-
热带珊瑚礁是地球上生物多样性最丰富的生态系统[1]。在海洋的众多生物圈中,珊瑚礁生态系统是非常重要的组成成分,因其具有高初级生产力和丰富的物种多样性,被誉为“海底热带雨林”[2]。珊瑚礁生态系统主要分布在海水表面温度20℃等温线内[3]。20 世纪珊瑚白化,世界各地珊瑚(Scleractinia)大量死亡,对海洋生态系统造成了严重影响[4]。引起珊瑚白化和死亡的原因有很多,海洋温度升高和极端气候事件频繁发生是最主要原因[5]。由于全球气候变暖,世界各地珊瑚礁白化频率和程度在增加,珊瑚礁覆盖大幅下降[6]。中国南海经常发生由于温度变化导致的珊瑚白化事件[7],在过去40 年间,中国南海北部东沙环礁水温升高2℃,导致 40% 的珊瑚以前所未有的速度白化[8]。随着气候变暖的加剧,未来,异常高温事件发生会越发频繁,海洋热浪频繁发生,对涠洲岛珊瑚礁造成的威胁将越来越严峻。分离纯化虫黄藻是研究珊瑚-虫黄藻共生关系的核心基础。通过获得单一藻株,能够精确分析虫黄藻与宿主之间的代谢互惠机制。
1962 年,Freudenthal[9] 从石珊瑚中分离出虫黄藻,首次正式提出虫黄藻属于共生甲藻属(Symbiodinium)。由于虫黄藻形态大小相似、种类繁多,鉴定效率和准确率很低,直至 1994年 ,研究者开始利用分子生物学方法进行物种分类[10]。此后,研究人员不断从石珊瑚(Scleractinia)、软珊瑚(Alcyonacea)、海葵(Actiniaria)、砗磲(Tridacna)、水母(Hydrozoa)、海绵(Porifera)等海洋生物中发现虫黄藻,并一直在进行对虫黄藻命名的不断研究和探索。直到 2018 年,LaJeunesse[11]根据分子、形态学、生理学和生态学数据,虫黄藻的 A—G 7 个系群已被正式定义,并重新描述为 7 个已定属:Symbiodinium、Breviolum、Cladocopium、Durusdinium、Effrenium、Fugacium、Gerakladium,以及 2 个未定属:clad H 和clade I。
Cladocopium 是具有功能多样性和遗传多样性的虫黄藻属[11],其分布最广泛,种类最多,主要存在于东太平洋和印度洋海域的亚热带-热带海洋的珊瑚礁中[12]。研究表明,在野生环境中,由不同系群虫黄藻主导的同一珊瑚物种,其生长速度和光合产物转运表现出显著差异[13-14],与以 Cladocopium 属为主导的珊瑚相比较,以 Durusdinium属为主导的珊瑚通常表现出较低的生长速率[15],原因是Durusdinium储存的脂质含量更低,导致其向宿主转运的光合产物减少[16-17]。此外,在多数珊瑚礁区,以 Durusdinium 属为主导的珊瑚普遍被认为具有更强的耐热性[18]。然而,对于高纬度礁区的珊瑚种群,内共生的 Cladocopium 和Durusdinium 属虫黄藻所赋予宿主的耐热能力则表现出相当的水平[19]。印度–太平洋海域中,许多珊瑚共生虫黄藻亚系群如C. goreaui、CladocopiumC7和 Symbiodinium A13 虫黄藻亚系群是热敏感的[20-22]。已有研究结果表明,南海北部礁区珊瑚普遍与 Cladocopium属的虫黄藻共生,且珊瑚的主导虫黄藻亚系群大多为 C. goreaui,属于南海北部、琉球群岛、韩国济州岛等礁区/群落的热敏感型虫黄藻。且南海北部珊瑚礁区(涠洲岛、雷州半岛、大亚湾、香港等)的丛生盔形珊瑚主导虫黄藻均为Cladocopium C1[23-27]。
1944年Kawaguti[28]对虫黄藻进行离体培养探究,1967年Ahles[29]尝试对来自 70个不同宿主的虫黄藻进行体外培养, 却发现只有不到25%的虫黄藻能够在 ASP-8A 培养基短暂存活。2005年之前,国内对共生藻的研究还是一片空白。直到朱葆华[30]尝试用 ASP-8A培养基进行虫黄藻离体培养, 但是培养7 d后,所有虫黄藻全部死亡。2014年之前,国内珊瑚共生虫黄藻的分离培养工作处在初步探索阶段,这个时期开展虫黄藻的分离培养主要存在两方面问题,1)细菌等微生物污染;2)分离的虫黄藻不能进行有效地传代[31-32]。目前,国内研究人员已经使用改良的L1、Z1等培养基培养出了不同系群的离体虫黄藻[33-36]。然而,如何高效有目标的分离共生虫黄藻、分离后的藻种在稳定传代后是否改变其生理活性和细胞结构,以及在极端温度胁迫中的功能仍有待进一步探索。本研究从美丽鹿角珊瑚上分离出与其共生的虫黄藻 Cladocopium C1 sp.,并对其进行稀释培养逐渐纯化,结合一代测序和高通量测序鉴定虫黄藻的分离和纯培养的程度。为我们深入了解它们在与珊瑚的共生中所扮演的角色以及它们与珊瑚之间相互作用的机制提供科学基础。也为我们应对珊瑚礁退化和保护珊瑚礁提供更加多的科学依据。
-
1)取样。采用水肺潜水的方式从南海北部的涠洲岛海域采集美丽鹿角珊瑚(Acropora formosa)。2)养殖缸养殖。将运回的活珊瑚进行过水处理,适应养殖室内的水环境。采集的活珊瑚样品放在300 L养殖缸中培育,方便后续取样。水质条件:盐度 35‰,温度 24 ~ 27℃,Ca2+含量为 360 ~ 420 mg·L−1,Mg2+含量 1 200 ~ 1 350 mg·L−1,碳酸氢根(HCO3−)浓度指数(KH)为 7 ~ 8 dKH,pH 为 8.1 ~ 8.2,NO3−为 5 mg·L−1以下,PO42−为 1 mg·L−1以下。每周换水 100 L,维持缸内水质稳定。3)分离纯化培养。取活珊瑚的组织至无菌海水中,在超净工作台中用冲洗或者剪切的方式,使珊瑚在六孔板的无菌海水中释放共生虫黄藻。使用倒置显微镜镜检观察海水,确定水样中有虫黄藻。通过离心的方式,4 000 r·min−1,5 min,4℃初步将共生虫黄藻与其它微藻以及细菌分离。使用改良的L1培养基,并在实验的过程每两周补充三联抗生素(青霉素200 μg·mL−1,链霉素100 μg·mL−1和卡那霉素100 μg·mL−1)处理,防止细菌等污染虫黄藻样品。将分离后的共生虫黄藻培养板放置在恒温光照培养箱中。培养条件:光照:50 μmol·m−2·s−1;温度:(25 ± 1)℃;光照时长为12 h(光照):12 h(黑暗)。每隔一周使用倒置显微镜镜检观察分离的共生虫黄藻的生长情况。每两周对虫黄藻进行进一步离心、稀释、转接和培养。4)分子鉴定。固体培养基、斜面上或者是液体培养基中取样镜检,观察是否有污染物。先进行形态学鉴定,正常状态的虫黄藻细胞核用光学显微镜可以清晰的观察到,有密集的核染色质。而裂解的虫黄藻,失去完整的细胞结构,并且不再是圆形,细胞增大两倍并且有空的气泡。观察确认是虫黄藻后,通过 Sanger 测序及高通量测序,鉴定分离的虫黄藻是否为共生虫黄藻,以及纯化情况。
-
提取珊瑚中的虫黄藻需要对样品进行预处理。用无菌的镊子从珊瑚珊瑚片段的新生触手区域剪取分离并收集约50 mg的纯净共生组织。将收集的组织置于适量的无菌过滤海水中,通过离心、重悬的方式进行预处理。每个独立的珊瑚样本(n = 3)均按此流程重复进行预处理和DNA提取。用 DNeasy Plant Mini Kit(QLAGEN,Hilden,Germany)试剂盒,按照制造商说明书进行提取 DNA。提取的 DNA 用液氮速冻30 min以上,再放置在-80℃冰箱保藏,以用来 PCR。将获取的DNA作为模板,通过琼脂糖凝胶电泳检测DNA的质量。DNA质检后,进行PCR扩增。 PCR的引物是ITSintfor2(F:5'-GAATTGCAGAACTCCGTG-3'和R:5'- GGGATCCATATGCTTAAGTTCAGCGGGT-3')[37]。PCR反应体系:20 μL的体 系,和10 ng模板DNA,珊瑚共生虫黄藻DNA片段大小为281 ~ 300 bp。PCR扩 增的条件:95℃变性3 min,35次循环30 s,55℃退火30 s,72℃扩增10 min。 PCR一式3份。将扩增的ITS2产物纯化后,用干冰运送至上海美吉生物公司,将样品在Illumina Miseq平台上进行成对末端测序。
-
使用高速冷冻离心机将虫黄藻细胞离心沉淀,离心条件500 g,5 min后,留 沉淀,弃培养基。离心收集体积为1 mL虫黄藻细胞,后使用磷酸缓冲盐溶液 (PBS)进行清洗,反复操作3次,留沉淀,转移至1.5 mL无菌无酶的冻存管中,置入液氮速冻。其余步骤见1.2。
-
悬浮的虫黄藻细胞通过离心500 g,5 min 留沉淀,弃培养基。离心收集体积为1 mL的虫黄藻细胞,后使用PBS进行清洗,反复操作3次,留沉淀,弃PBS。将处理后的细胞沉淀使用电子显微镜固定液吹打重悬,于室温固定2 h, 再转移至4℃保存。扫描电子显微镜:使用乙醇梯度脱水。干燥后将虫黄藻细胞样本贴于导电碳膜双面胶上,放入离子溅射仪样品台上喷金30 s,扫描电子显微镜下观察[38]。透射电子显微镜:室温脱水;包埋后将虫黄藻细胞样品放入37℃烤箱过夜。使用包埋板包埋,使用超薄切片机切片。制作铜网切片,透射电子显微镜下观察[39]。
-
用无菌剪刀从美丽鹿角珊瑚组织块上剪下1 cm3左右的组织,在无菌无酶离心管中剪碎。通过不同方式对含有共生虫黄藻原始溶液进行预处理,由图1可知,不经过任何预处理的共生虫黄藻溶液,密度高,但杂质较多,有大量的污染物(图1-A);使用高速冷冻离心机经过4 000 g,15 min,4℃处理,杂质的聚团现象明显,但细胞个数保留的相对比较多(图1-B);使用高速冷冻离心机经过7 000 g,10 min,4℃杂质比较少,单个细胞的距离较大,与黏液分离,后期生长粘连的可能性较小(图1-C);将生虫黄藻溶液的母液稀释1 000倍,培养中的虫黄藻密度较低,难以镜检到单个细胞(图1-D)。图1-E和图1-G分别为使用高速冷冻离心机经过7 000 g和4 000 g, 5 min,4℃处理后的虫黄藻。将不同预处理的细胞转接到六孔板,分别培养5 d,再次用倒置显微镜观察不同预处理的效果,如图1所示,经过7 000 g,5 min,4℃处理的共生虫黄藻细胞基本没有粘连现象,并且可以传代,没有明显的污染物(图1-F);经过4 000 g,5 min,4℃的共生虫黄藻细胞存在大量的污染物,并且细胞粘连严重,难以进行后续的单细胞分离试验(图1-H);而梯度稀释的共生虫黄藻没有传代,仍然无法镜检到细胞。

Figure 1. Symbiodiniaceae cultured after different pretreatment methods under inverted microscope
-
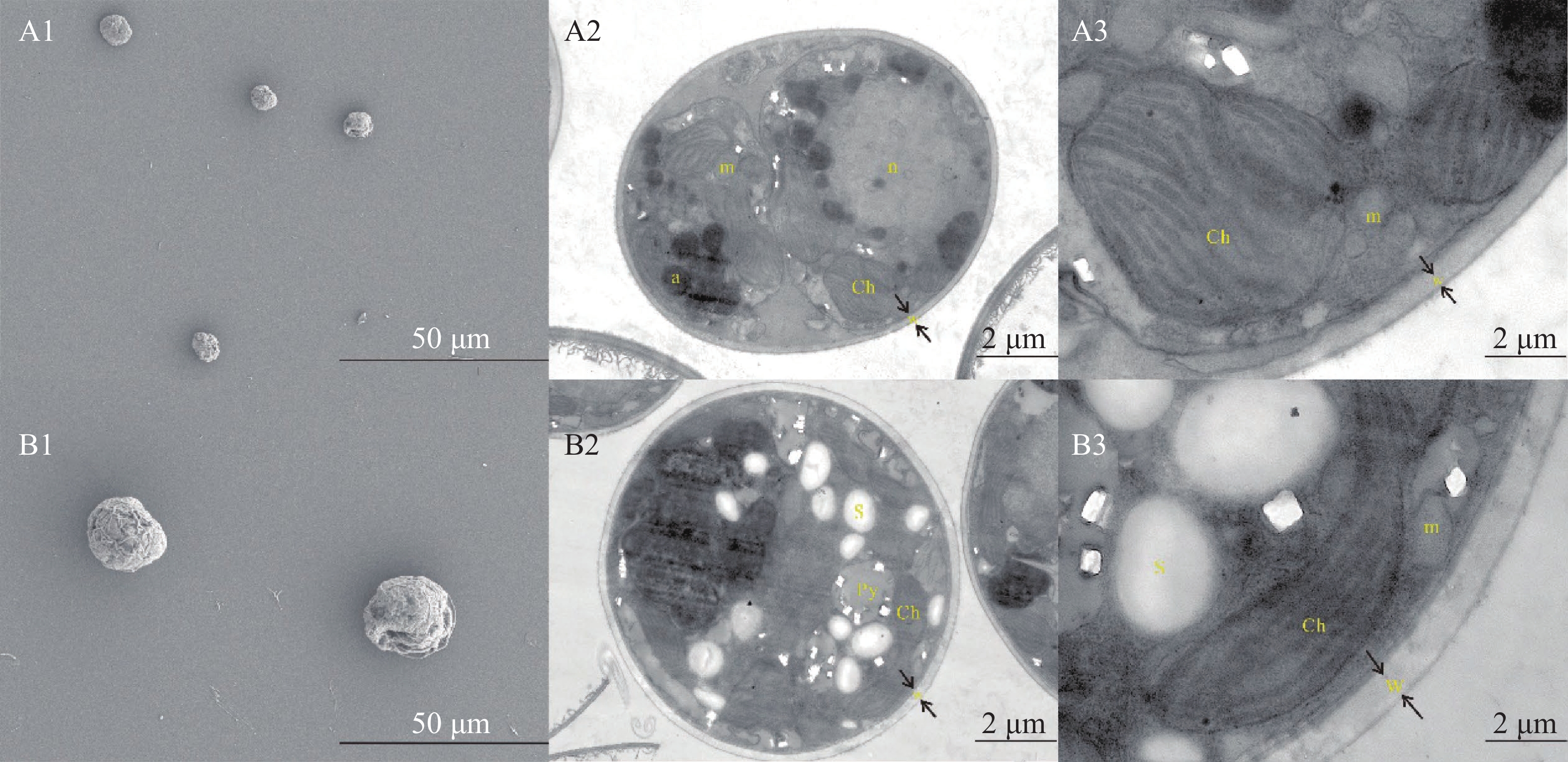
经过五代的分离纯化,半年后,仅单瓶虫黄藻培养液培养时间过长(一般为 2 个月以上),没有及时转接时,虫黄藻才会有代谢黏液,正常情况下,共生虫黄藻已经能够稳定的传代,并且未观察到其他微生物污染物(图2)。 经过分离培养的虫黄藻Cladocopium C1 sp.未观察到鞭毛(图3),但可以从不同的剖面观察细胞(图3- A2、图3-B2)。这些分离的虫黄藻Cladocopium C1 sp. 细胞有着良好健康的细胞结构,包括细胞核内含有多个椭圆形染色体;细胞质内,具有多个发达的叶绿体、线粒体和淀粉核等。虫黄藻Cladocopium C1 sp.细胞内的叶绿体附近存在许多的淀粉颗粒和线粒体,和一些离散分布的碳酸钙晶体。

Figure 2. Symbiodiniaceae cultured after half of a year
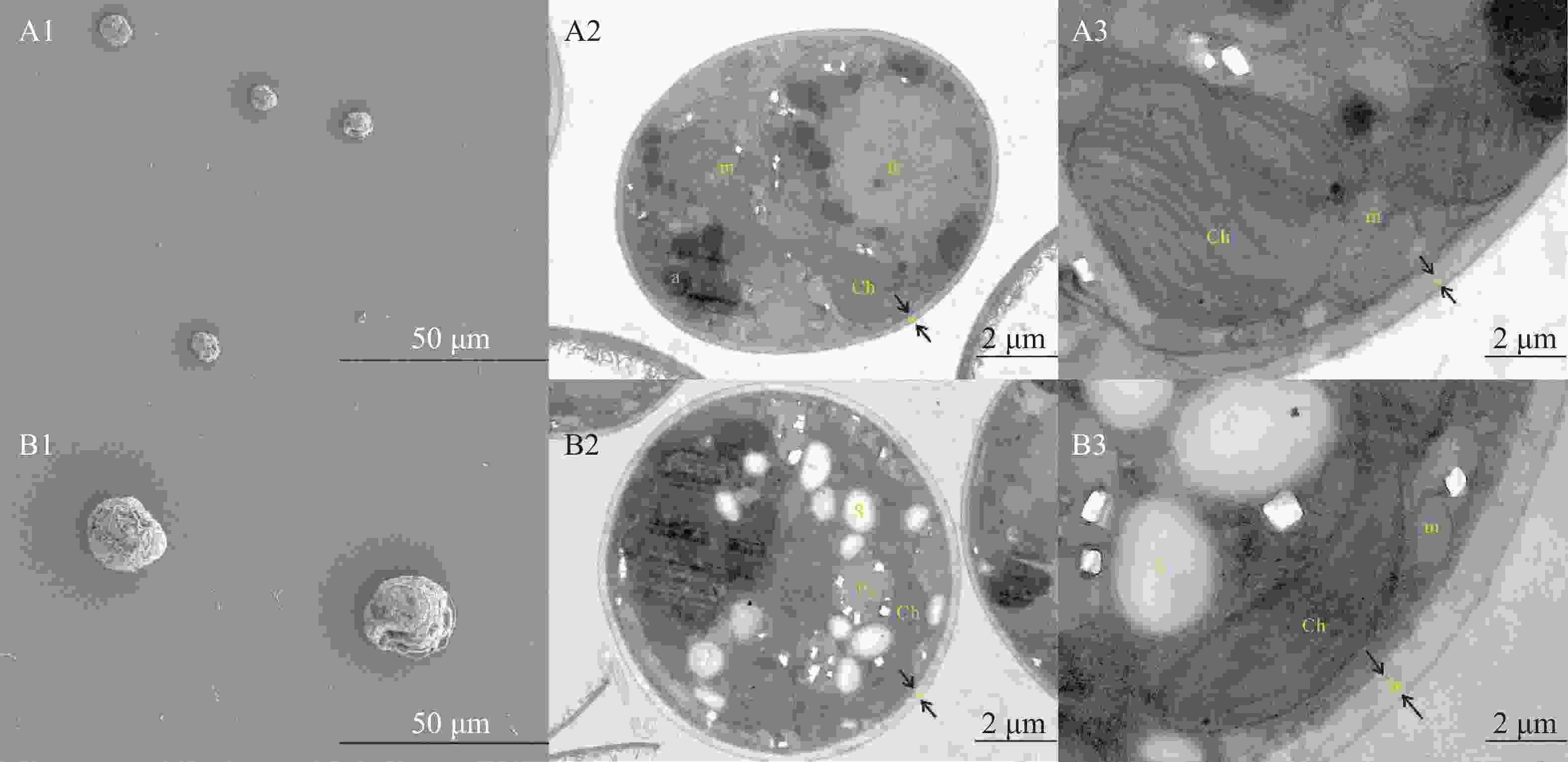
Figure 3. The cultured Symbiodiniaceae were observed under scanning electron microscope and transmission electron microscope
-
利用引物进行PCR扩增得到样品ITS序列片段 :GTCCATACGCGCATTGCACTCTTGGGACTTCCTGAGAGTATGTTTGCTTCAGTGCTTATTTTACCTCCTTGCAAGGTTCTGTCGCAACCTTGTGCCCTGGCCAGCCATGGGTTAACTTGCCCATGGCTTGCTGAGTAGTGATCTTTTAGAGCAAGCTCTGGCACGCTGTTGTTTGAGGCAGCCTATATTGAGGCTATTTCAAATGACGTTGCTACAAGCTTGATGTGTCCTTCTGCGCCGTTGCGCATCCCATAGCATGAAGTCAAACAAGAGAACCCGCTGAACTTAAGCTATTGGATCCCA。使用以上序列与 NCBI 数据库(
http://ncbi.nlm.nih.gov/ )进行比对。结果显示,该序列与数据库中Cladocopium C1 sp.(登录号长度:331 bp)的参考序列高度相似。具体的比对参数如下:最大得分(Max Score)为553,总得分(Total Score)为545,查询序列覆盖度(Query Cover)达到97%,匹配的E值为5e-151,序列一致性(Percent Identity)高达99.67%。以上数据综合表明,所测虫黄藻样品与Cladocopium C1具有极高的同源性。 -
第一次纯培养虫黄藻高通量测序结果如表1所示,第二次纯培养虫黄藻高通量测序结果如表3所示。表中的所有数据单位均为百分比,表示该物种在所测样品群落中的百分比。纯培养第一次的高通量检测结果显示了样品中微生物的数量和种类,结果包括了多个不同分类的虫黄藻系群。把所测样品编号为 A1、A2、A3,结果显示样品中第一主导共生虫黄藻为Cladocopium C1 sp.,且通过16S rRNA 并未检测出细菌污染。
分类单元 样本 A1 /% 样本 A2 /% 样本 A3/% Cladocopium C1 98.134 5 97.804 1 97.801 0 Effrenium E1 0.026 62 0 0.424 22 Symbiodinium A6 0.007 83 0 0.003 89 Durusdinium D1 0 0 0.233 51 Others 1.831 02 2.195 89 1.770 84 Table 1. Primary high-throughput sequencing of the pure culture
纯培养的第二次高通量测序检测结果相比于第一次高通量测序结果深度更大,覆盖更广。将第二次高通量测序的11个样品分别编号为S1~S11,从表2的数据中可以看出第二次高通量测序的11个样品中,第一优势种均是虫黄藻 Cladocopium C1 sp.,分别占据了总分类的97.28%、96.73 %、97.41%、97.38%、93.99%、97.26%、97.49%、97.75%、97.05%、96.88%、97.07%。
分类单元 样本S1/% 样本S2/% 样本S3/% 样本S4/% 样本S5/% 样本S6/% 样本S7/% 样本S8/% 样本S9/% 样本S10/% 样本S11/% Cladocopium C1 97.28 96.73 97.41 97.38 93.99 97.26 97.49 97.75 97.05 96.88 97.07 Durusdinium D1 0 0.72 0 0 3.39 0.63 0 0 0.35 0 0 其他 Others 2.7 2.56 2.56 3.84 2.64 2.71 2.52 2.26 2.6 3.12 2.92 Table 2. Second high-throughput sequencing of the pure culture
-
珊瑚礁作为海洋生态系统的核心组成部分,正面临气候变化、海水酸化、过度捕捞、陆源污染及旅游开发等多重威胁的叠加效应。其中,气候变暖引发的海水温度异常升高是导致珊瑚白化频发的关键因素。当海洋热浪事件持续超过珊瑚耐受阈值时,珊瑚会通过应激机制主动排出体内共生的虫黄藻(Symbiodiniaceae),导致共生关系破裂并引发白化现象[40]。这一过程中,虫黄藻作为珊瑚能量代谢的核心贡献者(提供宿主80%以上的光合产物)[41],其流失直接导致珊瑚能量匮乏、免疫力下降及钙化速率降低,进而引发珊瑚礁生态系统的级联崩溃。
本研究成功从美丽鹿角珊瑚中分离出虫黄藻Cladocopium C1 sp.,并且通过稀释培养后的虫黄藻Cladocopium C1 sp.纯度高达98%。并能在实验室内保持稳定的传代。实验中不同处理后的细胞分离和生长的初步结果表明:1)分离的过程会对虫黄藻细胞造成不同程度的机械损伤,可能让共生虫黄藻更加难以适应后期生长的环境;2)共生虫黄藻的种属繁多,具有遗传多样性会影响实验结果,这种遗传多样性可能导致没有对应的makers用来检测其种属;3)共生虫黄藻容易受到细菌、真菌、原生生物等污染,这些生物污染物及其代谢产生的化学污染会导致共生虫黄藻的生存环境受到影响,例如形成营养竞争,而共生虫黄藻在竞争关系中处于弱势地位。此前,共生虫黄藻分离过程中最大的挑战之一是消除培养系统中硅藻、原生动物和真菌的污染。尽管使用三联抗生素可以有效抑制培养物中的细菌生长,但是,在分离过程中源自珊瑚外骨骼或内部组织的真菌,硅藻和原生动物等污染物很难消除[42]。本实验通过对原始虫黄藻样品进行分离处理,让共生虫黄藻与其他污染物初步分离,并在实验的过程每两周补充三联抗生素处理,防止细菌等污染虫黄藻样品。尽管抗生素处理可抑制细菌增殖,但真菌与原生动物污染仍是离体培养的主要障碍[43]。近期研究表明,添加两性霉素B(0.25 μg·mL-1)或采用微流控芯片进行单细胞分离可提升污染控制效率[44]。
经过分离培养的虫黄藻Cladocopium C1 sp.未观察到鞭毛,这可能是由于共生状态下,虫黄藻褪去了鞭毛,从而形成珊瑚-虫黄藻共生体的状态,而分离的虫黄藻暂时未恢复游离的状态。在进化过程中,虫黄藻在宿主细胞内长期定居,鞭毛结构被认为在进化过程中逐渐退化或暂时抑制,以适应稳定的内共生环境[45]。Aranda等[46]通过比较三种共生虫黄藻基因组发现,编码鞭毛轴丝核心组分(如动力蛋白重链和驱动蛋白)的基因在3种虫黄藻基因组中均缺失或显著减少,这与它们在内共生阶段失去运动能力的现象一致。此外,有研究表明体外培养的共生虫黄藻的细胞壁可能比共生状态下的虫黄藻细胞壁明显厚(约两倍),这可能是共生的虫黄藻细胞为促进和珊瑚宿主之间营养物质的运输[47]。共生虫黄藻为适应共生体状态已经达到了微妙的代谢平衡和生理妥协(例如放弃有性生殖并与珊瑚的免疫系统兼容)[48]。 然而,它们最初是游离在海水中的生物,类似于裸子藻和其他密切相关的甲藻,在共生体进化的早期,它们是运动的、浮游的和自养的[49]。这表明共生状态和离体状态的虫黄藻会存在着细胞结构、运动方式、能量代谢及分裂方式的差异。当前环境压力下修复珊瑚礁的紧迫性, 珊瑚和幼体的培养和存活, 都必须从外界获得虫黄藻, 达到共生平衡[44]。所以, 离体培养虫黄藻, 大量培养后提供给共生体幼虫是珊瑚礁恢复的关键技术, 也有利于更好地研究虫黄藻与宿主间特殊的共生机制。但是, 复杂的共生关系和分离后变化的生存环境增加了虫黄藻离体培养的难度。
-
本研究经过半年的培养,确定分离的虫黄藻Cladocopium C1 sp.可以在实验室稳定的传代。倒置显微镜及电子显微的观察结果显示,虫黄藻Cladocopium C1 sp.细胞有着完整的细胞结构,细胞质内具有多个发达的叶绿体、线粒体和淀粉核等。在NCBI平台比对Sanger测序的结果,表明分离的虫黄藻为美丽鹿角珊瑚的主导虫黄藻Cladocopium C1 sp.,高通量测序结果表明虫黄藻Cladocopium C1 sp.细胞相对丰度为98%。成功分离出纯度高并能稳定传代虫黄藻为探索珊瑚与虫黄藻共生机制提供更多实验基础。
Isolation,culture and purification of the coral symbiotic alga Cladocopium C1 sp.
DOI: 10.15886/j.cnki.rdswxb.20250065
- Received Date: 2025-05-08
- Accepted Date: 2026-02-12
- Rev Recd Date: 2025-10-23
-
Key words:
- Symbiodiniaceae /
- Cladocopium C1 sp. /
- electron microscope /
- high-throughput sequencing /
- pure culture of isolates
Abstract: Coral reef ecosystem is one of the most biodiverse and productive ecosystems in the ocean.However, coral reefs have been degraded to different degrees in recent years due to the impact of global environmental changes.As microorganisms that live in symbiosis with corals, the isolation and purification of Symbiodiniaceae clades can provide a deeper understanding of their biological properties and provide an experimental basis for revealing their important physiology, such as coral-associated physiology mechanisms.In this context Cladocopium C1 sp.was isolated from Acropora formosa, and then rinsed or sheared to make the coral released symbiotic Symbiodiniaceae.Observations were made using an inverted microscope microscope to determine whether Symbiodiniaceae were isolated or not, and the Symbiodiniaceae were centrifuged, diluted, transferred, and cultured every two weeks.Sanger sequencing and high-throughput sequencing were performed on the cultured Symbiodiniaceae.Observations under inverted microscope and electron microscope showed that the cells of the Symbiodiniaceae Cladocopium C1 sp.had a complete structure with multiple well-developed chlorophylls, mitochondria, pyrenoids, etc in the cytoplasm.The Sanger sequencing results were compared on the NCBI platform, and the isolated Symbiodiniaceae were the dominant Symbiodiniaceae Cladocopium C1 sp.of A.formosa.The high-throughput sequencing results showed that the relative abundance of Symbiodiniaceae Cladocopium C1 sp.cells was 98%.The successful isolation of Cladocopium C1 sp.from A.formosa provides a scientific basis for a deeper understanding of the roles of Symbiodiniaceae in the symbiosis with corals and the mechanism of their interactions with corals.
| Citation: | Wei Lifei, Chen Shuchang, Lan Mengling, Yang Qizhi, Liang Hongye, Qin Zhenjun. Isolation,culture and purification of the coral symbiotic alga Cladocopium C1 sp.[J]. Journal of Tropical Biology. doi: 10.15886/j.cnki.rdswxb.20250065

|
 DownLoad:
DownLoad: