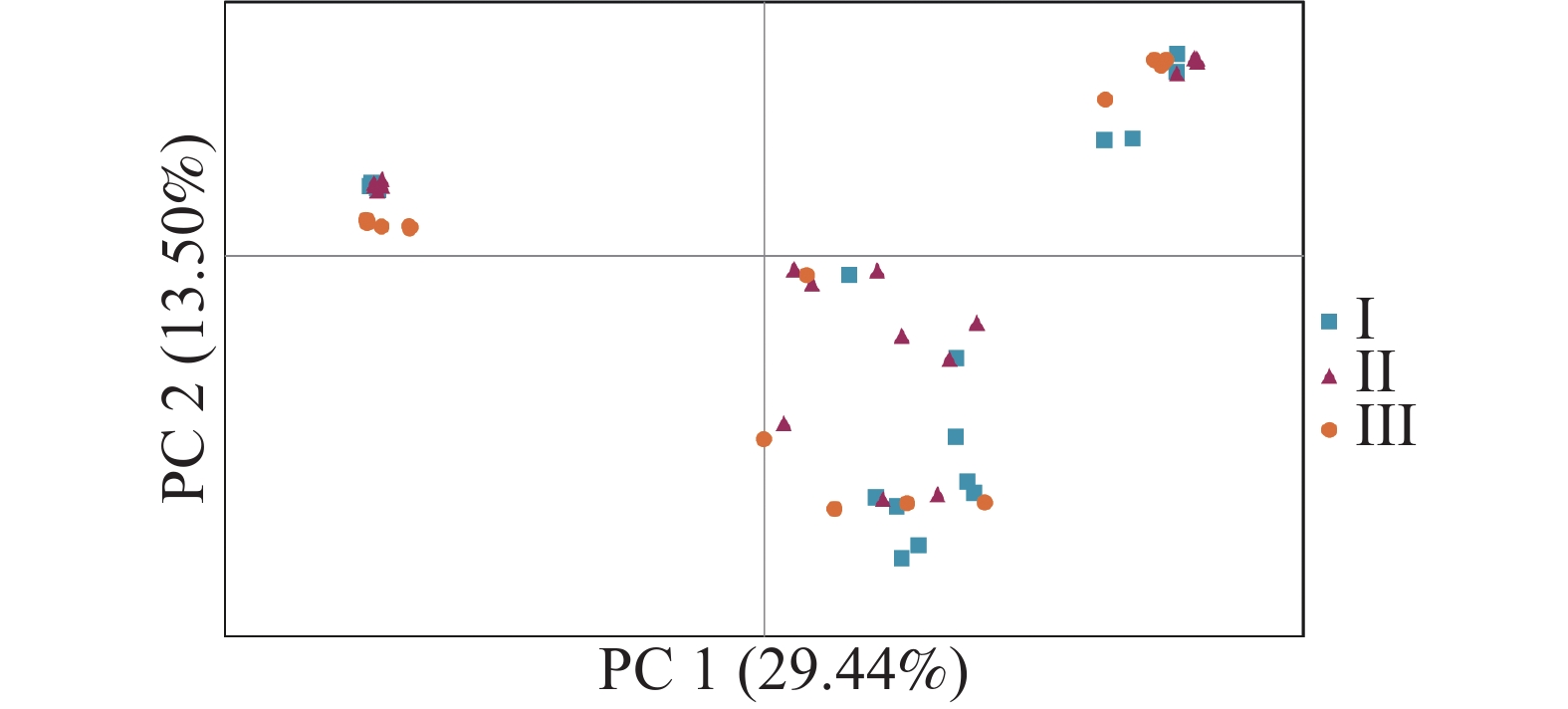
无翼坡垒遗传结构的Structure分析结果(图2)显示,delta K在K = 2时取得最大值,故最优K值为2,即祖先种群由2种遗传组分构成。3个龄级的遗传组成没有明显分化,均包含2种组分,除少量个体是2种组分的混合外,大部分个体只有单一遗传组分(图2)。为进一步分析3个龄级的关系,根据Cavalli -Sforza(1967)的公式计算遗传距离并构建NJ树(图3)。无翼坡垒3个龄级的个体聚为两大支,不同龄级的个体分散在两大支中,不同龄级之间未显示出明显分化,与Structure的结果一致。PCoA分析的第1主坐标(PC1)和第2主坐标(PC2)分别解释了29.44%和13.50%的变异,所有个体大致分为2~3个遗传组,每个遗传组中均包含3个龄级的个体(图4)。PCoA的结果与Structure和NJ树的结果基本一致,进一步说明无翼坡垒龄级间的遗传差异很小,随着龄级增加,没有出现遗传组成的明显变化。无翼坡垒低频等位基因(0~0.100,0.101~0.200)的比例与经历瓶颈的坡垒种群相似,与未经历瓶颈的其他龙脑香科种群相比显著减少[29](图5)。这种等位基因频率分布的变形,低频等位基因的丧失,是种群经历瓶颈的1个特征,表明无翼坡垒可能最近发生了种群规模的缩减,导致大量低频等位基因丢失,因此,等位基因频率分布出现偏移。
2.1. 无翼坡垒的遗传多样性
2.2. 无翼坡垒的遗传结构
-
遗传多样性对于物种进化和适应有重要影响[44-45]。物种的遗传多样性水平越低,适应环境的能力越差,灭绝的风险越大[46]。一般认为,濒危、特有和狭域分布的植物遗传多样性较低[47]。海南岛的坡垒属有坡垒和无翼坡垒2种,WANG等 [29]采集了10个位于海南热带雨林国家公园内不同地点的坡垒种群,利用12对微卫星标记评估坡垒的遗传多样性,结果显示,坡垒的期望杂合度(He)为0.409,与同属其他濒危种如狭叶坡垒(Hopea chinensis)(He=0.473)、H. odorata(He=0.392)[48]的遗传多样性水平相当。然而,海南岛无翼坡垒的期望杂合度的均值为0.567(GenoDive)或0.600(Polygene),明显高于同域分布的坡垒(He=0.409)。无翼坡垒微卫星位点的等位基因个数(Na=3.515, Ne=2.397)也比坡垒(Na =2.458, Ne =1.964)多。尽管伐木等干扰导致无翼坡垒数量减小,但因剩余个体较多,有效种群规模仍然较大,与零星分布、个体数量稀少的坡垒相比,无翼坡垒能够维持相对更丰富的遗传变异。当然与非濒危种H. dryobalanoides(He =0.67)[49]相比,无翼坡垒的遗传多样性仍是降低的。无翼坡垒与坡垒具有类似的微卫星等位基因频率分布,低频等位基因的数量都显著减少,显示出清晰的瓶颈信号。综上所述,无翼坡垒很可能经历了种群瓶颈,导致低频等位基因大量丢失,遗传多样性降低。
不同龄级的无翼坡垒代表了不同时期的种群。成熟无翼坡垒存活的时间长,代表了较早时期的种群;中龄个体可能是干扰时期或干扰停止后产生的;而幼龄无翼坡垒是经历干扰后存活个体繁殖的年轻后代。因此推测成熟无翼坡垒的遗传多样性比中龄和幼龄的更高。然而3个龄级的遗传多样性没有明显差异。这一结果的可能解释是种群瓶颈持续的时间长,直至保护区建立人为干扰才得以消除,因此,目前处于成熟龄的无翼坡垒,其遗传多样性已经遭到破坏,故没有表现出比中龄与幼龄个体更高水平的遗传变异。基于主坐标分析和贝叶斯聚类的遗传结构分析也再次证实不同龄级的无翼坡垒不存在明显的遗传分化。考虑到微卫星提供的进化信息有限,计划将使用全基因SNP变异数据进一步研究无翼坡垒瓶颈开始与结束时间,以及瓶颈强度[50]。
无翼坡垒濒危的主要原因是长期的人为干扰导致种群规模缩小,遗传变异大量丧失,适应环境变化的能力减弱。另一方面,无翼坡垒果实无翅,传播距离很短,使得幼苗密度大,竞争激烈,加上冠层遮挡,难以成长为成年个体。要保护无翼坡垒,实现长期存活,首先要恢复其遗传多样性,增加种群的进化适应潜力。建议在就地保护的基础上,进行远距离个体杂交授粉,提高种苗的遗传变异。同时进行迁地保护,增加无翼坡垒的生存地点,避免因偶然因素导致原生地种群破坏而引起物种灭绝。
 DownLoad:
DownLoad: