
-
植物在生长过程中会受到生物胁迫与非生物胁迫的影响。盐碱胁迫是非生物胁迫的一种,不但制约植物生长,同时也严重影响土地的利用。土壤的盐碱化是一个全球性的环境问题,据联合国教科文组织和粮农组织的不完全统计,世界上盐碱地已达到约9.5×108 hm2,相当于全球陆地总面积的7.23%,并且人类活动不断占用土地导致耕地面积逐渐减少,土壤盐碱化的程度正在逐年增加。中国是一个农业大国,盐碱地分布广泛,总面积约有9.9×107 hm2,土壤的盐碱化问题非常严峻[1]。土壤中可溶性盐分含量过高会对植物造成盐害,大部分植物在含盐量达0.3%的土壤中会受到伤害[2]。盐胁迫对植物造成的伤害主要有渗透胁迫、质膜伤害、离子平衡失调以及代谢紊乱[3]。在面对盐胁迫时,植物会采取一些相应的应答机制以减少对自身的危害,植物的应答机制是将细胞质内过多的Na+排出细胞外或将细胞质内的Na+区隔到液泡中来维持细胞质内正常的Na+浓度[4]。质膜上的Na+/H+逆转运蛋白SOS1(salt overly sensitive 1) 将Na+排出细胞,即SOS途径;而液泡膜上的Na+/H+逆转运蛋白NHX将Na+区隔到液泡中,即NHX途径[5]。植物利用PM-ATPase或V-ATPase和V-PPase所产生的跨膜H+浓度梯度,将细胞质中的Na+排到细胞外部或将其隔离到液泡中,从而调节细胞内Na+的浓度,调节细胞质的酸碱值及维持细胞内离子的稳态[6]。在植物中,Na+区隔化是植物抵抗盐碱的重要过程,主要依赖于液泡膜上的Na+/H+逆向转运蛋白,同时液泡膜上的H+-ATPases 和 PPases水解ATP释放的能量将H+泵出膜外,产生跨膜的电化学梯度,进而驱动Na+的逆向转运[7]。超表达液泡膜上Na+/H+逆向转运蛋白基因AtNHX1的拟南芥耐盐性得到明显提高[8]。将拟南芥AtNHX1基因转入棉花后,转基因植株能够在高浓度NaCl条件下正常生长[9]。将绿豆VrNHX1基因导入拟南芥,转基因拟南芥的耐盐性显著提高[10]。在番茄中超表达AtNHX1基因,可以促进番茄细胞在盐胁迫下K+从地下部向地上部运输,提高液泡中K+含量,使K+/Na+提高,减轻Na+毒害[11]。Paulo等用酵母双杂交的方法从人脑cDNA文库中找到NHE7的互作用蛋白,又用免疫共定位/沉淀的方法确定了其与SCAMP2可以相互作用[12]。GRAHAM等论证了哺乳动物中NHE5可以与SCAMP2相互作用[13]。之后在小麦中TaSCAMP1被证实可以与液泡膜的Na+/H+逆向转运蛋白发生相互作用,并被推测与植物耐盐相关[14],而在拟南芥中,利用BiFC实验和泛素酵母双杂实验证明AtSCAMP3可以与AtNHX2互作[15]。以上研究结果表明细胞中Na+/H+逆向转运蛋白的功能受到分泌载体膜蛋白SCAMP的调控。分泌载体膜蛋白(Secretory carrier membrane protein, SCAMP)是生物体内一种非常重要的膜蛋白,存在于许多真核生物中[16]。哺乳动物有5个SCAMP基因SCAMP1-SCAMP5,研究证明,哺乳动物中的SCAMP基因与神经递质的传递有关系,且涉及神经递质信号传递中膜的去极化和突触前膜分泌突触小泡或密集核心小泡(DCV)[17-18]。SCAMP3控制着晚期质体腔内囊泡的形成和多囊泡质体以及溶酶的分化[19],还在调节早期内体形成中起到重要作用[20]。植物SCAMP定位在根和花粉管等部分的质膜、初级质体或者反面高尔基体中[21]。为探究分泌载体膜蛋白(Secretory carrier membrane protein, SCAMP)的功能,笔者采用 PCR技术,从模式植物拟南芥中克隆出了5个SCAMP基因,利用生物信息学技术对拟南芥AtSCAMP蛋白家族结构进行了预测,并利用荧光定量PCR技术研究了它们在盐胁迫下的表达模式,旨在为进一步研究和完善植物的耐盐机理提供参考。
HTML
-
植物材料为哥伦比亚型拟南芥(Col-0),由本实验室保存。取野生型拟南芥种子清洗后用水浸泡,遮光放于4 ℃冰箱中3 d,然后把种子播种到1/2 MS固体培养基上,竖直培养10 d后,移栽幼苗至含有50 mmol·L−1 NaCl的1/2 MS培养基上盐胁迫培养,于盐胁迫培养0,1,3,5,7,9,11,13 h时分别取幼苗的根和叶,并立即放入液氮中速冻,−80 ℃保存以备提取RNA。
-
依据说明书,使用植物总RNA提取试剂盒(RNAplant Plus Reagent, DP437;TIANGEN公司生产)提取RNA;采用TransScript One-Step gDNA Removal 和 cDNA Synthesis SuperMix进行RNA反转录(TaKaRa, R047A)。将样品用手拨打轻轻混匀后42 ℃反应15 min;85 ℃反应5 s;4 ℃保存。获得的cDNA用Actin[22]引物扩增,以检测cDNA的质量。
-
根据拟南芥基因组网站 (www.arabidopsis.org) 上公布的AtSCAMP基因序列 [AtSCAMP1(基因登录号:AT1G61250),AtSCAMP2(基因登录号:AT1G11180),AtSCAMP3(基因登录号:AT2G20840),AtSCAMP4(基因登录号:AT1G03550),AtSCAMP5(基因登录号:AT1G32050)]设计全长引物。引物序列为:AtSCAMP1-F:ATGGCTAATCGTTATGATCC,AtSCAMP1-R:TCAAACAGCGGCTCTCAAG;AtSCAMP2-F:ATGGGTGGTCGTTACGAT,AtSCAMP2-R:TCATATGGCAGCTCTCATG;AtSCAMP3-F:ATGTCTCGCTACCAGTCTC,AtSCAMP3-R:TCAGAGAGCTGCCATCATT;AtSCAMP4-F:ATGGCACGACACGATCCT,AtSCAMP4-R:TCATAGTGCACGCATCAAGGT;AtSCAMP5-F:ATGAATCGCCACCACGATC,AtSCAMP5-R:TCACTTGTTTCCCCTAAAGTAG。将1.2中得到的cDNA混合作为模板进行扩增。将片段进行胶回收后连接T载体,转化大肠杆菌后选取菌落PCR正确的样品进行测序。
-
采用Protparam 在线软件(
http://www.expasy.org/tools/protparam.html )分析SCAMP蛋白理化性质。采用SOPMA 在线软件(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl )预测SCAMP蛋白的二级结构。采用TMHMM在线软件(http://www.cbs.dtu.dk/services/TMHMM-2.0/ )预测SCAMP蛋白的跨膜结构。采用MEGA7.0软件对SCAMP蛋白做进化树分析。 -
利用oligo7软件设计AtSCAMP基因的荧光定量PCR引物(表1)。以Actin作为内标基因进行荧光定量PCR分析。采用染料法实时荧光定量PCR试剂盒(TaKaRa, SYBR Premix EX TaqTM(Perfect Real time),R820A)进行试验,反应体系按照说明书进行。实时荧光定量PCR的扩增条件为94 ℃预变性30 s,94 ℃变性5 s,60 ℃退火30 s,40个循环,结束后读取荧光信号:95 ℃,15 s;60 ℃,15 s;95 ℃,15 s。采用比较CT法计算目的基因的相对表达量,目的基因的相对表达量=2−ΔΔCt,其中ΔΔCt= (Ct目的基因−Ct内参基因) 实验组−(Ct目的基因−Ct内参基因) 对照组。进行3次生物学重复,取平均值进行作图。
名称Primer 引物序列Primer sequence Actin F: TATGAATTACCCGATGGGCAAG R:TGGAACAAGACTTCTGGGCAT AtSCAMP1-qPCR F: TCGACCACTCTACCGTGCCTTC R: TCGCTGCTTCCCGTCTCATTTG AtSCAMP2-qPCR F: CTTTGCCTCCCGAACCTGCTG R: CCAAGCGGTAGTAACGGCGATG AtSCAMP3-qPCR F: TTGCGGCGGTGGCTCCTC R: GCCTCCTGCTTCATCTCTGCTG AtSCAMP4-qPCR F: ACGTGTTCCACATCGCGTTCTG R: TGCCACTCCCTCGGAAGTATGC AtSCAMP5-qPCR F: GCTTGCTGACTGGGAAGCTGAG R: TCCTCATGGCTCGGTAGAGTGG Table 1. Primer sequence for real-time quantitative PCR
1.1. 实验材料
1.2. 拟南芥RNA的提取和cDNA反转录
1.3. AtSCAMP基因的克隆
1.4. 生物信息学分析
1.5. 实时荧光定量PCR
-
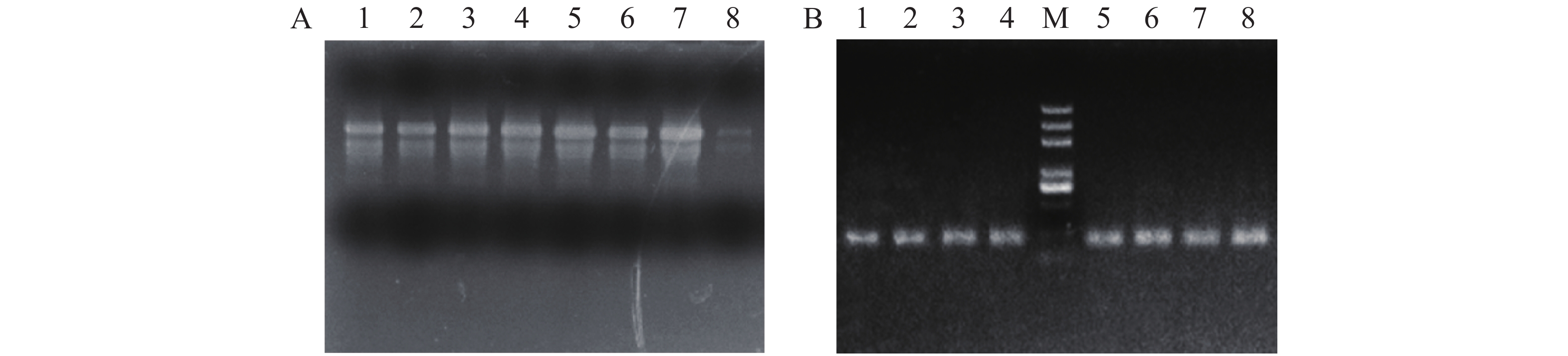
提取的RNA用1%的琼脂糖检测(图1A),从图1A可见,叶片中的RNA的28S 和18S 完整,说明 RNA 质量比较好。用反转录试剂盒进行cDNA反转录,得到的cDNA用内参基因Actin引物进行扩增(图1B),从图1B可见,产物没有非特异性扩增或者引物二聚体,表明反转录得到的cDNA质量较好。
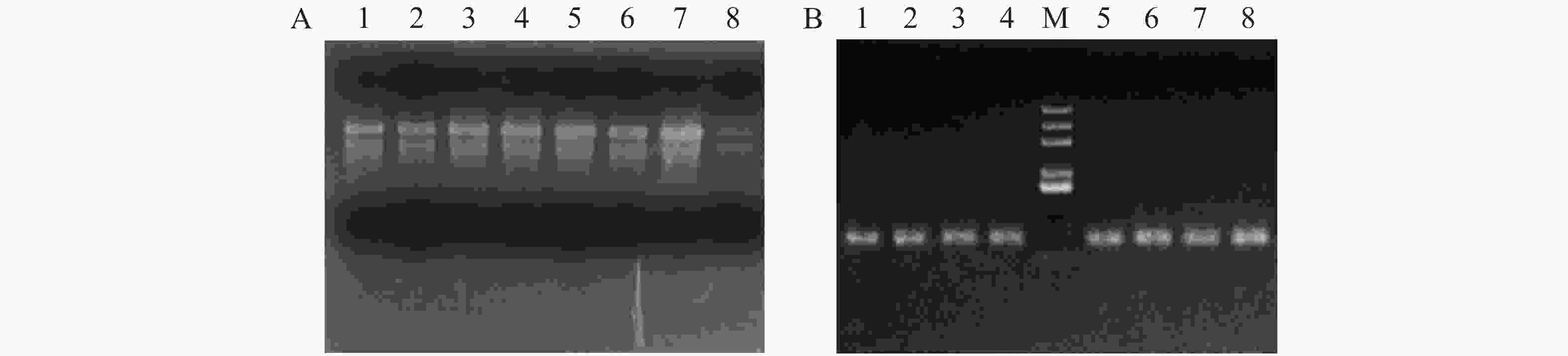
Figure 1. RNA and cDNA amplification of Arabidopsis thaliana leaves
-
在拟南芥基因组网站(www.arabidopsis.org) 上查找公布的AtSCAMP基因序列设计引物,扩增得到长度为750~1 000 bp之间的片段(图2)。测序结果表明得到的目的片段分别为870,894,849,852,795 bp,与AtSCAMP1-5序列比对正确,基因克隆成功。
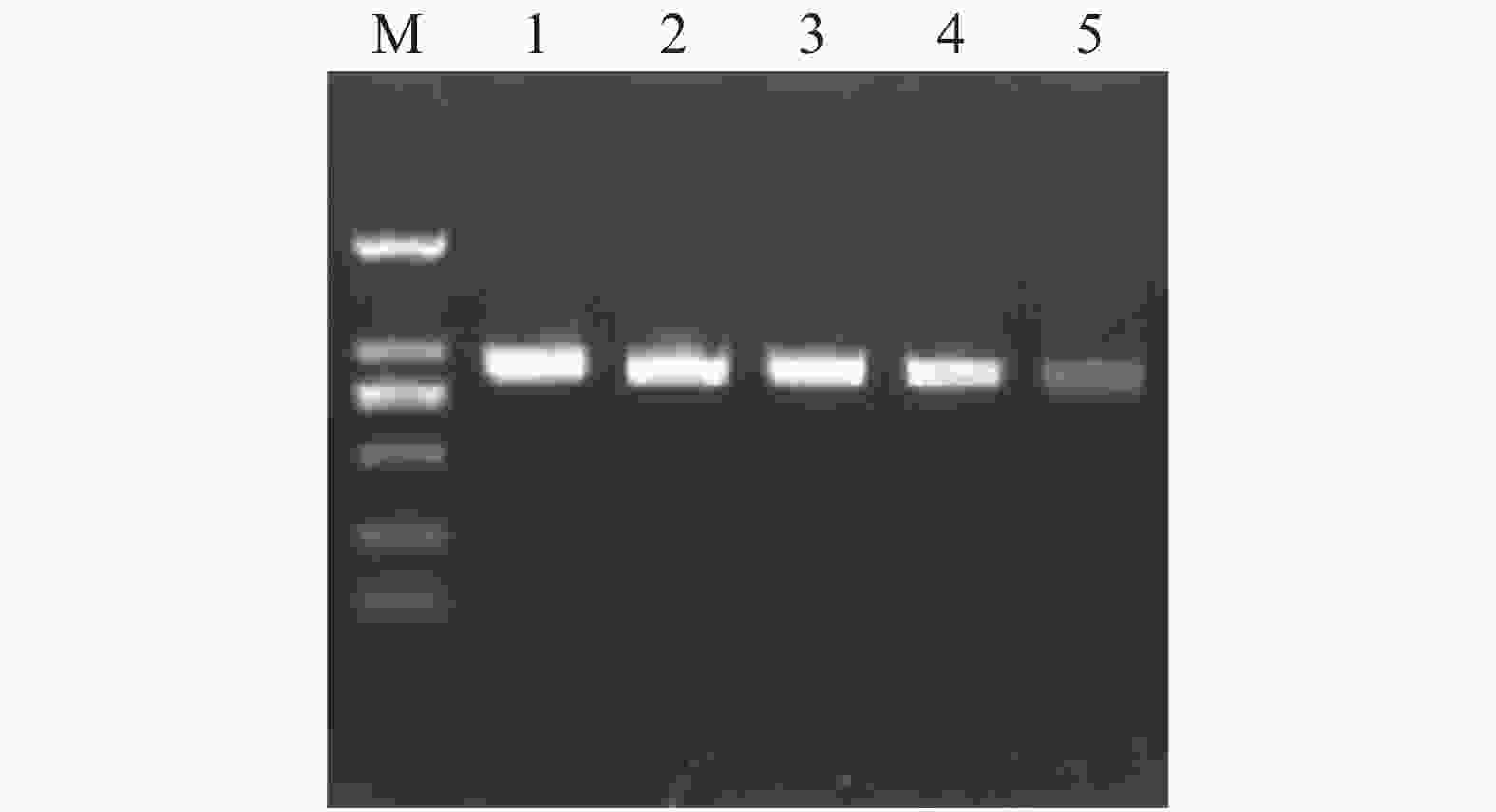
Figure 2. Amplification of AtSCAMP genes
-
AtSCAMP1-5蛋白的氨基酸组成见表2,用 Protparam 在线软件分析AtSCAMP1-5蛋白的理化性质。预测结果:AtSCAMP1-5蛋白质相对分子量分别约为32.6,33.2,32,31.9,30.1 kDa;AtSCAMP1-5蛋白质等电点(pI)分别是 8.87,9.18,8.32,6.60,7.66;AtSCAMP1-5蛋白质元素组成分别是C1526H2321N389O392S7,C1536H2354N402O404S8,C1491H2274N368O389S12,C1485H2264N372O393S8,C1409H2135N349O361S10;AtSCAMP1-5蛋白质亲水性分别是0.185,0.162,0.179,0.120,0.233,属于疏水蛋白,不稳定指数均大于40,表明它们均为不稳定蛋白。
SCAMP Ala(A) Arg (R) Asn (N) Asp (D) Cys (C) Gln (Q) Glu (E) Gly (G) His (H) Ile (I) AtSCAMP1 11.4 6.20 2.80 3.80 1.00 3.50 5.50 6.90 1.40 6.90 AtSCAMP2 11.4 2.80 3.70 5.10 1.00 3.40 3.70 8.40 1.00 7.10 AtSCAMP3 9.6 3.80 2.10 3.50 1.10 2.80 6.00 6.40 1.80 9.60 AtSCAMP4 10.6 1.00 3.90 3.20 1.10 2.50 6.70 6.40 2.10 7.80 AtSCAMP5 8.3 3.50 3.00 5.70 1.90 2.70 4.20 6.80 2.70 9.50 SCAMP Leu (L) Lys (K) Met (M) Phe (F) Pro (P) Ser (S) Thr (T) Trp (W) Tyr (Y) Val (V) AtSCAMP1 9.30 4.50 1.40 7.60 6.60 3.50 2.80 2.40 4.20 8.30 AtSCAMP2 10.80 4.00 1.70 7.40 5.40 4.40 2.70 2.00 4.00 6.10 AtSCAMP3 8.20 5.70 3.20 7.40 6.40 5.70 3.90 2.50 3.90 5.70 AtSCAMP4 9.90 4.90 1.80 7.10 7.10 3.20 5.30 2.10 3.90 6.00 AtSCAMP5 10.20 6.10 1.90 8.00 5.30 4.90 2.30 2.70 3.80 6.10 Table 2. Amino acid composition of SCAMP proteins in Arabidopsis thaliana
% 用 SOPMA 在线软件对AtSCAMP家族蛋白二级结构进行预测,分析发现,AtSCAMP蛋白家族二级结构中都包含有4种构象:α−螺旋(Alpha helix, Hh),β−折叠(Beta turn, Tt),直链延伸(Extended strand, Ee),无规则卷曲(Randon coil, Cc),其中含量最高的是α−螺旋(Hh),约占蛋白质二级结构总量44%~50%,含量第2的是无规则卷曲(Cc),约占总量32%~36%,含量第3的是直链延伸(Ee),约占11%~16%,含量最少的蛋白二级结构是β−折叠(Tt),含量约为4%~6%(表3)。
AtSCAMP α−螺旋(Hh) β−折叠(Tt) 直链延伸(Ee) 无规则卷曲(Cc) AtSCAMP 1 49.48 5.88 11.76 32.87 AtSCAMP 2 45.79 5.72 13.47 35.02 AtSCAMP 3 48.94 6.03 15.25 29.79 AtSCAMP 4 47.00 3.89 14.49 34.63 AtSCAMP 5 44.70 4.55 14.77 35.98 Table 3. Contents of secondary structures of AtSCAMP proteins in Arabidopsis thaliana
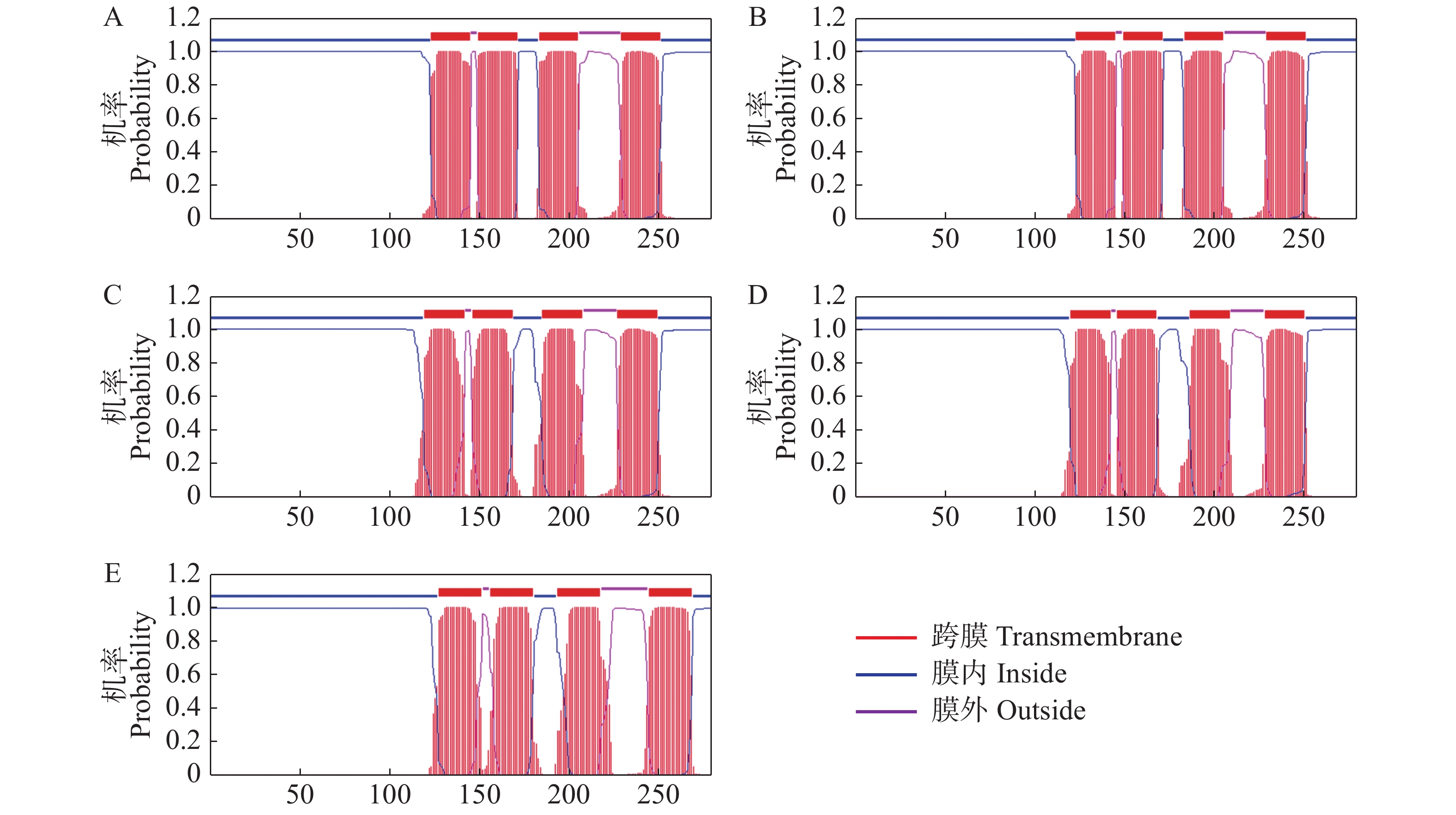
% 通过TMHMM在线软件对AtSCAMP蛋白家族进行跨膜结构域的预测,结果(图3)表明,AtSCAMP1-5蛋白均具有4个跨膜结构域,N端和C端均在细胞膜内。
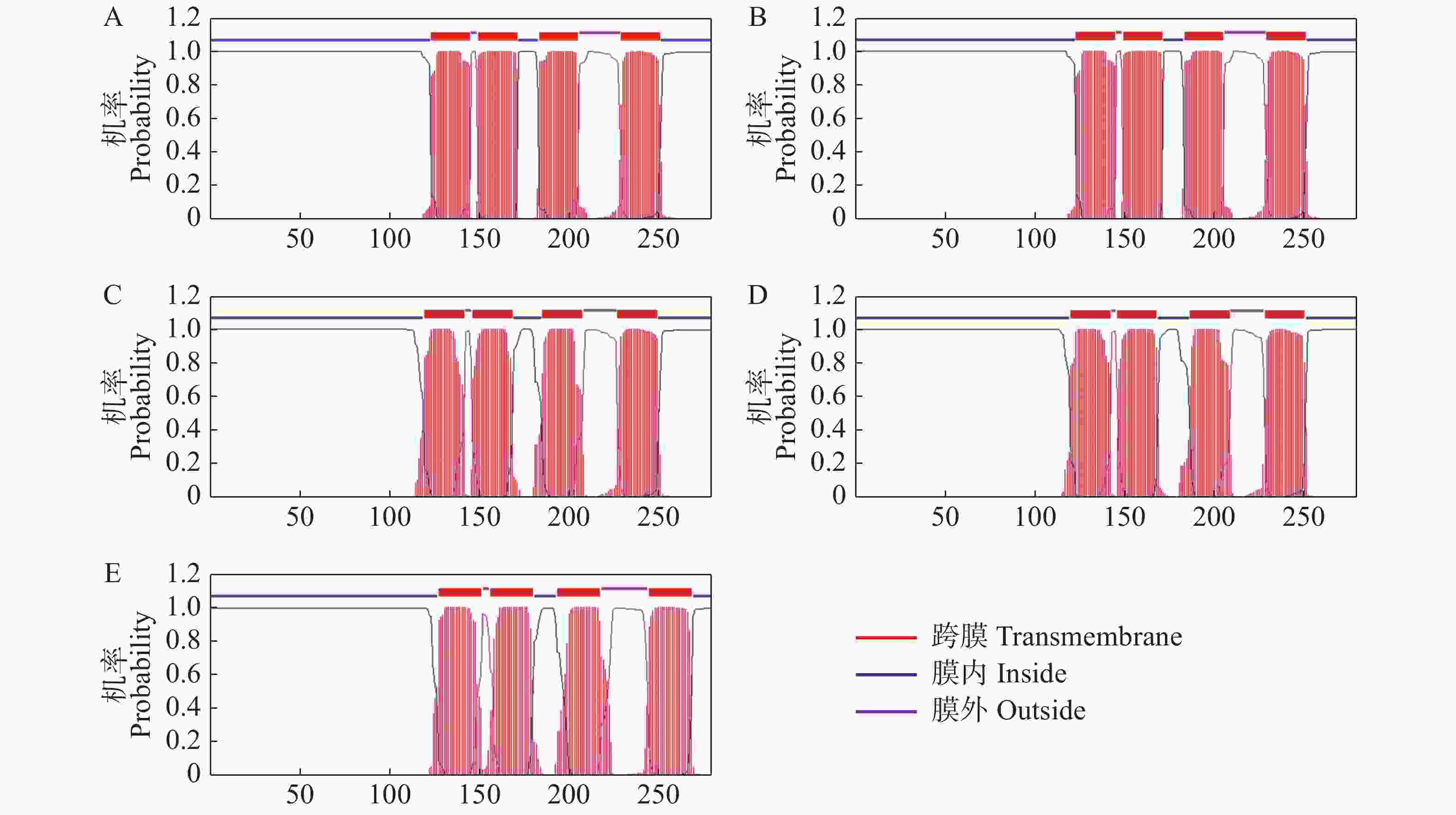
Figure 3. Transmembrane domain prediction of SCAMP proteins in Arabidopsi thaliana
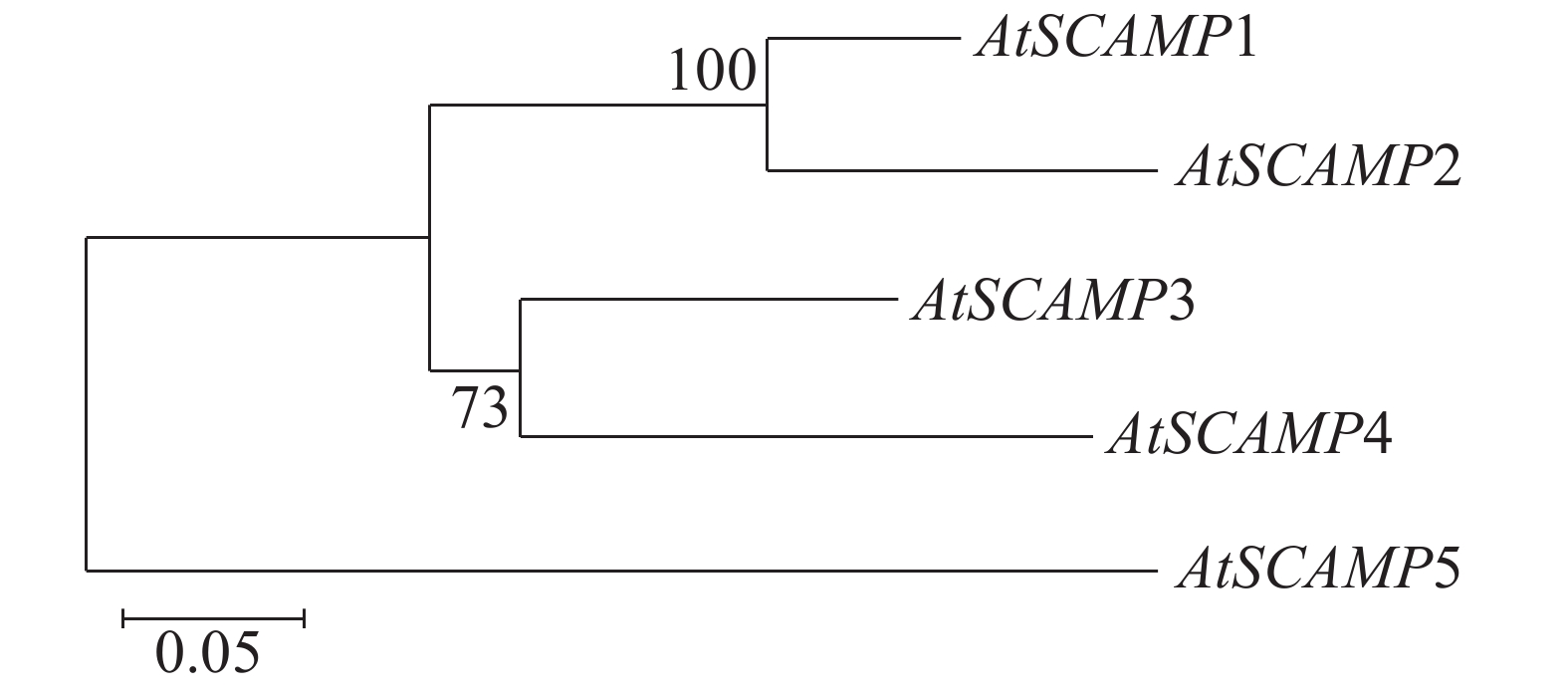
通过MEGA7.0软件对AtSCAMP蛋白家族进行序列比对,结果(图4)可见,AtSCAMP1与AtSCAMP2蛋白在进化关系上比较近,AtSCAMP3与AtSCAMP4蛋白在进化关系上较近,而AtSCAMP5蛋白与其他蛋白进化关系最远。
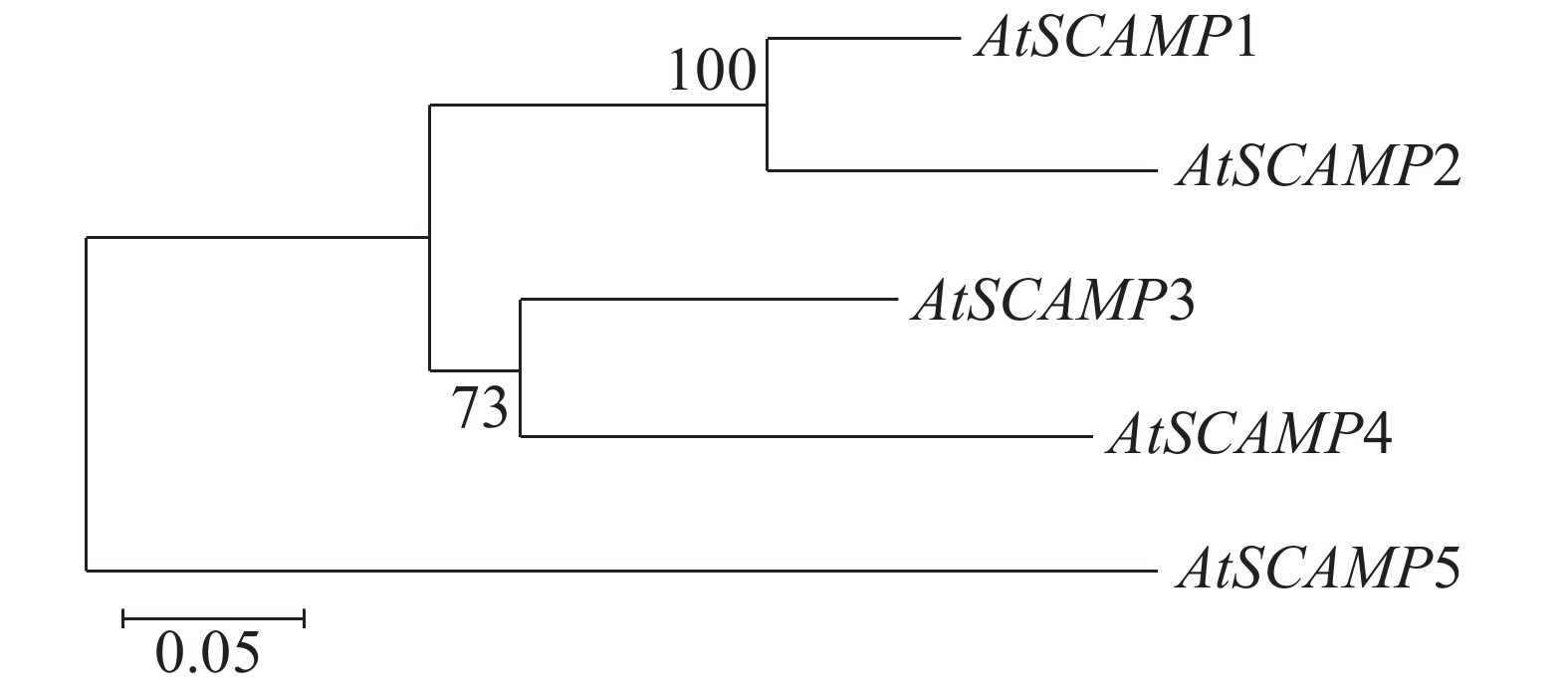
Figure 4. Phylogenetic tree analysis of SCAMP proteins in Arabidopsis thaliana
-
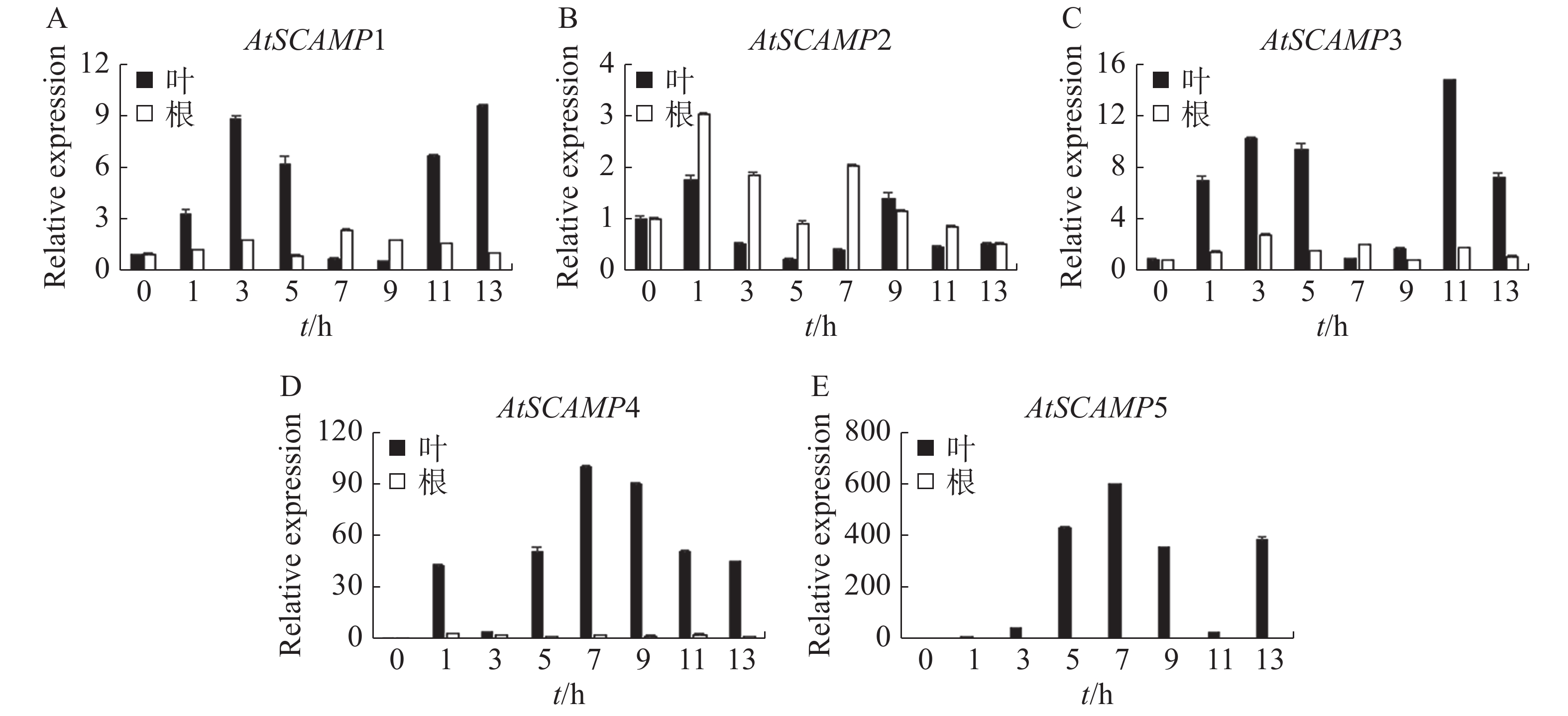
使用qRT-PCR方法研究AtSCAMP基因在50 mmol·L−1 NaCl胁迫下的表达情况。结果(图5)表明,在50 mmol·L−1 NaCl胁迫下,AtSCAMP1-5基因在拟南芥的根、叶中都呈上调表达,其中AtSCAMP1,AtSCAMP3,AtSCAMP4,AtSCAMP5基因在叶中的表达明显高于根中。AtSCAMP1在叶中初期表达量随时间升高而升高,3 h达到第1个峰值,约为起始表达量的9倍,此后表达量下降,在盐胁迫诱导9 h后表达量又逐渐上升;而根中AtSCAMP1基因的表达量没有明显的变化(图5A)。AtSCAMP2在叶中表达量高于根,初期随处理时间增加而增加,相对于其他AtSCAMP基因,AtSCAMP2在根和叶中的表达量受盐胁迫的变化都不太明显(图5B)。AtSCAMP3在根和叶中受盐胁迫诱导表达的模式与AtSCAMP1比较类似(图5C)。叶中AtSCAMP4和AtSCAMP5基因受盐胁迫诱导变化明显,盐胁迫处理后表达量迅速上升,而根中这2个基因在50 mmol·L−1 NaCl处理后的表达量变化都不明显(图5D-E)。
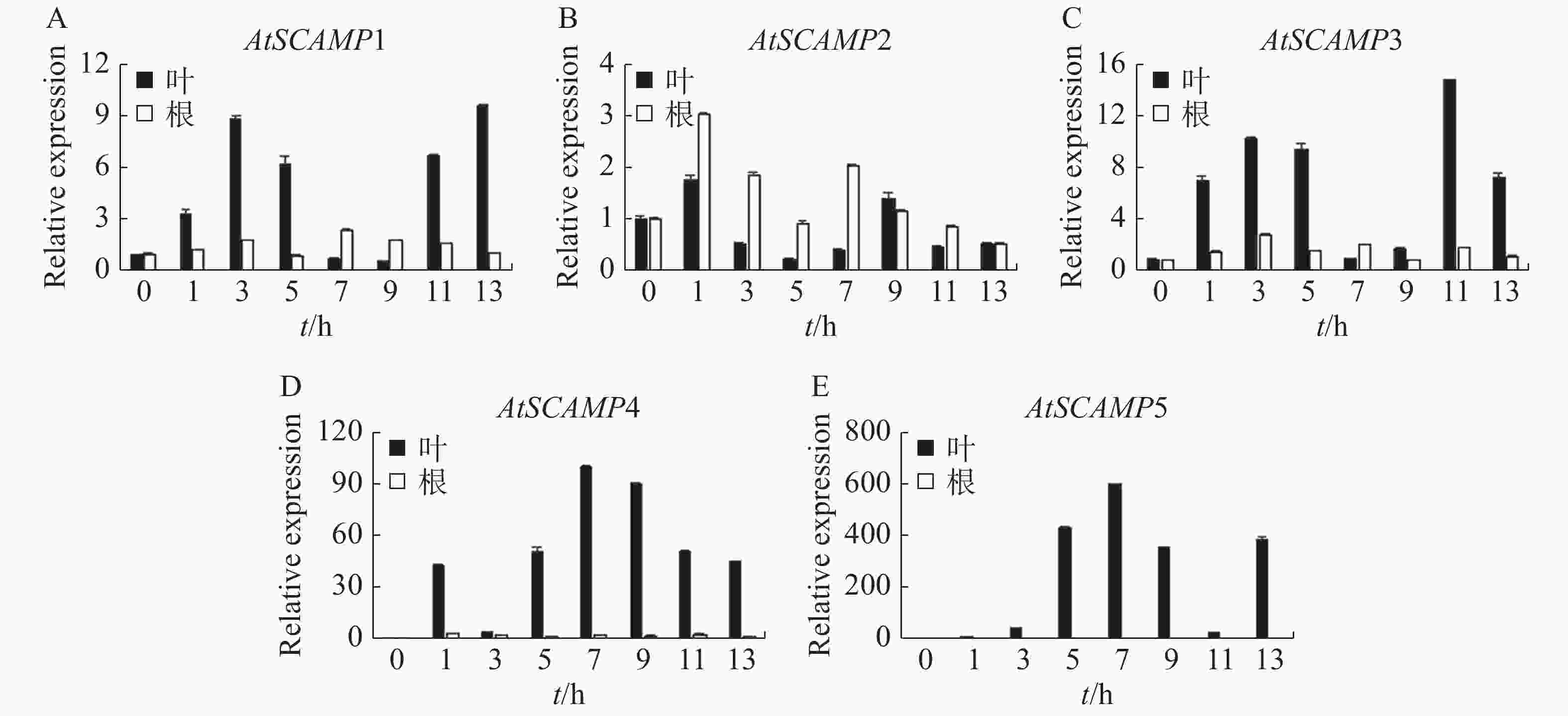
Figure 5. The expression analysis of AtSCAMP under salt stress
2.1. RNA提取及cDNA反转录
2.2. AtSCAMP家族基因的克隆
2.3. 拟南芥SCAMP的生物信息学分析
2.4. AtSCAMP基因在盐胁迫下的表达分析
-
研究植物的耐盐机制有助于培育耐盐植物新品种,提高盐碱地中植物的产量,合理利用盐碱地。SCAMP蛋白被证实是一种分泌载体膜蛋白质,不论在哺乳动物中还是植物中都被证明可以与Na+/H+转运蛋白进行互作,且生化分析显示NHE7的C端和SCAMP第2个、第3个跨膜结构域中的细胞质环优先结合,SCAMP2中缺少这一段的蛋白与NHE7的结合明显减弱[12-13]。笔者发现,AtSCAMP蛋白具有4个跨膜结构域,其N端和C端均在膜内,为此推测其定位在膜上,说明它们与细胞膜上的Na+/H+逆向转运蛋白互作是有可能的。已有研究[23-27]表明,植物体内液泡膜上的Na+/H+逆向转运蛋白(NHX)和细胞质膜上的Na+/H+逆向转运蛋白(SOS1)都在植物耐盐机制中发挥着重要的作用。小麦SCAMP基因转化到拟南芥和水稻中能够明显提高转基因拟南芥和水稻的耐盐抗性[15]。笔者发现从拟南芥中克隆的5个SCAMP基因均在受到盐胁迫诱导后上调表达(图5),由此推测SCAMP基因与植物耐盐性相关。
有研究[5]表明SCAMP3在调节早期内体形成中起到重要作用。笔者通过对盐胁迫下SCAMP基因表达量具体分析发现,在盐胁迫初期时,SCAMP1-5基因都呈现上升趋势,与该结果相似;实时荧光定量PCR结果还显示在盐胁迫下SCAMP基因表达量在开始上升后都存在1次下降趋势,由此推测SCAMP可能在应对盐胁迫时并没有单独发挥作用,而是可能与其他蛋白互作。文献[15]也发现在拟南芥中AtSCAMP3可以与AtNHX2互作。文献[14]也发现在小麦中TaSCAMP1可以与Na+/H+逆向转运蛋白产生荧光发生相互作用,且表达量在盐处理时升高。这些结果都说明,AtSCAMP可能参与了植物的耐盐调控途径。本研究克隆得到的AtSCAMP1-5基因均已构建到植物表达载体和酵母表达载体中,本实验室下一步将深入研究这些基因的功能。
 DownLoad:
DownLoad: