
-
文心兰是兰科(Orchidaceae)文心兰亚族(Subtribe Oncidiinae)文心兰属(Oncidium)的植物,又名舞女兰、跳舞兰、金蝶兰、瘤瓣兰,是具有极高观赏价值的热带兰花[1]。其中,开黄色花的南茜种文心兰(Oncidium Gower Ramsey)是栽培最广的热带兰切花品种。鲜切花的寿命长短是一个重要的品质指标,对文心兰切花的商品价值具有重要影响。因此,鉴定调控花瓣衰老的关键基因,研究其花朵衰老的分子机制,可为采用生物技术改良品种,在分子水平上控制切花衰老快慢和寿命长短打下基础[2-8]。反应调节因子(response regulator, RR)为His→Asp磷酸转移系统或双组分信号转导系统的下游组成成分。植物RR广泛影响植物的许多生长、发育和胁迫反应过程。研究表明,拟南芥中有23种拟南芥反应调节因子(Arabiopsis thaliana response regulators, ARRs),根据结构和功能分为A类ARRs和B类ARRs[9-12]。A类ARRs大多与细胞分裂素传导途径相关,表达受氮水平变化、盐处理、低温等影响,其中,拟南芥ARR4还与光信号传导以及下胚轴生长有关;B类ARRs是定位于细胞核中的转录因子,大多对细胞分裂素不敏感,但可作用于A类ARRs[10-13]。文心兰花瓣发育和衰老转录组学研究表明,文心兰RR基因可能参与了文心兰花瓣的衰老启动[14]。为了鉴定控制文心兰花瓣衰老的关键基因的结构和功能,首先需要对其进行克隆和表达分析。此前,笔者已对文心兰OnRR9基因的克隆与表达进行过研究[8],本实验研究了文心兰该基因家族的另一个成员——OnRR10基因的克隆和表达情况,旨在为解决兰花的采后保鲜及衰老问题提供理论依据。
-
本实验材料为文心兰南茜种‘黄金3代’(Oncidium Gower Ramsey ‘Gold 3’)的鲜切花,购自海南出入境检验检疫局热带植物隔离检疫中心。
-
参照文献[15]改良的CTAB-LiCL法提取文心兰花瓣总RNA,用w=1%琼脂糖凝胶电泳检测RNA完整性,并测量RNA浓度。按照PrimeScript RT reagent Kit with gDNA Eraser反转录试剂盒说明书合成cDNA第1条链。
-
根据NCBI上已提交的蝴蝶兰、石斛兰等兰科植物的保守序列设计兼并引物P1和P2 (表1),以合成的cDNA第1条链为模板,扩增OnRR10基因的保守区。反应体系:cDNA,0.5 μL;10 × LA BufferⅡ(Mg2+ plus),5 μL;dNTP Mix (2.5 mmol·L−1),8 μL;上游引物(10 µmol·L−1),0.5 μL;下游引物(10 µmol·L−1),0.5 μL;TaKaRa LA Taq (5 µmol·L−1),0.2 μL,总体积50 μL。反应条件:95 ℃,5 min;(95 ℃, 30 s; 49 ℃, 30 s; 72 ℃, 2 min) 35个循环;72 ℃,10 min;4 ℃,保存。
引物名称 Primer name 引物序列 Primer sequence 用途 Usage P1 5'-CTTCRGCTCCTTCTTCCAAG-3' 保守区扩增
Amplification of
conservative regionP2 5'-GTGGATGACAGCMTCATTGATMG-3' P3 5'-CTGTTGATCCTGGAAGGCAC-3' 5' RACE P4 5'-ATCATACCCAGTCATTYCWGGC-3' P5 5'-TTGACAGATAGCTCATCGAGAGA-3' 3' RACE P6 5'-TGGATTCTGGAAGTAAGGCG-3' QT 5'-CCAGTGAGCAGAGTGACGAGGACTCGAGCTCAAGC-17T-3' 5' RACE随机引物
5' RACE random primerQO 5'-CCAGTGAGCAGAGTGACG-3' QI 5'-GAGGACTCGAGCTCAAGC-3' OnRR10-A1 5'-GGACTAGTCGAGTTACTGCTTGATAATCGTTG-3' 全长序列的扩增
Full-length amplificationOnRR10-S1 5'-CATGCCATGGCCGTAGCTACGGAGG-3' OnRR10-A3 5'-ACTCTTCAAACCCTCGCAGA-3' 实时荧光定量PCR
Real-time fluorescence
quantitative PCROnRR10-S3 5'-TCCTTCCAAGTGACTGCAGT-3' Actin A1 5'-ATCCTCCAATCCAGACACTGT-3' 检测 cDNA Testing cDNA Actin S1 5'-CAGTGGTCGTACAACTGGTAT-3' Actin A2 5'-GGGCATATCCTTCGTAGATTGT-3' qPCR内参
Reference gene qPCRActin S2 5'-AATGTGCCTGCTATGTATGTTGCT-3' Table 1. List of primers used in this study
-
获得保守序列后设计5' RACE引物P3,P4,合成P3,P4以及随机引物QT,QO,QI (表1)。提取文心兰花瓣总RNA,方法同1.2,根据5'经典RACE方法[16]获得cDNA第1条链,并进行巢式反应,获得OnRR10基因5'端序列。
-
设计并合成3' RACE引物P5和P6。提取文心兰花瓣总RNA,方法同1.2。根据TaKaRa 3'-Full RACE Core set with PrimeScript RTase试剂盒说明书操作,获得OnRR10基因3'端序列。
-
将3',5'端片段与之前获得的基因保守区序列进行拼接得到基因全长序列,为了验证拼接得到的全长序列是否正确,分别设计并合成引物OnRR10-A1和OnRR10-S1,以cDNA第1条链为模板,进行PCR反应基因全长序列并测序。PCR反应体系:Premix Taq (TaKaRa Taq version 2.0 plus dye 5 μL,上游引物(10 µmol·L−1) 0.25 μL, 下游引物(10 µmol·L−1) 0.25 μL,cDNA 0.5 μL,总体积10 μL。反应程序: 95 ℃,3 min;30个循环(94 ℃, 30 s; 57 ℃, 30 s; 72 ℃, 30 s);72 ℃,5 min;4 ℃,保存。PCR产物用w=1%琼脂糖凝胶电泳检测。
-
利用SnapGene Viewer 2.2.8进行氨基酸序列预测;通过ExPASy服务器的在线软件ProtParam tool (
https://web.expasy.org/protparam/ )对推导的OnRR10基因的氨基酸序列进行理化性质分析;用ProtComp 9.0 (http://linux1.softberry.com/ )进行蛋白的亚细胞定位进行预测;运用NCBI的CD-Search service工具(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )基因进行蛋白序列的结构域分析;根据ProtFun在线分析程序(http://www.cbs.dtu.dk/services/ProtFun/ )对2个基因进行蛋白功能预测;采用SignalP 4.1程序(http://www.cbs.dtu.dk/services/SignalP-4.1/output.php )进行氨基酸序列的信号肽预测;根据TMHMM在线软件(http://www.cbs.dtu.dk/services/TMHMM/ )预测OnRR10蛋白跨膜区段。利用ProtScale(https://web.expasy.org/protscale/ )对氨基酸的疏水性/亲水性进行预测;用SOPMA SECONDARY STRUCTURE PREDICTION METHOD对蛋白进行2次结构分析;用NCBI的BLAST进行序列比对,并用MEGA6构建系统进化树。 -
根据文心兰南茜种花的形态变化特征,将文心兰南茜种花发育分为花苞期、绽口期、半开放期、盛开前期、盛开期、衰老初期、衰老期和脱落干枯期8个阶段[17]。连续8 d采集生长状况良好的不同时期的花瓣,装于无RNA酶的离心管中,−80 ℃保存备用;将150枝文心兰平均分到3个量杯中,分别在长日照(光照16 h·d)、全光照(光照24 h·d)、全黑暗(光照0 h·d)下处理,采样备用;配制60 mg·L−1的6-BA溶液处理文心兰,1 h内,每隔10 min采样备用。
按照1.2方法提取文心兰不同花期花瓣总RNA,反转录得到cDNA,用Actin A1和Actin S1引物检测cDNA质量。本研究内参基因为Actin A2和Actin S2,基因荧光定量PCR的上下游引物为OnRR10-A3和OnRR10-S3。PCR扩增反应体系:TB Green premix Ex Taq Ⅱ(Tli RNaseH Plus) 10.0 μL,Rox Reference DyeⅡ 0.4 µL,上下游引物(10 µmol·L−1)各0.8 μL,cDNA模板2.0 μL,dd H2O 6.0 μL,总体积20 μL。反应程序:95 ℃,30 s; (95 ℃, 5 s; Tm, 30 s) 40个循环;(95 ℃, 10 s; 0.5 ℃, 5 s)绘制溶解曲线。每对引物设3个无模板对照,不同模板都进行3次重复实验。
-
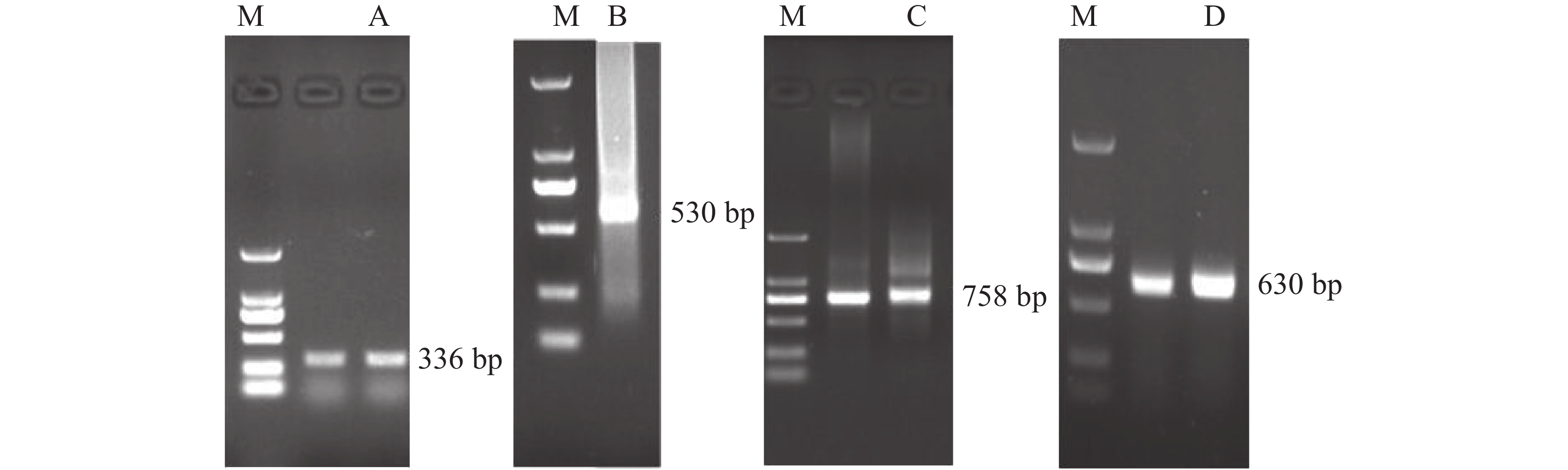
提取质量良好的花瓣总RNA后,反转录获得cDNA。以cDNA为模板,利用保守区引物进行PCR扩增得到OnRR10基因条带,大小约为336 bp (图1A)。经过BLAST比对,发现已知的兰科ORR基因均有较高的同源性,说明目的基因为ORR基因。
Figure 1. Electrophoresis of PCR products by OnRR10 gene of Oncidium
利用获得的保守区序列,分别设计5' RACE和3' RACE引物,经过两轮巢式反应,回收5'经典RACE二轮反应约为530 bp的条带并测序,得到大小为530 bp的序列(图1B)。经过DNAMAN和BLAST比对,为OnRR10基因5'端序列。同样地,回收3'-Full RACE第2轮反应的条带,测序分析,获得OnRR10基因大小为758 bp的3'端序列(图1C)。
经过DNAMAN比对和SnapGene Viewer2.2.8分析得到基因CDS区全长序列。设计引物OnRR10-A1,OnRR10-S1,以cDNA第1条链为模板,进行PCR反应,测序得到OnRR10基因全长约为630 bp (图1D),与预期结果一致。
-
OnRR10基因一级结构预测显示其编码的氨基酸个数为209,等电点为8.76,分子式是C999H1653N283O318S13,相对分子质量是23 133.60 ×103,平均疏水性是−0.40,表明该蛋白也为亲水蛋白,不稳定系数52.33,表明该蛋白不稳定。
用ProtComp 9.0进行OnRR10蛋白的亚细胞定位预测显示OnRR10蛋白定位于细胞核的概率为99.7%,由此推测,OnRR10是在细胞核中发挥转录激活作用的转录因子。
运用NCBI的CD-Search service工具分析OnRR10基因的CDS序列,结果显示,它有1个典型的YesN保守结构域,该保守结构域分布在OnRR10 CDS区的1~618 bp之间(图2),由此推测,OnRR10基因属于YesN家族。

Figure 2. Conserved domains of the OnRR10 gene of Oncidium
用ProtFun在线分析程序预测OnRR10编码蛋白的功能,从表2可知,激素功能的可能性高达17.538。
基因本体分类
Gene Ontology category可能性
Prob几率
Odds信号转导 0.272 1.271 受体 0.008 0.047 激素 0.114 17.538 结构蛋白 0.003 0.107 运载体 0.043 0.394 离子通道 0.079 1.386 电压门控离子通道 0.003 0.136 阳离子通道 0.010 0.217 转录 0.031 0.242 Transcription regulation(转录调控) 0.031 0.248 Stress response(胁迫应答) 0.223 2.534 Immune response(免疫应答) 0.123 1.447 Growth factor(生长因子) 0.040 2.857 Table 2. Prediction of protein functions of Oncidium OnRR10
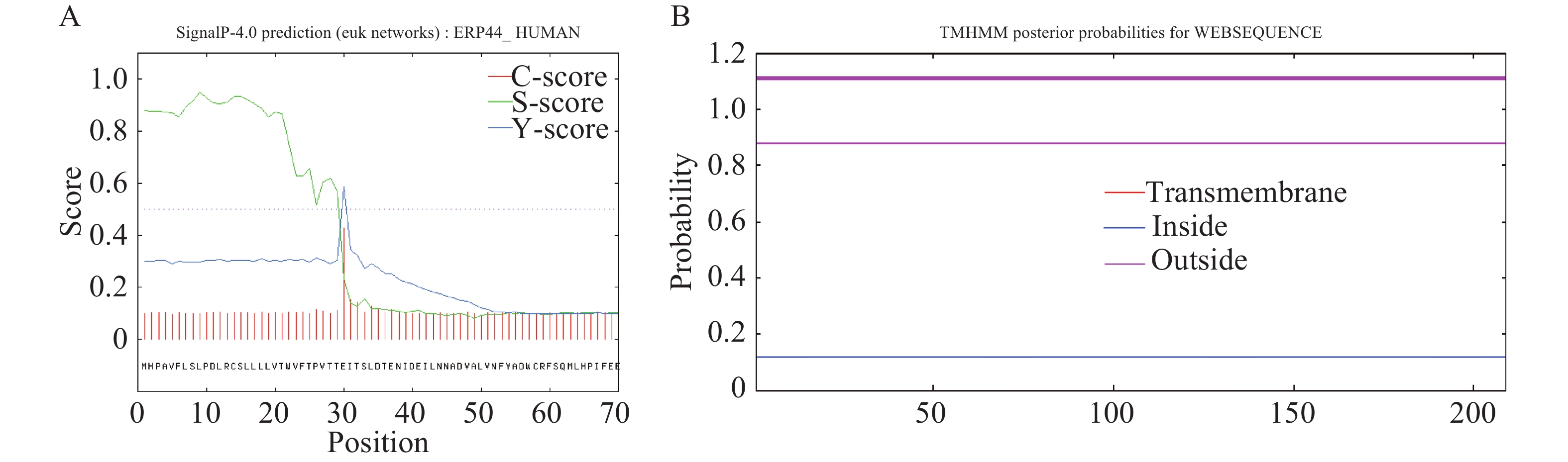
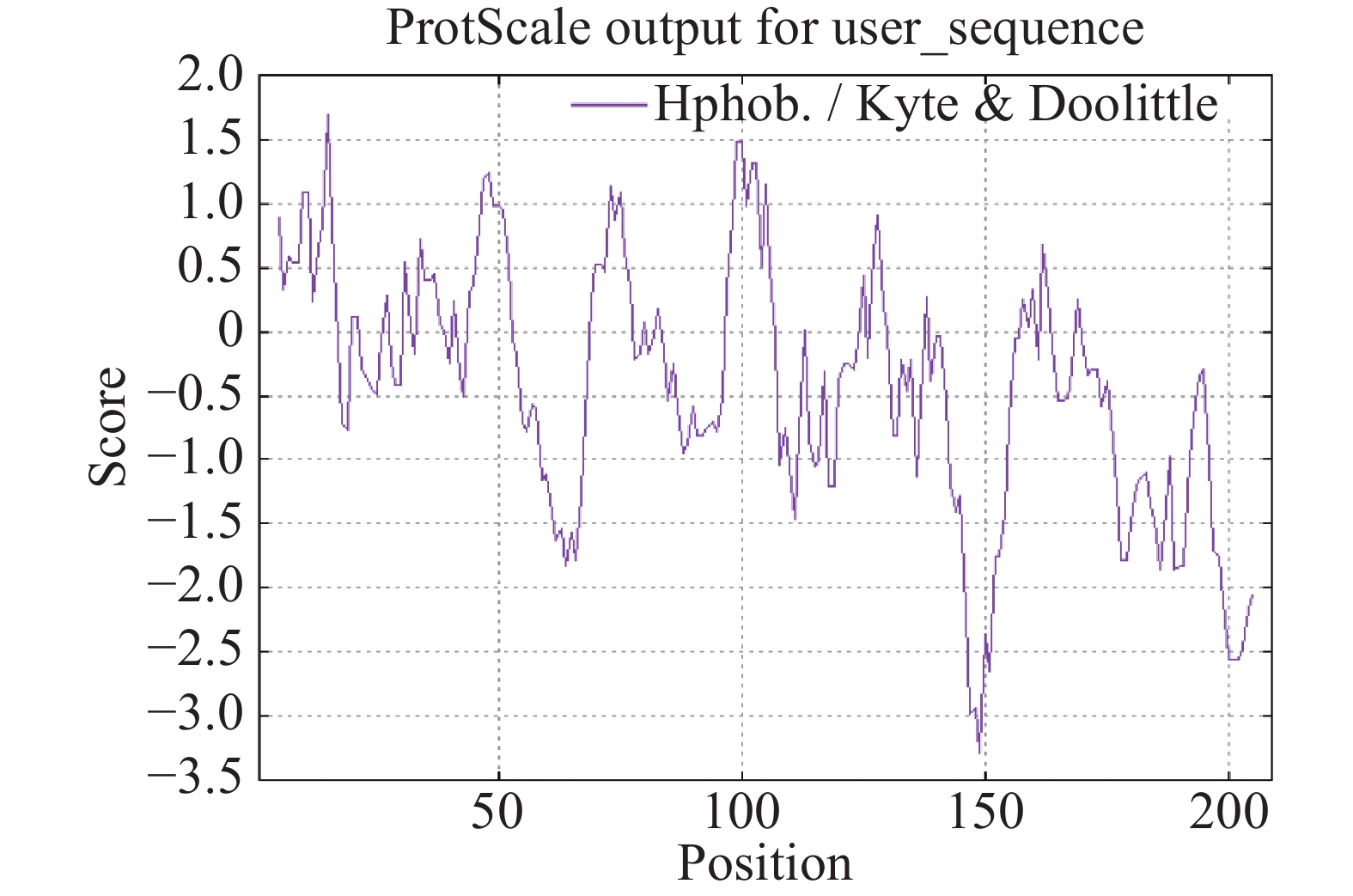
采用SignalP 4.1程序进行氨基酸序列的信号肽预测,结果显示,OnRR10基因在第30位氨基酸残基具有最高的原始剪切位点分值0.427,第9位氨基酸具有最高的信号肽分值0.950,由此可以看出,该基因编码的蛋白是分泌蛋白。根据TMHMM在线软件对OnRR10蛋白进行跨膜区段预测,结果表明,OnRR10基因编码的蛋白没有跨膜结构(图3),均属于非跨膜蛋白。利用ProtScale对氨基酸的疏水性/亲水性进行预测,结果表明,OnRR10基因编码的多肽链在第15号的天冬氨酸具有最高分值1.689,第149号的天冬酰胺具有最低分值−3.289;2条肽链均表现为亲水性,没有明显的疏水区(图4)。
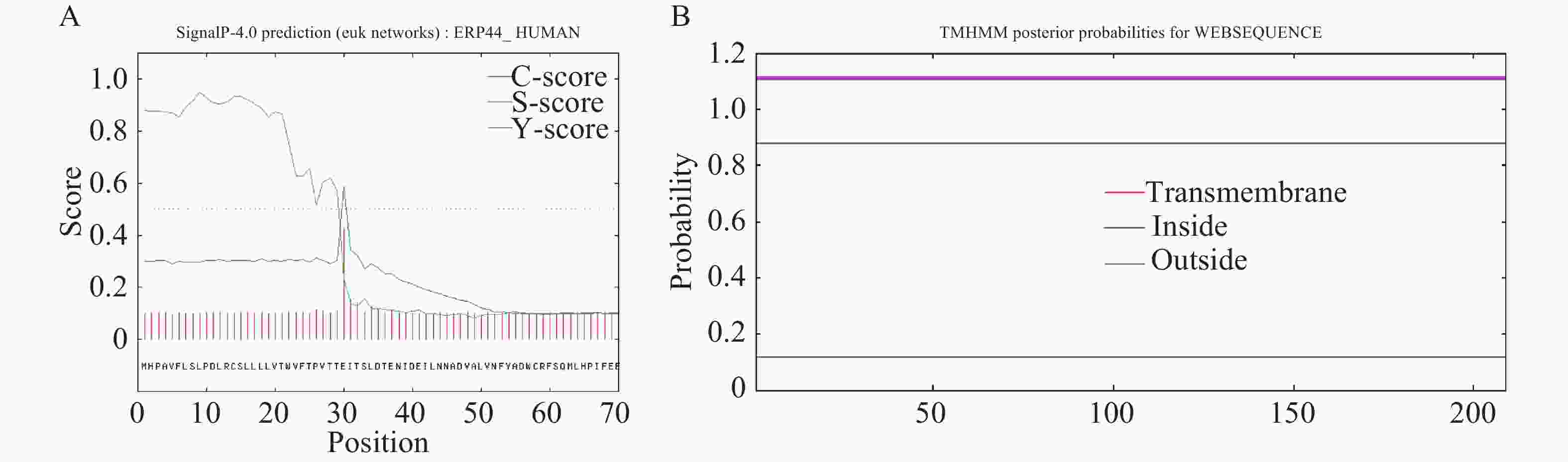
Figure 3. Prediction and analysis of signal peptides and prediction of transmembrane domains of the OnRR10 protein of Oncidium
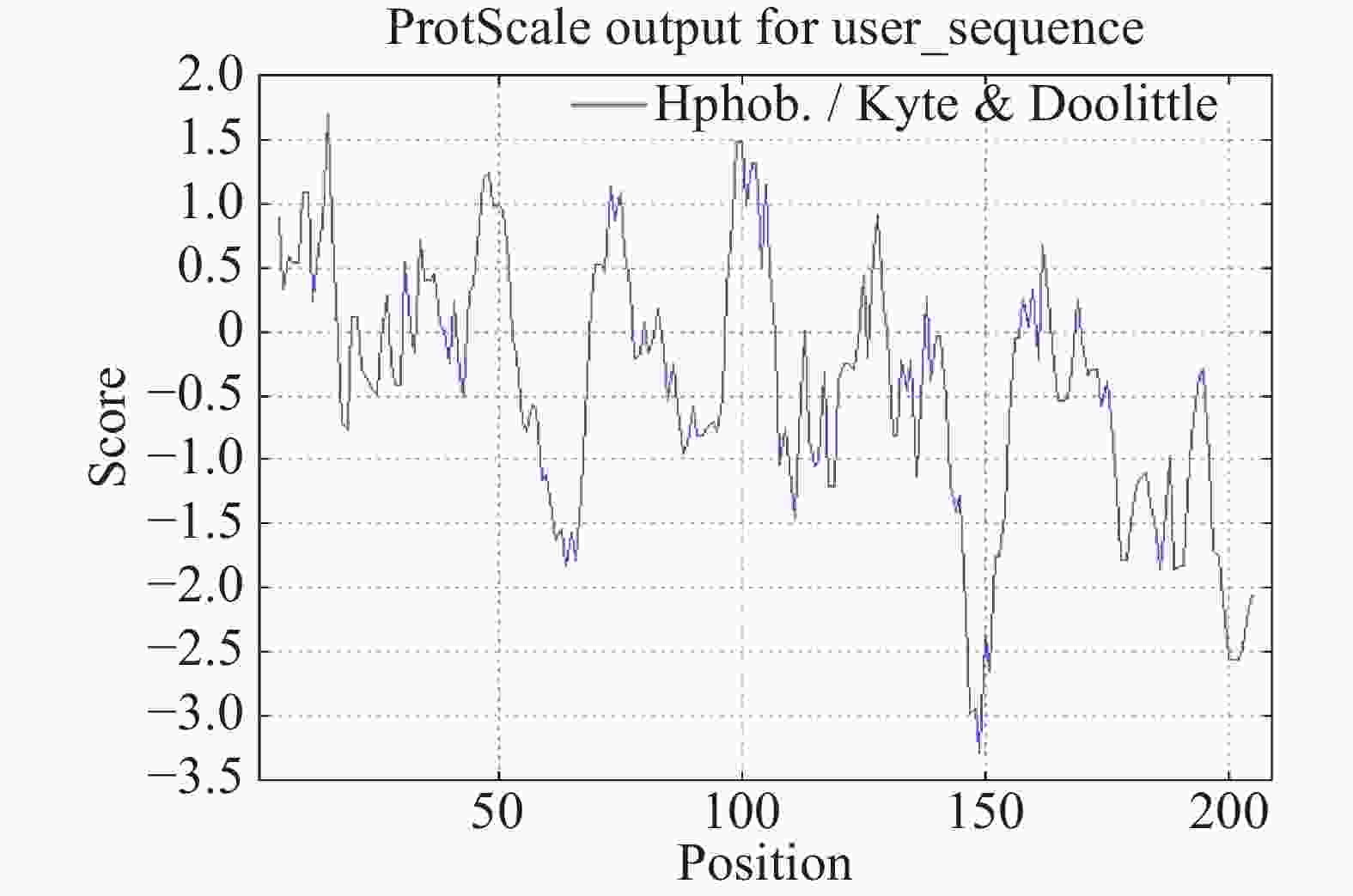
Figure 4. Prediction of hydrophilicity/hydrophobicity of Oncidium OnRR10 protein
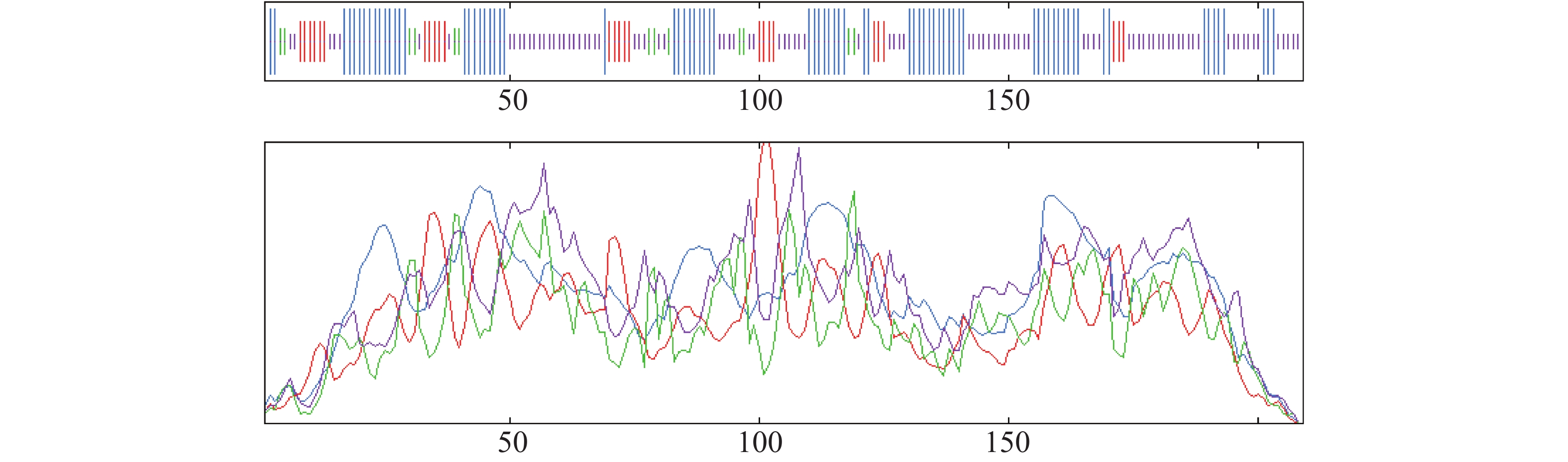
用SOPMA SECONDARY STRUCTURE PREDICTION METHOD对OnRR10蛋白进行2次结构分析;结果表明,该蛋白中无规卷曲占比最高为44.02%,α−螺旋、β−转角和延伸链占比分别为37.32%、6.22%和12.44%(图5)。
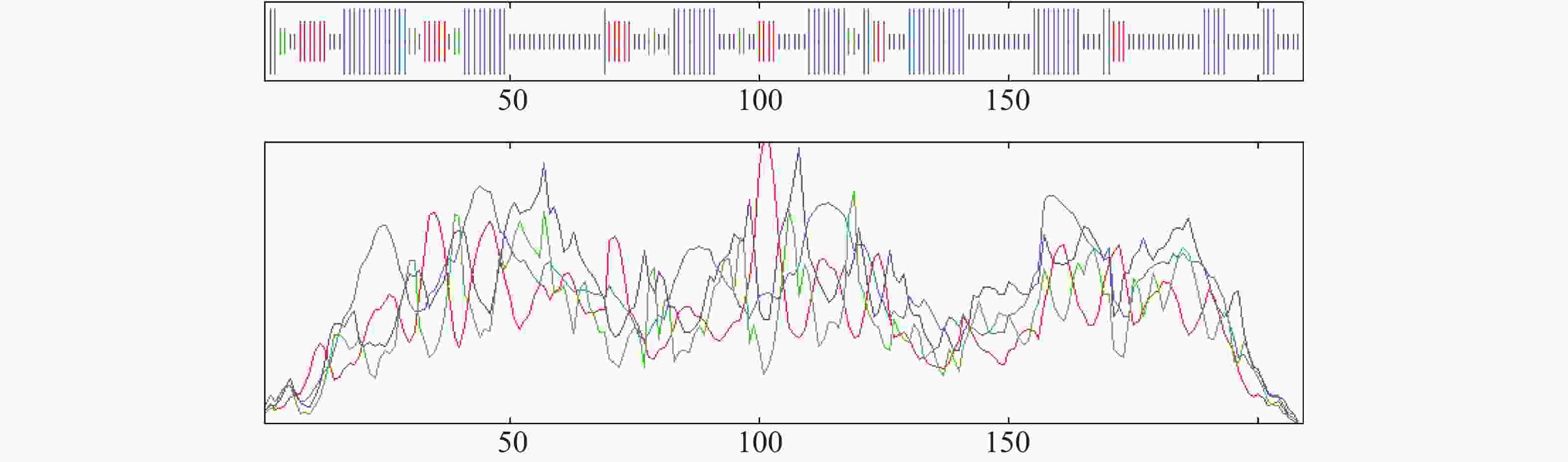
Figure 5. Prediction of secondary structure of the OnRR10 protein of Oncidium
用NCBI的BLAST进行序列比对,并用MEGA6基因及其同源序列构建进化树(图6)。结果显示,在与其他11个物种的比对中,OnRR10与石斛兰ORR9蛋白(Dendrobium catenatum, XP 020684569.1)及小兰屿蝴蝶兰ORR10蛋白(Phalaenopsis equestris, XP 020594130.1)亲缘关系最近。

Figure 6. Phylogenetic tree of the OnRR10 protein of Oncidium
-
分别采取文心兰花发育中7个时期(花苞期A、绽口期B、半开放期C、盛开前期D、盛开期E、衰老初期F、衰老期G)花瓣,提取总RNA,反转录得到cDNA进行Q-PCR反应,以A时期表达量为单位1,绘制不同花期OnRR10基因相对表达量柱状图。该基因在F时期表达量达到最高值,在D时期表达量最低(图7)。

Figure 7. Expressions of the OnRR10 gene of Oncidium in different flowering stages
-
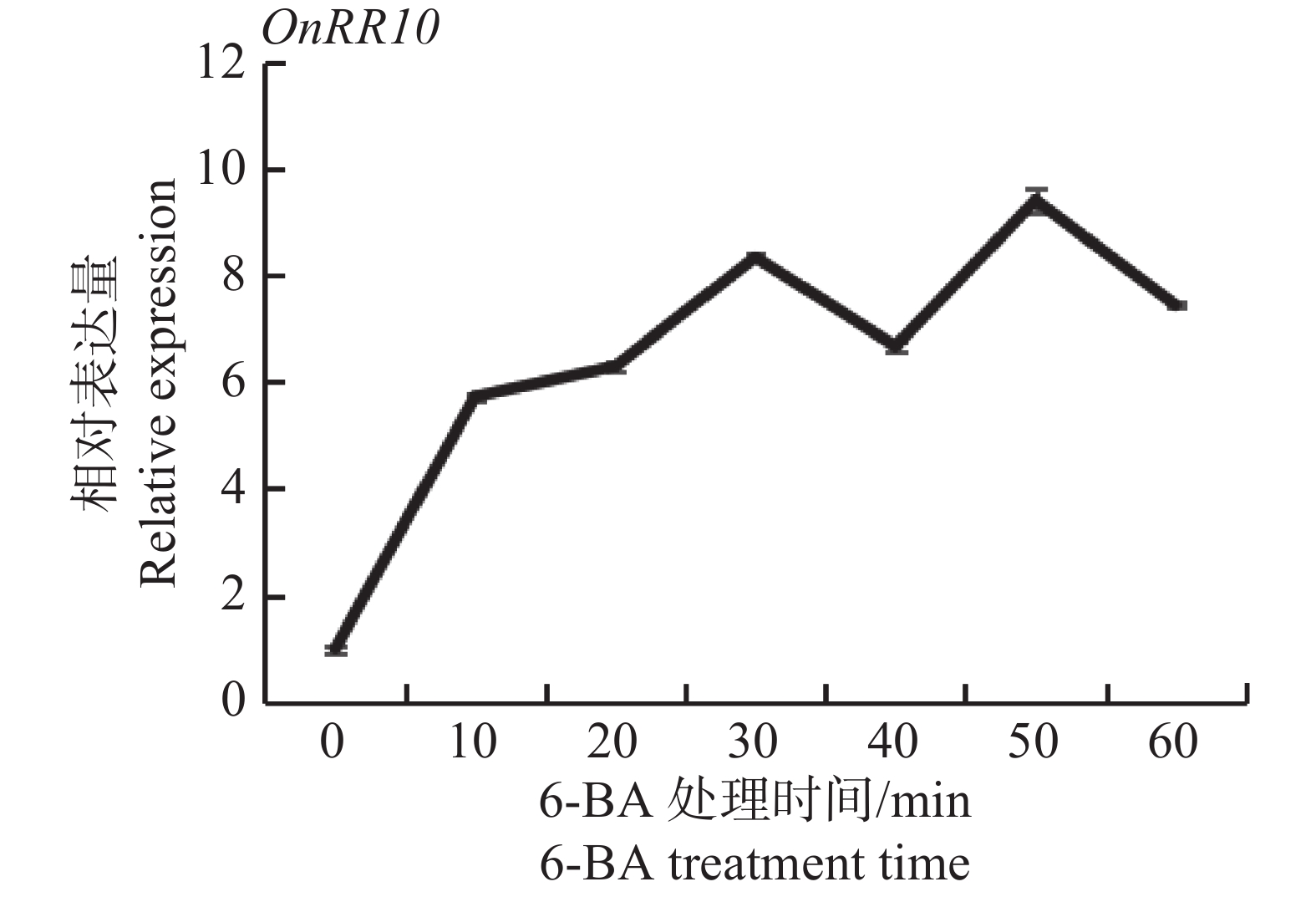
在60 mg·L−1 6-BA处理下,1 h内,OnRR10表达量整体呈上升趋势,在0~10 min内,OnRR10基因表达量急剧上升(图8)。这与D'AGOSTINO等[18]的研究结果基本一致(大部分A类ARRs受CTK转录诱导,外源CTK处理拟南芥,ARR基因在10~15 min内表达量急剧上升),表明该基因属于A型RR基因。
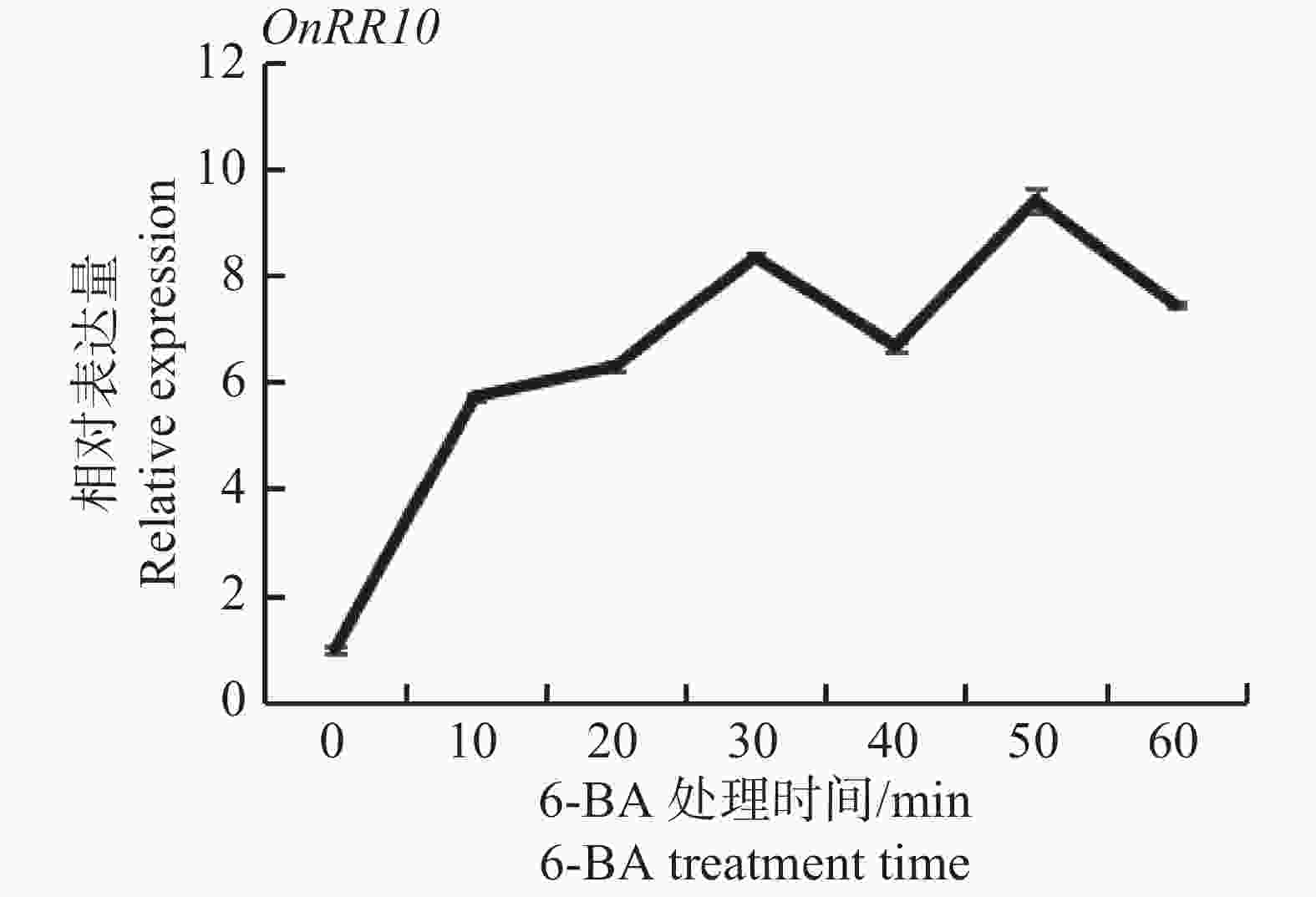
Figure 8. Changes in expressions of the OnRR10 gene under 6-BA treatments
-
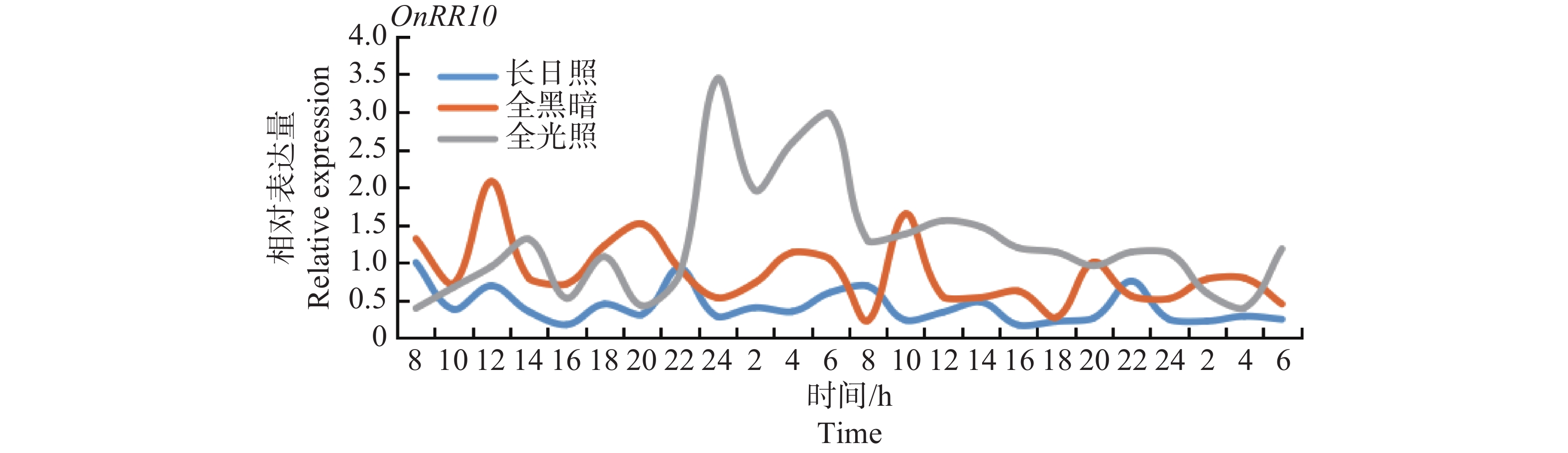
在全光照、长日照、全黑暗处理下,观察48 h内基因的表达量变化。结果显示:OnRR10基因在全光照下22~24 h及2~8 h出现表达量峰值;在长日照下20~22 h出现该基因表达量峰值;在全黑暗下10~12 h出现表达量峰值,该结果表明,该基因表达与生物钟无关,但同时也观察到,OnRR10基因在全光照下整体表达量最高,在长日照下整体表达量最低,说明该基因表达量受光照影响,全光照处理下,16 h后OnRR10基因表达量急速上升,持续约8 h,之后,表达量下降至平稳水平,但仍高于长日照下的表达量(图9)。

Figure 9. Changes in the expressions of the OnRR10 gene under different light treatments
-
本实验通过在NCBI上查找兰科RR基因序列,比对分析获得保守区域,设计兼并引物,并结合RACE技术获得OnRR10基因序列。Blast比对显示,OnRR10蛋白与蝴蝶兰、石斛兰等兰科植物ORR同源性在66%~83%之间,与其他物种同源性在60%~65%之间。同时,蛋白结构域分析表明,该基因属于YesN家族,与其他植物RR蛋白结构域分析结果一致[10-12],因此,克隆到的基因为RR基因。OnRR10基因在A时期(花苞期)和F(衰老早期)表达量最高,表明该基因可能在文心兰花开放及花瓣衰老早期发挥作用。在6-BA处理下,OnRR10在短期内急速上升,表达量达到峰值。由于拟南芥中A类型反应调节因子参与细胞分裂素信号转导系统,表达受外源细胞分裂素影响[18-20],结果表明,该基因属于A型RR基因,该基因表达并不受生物钟影响,但与光照有很大关系。相比于长日照、全黑暗条件,OnRR10基因全光照下表达量更高,但具体机制有待进一步研究。细胞分裂素是植物器官衰老的重要调控激素,细胞分裂素可延缓花瓣衰老[21-23]。有研究表明,参与双组分信号系统的蛋白参与到细胞分裂素信号转导,如拟南芥中ARR1,ARR6,ARR7,ARR10和ARR15均参与AHK调控的6-BA信号途径[11-12, 24-25]。因此,OnRR10基因很可能通过细胞分裂素信号转导途径在文心兰花瓣衰老启动中发挥调控作用。在后续的转化研究中将对其功能加以验证。
本实验通过RACE技术克隆获得OnRR10基因全长序列,对其编码蛋白质的氨基酸序列和蛋白质二级结构分析,对其亚细胞定位、疏水性、跨膜区段、功能进行预测,并对该基因在不同花发育阶段、6-BA处理、光照处理条件下进行表达分析,为进一步深入研究该基因功能打下了基础。
Cloning and Expression Analysis of OnRR10 Gene in Oncidium
doi: 10.15886/j.cnki.rdswxb.2020.03.005
- Received Date: 2019-11-07
- Rev Recd Date: 2020-02-27
- Available Online: 2019-11-28
- Publish Date: 2020-09-24
-
Key words:
- Oncidium /
- OnRR10 gene /
- expression analysis /
- real-time qPCR
Abstract: In order to characterize the key genes which control the petal senescence, a gene encoding an response regulator (RR), OnRR10 gene (GenBank accession number: MN337795), was isolated from the petals of Oncidium with RACE (rapid amplification of cDNA ends). The gene is 630 bp in length and encodes a protein of 209 amino acids in size. Protein secondary structure analysis showed that the OnRR10 protein is a hydrophilic protein and is localized in the nucleus. Homologous sequence alignment and phylogenetic analysis showed that the OnRR10 belongs to the YesN family. In Orchidaceae, OnRR10 protein is closely related to the ORR10 gene of Phalaenopsis equestris. Real-time quantitative analysis showed that the expression level of the OnRR10 gene was the highest at the bud swelling stage and flower initial senescence stage of Oncidium during the flower development, and that the expression of the OnRR10 gene was not regulated by the circadian clock, but increased rapidly in a short time under the 6-BA (6-Benzylaminopurine) treatment. These results implicated that the OnRR10 gene may be involved in the petal senescence through cytokine effect.
| Citation: | YAN Bingyu, GONG Xiaoxiao, TAN Yurong, WANG Dan, GAO Xuan, ZHANG Heng, LI Shuangjiang, WANG Peng, PAN Yingwen, LIU Jinping. Cloning and Expression Analysis of OnRR10 Gene in Oncidium[J]. Journal of Tropical Biology, 2020, 11(3): 288-295. doi: 10.15886/j.cnki.rdswxb.2020.03.005

|
 DownLoad:
DownLoad: