
-
溶珊瑚弧菌(Vibrio coralliilyticus)是一种革兰氏阴性、杆状的病原菌,它能引起印度洋和太平洋中普遍存在的鹿角杯形珊瑚(Pocillopora damicornis)的白化和组织裂解[1],研究显示,溶珊瑚弧菌也是柳珊瑚科的Pseudopterogorgia americana,鹿角珊瑚科的Acropora cytherea等白化的病原[2]。溶珊瑚弧菌还能感染多种海洋生物,如诱发东方牡蛎(Crassostrea virginica)和太平洋牡蛎(Crassostrea gigas)幼体大量死亡[3],也可导致虹鳟鱼(Oncorhynchus mykiss)患病[4]。群体感应(Quorum Sensing)是细菌通过感应种群密度的波动来调节自身基因表达的一种机制。群体感应细菌产生并释放被称为自诱导分子(Autoinducer)的化学物质来传递信号,其浓度随着细胞密度的增加而增加。通过感应这种信号分子的浓度,细菌可以调节多种生理活动,包括共生、竞争、毒力因子表达、接合、抗生素的合成、细菌运动、孢子形成以及生物膜形成[5]。弧菌一直以来都是研究群体感应系统的重要模型,因为在弧菌中,群体感应被证实可以参与调控多种基因的表达[6]。HapR家族蛋白作为响应种群密度变化的泛调控因子(global regulator),控制着上百个基因的表达,其中包括细胞的定殖、生物膜的形成、分泌系统和c-di-GMP信号传导、细胞毒素等基因[7]。HapR蛋白最早发现于哈维氏弧菌[8],后来相继在霍乱弧菌、副溶血弧菌、河流弧菌和溶藻弧菌中被发现[8-13],但尚未见HapR在溶珊瑚弧菌中的报道。在前期实验中,笔者发现,溶珊瑚弧菌的接种浓度必须达到107 cfu·mL−1时才能引起珊瑚的急性感染,推测群体感应系统可能是溶珊瑚弧菌毒力的调控因子。本研究中,笔者构建了溶珊瑚弧菌hapR基因缺失株△hapR-Vc450和回补株ChapR-Vc450,对溶珊瑚弧菌野生株Vc450,hapR缺失突变株和回补株的菌落形态、鞭毛结构、细菌运动性、生长代谢情况及生物膜的形成量进行了实验分析,同时比较了它们感染鹿角珊瑚的毒力,旨在探讨hapR在溶珊瑚弧菌中可能的生物学功能及其与毒力相关的机制,为珊瑚病害防治提供理论指导。
HTML
-
溶珊瑚弧菌野生株:Vc450;hapR缺失突变株:△hapR-Vc450;hapR回补株ChapR-Vc450;大肠杆菌:GEB802,GEB883(表1)。
菌株或质粒
Strain or plasmid基因型或表型
Genetype or phenotype参考或来源
Reference or source大肠杆菌 GEB802 LaclQ, thil, supE44, endA1, recA1, hsdR17, gyrA462,zei298::tn10[Tc], △thyA::(ermpir116), Thy-,intermediate host of suicide plasmid pSW7848 Annick Lab GEB883 WT E. coli K12 △dapA::erm pir RP4-2 △recAgyrA462, zei298::tn10,donor strain for conjugation Annick Lab 溶珊瑚弧菌 Vc450 Isolate from diseased coral Pocillopora damicornis in Mawi Island, Indian Ocean 美国菌种保藏中心ATCC △hapR-Vc450 hapR基因缺失株 本研究构建 ChapR-Vc450 Cmr; hapR基因完整开放阅读框回补株 本研究构建 Plasmid pSW7848 Cmr, T7, oriT RP4, araC, oriV R6K, catr Annick Lab pMMB207 Apr, Cmr, MobA, RecpA, RepC, lacI 赵哲实验室 Table 1. Bacterial strains and plasmids used for test
-
分子实验试剂:PCR反应试剂(高保真酶PrimeSTAR、常规rTaq DNA聚合酶及DNA marker)均购自TaKaRa公司;基因片段连接试剂购自诺唯赞生物科技有限公司;基因组提取试剂盒购自天根生物有限公司;PCR产物纯化试剂盒和DNA凝胶纯化试剂盒购自Axygen公司;质粒提取试剂盒购自Macherey nagel公司。细菌培养试剂:培养基LB购自Invitrogen公司;TCBS培养基购自广州环凯微生物科技有限公司;2216E培养基购自BD公司;三氟胸腺嘧啶核苷(Thy)、二氨基庚二酸(DAP)、氯霉素、IPTG诱导剂、D−(+)−葡萄糖(D-Glucose)、L−(+)阿拉伯糖(L-(+)-Arabinose)均购自sigma公司;氯霉素(Cm)使用浓度为5 μg·mL−1和20 μg·mL−1。表型分析用试剂及耗材:Agar琼脂粉、NaCl购自生工生物工程上海(股份)有限公司(简称:生工生物);5%苯酚溶液、甲醇、冰醋酸等常规试剂均为国产分析纯;24孔板购自美国Costar公司;染色制片用防脱载玻片及常规盖玻片均购自生工生物。常用培养基:LBS培养基为添加2% NaCl的LB培养基。BFM (Biofilm Formation Medium)培养基:KH2PO4 3 g·L−1,K2HPO4 7 g·L−1,(NH4)2SO4 2 g·L−1,FeSO4 0.5 mg·L−1,MgSO4 2 mmol·L−1,NaCl 10 g·L−1,Thiamine 5 mg·L−1,Casamino acids 15 g·L−1,Glucose 4 g·L−1。FASW(Filtered Artificial Sea Water)是用珊瑚盐配制的人工海水,盐度32,用0.2 μm滤膜过滤灭菌。
-
溶珊瑚弧菌Vc450敲除突变株构建过程:采用在线引物设计软件NEBuilder(
http://nebuilder.Neb.com/ )设计基因hapR上下游片段和质粒pSW7848线性化片段扩增引物,引物见表2。将上述纯化好的上下游同源臂片段和pSW7848线性载体进行重组片段的连接,重组质粒转化至大肠杆菌感受态细胞GEB883中,并与溶珊瑚弧菌野生株Vc450进行接合。在添加0.2%葡萄糖的TCBS平板和添加0.2%阿拉伯糖的TCBS平板上进行缺失突变菌株筛选,将可疑阳性克隆进行PCR和测序检验,最后挑选若干个单克隆进行PCR检测,筛选出成功突变的菌株△hapR-Vc450。溶珊瑚弧菌Vc450敲除回补株构建过程:根据溶珊瑚弧菌Vc450基因组中的hapR基因序列,设计线性化回补载体pMMB207引物以及hapR基因回补序列扩增引物,引物见表2。将已纯化的hapR基因片段与pMMB207线性载体进行重组连接,将重组质粒转化至大肠杆菌感受态细胞GEB883中,并与溶珊瑚弧菌hapR缺失株△hapR-Vc450进行接合。在添加0.2%葡萄糖的TCBS平板进行回补株筛选,将可疑阳性克隆进行PCR和测序检验,最后挑选若干个单克隆进行PCR检测,筛选出成功突变的菌株ChapR-Vc450。引物名称
Primer引物序列 Primer sequence 上游引物(5′→ 3′) Sense primer 下游引物(5′→ 3′) Antisense primer pSW7848 GTCTGATTCGTTACCAATTATGACAAC GAATTCGATATCAAGCTTATCGATAC Del-check-pSW7848 TCACTGTCCCTTATTCGCACC CTGCTTTTGAGCACTACCCG hapR-UP aagcttgatatcgaattcTACGCTGCTCAAGTAGAG catatctaggggtctcttagGGTCTCTTAGCTATAGAATCC hapR-DOWN ctaagagacccctagatatgCCTAGATATGCTTTGCATC taattggtaacgaatcagacACAGTTTACCAGTCACTAG Del-check-hapR CAGCAGGAGAAACGTTTGCT TATCCACGCTCACCCAACT pMMB207 AGAAGCGGTCTGATAAAACAGAATTTGC GCGCAACGCAATTAATGTAAGTTAG Vc_hapR attaattgcgttgcgcGTCATGAAATCAACTTGGTGAG tttatcagaccgcttctACGGTGGCGAACTACTAG com-PMMB207-check CTACTGAGCGCTGCCGCACA TCGTTTTATTTGATGCCTGGCAG 注:下划线加粗序列为上下游同源臂与质粒片段连接时重叠互补序列;斜体加粗序列为上下游同源臂之间连接时重叠互补序列。
Note: The sequences, underlined and bold, were overlapping fragments between up/down-segments and plasmid, and the sequences in italics and bold were overlapping fragments between up-segment and down-segment of target genes.Table 2. Nucleotide sequences of primers used for construction of deletion mutants
-
挑取溶珊瑚弧菌野生株Vc450和突变株△hapR-Vc450单菌落至LBS液体培养基中,将回补株ChapR-Vc450挑取至LBS+5 mg·L−1 Cm液体培养基中,在30 ℃ 200 r·min−1摇床中振荡过夜。连续稀释过夜培养的菌液至适当的浓度,取100 μL涂布于TCBS平板上,30 ℃静置培养16 h,在BIO-RAD图像系统Gel DocTM XR+下观察单个菌落形态。通过透射显微镜(Hitachi H-7650 TEM)观察细菌细胞形态及鞭毛结构。
-
将溶珊瑚弧菌Vc450和突变株△hapR-Vc450单菌落挑取至2216E液体培养基中,将回补株ChapR-Vc450挑取至2216E+5 mg·L−1 Cm液体培养基中,在30 ℃ 200 r·min−1摇床中振荡过夜。为了测试细菌在富营养条件下的生长情况,将培养过夜的菌液以1∶100的比例(体积比)接种至2216E液体培养基中,30 ℃ 200 r·min−1振荡培养,用分光光度计测定OD600。
-
Vc450和突变株△hapR-Vc450单菌落挑取至LBS液体培养基中,将ChapR-Vc450挑取至LBS+5 mg·L−1 Cm液体培养基中,以30 ℃ 200 r·min−1振荡过夜。调整培养液浓度至OD600=1.00,取5 μL平行3次点样于0.3%和1.5%琼脂的LBS半固体培养基上,30 ℃静置培养16 h,观察细菌的运动情况。
-
将溶珊瑚弧菌Vc450和突变株△hapR-Vc450单菌落挑取至LBS液体培养基中,将ChapR-Vc450挑取至LBS+5 mg·L−1 Cm液体培养基中,以30 ℃ 200 r·min−1振荡过夜。添加1 mL BFM培养基于24孔板中。调整菌液浓度至OD600=1.00后,取10 μL Vc450,△hapR-Vc450接种于含1 mL BFM培养基中,10 μL ChapR-Vc450接种于含5 mg·L−1 Cm的BFM培养基中,30 ℃静置培养,取样测定生物膜生成量OD562,生物膜生成量测定方法参照文献[14]。
-
将溶珊瑚弧菌野生株Vc450,突变株△hapR-Vc450单菌落接种于LBS液体培养基中,将回补株ChapR-Vc450挑取单菌落至LBS+5 mg·L−1cm液体培养基中,30 ℃ 200 r·min−1振荡培养过夜。再以1︰100的体积比接种至200 mL 2216E液体培养基中,30 ℃ 200 r·min−1振荡培养12 h。感染浓度为106 cfu·mL−1。感染方法:准备5个消毒后的10 L玻璃缸,添加9.5 L ASW并添置加热棒以维持水温在24 ℃,每个水缸配备1根温度计;每个水缸中分别放置6枝4~5 cm的珊瑚枝,以12 h∶12 h光照/黑暗暂养2 d;将培养12 h的菌液以3 800 r·min−1离心15 min,去上清;用10 mL FASW重悬,3 800 r·min−1离心15 min,去上清;用3 mL FASW过滤海水重悬添加至9.5 L ASW的玻璃缸中,对照组添加3 mL过滤海水;浸泡感染8 h后,换新鲜海水;观察并记录珊瑚状态。
1.1. 菌株及质粒
1.2. 主要试剂及培养基
1.3. 溶珊瑚弧菌Vc450 hapR基因敲除突变株及回补株的构建
1.4. 菌落和细胞形态观察
1.5. 细菌生长曲线测定
1.6. 细菌运动性分析将溶珊瑚弧菌
1.7. 生物膜形成能力检测
1.8. 鹿角珊瑚的感染
-
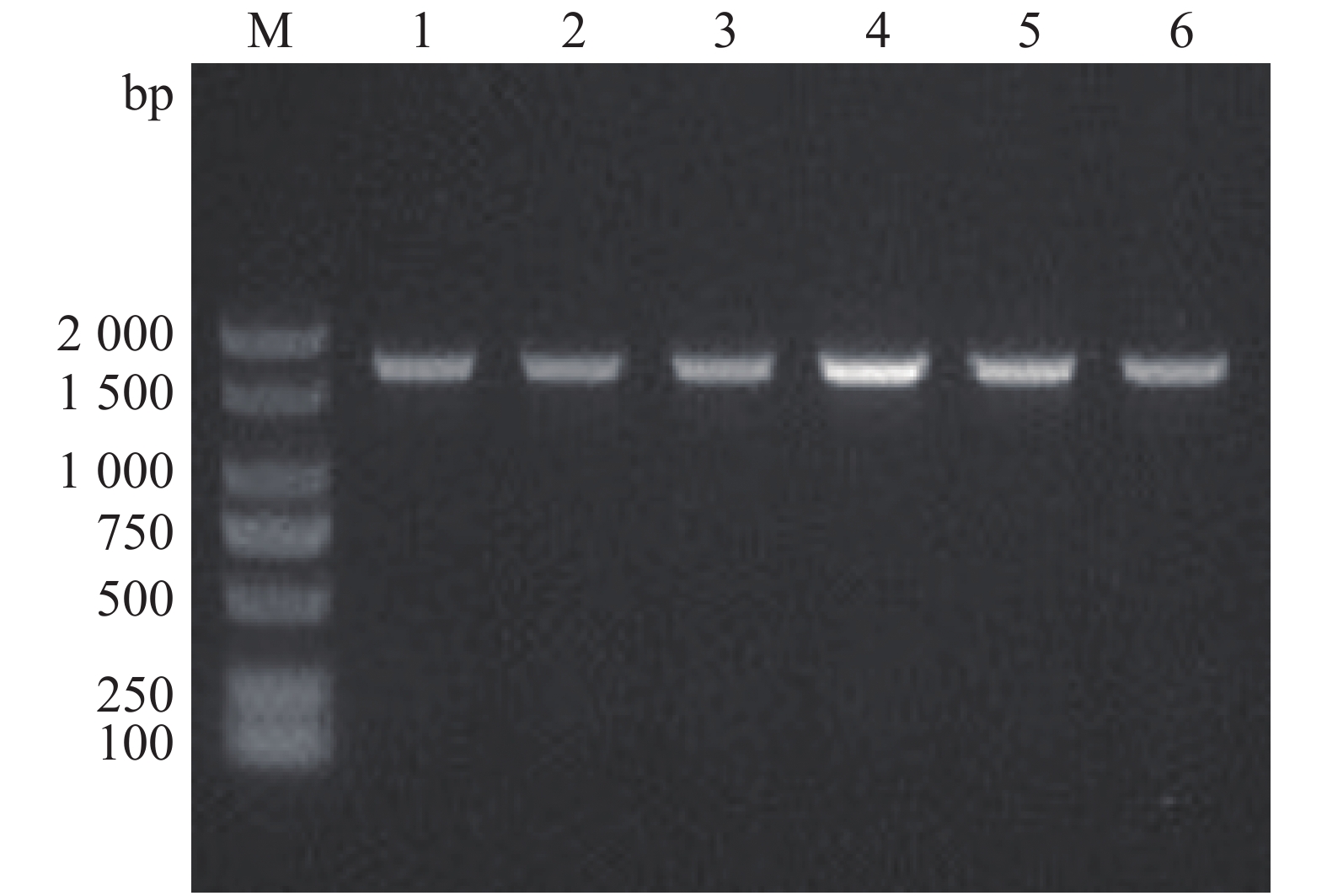
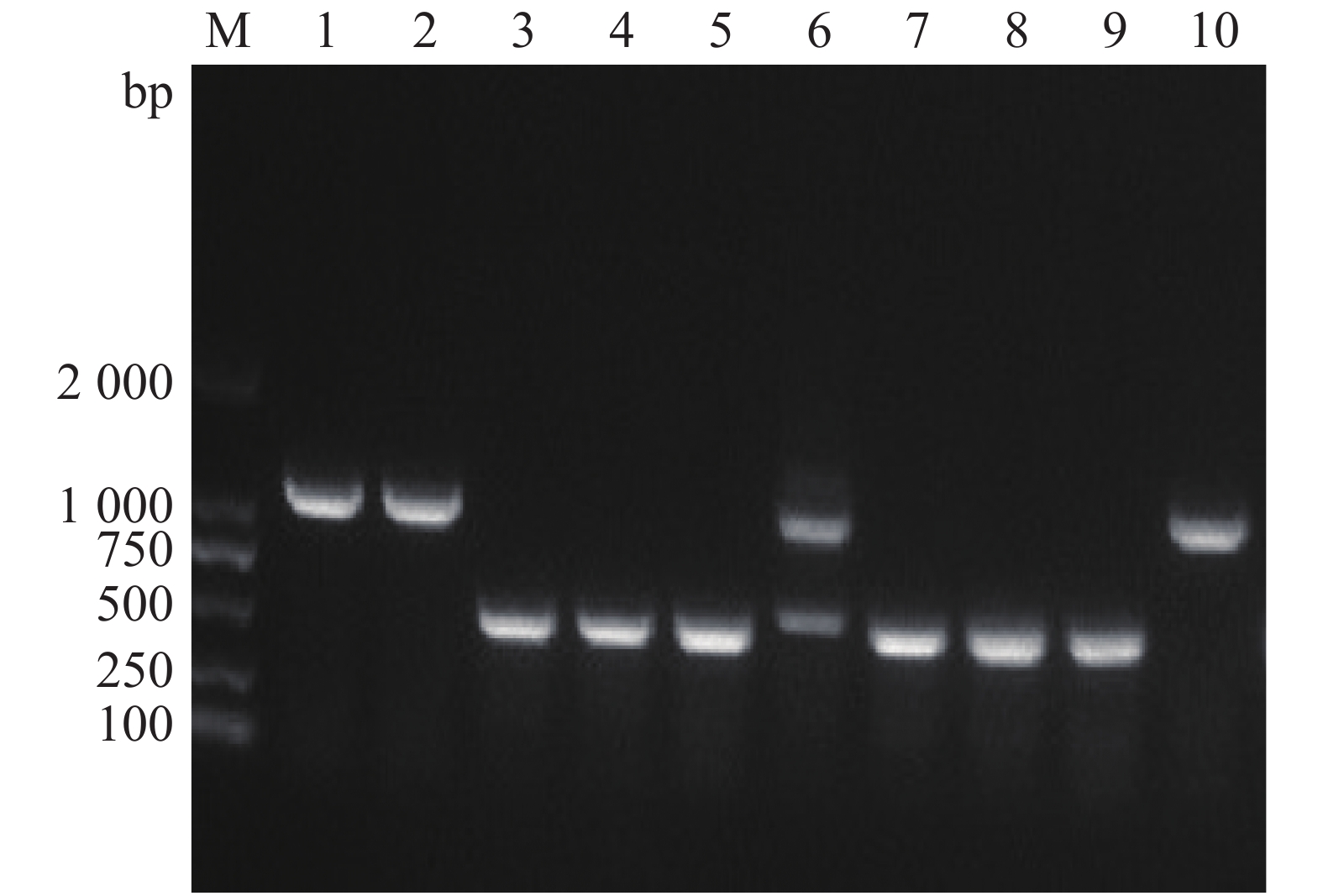
通过特异性引物设计、重组载体构建以及细菌接合和筛选,本实验成功构建了溶珊瑚弧菌Vc450 hapR基因敲除突变株ΔhapR-Vc450和回补株ChapR-Vc450。在hapR的两侧设计引物Del-check-hapR-F/R,突变株PCR检测电泳结果如图1。成功缺失掉hapR基因的突变株检测片段的大小为492 bp,而野生株检测片段大小为1 047 bp((hapR序列长度为555 bp),将条带大小吻合的可疑突变株送测,测序结果表明,hapR基因已被成功缺失。用检测引物com-PMMB207-check-F/R对回补株进行PCR检测验证,扩增得到约1 711 bp的片段(图2),测序结果表明,hapR回补株已被成功构建。

Figure 1. Agarose gel electrophoresis of the PCR products of the deletion mutant strain ΔhapR-Vc450
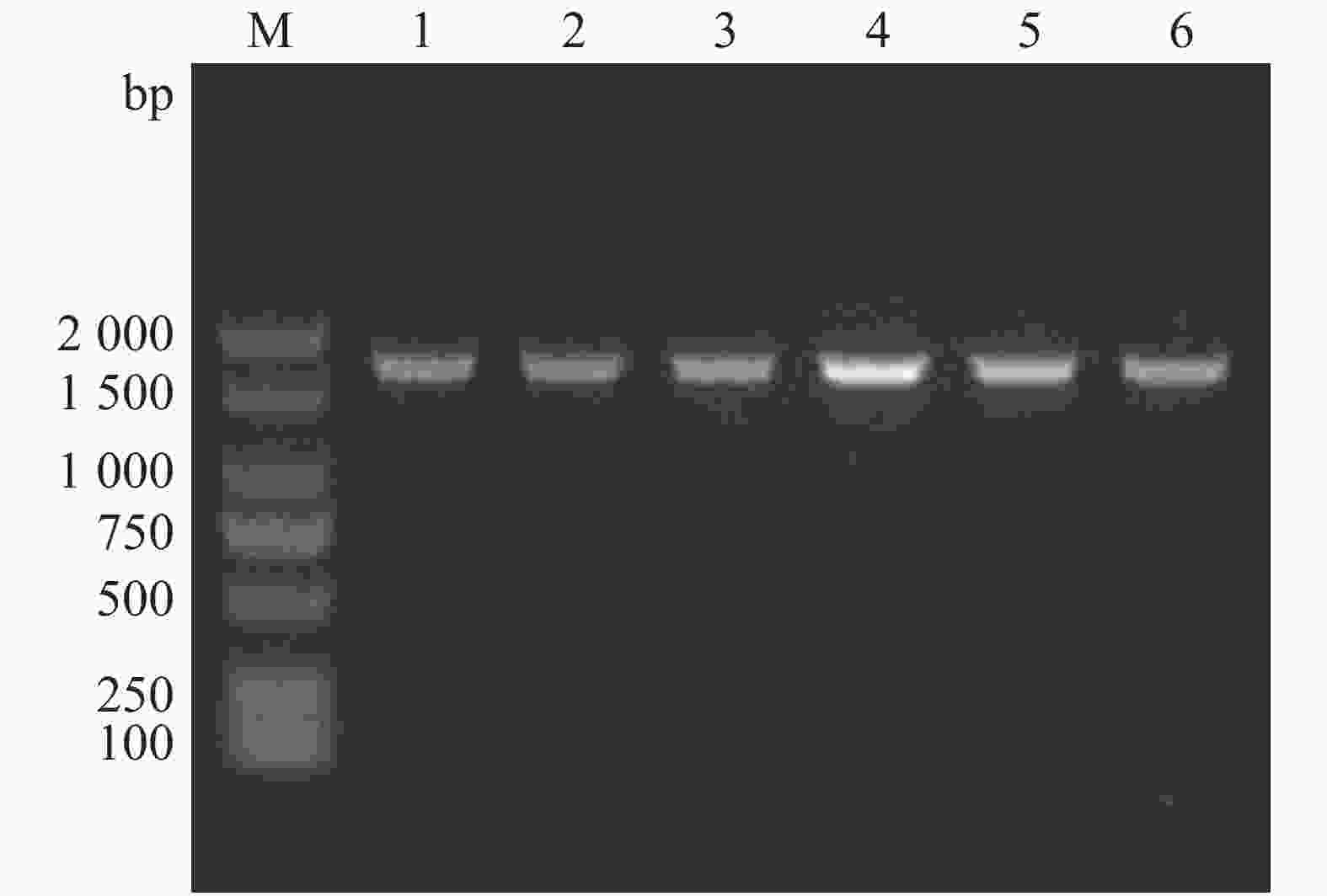
Figure 2. Agarose gel electrophoresis of the PCR products of the complementary strain ChapR-Vc450
-
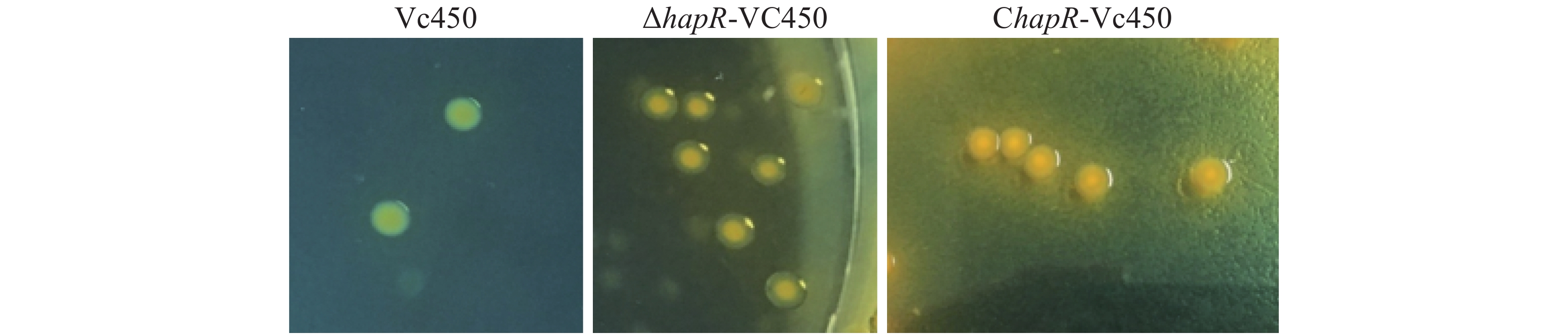
溶珊瑚弧菌野生株Vc450,突变株△hapR-Vc450和回补株ChapR-Vc450在TCBS平板上的菌落形态如图3。野生株Vc450在TCBS平板上的菌落表现不透明/光滑,hapR缺失株的菌落则表现为透明/光滑,当hapR基因回补后,菌株表型得到回补。

Figure 3. The colonial morphology of Vc450, mutant strain △hapR-Vc450 and complementary strain ChapR-Vc450 of Vibrio coralliilyticus
透射电镜结果如图4,图片显示,突变株△hapR-Vc450和回补株ChapR-Vc450具有与野生株一样的细胞形态和鞭毛结构,说明hapR基因对溶珊瑚弧菌的细胞形态和鞭毛结构没有影响。
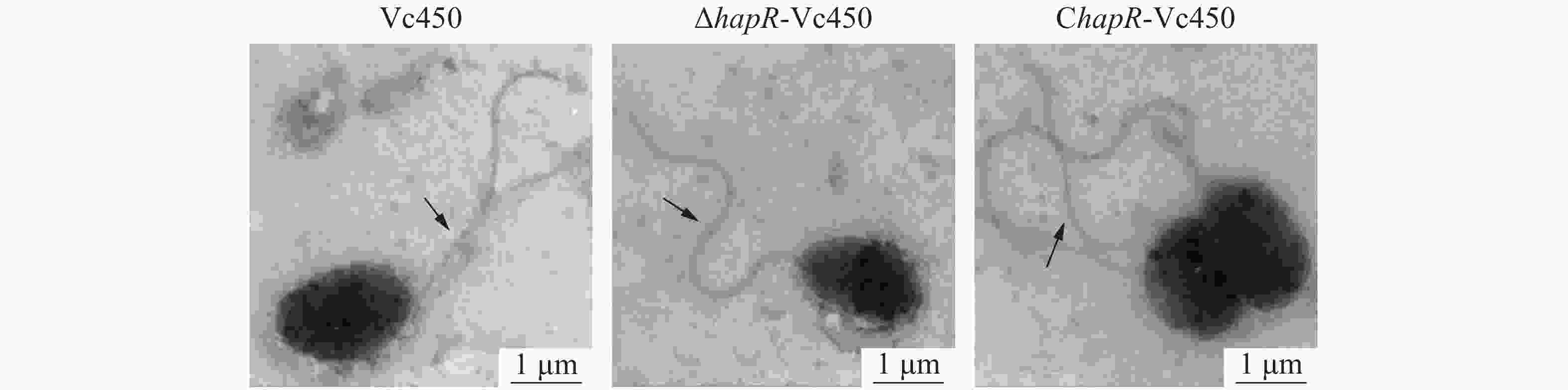
Figure 4. Observation of cellular morphology and flagella under Hitachi H-7650 TEM in Vc450, △hapR-Vc450 and ChapR-Vc450 of Vibrio coralliilyticus
-
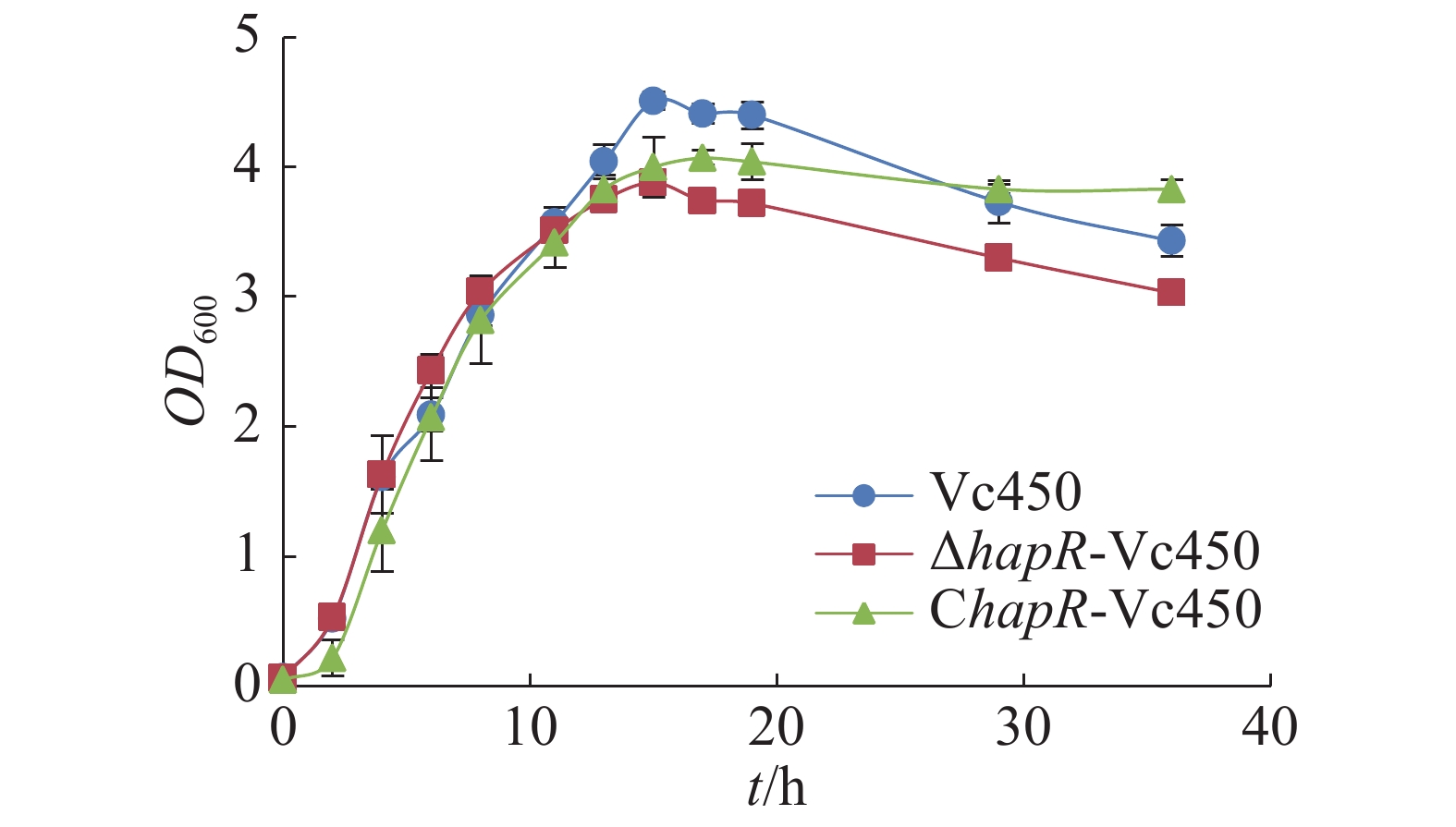
从图5可知,hapR敲除突变株与野生株在潜伏期及对数期并无显著差异,但在平台期,突变株的最大生长量明显减少;回补株在早期潜伏期及对数期的生长稍慢于突变株与野生株,到达平台期的最大生长量相较于突变株得到部分回补,说明hapR基因的缺失会影响溶珊瑚弧菌的最大生长量。
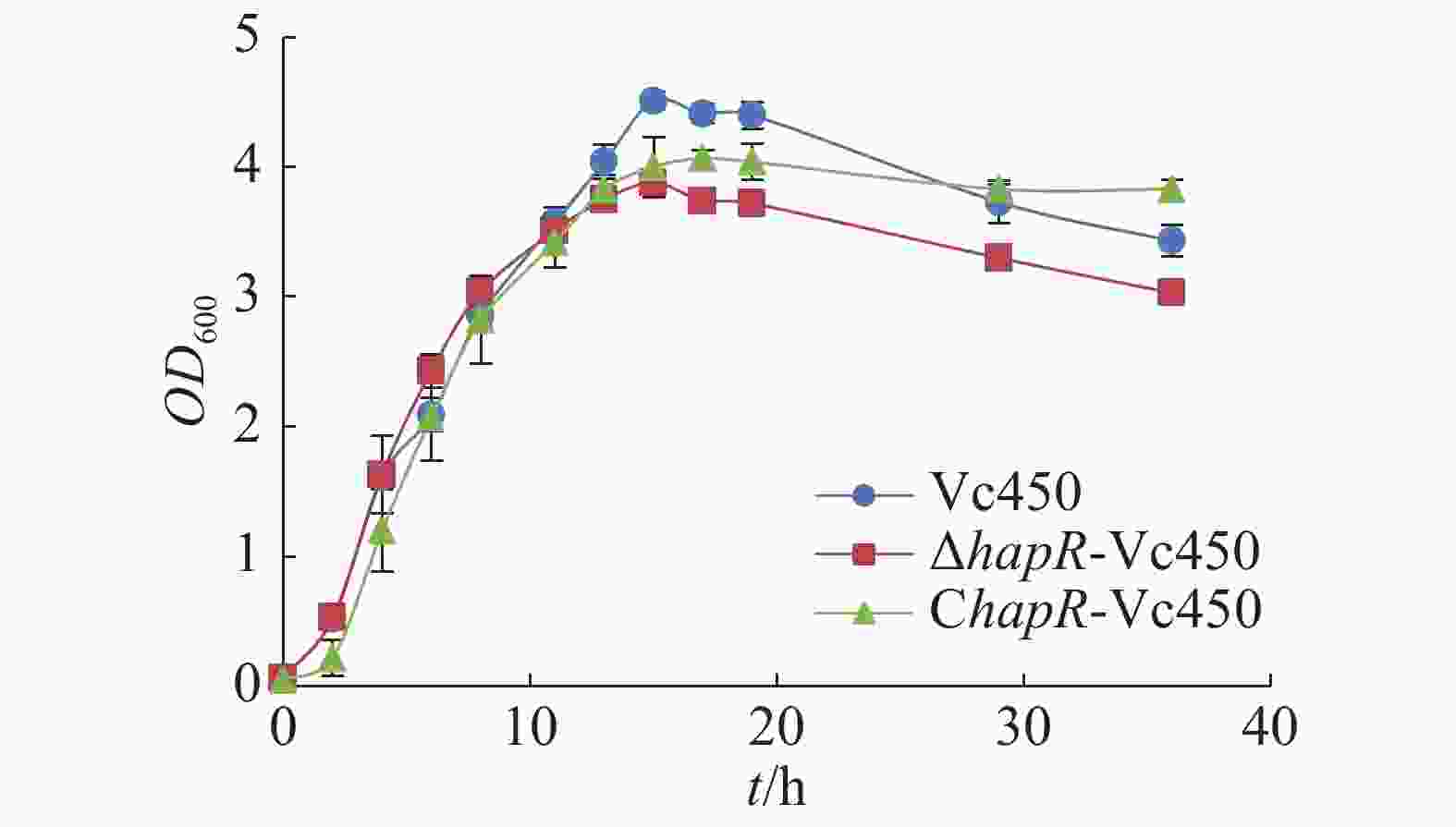
Figure 5. Effect of hapR deletion in Vc450 on growth of Vibrio coralliilyticus in 2216E medium
-
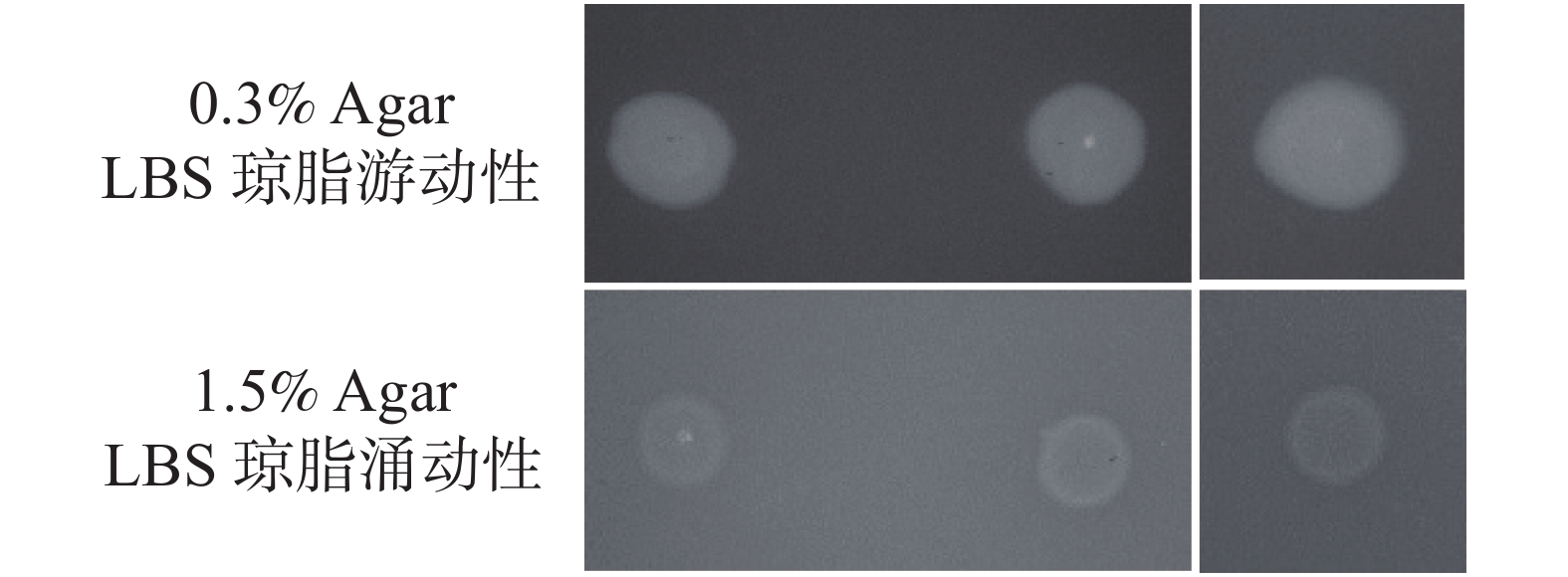
从图6可知,突变株与回补株在2种平板上的运动性,相较于野生株均无明显差异;而相比起1.5% Agar LBS涌动性,3株菌在0.3% Agar LBS表现出较强的游动性,但彼此间无明显差异,说明hapR基因不影响溶珊瑚弧菌的运动性。

Figure 6. The mobility of the Vc450 wild type, mutant and complementary strain of V. coralliilyticus on 0.3% Agar and 1.5%Agar LBS
-
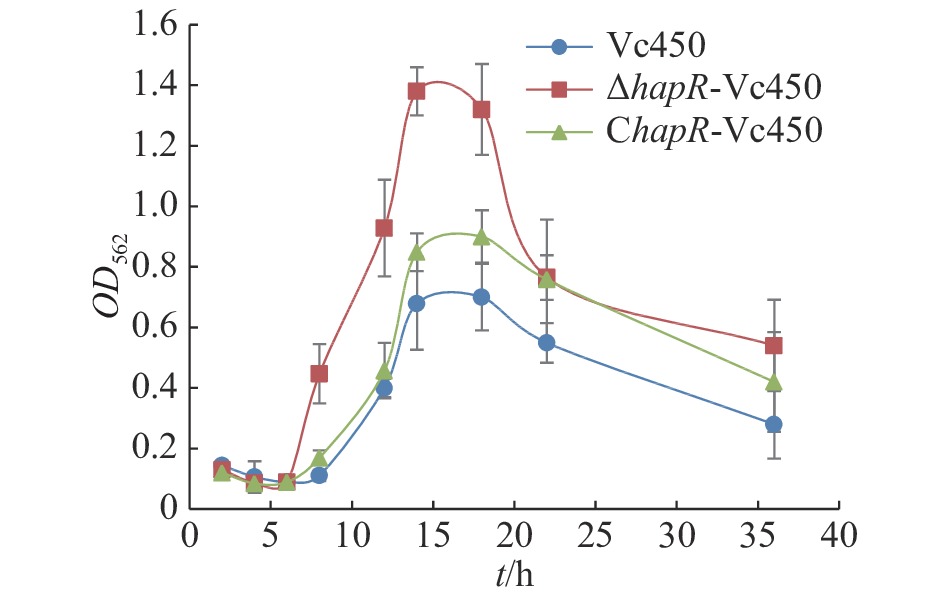
本研究采用结晶紫染色法定量测定溶珊瑚弧菌野生株Vc450,突变株△hapR-Vc450和回补株ChapR-Vc450在30 ℃静置培养条件下生物膜的形成量(图7)。从图7可知,在生物膜形成的起始阶段(0~6 h),3株菌生物膜的形成量无显著差异;在8 h后,△hapR-Vc450的生物膜形成速度显著高于野生株和回补株,并在14 h后形成量达到最大值;14 h后,回补株生物膜的形成量亦高于野生株,并在12~14 h时生物膜形成量达到最大值;野生株在12~14 h达到最大值。结果表明,hapR基因的缺失会显著影响溶珊瑚弧菌的生物膜形成能力,且主要参与了生物膜形成过程的起始阶段。
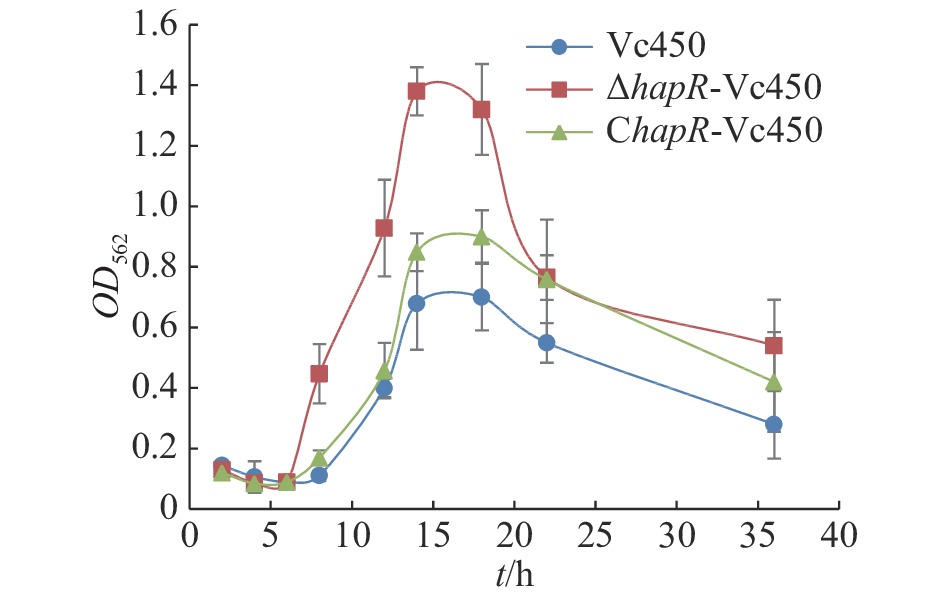
Figure 7. Detection of biofilm formation of the wild-type strain Vc450, mutant strain △hapR-Vc450 and complementary strain ChapR-Vc450 of Vibrio coralliilyticus
-
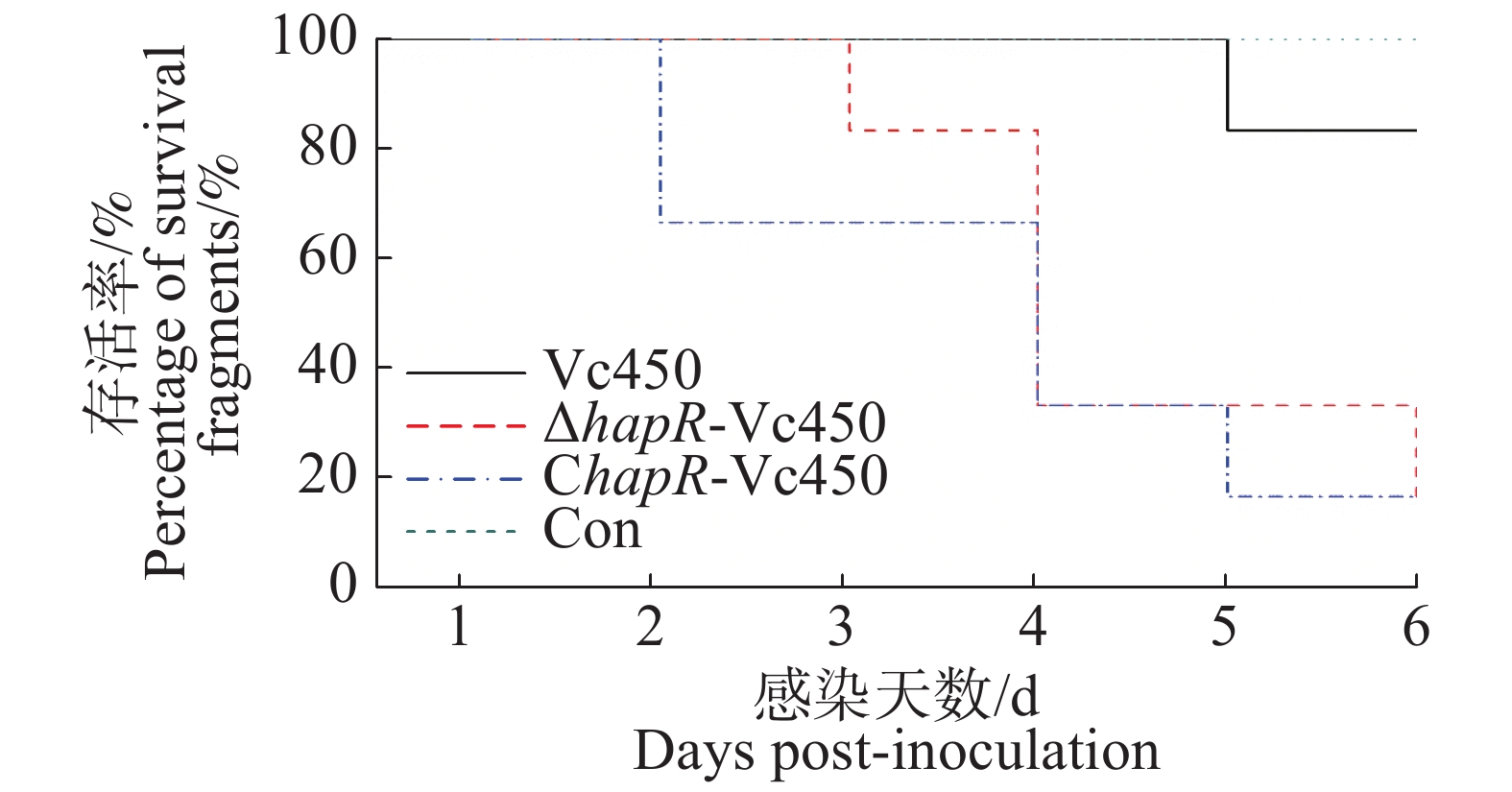
本研究用溶珊瑚弧菌对鹿角珊瑚进行感染。珊瑚的白化情况如图8及表3。实验结果表明,突变株△hapR-Vc450和回补株ChapR-Vc450的感染能力相对于野生株Vc450的感染能力较强。回补株感染后在第2天开始出现白化,在第5天,珊瑚白化和死亡的比例达到了83%;突变株感染则在第3天才出现白化,到第5天时,83%的珊瑚白化死亡;而野生株感染后,在感染5天后,只有17%的珊瑚白化死亡;空白对照组在观察期内未出现白化情况。这说明hapR基因突变之后,溶珊瑚弧菌毒力增强。
处理
Treatment感染浓度/
(cfu·mL−1) Concentration温度/℃Temp 被感染的枝条数/总测试枝条数
Number of infected fragments/total tested fragments第1天Day1 第2天Day2 第3天Day3 第4天Day4 第5天Day5 第6天Day6 Vc450 4.17×106 24 0/6 0/6 0/6 0/6 1/6 1/6 △hapR-Vc450 3.17×106 24 0/6 0/6 1/6 4/6 4/6 5/6 ChapR-Vc450 5.17×106 24 0/6 2/6 2/6 4/6 5/6 5/6 Con — 24 0/6 0/6 0/6 0/6 0/6 0/6 Table 3. Infection of Acropora pulchra with the wild-type strain Vc450, △hapR-Vc450 and ChapR-Vc450 of Vibrio coralliilyticus
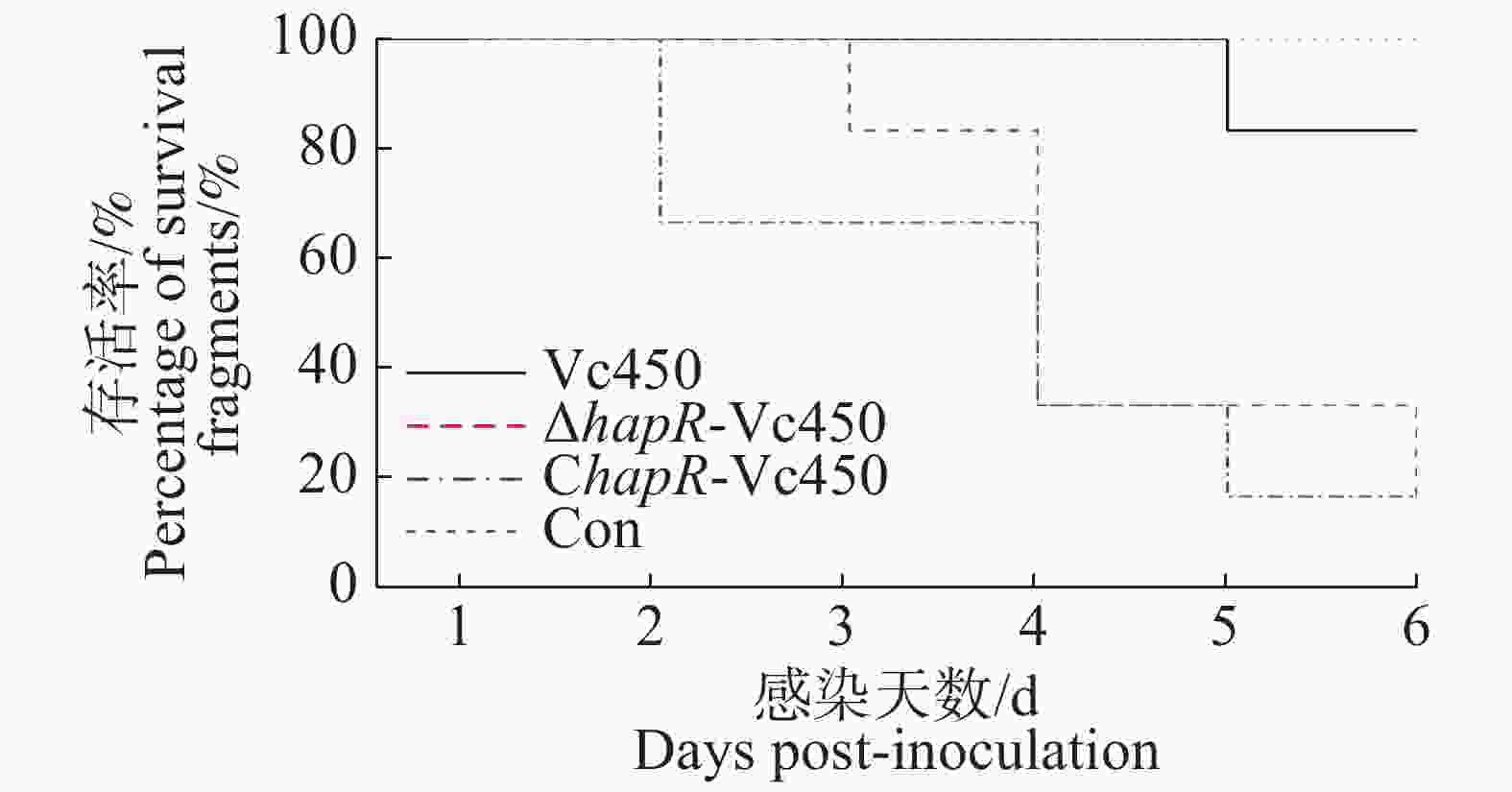
Figure 8. The percentage of survival fragments after inoculation of the Vc450, △hapR-Vc450 and ChapR-Vc450 of Vibrio coralliilyticus
2.1. 溶珊瑚弧菌Vc450 hapR基因敲除突变株及回补株的构建
2.2. hapR基因对溶珊瑚弧菌菌落及细胞形态的影响
2.3. hapR基因对溶珊瑚弧菌生长代谢的影响
2.4. hapR基因对溶珊瑚弧菌运动性的影响
2.5. hapR基因对溶珊瑚弧菌生物膜形成的影响
2.6. hapR基因对溶珊瑚弧菌毒力的影响
-
细菌的群体感应是细菌调控细胞密度和基因表达的一种细胞间的通讯方式,通过分泌信号分子协调细菌的基因表达及菌群的行为,如菌落相变、生物膜形成、生物发光和毒力因子表达等[5]。大多数细菌在细胞密度低时,通过群体感应系统抑制毒力或生物发光基因的表达,从而不产生毒力;在高于一定浓度后,细菌群体感应系统结合自诱导分子,启动hapR的表达,从而激活包括鞭毛合成、毒力因子基因等的转录,由此开启感染的过程。然而,在霍乱弧菌中,HapR能结合到aphA启动子结合区而抑制毒力基因的级联调控,最终阻止弧菌毒力岛上tcpPH启动子的激活[15],因此,在细胞密度低时,霍乱弧菌的hapR不表达,反倒激活了毒力因子如霍乱毒素CTX的表达;而在高细胞密度时,hapR被激活表达,从而抑制毒素的产生,但可促进细胞从生物膜中的脱离,从而开启快速的传染过程[16]。在本研究中,缺失hapR后溶珊瑚弧菌对鹿角珊瑚的毒力比野生型更高,显示溶珊瑚弧菌的群体感应系统对毒力基因的调控方式可能与霍乱弧菌相似,在hapR被敲除后,群体感应系统中细胞定殖和毒力基因被激活。此外,有报道显示,霍乱弧菌在自然环境中可以通过自发的hapR基因移码突变失活来抑制群体感应系统,获得较强的毒力,从而占据有利的生态位[17],但是在感染过程中,回补株ChapR-Vc450和突变株表现出同样的毒力,这可能是由于回补质粒上基因启动子是IPTG诱导性启动子,在感染的环境下由于没有IPTG诱导,从而不能表达HapR蛋白。
细菌的生物膜(Biofilm)是细胞分泌胞外基质的聚集体,包括细胞外多糖、蛋白质、核酸和脂质等[18],可以为细菌生长繁殖提供良好的微环境,保护细菌免受宿主免疫系统的攻击和外界环境的胁迫[19-20]。在霍乱弧菌中,hapR可以通过增强c-di-GMP的降解或直接抑制生物膜转录激活因子vpsT的表达来降低生物膜的合成量[21]。本研究中,溶珊瑚弧菌hapR基因缺失后,生物膜合成量增加,也证明了溶珊瑚弧菌中hapR的功能与霍乱弧菌hapR相类似。
 DownLoad:
DownLoad: